Submitted:
14 April 2026
Posted:
15 April 2026
You are already at the latest version
Abstract
Considering the daily importance of vegetables in the human diet, breeders are expected to find faster and more accurate methods of creating new varieties of vegetables. To more precisely meet the demands of vegetable producers and consumers, breeders are increasingly combining hybrid and molecular techniques. The integration of hybrid and molecular breeding represents a logical step towards the development of efficient vegetable breeding strategies. For this purpose, the aim of this review is to point out on several representative vegetable species the possibilities, advantages, and disadvantages of the integration of hybrid and molecular methods of breeding. While conventional breeding techniques are based on selective breeding, mass selection, pure line selection, backcrossing, and hybrid breeding that exploit the effects of heterosis, advanced techniques such as phenomics, molecular markers, genome-wide association studies, and next-generation sequencing facilitate the identification and selection of desirable traits and improve nutritional quality. Definitely, molecular techniques represent a faster and more precise part of this mentioned integration. However, classical-hybrid breeding still develops stable, uniform, and marketable varieties without the high costs and significant access of advanced laboratory infrastructure, without the regulatory barriers that accompany genetic engineering.
Keywords:
vegetables
; hybridization
; breeding
; molecular marker
; CRISPR/Cas9
; integration
1. Introduction
Vegetables are grown in nearly 200 countries and represent the largest portion of the human diet in many regions of the world. A global survey has shown that at least 402 types of vegetables from 69 families and 230 genera have been grown and commercialized worldwide [1].
In addition to their widespread production, they provide energy and, in their various forms, ensure an adequate intake of essential vitamins, minerals, dietary fiber, and phytochemicals [2,3,4]. Many vegetable products meet the daily calorie requirements owing to their carbohydrate content [5], while legumes represent a particularly valuable source of essential amino acids [6,7]. Leafy greens and other vegetables are good sources of vitamins, minerals, and dietary fiber [8,9,10,11]. Sharing information about the nutritional value of vegetables and the benefits of changes in human consumption habits can give vegetables an even more important role in the human diet [12,13,14].
The World Health Organization (WHO) has recognized the importance of vegetable consumption. It promotes and recommends consumption of at least 400 g of vegetables and fruits per day to ensure optimal health and intake of essential nutrients lacking in other food groups [15,16].
Higher living standards have also increased local demands for fresh vegetables, leading to off-season vegetable production [17]. However, the production of high-quality vegetables is challenged by abiotic and biotic factors, including the decrease of arable land, urbanization, and climate change, all of which affect product quality [18,19,20].
Biotic stresses are caused by insects, pests, bacteria, fungi, viruses, nematodes, weeds, and other living organisms responsible for a 20–40% reduction in annual crop productivity [21,22]. Abiotic stresses include drought, high salinity, temperature extremes (cold and heat), floods, and heavy metal toxicity. Almost 70% of annual yield loss worldwide is due to abiotic stresses [23,24]. Therefore, plant breeders are required to develop cultivars resistant or tolerant to both types of stress [25].
Conventional plant breeding focuses on the development of new cultivars through traditional methods that depend heavily on natural genetic variations and processes [26]. However, traditional breeding approaches have several limitations; they are time-consuming, often requiring 10–14 years to produce an improved cultivar, as well as being labor-intensive and expensive [27,28]. Advancements in genetics and genomics, along with the introduction of innovative methods and techniques, have significantly improved plant breeding programs. Molecular markers represent one such advancement used in modern breeding strategies to shorten the selection process [29,30]. Molecular markers facilitate the identification and selection of desirable traits [31,32,33]. Techniques such as marker-assisted selection (MAS), marker-assisted backcrossing (MABC), and marker-assisted recurrent selection (MARS) are widely used to achieve greater precision in plant breeding [34,35,36].
For the aforementioned reasons, the aim of this review is to emphasize, by using certain vegetable species as examples, the importance of integration of hybrid and molecular techniques in vegetable breeding. In this review, only a few representative vegetable species are highlighted to illustrate the importance and potential of integrating hybrid and molecular techniques in breeding programs.
2. Conventional Plant Breeding
The need for food and fiber turned farmers into the first plant breeders, who unwittingly created locally diverse varieties through several breeding approaches, namely selection, introduction, and domestication. Local genotypes possess diverse traits conferring resistance to insects, pests, and diseases, yet they generally have lower yield potential. Vegetable varieties have a long history, shaped over generations by the efforts of growers and breeders to improve their traits. Over the course of time, scientists found the genetic basis of selection that farmers had already utilized to select plants with desirable traits, giving researchers the motivation to develop cultivars with wider adaptability and higher yield potential under conventional production conditions [37].
Vegetable breeding is challenged to meet the demands of both consumers and producers. Consumers prefer vegetables of good appearance, quality, taste, color, texture, durability, and nutritional content, while producers prioritize cultivars with high yield, uniformity, high market value, resistance to multiple diseases and pests, and tolerance to abiotic stresses [38,39,40].
Conventional breeding techniques are becoming more expensive, time-consuming, and labor-intensive due to the increasing complexity of traits required in modern vegetable cultivars. In conventional breeding programs, phenotypic selection is a complex and lengthy process that combines desirable traits gradually, step by step. This is particularly true for traits controlled by multiple genes and influenced by environmental variation. Breeding a new and promising cultivar can take 10 or 15 years before it becomes available for commercial production [37,41].
Conventional breeding methods require five to six generations to transfer a trait to the locally adapted, high-yielding cultivars, with a large number of progeny produced in order to select the plants with the desired combination of traits. The improved lines must be subjected to a series of tests at multiple locations before producers can identify a cultivar suitable for production. This process takes at least 7 to 10 years [42].
Conventional breeding typically begins with the selection of parental traits, often from wild relatives or landraces [37], which involves recurrent backcrossing to maintain the crop’s genetic basis. Traditional breeding methods are based on strategies such as selective breeding, mass selection, pure line selection, backcrossing, and hybrid breeding, with F1 hybridization and heterosis occurrence that affects productivity and quality [40,41,42,43].
Hybrid breeding is manifested in the superiority of F1 hybrids in relation to parental lines and is based on the exploitation of heterosis, i.e., hybrid vigor. The method is widely used in breeding vegetable crops, such as tomatoes, peppers, cabbages, cucumbers, and onions [43]. The advantage of hybrid breeding is reflected in higher and more stable yields, crop uniformity, improved resistance to diseases and pests, as well as improved technological and nutritional traits [44]. Hybrid breeding requires the development of genetically stable inbred lines, a process of which is long and costly, but the investment is justified by the high performance of the obtained hybrids and their economic value (Figure 1).
One of the quality traits of hybrid vegetables is phenotypic uniformity, which relies on the genetic homozygosity of the inbred parents [43,44,45].
Screening of different sources of vegetable crops, such as germplasm and breeding lines, resulted in identifying various bioactive compounds. Bioactive compounds such as anthocyanin, isoflavonoids, quercetin, betalins, lycopene, glucosinolates, sulforaphane, carotenoids, chlorophylls, and capsaicin are present in many vegetable crops, and are commonly referred to as nutraceuticals [46]. Conventional breeding strategies, such as selection, backcrossing, and pedigree-followed hybridization, have been used to develop new nutraceutical cultivars of vegetable crops, some of which have been released for commercial cultivation. Genes determining the mentioned quality traits have also been discovered using several molecular approaches, such as marker-assisted selection and transgenic crops [47].
Conventional breeding strategies are heavily dependent on whether the target traits are qualitative or quantitative. Understanding inheritance and genetic associations is critical for quantitative traits, as these traits are influenced by multiple genes and environmental conditions. Many compounds related to the nutritional quality of vegetables are quantitative traits, determined by numerous genes and influenced by environmental conditions, such as the content of pigments in tomatoes [48,49].
The selection and development of cultivars adaptable to specific conditions is the first step in breeding, taken to ensure stable, high-quality vegetables with superior nutritional value. Great pressure is put on breeders to develop stable cultivars that can adapt to different environmental conditions [40,48,49] and achieve a high yield [37,50].
If conventional breeding represented one side of the coin, biotechnology would surely be found on the other [42].
Several morphological and phenological traits contribute to yield heterosis in cucumber, cabbage, and tomato (Figure 1). In cucumber, key traits include number of fruits, days to first female flowering, days to first harvest, first nodal position of the female flower, sex ratio (M/F), fruit length, fruit diameter, and fruit weight (Figure 1A, 1B). In cabbage, yield is primarily associated with fruit length, fruit diameter, and fruit weight (Figure 1A, 1C). In tomato, important traits include number of fruits, days to first female flowering, days to first harvest, number of flowers or fruits per cluster, fruit length, fruit diameter, and fruit weight (Figure 1A, 1D) [51].
3. Genetic Diversity Assessment
One of the main prerequisites for successful breeding is the existence of wide genetic variability in the starting material. On the other hand, the successful use of the existing genetic variability requires its reliable assessment. Thus, for example, the effect of hybrid heterosis is determined by the genetic diversity of the parents. The more genetically different the parents are, the better the chance of using heterosis. Therefore, the assessment of genetic diversity has a key role in the development of hybrids and the characterization of germplasm collected from different regions [52].
Molecular markers are a tool that can be used to precisely determine and analyze the connections between samples based on the assessment of genetic similarity. Therefore, characterizing genotypes at both the genetic and phenotypic levels is the first step towards efficiently conserving, maintaining, and utilizing the existing genetic diversity of plant germplasm [53]. Moreover, knowledge about the existence of genetic links between samples within the germplasm can be a reliable indicator when selecting parents for the development of cultivars and hybrids [54,55].
The study of genetic diversity using molecular markers is a widely practiced approach. Several researchers have evaluated genetic diversity in various vegetable crop species using molecular markers [30,56,57]. Vázquez et al. [58] determined the existence of a very narrow genetic base with more than 50% of deoxyribonucleic acid (DNA) fragments common to 11 pepper breeding lines using random amplified polymorphic DNA (RAPD) markers. Dijkhuizen et al. [59] used restriction fragment length polymorphism (RFLP) to investigate the genetic relationships in two collections of cucumber (Cucumis sativus L.) germplasm. Genetic relationships determined from RFLP markers were consistent with predictions based on fruit type and provenance data. During the assessment of world tomato collections, Villand et al. [60] found that samples from South America exhibited greater genetic diversity compared to European samples. Levi et al. [61] determined the existence of 80.2-97.8% genetic similarity among traditional watermelon cultivars using inter-simple sequence repeats (ISSR) and amplified fragment length polymorphism (AFLP) markers and concluded that these markers are very effective in determining genetic diversity among watermelon cultivars. Muminoric et al. [62] used 12 AFLP and 10 ISSR primers in a genetic diversity assessment of 68 radish cultivars. Blanca et al. [63] performed genotyping by sequencing using single nucleotide polymorphism (SNP) markers for 1,254 tomato samples that included European traditional and modern cultivars, early domesticated cultivars, and wild relatives. Very low genetic diversity was determined in the analyzed samples, with only 298 polymorphic loci (at the 95% threshold) out of a total of 64,943 variants.
Over the last few decades, simple sequence repeat (SSR) markers, also known as microsatellites, have gained popularity due to their high reproducibility, multiallelic nature, codominant inheritance, and broad genome coverage. SSR markers have been successfully used to analyze genetic diversity in different plant species. SSR and markers based on sequence-related amplified polymorphism (SRAP) were used to study the genetic variability of certain traditional tomato cultivars in Spain [52]. Singh et al. [53] evaluated the genetic diversity of 47 pea genotypes using 34 SSR markers specific to this plant species.
4. Marker-Assisted Selection (MAS)
Conventional plant breeding is based primarily on the phenotypic selection of superior individuals in segregating populations resulting from hybridization [64]. However, the development of improved cultivars in this way is limited because of its duration. Given the numerous stages of crossing, selection and testing involved in the conventional breeding process, it takes one or two decades to create a new cultivar [65]. This traditional approach is slow, labor-intensive, and expensive. Advances in genetics and genomics rely on new tools, techniques, and approaches that can be applied to improve breeding programs. Molecular markers, genetic linkage maps, marker assays, and whole genome sequencing have been developed in many plant species, including vegetable species. Methods such as marker-assisted selection, marker-assisted backcrossing, marker-assisted recurrent selection, and genomic selection, all applicable to precision breeding, are currently at various stages of development in certain vegetable species. Their use depends on available resources and the complexity of the genetics and breeding of the particular species [66] (Figure 2).
Marker-assisted selection involves the use of molecular markers to select plants carrying genes responsible for traits of interest [32]. With the development and availability of molecular markers and detailed molecular genetic maps, marker-assisted selection has become widely applied in plant breeding for both qualitative and quantitative traits [67]. Marker-assisted selection is a practical application of molecular markers in plant breeding. The process consists of several stages, including identification of sources of desirable alleles, detection of polymorphic markers through genome analysis, establishment of associations between marker allelic variation and target traits, estimation of the effects of candidate genes and loci controlling quantitative traits (QTL), and evaluation of their stability across different environments and genetic backgrounds [68]. On completion of the last MAS phase, it is possible to single out the markers useful for implementation of the so-called molecular breeding [54].
The fundamental principle of marker-assisted selection relies on linkage disequilibrium (LD) between the marker and the gene/QTL of interest. LD refers to the non-random association of alleles that tend to be inherited together due to genetic linkage. With sufficiently strong LD, selection can be carried out directly through markers, with the aim of indirectly increasing the frequency of certain alleles. In this way, marker-assisted selection would enable simpler, faster, cheaper, and more efficient selection [69].
In marker-assisted selection, DNA markers can be used for two primary purposes:
- Tracking favorable alleles (dominant or recessive) through generations;
- Identification of most suitable individuals among the offspring in the separation populations, based on the allelic representation in a part of the genome or the entire genome [64].
Most commonly used breeding strategies in MAS are the selection of desirable traits or QTL from breeding lines or populations, introduction of genes from breeding lines or wild relatives, marker-assisted backcrossing, recurrent marker-assisted selection, and gene pyramiding [67]. A schematic representation of conventional, advanced, and modern strategies for vegetable breeding is shown in Figure 3.
4.1. Introgression and Selection of Traits and QTL from Breeding Lines and Wild Relatives
One of the most practical applications of DNA-based markers in breeding programs is the ability to select phenotypic traits using markers tightly linked to the genes controlling the trait [35,70]. The ability to select plants based on genotype rather than phenotype is highly attractive to plant breeders, as it avoids issues associated with phenotypic selection. Interactions with other genetic and environmental factors limit the effectiveness of phenotypic evaluations [70]. On the other hand, the probability of identifying a gene through a marker is inversely proportional to the distance between the gene and the marker. In addition, a large proportion of individuals in segregating generations exhibit high heterozygosity, which complicates visual selection [71]. Selection based on genetic composition helps overcome this limitation [64].
Marker-assisted selection is based on the concept that it is possible to determine the presence of a gene from the presence of a marker if there is a close relationship between them. This technique is used in breeding programs for improvement and rapid selection in early generations [72]. Although markers can be used at any stage of the breeding process, marker-assisted selection is a major advantage in early generations, as plants with undesirable genes and combinations of genes can be eliminated [35]. This allows breeders to focus on a smaller number of priority lines in subsequent generations. An important prerequisite for successful selection in early generations using markers is working with large populations and low heritability of the traits of interest because, in individual selection, the relative efficiency of marker-assisted selection is greatest for traits with low heritability. Unfortunately, the biggest disadvantage of marker-assisted selection in early generations is the cost of genotyping a large number of plants [67].
Marker-assisted selection is an effective approach in which traditional plant breeding is improved by the use of molecular markers, thereby increasing the precision and reliability of selection [25]. Through marker-assisted selection, plant breeders can detect and select appropriate dominant and recessive alleles across generations of a breeding population [73]. Within MAS, QTL associated with significant agronomic traits and valuable resistance genes are used, together with their specific DNA markers that are tightly linked to the trait of interest [74].
Many fruit traits are controlled by a relatively large number of QTL, each contributing positively or negatively to the phenotype. DNA markers are particularly useful in the selection of such quantitative traits that are difficult to select for based on phenotypic assessment alone. QTL regions controlling such traits have been identified in several cultivated plant species, such as tomato [64].
Wild relatives of cultivated plants are important genetic resources, as potential sources of desirable alleles or genes lost through breeding over the years. In this regard, tomato provides a well-known example of the importance of wild relatives in improving agronomic traits, as they have been extensively used in tomato breeding programs. Lycopersicon pimpinellifolium has been an attractive source of germplasm for breeders due to its closeness to cultivated tomatoes and the simplicity of backcrossing. Genes for tolerance to Fusarium wilt and bacterial spot caused by Fusarium oxysporum f. sp. lycopersici and Pseudomonas syringae pv. tomato, respectively, have thus been transferred from wild relatives to cultivated tomato. Lycopersicon parviflorum and Lycopersicon chmielewskii are interesting because of the high sugar and vitamin C content in their fruits [75,76]. Furthermore, a series of backcrosses between L. chmielewskii (10-11% dry matter) and the Californian variety VF-145 (about 5% dry matter) resulted in lines with increased sugar content (about 7.5% dry matter) and acceptable fruit size and color [77]. Also, it has been noted that Lycopersicon hirsutum, in its natural habitat, is characterized by a high degree of resistance to insects, which was also confirmed by laboratory tests. For instance, during a mite attack, mites are trapped in the sticky exudate of the glandular scales that cover the L. hirsutum plant, which confirms a significant negative correlation between glandular malleat density and mite infestation. Moreover, L. hirsutum contains the natural insecticide 2-tridecanone in a 72-fold higher concentration compared to the tomato varieties cultivated for commercial purposes. As a result, L. hirsutum is resistant to a large number of insects and therefore is a valuable source of germplasm, enabling breeders to increase tolerance in commercially grown tomato cultivars. In addition to its well-documented insect resistance, L. hirsutum also contains genes for resistance to other pathogens. For example, certain specimens of Lycopersicon hirsutum f. glabratum are resistant to the bacterial spot and to the root rot caused by nematodes (Meloidogyne spp.). Furthermore, it serves as an important source of resistance to fungal diseases such as Septoria lycopersici and viral diseases such as the tobacco mosaic virus . Consequently, due to its preference for higher altitudes, L. hirsutum was investigated as a possible source of genes for tolerance to low temperatures [77,78].
Additionally, markers can also be used in the introgression of desirable traits from wild germplasm into cultivated varieties. In particular, markers associated with genes from the wild donor parent, together with markers distributed across the genome of the improved cultivar (recurrent parent), are used to select progeny derived from such crosses. Thus, markers can be used to track the favorable donor alleles and will also help reduce the genetic background of the donor in the offspring [64].
4.2. Marker-Assisted Backcrossing
In traditional backcrossing, the reconstruction of the recurrent parent genotype takes more than six generations, whereas marker-assisted backcrossing helps complete this process in three generations or less. Marker-assisted backcrossing is particularly effective for introducing individual alleles into a different genetic framework, for example, when improving an existing variety by introducing a specific trait [67].
Backcrossing is used in plant breeding to transfer favorable traits from a donor plant (donor parent) to an elite genotype (recurrent parent). During repeated crosses, the first original cross (F1) is crossed again with the recurrent parent until most of the genes from the donor plant have been eliminated. The purpose of backcrossing is to keep all the desirable traits already present in the recurrent parent and to “take” only the desired allele from the parent representing the donor of the gene of interest [79]. However, segments of the donor plant linked to the target allele may remain relatively large even after many generations of backcrossing, a limitation that can be addressed by using molecular markers [67].
In this context, markers can be used in backcrossing to control the target gene (foreground selection), accelerate reconstruction of the genotype of the recurrent parent (background selection), or select offspring with the target gene and the occurred recombinations (recombinant selection). Moreover, the transfer of recessive genes through conventional breeding techniques requires generations of inbreeding after each backcross, a procedure that is unacceptably slow for commercial breeding programs [64].
Marker-assisted backcrossing is likely to become a popular approach, largely for the same reasons that encouraged traditional backcrossing [80].
4.3. Marker-Assisted Recurrent Selection (MARS)
Recurrent selection is a key breeding strategy aimed at gradually increasing the frequency of desirable alleles in a population through repeated cycles of selection and recombination. In contrast, MARS applies molecular marker information to detect and select multiple genomic regions controlling complex traits, enabling the accumulation of superior alleles into high-performing genotypes within one or more related populations [81].
Marker-assisted recurrent selection is a breeding approach that aims to accumulate favorable alleles for a relatively large number of QTL in a given population, using a set of markers significantly associated with the traits of interest [82]. It is an important population improvement method that focuses on cyclical selection and enriching favorable alleles from biparental or multiparental introgression at several loci [83]. Marker-assisted recurrent selection begins with a heterogeneous base population and thereafter uses superior recombinants during each cycle to produce an improved population, inbred line, or hybrid. Recognizing its potential, breeding programs in both public and private sectors have successfully applied this approach to cultivated crops [25,83,84].
4.4. Cumulative Gene Pyramiding
The study of genetic resistance against disease in vegetable crops has seen tremendous progress over the past 3 decades. Research of qualitative resistance has had a head start compared with the more complex quantitative resistance, which still has many unanswered questions [85]. Crop plants often experience more than one biotic and abiotic stress. Stress tolerance is genetically complex and results from interplay of multiple genes [86].
Gene stacking, also referred to as pyramiding, represents an effective strategy for incorporating multiple target genes or QTL derived from distinct parental lines into a single genotype within a relatively short timeframe (approximately two to three generations). In contrast, traditional breeding approaches typically require at least six generations recovering about 99.2% of the recurrent parent genome [87,88]. One of the main limitations of conventional crop improvement is that, alongside the target traits, undesirable genes may persist even after multiple backcross generations, and identifying these residual genes can be challenging. In contrast, developments in molecular technologies have enabled more accurate, efficient, and accelerated breeding approaches through the use of molecular markers [89,90]. Marker-assisted selection represents an indirect breeding approach in which molecular markers closely associated with target genes are used to identify and select desirable traits. This strategy is particularly valuable for agronomic characteristics that are difficult to evaluate phenotypically, such as resistance to pathogens, diseases, and abiotic stresses, thereby contributing to the prevention of yield reduction and deterioration of quality traits [91].
To pyramid disease or pest resistance genes containing similar phenotypic effects, often without suitable breeds available, marker-assisted selection may be the only practical method – especially in cases where one gene masks the presence of others. The fact that pyramiding must be repeated after each cross must not be overlooked when applying these strategies in practical breeding, because the pyramided resistance genes segregate in the progeny [67].
The integration of gene pyramiding strategies, particularly those supported by MAS and related methodologies, has significantly expedited the production of durable, resistant or tolerant lines, enabling precise selection and substantially reducing the time required for their development [92]. A good example of gene pyramiding for biotic stress tolerance could be seen in the tomato trait, where resistance toleaf curl and spotted virus has been pyramided by genes Ty-1, Ty-3, and Sw-5 [93]. Another example of gene pyramiding is observed in pepper, where the genes ( the Me1 andMe2 have been used to confer resistance to root-knot nematode [94]. It is also a known fact that the probability of detecting a marker associated with a gene responsible for tolerance to a certain disease is inversely proportional to the genetic distance between the marker and the gene [95].
For a better estimate, the genetic distance between the markers and genes should be calculated using a large population or, even better, from multiple crosses. Associations between markers and monogenic resistance genes are typically identified through genetic mapping. Although RFLP and RAPD markers were frequently used in this strategy [72], kompetitive allele specific PCR (KASP) markers [96] and SNP markers [97] are now increasingly applied.
5. Examples of Marker-Assisted Selection in Vegetable Crops
5.1. Tomato
Tomato (Solanum lycopersicum L.), a member of the Solanaceae family, ranks as the second most important vegetable crop worldwide, following potato (Solanum tuberosum L.) [98]. Most of the existing tomato cultivars were developed by conventional breeding techniques. However, numerous problems of the breeding process have led to the development and application of new technologies in vegetable crop breeding, including tomato breeding [69,99].
Although tomato was one of the first cultivated plant species for which genetic markers and maps were used in breeding programs, until the early 1980s, almost all tomato breeding programs relied mainly on phenotypic selection. With the discovery of more suitable genetic markers for breeders, including polymerase chain reaction (PCR)-based markers and SNP, there has been an increased interest in the use of markers to facilitate the tomato breeding process. Marker-assisted selection is often utilized in tomato breeding programs to introduce and combine genes, especially when breeding for tolerance to multiple diseases. For instance, sequence characterized amplified region (SCAR), cleaved amplified polymorphic sequences (CAPS), and other PCR-based markers are used in most tomato breeding programs to select for resistance traits to major diseases controlled by a single gene [66]. Six genes (Ty1, Ty2, Ty3, Ty4, Ty5, Ty6) conferring resistance to tomato leaf curl virus, a major cause of crop losses, have been identified using molecular markers [52]. Markers are also routinely used for many other purposes, including testing hybrid purity and screening breeding populations for fruit quality traits. On the other hand, they are not typically used in breeding for certain complex traits, such as polygenic traits for disease resistance, abiotic stress tolerance, yield, and various fruit quality traits [66].
5.2. Pepper
Pepper (Capsicum annuum L.) plays a significant role in agriculture, yet its production remains highly susceptible to abiotic and biotic factors such as drought conditions, infectious diseases, and temperature fluctuations. Although the variety development process takes many years, advances in biotechnology over the past 25–30 years have resulted in a reduction in breeding time. The use of advanced DNA-based methods in breeding allows breeders to easily overcome the challenges of the traditional breeding process and has reduced the time required to release new varieties [100]. Genetic diversity studies using molecular methods allow breeders to rapidly characterize the genetic material of interest [101,102,103], develop markers linked to desired traits [104,105], make selections using developed markers [106], and map species genomes [107]. RFLP [108], Some of the molecular marker systems used in the diversity of Capsicum germplasms are RAPD [109], AFLP [110,111], microsatellite markers [112,113], SNP [114,115], and SRAP [116,117,118].
Several markers linked to specific traits have been developed for use in pepper breeding programs, including allele-specific CAPS markers for the pvr1, pvr11, pvr12, and pvr2 genes, which confer resistance to various strains of the tobacco mosaic virus. Also, several closely related markers have been developed for tolerance to important pepper diseases, including those caused by Phytophthora capsici, pepper mottle virus, tomato spotted wilt virus, and anthracnose [66].
Using derived cleaved amplified polymorphic sequences (dCAPS) and SCAR markers, two genes that controlled capsinoid biosynthesis (p-AMT and Pun1) were incorporated into a new pepper cultivar called Maru Salad, resulting in a higher concentration of capsinoids in this pepper cultivar compared to the control [106]. Capsaicinoids are a specific class of compounds found in hot peppers, responsible for their pungency [119].
By applying marker-assisted backcrossing and pyramiding with the recessive allele pun[1,120] succeeded in the introgression of the male sterility gene ms10 from the hot pepper (Capsicum annuum var. annuum) genotype into the genotype of bell pepper (C. annuum var. grossum). The selection of plants containing the gene of interest was aided by the use of the SSR marker linked to the ms10 gene located on chromosome I and the SCAR marker, indicating the presence of the no-heat gene pun1.
In short conclusion, by enabling precise tracking of desirable alleles for traits such as disease resistance, fruit quality, and abiotic stress tolerance, MAS accelerates breeding cycles and improves selection efficiency compared with traditional phenotypic approaches. Overall, MAS will remain a key component of pepper breeding strategies, helping to meet global demands for resilient, high-yielding, and high-quality pepper varieties.
5.3. Cabbage
Some of the most important objectives in cabbage breeding are to achieve crop uniformity, high yield, desirable appearance, and the appropriate size, shape, and color of economically important plant organs [39]. Today, most of the currently grown cabbage cultivars are F1 hybrids, characterized by high yields and early maturity as a result of heterosis [121]. Conventional breeding of head cabbage begins with crosses between parent lines carrying desirable traits, followed by successive generations of self-pollination and selection aimed at creating improved cultivars [122].
However, as with other Brassica species, the development of modern head cabbage (Brassica oleracea var. capitata L.) cultivars increasingly depends on molecular markers implemented via MAS. Further markers are use in hybrid breeding. Markers for Male Sterility Maintenance: PCR [123,124], CAPS [125], SSR [125,126], insertion-deletion (InDel) [127,128,129], RAPD [130], extended random primer amplified region (ERPAR), SCAR [126,131,132], AFLP [132], KASP [133].
Following markers for evaluation of combining ability, population genetic diversity and phylogenetic relationships: RAPD [134,135,136], SSR [136,137,138,139,140], ISSR [141], AFLP [142,143], expressed sequence tags–single sequence repeats (EST-SSR) [144], SNP [145], KASP [146].
Here are also markers for genetic purity testing of F1 hybrid seeds: RAPD markers [147], Combination of RAPD, ISSR and SSR markers [148], Combination of SSR, SRAP and ISSR markers [149], SSR [150].
Last in this group are fingerprinting markers sets: set of RAPD markers [151], set of KASP markers [152] and sets of SNP markers [153].
Next group are markers for morphological traits and it contains markers for petal color: InDel markers [154,155], cabbage waxiness: SSR markers [156,157,158,159], InDel markers [158,160], PCR markers [161], KASP markers [162], and leaf color: InDel markers [163,164].
Group of markers for resistance to abiotic factors is defined by markers for resistance to head splitting: SSR markers [165], InDel makers [166]. Next are bolting resistance and flowering time markers: SSR [167] and InDel markers [1,168]. Last are markers for tolerance to prolonged high and low temperatures: PCR markers [169,170], InDel markers [164] and SNP markers [171].
Last group contain markers for disease resistance. In this group are markers for Fusarium wilt resistance: PCR-RFLP markers [150], SCAR markers [172], SSR markers [173,174,175], SNP/InDel markers [173], InDel markers [176,177], CAPS markers [177], and dCAPS markers [177]. Further are Black rot resistance markers: SSR markers [178,179] and InDel markers [180]. Clubroot resistance markers are: SCAR, CAPS markers [181], SSR markers [182,183], and SCAR, SSR, CAPS markers [184]. Last in the group are Downy Mildew resistance markers: InDel [185] and KASP markers [186].
Therefore, although current markers already play a significant role in speeding up the breeding process, future research should prioritize a deeper genetic analysis of quantitative traits, the development of more advanced and efficient markers—such as KASP and SNP markers derived from whole-genome sequencing—and the expansion of available marker systems to encompass a wider range of valuable traits [187].
5.4. Lettuce
Lettuce (Lactuca sativa L.) is a widely cultivated vegetable of significant economic importance [188]. The species includes seven principal genotype groups grown in numerous regions, across varying altitudes, and represented by many traditional cultivars [189].
The development of new lettuce cultivars relies on both traditional and modern breeding strategies to fulfill the specific demands of producers and consumers. However, the generation of F1 hybrids has proven unsuccessful because the pollen of L. sativa is heavy, viscous, and poorly transferable.
The main aim of traditional lettuce breeding is to develop varieties that perform well in a wide range of environments. Given current global challenges, breeders must also account for changing agricultural land, biotic and abiotic stresses, and the need for better quality and higher yields [190,191].
Several drawbacks are associated with traditional breeding techniques, including difficulty in ensuring the genetic purity of progeny, slow breeding cycles, limited offspring production, and the introduction of undesirable traits that can only be removed through prolonged backcrossing.
Breeding programs for L. sativa mainly aim to improve plant quality and strengthen resistance to early wilting, pests, and diseases. These objectives are achieved through biotechnology and genetic engineering methods, such as gene transfer. Modern technologies have enabled the preservation of diverse genotypes, supported healthier growth environments, and reduced the time required for breeding compared with traditional in vivo approaches [192].
Different molecular markers techniques are use for various genetic purposes of L. sativa species. Usage of RFLP markers we found in investigations of Landry et al.[193], Kesseli et al. [194], Vermeulen et al.[195], Kesseli et al. [196] and El-Esawi [197]. Further, RAPD markers described Kesseli et al.[196], Yamamoto et al. [198], Tardin et al. [199] and El-Esawi [197]. Utilization of AFLP markers we could find in works from Hill et al. [200], Jeuken et al. [201], Koopman et al. [202], Van Treuren and Van Hintum [203] and El-Esawi [197]. Microsatellite makers in lettuce breeding were described by Witsenboer et al. [204], Van de Wiel et al. [205], Van de Wiel et al. [206], Van Treuren and Van Hintum [203], Rauscher and Simko [207], El-Esawi [197] and Rui et al. [208]. Next type of EST-SSR markerswere demonstrated by Simko [209] and Riar et al. [210]. SNP type of markers were listed in works of MorenoVa’zquez et al. [211], Simko et al. [212], El-Esawi [197] and Walley et al. [213]. Last type of markers, or SRAP and Sequence-specific Amplified Polymorphism (SSAP) were investigated [203,214].
Conventional breeding remains a primary strategy for the incorporation of novel alleles by crossing genotypes derived from diverse plant genetic resources to improve multiple agronomic traits. For example, modern varieties are hybridized with locally adapted genotypes to achieve early maturity, increased yield, improved quality, and enhanced resistance to abiotic and biotic stresses. Moreover, modern biotechnological approaches, including advanced DNA sequencing and molecular tools, are essential for the development of lettuce cultivars possessing desirable characteristics [215].
6. Genomic Selection as a Bridge Between Molecular and Hybrid Breeding
Genomic selection (GS) represents a major advancement that enables the integration of molecular and hybrid breeding techniques in vegetables. Unlike MAS, which targets markers closely linked to genes controlling traits of interest or major QTL, GS utilizes genome-wide marker information combined with phenotypic data to predict the breeding value of individuals based on genomic estimated breeding values (GEBVs) [216,217]. Breeders can use this method to capture the cumulative effects of numerous small-effect loci controlling complex quantitative traits, such as yield, fruit quality, and abiotic stress tolerance [218].
In hybrid vegetable breeding, GS provides substantial benefits by enabling the early prediction of parental line performance and potential hybrid combinations before extensive phenotypic evaluation. Genomic prediction typically involves establishing a relatively small training population alongside a larger virtual test population. High-density genotypic and phenotypic data are collected from the training population to develop genomic prediction models, which are then applied to candidate populations to estimate the effects of individual genetic markers on target traits or to determine the breeding values of prospective hybrids [219]. Kondo et al. [220] demonstrated that genomic prediction integrating SNP genotypic data and parental phenotypic traits enabled highly reliable prediction of F1 hybrid phenotypes in chili peppers. Genomic prediction models also allow for more precise estimation of general combining ability (GCA) and, increasingly, specific combining ability (SCA), facilitating the definition of heterotic groups based on genomic relationships rather than solely on pedigree information. This approach enhances parental line selection and improves hybrid performance prediction [221]. For example, Yue et al. [222] found that genomic distance between parental lines, as determined by SNP markers, improved the prediction of heterosis and F1 hybrid performance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). GS has the potential to accelerate the improvement of complex traits in line and hybrid development, particularly for traits with low heritability and pronounced genotype × environment interactions, where conventional phenotypic selection is less effective. This reduces the need for extensive field trials, shortens breeding cycles, and lowers operational costs [223].
The integration of GS and other modern vegetable breeding approaches offers additional benefits. Combining GS with doubled haploid (DH) technology enables rapid cycle breeding, as DH lines can simultaneously serve as training and selection populations, increasing prediction accuracy and shortening generation time while improving annual genetic gain [224]. Singh et al. [225] demonstrated in cauliflower that the combination of fully homozygous DH lines and microsatellite markers allows more accurate prediction of heterosis for key agronomic traits, since genetic distance derived from these tools provides a more reliable assessment of hybrid performance than phenotypic methods. Furthermore, combining GS with high-throughput phenotyping (HTPP) platforms [226] and machine learning algorithms [227] further enhances prediction accuracy. With the rapid development of artificial intelligence (AI), machine learning algorithms are increasingly applied to capture the effects of multiple genes, further improving prediction precision [228]. Among the most advanced applications of AI in this field are genomic language models (gLMs), which interpret genomic sequences as structured linguistic data. These models learn contextual relationships between genetic elements, thereby enabling precise gene annotation, regulatory motif identification, and accurate prediction of variant effects. Although the use of gLMs in vegetable crops is still at an early stage, models trained on the tomato genome have already shown notable potential in predicting promoter activity, guiding single guide ribonucleic acid (sgRNA) design for clustered regularly interspaced short palindromic repeats (CRISPR) editing, and identifying non-coding regulatory elements involved in stress responses [229].
With declining sequencing costs one of the main limiting factors, [223], GS is likely to become a key component of modern vegetable breeding programs, complementing MAS and enabling more efficient improvement of complex agronomic traits.
7. CRISPR/Cas9 Technology in Vegetable Breeding
The innovative breeding method called CRISPR-associated protein 9 (Cas9) enables precise and rapid enhancement of multiple crop traits, including yield, quality, resistance to diseases, tolerance to abiotic stresses, and nutritional value [230]. In the vegetable breeding CRISPR-Cas9 technology has the next improvements: improvement of biotic stress resistance, abiotic stress resistance improvement, herbicide resistance improvement, and quality improvement [230]. Compared with conventional breeding methods, this technology provides substantially greater precision and significantly reduces the time required for trait improvement [231].
7.1. Tomato
Tomato has emerged as an ideal model crop for evaluating CRISPR/Cas9 applications due to its significant economic importance, the feasibility of Agrobacterium-mediated transformation, and the successful development of new varieties without introducing foreign genes into its genome [230,232].
CRISPR-Cas9 technology through Agrobacterium-mediated transformation in tomato was modified for further traits: wiry phenotype [233], resistance against downy mildew [234,235], Fusarium wilt susceptibility [236], resistance to Botrytis cinerea [237], decrease in heat stress tolerance [238], decrease in chilling stress tolerance [239], decrease in drought stress tolerance [240], herbicide resistance [241], resistance against Phelipanche aegytiaca [242,243], introduction of desirable traits with morphology, flower number, fruit size and number, and ascorbic acid synthesis [244], fruit size, inflorescence branching, and plant architecture [245], yellow-coloured tomato [246], pink-coloured tomato [247], purple-coloured tomato [248], repression of genes controlling sugar and auxin metabolism [249], increase in gamma-aminobutyric acid content [250], lycopene content [251], MADS-box transcription factor regulating fruit ripening [252], activation and inhibition of fruit ripening [253], pectin degradation control [254], long shelf-life [255], jointless fruit stem [256], parthenocarpic [257,258,259], early flowering with simplified inflorescence architecture [260], day-length-sensitive flowering [261]. Furthermore, targeted mutation of the polygalacturonase (SlPG) gene using CRISPR/Cas9 led to a delay in fruit softening, thus extending postharvest firmness and shelf life [262]. To increase fruit firmness, [263] simultaneously knocked out the FIS1 and PL genes using CRISPR/Cas9, as these genes negatively affect fruit firmness. The obtained plants showed a significantly increased firmness of the fruits and better storage, without any negative impact on the basic traits of the fruit.
The results of increased lycopene content in tomatoes using CRISPR/Cas9 have been demonstrated in studies by Li et al. [251] and Ahmad et al. [264], who used the CRISPR/Cas9 technology to simultaneously mutate multiple genes involved in carotenoid metabolism in order to increase the concentration of lycopene, an antioxidant with important nutritional properties that affects the color of tomato fruit.
Traditional breeding techniques for major agronomic crops like tomato are unlikely to keep pace with emerging challenges, making the development of advanced, disease-resistant crop varieties essential for ensuring global food security. In this regard, advanced genome-editing tools such as CRISPR/Cas9 have become increasingly significant, as they have been applied in tomato to improve yield, enhance nutritional quality, and increase resistance to both biotic and abiotic stresses [265].
7.2. Pepper
To achieve long-term sustainability in production, it is essential to incorporate cutting-edge genetic and molecular strategies into pepper breeding and improvement programs. Innovations in genomic technologies—including CRISPR-Cas9 editing and integrated multi-omics approaches—are enabling the creation of varieties resilient to environmental stress [266].
Research conducted by [267] aimed to enhance pepper resistance to anthracnose diseases caused by Colletotrichum spp. through CaERF28 gene mutation. The resulting lines showed significantly improved resistance to infection compared to the wild type, and the T-DNA and marker-free mutants retained the edited phenotype in subsequent generations. Functional validation of CRISPR/Cas9 editing in pepper was performed using the phytoene desaturase (CaPDS) gene as a visual marker. Approximately 62.5% of the regenerated plants displayed albino or mosaic phenotypes due to loss of PDS protein function, confirming successful genome editing in Capsicum [268]. Parthenocarpic (seedless) pepper fruits were selectively generated by introducing the CRISPR/Cas9-RNP (ribonucleoprotein) complex into the protoplasts of the leaves of pepper lines, demonstrating potential applications for production under adverse climatic conditions [269].
Further, testing the delivery capability of the CRISPR/Cas9 system using Agrobacterium in two pepper cultivars provided the next example of how the system can be established for the future pepper breeding. The goal was to test the Agrobacterium-mediated CRISPR/Cas9 system for editing the CaMLO2 gene, which is associated with susceptibility to diseases [270].
On the other side, employing CRISPR genome editing to improve breeding program chili peppers, next stress responses were obtained: cold, high temperature, NaCl, and mannitol [271,272,273,274,275], drought and salt [276,277,278], chilling and salt [279], pathogen (R. solanacearum] and high temperature [280,281,282,283], pathogens (X. axonopodis, TMV and P. syringe) [284], R. solanacearum infection [285,286], high temperature [287,288,289], pathogen stress responses [290,291], heat shock and R. solanacearum[292], silencing of CaWRKY50-showed enhanced pepper resistance to C. scovillei [293], pathogen (P. capsici) [294], high temperature, salt and drought [295,296,297], pathogen (Pseudomonas syringae pv. tomato DC3000), drought and salt [298], pathogen (P. syringae pv. tomato DC3000) [299,300], water and salt [301].
With emerging pathogens and environmental challenges posing ongoing risks to crop production, the future of pepper improvement is being driven by the integration of breeding and molecular strategies [266].
7.3. Cabbage
The main goals of cabbage crop improvement include improving crop uniformity for maturity, increasing yield and yield-related traits, and developing climate-resilient varieties and hybrids that are resistant to biotic and abiotic stresses and rich in nutraceutical properties [302]. CRISPR/Cas technology has been widely applied for the modification of physiological and agronomic traits in Brassica species, including growth control, gene methylation and functional trait characterization [168].
The first CRISPR-Cas9 gene editing in cole vegetables was reported by Jansson [303], who used cabbage as a model plant and targeted the PsbS gene to create a Brassica deletion mutant.
Zhang et al. [228] used a modified CRISPR/Cas9 system to deactivate the BoBPM6 and BoDMR6 genes, resulting in broad-spectrum disease resistance in Brassica oleracea. This example illustrates how CRISPR/Cas9 technology can directly contribute to increased disease resistance in commercial cabbage varieties, an important trait for improving yield stability and reducing pesticide use.
In another research, the CRISPR/Cas9 system with an endogenous tRNA system was used to simultaneously target several genes in cabbage, such as: BoPDS (phytoene desaturase - related to pigment biosynthesis), BoSRK (S-receptor kinase - determinant of the autoincompatibility mechanism), and BoMS1 (gene related to male sterility) [304]. Multiple, inherited mutants were obtained in a single transformation, including lines that became self-compatible and male-sterile, which can be extremely useful for hybrid cabbage production [305]. This example shows how CRISPR/Cas9 can be used not only for functional analysis of genes but also for direct breeding of agronomic traits (e.g., removal of auto-incompatibility and fertility control).
Further, CRISPR/Cas9 ribonucleoprotein (RNP) complexes were introduced into cabbage protoplasts without the integration of foreign DNA. The approach enabled genome editing without transgenic sequences, which is important for both regulatory and consumer understanding [306]. The target genes were FRI and PDS, and the mutations were generated directly in the protoplasts. The approach represents an important breeding strategy, as it can result in varieties that are not legally classified as genetically modified organisms (GMOs) [306].
To illustrate another application, the CRISPR/Cas9 system has been successfully used to mutate the BoPDS gene in cabbage, causing clearly detectable albino phenotypes. The aim of the research was not to directly improve commercial properties but to serve as a technological foundation for later targeting of more important agronomic genes via the CRISPR/Cas9 system [304].
To improve black rot resistance in B. oleracea var. capitata CRISPR/Cas9 editing was used [307]. Black rot, a major disease of vegetable brassicas such as B. oleracea var. capitata, is caused by the bacterial pathogen Xanthomonas campestris pv. campestris (Xcc) [308,309]. Another study employed CRISPR/Cas9 to optimize the modification of a potential host susceptibility gene, BoSWEET15b (SWEET15b gene in B. oleracea), in the cabbage cultivar ‘Ohgane’ [310]. Recent progress in genomics, combined with CRISPR/Cas technologies, has facilitated faster identification, validation, and deployment of resistance genes for clubroot and black rot.
By applying CRISPR/Cas systems, researchers can better understand Brassica–pathogen interactions and the genetic processes involved in abiotic stress responses in Brassica plants.
7.4. Cucumber
Progress in transgenic cucumber breeding has been constrained by the absence of a reliable genetic transformation system, which restricts the use of tools such as CRISPR/Cas9 [311]. CRISPR/Cas9 is a precise gene editing method that enables targeted changes in the plant genome by introducing double-strand DNA breaks at specific loci, leading to mutations via non-homologous end joining (NHEJ) or homology-directed repair (HDR) pathways. This technology has proven highly useful for improving agronomic traits, such as disease resistance, flowering, or fruit quality traits [312].
Research using CRISPR/Cas9 mainly targets characteristics like resistance to viruses, fruit length, shoot growth, branching, and the production of gynoecious lines for hybrid breeding programs [313,314]. One strategy to optimize CRISPR/Cas9 systems is to utilize a stronger CsU6 promoter and include a GFP tag to aid in transformant detection [313].
Moreover, targeted mutations in Csa2G264590 led to a ten-fold increase in the number of spines on cucumber fruit compared to the wild type, which is an indication that structural traits can also be modified specifically [315]. Besides, mutating the MLO-type genes, or CsaMLO1, responsible for powdery mildew resistance, would be another potential strategy in breeding for resistance to mold, powdery mildew, or nematodes through mutation of the CsMS gene [316].
To develop viral resistance, CRISPR/Cas9 was used for the targeted mutation of the eIF4E (eukaryotic translation initiation factor 4E) gene in cucumber. This gene is known as the susceptibility (S) gene, which viruses use for infection. Gene editing resulted in plants resistant to several viral diseases, including cucumber vein yellowing virus (ipomovirus), zucchini yellow mosaic virus (potyvirus) and papaya ringspot virus-W (potyvirus). Homozygous progeny showed complete or significant resistance without other visible negative effects on plant development. The result shows how non-transgenic cucumbers with long-term virus resistance can be developed by using CRISPR/Cas9 [317].
Various breeding methods have been employed to achieve genetic enhancements in cucumbers, guided by specific breeding objectives. The primary goal of cucumber breeding programs is to develop varieties that are high-yielding, produce superior-quality fruits, and exhibit resistance to both biotic and abiotic stresses. The growing challenges in cultivation, along with rapid advancements in breeding technologies, have facilitated the development of cucumber varieties with improved resistance to diseases and tolerance to environmental stresses [318].
7.5. Lettuce
CRISPR/Cas9-based gene editing has been successfully demonstrated in lettuce, which is a model vegetable crop with a variety of characteristics. The CRISPR/Cas9 system was optimized to improve editing efficiency and stability by using the endogenous U6 promoter derived from the lettuce genome. The PHOT2 gene was targeted to check the efficiency of editing, i.e., the setting of gene efficiency and system optimization in lettuce [319]. However, the AtU6-26 promoter has been heterologously applied to drive sgRNAfor the CRISPR/Cas9 system in lettuce [320,321,322].
Nguyen et al. [322] use CRISPR gene editing for creating plants with novel phenotypes for faster plant production or marketing. The mentioned authors demonstrated the improved efficiency of a modified CRISPR/Cas9 construct through minimizing T-DNA positional effects and simplifying the process of transgene selection. In the study of 322], a CRISPR/Cas9 construct targeting the LsVAR2 gene, which controls leaf variegation, was used. Homozygous mutations produced an albino phenotype, whereas heterozygous plants displayed variegated or mottled leaves.
Choi et al. [323] developed the late-bolting lettuce varieties using the CRISPR/Cas9 system to introduce a precise single-base modification of the SOC1 gene, which regulates the onset of flowering in lettuce. Delayed bolting increases vegetative growth and may result in higher leaf yield.
Genetic engineering enables the direct manipulation of lettuce genes. By creating lines with enhanced tolerance to various environmental conditions, it can complement conventional breeding approaches, improving lettuce production efficiency and reducing losses caused by biotic and abiotic stresses [192]. Leveraging advances in molecular biology and genome sequencing, genetic engineering offers tools to increase both the yield and quality of lettuce. This approach has led to highly efficient and reliable transformation systems capable of directing nucleases to specific genetic sequences. In lettuce, transformation methods such as Agrobacterium tumefaciens-mediated techniques have shown strong success. These achievements have paved the way for genome editing approaches, including CRISPR/Cas9, which hold significant promise for enhancing performance and boosting yield. Consequently, genetic engineering in lettuce is poised to surpass traditional transformation techniques in effectiveness and efficiency [192].
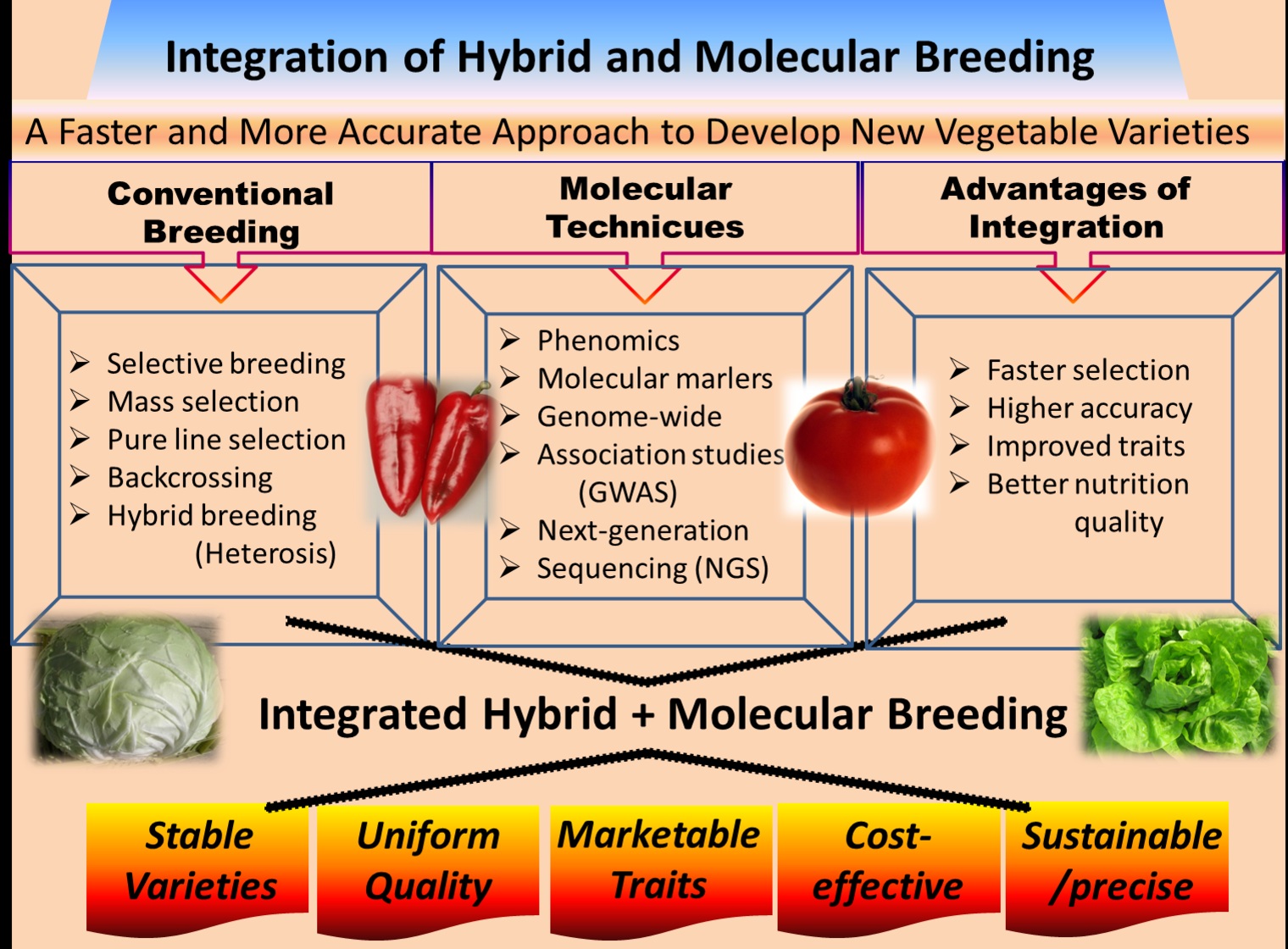
8. Advantages of Integrating Hybrid and Molecular Breeding of Vegetables
Traditional breeding is a method of selective breeding in which crops are chosen for propagation based on their superior performance. This approach is time-consuming and relies heavily on the observable characteristics (phenotype) of plants. By combining with hybrid breeding, where heterosis (hybrid vigor) is exploited, highly productive and high-quality vegetable hybrids can be developed more quickly [324,325]. However, selection based solely on phenotypic traits is often imprecise. Consequently, breeders began incorporating multiple disciplines of biology into plant breeding, leading to the development of modern breeding techniques [326].
Today, this incorporation or integration of hybrid and molecular breeding approaches in vegetables offers several key advantages, including:
- Improved stability and stress resistance of varieties [330],
- Optimized genetic resources and diversity [42],
This application of molecular markers in hybrid programs enables more precise introgression and pyramiding of beneficial traits, including resistance to diseases and stress conditions [20]. Combining molecular techniques with hybrid programs, it is easier to identify and use genetically diverse parental materials to create superior hybrids. Although initial costs in marker development may be higher, marker-assisted selection reduces the need for large field populations and extensive phenotypic evaluations, ultimately lowering overall costs and accelerating the process [20,51,324,325]. The integration of molecular methods enables more precise mapping and introduction of genes for resistance to biotic and abiotic stressors (diseases, drought, high temperatures), which increases the stability of vegetable yields [51].
Through germplasm assessment, gene discovery, and the development of molecular markers, researchers and plant breeders are collaborating to enhance breeding technologies aimed at producing superior cultivars. As extensive genomic data and advanced tools continue to emerge, deeper partnerships will be essential to advance next-generation strategies such as genome-based breeding by design to develop high-yielding, high-quality vegetable cultivars that are environmentally sustainable and well adapted to changing conditions [333].
9. Limitations and Challenges of Integrating Conventional Breeding and Molecular Techniques
Although the integration of hybrid and molecular breeding in vegetable production brings precision, accelerates selection, and potentially increases genetic progress, it also has several significant disadvantages, including:
- High complexity and technical demands [326],
- Challenges in translating theoretical approaches into practical applications, with limited applicability under field conditions [334],
- High costs and long duration [337],
Successful integration of hybridization and molecular techniques requires a multidisciplinary approach including classical genetics, molecular biology, statistics, and agronomy [28,326]. The lack of multidisciplinary teams can limit the possibilities for developing new cultivars or lines that successfully combine the aforementioned methods. The integration of molecular methods and hybrid breeding requires significantly greater technological and laboratory resources than conventional selection, including sophisticated tools for genotyping, gene analysis, and bioinformatic support, which can be an obstacle for smaller research teams or countries with more limited resources [327,339]. Furthermore, although molecular methods speed up the process, the initial investment is high, primarily in equipment, laboratory infrastructure, and experts for genetic analysis [340]. To achieve optimal profitability of such investments, long-term with a broad base of breeding programs must be developed.
Despite the fact that molecular methods can theoretically accelerate selection and improve accuracy in recognizing useful genes, their effectiveness in practice is often limited: it is difficult to predict how marker-related traits will behave in different climates and agroecosystems, and there is often a lack of clear strategies for transferring laboratory results into cultivated varieties with a realistically increased yield [334]. Marker-assisted selection (MAS) is a more efficient strategy for improving traits that have low heritability or are costly and labor-intensive to measure. However, it is most effective when applied to a limited number of quantitative traits that account for a significant proportion of the variation within a population [341].
Molecular techniques such as marker-assisted selection (MAS) have limitations, especially for complex, polygenically controlled traits (e.g., stress tolerance, nutritional quality). Marker-assisted selection is often inefficient for traits controlled by numerous small-effect genes, and precise mapping of all relevant quantitative traits is demanding and expensive. On the other hand, hybridization and selection with molecular markers can lead to a reduction in genetic variability if they focus only on a few high-potential genotypes. A lack of genetic variability can lead to increased susceptibility to diseases and stress factors in changing growing conditions [20,333,342].
These limitations must be addressed through appropriate funding, education, and multidisciplinary collaboration in order to realize the full potential of these approaches in vegetable production.
10. Conclusions
The integration of new technologies allows breeders to plan and develop inbred lines containing different combinations of desirable traits, based on predefined molecular profiles. Carefully selected parental lines, both genetically and phenotypically, then enable the development of F1 hybrids with optimally complementary traits. Advanced technological approaches thus accelerate the development of elite inbred lines and commercially valuable F1 hybrids with complex sets of traits, such as specific combinations of genes or QTL which correspond to specific growing conditions, that a significant reduction in breeding costs and time.
Similar to the marker-assisted selection (MAS), other modern technologies have also encouraged vegetable breeders to approach breeding with greater precision. In the past, breeders generally focused on one or a few traits that they improved over a long period of time. Conventional breeding techniques involved the development of numerous inbred lines and a detailed phenotypic evaluation, as well as testing innumerable experimental F1 hybrids before selecting those with market value potential.
The development of more advanced SNP chips will also enable early-stage genotyping of breeding material for a large number of traits and thus significantly reduce the time required for the identification of elite inbred lines and the development of desirable hybrid combinations.
If the breeder’s resources allow the full breadth of integration of conventional and molecular breeding to be fully exploited, genetic progress and the faster development of higher-quality vegetable varieties are proportionally increased compared to either of these approaches alone. The true values of integration are a combination of speed, precision, saving time and resources, greater accuracy of selection, and more efficient use of plant genetic resources.
Author Contributions
Conceptualization, J.Č., S.Z., G.T., and D.M.; methodology, J.Č. and S.Z.; validation, G.T., and J.M.; formal analysis, J.Č., G.T., Đ.V., and A.I.; investigation, J.Č.,S.Z.; G.T., and B.A.; resources, S.Z.; and G.T.; data curation, J.Č. and S.Z.; writing—original draft preparation, J.Č.; writing—review and editing, S.Z., G.T., D.M., B.A., Đ.V., J.M., and A.I.; visualization, B.A., and J.M.; supervision, D.M., Đ.V., and A.I. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was supported by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia, grant numbers: 451-03-33/2026-03/200032 and 451-03-34/2026-03/200117.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WHO | World Health Organization |
| MAS | Marker-Assisted Selection |
| MABC | Marker-Assisted Backcrossing |
| MARS DNA |
Marker-Assisted Recurrent Selection Deoxyribonucleic Acid |
| RAPD | Random Amplified Polymorphic DNA |
| ISSR | Inter-Simple Sequence Repeats |
| AFLP | Amplified Fragment Length Polymorphism |
| RFLP | Restriction Fragment Length Polymorphism |
| SNP SSR |
Single Nucleotide Polymorphism Simple Sequence Repeat |
| SRAP QTL |
Sequence-Related Amplified Polymorphism Quantitative Trait Loci |
| LD | Linkage Disequilibrium |
| KASP | Kompetitive Allele Specific PCR |
| SCAR | Sequence Characterized Amplified Region |
| CAPS | Cleaved Amplified Polymorphic Sequences |
| PCR dCAPS InDel ERPAR EST-SSR SSAP GS GEBVs GCA SCA DH HTPP AI gLMs sgRNA CRISPR CRISPR/Cas9 |
Polymerase Chain Reaction Derived Cleaved Amplified Polymorphic Sequences Insertion-Deletion Extended Random Primer Amplified Region Expressed Sequence Tag-Simple Sequence Repeats Sequence-specific Amplified Polymorphism Genomic Selection Genomic Estimated Breeding Values General Combining Ability Specific Combining Ability Doubled Haploid High-Throughput Phenotyping Artificial Intelligence Genomic Language Models Single Guide Ribonucleic Acid Clustered Regularly Interspaced Short Palindromic Repeats CRISPR- associated Protein 9 |
| RNP GMOs NHEJ HDR |
Ribonucleoprotein Genetically Modified Organisms Non-Homologous End Joining Homology-Directed Repair |
References
- Abuyusuf, M.; Nath, U. K.; Kim, H.-T.; Islam, M. R.; Park, J.-I.; Nou, I.-S. Molecular Markers Based on Sequence Variation in BoFLC1.C9 for Characterizing Early- and Late-Flowering Cabbage Genotypes. BMC Genet. 2019, 20, 42. [Google Scholar] [CrossRef]
- Dias, J. S. Nutritional Quality and Health Benefits of Vegetables: A Review. Food Nutr. Sci. 2012, 03, 1354–1374. [Google Scholar] [CrossRef]
- Dias, J. C. da S. Dias, J. C. da S. Nutritional Quality and Health Benefits of Vegetables. In Emerging Trends in Disease and Health Research Vol. 4; Book Publisher International (a part of SCIENCEDOMAIN International). 2022, 7–35. [CrossRef]
- Vojnović, Đ; Ilin, Ž; Kostić, A. Ž; Adamović, B.; Kilibarda, S.; Pezo, L.; Popović-Đorđević, J. Phytochemical Composition, Antioxidant Potential, and Bulb Mass of Differently Colored Onions under Distinct Sowing Methods. Qual. Assur. Saf. Crops Foods 2025, 17, 108–126. [Google Scholar] [CrossRef]
- Drewnowski, A.; Rehm, C. D. Energy Intakes of US Children and Adults by Food Purchase Location and by Specific Food Source. Nutr. J. 2013, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Boye, J.; Zare, F.; Pletch, A. Pulse Proteins: Processing, Characterization, Functional Properties and Applications in Food and Feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Pulses: Nutritious Seeds for a Sustainable Future; Food & Agriculture Organization of the United Nations (FAO): Rome, Italy, 2016. [Google Scholar] [CrossRef]
- Slavin, J. L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Batoola, U.; Nawaz, R.; Ahmad, S.; Irfan, A.; Irshad, M. A.; Arshad, M.; Salamatullah, A. M.; Shah, A. N.; Dauelbaiti, M.; Butt, A. R.; Bourhiak, M. Physicochemical properties and antioxidant potentials of leafy vegetables grown under different agroecological conditions. International Journal of Food Properties 2024, 2024(27), 1046–1063. [Google Scholar] [CrossRef]
- Soujanya, K.; Kumari, B. A.; Jyothsna, E. Evaluation of Nutritional Quality Characteristics of Selected Uncultivated Green Leafy Vegetables. Ann. Phytomed. 2024, 13. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Green Leafy Vegetables (GLVs) as Nutritional and Preventive Agents Supporting Metabolism. Metabolites 2025, 15 502. [Google Scholar] [CrossRef]
- Ramya, V.; Patel, P. Health Benefits of Vegetables. International Journal of Chemical Studies 2019, 7, 82–87. [Google Scholar]
- Ebabhi, A.; Adebayo, R. Nutritional Values of Vegetables. In Vegetable Crops - Health Benefits and Cultivation; IntechOpen, 2022. [CrossRef]
- Devirgiliis, C.; Guberti, E.; Mistura, L.; Raffo, A. Effect of Fruit and Vegetable Consumption on Human Health: An Update of the Literature. Foods 2024, 13, 3149. [Google Scholar] [CrossRef] [PubMed]
- Silva Dias, J. World Importance, Marketing and Trading of Vegetables. Acta Hortic. 2011, 921, 153–169. [Google Scholar] [CrossRef]
- Ramya, V.; Patel, P. Health Benefits of Vegetables. International Journal of Chemical Studies 2019, 7, 82–87. [Google Scholar]
- Ramasamy, S.; Lin, M.-Y.; Wu, W.-J.; Wang, H.-I.; Sotelo-Cardona, P. Evaluating the Potential of Protected Cultivation for Off-Season Leafy Vegetable Production: Prospects for Crop Productivity and Nutritional Improvement. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Zandalinas, S. I.; Balfagón, D.; Gómez-Cadenas, A.; Mittler, R. Plant Responses to Climate Change: Metabolic Changes under Combined Abiotic Stresses. J. Exp. Bot. 2022, 73, 3339–3354. [Google Scholar] [CrossRef]
- Sánchez-Bermúdez, M.; Del Pozo, J. C.; Pernas, M. Effects of Combined Abiotic Stresses Related to Climate Change on Root Growth in Crops. Front. Plant Sci. 2022, 13, 918537. [Google Scholar] [CrossRef]
- Weiss, J.; Gruda, N. S. Novel Breeding Techniques and Strategies for Enhancing Greenhouse Vegetable Product Quality. Agronomy (Basel) 2025a, 15, 207. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S. G. Ecological Intensification: Harnessing Ecosystem Services for Food Security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S. J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Tigchelaar, M.; Battisti, D. S.; Naylor, R. L.; Ray, D. K. Future Warming Increases Probability of Globally Synchronized Maize Production Shocks. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 6644–6649. [Google Scholar] [CrossRef]
- Zörb, C.; Geilfus, C.-M.; Dietz, K.-J. Salinity and Crop Yield. Plant Biol. (Stuttg.) 2019, 21 Suppl 1 (S1), 31–38. [Google Scholar] [CrossRef] [PubMed]
- Mehra, P.; Patel, S. S.; Pansare, U. D. Application of Molecular Markers in Vegetable Improvement : A Review. Plant Arch. 2024, 24. [Google Scholar] [CrossRef]
- Botstein, D.; White, R. L.; Skolnick, M. Construction of a Genetic Linkage Map in Man Using Restriction Fragment Length Pol-Ymorphisms. Am J Hum Genet 1980, 32, 314–331. [Google Scholar] [PubMed]
- Ceccarelli, S. Efficiency of Plant Breeding. Crop Sci. 2015, 55, 87–97. [Google Scholar] [CrossRef]
- He, R.; Ju, J.; Liu, K.; Song, J.; Zhang, S.; Zhang, M.; Hu, Y.; Liu, X.; Li, Y.; Liu, H. Technology of Plant Factory for Vegetable Crop Speed Breeding. Front. Plant Sci. 2024, 15, 1414860. [Google Scholar] [CrossRef]
- Williams, J. G.; Kubelik, A. R.; Livak, K. J.; Rafalski, J. A.; Tingey, S. V. DNA Polymorphisms Amplified by Arbitrary Primers Are Useful as Genetic Markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Sun, L.; Lai, M.; Ghouri, F.; Nawaz, M. A.; Ali, F.; Baloch, F. S.; Nadeem, M. A.; Aasim, M.; Shahid, M. Q. Modern Plant Breeding Techniques in Crop Improvement and Genetic Diversity: From Molecular Markers and Gene Editing to Artificial Intelligence-A Critical Review. Plants 2024, 13, 2676. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. AFLP: A New Technique for DNA Fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, S. S.; Ingle, K. P.; Das, U.; Ramteke, P. W.; Kurubar, A. R.; Shukla, P. K.; Geethika, P.; Madala, R. S. Molecular Marker-Assisted Selection in Plant Breeding. In Advances in Plant Breeding Strategies; Springer Nature Switzerland: Cham, 2024; pp. 95–111. [Google Scholar] [CrossRef]
- Fetisov, I.; Eizikovich, O.; Charles Diouf, D.; Romanova, E.; Kezimana, P. Advancements in Molecular Breeding Techniques for Soybeans. Plants 2025, 15, 5. [Google Scholar] [CrossRef]
- Charmet, G.; Robert, N.; Perretant, M. R.; Gay, G.; Sourdille, P.; Groos, C.; Bernard, S.; Bernard, M. Marker-Assisted Recurrent Selection for Cumulating Additive and Interactive QTLs in Recombinant Inbred Lines. Theor. Appl. Genet. 1999, 99, 1143–1148. [Google Scholar] [CrossRef]
- Collard, B. C. Y.; Mackill, D. J. Marker-Assisted Selection: An Approach for Precision Plant Breeding in the Twenty-First Century. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 557–572. [Google Scholar] [CrossRef]
- Jiang, G.L. Molecular Markers and Marker-Assisted Breeding in Plants. In Plant Breeding from Laboratories to Fields; InTech, 2013. [CrossRef]
- Thakur, M.; Thakur, H. Vegetable Breeding for Organic Farming: Opportunities and Perspectives. Genetika 2025, 57, 443–456. [Google Scholar] [CrossRef]
- Ebert, A. W. The Role of Vegetable Genetic Resources in Nutrition Security and Vegetable Breeding. Plants 2020, 9, 736. [Google Scholar] [CrossRef]
- Červenski, J.; Slobodan, F.V.; Adamovic, A.B.; Vojnovic, D.D.; Zec, S.S.D. Possibilities of Cabbage Production under Climatic Changes. J. Agric. Sci. Belgrade 2025, 70, 233–247. [Google Scholar] [CrossRef]
- Weiss, J.; Gruda, N. S. Enhancing Nutritional Quality in Vegetables through Breeding and Cultivar Choice in Protected Cultivation. Sci. Hortic. (Amsterdam) 2025b, 339, 113914. [Google Scholar] [CrossRef]
- Tabasum, S.; Nazir, G.; Hussain, K.; Gautam, A.; Mushtaq, F.; Nabi, A.; Ahsan, S. Climate Smart Breeding of Vegetable Crops : A Review. Plant Arch. 2025, 25. [Google Scholar] [CrossRef]
- Ghuge, M. B.; Mirza, A. Vegetable Breeding Strategies. Asian J. Agric. Hortic. Res. 2021, 28–37. [Google Scholar] [CrossRef]
- Singh, H.; Sekhon, B. S.; Kumar, P.; Dhall, R. K.; Devi, R.; Dhillon, T. S.; Sharma, S.; Khar, A.; Yadav, R. K.; Tomar, B. S.; Ntanasi, T.; Sabatino, L.; Ntatsi, G. Genetic Mechanisms for Hybrid Breeding in Vegetable Crops. Plants 2023, 12, 2294. [Google Scholar] [CrossRef]
- Rauf, S.; Basharat, T.; Iqbal, Z. Hybrid Breeding in Vegetable Crops. In Sustainable and Innovative Vegetable Production in times of Climate Change; Springer Nature Singapore: Singapore, 2025; pp. 403–420. [Google Scholar] [CrossRef]
- Riggs, T. J. Breeding F1hybrid Varieties of Vegetables. J. Hortic. Sci. 1988, 63, 369–382. [Google Scholar] [CrossRef]
- Carvalho, F.; Lahlou, R. A.; Silva, L. R. Exploring Bioactive Compounds from Fruit and Vegetable By-Products with Potential for Food and Nutraceutical Applications. Foods 2025, 14, 3884. [Google Scholar] [CrossRef]
- Pidigam, S.; Geetha, A.; Nagaraju, K.; Pandravada, S. R.; Khan, M. S.; Rajasekhar, M.; Sivraj, N.; Vishnukiran, T. Breeding Approaches for the Development of Nutraceutical Vegetables: A Review. Ann. Phytomed. 2022, 11. [Google Scholar] [CrossRef]
- Causse, M.; Saliba-Colombani, V.; Lecomte, L.; Duffé, P.; Rousselle, P.; Buret, M. QTL Analysis of Fruit Quality in Fresh Market Tomato: A Few Chromosome Regions Control the Variation of Sensory and Instrumental Traits. J. Exp. Bot. 2002, 53, 2089–2098. [Google Scholar] [CrossRef] [PubMed]
- Capel, C.; Yuste-Lisbona, F. J.; López-Casado, G.; Angosto, T.; Heredia, A.; Cuartero, J.; Fernández-Muñoz, R.; Lozano, R.; Capel, J. QTL Mapping of Fruit Mineral Contents Provides New Chances for Molecular Breeding of Tomato Nutritional Traits. Theor. Appl. Genet. 2017, 130, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Červenski, J.; Vlajić, S.; Ignjatov, M.; Tamindžić, G.; Zec, S. Agroclimatic Conditions for Cabbage Production. Ratar. Povrt. 2022, 59, 43–50. [Google Scholar] [CrossRef]
- Yu, D.; Gu, X.; Zhang, S.; Dong, S.; Miao, H.; Gebretsadik, K.; Bo, K. Molecular Basis of Heterosis and Related Breeding Strategies Reveal Its Importance in Vegetable Breeding. Hortic. Res. 2021, 8, 120. [Google Scholar] [CrossRef]
- Sahu, D.; Tripathy, S. K.; Sahoo, B.; Nayak, L.; Swarajya, R.; Dalei, D. Role of Molecular Markers in Vegetables Improvement. Journal of Pharmacognosy and Phytochemistry 2019, 8, 456–460. [Google Scholar]
- Singh, J.; Dhall, R. K.; Vikal, Y. Genetic Diversity Studies in Indian Germplasm of Pea (Pisum Sativum L.) Using Morphological and Microsatellite Markers. Genetika 2021, 53, 473–491. [Google Scholar] [CrossRef]
- Kondić-Špika, A.; Kobiljski, B. Biotechnology in modern breeding and agriculture. In Book of the Proceedings, International Conference on BioScience: Biotechnology and Biodiversity,” Step in the Future”. 2012, 201-210. https://fiver.ifvcns.rs/handle/123456789/4822.
- Singh, H. P.; Raigar, O. P.; Chahota, R. K. Estimation of Genetic Diversity and Its Exploitation in Plant Breeding. Bot. Rev. 2022, 88, 413–435. [Google Scholar] [CrossRef]
- Salgotra, R. K.; Chauhan, B. S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes (Basel) 2023, 14, 174. [Google Scholar] [CrossRef]
- Bidyananda, N.; Jamir, I.; Nowakowska, K.; Varte, V.; Vendrame, W. A.; Devi, R. S.; Nongdam, P. Plant Genetic Diversity Studies: Insights from DNA Marker Analyses. Int. J. Plant Biol. 2024, 15, 607–640. [Google Scholar] [CrossRef]
- Vázquez, H.; Jimenez, F. L.; Vico, J. M. C. RAPD Fingerprinting of Pepper (Capsicum Annuum L.) Breeding Lines. Capsicum and Eggplant Newslette 1996, 15, 37–40. [Google Scholar]
- Dijkhuizen, A.; Kennard, W. C.; Havey, M. J.; Staub, J. E. RFLP Variation and Genetic Relationships in Cultivated Cucumber. Euphytica 1996, 90, 79–87. [Google Scholar] [CrossRef]
- Villand, J.; Skroch, P. W.; Lai, T.; Hanson, P.; Kuo, C. G.; Nienhuis, J. Genetic Variation among Tomato Accessions from Primary and Secondary Centers of Diversity. Crop Sci. 1998, 38, 1339–1347. [Google Scholar] [CrossRef]
- Levi, A.; Thomas, C. E.; Newman, M.; Reddy, O. U. K.; Zhang, X.; Xu, Y. ISSR and AFLP Markers Differ among American Watermelon Cultivars with Limited Genetic Diversity. J. Am. Soc. Hortic. Sci. 2004, 129, 553–558. [Google Scholar] [CrossRef]
- Muminović, J.; Merz, A.; Melchinger, A. E.; Lübberstedt, T. 3 Genetic Structure and Diversity among Radish Varieties as Inferred from AFLP and ISSR Analyses. Journal of the American Society for Horticultural Science 2005, 130, 79–87. [Google Scholar] [CrossRef]
- Blanca, J.; Pons, C.; Montero-Pau, J.; Sanchez-Matarredona, D.; Ziarsolo, P.; Fontanet, L.; Fisher, J.; Plazas, M.; Casals, J.; Rambla, J. L.; Riccini, A.; Pombarella, S.; Ruggiero, A.; Sulli, M.; Grillo, S.; Kanellis, A.; Giuliano, G.; Finkers, R.; Cammareri, M.; Grandillo, S.; Mazzucato, A.; Causse, M.; Díez, M. J.; Prohens, J.; Zamir, D.; Cañizares, J.; Monforte, A. J.; Granell, A. European Traditional Tomatoes Galore: A Result of Farmers’ Selection of a Few Diversity-Rich Loci. J. Exp. Bot. 2022, 73, 3431–3445. [Google Scholar] [CrossRef]
- Ibitoye, D.; Akin-Idowu, P. Marker-assisted-selection (MAS): A fast track to increase genetic gain in horticultural crop breeding. African Journal of Biotechnology 2013, 10, 11333–11339. [Google Scholar] [CrossRef]
- Ahmar, S.; Gill, R. A.; Jung, K.-H.; Faheem, A.; Qasim, M. U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef]
- Simko, I.; Jia, M.; Venkatesh, J.; Kang, B.-C.; Weng, Y.; Barcaccia, G.; Lanteri, S.; Bhattarai, G.; Foolad, M. R. Genomics and Marker-Assisted Improvement of Vegetable Crops. CRC Crit. Rev. Plant Sci. 2021, 40, 303–365. [Google Scholar] [CrossRef]
- Lema, M. Marker Assisted Selection in Comparison to Conventional Plant Breeding: Review Article. Agric. Res. Technol. 2018, 14. [Google Scholar] [CrossRef]
- Collard, B. C. Y.; Jahufer, M. Z. Z.; Brouwer, J. B.; Pang, E. C. K. An Introduction to Markers, Quantitative Trait Loci (QTL) Mapping and Marker-Assisted Selection for Crop Improvement: The Basic Concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Guberac, S. Strossmayer University of Osijek, Faculty of Agriculture in Osijek, Kralja Petra Svačića 1d, 31000 Osijek, Croatia; Petrović, S.; Guberac, V.; Marić, S. Molecular Markers in Tomato Breeding. Poljoprivreda 2015, 21, 55–60. [Google Scholar] [CrossRef]
- Xu, Y.; Crouch, J. H. Marker-assisted Selection in Plant Breeding: From Publications to Practice. Crop Sci. 2008, 48, 391–407. [Google Scholar] [CrossRef]
- Bernardo, R. Molecular Markers and Selection for Complex Traits in Plants: Learning from the Last 20 Years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef]
- Ansari, A. Molecular Markers in Vegetable Improvement. Horticultural Biotechnology Research 2015, 1, 5–10. [Google Scholar] [CrossRef]
- Kalendar, R.; Grob, T.; Regina, M. IRAP and REMAP: two new retrotransposon-based DNA fingerprinting techniques. Theor Appl Genet. 1999, 98, 704–711. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity Arrays: A Solid State Technology for Sequence Information Independent Genotyping. Nucleic Acids Res. 2001, 29, E25. [Google Scholar] [CrossRef]
- Kulus, D. Genetic Diversity for Breeding Tomato. In Cash Crops; Springer International Publishing: Cham, 2022; pp. 505–521. [Google Scholar] [CrossRef]
- Khazaei, H.; Madduri, A. Role of Tomato Wild Relatives in Breeding Disease-Free Varieties. GenResJ 2022, 3, 64–73. [Google Scholar] [CrossRef]
- Taylor, B. Biosystematics of the tomato. In: Atherton, J. G., and J. Rudich, editors. The Tomato Crop : A Scientific Basis for Improvement. 1st ed. Springer Netherlands, 1986. [CrossRef]
- John Goodstal, F.; Kohler, G. R.; Randall, L. B.; Bloom, A. J.; St Clair, D. A. A Major QTL Introgressed from Wild Lycopersicon Hirsutum Confers Chilling Tolerance to Cultivated Tomato (Lycopersicon Esculentum). Theor. Appl. Genet. 2005, 111, 898–905. [Google Scholar] [CrossRef]
- Mladenov, V.; Boćanski, Ј; Banjac, B. Oplemenjivanje Biljaka: Praktikum; Novi Sad; 2021.1-68.
- Shrestha, S.; Subedi, S.; Jiban, J. Marker-Assisted Selection: A Smart Biotechnological Strategy for Modern Plant Breeding. Peruvian Journal of Agronomy 2020, 4, 104–120. [Google Scholar] [CrossRef]
- Gokidi, Y. U.; Bhanu, A. N.; Singh, M. N. Marker Assisted Recurrent Selection: An Overview. Adv Life Sci 2016, 5, 6493–6499. [Google Scholar]
- Beyene, Y.; Semagn, K.; Mugo, S.; Prasanna, B. M.; Tarekegne, A.; Gakunga, J.; Sehabiague, P.; Meisel, B.; Oikeh, S. O.; Olsen, M.; Crossa, J. Performance and Grain Yield Stability of Maize Populations Developed Using Marker-Assisted Recurrent Selection and Pedigree Selection Procedures. Euphytica 2016, 208, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Cholin, S. S.; Kulkarni, C. C. Prospects of Marker-Assisted Recurrent Selection: Current Insights and Future Implications. Trop. Plant Biol. 2023. [Google Scholar] [CrossRef]
- Na Jinda, A.; Nikornpun, M.; Jeeatid, N.; Thumdee, S.; Thippachote, K.; Pusadee, T.; Kumchai, J. Marker-Assisted Selection of Male-Sterile and Maintainer Line in Chili Improvement by Backcross Breeding. Horticulturae 2023, 9, 357. [Google Scholar] [CrossRef]
- Thomas, W. J. W.; Amas, J. C.; Dolatabadian, A.; Huang, S.; Zhang, F.; Zandberg, J. D.; Neik, T. X.; Edwards, D.; Batley, J. Recent Advances in the Improvement of Genetic Resistance against Disease in Vegetable Crops. Plant Physiol. 2024, 196, 32–46. [Google Scholar] [CrossRef]
- Husaini, A. M. High-Value Pleiotropic Genes for Developing Multiple Stress-Tolerant Biofortified Crops for 21st-Century Challenges. Heredity (Edinb.) 2022, 128, 460–472. [Google Scholar] [CrossRef]
- Suresh, S.; Malathi, D. Gene Pyramiding for Biotic Stress Tolerance In Crop Plants. Wkly. Sci. Res. J 2013, 1, 1–14. [Google Scholar]
- Bai, Y.; Kissoudis, C.; Yan, Z.; Visser, R. G. F.; van der Linden, G. Plant Behaviour under Combined Stress: Tomato Responses to Combined Salinity and Pathogen Stress. Plant J. 2018, 93, 781–793. [Google Scholar] [CrossRef]
- Kage, U.; Kumar, A.; Dhokane, D.; Karre, S.; Kushalappa, A. C. Functional Molecular Markers for Crop Improvement. Crit. Rev. Biotechnol. 2016, 36, 917–930. [Google Scholar] [CrossRef]
- Hasan, M. M.; Rafii, M. Y.; Ismail, M. R.; Mahmood, M.; Rahim, H. A.; Alam, M. A.; Ashkani, S.; Malek, M. A.; Latif, M. A. Marker-Assisted Backcrossing: A Useful Method for Rice Improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.; Sood, A.; Hussain, W.; Kaldate, R.; Sharma, T. R.; Gill, R. K.; Kumar, S.; Singh, S. Chapter 6-Gene Pyramiding and Multiple Character Breeding. In Lentils; Singh, M; Academic Press: Cambridge, MA, USA, 2019, 83–124.
- Dormatey, R.; Sun, C.; Ali, K.; Coulter, J. A.; Bi, Z.; Bai, J. Gene Pyramiding for Sustainable Crop Improvement against Biotic and Abiotic Stresses. Agronomy (Basel) 2020, 10, 1255. [Google Scholar] [CrossRef]
- Verlaan, M. G.; Hutton, S. F.; Ibrahem, R. M.; Kormelink, R.; Visser, R. G. F.; Scott, J. W.; Edwards, J. D.; Bai, Y. The Tomato Yellow Leaf Curl Virus Resistance Genes Ty-1 and Ty-3 Are Allelic and Code for DFDGD-Class RNA-Dependent RNA Polymerases. PLoS Genet. 2013, 9, e1003399. [Google Scholar] [CrossRef] [PubMed]
- Djian-Caporalino, C.; Palloix, A.; Fazari, A.; Marteu, N.; Barbary, A.; Abad, P.; Sage-Palloix, A.-M.; Mateille, T.; Risso, S.; Lanza, R.; Taussig, C.; Castagnone-Sereno, P. Pyramiding, Alternating or Mixing: Comparative Performances of Deployment Strategies of Nematode Resistance Genes to Promote Plant Resistance Efficiency and Durability. BMC Plant Biol. 2014, 14, 53. [Google Scholar] [CrossRef]
- Mapari, A. R.; Mehandi, S. Enhancing Crop Resilience: Advances and Challenges in Marker-Assisted Selection for Disease Resistance. J. Adv. Biol. Biotechnol. 2024, 27, 569–580. [Google Scholar] [CrossRef]
- Ijaz, U.; Sudheesh, S.; Kaur, S.; Sadeque, A.; Bariana, H.; Bansal, U.; Adhikari, K. Mapping of Two New Rust Resistance Genes Uvf-2 and Uvf-3 in Faba Bean. Agronomy (Basel) 2021, 11, 1370. [Google Scholar] [CrossRef]
- Purkaystha, S.; Das, P.; Rashmi, K.; Rout, S.; Nanda, S. Advances in Genetic Mapping of Loci Governing Disease Resistance in Plants. In Biotechnological Advances for Disease Tolerance in Plants; Springer Nature Singapore: Singapore, 2024; pp. 1–27. [Google Scholar]
- Zec, S.; Tamindžić, G.; Azizbekian, S.; Ignjatov, M.; Danojević, D.; Červenski, J.; Vlajić, S.; Vojnović, Đ; Banjac, B. Foliar Application of Ca-Based Fertilizers (Conventional vs. Nanofertilizers): Effects on Fruit Traits, Seed Quality Parameters and Initial Plant Growth of Tomato Genotypes. Horticulturae 2025, 11, 1303. [Google Scholar] [CrossRef]
- Glogovac, S.; Takac, A.; Brbaklic, L.; Trkulja, D.; Cervenski, J.; Gvozdanovic-Varga, J.; Popovic, V. 10.5937/Ratpov50-4344 = Molecular Evaluation of Genetic Variability in Tomato (Lycopersicon Esculentum Mill.) Genotypes by Microsatellite Markers. Ratar. Povrt. 2013, 50, 1–5. [Google Scholar] [CrossRef]
- Ekbiç, E.; Okay, C. Ö. Assessment of Elite Pepper Breeding Lines Using Molecular Markers. Plant Biotechnol. Rep. 2024, 18, 515–524. [Google Scholar] [CrossRef]
- Aktas, H.; Abak, K.; Sensoy, S. Genetic Diversity in Some Turkish Pepper (Capsicum Annuum L.) Genotypes Revealed by AFLP Analyses. African Journal of Biotechnology 2009, 8, 4378–4386. [Google Scholar]
- Bozkalfa, M. K.; Kaygısız Aşcıoğlu, T.; Eşiyok, D. Biber Genotiplerinin Genetik Çeşitliliklerinin Srap Markörleri Kullanılarak Belirlenmesi. ANADOLU J. OF AGRIC. SCI. 2017, 32, 321–329. [Google Scholar] [CrossRef]
- Yildiz, M.; Koçak, M.; Nadeem, M. A.; Cavagnaro, P.; Barboza, K.; Baloch, F. S.; Argün, D.; Keleş, D. Genetic Diversity Analysis in the Turkish Pepper Germplasm Using iPBS Retrotransposon- Based Markers. Turk J. Agric. For. 2020, 44, 1–14. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.; Buirchell, B.; Yan, G. Development of a DNA Marker Tightly Linked to Low-Alkaloid Gene Iucundus in Narrow-Leafed Lupin (Lupinus Angustifolius L.) for Marker-Assisted Selection. Crop Pasture Sci. 2011, 62, 218–224. [Google Scholar] [CrossRef]
- Barka, G. D.; Lee, J. Molecular Marker Development and Gene Cloning for Diverse Disease Resistance in Pepper (Capsicum AnnuumL.): Current Status and Prospects. Plant Breed. Biotechnol. 2020, 8, 89–113. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yoneda, H.; Hosokawa, M.; Miwa, T.; Yazawa, S. Application of Marker-Assisted Selection in Breeding of a New Fresh Pepper Cultivar (Capsicum Annuum) Containing Capsinoids, Low-Pungent Capsaicinoid Analogs. Sci. Hortic. (Amsterdam) 2014, 165, 242–245. [Google Scholar] [CrossRef]
- Paran, I.; van der Voort, J. R.; Lefebvre, V.; Jahn, M.; Landry, L.; van Schriek, M.; Tanyolac, B.; Caranta, C.; Chaim, A. B.; Livingstone, K.; Palloix, A.; Peleman, J. An Integrated Genetic Linkage Map of Pepper (Capsicum Spp.). Mol. Breed. 2004, 13, 251–261. [Google Scholar] [CrossRef]
- Hyeon, N. S.; Woong, Y. J.; Cheorl, K. B.; Byung-Dong, K. Selection of Parental Lines for Hot Pepper Mapping Population Using RFLP and AFLP Analyses. Hortic Environ Biotechnol 1997, 38, 693–697. [Google Scholar]
- Rêgo, E. R.; Rego, M. M.; Farias-Filho, L. P. Genetic Diversity in Pepper (Capsicum Spp.) by Rapd Marker. Acta Hortic. 2011, 918, 341–347. [Google Scholar] [CrossRef]
- Lefebvre, V.; Goffinet, B.; Chauvet, J. C.; Caromel, B.; Signoret, P.; Brand, R.; Palloix, A. Evaluation of Genetic Distances between Pepper Inbred Lines for Cultivar Protection Purposes: Comparison of AFLP, RAPD and Phenotypic Data. Theor. Appl. Genet. 2001, 102, 741–750. [Google Scholar] [CrossRef]
- Tam, S. M.; Mhiri, C.; Vogelaar, A.; Kerkveld, M.; Pearce, S. R.; Grandbastien, M.-A. Comparative Analyses of Genetic Diversities within Tomato and Pepper Collections Detected by Retrotransposon-Based SSAP, AFLP and SSR. Theor. Appl. Genet. 2005, 110, 819–831. [Google Scholar] [CrossRef]
- Dhaliwal, M.S.; Abhay, Y.; S., K. J. Molecular Characterization and Diversity Analysis in Chilli Pepper Using Simple Sequence Repeats (SSR) Markers. Afr. J. Biotechnol. 2014, 13, 3137–3143. [Google Scholar] [CrossRef]
- Villela, J. C. B.; Barbieri, R. L.; Castro, C. M.; Neitzke, R. S.; Vasconcelos, C. S.; Carbonari, T.; Mistura, C. C.; Priori, D. Caracterização Molecular de Variedades Crioulas de Pimentas (Capsicum Baccatum) Com Marcadores Microssatélites. Hortic. Bras. 2014, 32, 131–137. [Google Scholar] [CrossRef]
- Du, H.; Yang, J.; Chen, B.; Zhang, X.; Zhang, J.; Yang, K.; Geng, S.; Wen, C. Target Sequencing Reveals Genetic Diversity, Population Structure, Core-SNP Markers, and Fruit Shape-Associated Loci in Pepper Varieties. BMC Plant Biol. 2019, 19, 578. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, A.; Choi, S.; Jun, T.H.; Yang, E.Y.; Kim, J.H.; Lee, E.S.; Lee, H.E.; Kim, D.S.; Ahn, Y.K. Genotyping by Sequencing-Based Discovery of SNP Markers and Construction of Linkage Map from F5 Population of Pepper with Contrasting Powdery Mildew Resistance Trait. Biomed Res. Int. 2021, 2021, 6673010. [Google Scholar] [CrossRef]
- Xu, X.; Liu, Z.; Lin, X.; Mou, S.; Guan, D.; He, S. Genetic Diversity and Relationship Analysis of Pepper Germplasm Resources Based on Phenotype Traits and SRAP Molecular Markers. J Fujian Agric For Univ 2011, 40, 48–53. [Google Scholar]
- Qiu, Y.H.; Lin, S.T.; Zeng, S.G.; Luo, Y.; Zhu, B.T.; Li, Y.Q. Phenotypic and SRAP genetic diversity analysis of 41 yellow pepper germplasm resources. J South Agric 2021, 52, 2165–2173. [Google Scholar] [CrossRef]
- Maulani, R.; Murti, R. H.; Purwantoro, A. Molecular Diversity in Populations of Chili (Capsicum Annuum l.). SABRAO J. Breed. Genet. 2023, 55, 15–24. [Google Scholar] [CrossRef]
- Danojević, D.; Červenski, J.; Gvozdanović-Varga, J.; Ignjatov, M.; Medić-Pap, S.; Ilić, A.; Bugarski, D.; Takač, A.; Vlajić, S.; Popović, V.; Kiprovski, B.; Bajić, I.; Glogovac, S.; Milošević, D.; Stojanov, N.; Zeremski, T. Nove NS sorte povrtarskih biljnih vrsta, Zbornik referata 56 Savetovanja agronoma i poljoprivrednika Srbije (SAPS) i 2 Savetovanje agronoma Republike Srbije i Republike Srpske. 2022, 121-129, http://fiver.ifvcns.rs/handle/123456789/2548.
- Rani, M.; Jindal, S. K.; Vikal, Y.; Meena, O. P. Genetic Male Sterility Breeding in Heat Tolerant Bell Pepper: Introgression of Ms10 Gene from Hot Pepper through Marker-Assisted Backcrossing. Sci. Hortic. (Amsterdam) 2021, 285, 110172. [Google Scholar] [CrossRef]
- Červenski, J.; Medić-Pap, S.; Danojević, D.; Stojanović, A.; Savić, A. Technological quality of domestic cabbage (Brassica oleracea var. capitata L.) populations and varieties from the Vojvodina Province-Serbia. Contemporary Agriculture. 2014, 63, 473–479, http://fiver.ifvcns.rs/handle/123456789/1305. [Google Scholar]
- Fang, Z.; Liu, Y.; Lou, P.; Liu, G. Current Trends in Cabbage Breeding. J. New Seeds 2004, 6, 75–107. [Google Scholar] [CrossRef]
- Bin, L. J.; Zhang, H. J.; Lin, Y. X.; Wang, S. Y.; Cao, C. S. Molecular Identification of Cytoplasmic Male Sterile (CMS) Type in Common Head Cabbage. Mol. Plant Breed 2009, 7, 1149–1153. [Google Scholar]
- Zhang, Y.; Wang, X. J.; Li, O. C.; Song, H. Y.; Ren, X. S.; Si, J. Molecular Identification of Brassica Oleracea CMS and the Morphologyresponse of Flower to Nuclear Background. Acta Hortic. Sin 2010, 37, 915–922. [Google Scholar]
- Zhang, Y. Y.; Fang, F. Y.; Wang, Q. B.; Liu, Y. M.; Yang, L. M.; Zhuang, M.; Sun, P. T. Molecular Distinction of Two Ogura CMS Sources in Brassica Oleracea Var. Capitata L. Acta Hortic. Sin 2011, 44, 2959–2965. [Google Scholar]
- Chen, C.; Zhuang, M.; Fang, Z.-Y.; Wang, Q.-B.; Zhang, Y.-Y.; Liu, Y.-M.; Yang, L.-M.; Cheng, F. A Co-Dominant Marker BoE332 Applied to Marker-Assisted Selection of Homozygous Male-Sterile Plants in Cabbage (Brassica Oleracea Var. Capitata L.). J. Integr. Agric. 2013, 12, 596–602. [Google Scholar] [CrossRef]
- Yu, H.L.; Li, Z.Y.; Ren, W.J.; Han, F.Q.; Yang, L.M.; Zhuang, M.; Lv, H.H.; Liu, Y.M.; Fang, Z.Y.; Zhang, Y.Y. Creation of Fertility-Restored Materials for Ogura CMS in Brassica Oleracea by Introducing Rfo Gene from Brassica Napus via an Allotriploid Strategy. Theor. Appl. Genet. 2020, 133, 2825–2837. [Google Scholar] [CrossRef]
- Han, F.; Yuan, K.; Kong, C.; Zhang, X.; Yang, L.; Zhuang, M.; Zhang, Y.; Li, Z.; Wang, Y.; Fang, Z.; Lv, H. Fine Mapping and Candidate Gene Identification of the Genic Male-Sterile Gene Ms3 in Cabbage 51S. Theor. Appl. Genet. 2018, 131, 2651–2661. [Google Scholar] [CrossRef]
- Ji, J.L.; Yang, L.M.; Fang, Z.Y.; Zhuang, M.; Zhang, Y.Y.; Lv, H.-H.; Liu, Y.M.; Li, Z.S. Recessive Male Sterility in Cabbage (Brassica Oleracea Var. Capitata) Caused by Loss of Function of BoCYP704B1 Due to the Insertion of a LTR-Retrotransposon. Theor. Appl. Genet. 2017, 130, 1441–1451. [Google Scholar] [CrossRef]
- Wang, X.; Fang, Z.; Sun, P.; Liu, Y.; Yang, L. Identification of a RAPD Marker Linked to a Dominant Male Sterile Gene in Cabbage. Acta Hortic. Sin 1998, 25, 197–198. [Google Scholar]
- Wang, X.; Fang, Z.; Huang, S.; Sun, P.; Liu, Y.; Yang, L.; Zhuang, M.; Qu, D. An Extended Random Primer Amplified Region (ERPAR) Marker Linked to a Dominant Male Sterility Gene in Cabbage (Brassica Oleracea Var. Capitata). Euphytica 2000, 112, 267–273. [Google Scholar] [CrossRef]
- Wang, X.W.; Fang, Z.; Sun, P.; Liu, Y.; Yang, L.; Zhuang, M. A SCAR marker applicable in marker assisted selection of a dominant male sterility gene in cabbage. Acta Hortic. Sin. 2000a, 27, 143–144. [Google Scholar]
- Han, F.; Cui, H.; Zhang, B.; Liu, X.; Yang, L.; Zhuang, M.; Lv, H.; Li, Z.; Wang, Y.; Fang, Z.; Song, J.; Zhang, Y. Map-Based Cloning and Characterization of BoCCD4, a Gene Responsible for White/Yellow Petal Color in B. Oleracea. BMC Genomics 2019a, 20, 242. [Google Scholar] [CrossRef]
- Margalé, E.; Hervé, Y.; Hue, J.; Quiros, C. F. Determination of Genetic Variability by RAPD Markers in Cauliflower, Cabbage and Kale Local Cultivars from France. Genet. Resour. Crop Evol. 1995, 42, 281–289. [Google Scholar] [CrossRef]
- Koutita, O.; Tertivanidis, K.; Koutsos, T. V.; Koutsika-Sotiriou, M.; Skaracis, G. N. Genetic Diversity in Four Cabbage Populations Based on Random Amplified Polymorphic DNA Markers. J. Agric. Sci. 2005, 143, 377–384. [Google Scholar] [CrossRef]
- Saxena, B.; Kaur, R.; Bhardwaj, S. V. Assessment of Genetic Diversity in Cabbage Cultivars Using RAPD and SSR Markers. J. Crop Sci. Biotechnol. 2011, 14, 191–196. [Google Scholar] [CrossRef]
- Li, X.; Yu, H.; Li, Z.; Liu, X.; Fang, Z.; Liu, Y.; Yang, L.; Zhuang, M.; Lv, H.; Zhang, Y. Heterotic Group Classification of 63 Inbred Lines and Hybrid Purity Identification by Using SSR Markers in Winter Cabbage (Brassica Oleracea L. Var. Capitata). Hortic. Plant J 2018a, 4, 158–164. [Google Scholar] [CrossRef]
- Louarn, S.; Torp, A. M.; Holme, I. B.; Andersen, S. B.; Jensen, B. D. Database Derived Microsatellite Markers (SSRs) for Cultivar Differentiation in Brassica Oleracea. Genet. Resour. Crop Evol. 2007, 54, 1717–1725. [Google Scholar] [CrossRef]
- Flannery, M. L.; Mitchell, F. J. G.; Coyne, S.; Kavanagh, T. A.; Burke, J. I.; Salamin, N.; Dowding, P.; Hodkinson, T. R. Plastid Genome Characterisation in Brassica and Brassicaceae Using a New Set of Nine SSRs. Theor. Appl. Genet. 2006, 113, 1221–1231. [Google Scholar] [CrossRef]
- El-Esawi, M. A.; Germaine, K.; Bourke, P.; Malone, R. Genetic Diversity and Population Structure of Brassica Oleracea Germplasm in Ireland Using SSR Markers. C. R. Biol. 2016, 339, 133–140. [Google Scholar] [CrossRef]
- Leroy, X. J.; Leon, K.; Branchard, M. Characterisation of Brassica Oleracea L. by Microsatellite Primers. Plant Syst. Evol 2000, 225, 235–240. [Google Scholar] [CrossRef]
- Faltusová, Z.; Kučera, L.; Ovesná, J. Genetic Diversity of Brassica Oleracea Var. Capitata Gene Bank Accessions Assessed by AFLP. Electron. J. Biotechnol. 2011, 14. [Google Scholar] [CrossRef]
- Van Hintum, T. J. L.; van de Wiel, C. C. M.; Visser, D. L.; van Treuren, R.; Vosman, B. The Distribution of Genetic Diversity in a Brassica Oleracea Gene Bank Collection Related to the Effects on Diversity of Regeneration, as Measured with AFLPs. Theor. Appl. Genet. 2007, 114, 777–786. [Google Scholar] [CrossRef]
- Shapturenko, M. N.; Pechkovskaya, T. V.; Vakula, S. I.; Jakimovich, A. V.; Zabara, Y. M.; Khotyleva, L. V. Informative EST-SSR Markers for Genotyping and Intraspecific Differentiation of Brassica Oleracea Var. Capitata L. Russ. J. Genet. Appl. Res. 2017, 7, 14–20. [Google Scholar] [CrossRef]
- Palmé, A. E.; Hagenblad, J.; Solberg, S. Ø; Aloisi, K.; Artemyeva, A. SNP Markers and Evaluation of Duplicate Holdings of Brassica Oleracea in Two European Genebanks. Plants 2020, 9, 925. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, H.; Li, X.; Zhang, B.; Ren, W.; Liu, X.; Fang, Z.; Yang, L.; Zhuang, M.; Lv, H.; Zhang, Y. Kompetitive Allele-Specific PCR (KASP) Genotyping and Heterotic Group Classification of 244 Inbred Lines in Cabbage (Brassica Oleracea L. Var. Capitata). Euphytica 2020a, 216. [Google Scholar] [CrossRef]
- Crockett, P. A.; Bhalla, P. L.; Lee, C. K.; Singh, M. B. RAPD Analysis of Seed Purity in a Commercial Hybrid Cabbage (Brassica Oleracea Var. Capitata) Cultivar. Genome 2000, 43, 317–321. [Google Scholar] [CrossRef]
- Liu, G.; Liu, L.; Gong, Y.; Wang, Y.; Yu, F.; Shen, H.; Gui, W. Seed Genetic Purity Testing of F1 Hybrid Cabbage (Brassica Oleracea Var. Capitata) with Molecular Marker Analysis. Seed Sci. Technol. 2007, 35, 477–486. [Google Scholar] [CrossRef]
- Ye, S.; Wang, Y.; Huang, D.; Li, J.; Gong, Y.; Xu, L.; Liu, L. Genetic Purity Testing of F1 Hybrid Seed with Molecular Markers in Cabbage (Brassica Oleracea Var. Capitata). Sci. Hortic. (Amsterdam) 2013, 155, 92–96. [Google Scholar] [CrossRef]
- Kawamura, K.; Shimizu, M.; Kawanabe, T.; Pu, Z.; Kodama, T.; Kaji, M.; Osabe, K.; Fujimoto, R.; Okazaki, K. Assessment of DNA Markers for Seed Contamination Testing and Selection of Disease Resistance in Cabbage. Euphytica 2017, 213. [Google Scholar] [CrossRef]
- Cansian, R. L.; Echeverrigaray, S. Discrimination among Cultivars of Cabbage Using Randomly Amplified Polymorphic DNA Markers. HortScience 2000, 35, 1155–1158. [Google Scholar] [CrossRef]
- Li, Z.; Yu, H.; Fang, Z.; Yang, L.; Liu, Y.; Zhuang, M.; Lü, H.; Zhang, Y. Development of SNP Markers in Cabbage and Construction of DNA Fingerprinting of Main Varieties. China J. Agric. Sci 2018b, 51, 2771–2787. [Google Scholar]
- Jo, J.; Kang, M.-Y.; Kim, K. S.; Youk, H. R.; Shim, E.-J.; Kim, H.; Park, J.-S.; Sim, S.-C.; Yu, B. C.; Jung, J.-K. Genome-Wide Analysis-Based Single Nucleotide Polymorphism Marker Sets to Identify Diverse Genotypes in Cabbage Cultivars (Brassica Oleracea Var. Capitata). Sci. Rep. 2022, 12, 20030. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhang, X.; Yuan, K.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Liu, Y.; Li, Z.; Lv, H. A User-Friendly KASP Molecular Marker Developed for the DGMS-Based Breeding System in Brassica Oleracea Species. Mol. Breed. 2019b, 39. [Google Scholar] [CrossRef]
- Han, F.Q.; Yang, C.; Fang, Z.Y.; Yang, L.M.; Zhuang, M.; Lv, H.H.; Liu, Y.M.; Li, Z.S.; Liu, B.; Yu, H.L.; Liu, X.P.; Zhang, Y.Y. Inheritance and InDel Markers Closely Linked to Petal Color Gene (Cpc-1) in Brassica Oleracea. Mol. Breed. 2015, 35. [Google Scholar] [CrossRef]
- Liu, D.; Tang, J.; Liu, Z.; Dong, X.; Zhuang, M.; Zhang, Y.; Lv, H.; Sun, P.; Liu, Y.; Li, Z. Fine Mapping of BoGL1, a Gene Controlling the Glossy Green Trait in Cabbage (Brassica Oleracea L. Var. Capitata). Mol. Breed 2017a, 37, 69. [Google Scholar] [CrossRef]
- Liu, Z.; Fang, Z.; Zhuang, M.; Zhang, Y.; Lv, H.; Liu, Y.; Li, Z.; Sun, P.; Tang, J.; Liu, D. Fine-Mapping and Analysis of Cgl1, a Gene Conferring Glossy Trait in Cabbage (Brassica Oleracea L. Var. Capitata). Front. Plant Sci 2017b, 8. [Google Scholar] [CrossRef]
- Liu, D.; Tang, J.; Liu, Z.; Dong, X.; Zhuang, M.; Zhang, Y.; Lv, H.; Sun, P.; Liu, Y.; Li, Z.; Ye, Z.; Fang, Z.; Yang, L. Cgl2 Plays an Essential Role in Cuticular Wax Biosynthesis in Cabbage (Brassica Oleracea L. Var. Capitata). BMC Plant Biol. 2017c, 17, 223. [Google Scholar] [CrossRef]
- Liu, D.; Dong, X.; Liu, Z.; Tang, J.; Zhuang, M.; Zhang, Y.; Lv, H.; Liu, Y.; Li, Z.; Fang, Z.; Yang, L. Fine Mapping and Candidate Gene Identification for Wax Biosynthesis Locus, BoWax1 in Brassica Oleracea L. Var. Capitata. Front. Plant Sci. 2018, 9, 309. [Google Scholar] [CrossRef]
- Ji, J.; Cao, W.; Dong, X.; Liu, Z.; Fang, Z.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; Sun, P.; Liu, Y.; Li, Z.; Yang, L. A 252-Bp Insertion in BoCER1 Is Responsible for the Glossy Phenotype in Cabbage (Brassica Oleracea L. Var. Capitata). Mol. Breed. 2018, 38. [Google Scholar] [CrossRef]
- Dong, X.; Ji, J.; Yang, L.; Fang, Z.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; Sun, P.; Tang, J.; Liu, D.; Liu, Y.; Li, Z. Fine-Mapping and Transcriptome Analysis of BoGL-3, a Wax-Less Gene in Cabbage (Brassica Oleracea L. Var. Capitata). Mol. Genet. Genomics 2019, 294, 1231–1239. [Google Scholar] [CrossRef]
- Ji, J.; Cao, W.; Tong, L.; Fang, Z.; Zhang, Y.; Zhuang, M.; Wang, Y.; Yang, L.; Lv, H. Identification and Validation of an ECERIFERUM2- LIKE Gene Controlling Cuticular Wax Biosynthesis in Cabbage (Brassica Oleracea L. Var. Capitata L.). Theor. Appl. Genet. 2021, 134, 4055–4066. [Google Scholar] [CrossRef]
- Liu, X.; Yang, C.; Han, F.; Fang, Z.; Yang, L.; Zhuang, M.; Lv, H.; Liu, Y.; Li, Z.; Zhang, Y. Genetics and Fine Mapping of a Yellow-Green Leaf Gene (Ygl-1) in Cabbage (Brassica Oleracea Var. Capitata L.). Mol. Breed 2016. [Google Scholar] [CrossRef]
- Song, H.; Yi, H.; Lee, M.; Han, C.T.; Lee, J.; Kim, H.; Park, J.I.; Nou, I.S.; Kim, S.J.; Hur, Y. Purple Brassica Oleracea Var. Capitata F. Rubra Is Due to the Loss of BoMYBL2-1 Expression. BMC Plant Biol. 2018a, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Li, X.; Choi, S. R.; Nguyen, V. D.; Dhandapani, V.; Kim, Y. Y.; Ramchiary, N.; Kim, J. G.; Edwards, D.; Batley, J. Mapping QTLs of Resistance to Head Splitting in Cabbage (Brassica Oleracea Var. Capitata L.). Mol. Breed 2015, 35, 126. [Google Scholar] [CrossRef]
- Zhu, X.; Tai, X.; Ren, Y.; Wang, M.; Chen, J.; Bo, T. QTL-Seq and Marker Development for Resistance to Head Splitting in Cabbage. Euphytica 2022a, 218. [Google Scholar] [CrossRef]
- Zhao, T.; Miao, L.; Zou, M.; Hussain, I.; Yu, H.; Li, J.; Sun, N.; Kong, L.; Wang, S.; Li, J. Development of SSRs Based on the Whole Genome and Screening of Bolting-Resistant SSR Marker in Brassica Oleracea L. Horticulturae 2024, 10. [Google Scholar] [CrossRef]
- Li, Q.; Peng, A.; Yang, J.; Zheng, S.; Li, Z.; Mu, Y.; Chen, L.; Si, J.; Ren, X.; Song, H. A 215-Bp Indel at Intron I of BoFLC2 Affects Flowering Time in Brassica Oleracea Var. Capitata during Vernalization. Theor. Appl. Genet. 2022, 135, 2785–2797. [Google Scholar] [CrossRef]
- Song, H.; Lee, M.; Hwang, B. H.; Han, C. T.; Park, J. I.; Hur, Y. Development and Application of a PCR-Based Molecular Marker for the Identification of High Temperature Tolerant Cabbage (Brassica Oleracea Var. Capitata) Genotypes. Agronomy 2020a, 10. [Google Scholar] [CrossRef]
- Song, H.; Kim, H.; Hwang, B.-H.; Yi, H.; Hur, Y. Natural Variation in Glycine-Rich Region of Brassica Oleracea Cold Shock Domain Protein 5 (BoCSDP5) Is Associated with Low Temperature Tolerance. Genes Genomics 2020b, 42(12), 1407–1417. [Google Scholar] [CrossRef]
- Song, H.; Yi, H.; Han, C.T.; Park, J.I.; Hur, Y. Allelic Variation in Brassica Oleracea CIRCADIAN CLOCK ASSOCIATED 1 (BoCCA1) Is Associated with Freezing Tolerance. Hortic. Environ. Biotechnol. 2018b, 59, 423–434. [Google Scholar] [CrossRef]
- Jiang, M.; Zhao, Y.; Xie, J.; Tian, R.; Chen, Y.; Kang, J. Development of a SCAR Marker for Fusarium Wilt Resistance in Cabbage. Sci. Agric. Sin 2011, 44, 3053–3059. [Google Scholar]
- Pu, Z.J.; Shimizu, M.; Zhang, Y.J.; Nagaoka, T.; Hayashi, T.; Hori, H.; Matsumoto, S.; Fujimoto, R.; Okazaki, K. Genetic Mapping of a Fusarium Wilt Resistance Gene in Brassica Oleracea. Mol. Breed. 2012, 30, 809–818. [Google Scholar] [CrossRef]
- Liu, X.; Han, F.; Kong, C.; Fang, Z.; Yang, L.; Zhang, Y.; Zhuang, M.; Liu, Y.; Li, Z.; Lv, H. Rapid Introgression of the Fusarium Wilt Resistance Gene into an Elite Cabbage Line through the Combined Application of a Microspore Culture, Genome Background Analysis, and Disease Resistance-Specific Marker Assisted Foreground Selection. Front. Plant Sci. 2017, 8, 354. [Google Scholar] [CrossRef]
- Dubina, E. V.; Makukha, Y. A.; Artemeva, A. M.; Fateev, D. A.; Garkusha, S. V.; Gorun, O. L.; Lesnyak, S. A. Molecular Marking in Brassica Oleracea L. Breeding for Resistance to Fusarium Wilt. Russ. J. Genet 2023, 59, 1004–1010. [Google Scholar] [CrossRef]
- Lv, H.H.; Yang, L.M.; Kang, J.G.; Wang, Q.B.; Wang, X.W.; Fang, Z.Y.; Liu, Y.M.; Zhuang, M.; Zhang, Y.Y.; Lin, Y.; Yang, Y.H.; Xie, B.Y.; Liu, B.; Liu, J.S. Development of InDel Markers Linked to Fusarium Wilt Resistance in Cabbage. Mol. Breed. 2013, 32, 961–967. [Google Scholar] [CrossRef]
- Sato, M.; Shimizu, M.; Shea, D. J.; Hoque, M.; Kawanabe, T.; Miyaji, N.; Fujimoto, R.; Fukai, E.; Okazaki, K. Allele Specific DNA Marker for Fusarium Resistance Gene FocBo1 in Brassica Oleracea. Breed. Sci. 2019, 69, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Afrin, K. S.; Rahim, M. A.; Park, J.-I.; Natarajan, S.; Rubel, M. H.; Kim, H.-T.; Nou, I.-S. Screening of Cabbage (Brassica Oleracea L.) Germplasm for Resistance to Black Rot. Plant Breed. Biotechnol. 2018, 6, 30–43. [Google Scholar] [CrossRef]
- Makukha, Y. PCR Identification of Genes of Resistance to Black Rot in White Cabbage Using SSR-Markers. BIO Web Conf. 2020, 21, 00013. [Google Scholar] [CrossRef]
- Hong, J.E.; Afrin, K. S.; Rahim, M. A.; Jung, H.J.; Nou, I.S. Inheritance of Black Rot Resistance and Development of Molecular Marker Linked to Xcc Races 6 and 7 Resistance in Cabbage. Plants 2021, 10, 1940. [Google Scholar] [CrossRef]
- Nomura, K.; Minegishi, Y.; Kimizuka-Takagi, C.; Fujioka, T.; Moriguchi, K.; Shishido, R.; Ikehashi, H. Evaluation of F2 and F3 Plants Introgressed with QTLs for Clubroot Resistance in Cabbage Developed by Using SCAR Markers. Plant Breed. 2005, 124, 371–375. [Google Scholar] [CrossRef]
- Tomita, H.; Shimizu, M.; Asad-Ud Doullah, M. D.; Fujimoto, R.; Okazaki, K. Accumulation of Quantitative Trait Loci Confer-Ring Broad-Spectrum Clubroot Resistance in Brassica Oleracea. Mol. Breed 2013, 32, 889–900. [Google Scholar] [CrossRef]
- Nagaoka, T.; Doullah, M. A. U.; Matsumoto, S.; Kawasaki, S.; Ishikawa, T.; Hori, H.; Okazaki, K. Identification of QTLs That Control Clubroot Resistance in Brassica Oleracea and Comparative Analysis of Clubroot Resistance Genes between B. Rapa and B. Oleracea. Theor. Appl. Genet. 2010, 120, 1335–1346. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, L.; Zhang, Y.; Zhuang, M.; Ji, J.; Hou, X.; Li, Z.; Han, F.; Fang, Z.; Lv, H.; Wang, Y. Introgression of Clubroot Resistant Gene into Brassica Oleracea L. from Brassica Rapa Based on Homoeologous Exchange. Hortic. Res. 2022b, 9, uhac195. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, B.; Yang, L.; Zhuang, M.; Lv, H.; Wang, Y.; Ji, J.; Hou, X.; Zhang, Y. Fine Mapping and Identification of the Downy Mildew Resistance Gene BoDMR2 in Cabbage (Brassica Oleracea L. Var. Capitata). BMC Plant Biol. 2024, 24, 987. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.A. Downy Mildew Resistance in B. oleracea. Ph.D. Thesis, The University of East Anglia, Norwich, UK, 2022. [Google Scholar]
- Strembovskiy, I. V.; Kroupin, P. Y. The Current Status and Prospects of Molecular Marker Applications in Head Cabbage (Brassica Oleracea Var. Capitata L.): A Review. Agronomy (Basel) 2025, 15, 2644. [Google Scholar] [CrossRef]
- Shatilov, M. V.; Razin, A. F.; Ivanova, M. I. Analysis of the World Lettuce Market. IOP Conf. Ser. Earth Environ. Sci. 2019, 395, 012053. [Google Scholar] [CrossRef]
- Lebeda, A.; Ryder, E. J.; Grube, R. Lettuce (Asteraceae;Lactuca Spp). In Genetic resources, chromosome engineering and crop improvement; Singh, R. J., Ed.; CRC Press, 2007, 3, 377–472.
- Hartman, Y.; Hooftman, D.; Uwimana, B. Abiotic Stress QTL in Lettuce Crop-Wild Hybrids: Comparing Green-House and Field Experiments. Ecology Evolution 2014, 4, 2395–2409. [Google Scholar] [CrossRef]
- McCabe, M. S.; Garratt, L. C.; Schepers, F.; Jordi, W. J. R. M.; Stoopen, G. M.; Davelaar, E.; van Rhijn, J. H. A.; Power, J. B.; Davey, M. R. Effects of PSAG12-IPT Gene Expression on Development and Senescence in Transgenic Lettuce. Plant Physiol. 2001, 127, 505–516. [Google Scholar] [CrossRef]
- Hassan, M. N.; Mekkawy, S. A.; Mahdy, M.; Salem, K. F. M.; Tawfik, E. Recent Molecular and Breeding Strategies in Lettuce (Lactuca Spp.). Genet. Resour. Crop Evol. 2021, 68, 3055–3079. [Google Scholar] [CrossRef]
- Landry, B. S.; Kesseli, R.; Farrara, B.; Michelmore, R. W. A Genetic Map of Lettuce (Lactuca Sativa L.) with Restriction Frag-Ment Length Polymorphism, Isozymes, Disease Resistance and Morphological Markers. Genetics 1987, 116, 331–337. [Google Scholar] [CrossRef]
- Kesseli, R.; Ochoa, O.; Michelmore, R. Variation at RFLP Loci in Lactuca Spp. and Origin of Cultivated Lettuce (L.Sativa). Genome 1991, 34, 430–436. [Google Scholar] [CrossRef]
- Vermeulen, A.; Desprez, B.; Lancelin, D.; Bannerot, H. Relationship among Cichorium Species and Related Genera as De-Termined by Analysis of Mitochondrial RFLPs. Theor Appl Genet 1994, 88, 159–166. [Google Scholar] [CrossRef]
- Kesseli, R. V.; Paran, I.; Michelmore, R. W. Analysis of a Detailed Genetic Linkage Map of Lactuca Sativa (Lettuce) Con-Structed from RFLP and RAPD Markers. Genetics 1994, 136, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M. Molecular Genetic Markers for Assessing the Genetic Variation and Relationships in Lactuca Germplasm. Annu. Res. Rev. Biol. 2015, 8, 1–13. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nishikawa, A.; Oeda, K. DNA Polymorphisms in Oryza Sativa L. and Lactuca Sativa L. Amplified by Arbitrary Primed PCR. Euphytica 1994, 78, 143–148. [Google Scholar] [CrossRef]
- Tardin, F. D.; Junior, A.; Pereira, M. G. Genetic Diversity and Determination of the Optimum Number of RAPD Markers in Lettuce (Lactuca Sativa L.). Acta Scientiarum: Agronomy Maringa’ 2003, 25, 1–5. [Google Scholar]
- Hill, M.; Witsenboer, H.; Zabeau, M.; Vos, P.; Kesseli, R.; Michelmore, R. PCR-Based Fingerprinting Using AFLPs as a Tool for Studying Genetic Relationships in Lactuca Spp. Theor. Appl. Genet. 1996, 93, 1202–1210. [Google Scholar] [CrossRef]
- Jeuken, M.; Van Wijk, R.; Peleman, J.; Lindhout, P. An Integrated Interspecific AFLP Map of Lettuce (Lactuca) Based on Two L. Sativa 9 L. Saligna F2 Populations. Theor Appl Genet 2001, 103, 638–647. [Google Scholar] [CrossRef]
- Koopman, W.; Zevenbergen, M. J.; Ronald, G.; Van Den Berg, R. G. Species Relationship in Lactuca s. l. (Lactuceae, Asteraceae) Inferred from AFLP Fingerprints. Am J Bot 2001, 88, 1881–1887. [Google Scholar] [CrossRef]
- Van Treuren, R.; Hintum, V. Comparison of Anonymous and Targeted Molecular Markers for the Estimation of Genetic Diversity in Ex-Situ Conserved Lactuca. Theor Appl Genet 2009, 119, 1265–1279. [Google Scholar] [CrossRef]
- Witsenboer, H.; Vogel, J.; Michelmore, R. W. Identification, Genetic Localization, and Allelic Diversity of Selectively Am-Plified Microsatellite Polymorphic Loci in Lettuce and Wild Relatives (Lactuca Spp). Genome 1997, 40, 923–936. [Google Scholar] [CrossRef]
- Van de Wiel, C.; Arens, P.; Vosman, B. Microsatellite Fingerprinting in Lettuce (Lactuca Sativa L.) and Wild Relatives. Plant Cell Rep. 1998, 17, 837–842. [Google Scholar] [CrossRef]
- Van de Wiel, C.; Arens, P.; Vosman, B. Microsatellite Retrieval in Lettuce (Lactuca Sativa L.). Genome 1999, 42, 139–149. [Google Scholar] [CrossRef]
- Rauscher, G.; Simko, I. Development of Genomic SSR Markers for Fingerprinting Lettuce (Lactuca Sativa L.) Cultivars and Mapping Genes. BMC Plant Biol. 2013, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Rui, S.; Qi, G.; Shuangxi, F.; Chaojie, L.; Yingyan, H. Analysis of Genetic Diversity in Purple Lettuce (Lactuca Sativa L.) by SSR Markers. Pak. J. Bot. 2020, 52. [Google Scholar] [CrossRef] [PubMed]
- Simko, I. Development of EST-SSR Markers for the Study of Population Structure in Lettuce (Lactuca Sativa L.). J. Hered. 2009, 100, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Riar, D. S.; Rustgi, S.; Burke, I. C.; Gill, K. S.; Yenish, J. P. EST-SSR Development from 5 Lactuca Species and Their Use in Studying Genetic Diversity among L. Serriola Biotypes. J. Hered. 2011, 102, 17–28. [Google Scholar] [CrossRef]
- Moreno-Vazquez, S.; Ochoa, O. E.; Faber, N. SNP-Based Codominant Markers for a Recessive Gene Conferring Re-Sistance to Corky Root Rot (Rhizomonas Suberifaciens) in Lettuce (Lactuca Sativa). Genome 2003, 46, 1059–1069. [Google Scholar] [CrossRef]
- Simko, I.; Pechenick, D. A.; McHale, L. K.; Truco, M. J.; Ochoa, O. E.; Michelmore, R. W.; Scheffler, B. E. Development of Molecular Markers for Marker-Assisted Selection of Dieback Disease Resistance in Lettuce (Lactuca Sativa). Acta Hortic. 2010, 859, 401–408. [Google Scholar] [CrossRef]
- Walley, P. G.; Hough, G.; Moore, J. D.; Carder, J.; Elliott, M.; Mead, A.; Jones, J.; Teakle, G.; Barker, G.; Buchanan-Wollaston, V.; Hand, P.; Pink, D.; Collier, R. Towards New Sources of Resistance to the Currant-Lettuce Aphid (Nasonovia Ribisnigri). Mol. Breed. 2017, 37, 4. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Z.; Chen, H.; Zhou, L.; Liu, Y.; Luo, L. SRAP Markers and Morphological Traits Could Be Used in Test of Distinctiveness, Uniformity, and Stability (DUS) of Lettuce (Lactuca Sativa) Varieties. J. Agric. Sci. 2011, 4. [Google Scholar] [CrossRef]
- Salem, K. F. M.; Alghuthaymi, M. A.; Mahdy, M.; Mekkawy, S. A.; Hassan, M. N.; Ibrahim, A. A.; Tawfik, E. Advances in Lettuce (Lactuca Spp.) Molecular Breeding Strategies. In Smart Plant Breeding for Vegetable Crops in Post-genomics Era; Springer Nature Singapore: Singapore, 2023, 251–277. [CrossRef]
- Meuwissen, T. H.; Hayes, B. J.; Goddard, M. E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Heffner, E. L.; Sorrells, M. E.; Jannink, J.-L. Genomic Selection for Crop Improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]
- Varshney, R. K.; Bohra, A.; Yu, J.; Graner, A.; Zhang, Q.; Sorrells, M. E. Designing Future Crops: Genomics-Assisted Breeding Comes of Age. Trends Plant Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Du, S.; Wei, A.; Cheng, Z.; Meng, H.; Han, Y. Hybrid Prediction in Horticulture Crop Breeding: Progress and Challenges. Plants 2024, 13, 2790. [Google Scholar] [CrossRef]
- Kondo, F.; Kumanomido, Y.; D’Andrea, M.; Palombo, V.; Ahmed, N.; Futatsuyama, S.; Nemoto, K.; Matsushima, K. Phenotypic Simulation for Fruit-Related Traits in F1 Progenies of Chili Peppers (Capsicum Annuum) Using Genomic Prediction Based Solely on Parental Information. Mol. Genet. Genomics 2025, 300, 15. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Z.; Liu, G.; Jiang, Y.; Maurer, H. P.; Würschum, T.; Mock, H.-P.; Matros, A.; Ebmeyer, E.; Schachschneider, R.; Kazman, E.; Schacht, J.; Gowda, M.; Longin, C. F. H.; Reif, J. C. Genome-Based Establishment of a High-Yielding Heterotic Pattern for Hybrid Wheat Breeding. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 15624–15629. [Google Scholar] [CrossRef]
- Yue, L.; Zhang, S.; Zhang, L.; Liu, Y.; Cheng, F.; Li, G.; Zhang, S.; Zhang, H.; Sun, R.; Li, F. Heterotic Prediction of Hybrid Performance Based on Genome-Wide SNP Markers and the Phenotype of Parental Inbred Lines in Heading Chinese Cabbage (Brassica Rapa L. Ssp. Pekinensis). Sci. Hortic. (Amsterdam) 2022, 296, 110907. [Google Scholar] [CrossRef]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de Los Campos, G.; Burgueño, J.; González-Camacho, J. M.; Pérez-Elizalde, S.; Beyene, Y.; Dreisigacker, S.; Singh, R.; Zhang, X.; Gowda, M.; Roorkiwal, M.; Rutkoski, J.; Varshney, R. K. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Paspureddy, S. L.; Sasirekhamani, M.; Sneha, P. I.; Deepanshu; Sharma, A.; Harshavardhan, S. S.; Chandravanshi, R.; Sahu, A. Advances in Haploid and Doubled Haploid Technology for Accelerating Breeding Cycles in Crop Improvement. Plant Cell Biotechnol. Mol. Biol. 2025, 26, 61–77. [Google Scholar] [CrossRef]
- Singh, S.; Dey, S. S.; Bhatia, R.; Kumar, R.; Sharma, K.; Behera, T. K. Heterosis and Combining Ability in Cytoplasmic Male Sterile and Doubled Haploid Based Brassica Oleracea Progenies and Prediction of Heterosis Using Microsatellites. PLoS One 2019, 14, e0210772. [Google Scholar] [CrossRef]
- Persa, R.; Ribeiro, P. C. de O.; Jarquin, D. The Use of High-Throughput Phenotyping in Genomic Selection Context. Crop Breed. Appl. Biotechnol. 2021, 21. [Google Scholar] [CrossRef]
- Yoosefzadeh Najafabadi, M.; Hesami, M.; Eskandari, M. Machine Learning-Assisted Approaches in Modernized Plant Breeding Programs. Genes (Basel) 2023, 14, 777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yang, F.; Li, J.; Liu, Z.; Han, Y.; Zhang, Q.; Pan, S.; Zhao, X.; Wang, K. Progress and Perspectives on Genomic Selection Models for Crop Breeding. Technology in Agronomy 2025, 5, 0–0. [Google Scholar] [CrossRef]
- Jaganathan, D.; Dutta, B.; Basu, S.; Bag, S.; Srinivasan, R.; Eybishitz, A.; Barchenger, D. W.; Caarls, L.; Simmons, A. M.; Nankar, A. N. Breeding Vegetables for Whitefly Resistance: Past, Present, and Future in the AI Era. Front. Plant Sci. 2025, 16, 1724403. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Wang, Z.; Tang, M.; Hong, D.; Sun, Y.; Ren, J.; Zhang, N.; Zeng, H. CRISPR-Cas9 Gene Editing for Fruit and Vegetable Crops: Strategies and Prospects. Horticulturae 2021, 7, 193. [Google Scholar] [CrossRef]
- Li, H.; Yang, X.; Shang, Y.; Zhang, Z.; Huang, S. Vegetable Biology and Breeding in the Genomics Era. Sci. China Life Sci. 2023, 66, 226–250. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Halder, K.; Datta, A. Classification of CRISPR/Cas System and Its Application in Tomato Breeding. Theor. Appl. Genet. 2022, 135, 367–387. [Google Scholar] [CrossRef]
- Brooks, C.; Nekrasov, V.; Lippman, Z. B.; Van Eck, J. Efficient Gene Editing in Tomato in the First Generation Using the Clus-Tered Regularly Interspaced Short Palindromic Repeats/CRISPR-Associated9 System. Plant Physiol 2014, 166, 1292–1297. [Google Scholar] [CrossRef]
- De Toledo Thomazella, P.; Brail, D.; Dahlbeck, Q.; Staskawicz, D. CRISPR-Cas9 Mediated Mutagenesis of a DMR6 Ortholog in Tomato Confers Broad-Spectrum Disease Resistance. BioRxiv 2016. [Google Scholar]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid Generation of a Transgene-Free Powdery Mildew Resistant Tomato by Genome Deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Prihatna, C.; Barbetti, M. J.; Barker, S. J. A Novel Tomato Fusarium Wilt Tolerance Gene. Front. Microbiol. 2018, 9, 1226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, L.; Zhao, R.; Yu, W.; Li, R.; Li, Y.; Sheng, J.; Shen, L. Knockout of SlMAPK3 Reduced Disease Resistance to Botrytis Cinerea in Tomato Plants. J. Agric. Food Chem. 2018, 66, 8949–8956. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Qin, K.; Song, X.; Zhang, Q.; Zhou, Y.; Xia, X.; Yu, J. BZR1 Transcription Factor Regulates Heat Stress Tolerance Through FERONIA Receptor-Like Kinase-Mediated Reactive Oxygen Species Signaling in Tomato. Plant Cell Physiol 2018, 59, 2239–2254. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Wang, L.; Chen, L.; Zhao, R.; Sheng, J.; Shen, L. Reduction of Tomato-Plant Chilling Tolerance by CRISPR-Cas9-Mediated SlCBF1 Mutagenesis. J. Agric. Food Chem. 2018c, 66, 9042–9051. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef]
- Butler, N. M.; Atkins, P. A.; Voytas, D. F.; Douches, D. S. Generation and Inheritance of Targeted Mutations in Potato (Sola-Num Tuberosum L.) Using the CRISPR/Cas System. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Bari, V. K.; Nassar, J. A.; Kheredin, S. M.; Gal-On, A.; Ron, M.; Britt, A.; Steele, D.; Yoder, J.; Aly, R. CRISPR/Cas9-Mediated Mutagenesis of CAROTENOID CLEAVAGE DIOXYGENASE 8 in Tomato Provides Resistance against the Parasitic Weed Phelipanche Aegyptiaca. Sci. Rep. 2019, 9, 11438. [Google Scholar] [CrossRef]
- Bari, V. K.; Nassar, J. A.; Aly, R. CRISPR/Cas9 Mediated Mutagenesis of MORE AXILLARY GROWTH 1 in Tomato Confers Resistance to Root Parasitic Weed Phelipanche Aegyptiaca. Sci. Rep. 2021, 11, 3905. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of Wild Tomato Is Accelerated by Genome Editing. Nat. Biotechnol. 2018d, 36, 1160–1163. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z. H.; Man, J.; Bartlett, M. E.; Lippman, Z. B. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480.e8. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N. J.; Čegan, R.; Zhang, Y.; Voytas, D. F. High-Frequency, Precise Modification of the Tomato Genome. Genome Biol. 2015, 16(1), 232. [Google Scholar] [CrossRef]
- Deng, L.; Wang, H.; Sun, C.; Li, Q.; Jiang, H.; Du, M.; Li, C.B.; Li, C. Efficient Generation of Pink-Fruited Tomatoes Using CRISPR/Cas9 System. J. Genet. Genomics 2018, 45, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Filler Hayut, S.; Melamed Bessudo, C.; Levy, A. A. Targeted Recombination between Homologous Chromosomes for Precisebreeding in Tomato. Nat. Commun 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, D.; Zhang, Y.; Wu, L.; Zhang, Y.; Ye, L.; Pan, C.; He, Y.; Huang, L.; Ruan, Y.L.; Lu, G. Evidence for a Specific and Critical Role of Mitogen-Activated Protein Kinase 20 in Uni-to-Binucleate Transition of Microgametogenesis in Tomato. New Phytol. 2018, 219, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient Increase of Ɣ-Aminobutyric Acid (GABA) Content in Tomato Fruits by Targeted Mutagenesis. Sci. Rep. 2017, 7, 7057. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Chen, S.; Tian, H.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Lycopene Is Enriched in Tomato Fruit by CRISPR/Cas9- Mediated Multiplex Genome Editing. Front. Plant Sci 2018e, 9. [Google Scholar] [CrossRef]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Toki, S. CRISPR/Cas9-Mediated Mutagenesis of the RIN Locus That Regulates Tomato Fruit Ripening. Biochem. Biophys. Res. Commun. 2015, 467, 76–82. [Google Scholar] [CrossRef]
- Lang, Z.; Wang, Y.; Tang, K.; Tang, D.; Datsenka, T.; Cheng, J.; Zhang, Y.; Handa, A. K.; Zhu, J.K. Critical Roles of DNA Demethylation in the Activation of Ripening-Induced Genes and Inhibition of Ripening-Repressed Genes in Tomato Fruit. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E4511–E4519. [Google Scholar] [CrossRef]
- Wang, D.; Samsulrizal, N. H.; Yan, C.; Allcock, N. S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S. E.; Bagheri, H. M.; Perez Fons, L.; Fraser, P. D.; Foster, T.; Fray, R.; Knox, J. P.; Seymour, G. B. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar] [CrossRef]
- Yu, Q.H.; Wang, B.; Li, N.; Tang, Y.; Yang, S.; Yang, T.; Xu, J.; Guo, C.; Yan, P.; Wang, Q.; Asmutola, P. CRISPR/Cas9-Induced Targeted Mutagenesis and Gene Replacement to Generate Long-Shelf Life Tomato Lines. Sci. Rep. 2017, 7, 11874. [Google Scholar] [CrossRef]
- Roldan, M. V. G.; Périlleux, C.; Morin, H.; Huerga-Fernandez, S.; Latrasse, D.; Benhamed, M.; Bendahmane, A. Natural and Induced Loss of Function Mutations in SlMBP21 MADS-Box Gene Led to Jointless-2 Phenotype in Tomato. Sci. Rep. 2017, 7, 4402. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Yeshayahou, E.; Bolger, A. M.; Arazi, T.; Gupta, S. K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato Facultative Parthenocarpy Results from SlAGAMOUS-LIKE 6 Loss of Function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Ueta, R.; Abe, C.; Watanabe, T.; Sugano, S. S.; Ishihara, R.; Ezura, H.; Osakabe, Y.; Osakabe, K. Rapid Breeding of Parthenocarpic Tomato Plants Using CRISPR/Cas9. Sci. Rep. 2017, 7, 507. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Alon, I.; Naomi, O.; Sun, T. P. DELLA-ARF/IAA Interaction Mediates Crosstalk between Gibberellin and Auxin Signaling in Controlling Fruit Initiation in Solanum Lycopersicum. Plant Cell 2018, 30, 1710–1728. [Google Scholar] [CrossRef]
- Xu, C.; Park, S. J.; Van Eck, J.; Lippman, Z. B. Control of Inflorescence Architecture in Tomato by BTB/POZ Transcriptional Regulators. Genes Dev. 2016, 30, 2048–2061. [Google Scholar] [CrossRef]
- Soyk, S.; Müller, N. A.; Park, S. J.; Schmalenbach, I.; Jiang, K.; Hayama, R.; Zhang, L.; Van Eck, J.; Jiménez-Gómez, J. M.; Lippman, Z. B. Variation in the Flowering Gene SELF PRUNING 5G Promotes Day-Neutrality and Early Yield in Tomato. Nat. Genet. 2017, 49, 162–168. [Google Scholar] [CrossRef]
- Nie, H.; Shi, Y.; Geng, X.; Xing, G. CRISRP/Cas9-Mediated Targeted Mutagenesis of Tomato Polygalacturonase Gene (SlPG) Delays Fruit Softening. Front. Plant Sci. 2022, 13, 729128. [Google Scholar] [CrossRef]
- Yang, Q.; Cai, L.; Wang, M.; Gan, G.; Li, W.; Li, W.; Jiang, Y.; Yuan, Q.; Qin, C.; Yu, C.; Wang, Y. CRISPR/Cas9 Allows for the Quick Improvement of Tomato Firmness Breeding. Curr. Issues Mol. Biol. 2024, 47, 9. [Google Scholar] [CrossRef]
- Ahmad, S.; Tang, L.; Shahzad, R.; Mawia, A. M.; Rao, G. S.; Jamil, S.; Wei, C.; Sheng, Z.; Shao, G.; Wei, X.; Hu, P.; Mahfouz, M. M.; Hu, S.; Tang, S. CRISPR-Based Crop Improvements: A Way Forward to Achieve Zero Hunger. J. Agric. Food Chem. 2021, 69, 8307–8323. [Google Scholar] [CrossRef]
- Chaudhary, N.; Sandhu, R. A Comprehensive Review on Speed Breeding Methods and Applications. Euphytica 2024, 220(3). [Google Scholar] [CrossRef]
- Ullah, I.; Zhang, H.; Khan, M. R.; Mateen, A.; Pei, Y.; Shakeel, A.; Bhat, A. H.; Fu, C.; Chen, R. Molecular Mecha-Nisms and Genomic Strategies for Enhancing Stress Resilience in Pepper Crop. Scientia Horticulturae 2025, 352. [Google Scholar]
- Mishra, R.; Mohanty, J. N.; Mahanty, B.; Joshi, R. K. A Single Transcript CRISPR/Cas9 Mediated Mutagenesis of CaERF28 Confers Anthracnose Resistance in Chilli Pepper (Capsicum Annuum L.). Planta 2021, 254, 5. [Google Scholar] [CrossRef] [PubMed]
- Bulle, M.; Venkatapuram, A. K.; Abbagani, S.; Kirti, P. B. CRISPR/Cas9 Based Genome Editing of Phytoene Desaturase (PDS) Gene in Chilli Pepper (Capsicum Annuum L.). J. Genet. Eng. Biotechnol. 2024, 22, 100380. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Shin, H.; Kim, C. Y.; Park, J.; Kim, H. Highly Efficient CRISPR/Cas9-RNP Mediated CaPAD1 Editing in Protoplasts of Three Pepper (Capsicum Annuum L.) Cultivars. Plant Signal. Behav. 2024, 19, 2383822. [Google Scholar] [CrossRef]
- Park, S.-I.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-Mediated Capsicum Annuum Gene Editing in Two Cultivars, Hot Pepper CM334 and Bell Pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef]
- Hou, X.M.; Zhang, H.F.; Liu, S.Y.; Wang, X.K.; Zhang, Y.M.; Meng, Y.C.; Luo, D.; Chen, R.G. The NAC Transcription Factor CaNAC064 Is a Regulator of Cold Stress Tolerance in Peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- Kong, X.M.; Zhou, Q.; Zhou, X.; Wei, B.D.; Ji, S.J. Transcription Factor CaNAC1 Regulates Low-Temperature-Induced Phospholipid Degradation in Green Bell Pepper. J. Exp. Bot. 2020, 71, 1078–1091. [Google Scholar] [CrossRef]
- Ma, X.; Yu, Y.N.; Jia, J.H.; Li, Q.H.; Gong, Z.H. The Pepper MYB Transcription Factor CaMYB306 Accelerates Fruit Coloration and Negatively Regulates Cold Resistance. Sci. Hortic. (Amsterdam) 2022, 295, 110892. [Google Scholar] [CrossRef]
- Zhang, H.X.; Zhu, W.C.; Feng, X.H.; Wei, J. J.H.; Gong, A.M. Transcription Factor CaSBP12 Negatively Regulates Salt Stress Tolerance in Pepper (Capsicum Annuum L). Int J Mol Sci 2020b, 21. [Google Scholar] [CrossRef]
- Zhang, H.; Pei, Y.; Zhu, F.; He, Q.; Zhou, Y.; Ma, B.; Chen, X.; Guo, J.; Khan, A.; Jahangir, M. CaSnRK2. 4-Mediated Phosphorylation of CaNAC035 Regulates Abscisic Acid Synthesis in Pepper (Capsicum Annuum L.) Responding to Cold Stress. Plant J. 2024a, 117, 1377–1391. [Google Scholar] [CrossRef]
- Ma, J.; Wang, L.Y.; Dai, J.X.; Wang, Y.; Lin, D. The NAC-Type Transcription Factor CaNAC46 Regulates the Salt and Drought Tolerance of Transgenic Arabidopsis Thaliana. BMC Plant Biol 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lim, C. W.; Lee, S. C. Role of Pepper MYB Transcription Factor CaDIM1 in Regulation of the Drought Response. Front. Plant Sci. 2022, 13, 1028392. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.; Lim, C. W.; Lee, S. C. GRAS-Type Transcription Factor CaGRAS1 Functions as a Positive Regulator of the Drought Response in Capsicum Annuum. Environ. Exp. Bot. 2022, 198, 104853. [Google Scholar] [CrossRef]
- Guo, W.L.; Wang, S.B.; Chen, R.G.; Chen, B.H.; Du, X.H.; Yin, Y.X.; Gong, Z.H.; Zhang, Y.Y. Characterization and Expression Profile of CaNAC2 Pepper Gene. Front. Plant Sci. 2015b, 6, 755. [Google Scholar] [CrossRef]
- Wang, Y.; Dang, F.; Liu, Z.; Wang, X.; Eulgem, T.; Lai, Y.; Yu, L.; She, J.; Shi, Y.; Lin, J. CaWRKY 58, Encoding a Group I WRKY Transcription Factor of Capsicum Annuum, Negatively Regulates Resistance to Ralstonia Solanacearum Infection. Mol Plant Pathol 2013, 14, 131–144. [Google Scholar] [CrossRef]
- Noman, A.; Hussain, A.; Adnan, M.; Khan, M. I.; Ashraf, M. F.; Zainab, M.; Khan, K. A.; Ghramh, H. A.; He, S. A Novel MYB Transcription Factor CaPHL8 Provide Clues about Evolution of Pepper Immunity Againstsoil Borne Pathogen. Microb. Pathog. 2019, 137, 103758. [Google Scholar] [CrossRef]
- Cai, W.; Yang, S.; Wu, R.; Cao, J.; Shen, L.; Guan, D.; Shuilin, H. Pepper NAC-Type Transcription Factor NAC2c Balances the Trade-off between Growth and Defense Responses. Plant Physiol. 2021, 186, 2169–2189. [Google Scholar] [CrossRef]
- Hussain, A.; Qayyum, A.; Farooq, S.; Almutairi, S. M.; Rasheed, R. A.; Qadir, M.; Vyhnánek, T.; Sun, Y. Pepper Immunity against Ralstonia Solanacearum Is Positively Regulated by CaWRKY3 through Modulation of Different WRKY Transcription Factors. BMC Plant Biol. 2024, 24, 522. [Google Scholar] [CrossRef]
- Oh, S.K.; Baek, K.H.; Park, J. M.; Yi, S. Y.; Yu, S. H.; Kamoun, S.; Choi, D. Capsicum Annuum WRKY Protein CaWRKY1 Is a Negative Regulator of Pathogen Defense. New Phytol. 2008, 177, 977–989. [Google Scholar] [CrossRef]
- Dang, F.; Wang, Y.; She, J.; Lei, Y.; Liu, Z.; Eulgem, T.; Lai, Y.; Lin, J.; Yu, L.; Lei, D. Overexpression of CaWRKY27, a Subgroup Iie WRKY Transcription Factor of Capsicum Annuum, Positively Regulates Tobacco Resistance to Ralstonia Solanacearum in-Fection. Physiol Plant 2014, 150, 397–411. [Google Scholar] [CrossRef]
- Hussain, A.; Li, X.; Weng, Y.; Liu, Z.; Ashraf, M. F.; Noman, A.; Yang, S.; Ifnan, M.; Qiu, S.; Yang, Y.; Guan, D.; He, S. CaWRKY22 Acts as a Positive Regulator in Pepper Response to RalstoniaSolanacearum by Constituting Networks with CaWRKY6, CaWRKY27, CaWRKY40, and CaWRKY58. Int. J. Mol. Sci. 2018, 19, 1426. [Google Scholar] [CrossRef]
- Guo, M.; Zhai, Y.F.; Lu, J.P.; Chai, L.; Chai, W.G.; Gong, Z.H.; Lu, M.H. Characterization of CaHsp70-1, a Pepper Heat-Shock Protein Gene in Response to Heat Stress and Some Regulation Exogenous Substances in Capsicum Annuum L. Int. J. Mol. Sci. 2014, 15, 19741–19759. [Google Scholar] [CrossRef]
- Guo, M.; Lu, J.P.; Zhai, Y.F.; Chai, W.G.; Gong, Z.H.; Lu, M.H. Genome-Wide Analysis, Expression Profile of Heat Shock Factor Gene Family (CaHsfs) and Characterisation of CaHsfA2 in Pepper (Capsicum Annuum L.). BMC Plant Biol. 2015a, 15, 151. [Google Scholar] [CrossRef]
- Dang, F.; Lin, J.; Xue, B.; Chen, Y.; Guan, D.; Wang, Y.; He, S. CaWRKY27 Negatively Regulates H2O2-Mediated Thermotol-Erance in Pepper (Capsicum Annuum). Front Plant Sci 2018, 9. [Google Scholar] [CrossRef]
- Jingyuan, Z.; Xuexiao, Z.; Zhenchuan, M.; Bingyan, X. A Novel Pepper (Capsicum Annuum L.) WRKY Gene, CaWRKY30, Is Involved in Pathogen Stress Responses. J. Plant Biol. 2011, 54, 329–337. [Google Scholar] [CrossRef]
- Hussain, A.; Khan, M. I.; Albaqami, M.; Mahpara, S.; Noorka, I. R.; Ahmed, M. A. A.; Aljuaid, B. S.; El-Shehawi, A. M.; Liu, Z.; Farooq, S.; Zuan, A. T. K. CaWRKY30 Positively Regulates Pepper Immunity by Targeting CaWRKY40 against Ralstonia Solanacearum Inoculation through Modulating Defense-Related Genes. Int. J. Mol. Sci. 2021, 22, 12091. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.F.; Wang, Y.N.; Yu, L.; Eulgem, T.; Lai, Y.; Liu, Z.Q.; Wang, X.; Qiu, A.L.; Zhang, T.X.; Lin, J.; Chen, Y.S.; Guan, D.Y.; Cai, H.Y.; Mou, S.L.; He, S.L. CaWRKY40, a WRKY Protein of Pepper, Plays an Important Role in the Regulation of Tolerance to Heat Stress and Resistance to Ralstonia Solanacearum Infection: CaWRKY40 in Heat andR. Solanacearuminfection. Plant Cell Environ. 2013, 36, 757–774. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, X.; Xiao, L.-D.; Yu, Y.-N.; Yan, H.-L.; Gong, Z.-H. CaWRKY5 Acts as a Negative Regulator in Response to Colleto-Trichumscovillei Infection in Pepper. Plants 2023, 12(10). [Google Scholar]
- Cheng, W.; Wang, N.; Li, Y.; Zhou, X.; Bai, X.; Liu, L.; Ma, X.; Wang, S.; Li, X.; Gong, B.; Jiang, Y.; Azeem, M.; Zhu, L.; Chen, L.; Wang, H.; Chu, M. CaWRKY01-10 and CaWRKY08-4 Confer Pepper’s Resistance to Phytophthora Capsici Infection by Directly Activating a Cluster of Defense-Related Genes. J. Agric. Food Chem. 2024, 72, 11682–11693. [Google Scholar] [CrossRef]
- Feng, X.H.; Zhang, H.X.; Gai, A. M.; Cheng, W.X.; Li, Y. S.B.; Gong, X.X. A Small Heat Shock Protein CaHsp25. 9 Positively Regulates Heat, Salt, and Drought Stress Tolerance in Pepper (Capsicum Annuum L.). Plant Physiol Biochem 2019, 142, 151–162. [Google Scholar] [CrossRef]
- Liu, R.; Song, J.; Liu, S.; Chen, C.; Zhang, S.; Wang, J.; Xiao, Y.; Cao, B.; Lei, J.; Zhu, Z. Genome-Wide Identification of the Capsicum bHLH Transcription Factor Family: Discovery of a Candidate Regulator Involved in the Regulation of Species-Specific Bioactive Metabolites. BMC Plant Biol 2021a, 21. [Google Scholar] [CrossRef]
- Liu, Y.L.; Liu, S.; Xiao, J.J.; Cheng, G.X.; Ul; H. S. Gong, Z.H. CaHSP18. 1a, a Small Heat Shock Protein from Pepper (Capsicum Annuum L) Positively Responds to Heat Drought and Salt Tolerance. Horticulturae 2021a, 7. [Google Scholar] [CrossRef]
- Lee, S. C.; Choi, H. W.; Hwang, I. S.; Choi, D. S.; Hwang, B. K. Functional Roles of the Pepper Pathogen-Induced bZIP Transcription Factor, CAbZIP1, in Enhanced Resistance to Pathogen Infection and Environmental Stresses. Planta 2006, 224, 1209–1225. [Google Scholar] [CrossRef] [PubMed]
- Lim, C. W.; Baek, W.; Lim, S.; Han, S.-W.; Lee, S. C. Expression and Functional Roles of the Pepper Pathogen-Induced bZIP Transcription Factor CabZIP2 in Enhanced Disease Resistance to Bacterial Pathogen Infection. Mol. Plant. Microbe. Interact. 2015, 28, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Gai, W.X.; Ma, X.; Qiao, Y.M.; Shi, B.H.; Ul Haq, S.; Li, Q.H.; Wei, A.M.; Liu, K.K.; Gong, Z.H. Characterization of the bZIP Transcription Factor Family in Pepper (Capsicum Annuum L.): CabZIP25 Positively Modulates the Salt Tolerance. Front. Plant Sci. 2020, 11, 139. [Google Scholar] [CrossRef]
- Hong, J.P.; Kim, W. T. Isolation and Functional Characterization of the Ca-DREBLP1 Gene Encoding a Dehydra-Tion-Responsive Element Binding-Factor-like Protein 1 in Hot Pepper (Capsicum Annuum L. Cv. Pukang). Planta 2005, 220, 875–888. [Google Scholar] [CrossRef]
- Parkash, C.; Kumar, S.; Thakur, N.; Singh, S.; Sharma, B. B. Cabbage: Breeding and Genomics. isvs 2023, 50 (Special), 231–243. [Google Scholar] [CrossRef]
- Jansson, S. Gene-Edited Plants on the Plate: The “CRISPR Cabbage Story.”. Physiol. Plant. 2018, 164, 396–405. [Google Scholar] [CrossRef]
- Ma, C.; Zhu, C.; Zheng, M.; Liu, M.; Zhang, D.; Liu, B.; Li, Q.; Si, J.; Ren, X.; Song, H. CRISPR/Cas9-Mediated Multiple Gene Editing in Brassica Oleracea Var. Capitata Using the Endogenous tRNA-Processing System. Hortic. Res. 2019a, 6, 20. [Google Scholar] [CrossRef]
- Ma, C.; Liu, M.; Li, Q.; Si, J.; Ren, X.; Song, H. Efficient BoPDS Gene Editing in Cabbage by the CRISPR/Cas9 System. Hortic. Plant J. 2019b, 5, 164–169. [Google Scholar] [CrossRef]
- Murovec, J.; Guček, K.; Bohanec, B.; Avbelj, M.; Jerala, R. DNA-Free Genome Editing of Brassica Oleracea and B. Rapa Protoplasts Using CRISPR-Cas9 Ribonucleoprotein Complexes. Front. Plant Sci. 2018, 9, 1594. [Google Scholar] [CrossRef]
- Kaur, C.; Lee, M.; Yao, J.; Nam, D.; Lee, G. J. CRISPR/Cas12-Mediated Detection of Xanthomonas Campestris Pv. Campestris (Race 1 and 4) for Improved Pathogen Diagnosis in Cabbage. Horticulturae Sci. Technol 2023a, 41, 338. [Google Scholar]
- Vicente, J. G.; Holub, E. B. X Anthomonas Campestris Pv. Campestris (Cause of Black Rot of Crucifers) in the Genomic Era Is Still a Worldwide Threat to Brassica Crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, S.; Chen, W.; Zhang, J.; Zhang, L.; Sun, W.; Wang, Z. Plant Dehydrins: Expression, Regulatory Networks, and Protective Roles in Plants Challenged by Abiotic Stress. Int. J. Mol. Sci. 2021, 22, 12619. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Lee, M.; Jeon, Y.; Lee, G. J. Enhancing Black-Rot Resistance in Cabbage Using CRISPR/Cas9 System. Horticulturae Sci. Technol. 2023b, 41, 423. [Google Scholar]
- Zhao, Z.; Qi, Y.; Yang, Z.; Cheng, L.; Sharif, R.; Raza, A.; Chen, P.; Hou, D.; Li, Y. Exploring the Agrobacterium-Mediated Transformation with CRISPR/Cas9 in Cucumber (Cucumis Sativus L.). Mol. Biol. Rep. 2022, 49, 11481–11490. [Google Scholar] [CrossRef]
- Feng, J.; Wang, N.; Li, Y.; Wang, H.; Zhang, W.; Wang, H.; Chai, S. Recent Progress in Genetic Transformation and Gene Editing Technology in Cucurbit Crops. Agronomy (Basel) 2023, 13, 755. [Google Scholar] [CrossRef]
- Hu, B.; Li, D.; Liu, X.; Qi, J.; Gao, D.; Zhao, S.; Huang, S.; Sun, J.; Yang, L. Engineering Non-Transgenic Gynoecious Cucumber Using an Improved Transformation Protocol and Optimized CRISPR/Cas9 System. Mol. Plant 2017, 10(12), 1575–1578. [Google Scholar] [CrossRef]
- Feng, S.; Zhang, J.; Mu, Z.; Wang, Y.; Wen, C.; Wu, T.; Yu, C.; Li, Z.; Wang, H. Recent Progress on the Molecular Breeding of Cucumis Sativus L. in China. Theor. Appl. Genet. 2020, 133, 1777–1790. [Google Scholar] [CrossRef]
- Ma, Z.; Ma, L.; Zhou, J. Applications of CRISPR/Cas Genome Editing in Economically Important Fruit Crops: Recent Advances and Future Directions. Mol. Hortic. 2023, 3, 1. [Google Scholar] [CrossRef]
- Pawełkowicz, M.; Zieniuk, B.; Staszek, P.; Przybysz, A. From Sequencing to Genome Editing in Cucurbitaceae: Application of Modern Genomic Techniques to Enhance Plant Traits. Agriculture 2024, 14, 90. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of Broad Virus Resistance in Non-Transgenic Cucumber Using CRISPR/Cas9 Technology: Virus Resistance in Cucumber Using CRISPR/Cas9. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Dey, S. S.; Singh, S.; Munshi, A. D.; Behera, T. K. Classical Genetics and Traditional Breeding. In Compendium of Plant Genomes; Springer International Publishing: Cham, 2022; pp. 159–183. [Google Scholar] [CrossRef]
- Riu, Y.-S.; Kim, G. H.; Chung, K. W.; Kong, S.-G. Enhancement of the CRISPR/Cas9-Based Genome Editing System in Lettuce (Lactuca Sativa L.) Using the Endogenous U6 Promoter. Plants 2023, 12, 878. [Google Scholar] [CrossRef] [PubMed]
- Bertier, L. D.; Ron, M.; Huo, H.; Bradford, K. J.; Britt, A. B.; Michelmore, R. W. High-Resolution Analysis of the Efficiency, Heritability, and Editing Outcomes of CRISPR/Cas9-Induced Modifications of NCED4 in Lettuce (Lactuca Sativa). G3 (Bethesda) 2018, 8, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Wang, S.; Ning, K.; Chen, Z.; Wang, Y.; Yang, J.; Wang, Q. LsAP2 Regulates Leaf Morphology by Inhibiting CIN-like TCP Transcription Factors and Repressing LsKAN2 in Lettuce. Hortic. Res. 2021, 8, 184. [Google Scholar] [CrossRef]
- Nguyen, C. D.; Li, J.; Mou, B.; Gong, H.; Huo, H. A Case Study of Using an Efficient CRISPR/Cas9 System to Develop Variegated Lettuce. Vegetable Research 2021, 1, 1–10. [Google Scholar] [CrossRef]
- Choi, S. H.; Ahn, W. S.; Jie, E. Y.; Cho, H.-S.; Kim, S. W. Development of Late-Bolting Plants by CRISPR/Cas9-Mediated Genome Editing from Mesophyll Protoplasts of Lettuce. Plant Cell Rep. 2022, 41, 1627–1630. [Google Scholar] [CrossRef]
- Ahmad, F.; Akram, A.; Farman, K.; Abbas, T.; Bibi, A.; Khalid, S.; Waseem, M. Molecular Markers and Marker Assisted Plant Breeding: Current Status and Their Applications in Agricultural Development. Journal of Environmental and Agricultural Sciences 2017, 11, 35–50. [Google Scholar]
- Pejić, I.; Šatović, Z. Konvencionalno i Molekularno Oplemenjivanje Bilja / Molekularno Oplemenjivanje Bilja, Agronomski Fakultet, Zagreb - Pregled Integracije Molekularnih i Klasičnih Metoda. 2022, 3–20.
- Lamichhane, S.; Thapa, S. Advances from Conventional to Modern Plant Breeding Methodologies. Plant Breed. Biotechnol. 2022, 10, 1–14. [Google Scholar] [CrossRef]
- Wanga, M. A.; Shimelis, H.; Mashilo, J.; Laing, M. D. Opportunities and Challenges of Speed Breeding: A Review. Plant Breed. 2021, 140, 185–194. [Google Scholar] [CrossRef]
- Datta, A. Genetic Engineering for Improving Quality and Productivity of Crops. Agric. & Food Secur. 2013, 2, 15. [Google Scholar] [CrossRef]
- Singh, R. K.; Jamir, S. Advancements in vegetable breeding techniques for quality enhancement. Plant Archives 2025, 25, 26–35. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, X.; Li, H.; Zheng, H.; Zhang, J.; Olsen, M. S.; Varshney, R. K.; Prasanna, B. M.; Qian, Q. Smart Breeding Driven by Big Data, Artificial Intelligence, and Integrated Genomic-Enviromic Prediction. Mol. Plant 2022, 15, 1664–1695. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Namdeo, A.; Mahadik, K. Molecular Markers: New Prospects in Plant Genome Analysis. Pharmacognosy reviews 2008, 2. [Google Scholar]
- Nadeem, M. A.; Nawaz, M. A.; Shahid, M. Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; Özkan, H.; Chung, G.; Baloch, F. S. DNA Molecular Markers in Plant Breeding: Current Status and Recent Advancements in Genomic Selection and Genome Editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y. Advances in Molecular Breeding of Vegetable Crops. Horticulturae 2022, 8, 821. [Google Scholar] [CrossRef]
- Kumar, J.; Choudhary, A. K.; Solanki, R. K.; Pratap, A. Towards Marker-Assisted Selection in Pulses: A Review: Towards Marker-Assisted Selection in Pulses. Plant Breed. 2011, 130, 297–313. [Google Scholar] [CrossRef]
- Fu, Y.B.; Dong, Y.B. Genetic Erosion under Modern Plant Breeding: Case Studies in Canadian Crop Gene Pools. In Sustainable Development and Biodiversity; Springer International Publishing: Cham, 2015; pp. 89–104. [Google Scholar]
- Behera, T. K.; Devi, J.; Tiwari, J. K.; Singh, B. K. Vegetable Breeding: Status and Strategies. isvs 2023, 50 (Special), 131–145. [Google Scholar] [CrossRef]
- Song, S.; Ong, E. J. K.; Lee, A. M. J.; Chew, F. T. How Crop Breeding Programs Can Improve Plant Factories’ Business and Environmental Sustainability: Insights from a Farm Level Analysis. Sustain. Prod. Consum. 2024, 44, 298–311. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, H. Development and Prospect of Gene-Edited Fruits and Vegetables. Food Qual. Saf. 2024, 8, fyad045. [Google Scholar] [CrossRef]
- Brennan, J. P.; Martin, P. J.; Brennan, J. P.; Martin, P. J. Developing Cost Functions for a Wheat Breeding Program. Unknown 2006. [CrossRef]
- Morris, M. L.; Bellon, M. R. Participatory Plant Breeding Research: Opportunities and Challenges for the International Crop Improvement System. Euphytica 2004, 136, 21–35. [Google Scholar] [CrossRef]
- Shu, Q.; Wu, D. Rice: Breeding. In Encyclopedia of Food Grains; Elsevier, 2016, 304–310.
- Sharma, R.; Yang, C. J.; Rossi, N.; Irving, E.; Tuffin, A.; Aliki, H.; Powell, W.; Dawson, I. K. Integrating Molecular Genetics with Plant Breeding to Deliver Impact. Plant Physiol. 2025, 198. [Google Scholar] [CrossRef]
Figure 1.
Morphological and phenological traits that contribute to yield heterosis in cucumber, cabbage, and tomato. (A) Key traits associated with yield heterosis. (B) A gynoecious cucumber line developed for commercial production, characterized by a small number of branches. (C) A model of cabbage showing both the aerial view and a cross-section, including the leaves and the head. (D) A cross between a tomato plant with single inflorescences and indeterminate growth and one with compound inflorescences and determinate growth yields an F1 hybrid with earlier fruit production, a higher number of compound inflorescences, and determinate growth [51].
Figure 1.
Morphological and phenological traits that contribute to yield heterosis in cucumber, cabbage, and tomato. (A) Key traits associated with yield heterosis. (B) A gynoecious cucumber line developed for commercial production, characterized by a small number of branches. (C) A model of cabbage showing both the aerial view and a cross-section, including the leaves and the head. (D) A cross between a tomato plant with single inflorescences and indeterminate growth and one with compound inflorescences and determinate growth yields an F1 hybrid with earlier fruit production, a higher number of compound inflorescences, and determinate growth [51].

Figure 2.
Vegetable breeding strategies [42].
Figure 2.
Vegetable breeding strategies [42].

Figure 3.
Crop improvement methods categorized as conventional breeding, advanced breeding, and plant transformation and gene editing. (A) A conventional strategy involving successive backcrossing and visual phenotypic evaluation of (BC) populations. (B) Advanced approaches for screening BC populations using molecular markers and phenomics. (C) Genetic engineering approaches such as Agrobacterium-mediated plant transformation, cis- and transgenesis, and CRISPR/Cas9-mediated genome editing [40].
Figure 3.
Crop improvement methods categorized as conventional breeding, advanced breeding, and plant transformation and gene editing. (A) A conventional strategy involving successive backcrossing and visual phenotypic evaluation of (BC) populations. (B) Advanced approaches for screening BC populations using molecular markers and phenomics. (C) Genetic engineering approaches such as Agrobacterium-mediated plant transformation, cis- and transgenesis, and CRISPR/Cas9-mediated genome editing [40].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



