Submitted:
13 April 2026
Posted:
15 April 2026
You are already at the latest version
Abstract
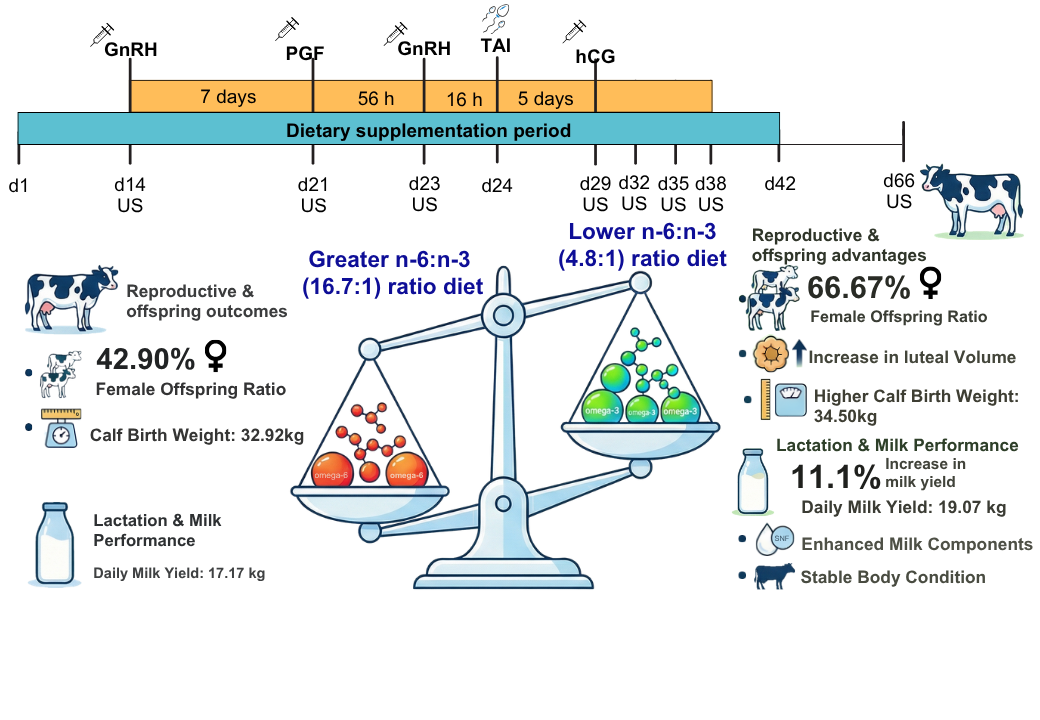
The objectives were to evaluate the effects of different dietary omega-6 to omega-3 (n-6:n-3) fatty acid ratios during timed AI on fertility and offspring sex ratio in lactating dairy cows. Holstein cows (n=60) averaging 109 ± 10 days in milk (DIM) were randomly assigned to be fed either a greater n-6:n-3 diet or a lower n-6:n-3 diet for 42 days. All cows were synchronized using a modified Ovsynch protocol. Although synchronized ovulation rates and pregnancy outcomes were not significantly affected by treatment, cows fed the lower n-6:n-3 ratio exhibited significantly greater corpus luteum (CL) volume and cross-sectional area on days 11 and 14 post-TAI, indicating enhanced luteal development. Pregnancy rates following the first AI and cumulative AI, as well as the number of services per conception, did not differ between treatments. However, maternal dietary fatty acid composition influenced offspring characteristics. Cows fed the lower n-6:n-3 ratio tended to produce a greater proportion of female offspring (66.67% vs. 42.90%; p = 0.09). In addition, calves born to cows receiving the lower n-6:n-3 ratio had greater birth weight than those from cows fed the greater n-6:n-3 ratio diet. Additionally, the lower n-6:n-3 diet tended to increase milk yield and significantly increased lactose and solids-not-fat yields. In conclusion, a lower dietary n-6:n-3 ratio during a TAI program improves fertility potential by enhancing luteal development and may favor female offspring production while supporting lactation performance.
Keywords:
polyunsaturated fatty acids
; omega-3
; omega-6
; fertility
; dairy cows
; sex ratio
1. Introduction
Reproductive efficiency is a central determinant of sustainability and profitability in dairy production systems, as it directly influences herd replacement rate, genetic progress, and lifetime productivity of dairy cows. Infertility remains a leading cause of involuntary culling in dairy herds worldwide, with rates often exceeding 35% of total herd removals in many production systems [1]. Impaired reproductive performance is commonly associated with postpartum disorders, such as retained placenta and uterine infections, as well as prolonged negative energy balance (NEB) during early lactation. NEB disrupts endocrine homeostasis by reducing circulating concentrations of luteinizing hormone (LH), glucose, insulin, and insulin-like growth factor I (IGF-I), while increasing non-esterified fatty acids (NEFA), thereby impairing ovarian steroidogenesis and compromising fertility [2]. These metabolic and physiological disturbances contribute to early embryonic loss, particularly within the first 28 days of pregnancy, resulting in reduced conception rates and extended calving intervals [3,4]. Beyond its direct impact on fertility, reproductive failure also indirectly constrains milk production and limits opportunities for herd expansion through reduced availability of female replacements.
Nutritional management plays a pivotal role in regulating reproductive performance, particularly during early and mid-lactation when cows experience high metabolic demands and are susceptible to NEB. Among nutritional factors, dietary fatty acids (FAs) have gained increasing attention due to their involvement in metabolic regulation, hormone synthesis, and reproductive physiology. Although energy balance is widely recognized as a primary determinant of reproductive outcomes [5,6,7,8], accumulating evidence indicates that fat supplementation can enhance reproductive performance independently of increased energy intake [9,10,11]. These findings suggest that specific FAs, beyond their caloric contribution, may directly modulate reproductive biology in cattle. Fatty acids serve not only as concentrated energy sources but also as key regulators of cellular function [12], metabolic pathways [13], immune system [14], and hormone responsiveness [15,16]. Structurally, FAs are classified according to the number of double bonds into saturated, monounsaturated, and polyunsaturated fatty acids (PUFAs) [17]. PUFAs are further categorized into n-6 and n-3 families based on the position of the first double bond from the methyl (omega) end [17,18]. PUFAs, particularly omega-6 and omega-3, exert distinct and often opposing effects on reproductive processes through their influence on steroidogenesis and prostaglandin metabolism [19]. Omega-6 PUFAs, such as linoleic acid (LA; C18:2 n-6) and its derivative arachidonic acid (C20:4n-6), serve as precursors for prostaglandin F₂α (PGF₂α), a key mediator of luteolysis and estrous cycle regulation. In contrast, omega-3 PUFAs, including α-linolenic acid (ALA; C18:3n-3) and its longer-chain derivatives eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), have been shown to reduce PGF₂α synthesis, prolong corpus luteum lifespan, enhance progesterone production, and improve embryo survival [10,20,21]. Mechanistically, LA and ALA compete for the same metabolic pathways and cyclooxygenase (COX) enzymes. Increasing dietary ALA may reduce the availability of LA for conversion into PGF₂α, thereby lowering PGF₂α concentrations. Reduced PGF₂α has been associated with elevated progesterone levels and delayed CL regression [22,23], both of which are essential for the establishment and maintenance of early pregnancy. Because pregnancy loss in dairy cattle has been linked to CL regression and embryonic failure [24], modulation of the dietary n-6:n-3 ratio may represent a practical strategy to enhance reproductive success.
In practical feeding systems, particularly in tropical dairy production systems such as those in Thailand, concentrate feeds are predominantly formulated using locally available ingredients, including soybean meal, rice bran, and palm oil by-products. These feedstuffs are inherently high in n-6:n-3 ratio, resulting in diets characterized by a high n-6:n-3 ratio. Similarly, concentrate-based rations frequently exhibit an imbalanced n-6:n-3 ratio due to the high inclusion of n-6-rich ingredients and the limited availability of omega-3 sources [21]. Given that omega-6 and omega-3 PUFAs are associated with pro-inflammatory and anti-inflammatory pathways, respectively [25], such imbalance may favor the synthesis of pro-inflammatory mediators and potentially compromise reproductive performance. Conversely, a more balanced n-6:n-3 ratio has been associated with improved ovarian function, altered follicular dynamics, enhanced oocyte and embryo quality, and reduced pregnancy loss [21,26]. Furthermore, supplementation with n-3 and n-6 PUFAs during the period of maternal recognition of pregnancy has been reported to optimize uterine–conceptus interactions, enhance early embryo development [27] and elongation [28], first-service conception in primiparous and multiparous dairy cows [29], reduce embryonic mortality [26,30,31,32].
Beyond fertility traits, the sex ratio of offspring has important implications for herd sustainability, as female calves are essential for herd replacement and long-term productivity. Under natural conditions, the calf sex ratio remains close to 1:1 [33]. Although sexed semen technology can increase the proportion of female offspring, it is relatively costly and often associated with reduced conception rates [34]. Emerging evidence suggests that omega-3 fatty acid supplementation during the breeding period may influence offspring sex ratio in favor of female calves [20]. Although previous studies have demonstrated the effects of PUFAs on reproductive performance in dairy cows, limited information is available regarding the effects of different dietary n-6:n-3 fatty acid ratios under practical feeding conditions. Moreover, only a few studies have investigated the influence of PUFAs supplementation on offspring sex ratio in dairy cattle.
Therefore, the objective of this study was to evaluate the effects of dietary supplementation with differing n-6:n-3 ratios on reproductive performance, pregnancy rate, and offspring sex ratio in lactating dairy cows. We hypothesized that reducing the dietary n-6:n-3 ratio through targeted omega-3 supplementation would improve reproductive performance and increase the proportion of female offspring. The findings of this study aim to provide practical nutritional insights for optimizing reproductive efficiency and herd sustainability in dairy production systems that rely on feed ingredients with high n-6:n-3 ratios.
2. Materials and Methods
2.1. Animal Ethics and Experimental Design
All procedures were conducted in accordance with the guidelines approved by the Institutional Animal Care and Use Committee of Khon Kaen University, based on the Ethics of Animal Experimentation of the National Research Council of Thailand (IACUC-KKU-141/67). The study was carried out on dairy farms located in Ban-Kho Subdistrict, Mueang District, Khon Kaen Province, Thailand.
The study was conducted on four smallholder dairy farms that are members of the Khon Kaen Dairy Cooperative, located within a 5 km radius in Khon Kaen Province, Thailand. All farms operated under comparable housing, feeding, and reproductive management systems in accordance with cooperative training programs. Cows were housed in individual pens (3 × 4 m) in a free-stall barns with open-sided, natural ventilation, and concrete floors. To ensure animal comfort, each pen was bedded with sand and provided with adequate space for free movement. All cows were offered the same forage sources across all farms, consisting of corn silage and rice straw (57% moisture) provided ad libitum. Free access to clean drinking water and mineralized salt blocks was ensured within each pen. Milking was performed twice daily in herringbone parlors at 06:00 and 16:00 h. Concentrate was individually offered during milking based on milk yield at a ratio of 1 kg concentrate per 2 kg of milk produced. Routine veterinary services were provided through the cooperative, and herd health records confirmed a low prevalence of mastitis, metritis, and metabolic disorders during the study period. Prior to the experiment, participating farmers received training in feeding and milking management. Health monitoring was conducted throughout the study to ensure uniform management across locations. These management practices minimized potential variation due to environmental or management differences, allowing the animals to be considered a single experimental population.
A total of 60 lactating Holstein cows (109 ± 10 DIM) were enrolled in the trial. This DIM range corresponds to the period after the voluntary waiting period (VWP), when cows are eligible for breeding and metabolic status is relatively stable, making it suitable for evaluating reproductive responses. Cows were blocked by lactation number (1–2 vs. 3–4) and randomly assigned in a randomized complete block design (RCBD) to receive either a greater n-6:n-3 diet (n = 30) or a lower n-6:n-3 diet (n = 30) for 42 days (6 weeks), covering the breeding period. However, one cow from the lower n-6:n-3 diet group was excluded due to health issues unrelated to the treatment, leaving 29 cows in the n-3 group for final analysis. Diets were formulated to meet NRC recommendations [9]. The lower n-6:n-3 diet was formulated by partially replacing conventional fat sources with a rumen-protected (bypass) fat rich in n-3 fatty acids, specifically the commercial product HI-FLAX (NOREL, S.A., Spain), distributed by Natural Nutrients Co., Ltd. In contrast, the control diet reflected typical on-farm feeding practices and was based on locally available feed ingredients high in n-6:n-3 ratio.
Fatty acid compositions of the experimental diets were analyzed by gas chromatography at the Central Laboratory (Thailand) Co., Ltd., Bangkok. The greater n-6:n-3 diet contained 7.99 g/kg n-6 and 0.48 g/kg n-3 fatty acids (16.7:1 n-6:n-3 ratio), whereas the lower n-6:n-3 diet contained 6.45 g/kg n-6 and 1.35 g/kg n-3 fatty acids (4.8:1 n-6:n-3 ratio). The chemical compositions of the experimental diets are presented in Table 1.
2.2. Reproductive Management
The reproductive synchronization and monitoring schedule are illustrated in Figure 1. On approximately day 14 of the experimental period, all cows commenced a modified Ovsynch protocol. This protocol began with an intramuscular injection of 10 µg of GnRH agonist (Buserelin; Receptal®, Intervet, New Zealand). Seven days later (day 21), luteolysis was induced with 25 mg of PGF2α (Lutalyse®, Pfizer, USA). A second GnRH dose (10 µg) was administered 56 hours after the PGF2α treatment (day 23), followed by timed artificial insemination (TAI) 16 hours thereafter (day 24). To support the subsequent luteal phase and potential pregnancy, all cows received 1,500 IU of hCG (Choluron®, Pfizer, USA) on day 29 (day 5 post-TAI) [10].
2.3. Data and Sample Collection
Feed intake was measured daily using the weigh-back method by recording the feed offered and refusals to calculate dry matter intake (DMI). Body weight (BW) and body condition score (BCS) were recorded weekly throughout the 42 days experimental period. BW was estimated using a heart-girth measuring tape after the morning milking, while BCS was assessed by a trained evaluator using a 5-point scale with 0.25-unit increments [35]. For a comprehensive metabolic assessment, DMI was also expressed as a percentage of BW and relative to metabolic body weight (g/kg BW0.75). Individual milk yield was recorded dairy by weighing the milk collected during both morning and afternoon milking. Milk composition, including fat, protein, lactose, somatic cell counts (SCC), was analyzed biweekly at the Veterinary Research and Development Center (Upper Northeastern Region), Khon kaen, Thailand, using MilkoScan 7 RM (Foss Analytical A/S, Hillerod, Denmark). 3.5% fat-corrected milk (FCM) was calculated as 0.4324 x milk yield (Kg) + 16.218 x fat yield (Kg). Energy-corrected milk (ECM) was calculated using the equation: ECM (kg/day) = Milk yield × (0.124 × fat% + 0.073 × protein% + 0.256), as recommended by NRC [9].
Ovarian activity was assessed using real-time B-mode ultrasonography with a 7.4 MHz transrectal probe (DUS 60 Vet, Edan, USA). Ovulation was confirmed by the disappearance of the dominant follicle and the subsequent formation of the CL following GnRH administration. Reproductive parameters, including ovulation, follicle size, luteolysis, and CL development, were evaluated on days 14, 21, and 23 of the experimental periods, and days 5, 8, 11, and 14 post-TAI (days 29, 32, 35, and 38 of the study). Conception was diagnosed on day 42 after TAI (day 66 of the study) via transrectal ultrasonography. Offspring sex and birth weight were recorded at parturition.
2.4. Statistical Analysis
For variables with repeated measurements, including DMI, milk yield, and milk composition, data were analyzed using the MIXED procedure of SAS software (version 9.0; SAS Institute Inc., Cary, NC, USA) with the individual cow as the experimental unit. Reproductive parameters such as conception rate and offspring sex ratio were analyzed using the GLIMMIX procedure for logistic regression and chi-square tests. For all continuous data, differences among treatment means were compared using the least significant difference (LSD) test as described by Steel et al. [36]. Results are expressed as means and the standard error of the mean (SEM). Statistical significance was declared at p < 0.05, while a tendency was considered at 0.05 ≥ p < 0.10.
3. Results
3.1. Feed intake, Body Weight, and Body Condition Score
The effects of different dietary n-6 to n-3 fatty acid ratio during timed AI on DMI, BW, and BCS are presented in Table 2 and Figure 2. Overall, the average DMI of concentrate, roughage, and total intake did not differ significantly between greater n-6:n-3 and lower n-6:n-3 groups when expressed as absolute intake (kg/d), percentage of BW (% BW), or relative to metabolic body weight (g/kg BW0.75) (p ≥ 0.13; Table 2). Specifically, the total DMI for the greater n-6:n-3 and lower n-6:n-3 groups was 13.34 ± 0.32 and 14.02 ± 0.32 kg/d, respectively (p = 0.20).
However, longitudinal analysis of weekly total DMI revealed a tendency for a treatment effect (p = 0.09; Figure 2), with cows in the lower n-6:n-3 group consistently maintaining numerically higher intake levels throughout the 6-week experimental period compared to the n-6 group. A significant effect of week was observed for total DMI (p < 0.0001), reflecting a progressive increase in feed consumption over time in both groups. However, no significant treatment x week interaction was detected (p > 0.05), indicating that the pattern of DMI increase over time was similar across both dietary treatments.
Regarding body reserves, the mean BW remained comparable between the greater n-6:n-3 and lower n-6:n-3 groups, recorded at 539.62 and 541.13 ± 6.85 kg, respectively (p = 0.99). Similarly, dietary treatments did not significantly influence the initial BCS (2.50 ± 0.07 for both groups; p = 0.77), the final BCS (2.65 and 2.71 ± 0.07; p = 0.51), or the net BCS change during the study (0.16 and 0.22 ± 0.03; p = 0.23). These results suggest that the source of PUFAs supplementation provided comparable nutrient intake and maintained a stable energy status across both experimental groups.
3.2. Milk Yield and Composition
The effects of different dietary n-6 to n-3 fatty acid ratio on milk production and its composition are summarized in Table 3 and Figure 3. Over the entire experimental period, cows supplemented with lower n-6:n-3 group showed a tendency toward higher daily milk yield compared to the greater n-6:n-3 group (19.07 vs. 17.17 kg/d; p = 0.08; Table 3). Analysis of weekly production data revealed a significant treatment × week interaction (p = 0.03; Figure 3), with the lower n-6:n-3 group maintaining significantly higher production during the first three weeks of the study (p < 0.05). However, no significant differences were observed between treatments for 3.5% FCM (p = 0.15), ECM (p = 0.13), or feed efficiency (ECM/DMI; p = 0.41).
Regarding milk composition, no significant differences were observed between the greater n-6:n-3 group and lower n-6:n-3 group for the percentages of fat, protein, lactose, solids-not-fat (SNF), and total solids (p ≥ 0.48). Somatic cell counts (SCC) were also similar between groups (p = 0.55), suggesting that n-6:n-3 ratios did not adversely affect mammary gland health.
Notably, the total daily yields of several milk components were significantly improved by lower n-6:n-3 group. Cows in lower n-6:n-3 group had a significantly higher lactose yield (0.94 vs. 0.80 kg/d; p = 0.03) and SNF yield (1.67 vs. 1.45 kg/d; p = 0.05) compared to the greater n-6:n-3 group. Additionally, lower n-6:n-3 group tended to increase protein yield (0.60 vs. 0.53 kg/d; p = 0.10), while the difference in total solids yield did not reach statistical significance (p = 0.14).
3.3. Ovarian Dynamics, Reproductive Performance, and Offspring Characteristics
The effects of different dietary n-6 to n-3 fatty acid ratio on ovarian functions, fertility, and offspring traits are summarized in Table 4 and Table 5. Synchronized ovulation rates following the first and second GnRH injections, as well as luteolysis rates and pre-ovulatory follicle diameter, did not differ significantly between the greater n-6:n-3 ratio and Lower (n-6:n-3 ratio 4.8:1) groups (p > 0.05; Table 4). Regarding CL development, the number of CL at all measured time points (days 5, 8, 11, and 14 post-TAI) was comparable between treatments (p > 0.05). Similarly, CL diameter did not show significant differences between groups, although a tendency for a larger diameter was observed in the lower n-6:n-3 ratio on day 11 (p = 0.05) and day 14 (p = 0.10) post-TAI. In contrast, dietary treatment significantly influenced CL morphometry during the late luteal phase. While no differences were observed in CL cross-sectional area and volume on days 5 and 8 post-TAI (p > 0.05), cows in the lower n-6:n-3 ratio group exhibited significantly higher CL cross-sectional areas on day 11 (461.94 vs. 380.10 mm2; p = 0.04) and day 14 (560.32 vs. 474.14 mm2; p = 0.03) compared to the greater dietary n-6:n-3 ratio group. Consistently, CL volume was significantly greater in the lower n-6:n-3 ratio group on day 11 (7,774.80 vs. 5,864.44 mm3; p = 0.03) and day 14 (10,395.00 vs. 7,978.92 mm3; p = 0.007) post-TAI.
In terms of reproductive success, pregnancy rates for both the first AI and cumulative AI, as well as the number of services per conception, were not significantly influenced by the source of PUFAs (p ≥ 0.37; Table 5). Notably, dietary supplementation differentially n-6:n-3 ratio influenced offspring characteristics. The proportion of female offspring in the lower n-6:n-3 ratio group was 66.67 % (18/27), whereas it was 42.90 % (12/28) in the greater n-6:n-3 ratio group, representing a tendency toward a female-skewed sex ratio (p = 0.09). Furthermore, supplementation with lower n-6:n-3 ratio significantly increased the birth weight of offspring compared to the greater n-6:n-3 ratio group (34.50 vs. 32.92 kg; p = 0.02).
4. Discussion
The findings of the present study demonstrate that reducing the dietary n-6:n-3 fatty acid ratio to 4.8:1 through rumen-protected n-3 supplementation significantly enhances early lactation milk production and modifies ovarian dynamics in dairy cows. These improvements, coupled with a notable shift in the offspring sex ratio toward females, suggest that strategic PUFAs nutrition is a potent modulator of both metabolic and reproductive functions in tropical dairy production systems.
4.1. Feed Intake and Lactation Performance
Our results indicate that the lower n-6:n-3 ratio dietary did not adversely affect total DMI. This stability in feed consumption suggests that the use of rumen-protected (bypass) fat effectively mitigated the typical inhibitory effects of unsaturated fatty acids on rumen microbial fermentation and fiber digestibility, which are often reported as limiting factors in fat supplementation strategies [10].
While some studies suggest that an increase in dietary fat, particularly unsaturated fatty acids, may cause a decrease in DMI [37,38], our findings align with Huang et al. [39], who reported that supplementation with ground flaxseed (GF) or whole flaxseed (WF) did not affect DMI. This observation is further supported by a meta-analysis by Leduc et al. [40] and various studies (Do Prado et al. [41]; Isenberg et al. [42]), suggesting that the amount and form of supplementation are key factors; in our study, the inclusion level was likely not sufficient to trigger a reduction in DMI.
Interestingly, the longitudinal analysis of weekly total DMI revealed a tendency for a treatment effect, with cows in the lower n-6:n-3 ratio group consistently maintaining numerically higher intake levels. This trend is consistent with Sun et al. [43], who found that high n-3 PUFAs supplementation significantly increased DMI during the transition period. Such an increase in DMI may be associated with various internal factors, including optimized fermentation parameters, bacterial abundance, and specific gene expression [43].
Moreover, previous reports by Petit et al. [44] and Zachut et al. [45] also noted that cows fed diets rich in unsaturated fatty acids (FA) showed higher DMI and milk yield compared to those fed saturated FA. However, the effect of n-3 PUFA on intake remains variable across the literature; for instance, Chilliard et al. [46] reported a decrease in DMI with high levels of extruded flaxseed in late lactation. These inconsistencies likely stem from differences in supplement concentration, the lactation stage of the cows, or interactions between dietary supplements and other components [43,46].
Regarding lactation performance, the lower n-6:n-3 ratio group in the present study exhibited an 11.1% increase in average milk yield compared to the greater n-6:n-3 ratio group (19.07 vs. 17.17 kg/d). This production boost corroborates the dramatic increase in milk production reported by Sun et al. [43] and findings by Moallem et al. [47]. While Huang et al. [39] observed no significant effect on milk yield in their specific dietary setup, they noted that the high energy content of flaxseed oil can often balance dietary energy, but this effect is highly dependent on the basal diet type. In contrast, our study found a significant elevation in lactose yield and SNF yield in the lower n-6:n-3 ratio group.
Physiologically, lactose serves as the primary osmotic regulator of milk volume by drawing water into the mammary alveolar lumen through an osmotic gradient. Consequently, the higher lactose yield observed in the lower n-6:n-3 ratio group provides a mechanistic explanation for the increased milk volume. This suggests that the lower n-6:n-3 ratio, particularly EPA and DHA derived from the conversion of ALA, may enhance the biosynthetic activity of mammary epithelial cells or improve mammary glucose uptake, rather than merely altering milk component synthesis pathways [21]. Furthermore, because this improvement in milk production was achieved without compromising body reserves, as evidenced by the comparable BW and final BCS between groups, it indicates a superior metabolic efficiency in nutrient utilization for lactation in cows receiving a reduced n-6:n-3 ratio. These results suggest that the specific fatty acid composition, rather than total fat intake, is the primary driver of these divergent production responses.
4.2. Ovarian Dynamics and Corpus Luteum Development
The reproductive outcomes in this study reveal important temporal and structural patterns in CL development as influenced by the dietary n-6:n-3 ratio. While synchronized ovulation and luteolysis rates remained comparable between groups, the morphometric characteristics of the resulting CL showed significant divergent responses during the mid-to-late luteal phase.
During the early luteal phase (Day 5 to Day 8 post-TAI), no significant differences were observed between the High and Low dietary n-6:n-3 ratio groups regarding CL cross-sectional area and volume. This indicates that the initial luteogenesis and early structural formation of the CL were not immediately impacted by the fatty acid profile of the diet. This is consistent with the broader understanding that while fat supplementation generally improves caloric density, its specific fatty acid effects often manifest more clearly as the cycle progresses [10]. A clear divergence emerged during the mid-to-late luteal phase (Days 11 to 14 post-TAI), where cows in the lower n-6:n-3 ratio group exhibited significantly greater CL cross-sectional areas and volumes compared to the greater n-6:n-3 group. This finding is of high physiological relevance, as CL volume is a primary determinant of its functional capacity. According to Santos et al. [10], feeding PUFAs can influence reproduction by increasing progesterone (P4) concentrations during the luteal phase and improving embryo quality. Our results support this, as a larger CL mass provides a greater population of steroidogenic luteal cells responsible for P4 synthesis. This is further corroborated Dirandeh et al. [48], who demonstrated that cows supplemented with omega-3 (linseed) maintained higher plasma P4 concentrations (15.0 ng/mL) compared to control groups (12.0 ng/mL) during the synchronized cycle, which matches the increased CL morphometry observed in our Low group.
The superior CL development and persistence in the lower n-6:n-3 ratio group may be explained by the biochemical inhibitory effects of lower n-6:n-3 ratio on the luteolytic pathway. It is well-documented that n-3 fatty acids, such as α-linolenic acid (ALA) and its derivatives (EPA and DHA), can inhibit the expression of cyclooxygenase-2 (COX-2) and competitively displace arachidonic acid (an n-6 PUFA) from cell membranes. As noted by both Santos et al. [10] and Dirandeh et al. [48], this mechanism leads to a significant reduction in the synthesis and pulsatile secretion of endometrial PGF2α, thereby preventing premature luteolysis and supporting CL persistence during the critical window of maternal recognition of pregnancy [49,50].
Although the enhanced luteal presence in the lower n-6:n-3 ratio group did not translate into significantly higher pregnancy rates in the current study, the results suggest a superior physiological environment for the developing embryo. The lack of a significant difference in conception rates may be attributed to the relatively short duration of supplementation or the moderate sample size, as previous reports suggesting higher fertility often utilized longer pre-calving supplementation periods [5]. Nevertheless, the significantly higher birth weight of offspring in the lower n-6:n-3 ratio group, further suggests that the lower n-6:n-3 diet maternal environment may have supported better embryonic or fetal development. These findings underscore the potential of the lower n-6:n-3 ratio as a precision nutritional tool to modulate the endocrine environment and enhance reproductive sustainability in dairy cattle.
4.3. Maternal Nutrition and Offspring Sex Ratio
The most striking observation in this study emerged from the offspring characteristics, where maternal dietary fatty acid composition significantly influenced both the sex ratio and birth weight. While reproductive success, measured by pregnancy rates per AI and the number of services per conception, was not significantly affected by the PUFA source, the phenotypic outcomes of the calves suggest that the specific n-6:n-3 ratio during the periconceptional and gestational periods plays a critical role in offspring development.
The strong trend toward a female-skewed sex ratio in the lower n-6:n-3 ratio group (66.67% vs. 42.90%) provides compelling evidence that maternal fatty acid profiles can influence sex allocation. This aligns with emerging research demonstrating that the n-6:n-3 ratio may exert opposing effects on sex ratio modulation [20]. Traditionally, the Trivers-Willard hypothesis [51] provides an evolutionary framework suggesting that maternal condition biases sex ratios; however, our results suggest that fatty acid composition may override general nutritional status, as the lower n-6:n-3 ratio produced more female offspring despite both groups maintaining adequate energy balance (stable BW and BCS).
Mechanistically, n-6 PUFAs like LA have been associated with a biochemical environment favoring Y-chromosome-bearing sperm or male embryo development. In contrast, n-3 fatty acids may counteract these effects through their anti-inflammatory properties and differential modulation of prostaglandin pathways [17]. Furthermore, male embryos are known to be more vulnerable to oxidative stress and developmental perturbations [52]. The antioxidant properties of n-3 PUFAs might selectively benefit female embryo survival or modulate epigenetic regulation affecting X-chromosome inactivation and stress response pathways [53].
Furthermore, the significant 4.8% increase in calf birth weight in the n-3 group highlights the profound role of n-3 PUFAs in fetal programming. Long-chain PUFAs, such as DHA and EPA, are essential structural components of cell membranes and are vital for fetal brain and retinal development. These fatty acids can alter placental gene expression (e.g., FATP1, FFAR4) to enhance angiogenesis and nutrient transfer across the placental barrier [16,54]. The higher maternal lactose yield observed in our n-3 group likely reflects an enhanced systemic nutrient flux that optimizes fetal growth trajectories.
In summary, n-3 fatty acid supplementation at an n-6:n-3 ratio of 4.8:1 offers substantial reproductive and production benefits. While n-6 supplementation may promote early luteal development, these effects were transient and did not persist into later reproductive outcomes. In contrast, the lower n-6:n-3 ratio resulted in improved milk yield, sustained mid-luteal maintenance, and superior offspring outcomes, specifically higher birth weights and a female-biased sex ratio. These findings suggest that targeted lower n-6:n-3 ratio is a valuable strategic tool for dairy operations, particularly for herd replacement programs where female calves carry higher economic value.
5. Conclusions
The findings of this study demonstrate that reducing the dietary n-6:n-3 ratio to 4.8:1 through rumen-protected n-3 supplementation during a timed AI program represents a strategic nutritional intervention to improve fertility potential and modulate offspring characteristics in lactating dairy cows. Lowering the dietary n-6:n-3 ratio significantly enhanced luteal development, as evidenced by the significantly greater corpus luteum volume and cross-sectional area during the critical mid-to-late luteal phase (days 11 and 14 post-TAI). This robust CL morphometry is indicative of an improved endocrine environment, likely mediated by increased progesterone (P4) synthesis and the potential suppression of luteolytic PGF2α secretion, thereby providing a superior physiological foundation for the establishment and maintenance of pregnancy.
Most notably, the Lower n-6:n-3 ratio exerted a profound influence on offspring outcomes, showing a strong tendency toward a female-skewed sex ratio (66.67% vs. 42.90%) and a significant 4.8% increase in calf birth weight. These results suggest that targeted fatty acid ratios can modulate fetal programming and sex allocation independently of general nutrition status, providing a non-invasive strategy to enhance herd replacement through the production of robust female calves.
As a secondary advantage, this nutritional strategy significantly enhances lactation performance, characterized by an 11.1% increase in milk yield and elevated lactose and SNF yields, without compromising the cows’ body condition or dry matter intake. Overall, targeted a lower n-6:n-3 ratio is a potent modulator of both reproductive and metabolic functions, providing a multifaceted approach to optimize production efficiency and sustainability in dairy operations.
Author Contributions
Conceptualization, J.N., P.N., T.N., J.P., and C.N.; methodology, J.N., S.N., N.L., P.N., T.N., J.P., and C.N.; software, J.N. and C.N.; validation, J.N. and C.N.; formal analysis, J.N. and C.N.; investigation, J.N., S.N., N.L. and C.N.; resources, J.N. and C.N.; data curation, C.N. and J.N.; writing—original draft preparation, J.N.; writing—review and editing, J.N. and C.N.; visualization, J.N. and C.N.; supervision, C.N.; project administration, J.N. and C.N.; funding acquisition, C.N. All authors have read and agreed to the published version of the manuscript.
Funding
The authors express their sincerest gratitude to the Fundamental Fund of Khon Kaen University, which has received funding support from the National Science, Research and Innovation Fund (NSRF) for providing financial support. Juthamas Nabthonglang received a grant from the National Research Council of Thailand (NRCT) through the Royal Golden Jubilee Ph.D. Program (Grant number: N41A661105).
Institutional Review Board Statement
The study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Khon Kaen University, in accordance with the Ethics of Animal Experimentation guidelines established by the National Research Council of Thailand. Approval record no. IACUC-KKU-141/67; reference no. 660201.2.31/10 (146).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request. In accordance with institutional regulations for the author’s doctoral thesis, the dataset will remain unpublished until the thesis evaluation process is complete. After this process, the data may be made publicly available.
Acknowledgments
The authors would like to express their sincere gratitude to the Department of Animal Science, Faculty of Agriculture, Khon Kaen University, for providing research facilities and technical support. We are also grateful to Betagro Public Company Limited, Thailand, for providing the concentrate diet used in the experiment. This manuscript was edited for clarity and style with the assistance of ChatGPT 5 (OpenAI, San Francisco, CA), which was used solely to improve grammar and readability. The authors reviewed and approved all content.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Diniso, Y.S.; Jaja, I.F. A Retrospective Survey of the Factors Responsible for Culling and Mortality in Dairy Farms in the Eastern Cape Province, South Africa. Sci. Afr. 2021, 12, e00838. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.G.; Bromfield, J.J. Tolerance and Innate Immunity Shape the Development of Postpartum Uterine Disease and the Impact of Endometritis in Dairy Cattle. Annu. Rev. Anim. Biosci. 2019, 7, 361–384. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Baez, G.M.; Garcia-Guerra, A.; Toledo, M.Z.; Monteiro, P.L.; Melo, L.F.; Ochoa, J.C.; Santos, J.E.; Sartori, R. Pivotal Periods for Pregnancy Loss during the First Trimester of Gestation in Lactating Dairy Cows. Theriogenology 2016, 86, 239–253. [Google Scholar] [CrossRef]
- Crouse, M.S.; Greseth, N.P.; McLean, K.J.; Crosswhite, M.R.; Pereira, N.N.; Ward, A.K.; Reynolds, L.P.; Dahlen, C.R.; Neville, B.W.; Borowicz, P.P. Maternal Nutrition and Stage of Early Pregnancy in Beef Heifers: Impacts on Hexose and AA Concentrations in Maternal and Fetal Fluids. J. Anim. Sci. 2019, 97, 1296–1316. [Google Scholar] [CrossRef]
- Staples, C.R.; Burke, J.M.; Thatcher, W.W. Influence of Supplemental Fats on Reproductive Tissues and Performance of Lactating Cows. J. Dairy Sci. 1998, 81, 856–871. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.R. Nutritional Interactions with Reproductive Performance in Dairy Cattle. Anim. Reprod. Sci. 2000, 60–61, 449–457. [Google Scholar] [CrossRef]
- Roche, J.R.; Friggens, N.C.; Kay, J.K.; Fisher, M.W.; Stafford, K.J.; Berry, D.P. Invited Review: Body Condition Score and Its Association with Dairy Cow Productivity, Health, and Welfare. J. Dairy Sci. 2009, 92, 5769–5801. [Google Scholar] [CrossRef]
- Mekuriaw, Y. Negative Energy Balance and Its Implication on Productive and Reproductive Performance of Early Lactating Dairy Cows: Review Paper. J. Appl. Anim. Res. 2023, 51, 220–228. [Google Scholar] [CrossRef]
- Mattos, R.; Staples, C.R.; Thatcher, W.W. Effects of Dietary Fatty Acids on Reproduction in Ruminants. J. Reprod. Fertil. 2000, 5, 38–45. [Google Scholar] [CrossRef]
- Santos, J.; Bilby, T.; Thatcher, W.; Staples, C.; Silvestre, F. Long Chain Fatty Acids of Diet as Factors Influencing Reproduction in Cattle. Reprod. Domest. Anim. 2008, 43, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, W.; Santos, J.E.P.; Staples, C.R. Dietary Manipulations to Improve Embryonic Survival in Cattle. Theriogenology 2011, 76, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Vargas-Bello-Pérez, E.; Busato, S. Advances in Fatty Acids Nutrition in Dairy Cows: From Gut to Cells and Effects on Performance. J. Anim. Sci. Biotechnol. 2020, 11. [Google Scholar] [CrossRef]
- Sordillo, L.M. Nutritional Strategies to Optimize Dairy Cattle Immunity. J. Dairy Sci. 2016, 99, 4967–4982. [Google Scholar] [CrossRef]
- Hess, B.W.; Lake, S.L.; Scholljegerdes, E.J.; Weston, T.R.; Nayigihugu, V.; Molle, J.D.C.; Moss, G.E. Nutritional Controls of Beef Cow Reproduction1. J. Anim. Sci. 2005, 83, E90–E106. [Google Scholar] [CrossRef]
- Zeng, X.; Li, S.; Liu, L.; Cai, S.; Ye, Q.; Xue, B.; Wang, X.; Zhang, S.; Chen, F.; Cai, C.; et al. Role of Functional Fatty Acids in Modulation of Reproductive Potential in Livestock. J. Anim. Sci. Biotechnol. 2023, 14, 24. [Google Scholar] [CrossRef]
- Wathes, D.C.; Abayasekara, D.R.E.; Aitken, R.J. Polyunsaturated Fatty Acids in Male and Female Reproduction1. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- Rincón Cervera, M.Á.; Bravo Sagua, R.; Soares-Freitas, R.; Lopez Arana, S.L.; de Camargo, A. Monounsaturated and Polyunsaturated Fatty Acids: Structure, Food Sources, Biological Functions, and Their Preventive Role against Noncommunicable Diseases. In; 2022; pp. 185–210 ISBN 978-0-12-823569-0.
- Schmitz, G.; Ecker, J. The Opposing Effects of n−3 and n−6 Fatty Acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Marei, W.F.A.; Khalil, W.A.; Pushpakumara, A.P.G.; El-Harairy, M.A.; Abo El-Atta, A.M.A.; Wathes, D.C.; Fouladi-Nashta, A. Polyunsaturated Fatty Acids Influence Offspring Sex Ratio in Cows. Int. J. Vet. Sci. Med. 2018, 6, S36–S40. [Google Scholar] [CrossRef] [PubMed]
- Moallem, U. Invited Review: Roles of Dietary n-3 Fatty Acids in Performance, Milk Fat Composition, and Reproductive and Immune Systems in Dairy Cattle. J. Dairy Sci. 2018, 101, 8641–8661. [Google Scholar] [CrossRef] [PubMed]
- Mattos, R.; Guzeloglu, A.; Badinga, L.; Staples, C.R.; Thatcher, W.W. Polyunsaturated Fatty Acids and Bovine Interferon-τ Modify Phorbol Ester-Induced Secretion of Prostaglandin F2α and Expression of Prostaglandin Endoperoxide Synthase-2 and Phospholipase-A2 in Bovine Endometrial Cells1. Biol. Reprod. 2003, 69, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.; Hammond, A.; Wathes, D.; Hunter, M.; Mann, G. Corpus Luteum–Endometrium–Embryo Interactions in the Dairy Cow: Underlying Mechanisms and Clinical Relevance. Reprod. Domest. Anim. 2008, 43, 104–112. [Google Scholar] [CrossRef]
- Domingues, R.R.; Andrade, J.P.N.; Cunha, T.O.; Madureira, G.; Moallem, U.; Gomez-Leon, V.; Martins, J.P.N.; Wiltbank, M.C. Is Pregnancy Loss Initiated by Embryonic Death or Luteal Regression? Profiles of Pregnancy-Associated Glycoproteins during Elevated Progesterone and Pregnancy Loss. JDS Communications 2023, 4, 149–154. [Google Scholar] [CrossRef]
- Calder, P.C. Long-Chain Fatty Acids and Inflammation. Proc. Nutr. Soc. 2012, 71, 284–289. [Google Scholar] [CrossRef]
- Silvestre, F.T.; Carvalho, T.S.M.; Crawford, P.C.; Santos, J.E.P.; Staples, C.R.; Jenkins, T.; Thatcher, W.W. Effects of Differential Supplementation of Fatty Acids during the Peripartum and Breeding Periods of Holstein Cows: II. Neutrophil Fatty Acids and Function, and Acute Phase Proteins. J. Dairy Sci. 2011, 94, 2285–2301. [Google Scholar] [CrossRef]
- Thangavelu, G.; Colazo, M.G.; Ambrose, D.J.; Oba, M.; Okine, E.K.; Dyck, M.K. Diets Enriched in Unsaturated Fatty Acids Enhance Early Embryonic Development in Lactating Holstein Cows. Theriogenology 2007, 68, 949–957. [Google Scholar] [CrossRef]
- Giller, K.; Drews, B.; Berard, J.; Kienberger, H.; Schmicke, M.; Frank, J.; Spanier, B.; Daniel, H.; Geisslinger, G.; Ulbrich, S.E. Bovine Embryo Elongation Is Altered Due to Maternal Fatty Acid Supplementation. Biol. Reprod. 2018, 99, 600–610. [Google Scholar] [CrossRef]
- Sinedino, L.D.P.; Honda, P.; Souza, L.; Lock, A.; Boland, M.; Staples, C.; Thatcher, W.; Santos, J. Effects of Supplementation with Docosahexaenoic Acid on Reproduction of Dairy Cows. Reproduction 2017, 153, REP–16. [Google Scholar] [CrossRef]
- Ambrose, D.J.; Kastelic, J.P.; Corbett, R.; Pitney, P.A.; Petit, H.V.; Small, J.A.; Zalkovic, P. Lower Pregnancy Losses in Lactating Dairy Cows Fed a Diet Enriched in α-Linolenic Acid. J. Dairy Sci. 2006, 89, 3066–3074. [Google Scholar] [CrossRef] [PubMed]
- Fabjanowska, J.; Kowalczuk-Vasilev, E.; Klebaniuk, R.; Milewski, S.; Gümüş, H. N-3 Polyunsaturated Fatty Acids as a Nutritional Support of the Reproductive and Immune System of Cattle—A Review. Animals 2023, 13, 3589. [Google Scholar] [CrossRef]
- Mattos, R.; Staples, C.R.; Williams, J.; Amorocho, A.; McGuire, M.A.; Thatcher, W.W. Uterine, Ovarian, and Production Responses of Lactating Dairy Cows to Increasing Dietary Concentrations of Menhaden Fish Meal. J. Dairy Sci. 2002, 85, 755–764. [Google Scholar] [CrossRef]
- Iwata, H. Factors Affecting the Primary Sex Ratio in Cows. J. Mamm. Ova Res. 2012, 29, 96–102. [Google Scholar] [CrossRef]
- Seidel, G.E.; Schenk, J.L. Pregnancy Rates in Cattle with Cryopreserved Sexed Sperm: Effects of Sperm Numbers per Inseminate and Site of Sperm Deposition. Anim. Reprod. Sci. 2008, 105, 129–138. [Google Scholar] [CrossRef]
- Jones, C.; Heinrichs, J. Learn To Score Body Condition Available online: https://extension.psu.edu/learn-to-score-body-condition (accessed on 16 March 2022).
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics, A Biometrical Approach. 3rd Edition; McGraw Hill, Inc. Book Co., New York, 1997.
- Allred, S.L.; Dhiman, T.R.; Brennand, C.P.; Khanal, R.C.; McMahon, D.J.; Luchini, N.D. Milk and Cheese from Cows Fed Calcium Salts of Palm and Fish Oil Alone or in Combination with Soybean Products. J. Dairy Sci.e 2006, 89, 234–248. [Google Scholar] [CrossRef] [PubMed]
- NRC, N.R.C. Nutrient Requirements of Dairy Cattle, 7th Rev. Ed.; The National Academies Press: Washington, DC, USA, 2001.
- Huang, G.; Wang, J.; Liu, K.; Wang, F.; Zheng, N.; Zhao, S.; Qu, X.; Yu, J.; Zhang, Y.; Wang, J. Effect of Flaxseed Supplementation on Milk and Plasma Fatty Acid Composition and Plasma Parameters of Holstein Dairy Cows. Animals 2022, 12, 1898. [Google Scholar] [CrossRef]
- Leduc, M.; Létourneau-Montminy, M.-P.; Gervais, R.; Chouinard, P.Y. Effect of Dietary Flax Seed and Oil on Milk Yield, Gross Composition, and Fatty Acid Profile in Dairy Cows: A Meta-Analysis and Meta-Regression. J. Dairy Sci. 2017, 100, 8906–8927. [Google Scholar] [CrossRef]
- Do Prado, R.M.; Palin, M.F.; Do Prado, I.N.; Dos Santos, G.T.; Benchaar, C.; Petit, H.V. Milk Yield, Milk Composition, and Hepatic Lipid Metabolism in Transition Dairy Cows Fed Flaxseed or Linola. J. Dairy Sci. 2016, 99, 8831–8846. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, B.J.; Soder, K.J.; Pereira, A.B.D.; Standish, R.; Brito, A.F. Production, Milk Fatty Acid Profile, and Nutrient Utilization in Grazing Dairy Cows Supplemented with Ground Flaxseed. J. Dairy Sci. 2019, 102, 1294–1311. [Google Scholar] [CrossRef]
- Sun, X.; Guo, C.; Wang, Q.; Zhang, Y.; Wang, Z.; Cao, Z.; Wang, W.; Li, S. Dietary N-3 Polyunsaturated Fatty Acids Enhance Dry Matter Intake in Transition Cows by Regulating Taste Transduction Gene Expression in the Liver Associated with Rumen Microbial Alteration. Anim. Nutr. 2025, 22, 502–513. [Google Scholar] [CrossRef]
- Petit, H.V.; Palin, M.F.; Doepel, L. Hepatic Lipid Metabolism in Transition Dairy Cows Fed Flaxseed. J. Dairy Sci. 2007, 90, 4780–4792. [Google Scholar] [CrossRef]
- Zachut, M.; Arieli, A.; Lehrer, H.; Livshitz, L.; Yakoby, S.; Moallem, U. Effects of Increased Supplementation of N-3 Fatty Acids to Transition Dairy Cows on Performance and Fatty Acid Profile in Plasma, Adipose Tissue, and Milk Fat. J. Dairy Sci. 2010, 93, 5877–5889. [Google Scholar] [CrossRef]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk Fatty Acids in Dairy Cows Fed Whole Crude Linseed, Extruded Linseed, or Linseed Oil, and Their Relationship with Methane Output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef]
- Moallem, U.; Lehrer, H.; Livshits, L.; Zachut, M. The Effects of Omega-3 α-Linolenic Acid from Flaxseed Oil Supplemented to High-Yielding Dairy Cows on Production, Health, and Fertility. Livest. Sci. 2020, 242, 104302. [Google Scholar] [CrossRef]
- Dirandeh, E.; Towhidi, A.; Ansari Pirsaraei, Z.; Adib Hashemi, F.; Ganjkhanlou, M.; Zeinoaldini, S.; Rezaei Roodbari, A.; Saberifar, T.; Petit, H.V. Plasma Concentrations of PGFM and Uterine and Ovarian Responses in Early Lactation Dairy Cows Fed Omega-3 and Omega-6 Fatty Acids. Theriogenology 2013, 80, 131–137. [Google Scholar] [CrossRef]
- Doyle, D.N.; Lonergan, P.; Diskin, M.G.; Pierce, K.M.; Kelly, A.K.; Stanton, C.; Waters, S.M.; Parr, M.H.; Kenny, D.A. Effect of Dietary N-3 Polyunsaturated Fatty Acid Supplementation and Post-Insemination Plane of Nutrition on Systemic Concentrations of Metabolic Analytes, Progesterone, Hepatic Gene Expression and Embryo Development and Survival in Beef Heifers. Theriogenology 2019, 127, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Jaśkowski, B.M.; Opałka, A.; Gehrke, M.; Herudzińska, M.; Czeladko, J.; Baumgartner, W.; Jaśkowski, J.M. A Critical Overview on Prostaglandin Inhibitors and Their Influence on Pregnancy Results after Insemination and Embryo Transfer in Cows. Animals 2021, 11, 3368. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L.; Willard, D.E. Natural Selection of Parental Ability to Vary the Sex Ratio of Offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, X.; Li, H. Mechanisms of Oxidative Stress-Induced Sperm Dysfunction. Front. Endocrinol. 2025, 16. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [PubMed]
- Roque-Jiménez, J.A.; Rosa-Velázquez, M.; Pinos-Rodríguez, J.M.; Vicente-Martínez, J.G.; Mendoza-Cervantes, G.; Flores-Primo, A.; Lee-Rangel, H.A.; Relling, A.E. Role of Long Chain Fatty Acids in Developmental Programming in Ruminants. Animals 2021, 11, 762. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flowchart of an experiment design indicating dietary treatment, modified Ovsynch protocol with TAI, and sample collection. US = ultrasonography, GnRH = gonadotropin releasing hormone (Receptal®), PGF = prostaglandin F2α (Lutalyse®), hCG = human Chorionic Gonadotropin (Chorulon®).
Figure 1.
Flowchart of an experiment design indicating dietary treatment, modified Ovsynch protocol with TAI, and sample collection. US = ultrasonography, GnRH = gonadotropin releasing hormone (Receptal®), PGF = prostaglandin F2α (Lutalyse®), hCG = human Chorionic Gonadotropin (Chorulon®).

Figure 2.
Total feed intake in dairy cows supplemented with greater dietary n-6:n-3 ratio (16.7:1) (∆) or lower dietary n-6:n-3 ratio (4.8:1) (○). Week had a significant effect (p < 0.0001). Treatment tended to affect DMI (p = 0.0989), whereas the treatment × week interaction was not significant (p > 0.05). Error bars represent SEM.
Figure 2.
Total feed intake in dairy cows supplemented with greater dietary n-6:n-3 ratio (16.7:1) (∆) or lower dietary n-6:n-3 ratio (4.8:1) (○). Week had a significant effect (p < 0.0001). Treatment tended to affect DMI (p = 0.0989), whereas the treatment × week interaction was not significant (p > 0.05). Error bars represent SEM.

Figure 3.
Milk yield in dairy cows supplemented with greater dietary n-6:n-3 ratio (16.7:1) (∆) or lower dietary n-6:n-3 ratio (4.8:1) (○). A significant treatment × week interaction was observed (p = 0.03). Asterisks (*) indicate weeks in which milk production differed between treatments (p < 0.05). Error bars represent SEM.
Figure 3.
Milk yield in dairy cows supplemented with greater dietary n-6:n-3 ratio (16.7:1) (∆) or lower dietary n-6:n-3 ratio (4.8:1) (○). A significant treatment × week interaction was observed (p = 0.03). Asterisks (*) indicate weeks in which milk production differed between treatments (p < 0.05). Error bars represent SEM.


Table 1.
Chemical composition and fatty acids profiles of the experimental diets
| Item | Treatments | |
|---|---|---|
| Greater n-6:n-3 (16.7:1) |
Lower n-6:n-3 (4.8:1) |
|
| Chemical composition (% of DM) | ||
| Moist | 9.90 | 10.10 |
| Dry matter | 90.10 | 89.90 |
| Crude protein | 24.00 | 24.48 |
| Fat | 4.05 | 4.13 |
| NDF | 28.26 | 30.70 |
| ADF | 15.27 | 16.59 |
| ADL | 5.23 | 5.34 |
| Ash | 9.19 | 8.43 |
| Fatty acids composition (g/100g) | ||
| Caprylic acid (C8:0) | 0.07 | 0.06 |
| Capric acid (C10:0) | 0.07 | 0.07 |
| Lauric acid (C12:0) | 0.93 | 0.99 |
| Myristic acid (C14:0) | 0.33 | 0.35 |
| Palmitic acid (C16:0) | 0.89 | 0.73 |
| Stearic acid (C18:0) | 0.15 | 0.35 |
| Arachidic acid (C20:0) | 0.02 | 0.02 |
| Lignoceric acid (C24:0) | 0.01 | 0.01 |
| Saturated Fat | 2.47 | 2.58 |
| Palmitoleic acid (C16:1n7) | 0.01 | 0.01 |
| Oleic acid (C18:1n9c; Omega-9) | 1.11 | 0.82 |
| Eicosenoic acid (C20:1n9; Omega-9) | 0.01 | - |
| Monounsaturated fatty acid | 1.13 | 0.83 |
| Linoleic acid (C18:2n6c; Omega-6) | 0.80 | 0.65 |
| Alpha-Linolenic acid (C18:3n3; ALA, Omega-3) | 0.05 | 0.13 |
| Polyunsaturated fatty acid | 0.85 | 0.78 |
| Unsaturated fat | 1.98 | 1.61 |
| Omega-3 (mg/100g) | 47.90 | 134.54 |
| Omega-6 (mg/100g) | 798.73 | 645.45 |
| Omega-9 (mg/100g) | 1115.76 | 824.82 |
| Omega-6 : Omega-3 | 16.70 | 4.80 |
Greater n-6:n-3 = greater dietary n-6:n-3 ratio (16.7:1); Lower n-6:n-3 = lower dietary n-6:n-3 ratio (4.8:1); NDF = neutral detergent fiber; ADF = acid detergent fiber; ADL = acid detergent lignin.
Table 2.
Effect of greater and lower dietary n-6:n-3 ratios on dry matter intake, body weight, and body condition score in lactating dairy cows.
Table 2.
Effect of greater and lower dietary n-6:n-3 ratios on dry matter intake, body weight, and body condition score in lactating dairy cows.
| Item | Treatments | SEM | p-value | |
|---|---|---|---|---|
| Greater n-6:n-3 (16.7:1) |
Lower n-6:n-3 (4.8:1) |
|||
| Feed intake (DM) | ||||
| Concentrate | ||||
| kg/day | 7.82 | 8.30 | 0.25 | 0.35 |
| % BW | 1.45 | 1.55 | 0.04 | 0.25 |
| g/kg BW0.75 | 69.58 | 74.15 | 2.05 | 0.30 |
| Roughage | ||||
| kg/d | 5.52 | 5.72 | 0.12 | 0.16 |
| % BW | 1.02 | 1.06 | 0.02 | 0.17 |
| g/kg BW0.75 | 49.18 | 51.17 | 1.06 | 0.13 |
| Total | ||||
| kg/d | 13.34 | 14.02 | 0.32 | 0.20 |
| % BW | 2.46 | 2.60 | 0.05 | 0.17 |
| g/kg BW0.75 | 118.76 | 125.31 | 2.54 | 0.14 |
| BW (kg) | 539.62 | 541.13 | 6.85 | 0.99 |
| BCS | ||||
| Initial | 2.50 | 2.50 | 0.07 | 0.77 |
| Final | 2.65 | 2.71 | 0.07 | 0.51 |
| BCS Change | 0.16 | 0.22 | 0.03 | 0.23 |
Greater n-6:n-3 = greater dietary n-6:n-3 ratio (16.7:1); Lower n-6:n-3 = lower dietary n-6:n-3 ratio (4.8:1); SEM = pooled standard error of the mean. DM = dry matter; BW = body weight; BCS = body condition score.
Table 3.
Effects of greater and lower dietary n-6:n-3 ratios on milk yield and composition in lactating dairy cows.
Table 3.
Effects of greater and lower dietary n-6:n-3 ratios on milk yield and composition in lactating dairy cows.
| Item | Treatments | SEM | P-value | |
|---|---|---|---|---|
| Greater n-6:n-3 (16.7:1) |
Lower n-6:n-3 (4.8:1) |
|||
| Milk yield (Kg/d) | 17.17 | 19.07 | 0.76 | 0.08 |
| 3.5% FCM (Kg/d) | 17.17 | 19.28 | 1.02 | 0.15 |
| ECM (Kg/d) | 15.68 | 17.60 | 0.88 | 0.13 |
| Efficiency, ECM/DMI | 1.14 | 1.20 | 0.05 | 0.41 |
| Milk composition, % | ||||
| Fat | 3.28 | 3.12 | 0.19 | 0.52 |
| Protein | 2.99 | 2.92 | 0.07 | 0.48 |
| Lactose | 4.49 | 4.53 | 0.07 | 0.58 |
| Solid not fat | 8.08 | 8.06 | 0.11 | 0.98 |
| Total solid | 12.47 | 12.69 | 0.63 | 0.96 |
| SCC (x103 cells/mL) | 440.7 | 356.3 | 153.97 | 0.55 |
| Milk production, kg/d | ||||
| Fat yield | 0.59 | 0.64 | 0.05 | 0.43 |
| Protein yield | 0.53 | 0.60 | 0.03 | 0.10 |
| Lactose yield | 0.80 | 0.94 | 0.04 | 0.03 |
| Solid not fat yield | 1.45 | 1.67 | 0.08 | 0.05 |
| Total solid yield | 2.24 | 2.64 | 0.17 | 0.14 |
Greater n-6:n-3 = greater dietary n-6:n-3 ratio (16.7:1); Lower n-6:n-3 = lower dietary n-6:n-3 ratio (4.8:1); SEM = pooled standard error of the mean. FCM = fat-corrected milk; ECM = energy-corrected milk; SCC = somatic cell count. 3.5% FCM was calculated as 0.4324 x milk yield (Kg) + 16.218 x fat yield (Kg); ECM was corrected for 4% fat and 3.4% protein using the equation: Milk yield × (0.124 × fat% + 0.073 × protein% + 0.256), according to NRC [9].
Table 4.
Effect of treatment on fertility parameters of dairy cows.
| Item | Treatments | SEM | p-value | |
|---|---|---|---|---|
| Greater n-6:n-3 (16.7:1) |
Lower n-6:n-3 (4.8:1) |
|||
| Synchronized ovulation rate | ||||
| first GnRH, % (no./no.) | 53.3 (16/30) | 58.6 (17/29) | 0.09 | 0.68 |
| second GnRH, % (no./no.) | 90.0 (27/30) | 96.6 (28/29) | 0.04 | 0.32 |
| Luteolysis (%) | 73.3 (22/30) | 72.4 (21/29) | 0.08 | 0.94 |
| pre-ovulatory follicle (mm) | 14.0 | 13.1 | 0.65 | 0.21 |
| Number of CL (n) | ||||
| d5 post TAI | 0.71 | 0.93 | 0.11 | 0.86 |
| d8 post TAI | 0.74 | 1.00 | 0.11 | 0.73 |
| d11 post TAI | 0.84 | 1.14 | 0.11 | 0.58 |
| d14 post TAI | 1.01 | 1.31 | 0.11 | 0.55 |
| CL diameter (mm) | ||||
| d5 post TAI | 16.97 | 15.35 | 0.84 | 0.17 |
| d8 post TAI | 19.51 | 21.21 | 0.84 | 0.15 |
| d11 post TAI | 21.66 | 24.04 | 0.86 | 0.05 |
| d14 post TAI | 24.61 | 26.58 | 0.89 | 0.10 |
| CL cross-sectional area (mm2) | ||||
| d5 post TAI | 246.38 | 196.04 | 26.49 | 0.18 |
| d8 post TAI | 313.71 | 366.87 | 30.52 | 0.16 |
| d11 post TAI | 380.10b | 461.94a | 30.90 | 0.04 |
| d14 post TAI | 474.14b | 560.32a | 32.41 | 0.03 |
| CL volume (mm3) | ||||
| d5 post TAI | 3,195.94 | 2,316.63 | 581.28 | 0.28 |
| d8 post TAI | 4,405.69 | 5,657.99 | 594.62 | 0.14 |
| d11 post TAI | 5,864.44 b | 7,774.80 a | 610.91 | 0.03 |
| d14 post TAI | 7,978.92 b | 10,395.00 a | 627.02 | 0.007 |
Greater n-6:n-3 = greater dietary n-6:n-3 ratio (16.7:1); Lower n-6:n-3 = lower dietary n-6:n-3 ratio (4.8:1); GnRH = gonadotropin-releasing hormone; TAI = timed artificial insemination; CL = corpus luteum; SEM = pooled standard error of the mean. a,b least square means within a row with different superscripts differ significantly (P < 0.05).
Table 5.
Reproductive performance, pregnancy rate per AI, offspring sex ratio and birth weight of offspring from cows supplemented with greater and lower dietary n-6:n-3 ratio.
Table 5.
Reproductive performance, pregnancy rate per AI, offspring sex ratio and birth weight of offspring from cows supplemented with greater and lower dietary n-6:n-3 ratio.
| Item | Treatments | SEM | p-value | |
|---|---|---|---|---|
| Greater n-6:n-3 (16.7:1) |
Lower n-6:n-3 (4.8:1) |
|||
| PR/AI, % (n/n) | ||||
| First AI | 23.33 (7/30) | 13.80 (4/29) | 0.08 | 0.37 |
| All AI | 96.67 (29/30) | 96.55 (28/29) | 0.01 | 1.00 |
| Number of services per conception, no. | 3.33 | 3.15 | 0.44 | 0.73 |
| Female offspring ratio, % (n/n) | 42.90 (12/28) | 66.67 (18/27) | 0.10 | 0.09 |
| Weight of offspring (Kg) | 32.92b | 34.50a | 0.54 | 0.02 |
Greater n-6:n-3 = greater dietary n-6:n-3 ratio (16.7:1); Lower n-6:n-3 = lower dietary n-6:n-3 ratio (4.8:1); PR/AI = pregnancy rate per artificial insemination. SEM = pooled standard error of the mean. a,b least square means within a row with different superscripts differ significantly (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



