Submitted:
28 April 2026
Posted:
29 April 2026
You are already at the latest version
Abstract
Background/Objectives: Elastin-derived peptides (EPs) from food sources may be multifunctional dietary components that support metabolic and vascular health. However, their in vivo physiological actions remain incompletely understood. This study investigated the effects of bonito bulbus arteriosus-derived EPs on glucose metabolism, GLP‑1 elevation and enhanced early-phase insulin secretion, and renal vascular integrity in stroke-prone spontaneously hypertensive rats (SHRSP) with glucose intolerance. Methods: Male SHRSP were administered EPs orally as a single dose (1,000 mg/kg) or 4-week regimen (600 mg/kg/day). Glucose tolerance, plasma GLP‑1 and insulin levels, and blood glucose levels were measured following glucose loading. Renal morphology was assessed histologically. Dpp4, Icam‑1, and Agtr1 expression was quantified in glomerular and leukocyte fractions. Leukocyte oxidative signaling was evaluated by quantifying reactive oxygen species production associated with inducible nitric oxide synthase (iNOS). Age‑matched Wistar-Kyoto rats were included as normotensive controls. Results: A single dose increased plasma GLP‑1 and insulin levels and improved glucose tolerance compared with controls. The 4‑week regimen resulted in sustained improvements in glucose tolerance, without changes in blood pressure, a lower nephrosclerosis incidence, and reduced renal and leukocytic inflammatory marker expression. Dpp4, Icam‑1, and Agtr1 expression was downregulated and leukocyte iNOS‑driven oxidative signaling was reduced. These effects occurred despite the modest DPP‑IV inhibitory activity of EPs. Conclusions: Food-derived EPs exert multi-target physiological actions, including GLP‑1 elevation with enhanced early-phase insulin secretion and leukocyte oxidative and inflammatory response suppression, that improve metabolic and renal vascular outcomes. EPs warrant further investigation as candidate functional food ingredients for metabolic and vascular health.
Keywords:
functional food
; bioreactive peptide
; elastin peptide
1. Introduction
Japan has approximately 350,000 chronic dialysis patients, according to the Japanese Society for Dialysis Therapy Renal Data Registry [1]. A shift in primary renal diagnoses in Japan was reported between 2006 and 2020, during which the number of nephrosclerosis diagnoses increased, whereas that of glomerulonephritis and diabetic nephropathy diagnoses declined [2]. The incidence of nephrosclerosis has surpassed that of glomerulonephritis, making it the second most common cause of chronic dialysis and end-stage kidney disease. Hypertensive nephrosclerosis is the leading cause of end-stage renal disease in Japan. Hypertensive kidney disease primarily affects the afferent arterioles and glomeruli, and activation of the renin–angiotensin system is well-established in its pathogenesis [3].
Dipeptidyl peptidase-IV (DPP-IV; CD26, EC 3.4.14.5) is a homodimeric type II transmembrane glycoprotein with a large extracellular catalytic domain. The soluble form of DPP-IV is also present in body fluids such as plasma and saliva [4]. DPP-IV cleaves X-Pro/X-Ala dipeptides from the N-terminus of substrates, thereby modulating the activity of various peptide hormones and cytokines [5,6]. DPP-IV rapidly inactivates incretins such as glucagon-like peptide-1 (GLP-1), which regulate postprandial insulin secretion [7]. Several DPP-IV inhibitors are widely used to treat type 2 diabetes because they have a low risk of hypoglycemia and neutral effects on body weight [8]. Potential renoprotective effects of DPP-IV inhibitors via blood vessel regulation, possibly via increased GLP-1 bioavailability and anti-inflammatory or antioxidant mechanisms, have been reported [9].
Clinically used DPP-IV inhibitors are generally well-tolerated but can cause gastrointestinal and dermatologic adverse effects [10]. Natural peptides with DPP-IV inhibitory activity are being investigated as dietary bioactive compounds with potential metabolic benefits. We previously reported that elastin peptides (EPs) derived from fish bulbus arteriosus exert vasculoprotective effects that improve vascular reactivity without lowering blood pressure in spontaneously hypertensive rats [11]. We also reported that solubilized EPs suppress cellular senescence and inflammation through DPP-IV inhibition [12]. However, the mechanism of action of EPs in blood pressure-independent vascular protection has not yet been fully elucidated.
In this study, we aimed to elucidate the beneficial effects of EPs and the mechanisms underlying the prevention of hypertensive arterial changes without changes in blood pressure.
2. Materials and Methods
2.1. Materials
2.2. Animals
Male Wistar-Kyoto (WKY/KPO, WKY) rats were used as normotensive controls, and stroke-prone spontaneously hypertensive rats (SHRSP/KPO, SHRSP) were used as the experimental model (Kindai University, Osaka, Japan) [14]. The animals were maintained at 23 ± 1 °C and 55% ± 5% relative humidity under a 12 h light-dark cycle, housed in standard cages (two rats per cage), and given free access to a Funabashi SP diet (Funabashi Farm Co., Ltd, Chiba, Japan) and water. All animals were acclimatized for 7 days before the experiment. The experimental protocol was approved by the Kindai University Animal Experiment Committee and conducted in strict accordance with the Kindai University Animal Experiment Regulations (approval no. KAAG-2024-003) and ARRIVE guidelines [15].
2.3. Peptide Derivative Identification in the Plasma of Rats Administered EP
EP (1 g/kg) was administered to SHRSP at 12 weeks of age by oral gavage using a gastric tube. Portal blood samples were collected using a syringe treated with 5% EDTA-2Na/0.9% NaCl under anesthesia induced 5 min before collection with a combination of pentobarbital sodium (Nacalai Tesque Inc., Kyoto, Japan) and isoflurane (FUJIFILM Wako Pure Chemical Co., Osaka, Japan). Blood samples were centrifuged at 16 500 × g and 4 °C for 10 min to obtain plasma. Plasma was mixed with three volumes of ethanol and centrifuged again at 16 500 × g and 4 °C for 10 min. The supernatant was used as the deproteinized plasma fraction.
Peptide derivatives detected in rat portal plasma after EP administration were quantified using liquid chromatography–tandem mass spectrometry in multiple reaction monitoring mode using a previously established method [16].
2.4. Oral Glucose Tolerance Test
The rats were fasted for 15 h before oral administration of a 20% glucose solution (1 mL/100 g body weight) by oral gavage via a gastric tube. The EP-treated group was administered 1 g/kg EP in a 20% glucose solution. Rats were kept in an incubator (NK-210-3; Natsume Seisakusho Co., Ltd., Tokyo, Japan) at 40 °C for 10 min, and blood samples were collected from the tail vein at specific time (0, 15, 30, 60, and 120 min) intervals following glucose administration. A solution containing 1.5% EDTA-2Na (DOJINDO LABORATORIES, Kumamoto, Japan) in phosphate-buffered saline (PBS; FUJIFILM Wako Pure Chemical Co.) and 100 KIU/mL aprotinin (FUJIFILM Wako Pure Chemical Co.) in 0.9% NaCl was used as an anticoagulant and protease inhibitor. Blood samples were centrifuged at 820 × g and 4 °C for 10 min, and the resulting plasma was stored at −80 °C until analysis. Plasma GLP-1 levels (GLP-1 ELISA Kit Wako, High Sensitive, 299-75501; FUJIFILM Wako Pure Chemical Co.) and insulin concentrations (LBIS Rat Insulin ELISA Kit, AKRIN-010T; FUJIFILM Wako Shibayagi Co., Gunma, Japan) were measured using commercially available kits. Blood glucose levels were measured using a compact blood glucose meter (Glutest Aqua; SANWA KAGAKU KENKYUSHO Co., Ltd., Aichi, Japan).
2.5. Effects of EP Intake on Hypertensive Kidney Failure
2.5.1. Animals
Twelve-week-old male SHRSP (n = 19) and WKY (n = 9) rats were used. The SHRSP were randomly divided into two groups. One group received a stock chow diet without EP (Control group; n = 9), whereas the other group received a diet with EP at 600 mg/kg body weight/day (EP group; n = 10), administered twice daily via oral gavage with a gastric tube. The content and weight of administration were chosen because Pro-Gly (PG), the main degradation product of EP, is metabolized within approximately 4 h [11]. Blood pressure and body weight were measured once weekly. Tail arterial blood pressure was measured using a non-invasive blood pressure monitoring system (BP-98A; Softron Co., Ltd., Tokyo, Japan). The experiment was conducted for 4 weeks. At the end of the experiment, blood samples were collected from the abdominal vena cava under anesthesia using a heparinized syringe (Heparin Sodium Injection-N “AY”; AY PHARMACEUTICALS Co., Ltd., Tokyo, Japan). The brain, heart, and kidneys were excised, weighed, and examined macroscopically. Gross examination of the excised kidneys was performed, and cases showing renal atrophy together with fine granular changes on the kidney surface were regarded as nephrosclerosis.
2.5.2. Histological Examination
The right kidney from each rat was fixed in 10% neutral buffered formalin (FUJIFILM Wako Pure Chemical Co.). Paraffin-embedded tissues were sectioned into 2.5 µm-thick slices using a sliding microtome (Sakura Finetek Japan Co., Ltd., Tokyo, Japan), deparaffinized with xylene, and rehydrated through a graded alcohol series. Structural changes in the kidney were evaluated microscopically after hematoxylin–eosin staining. Histological observation was performed using the BZ-H3 application on an All-in-One Fluorescence Microscope (BZ-X800E; Keyence, Osaka, Japan), and the number of vascular necroses per section per individual was counted. All pathological evaluations were performed under blinded conditions. The pathologists who assessed the specimens were unaware of the experimental groups. This blinding procedure was implemented to minimize observer bias and ensure the objectivity of the morphological assessment.
2.5.3. Real-Time Polymerase Chain Reaction
The glomerular fraction from the left kidney was isolated as previously described [17]. The leukocyte-rich fraction (LRF) was isolated from blood as previously described [18]. Total RNA was extracted from both the glomerular and LRFs using the ReliaPrep™ RNA Cell Miniprep System (Promega, Z6010, Madison, WI, USA). Sample optical density was measured at 230, 260, and 280 nm using an e-Spect ES2 Spectrophotometer (BM Equipment Co., Ltd., Tokyo, Japan). Total RNA was reverse transcribed using the ExScript RT Reagent Kit (Applied Biosystems, Carlsbad, CA, USA) under the following conditions: 25 °C for 10 min, 30 °C for 120 min, 85 °C for 5 s; and stored at 4 °C. Real-time polymerase chain reaction was performed using an ABI PRISM® 7900HT system (Applied Biosystems) with TB Green® Premix Ex Taq™ (Tli RNaseH Plus; Takara Bio Inc., Shiga, Japan) under the following conditions: 95 °C for 30 s (one cycle), followed by 40 cycles of denaturation at 95 °C for 5 s and extension at 60 °C for 30 s. Specific primers were purchased from Takara Bio Inc., and mRNA expression levels were normalized to those of 18S rRNA.
2.5.4. Measurement of Nitric Oxide Release from the LRF-Rich Fraction
The LRF-rich fraction was isolated from 20-week-old SHRSP (six rats per group). Fluorescent indicator diaminofluoresceins (DAFs) enable bioimaging of nitric oxide (NO) production [19]. To measure NO release from leukocytes into the suspension medium, the membrane-impermeable indicator DAF-2 (1 μM; Cayman Chemical Co., Ann Arbor, MI, USA) was added to the LRF suspension (1 × 105 cells/mL in PBS) in a 96-well black plate (SPL Life Science Co., Ltd., Pocheon-si, Gyeonggi-do, Korea) and incubated for 30 min at 37 °C. Fluorescence was measured using a multilabel counter (Wallac ARVO 1420; PerkinElmer Inc., Waltham, MA, USA; excitation 495 nm, emission 515 nm). For inducible NO synthase (iNOS) stimulation experiments, LRFs were treated with 10 ng/mL lipopolysaccharide (LPS), a major component of the outer membranes of gram-negative bacteria (FUJIFILM Wako Pure Chemical Co.). Furthermore, for DPP-IV stimulation or inhibition experiments, LRFs were treated with 2.5 mU/100 µL DPP-IV (Sigma-Aldrich Co. LLC., St. Louis, MO, USA). For peptide inhibition experiments using EP hydrolysate products (100 µM), the following peptides were used: H-Gly-Hyp-Gly-OH (GOG; ABclonal Technology, Woburn, MA, USA), H-Val-Pro-OH (VP), cyclo(-Gly-Pro) (cGP; Bachem Holding AG, Basel-Landschaft, Switzerland), cyclo(L-Pro-L-Val) (cPV; Tokyo Chemical Industry Co., Ltd., Tokyo, Japan), and H-Pro-Gly-OH (PG; KOKUSAN CHEMICAL Co., Ltd., Tokyo, Japan). These peptides were added simultaneously to the LRF suspension.
2.6. Statistical Analysis
Results are expressed as mean ± standard deviation. SPSS Statistics Base version 29 (IBM, Armonk, NY, USA) was used for statistical analysis. For two-group comparisons, Student’s t-test or Welch’s t-test was applied according to the results of the F-test. For analyses involving three or more groups, Tukey’s honestly significant difference test or Dunnett’s test was used following the Shapiro–Wilk normality test and analysis of variance. Differences were considered statistically significant at p < 0.05.
2.7. Artificial Intelligence Declaration
During the preparation of this manuscript, the authors used Copilot, developed by OpenAI (Microsoft Co., Redmond, WA, USA), for the purposes of proofreading and enhancing the manuscript’s clarity. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
3. Results
3.1. Quantification in Portal Vein Plasma Samples After EP Administration
Figure 1 shows the plasma concentrations of EP degradation products over time following oral administration of EPs in SHRSP. Five specific peptides—GOG (Figure 1a), VP (Figure 1b), PG (Figure 1c), cGP (Figure 1d), and cPV (Figure 1e)—were detected following oral administration. Their concentrations increased rapidly, peaked between 30 and 60 min, and then gradually decreased.
The DPP-IV inhibitory activity of each peptide was evaluated at concentrations corresponding to their plasma levels. The observed inhibitory activities were as follows: GOG (0.25–0.3 µM), 8.73%; VP (0.02–0.04 µM), 11.1%; PG (2 µM), 10.4%–11.4%; cGP (1.5 µM), 10.8%; and cPV (0.1 µM), 0.67%. Half maximal inhibitory concentration (IC50) values were as follows: sitagliptin, 9.5 × 10⁻¹⁰ µM; GOG, 163.2 µM; VP, 10.68 µM; those for PG, cGP, and cPV were not detected because of low activity.
3.2. Comparison of Strain Differences in Insulin and Glucose Levels
Figure 2 shows a comparison of plasma insulin and blood glucose levels between WKY rats and SHRSP. In WKY rats, plasma insulin reached its maximum level 15 min after glucose loading, followed by a sharp decrease in blood glucose levels. In contrast, insulin secretion in SHRSP was significantly lower than that in WKY rats at 15 and 30 min (p < 0.05), and slightly increased at 60 min. Blood glucose levels in SHRSP were significantly higher than those in WKY rats from baseline to 60 min after glucose loading (p < 0.05); however, the difference disappeared at 120 min.
3.3. Effect of EPs on Glucose Metabolism
The effects of simultaneous administration of glucose and EP on glucose metabolism were examined (Figure 3). In the SHRSP EP group, GLP-1 (an incretin) and insulin levels were significantly higher than those in the SHRSP Control group 15 min after glucose loading (p < 0.05; Figure 3a, b). Blood glucose levels in the SHRSP EP group increased more slowly than those in the SHRSP Control group and were significantly lower than those in the SHRSP Control group from 15 to 60 min (p < 0.05; Figure 3c).
3.4. Effects of EP Intake on Hypertensive Kidney Failure
3.4.1. Chronological Changes in Blood Pressure
Figure 4 shows changes in blood pressure and body weight. Blood pressure was significantly higher in both SHRSP groups (Control and EP) than in the WKY group at all ages (p < 0.05; Figure 4a). Body weight throughout the experimental period was significantly lower in both SHRSP groups than in the WKY group (p < 0.05; Figure 4b). No significant differences in blood pressure or body weight were observed between the SHRSP Control and SHRSP EP groups, indicating that EP administration did not affect these parameters.

3.4.2. Organ Weight and Morphological Observations
Table 1 shows the organ weights. Although no differences were observed in brain and heart weights among the three groups, kidney weight was significantly lower in the two SHRSP groups than in the WKY group (p < 0.01).
No cases of nephrosclerosis were observed in the normotensive WKY rats (Figure 5a, d). Atrophy and structural irregularities of the renal cortex were observed in the SHRSP Control group (Figure 5b, e). Fewer cases of renal atrophy were observed in the SHRSP EP group than in the SHRSP Control group (Figure 5c, f). Moreover, the number of vascular necroses in the SHRSP EP group was significantly lower than in the SHRSP Control group (21 ± 12 vs. 12 ± 12; p < 0.05).
3.4.3. mRNA Expression of Dpp4, Icam-1, and Agtr1 in the Renal Glomerular Fraction
To estimate the effects of EP and its degradation products on the expression of factors related to inflammatory and immune responses, we investigated the mRNA expression of the following genes: Dpp4 (a marker of inflammation, Figure 6a), intercellular adhesion molecule 1 (Icam-1; an endothelial cell adhesion molecule, Figure 6b), and angiotensin receptor type 1 (Agtr1; an indicator of the renin–angiotensin system; Figure 6c). The Dpp4 mRNA level in the renal glomerular fraction was significantly higher in the SHRSP Control group than in the WKY group (p < 0.05) and significantly lower in the SHRSP EP group than in the SHRSP Control group (p < 0.05); Dpp4 expression was almost suppressed to the same level as that in the WKY group (Figure 6a). The expression patterns of Icam-1 and Agtr1 also showed changes similar to those of Dpp4 (Icam-1: WKY vs. SHRSP Control; p < 0.01, SHRSP Control vs. SHRSP EP; p < 0.01 Agtr1: Icam-1: WKY vs. SHRSP Control; p < 0.01, SHRSP Control vs. SHRSP EP; p < 0.01, Figure 6b, c).
3.4.4. mRNA Expression of Dpp4, Mac-1, Agtr1, and iNOS from the LRF-Rich Fraction
The expression levels of Dpp4, Mac-1, Agtr1, and iNOS in the leukocyte fraction were significantly lower in the SHRSP EP group than in the SHRSP Control group (p < 0.01; Figure 6d–g).
3.4.5. In Vitro Examination of Leukocyte NO Production
To investigate NO release from leukocytes into the culture medium, fluorescence intensity was measured using a multilabel counter with DAF-2, a membrane-impermeable NO indicator (Figure 7). To elucidate the mechanisms underlying leukocyte activation, we examined the effects of LPS, DPP-IV, and EP degradation products on leukocyte stimulation or inhibition. Thirty minutes after DAF-2 addition, fluorescence intensity in the culture medium of LPS- or DPP-IV-stimulated leukocytes was significantly higher than that of control leukocytes (p < 0.01). Although EP hydrolysate products had no effect on LPS-stimulated NO production, GOG, VP, cGP, and cPV inhibited NO production in DPP-IV-stimulated leukocytes.
4. Discussion
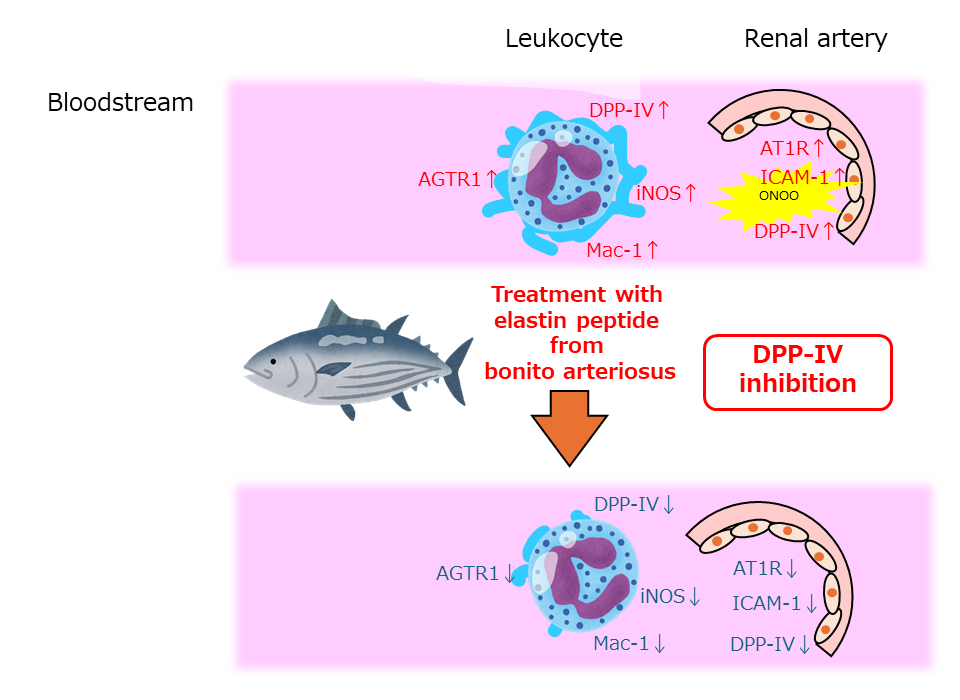
Bioactive peptides derived from dietary proteins have gained increasing attention as multifunctional food components capable of modulating metabolic and vascular functions. Although peptides are often limited by rapid gastrointestinal degradation, accumulating evidence indicates that certain food-derived peptides can reach the circulation and exert physiological effects [20]. In the present study, we demonstrated that EPs from bonito bulbus arteriosus undergo in vivo hydrolysis, enter the bloodstream, and contribute to vascular and metabolic regulation. These findings support the concept that food-derived peptides can act as physiologically relevant modulators rather than simple nutrient sources.
The DPP-IV inhibitory activity of EPs was modest, indicating that strong enzymatic inhibition alone is unlikely to account for the observed physiological effects. Among dietary proteins, DPP-IV inhibitory peptides have been reported [21]. The VP peptide identified in the present study exhibited stronger inhibitory activity than previously reported sequences [21]. Despite this limited inhibitory activity, EP administration improved glucose tolerance and increased GLP-1 and insulin levels, suggesting that mechanisms beyond direct DPP-IV inhibition contribute to the metabolic effects.
The insulinotropic and vasculoprotective actions of EPs are better interpreted as pleiotropic effects involving multiple pathways rather than a single dominant mechanism. The insulinotropic effect of EP intake may involve stabilization of endogenous GLP-1 and modulation of incretin signaling [22]. Additionally, the absence of significant weight reduction suggests that EPs act through pathways distinct from those of GLP-1 receptor agonists, warranting additional investigation into the relative contributions of each mechanism.
We confirmed that circulating EP-derived peptides We confirmed that circulating EP derived peptides were associated with suppression of DPP-IV–mediated leukocyte activation. Sequences containing hydroxyproline, such as GOG, are known to exhibit high resistance to proteolysis and prolonged persistence in vivo [23]. Tripeptides such as VPP exert antioxidant, antihypertensive, and gene-regulatory effects [24], whereas cyclic peptides such as cGP—one of the target peptides in the present study—exhibit high stability and membrane permeability, with potential roles as anti-inflammatory neuropeptides [25]. In our study, EP-derived peptides selectively suppressed NO production induced by DPP-IV stimulation but not by LPS, suggesting pathway-specific rather than broad-spectrum anti-inflammatory activity. This finding aligns with those of Martins et al., who reported that angiotensin II and DPP-IV activation contribute to vascular injury through shared inflammatory and oxidative stress pathways [26].
Collectively, these findings highlight that food-derived bioactive peptides function as multi-target modulators of metabolic and vascular homeostasis. EP administration in hypertensive rats enhanced GLP-1 and insulin secretion, improved glucose tolerance, and attenuated renal injury. These effects may be mediated through multiple mechanisms, including incretin regulation, suppression of inflammatory markers, and modulation of the RAS. The pathogenesis of endothelial injury in SHRSP likely involves inflammatory processes, such as leukocyte–endothelial adhesion, which are mediated by salt- or angiotensin II-induced hypertension [27,28].
The observed reduction in the expression of DPP-IV, ICAM-1, and AGTR1 in glomeruli, as well as the decreased expression of DPP-IV, Mac-1, and AGTR1 in leukocytes, substantiated the vasculoprotective and immunomodulatory effects of EPs, consistent with prior evidence showing that DPP-4 inhibition attenuates renal immune activation, inflammatory adhesion signaling, and RAAS-mediated vascular injury [29,30].
This study has some limitations. First, only one disease model (SHRSP) was used, and the treatment period was limited to 4 weeks at a relatively high dose (600 mg/kg). As SHRSP inherently exhibits glucose intolerance,14 it is difficult to fully exclude the contribution of improved glucose metabolism to the observed renoprotective effects. Using hypertensive models without metabolic abnormalities would provide clearer mechanistic insights. Second, longer-term studies evaluating renal function markers are needed to validate the vasculoprotective effects of EPs and determine whether lower, more practical intake levels are effective.
5. Conclusions
Although EPs from bonito bulbus arteriosus exhibited only modest DPP-IV inhibitory activity, they improved glucose metabolism and attenuated renal vascular injury in SHRSP, indicating pleiotropic metabolic and vascular effects. These results underscore the potential of food-derived peptides as bioactive compounds that contribute to metabolic and vascular regulation. Rather than acting through strong DPP IV inhibition or GLP 1 receptor agonism, EPs appear to modulate multiple interconnected physiological pathways as dietary bioactive compounds, including incretin signaling, inflammatory responses, and RAS related mechanisms.
Accordingly, EPs may hold promise as multi-target functional food ingredients for supporting metabolic and renal vascular health, rather than as therapeutic agents. Nevertheless, multiple EP-derived peptides were detected in the bloodstream, the stability and bioavailability of which require optimization to achieve consistent physiological effects. Future studies should focus on validating the efficacy of EPs in humans, elucidating the predominant mechanisms of action, and determining optimal intake levels for practical application.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Amino acid composition of bonito elastin peptide (residues/1000 residues).
Author Contributions
Conceptualization, K.T. and Y.N.; methodology, K.T., Y.N., and K.S.; validation, K.T. and K.S.; formal analysis, K.T. and Y.N.; investigation, K.T. and Y.N.; resources, E.S.; data curation, K.T. and Y.N.; writing—original draft preparation, Y.N.; writing—review and editing, K.T., K.S., E.S., and T.K.; visualization, K.T. and K.S.; supervision, K.S., T.K., and S.M.; project administration, K.S.; funding acquisition, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Grant-in-Aid for Scientific Research (C; General), grant numbers 22K11791 and 25K14926.
Institutional Review Board Statement
The animal study protocol was approved by the Kindai University Animal Experiment Committee (approval no. KAAG-2024-003 and date of approval Feb22/2024).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We sincerely thank Professor Emeritus Hiroyuki Ito (Kindai University) and Yasumitsu Akahoshi (Life Science Research Institute, Kindai University) for their technical support. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. During the preparation of this manuscript, the authors used Copilot, developed by OpenAI (Microsoft Co., Redmond, WA, USA), for the purposes of proofreading and enhancing the manuscript’s clarity. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AGTR1 | angiotensin receptor type 1 |
| cGP | cyclo(-Gly-Pro) |
| cPV | cyclo(L-Pro-L-Val) |
| DAF | diaminofluorescein |
| DPP-IV | dipeptidyl peptidase-IV |
| EP | elastin peptide |
| GLP-1 | glucagon-like peptide-1 |
| GOG | H-Gly-Hyp-Gly-OH |
| IC50 | half maximal inhibitory concentration |
| ICAM-1 | intercellular adhesion molecule 1 |
| iNOS | inducible NO synthase |
| LPS | lipopolysaccharide |
| LRF | leukocyte-rich fraction |
| NO | nitric oxide |
| PBS | phosphate-buffered saline |
| PG | H-Pro-Gly-OH |
| SHRSP | stroke-prone spontaneously hypertensive rats |
| VP | H-Val-Pro-OH |
| WKY/KPO | male Wistar-Kyoto rats |
References
- Hanafusa, N.; Abe, M.; Joki, N.; Hoshino, J.; Taniguchi, M.; Kikuchi, K.; Hasegawa, T.; Goto, S.; Ogawa, T.; Kanda, E.; Nakai, S.; Naganuma, T.; Miura, K.; Wada, A.; Takemoto, Y. Annual dialysis data report 2022, Japanese Society for Dialysis Therapy (JSDT) renal data registry. Ren. Replace. Ther. 2025, 11, 54. [Google Scholar] [CrossRef]
- Wakasugi, M.; Narita, I. Trends in the incidence of renal replacement therapy by type of primary kidney disease in Japan, 2006–2020. Nephrology 2023, 28, 119–129. [Google Scholar] [CrossRef]
- Costantino, V.V.; Gil Lorenzo, A.F.; Bocanegra, V.; Vallés, P.G. Molecular mechanisms of hypertensive nephropathy: renoprotective effect of losartan through Hsp70. Cells 2021, 10, 3146. [Google Scholar] [CrossRef] [PubMed]
- Durinx, C.; Lambeir, A.M.; Bosmans, E.; Falmagne, J.B.; Berghmans, R.; Haemers, A.; Scharpé, S.; De Meester, I. Molecular characterization of dipeptidyl peptidase activity in serum: soluble CD26/dipeptidyl peptidase IV is responsible for the release of X-Pro dipeptides. Eur. J. Biochem. 2000, 267, 5608–5613. [Google Scholar] [CrossRef] [PubMed]
- Elmansi, A.M.; Awad, M.E.; Eisa, N.H.; Kondrikov, D.; Hussein, K.A.; Aguilar-Pérez, A.; Herberg, S.; Periyasamy-Thandavan, S.; Fulzele, S.; Hamrick, M.W.; McGee-Lawrence, M.E.; Isales, C.M.; Volkman, B.F.; Hill, W.D. What doesn’t kill you makes you stranger: dipeptidyl peptidase-4 (CD26) proteolysis differentially modulates the activity of many peptide hormones and cytokines generating novel cryptic bioactive ligands. Pharmacol. Ther. 2019, 198, 90–108. [Google Scholar] [CrossRef]
- Klemann, C.; Wagner, L.; Stephan, M.; von Hörsten, S. Cut to the chase: a review of CD26/dipeptidyl peptidase-4’s (DPP4) entanglement in the immune system. Clin. Exp. Immunol. 2016, 185, 1–21. [Google Scholar] [CrossRef]
- Campbell, J.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of incretin hormone action. Cell. Metab. 2013, 17, 819–837. [Google Scholar] [CrossRef]
- Deacon, C.F. A review of dipeptidyl peptidase-4 inhibitors. Hot topics from randomized controlled trials. Diabetes Obes. Metab. 2018, 20, 34–46. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Q.; Zhong, J.; Liu, C.; Zheng, B.; Gong, Q. DPP-4 inhibitors as potential candidates for antihypertensive therapy: improving vascular inflammation and assisting the action of traditional antihypertensive drugs. Front. Immunol. 2019, 10, 1050. [Google Scholar] [CrossRef]
- Jones, L.; Jones, A.M. Suspected adverse drug reactions of the type 2 antidiabetic drug class dipeptidyl-peptidase IV inhibitors (DPP4i): can polypharmacology help explain? Pharmacol. Res. Perspect. 2022, 10, e01029. [Google Scholar] [CrossRef]
- Takemori, K.; Yamamoto, E.; Ito, H.; Kometani, T. Prophylactic effects of elastin peptide derived from the bulbus arteriosus of fish on vascular dysfunction in spontaneously hypertensive rats. Life Sci. 2015, 120, 48–53. [Google Scholar] [CrossRef]
- Takemori, K.; Yamamoto, E.; Chikugo, T.; Shiratsuchi, E.; Kometani, T. DPP-IV inhibition by solubilized elastin peptides from yellowtail bulbus arteriosus suppresses ultraviolet-B-induced photoaging in hairless mice. Nutraceuticals 2024, 4, 683–694. [Google Scholar] [CrossRef]
- Shiratsuchi, E.; Nakaba, M.; Shigemura, Y.; Yamada, M.; Sato, K. Fish-elastin hydrolysate: development and impact on the skin and blood vessels. In Marine Proteins and Peptides: Biological Activities and Applications; Kim, S.K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 467–486. [Google Scholar]
- Okamoto, K.; Yamori, Y.; Nagaoka, A. Establishment of the stroke-prone spontaneously hypertensive rat (SHR). Circ. Res. 1974, 34–35, 143–153. [Google Scholar]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; Garner, P.; Holgate, S.T.; Howells, D.W.; Karp, N.A.; Lazic, S.E.; Lidster, K.; MacCallum, C.J.; Macleod, M.; Pearl, E.J.; Petersen, O.H.; Rawle, F.; Reynolds, P.; Rooney, K.; Sena, E.S.; Silberberg, S.D.; Steckler, T.; Würbel, H. The ARRIVE guidelines 2.0: updated guidelines for reporting animal research. Br. J. Pharmacol. 2020, 177, 3617–3624. [Google Scholar] [CrossRef]
- Nagao, A.; Nakamoto, Y.; Miyauchi, S.; Sato, K. Presence of modified peptides with high bioavailability and angiotensin-converting enzyme inhibitory activity in Japanese fermented soybean paste (Miso). J. Agric. Food Chem. 2024, 72, 18942–18956. [Google Scholar] [CrossRef]
- Yamamoto, T. Isolation and enrichment of glomeruli using sieving techniques. In Renal and Urinary Proteomics: Methods and Protocols; Thongboonkerd, V., Ed.; John Wiley & Sons: Chichester, 2009; pp. 1–7. [Google Scholar]
- Takemori, K.; Ito, H.; Suzuki, T. Effects of the AT1 receptor antagonist on adhesion molecule expression in leukocytes and brain microvessels of stroke-prone spontaneously hypertensive rats. Am. J. Hypertens. 2000, 13, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Nakatsubo, N.; Kikuchi, K.; Kawahara, S.; Kirino, Y.; Nagoshi, H.; Hirata, Y.; Nagano, T. Detection and imaging of nitric oxide with novel fluorescent indicators: diaminofluoresceins. Anal. Chem. 1998, 70, 2446–2453. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Sato, M.; Katakura, Y.; Sugawara, Y.; Shigemura, Y. Increase in blood-transferable linear and cyclic dipeptides in human plasma following ingestion of elastin hydrolysate. Food Hydrocoll. Health 2024, 6, 100188. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef]
- Trivedi, K.; Dolinsky, V. Role of endogenous incretin hormones, GLP-1 and GIP, in cardiovascular physiology. Can. J. Physiol. Pharmacol. 2026, 104, 1–13. [Google Scholar] [CrossRef]
- Taga, Y.; Iwasaki, Y.; Tometsuka, C.; Funato, N.; Shigemura, Y.; Kusubata, M.; Mizuno, K. Identification of a highly stable bioactive 3-hydroxyproline-containing tripeptide in human blood after collagen hydrolysate ingestion. NPJ Sci. Food 2022, 6, 29. [Google Scholar] [CrossRef]
- Pérez-Vielma, N.M.; Gómez-López, M.; Maldonado, J.; Correa-Basurto, J.; Martínez-Godínez, M.L.Á.; Miliar-García, Á. Recognition of the interaction between the bioactive peptide Val–Pro–Pro and the minimal promoter region of genes SOD and CAT using QCM-D and docking studies. Anal. Methods 2023, 15, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Melo, I.S.; Candeia-Medeiros, N.; Ferro, J.N.S.; Cavalcante-Araújo, P.M.; Oliveira, T.L.; Santos, C.E.A.; Cardoso-Sousa, L.; Aguiar, E.M.G.; Wutke Oliveira, S.; Castro, O.W.; Alves-Balvedi, R.P.; Rodrigues, L.P.; Hickmann, J.M.; Alves, D.A.; Santos, I.A.; Jardim, A.C.G.; Siqueira, W.L.; Pipi, A.R.F.; Goulart, L.R.; Barreto, E.O.; Sabino-Silva, R. Restoration of Cyclo-Gly–Pro-induced salivary hyposecretion and submandibular composition by naloxone in mice. PLoS ONE 2020, 15, e0229761. [Google Scholar] [CrossRef]
- Martins, F.L.; Ribeiro-Silva, J.C.; Nistala, R.; Girardi, A.C.C. Bidirectional relation between dipeptidyl peptidase 4 and angiotensin II type I receptor signaling. Am. J. Physiol. Cell. Physiol. 2024, 326, C1203–C1211. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Nakagawa, S.; Wu, Y.; Kawabata, Y.; Numabe, A.; Yanagi, Y.; Tamaki, Y.; Uehara, Y.; Araie, M. A high-salt diet enhances leukocyte adhesion in association with kidney injury in young Dahl salt-sensitive rats. Hypertens. Res. 2017, 40, 912–920. [Google Scholar] [CrossRef]
- Takemori, K.; Inoue, T.; Ito, H. Possible role of nitric oxide generated by leukocytes in the pathogenesis of hypertensive cerebral edema in stroke-prone spontaneously hypertensive rats. Brain Res. 2011, 1417, 137–145. [Google Scholar] [CrossRef]
- Nistala, R.; Meuth, A.I.; Smith, C.; et al. DPP4 inhibition mitigates ANG II-mediated kidney immune activation and injury in male mice. Am. J. Physiol. Ren. Physiol. 2021, 320, F505–F517. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sen, U. More than just an enzyme: Dipeptidyl peptidase-4 (DPP-4) and its association with diabetic kidney remodelling. Pharmacol. Res. 2019, 147, 104391. [Google Scholar] [CrossRef]
Figure 1.
Plasma concentrations of elastin peptide degradation products. (a) H-Gly-Hyp-Gly-OH (GOG); (b) H-Val-Pro-OH (VP); (c) H-Pro-Gly-OH (PG); (d) cyclo(-Gly-Pro) (cGP); (e) cyclo(L-Pro-L-Val) (cPV). Mean ± standard deviation (SD), n = 3.
Figure 1.
Plasma concentrations of elastin peptide degradation products. (a) H-Gly-Hyp-Gly-OH (GOG); (b) H-Val-Pro-OH (VP); (c) H-Pro-Gly-OH (PG); (d) cyclo(-Gly-Pro) (cGP); (e) cyclo(L-Pro-L-Val) (cPV). Mean ± standard deviation (SD), n = 3.

Figure 2.
Comparison of blood glucose and plasma insulin levels in male Wistar-Kyoto rats (WKY) and stroke-prone spontaneously hypertensive rats (SHRSP). (a) Plasma insulin; (b) blood glucose. ●: WKY; 〇: SHRSP. Mean ± SD, n = 6, Student’s t-test, *p < 0.05.
Figure 2.
Comparison of blood glucose and plasma insulin levels in male Wistar-Kyoto rats (WKY) and stroke-prone spontaneously hypertensive rats (SHRSP). (a) Plasma insulin; (b) blood glucose. ●: WKY; 〇: SHRSP. Mean ± SD, n = 6, Student’s t-test, *p < 0.05.

Figure 3.
Effect of simultaneous administration of glucose and elastin peptides on glucose metabolism. (a) Glucagon-like peptide-1 (GLP-1); (b) plasma insulin; (c) blood glucose. ●: SHRSP Control; 〇: SHRSP elastin peptides (EPs). Mean ± SD, n = 6, Student’s t-test, *p < 0.05.
Figure 3.
Effect of simultaneous administration of glucose and elastin peptides on glucose metabolism. (a) Glucagon-like peptide-1 (GLP-1); (b) plasma insulin; (c) blood glucose. ●: SHRSP Control; 〇: SHRSP elastin peptides (EPs). Mean ± SD, n = 6, Student’s t-test, *p < 0.05.

Figure 4.
Chronological changes in blood pressure and body weight. (a) Blood pressure, (b) body weight. ●: WKY; 〇: SHRSP Control; : SHRSP EP. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s honest significant difference (HSD) test, *p < 0.05 WKY vs. SHRSP Control, +p < 0.05 WKY vs. SHRSP EP.
: SHRSP EP. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s honest significant difference (HSD) test, *p < 0.05 WKY vs. SHRSP Control, +p < 0.05 WKY vs. SHRSP EP.
Figure 4.
Chronological changes in blood pressure and body weight. (a) Blood pressure, (b) body weight. ●: WKY; 〇: SHRSP Control; : SHRSP EP. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s honest significant difference (HSD) test, *p < 0.05 WKY vs. SHRSP Control, +p < 0.05 WKY vs. SHRSP EP.
: SHRSP EP. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s honest significant difference (HSD) test, *p < 0.05 WKY vs. SHRSP Control, +p < 0.05 WKY vs. SHRSP EP.
Figure 5.
Morphological observations of the kidneys. Macroscopic findings: (a) WKY; (b) SHRSP Control; (c) SHRSP EP. Histological observations of the kidneys: (d) WKY; (e) SHRSP Control; (f) SHRSP EP. Scale bar: 250 μm.
Figure 5.
Morphological observations of the kidneys. Macroscopic findings: (a) WKY; (b) SHRSP Control; (c) SHRSP EP. Histological observations of the kidneys: (d) WKY; (e) SHRSP Control; (f) SHRSP EP. Scale bar: 250 μm.

Figure 6.
mRNA expression in the renal glomerular fraction and leukocyte-rich fraction (LRF). (a–c): Renal glomerular fraction, (d–g): LRF. (a) Dipeptidyl-peptidase-IV (Dpp4); (b) intercellular adhesion molecule 1 (Icam-1); (c) angiotensin receptor type 1 (Agtr1); (d) Dpp4; (e) Mac-1; (f) Agtr1; (g) iNOS. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s HSD, *p < 0.05, **p < 0.01.
Figure 6.
mRNA expression in the renal glomerular fraction and leukocyte-rich fraction (LRF). (a–c): Renal glomerular fraction, (d–g): LRF. (a) Dipeptidyl-peptidase-IV (Dpp4); (b) intercellular adhesion molecule 1 (Icam-1); (c) angiotensin receptor type 1 (Agtr1); (d) Dpp4; (e) Mac-1; (f) Agtr1; (g) iNOS. Mean ± SD, WKY n = 9, SHRSP Control n = 9, SHRSP EP n = 10, Tukey’s HSD, *p < 0.05, **p < 0.01.

Figure 7.
Nitric oxide (NO) production in the LRF. Mean ± SD, n = 6, Tukey’s HSD, *p < 0.05, **p < 0.01. Control (Cont); H-Gly-Hyp-Gly-OH (GOG); lipopolysaccharide (LPS); H-Val-Pro-OH (VP); cyclo(-Gly-Pro) (cGP); cyclo(L-Pro-L-Val) (cPV).
Figure 7.
Nitric oxide (NO) production in the LRF. Mean ± SD, n = 6, Tukey’s HSD, *p < 0.05, **p < 0.01. Control (Cont); H-Gly-Hyp-Gly-OH (GOG); lipopolysaccharide (LPS); H-Val-Pro-OH (VP); cyclo(-Gly-Pro) (cGP); cyclo(L-Pro-L-Val) (cPV).


Table 1.
Comparison of organ weights.
| Organ | WKY | SHRSP Control | SHRSP EP |
| Brain | 1.923±0.041 | 2.048±0.112 | 1.981±0.132 |
| Heart | 1.276±0.042 | 1.351±0.071 | 1.233±0.218 |
| Kidney (right) | 1.410±0.089 | 1.091±0.117** | 1.013±0.155** |
| Kidney (left) | 1.351±0.122 | 1.118±0.079** | 1.029±0.171** |
Mean ± standard deviation (SD), Dunnett’s test; **p < 0.01 vs. WKY. Elastin peptides (EPs); stroke-prone spontaneously hypertensive rats (SHRSP); male Wistar-Kyoto rats (WKY/KPO). Values are shown in grams.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



