Submitted:
29 April 2026
Posted:
04 May 2026
You are already at the latest version
Abstract
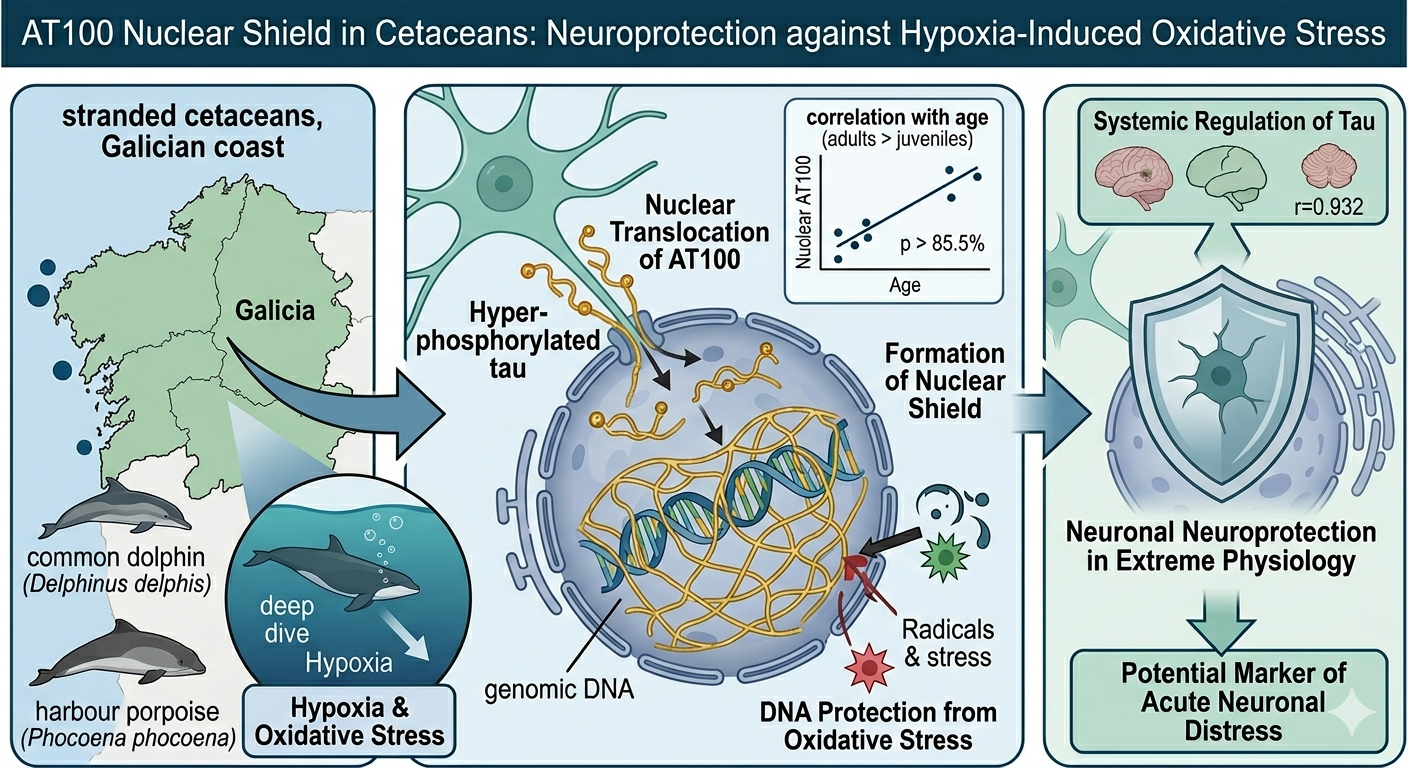
This study investigates the nuclear distribution of phosphorylated tau (AT100) in the frontal cortex and cerebellum of 12 cetaceans stranded along the Galician coast (NW Spain). Using Bayesian beta regression, we identified a strong positive correlation between aging and tau phosphorylation, with posterior probabilities of 85.5% in the cortex and 89.5% in the cerebellum. This age-associated increase suggests a systemic and coordinated regulation of tau as the brain matures. We propose that the translocation of tau to the nucleus could act as a nuclear shield against cumulative oxidative stress, a process potentially intensified by the intermittent hypoxia characteristic of diving in these mammals. Furthermore, the elevated AT100 levels observed in adults and in a case of cerebral necrosis reinforce its protective role and its potential as a marker for neuronal distress. While this exploratory pilot study is limited by sample size (n=12), the Bayesian analysis confirms biological consistency across the species analyzed. These findings establish a baseline for understanding neuroprotection in cetaceans, suggesting that tau phosphorylation is a key evolutionary mechanism for preserving neuronal genomic integrity under extreme physiological conditions.
Keywords:
cetaceans
; tau protein
; AT100
; brain aging
; nuclear translocation
; oxidative stress
; bayesian analysis
; neuroprotection
1. Introduction
Brain aging is a complex and multifaceted process involving structural and functional changes that affect cognition, memory, and behavior. In humans, age-related declines in neuroplasticity have been reported and are thought to contribute to cognitive changes associated with normal aging [1,2,3,4]. From a comparative perspective, cetaceans and humans exhibit notable similarities in brain organization, including high levels of encephalization, cortical complexity, and refined neuronal architecture [5,6,7,8]. These convergent traits are associated with complex social behavior and have been proposed to reflect shared principles of neural organization that may extend to aspects of brain development and aging [9,10,11]. In this context, cetaceans constitute valuable models for comparative studies, particularly given the exceptional longevity observed in some species [12].
At the cellular level, tau protein regulates cytoskeletal stability through its interaction with tubulin [13]. Beyond its structural role, tau is implicated in cellular stress responses and age-related neurodegenerative conditions, including Alzheimer’s disease [14,15]. Experimental studies have demonstrated that tau can bind DNA and exert protective effects against oxidative stress [16,17,18,19]. Under physiological conditions, tau phosphorylation is a dynamic process that modulates cytoskeletal organization [20,21]. Among phosphorylated tau species, the AT100 epitope (Thr212/Ser214) is recognized by specific conformational changes [22,23]. In mammalian brains, nuclear localization of AT100 has been associated with neuronal aging and DNA protection [24,25,26]. Conversely, in pathological states, nuclear AT100 immunoreactivity decreases as cytoplasmic aggregation increases [15,24,27]. This duality highlights its relevance as a marker of neuronal state [28]. The present study aimed to evaluate the nuclear localization of AT100 in the frontal cortex and cerebellum of several cetacean species stranded along the Galician coast.
2. Materials and Methods
2.1. Study Subjects
A total of 12 stranded cetaceans from the Galician coast were analyzed between February and August 2025, representing four species: Delphinus delphis (), Phocoena phocoena (), Tursiops truncatus (), and Stenella coeruleoalba ().
Samples were provided by the Coordinadora para o Estudo dos Mamíferos Mariños (CEMMA). Individuals were classified by sex and age category based on total body length, external development, and dental eruption patterns. Dental age estimation was performed by counting Growth Layer Groups (GLGs) [29,30,31]. Individuals were grouped into three classes: juveniles (minimal dental wear, incomplete sexual development), subadults (moderate wear, near-maximum body length), and adults (marked dental wear, complete sexual maturity) [32,33]. Frontal cortex and cerebellum samples were available for all specimens as detailed in Table 1.
2.2. Sample Analysis
All biological samples were immediately fixed after collection in 4% (v/v) formaldehyde solution to ensure proper preservation of tissue architecture. Subsequently, tissues were processed using conventional histological techniques, including dehydration through ascending ethanol series and embedding in paraffin blocks. Serial sections were then cut from these blocks using a microtome for histological and molecular analyses, specifically following standardized and validated immunohistochemistry protocols established in this study.
For immunohistochemical studies, sections were deparaffinized in xylene and rehydrated through graded ethanol series. Antigen retrieval was performed by heat-induced epitope retrieval in citrate buffer (pH 6.0) at 95°C for 20 minutes. Endogenous peroxidase activity was subsequently blocked with 3% hydrogen peroxide in methanol for 10 minutes, followed by blocking of nonspecific binding sites with 5% normal goat serum in PBS for 1 hour at room temperature.
Sections were incubated with the primary antibody AT100 (Invitrogen™ Phospho-Tau (Thr212, Ser214) Monoclonal Antibody (clone AT100, cat. MN1060)) at a 1:100 dilution in PBS containing 0.2% Triton X-100 for 2 hours at room temperature, according to the manufacturer’s instructions. Antibody binding was visualized using 0.025% 3,3′-diaminobenzidine (DAB Peroxidase Substrate Kit; Vector Laboratories) with 0.01% H2O2 as the catalytic agent. Finally, sections were mounted with Entellan (Merck).
Neuronal nuclei in the frontal cortex and cerebellum were quantified independently by two observers from the Pathology Service of the Veterinary Clinical Hospital of Universidad Alfonso X El Sabio (HCV-UAX) to minimize counting errors. Both observers were blinded to the age and sex of the individuals. Neuronal nuclei were specifically counted in each sample of frontal cortex and cerebellum. Five fields per immunohistochemical preparation were analyzed, using three samples per animal, at magnification in sections of the frontal cortex and lateral cerebellar hemispheres.
2.3. Statistical Analysis
Due to the small sample size () and the proportional nature of AT100 expression data (percent values ranging from 0 to 100%), Bayesian statistical methods were employed. These are particularly suitable for limited datasets as they allow the incorporation of prior biological knowledge and provide full uncertainty quantification via credible intervals [34,35]. It should be noted, however, that with small samples, posterior estimates remain sensitive to prior specification; poorly chosen priors can introduce bias that may exceed that of frequentist alternatives. To address this concern, informative priors were grounded in published mammalian tau literature, and a formal sensitivity analysis was conducted (Section 3.6) to confirm that the direction and magnitude of the estimated effects were robust across a range of prior specifications.
2.3.1. Bayesian Beta Regression Models
AT100 expression data were modeled using Bayesian Beta regression, appropriate for proportional data bounded between 0 and 1 [36]. The Beta distribution naturally accommodates the constraints of percentage data and the various distributional shapes commonly observed in biological proportions. Separate models were fitted for AT100 expression in the cerebral cortex and cerebellum:
where represents the expected proportion of AT100-positive neurons, is the concentration parameter (inversely related to variance), Age is an ordinal variable ( juvenile, subadult, adult), and Sex is a binary indicator ( male, female).
2.3.2. Prior Specification
Informative priors were defined based on existing literature on tau protein expression in mammalian brains [15,38]. For cortex models, the prior for the intercept was set as , reflecting expected baseline AT100 levels of ∼60% in young cetaceans. For cerebellum models, a slightly lower baseline was assumed: . Age effects received weakly informative positive priors: , based on literature suggesting increased tau phosphorylation with age. Sex effects were assigned noninformative priors centered at zero: . The concentration parameter was assigned an prior, favoring moderate dispersion while allowing high variability if supported by the data.
2.3.3. Model Fitting and Diagnostics
Models were fitted using PyMC (version 5.0+) with No-U-Turn Sampling (NUTS), a Hamiltonian Monte Carlo variant effective for complex hierarchical models [37]. Four chains were run with 2,000 warm-up and 2,000 posterior samples each, yielding a total of 8,000 posterior samples. Convergence was assessed via the Gelman–Rubin statistic () and effective sample sizes (). Model fit was evaluated through posterior predictive checks, comparing observed data distributions with distributions predicted from posterior parameter samples.
2.3.4. Interpretation Framework
Bayesian results were interpreted probabilistically rather than via classical null hypothesis testing. Posterior means with 95% credible intervals (CrI) are reported, representing the range containing the true parameter with 95% probability given the data and priors. The probability of positive effects, , is also reported, providing direct measures of evidence for age-related increases in AT100 expression. Effect sizes are reported on the logit scale, where positive values indicate increases in AT100 proportion and negative values indicate decreases.
3. Results
3.1. Immunohistochemical Analysis and AT100 Quantification
Manual quantitative evaluation of AT100-positive nuclei is summarized in Table 2.
The immunohistochemical analysis of AT100 protein generally revealed age-related differences in its subcellular distribution in both frontal cortex and cerebellum. In the cortex, quantification of positive nuclei showed a higher proportion of nuclear AT100 immunoreactivity in adult individuals, followed by subadults, whereas juveniles exhibited lower nuclear immunoreactivity and a more restricted distribution (Figure 1 and Figure 2).
As previously mentioned, the cerebellum exhibited the same age-dependent pattern, with adults showing the highest proportion of AT100 positive nuclei, followed by subadults, while juveniles displayed the lowest nuclear expression. Although quantification of AT100 in the cerebellum, as in the cortex, showed a progressive increase with age, the values obtained in the cerebellum were generally lower than those recorded in the cortex for the same age groups. This finding indicates that, while the age-associated increase pattern is maintained in both structures, the magnitude of nuclear staining in the cerebellum is comparatively lower than in the cortex. AT100 immunolabeling in the cerebellum revealed phosphorylated tau predominantly in granule cells and, to a lesser extent, in Purkinje cells (Figure 3 and Figure 4).
Overall, these results indicate a progressive increase in nuclear AT100 localization associated with advancing age in both analyzed brain regions.
3.2. Atypical AT100 Expression in Two Subadult D. delphis
Two subadult male D. delphis exhibited nuclear AT100 immunoreactivity levels indistinguishable from those of adult individuals: the first reached 96.55% in CC and 62.41% in Cb; the second, 88.7% in CC and 78.4% in Cb (Table 2, Figure 5). These values lie 1.4–2.1 standard deviations above the subadult group mean and overlap entirely with the adult range. One of these individuals presented macroscopic cerebral necrosis at necropsy. Necrotic processes trigger energy failure, ionic imbalance, and activation of stress-sensitive kinases (GSK-3, CDK5), all of which promote tau hyperphosphorylation and nuclear redistribution independently of age [20,21]. The second atypical individual showed no gross neuropathology, raising the possibility of subclinical injury or individual biological precocity. Taken together, these cases suggest that nuclear AT100 accumulation reflects both a physiological aging trajectory and an acute neuropathological response, supporting its dual utility as an aging marker and a potential indicator of neuronal distress.
3.3. Descriptive Statistics
AT100 expression was successfully quantified in all 12 cetacean individuals across both brain regions. In the cerebral cortex, AT100 expression ranged from 17.9% to 96.7% (mean ± SD: %), while in the cerebellum it ranged from 19.5% to 91.7% (mean ± SD: %). The sample included 3 juveniles, 5 subadults, and 4 adults, with 7 males and 5 females distributed across four cetacean species, predominantly Delphinus delphis ().
A notably strong positive correlation was observed between AT100 expression in the cerebral cortex and cerebellum (; 95% CI: 0.82–0.98), indicating highly coordinated patterns of tau phosphorylation across brain regions within individuals. This correlation suggests systemic rather than region-specific mechanisms underlying AT100 expression in cetacean brains. The distribution of individual values, differentiated by sex and age category, is illustrated in Figure 6, showing that the cortex–cerebellum relationship remains consistent across all age groups and both sexes.
3.4. Bayesian Regression Results
3.4.1. Model Convergence and Fit
Both models (cerebral cortex and cerebellum) showed excellent convergence diagnostics. All values were , well below the conventional threshold of 1.05, and effective sample sizes exceeded 4,000 for all parameters, indicating robust sampling of the posterior distribution. Posterior predictive checks confirmed good model fit, with predicted distributions closely matching observed patterns in both brain regions.
3.4.2. Age Effects on AT100 Expression
Bayesian analysis revealed consistent positive age effects in both brain regions, providing credible probabilistic evidence of age-related increases in AT100 expression in cetaceans. In the cerebral cortex, the estimated age effect was (95% CrI: ; ), with an 85.5% probability of a positive age effect. In the cerebellum, the age effect was slightly stronger: (95% CrI: ; ), with an 89.5% probability of a positive effect.
Although the 95% credible intervals include zero, reflecting the small sample size, the consistently high probabilities (>85%) of positive age effects provide substantial evidence of age-related tau accumulation. Effect sizes on the logit scale correspond to substantial increases in the proportion of AT100 across age categories, with adults showing markedly higher expression than juveniles in both brain regions.
3.4.3. Sex Effects on AT100 Expression
Sex effects were minimal and centered near zero in both brain regions. In the cerebral cortex, (95% CrI: ; ), and in the cerebellum, (95% CrI: ; ). These results provide strong evidence against sexual dimorphism in AT100 expression, consistent with the hypothesis that tau accumulation follows similar patterns in male and female cetaceans. Age and sex effects in both brain regions, along with their 80% and 95% credible intervals, are shown comparatively in Figure 7.
3.5. Regional Comparison
Comparison between the cerebral cortex and cerebellum revealed remarkably similar patterns of AT100 expression. Although mean expression was slightly higher in the cerebral cortex (57.3%) compared to the cerebellum (49.0%), age-related trends were nearly identical, and the strong inter-regional correlation () suggests coordinated tau phosphorylation mechanisms operating across different brain regions. Both regions displayed comparable age effect sizes and a similar absence of sex effects, indicating that AT100 expression patterns are consistent across distinct neural tissues in cetaceans. The distribution of values by age category and sex in both regions is shown in Figure 8.
3.6. Sensitivity Analysis for Prior Specification
To assess the robustness of the Bayesian results to prior choice, a sensitivity analysis was conducted comparing three specifications for the prior of : the Original prior used in the main analysis [], an Agnostic prior centered at zero with greater diffusivity [], and a Skeptical prior of equal precision to the original but with no directional bias []. Figure 9 shows the posterior distributions of under each scenario for both brain regions. All three posteriors exhibited substantial overlap and consistently pointed toward positive values, with probabilities ranging from 76.2% (Skeptical, cortex) to 92.1% (Agnostic, cerebellum). This stability indicates that the direction of the age effect is supported by the data and is not an artifact of the informative prior, although the estimated magnitude shows some sensitivity to prior specification, particularly in the cortex under the Skeptical prior.
Figure 10 presents model predictions back-transformed to percentage points (pp) for direct interpretation of the age effect. Under the Agnostic prior—the most neutral—the predicted difference in AT100 expression between a juvenile and an adult is pp in the cerebral cortex and pp in the cerebellum. Using the Original prior, these differences are pp and pp, respectively. The 95% credible interval bands are wide, reflecting the uncertainty inherent to the small sample size (), but in all scenarios, the predicted interval for adults lies clearly above that of juveniles, reinforcing the qualitative consistency of the observed age effect.
4. Discussion
The present study demonstrates that adult cetaceans exhibit a higher proportion of neurons with nuclear immunoreactivity for phosphorylated tau (p-tau) at the AT100 epitope in both the frontal cortex and cerebellum compared with subadult and juvenile individuals. Our Bayesian analysis, yielding posterior probabilities of 85.5% and 89.5% for positive age effects in the cortex and cerebellum respectively, provides robust evidence of an age-associated increase in tau phosphorylation, even within the constraints of a limited sample size () [34,35].
The pattern of AT100 immunolabeling observed in the frontal cortex is consistent with findings reported in humans and other mammals [28,29]. Similarly, age-related increases in hyperphosphorylated tau and changes in its subcellular localization have been described in non-human primates such as Callithrix jacchus and in murine models [25,30,42]. The presence of comparable patterns across phylogenetically distant mammalian lineages suggests that tau phosphorylation may be linked to conserved biological processes. In this context, the AT100 epitope (Thr212/Ser214), associated with conformational changes in tau [21], may represent a relevant indicator of age-related neuronal modifications, although its role as a biomarker should be interpreted with caution.
The progressive increase in phosphorylated tau with age is likely driven by multiple interacting mechanisms. Imbalances between kinase activity (including GSK-3 and CDK5) and phosphatase function (such as PP2A), together with reduced efficiency of proteasomal and autophagic degradation systems and increased oxidative stress, have been implicated in tau dysregulation [21,32]. These processes may not only promote hyperphosphorylation but also facilitate the redistribution of tau from axonal compartments to the soma and nucleus. Although these mechanisms were not directly assessed in the present study, they provide a biologically plausible framework for interpreting the observed patterns. Nuclear localization of tau has been proposed to play a protective role in maintaining DNA integrity and regulating chromatin organization under conditions of cellular stress [15,17,43], although this function may vary depending on the physiological or pathological context [20]. In the specific case of cetaceans, this nuclear migration may be fundamentally linked to the extreme physiological demands of their environment. Unlike terrestrial mammals, cetaceans are frequently exposed to ischemia-reperfusion events during diving, which trigger significant production of reactive oxygen species (ROS). This suggests that the nuclear translocation of p-tau could be a specialized response to an oxidative stress environment that is inherently more severe than that of terrestrial mammals, acting as a “nuclear shield” to protect the genome against hypoxia-induced damage throughout their long lifespans [18,19].
A particularly notable finding was the strong correlation between AT100 immunoreactivity in the frontal cortex and cerebellum (). This association suggests that tau phosphorylation may be regulated through coordinated, organism-level mechanisms rather than strictly region-specific processes. Such coordination may reflect shared regulatory pathways, common responses to age-related stressors, or synchronized developmental and aging programs affecting tau metabolism across distinct brain regions. From an evolutionary perspective, this type of systemic regulation could confer adaptive advantages by ensuring consistent neuroprotective responses in functionally interconnected neural circuits.
In the cerebellum, phosphorylated tau was predominantly localized in granule cells and, to a lesser extent, in Purkinje neurons, a pattern consistent with previous observations in other mammals [33]. Granule cells, as the principal excitatory interneurons of the cerebellum, play a central role in transmitting sensorimotor information and modulating Purkinje cell activity, which constitutes the primary output of the cerebellar cortex [34,35,36]. This distribution suggests that tau phosphorylation may influence cerebellar circuits involved in motor integration and coordination.
Furthermore, we observed that two subadult individuals of D. delphis exhibited nuclear AT100 immunoreactivity levels comparable to those observed in adults. Moreover, one of these individuals presented cerebral necrosis, with particularly elevated levels of nuclear labeling in both the cortex and cerebellum. Necrotic processes are associated with energy failure, ionic imbalance, and activation of stress-sensitive kinases, all of which can promote tau redistribution from axonal compartments to the soma and nucleus [20,32]. This finding suggests that increased nuclear tau may reflect not only physiological aging but also acute responses to neuronal injury. However, the potential contribution of infectious or concomitant pathologies cannot be excluded, as agents such as morbillivirus, Brucella ceti, and Toxoplasma gondii have been associated with central nervous system lesions in marine mammals [45,46,47,48].
Regarding biological traits, no significant differences were observed between males and females in AT100 immunolabeling patterns. This lack of variation suggests that tau regulation and function may be conserved across sexes, consistent with its fundamental role in neuronal homeostasis [20,21]. Our findings complement those reported by Orekhova et al. [38], who investigated -amyloid and phosphorylated tau in Mediterranean cetaceans using the AT180 and AT8 epitopes. While their study focused on pathological tau aggregates potentially associated with neurodegeneration, the AT100 pattern observed in our samples appears more consistent with physiological aging processes. These differences between epitopes (AT100 versus AT180 and AT8) may explain such discrepancies, highlighting the importance of epitope-specific approaches when interpreting tau pathology across different species.
The Bayesian analytical framework was appropriate for this study given the small sample size and the proportional nature of the outcome variables. The use of Beta regression ensured appropriate modelling of bounded percentage data, and sensitivity analyses across different prior specifications consistently supported the direction of the age effect. Nevertheless, it is critical to emphasize that this work represents an exploratory or pilot study. While our Bayesian analysis suggests biological consistency across the individuals analyzed—showing no evidence of notable differences between the species included (D. delphis, P. phocoena, T. truncatus, S. coeruleoalba)—the limited sample size () precludes definitive generalizations about all cetaceans. Several limitations should therefore be acknowledged, including the small sample size, the cross-sectional design, and the multispecies composition of the dataset. The absence of species as a covariate represents a potential source of confounding. However, a post-hoc inspection of residuals from the main models revealed no systematic clustering by species, and the two non-Delphinus adults (T. truncatus, S. coeruleoalba) fell within the range predicted for their age class, suggesting that species-level differences, if present, are smaller than age-related variance in this dataset. Formal multi-species modelling remains a priority for future studies with adequate per-species sample sizes.
5. Conclusions
This pioneering study provides the first description of the nuclear distribution of the AT100 phosphorylated tau epitope in the cetacean brain, further distinguished as the first work to analyze this protein within the cerebellum of these species. Our results demonstrate that the increase in nuclear AT100 immunoreactivity is significantly correlated with age in both the frontal cortex and the cerebellum. This suggests a coordinated, systemic regulatory mechanism within the central nervous system as the animals mature.
The high posterior probability obtained through Bayesian analysis confirms that the nuclear translocation of tau is a robust biological event in cetaceans stranded along the Galician coast. We propose that this phenomenon acts as an adaptive “nuclear shield”[17,43], intended to preserve genomic integrity against chronic oxidative stress and intermittent hypoxia—factors inherent to the extreme diving physiology of these marine mammals. Furthermore, the observation of elevated AT100 levels in a single individual presenting cerebral necrosis suggests that this protein may also serve as a sensitive marker of acute neuronal distress in the presence of severe injury.
Despite the exploratory nature of this pilot study and the inherent limitations of the sample size, the consistency of the patterns observed across the different species analyzed highlights the importance of this neuroprotective mechanism. This work not only fills a gap in the comparative neuroanatomy literature but also establishes a fundamental starting point for future research on brain aging and neuronal resilience strategies in marine mammals.
Abbreviations
The following abbreviations are used in this manuscript:
| AT100 | Phospho-Tau (Thr212/Ser214) monoclonal antibody clone AT100 |
| CC | Cerebral cortex |
| Cb | Cerebellum |
| CEMMA | Coordinadora para o Estudo dos Mamíferos Mariños |
| CrI | Credible interval |
| DAB | 3,3′-Diaminobenzidine |
| ESS | Effective sample size |
| GLG | Growth Layer Group |
| NUTS | No-U-Turn Sampler |
| pp | Percentage points |
| ROS | Reactive oxygen species |
Author Contributions
Conceptualization, B.A.-E.; methodology, B.A.-E. and M.O.; software, M.O.; validation, M.O., F.V.; formal analysis, B.A.-E. and M.O.; investigation, B.A.-E.; resources, X.P. and A.L.; data curation, F.V.; writing—original draft preparation, B.A.-E.; writing—review and editing, M.O. and F.V.; visualization, F.V.; supervision, F.V.; project administration, B.A.-E. and F.V.; funding acquisition, F.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Fundación Alfonso X el Sabio (Project No. 1018023).
Acknowledgments
The authors would like to thank the Coordinadora para o Estudo dos Mamíferos Mariños (CEMMA) and the staff of the stranding network for their invaluable assistance in sample collection along the Galician coast. Their expertise and logistical support were essential for the successful completion of this study. The authors carefully reviewed and edited the manuscript and take full responsibility for its content.
Conflicts of Interest
The authors declare no conflicts of interest.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the fact that all samples were obtained from carcasses of wild stranded animals during official necropsies conducted by the regional stranding network (CEMMA). No animals were sacrificed or captured for the purposes of this research, and the handling of the remains complied with current national and international regulations for marine mammals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to the preliminary nature of this pilot study.
References
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nature 2012, 489, 229–235. [Google Scholar] [CrossRef]
- Park, D.C.; Reuter-Lorenz, P. The adaptive brain: Aging and neurocognitive scaffolding. Annu. Rev. Psychol. 2009, 60, 173–196. [Google Scholar] [CrossRef]
- Raz, N.; Lindenberger, U.; Rodrigue, K.M.; Kennedy, K.M.; Head, D.; Williamson, A.; Dahle, C.; Gerstorf, D.; Acker, J.D. Regional brain changes in aging healthy adults: General trends, individual differences and modifiers. Cereb. Cortex 2005, 15, 1676–1689. [Google Scholar] [CrossRef]
- Greenwood, P.M.; Parasuraman, R. Neuronal plasticity and genetic variation in dopamine and acetylcholine receptors as determinants of individual differences in cognitive aging. Neuropsychol. Rev. 2010, 20, 111–126. [Google Scholar]
- Marino, L. Convergence of complex cognitive abilities in cetaceans and primates. Brain Behav. Evol. 2002, 59, 21–32. [Google Scholar] [CrossRef]
- Marino, L.; Connor, R.C.; Fordyce, R.E.; Herman, L.M.; Hof, P.R.; Lefebvre, L.; Lusseau, D.; McCowan, B.; Nimchinsky, E.A.; Pack, A.A.; et al. Cetaceans have complex brains for complex lives. PLoS Biol. 2007, 5, e139. [Google Scholar] [CrossRef] [PubMed]
- Manger, P.R. An examination of cetacean brain structure with a novel hypothesis on the evolution of cetacean brains. Biol. Rev. 2006, 81, 293–338. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.; Dicke, U. Evolution of the brain and intelligence. Trends Cogn. Sci. 2005, 9, 250–257. [Google Scholar] [CrossRef]
- Hof, P.R.; Van der Gucht, E. Structure of the cerebral cortex of the humpback whale, Megaptera novaeangliae (Cetacea, Mysticeti, Balaenopteridae). Anat. Rec. 2007, 290, 1331–1359. [Google Scholar] [CrossRef]
- Striedter, G.F. Principles of Brain Evolution; Sinauer Associates: Sunderland, MA, USA, 2005. [Google Scholar]
- Whitehead, H.; Rendell, L. The Cultural Lives of Whales and Dolphins; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- George, J.C.; Bada, J.; Zeh, J.; Scott, L.; Brown, S.E.; O’Hara, T.; Suydam, R. Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization. Can. J. Zool. 1999, 77, 571–580. [Google Scholar] [CrossRef]
- Povlishock, J.T.; Christman, C.W.; Ellis, E.F. The cytoskeleton in health and disease. J. Neuropathol. Exp. Neurol. 1999, 58, 715–732. [Google Scholar]
- Panegyres, P.K.; Zafiris-Toufexis, K. The biology of tau protein. J. Clin. Neurosci. 2002, 9, 6–12. [Google Scholar]
- Gil, L.; Hernández-Ortega, K.; García-Magro, N.; Martínez-Valbuena, I.; Al-Shahrour, F.; Ávila, J.; Rabano, A.; Hernández, F. AT100-tau immunoreactivity in the neuronal nucleus: An aging-related marker in the human brain. Front. Cell. Neurosci. 2017, 11, 352. [Google Scholar]
- Wei, Y.; Qu, M.H.; Wang, X.S.; Chen, L.; Wang, D.L.; Liu, Y.; Hua, Q.; He, R.Q. Binding to the minor groove of DNA by cytoskeletal protein tau at low pH. J. Biol. Chem. 2008, 283, 36390–36398. [Google Scholar]
- Violet, M.; Delattre, L.; Dewachter, I.; Lefebvre, B.; Galas, M.C. A major role for tau in neuronal DNA and RNA protection in vivo under physiological and hyperthermic conditions. Nucleic Acids Res. 2014, 42, 2961–2978. [Google Scholar] [CrossRef]
- Rossi, G.; Dalprà, L.; Crosti, F.; Lissoni, S.; Sciacca, F.L.; Catania, M.; Di Fede, G.; Tagliavini, F. Tau protein binds to DNA and chromatin and its chromosomal distribution is dependent on its phosphorylation state. J. Biol. Chem. 2008, 283, 30332–30342. [Google Scholar]
- Hua, Q.; He, R.Q. Tau protein binds to the distal end of the microtubule. FEBS Lett. 2003, 540, 95–99. [Google Scholar]
- Avila, J.; Lucas, J.J.; Pérez, M.; Hernández, F. Role of tau protein in both physiological and pathological conditions. Physiol. Rev. 2004, 84, 361–403. [Google Scholar] [CrossRef]
- Wang, Y.; Mandelkow, E. Tau in physiology and pathology. Nat. Rev. Neurosci. 2016, 17, 5–21. [Google Scholar] [CrossRef]
- Goedert, M.; Jakes, R.; Vanmechelen, E. Monoclonal antibody AT100 recognizes a conformational epitope of highly phosphorylated tau from Alzheimer’s disease brain. Biochem. J. 1994, 301, 871–877. [Google Scholar] [CrossRef]
- Gärtner, U.; Holzer, M.; Heumann, R.; Arendt, T. Induction of Alzheimer-like phosphorylation of the microtubule-associated protein tau in rat brain and liver by the inhibition of protein phosphatase 2A. Neuroscience 1998, 87, 567–575. [Google Scholar]
- Hernández-Ortega, K.; Ferrera, P.; Blanco-Alvarez, V.M.; Arias, C. Subcellular distribution of tau protein in the human brain: Nuclear tau as a hallmark of neuronal health. J. Alzheimers Dis. 2015, 45, 1169–1181. [Google Scholar]
- Rodríguez-Callejas, J.D.; Fuchs, E.; Pérez-Cruz, C. Evidence of tau hyperphosphorylation and dystrophic microglia in the common marmoset as a naturally occurring model for brain aging and tauopathy. Front. Aging Neurosci. 2016, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- Gil, L.; Ávila, J.; Hernández, F. Nuclear tau in Alzheimer’s disease and other tauopathies. Neurobiol. Aging 2021, 100, 1–12. [Google Scholar]
- Mansuroglu, Z.; Hummel, A.; Thiry, M.; Sergeant, N.; Delattre, L.; Dewachter, I.; Galas, M.C.; Lefebvre, B. Loss of nuclear tau in Alzheimer’s disease: A contributing factor to neurodegeneration? Front. Mol. Neurosci. 2016, 9, 146. [Google Scholar]
- Baquero, M.; Andreu-Reinón, M.E. The brain aging process. Front. Aging Neurosci. 2019, 11, 304. [Google Scholar]
- Lockyer, C. A review of factors involved in the efficiency of age determination from teeth of odontocetes. Rep. Int. Whal. Comm. 1995, 16, 95–109. [Google Scholar]
- Hohn, A.A.; Scott, M.D.; Wells, R.S.; Sweeney, J.C.; Irvine, A.B. Growth layers in teeth from known-age free-ranging bottlenose dolphins. Mar. Mammal Sci. 1989, 5, 315–342. [Google Scholar] [CrossRef]
- Perrin, W.F.; Myrick, A.C. Age Determination of Toothed Whales and Sirenians; IWC Special Issue 3; International Whaling Commission: Cambridge, UK, 1980. [Google Scholar]
- Perrin, W.F.; Reilly, S.B. Reproductive parameters of dolphins and small whales of the family Delphinidae. Rep. Int. Whal. Comm. 1984, 6, 97–133. [Google Scholar]
- Jefferson, T.A.; Webber, M.A.; Pitman, R.L. Marine Mammals of the World: A Comprehensive Guide to Their Identification; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- McElreath, R. Statistical Rethinking: A Bayesian Course with Examples in R and Stan, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Ferrari, S.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Hoffman, M.D.; Gelman, A. The No-U-Turn Sampler: Adaptively setting path lengths in Hamiltonian Monte Carlo. J. Mach. Learn. Res. 2014, 15, 1593–1623. [Google Scholar]
- Orekhova, K.; et al. Amyloid-beta and tau pathology in Mediterranean cetaceans. Sci. Rep. 2024, 14, 4521. [Google Scholar]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Ittner, L.M.; Götz, J. Amyloid-β and tau — a toxic pas de deux in Alzheimer’s disease. Nat. Rev. Neurosci. 2011, 12, 65–72. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Rodríguez-Callejas, J.D. Non-human primate models of Alzheimer’s disease. Methods Mol. Biol. 2023, 2561, 145–160. [Google Scholar]
- Schindowski, K.; Belarbi, K.; Buée, L. Neurogenesis: Reduction in Alzheimer’s disease or during physiological aging? Am. J. Pathol. 2008, 172, 1373–1386. [Google Scholar]
- Sultan, A.; Nesslany, F.; Violet, M.; Bégard, S.; Loyens, A.; Tulasne, D.; Buée-Scherrer, V.; Ittner, L.M.; Buée, L.; Galas, M.C. Nuclear tau, a key player in neuronal DNA protection. J. Biol. Chem. 2011, 286, 4566–4575. [Google Scholar] [CrossRef]
- Ávila, J.; Llorens-Martín, M.; Pallas-Bazarra, N.; Hernández, F. Role of tau protein in both physiological and pathological conditions. Front. Aging Neurosci. 2012, 4, 23. [Google Scholar]
- Domingo, M.; Visa, J.; Pumarola, M.; Marco, A.J.; Ferrer, L.; Llobera, R.; Kennedy, S. Pathologic and immunocytochemical studies of morbillivirus infection in striped dolphins (Stenella coeruleoalba). Vet. Pathol. 1992, 29, 1–10. [Google Scholar] [CrossRef]
- Di Guardo, G.; Marruchella, G.; Agrimi, U.; Kennedy, S. Morbillivirus infections in marine mammals: A review. Vet. Pathol. 2005, 42, 105–123. [Google Scholar]
- Guzmán-Verri, C.; González-Barrientos, R.; Hernández-Mora, G.; Morales, J.A.; Baquero-Calvo, E.; Chaves-Olarte, E.; Moreno, E. Brucella ceti and brucellosis in cetaceans. Emerg. Infect. Dis. 2012, 18, 175–176. [Google Scholar] [CrossRef]
- Dubey, J.P.; Zarnke, R.; Thomas, N.J.; Wong, S.K.; Van Bonn, W.; Reed, M.; Dierauf, L.; Stephens, I.D. Toxoplasmosis in marine mammals. J. Parasitol. 2003, 89, 752–757. [Google Scholar]
Figure 1.
AT100 immunolabeling in the frontal cortex () (D. delphis). Brown staining (DAB) in the neuronal nucleus indicates the nuclear localization of phosphorylated tau. (A) Juvenile individual showing low nuclear immunoreactivity. (B) Adult individual displaying abundant and intense nuclear staining (brown). Scale bars: A, B = 100 µm.
Figure 1.
AT100 immunolabeling in the frontal cortex () (D. delphis). Brown staining (DAB) in the neuronal nucleus indicates the nuclear localization of phosphorylated tau. (A) Juvenile individual showing low nuclear immunoreactivity. (B) Adult individual displaying abundant and intense nuclear staining (brown). Scale bars: A, B = 100 µm.

Figure 2.
AT100 immunolabeling in the frontal cortex () (P. phocoena). (A) Juvenile individual showing low nuclear immunoreactivity. (B) subadult individual displaying abundant and intense nuclear staining (brown). The qualitative difference between age groups is evident at this magnification. Scale bars: A, B = 100 µm.
Figure 2.
AT100 immunolabeling in the frontal cortex () (P. phocoena). (A) Juvenile individual showing low nuclear immunoreactivity. (B) subadult individual displaying abundant and intense nuclear staining (brown). The qualitative difference between age groups is evident at this magnification. Scale bars: A, B = 100 µm.

Figure 3.
AT100 immunolabeling in the cerebellum () (D. delphis). (A) Juvenile individual showing faint nuclear signal. (B) Adult individual with intense nuclear staining, mainly in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.
Figure 3.
AT100 immunolabeling in the cerebellum () (D. delphis). (A) Juvenile individual showing faint nuclear signal. (B) Adult individual with intense nuclear staining, mainly in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.

Figure 4.
AT100 immunolabeling in the cerebellum () (P. phocoena). (A) Juvenile individual showing faint nuclear signal. (B) Subadult individual with more extensive nuclear staining, mainly in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.
Figure 4.
AT100 immunolabeling in the cerebellum () (P. phocoena). (A) Juvenile individual showing faint nuclear signal. (B) Subadult individual with more extensive nuclear staining, mainly in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.

Figure 5.
AT100 immunolabeling in the cerebellum (A) and cortex (B) () of a subadult D. delphis individual, showing intense nuclear signal in cerebellum and frontal cortex, primarily in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.
Figure 5.
AT100 immunolabeling in the cerebellum (A) and cortex (B) () of a subadult D. delphis individual, showing intense nuclear signal in cerebellum and frontal cortex, primarily in granule cells and, to a lesser extent, in Purkinje cells. Scale bars: A, B = 100 µm.

Figure 6.
Correlation of AT100 expression between the cerebral cortex and cerebellum (, Pearson ). Each point represents an individual, differentiated by age category (marker shape) and sex (color: blue = male, orange = female). The gray dashed line represents the regression line, and the dotted line represents the 1:1 relationship. The strong positive correlation indicates systemic coordination of tau phosphorylation patterns across both brain regions, independent of sex or age class.
Figure 6.
Correlation of AT100 expression between the cerebral cortex and cerebellum (, Pearson ). Each point represents an individual, differentiated by age category (marker shape) and sex (color: blue = male, orange = female). The gray dashed line represents the regression line, and the dotted line represents the 1:1 relationship. The strong positive correlation indicates systemic coordination of tau phosphorylation patterns across both brain regions, independent of sex or age class.

Figure 7.
Bayesian effect estimates for the cerebral cortex (blue) and cerebellum (red). Thick lines represent the 80% credible interval (80% CrI), and thin lines represent the 95% CrI; circles indicate the posterior mean. Upper panel: age effect (), showing positive values in both regions with greater magnitude in the cerebellum. Lower panel: sex effect (), centered around zero in both regions, indicating no sexual dimorphism in AT100 expression. The vertical dashed line at zero serves as a reference. All effects are reported on the logit scale.
Figure 7.
Bayesian effect estimates for the cerebral cortex (blue) and cerebellum (red). Thick lines represent the 80% credible interval (80% CrI), and thin lines represent the 95% CrI; circles indicate the posterior mean. Upper panel: age effect (), showing positive values in both regions with greater magnitude in the cerebellum. Lower panel: sex effect (), centered around zero in both regions, indicating no sexual dimorphism in AT100 expression. The vertical dashed line at zero serves as a reference. All effects are reported on the logit scale.

Figure 8.
AT100 expression (% of positive nuclei) by age category (Juvenile, Subadult, Adult) in the cerebral cortex (left panel) and cerebellum (right panel). Circles represent individual data points, and squares indicate additional observations; males are shown in blue and females in orange. A progressive increase in AT100 expression with age is observed in both brain regions, most pronounced during the transition from subadult to adult. Values are consistent across sexes, supporting the absence of sexual dimorphism. Note the considerable inter-individual variability within each age category, especially among subadults.
Figure 8.
AT100 expression (% of positive nuclei) by age category (Juvenile, Subadult, Adult) in the cerebral cortex (left panel) and cerebellum (right panel). Circles represent individual data points, and squares indicate additional observations; males are shown in blue and females in orange. A progressive increase in AT100 expression with age is observed in both brain regions, most pronounced during the transition from subadult to adult. Values are consistent across sexes, supporting the absence of sexual dimorphism. Note the considerable inter-individual variability within each age category, especially among subadults.

Figure 9.
Sensitivity analysis: posterior distributions of under three prior specifications (Original, Agnostic, Skeptical) for the cerebral cortex (left) and cerebellum (right). Thin dashed lines indicate the reference priors; shaded bands represent the 95% credible interval of each posterior. The stability of the posteriors across scenarios indicates that the data support the direction of the effect independently of the prior used.
Figure 9.
Sensitivity analysis: posterior distributions of under three prior specifications (Original, Agnostic, Skeptical) for the cerebral cortex (left) and cerebellum (right). Thin dashed lines indicate the reference priors; shaded bands represent the 95% credible interval of each posterior. The stability of the posteriors across scenarios indicates that the data support the direction of the effect independently of the prior used.

Figure 10.
Predicted AT100 expression (%) by age category under the Original prior (red) and Agnostic prior (blue), with 95% credible interval (CrI) bands. Observed data points are overlaid (color = sex; shape = age category). The width of the bands reflects the true uncertainty associated with the sample size (). Both priors agree on the direction of the age effect; differences in magnitude are moderate.
Figure 10.
Predicted AT100 expression (%) by age category under the Original prior (red) and Agnostic prior (blue), with 95% credible interval (CrI) bands. Observed data points are overlaid (color = sex; shape = age category). The width of the bands reflects the true uncertainty associated with the sample size (). Both priors agree on the direction of the age effect; differences in magnitude are moderate.


Table 1.
Distribution of cetacean specimens by species, sex, and age class. All individuals were classified with a decomposition code of 1 (fresh).
Table 1.
Distribution of cetacean specimens by species, sex, and age class. All individuals were classified with a decomposition code of 1 (fresh).
| Species | Age class * | Sex | Length (cm) | Weight (kg) |
|---|---|---|---|---|
| Delphinus delphis | Adult | M | 218 | 95.2 |
| Delphinus delphis | Adult | M | 224 | 200.0 |
| Delphinus delphis | Adult | F | 194 | 110.3 |
| Delphinus delphis | Subadult | M | 159 | 58.3 |
| Delphinus delphis | Subadult | M | 196 | 75.6 |
| Delphinus delphis | Subadult | M | 196 | 74.8 |
| Delphinus delphis | Subadult | F | 170 | 53.2 |
| Delphinus delphis | Juvenile | M | 148 | 35.4 |
| Tursiops truncatus | Adult | F | 278 | 223.0 |
| Stenella coeruleoalba | Juvenile | F | 126 | 23.1 |
| Phocoena phocoena | Subadult | M | 164 | 52.6 |
| Phocoena phocoena | Juvenile | F | 116 | 29.0 |
* Age categories were assigned according to body size and sexual maturity: juvenile, subadult, adult.
Table 2.
AT100 expression data in cerebral cortex (CC) and cerebellum (Cb). Counts are expressed as total nuclei counted / AT100-positive nuclei (5 fields at magnification); AT100 (%) = percentage of positive nuclei.
Table 2.
AT100 expression data in cerebral cortex (CC) and cerebellum (Cb). Counts are expressed as total nuclei counted / AT100-positive nuclei (5 fields at magnification); AT100 (%) = percentage of positive nuclei.
| Scientific name | Age class | Sex | CC count | AT100 CC (%) | Cb count | AT100 Cb (%) |
|---|---|---|---|---|---|---|
| D. delphis | Adult | M | 156/122 | 78.2 | 156/120 | 76.9 |
| D. delphis | Adult | F | 140/96 | 68.5 | 159/78 | 49.0 |
| D. delphis | Adult | M | 140/78 | 55.7 | 156/75 | 48.0 |
| D. delphis | Juvenile | M | 140/25 | 17.9 | 154/30 | 19.5 |
| D. delphis | Subadult | M | 142/67 | 47.2 | 139/68 | 48.9 |
| D. delphis | **Subadult | M | 145/140 | 96.55* | 149/93 | 62.41* |
| D. delphis | **Subadult | M | 142/126 | 88.7* | 156/123 | 78.4* |
| D. delphis | Subadult | F | 147/47 | 32.2 | 158/32 | 20.5 |
| D. delphis | Juvenile | M | 140/25 | 17.9 | 154/30 | 19.5 |
| T. truncatus | Adult | F | 156/151 | 96.7 | 170/156 | 91.7 |
| S. coeruleoalba | Juvenile | F | 140/30 | 21.4 | 123/28 | 22.7 |
| P. phocoena | Subadult | M | 136/72 | 52.94 | 142/70 | 49.2 |
| P. phocoena | Juvenile | F | 147/47 | 32.2 | 156/32 | 20.5 |
* CC = cerebral cortex; Cb = cerebellum. ** Subadult individuals showing adult-like AT100 expression levels.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



