Submitted:
10 April 2026
Posted:
13 April 2026
You are already at the latest version
Abstract
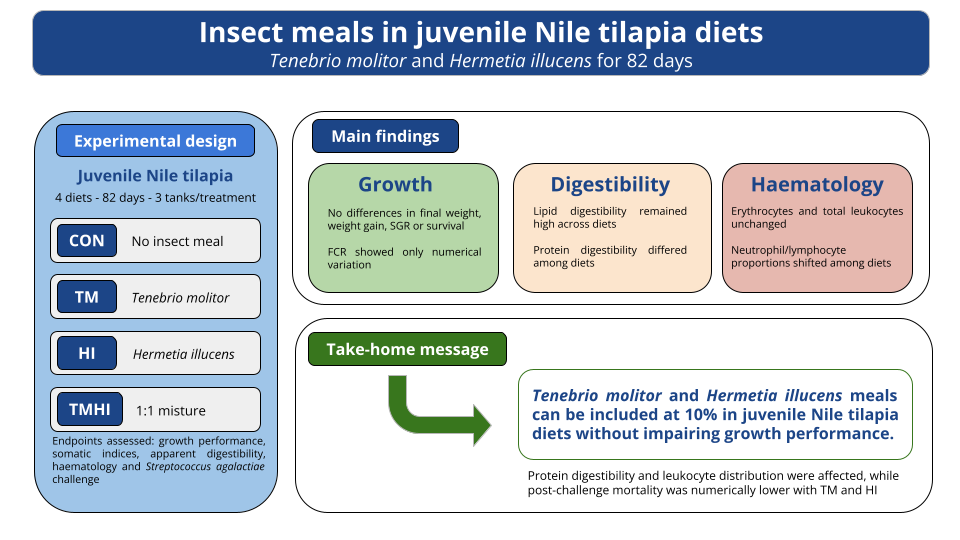
Insect meals are promising alternatives to conventional protein sources in aquafeeds, but comparative evidence in Nile tilapia (Oreochromis niloticus) remains limited. This study evaluated the effects of partially replacing an insect-free control diet with larvae meals from Tenebrio molitor and Hermetia illucens on growth performance, nutrient digestibility, haematological profile, and resistance to Streptococcus agalactiae in juvenile Nile tilapia. Fish were fed isonitrogenous and isoenergetic diets for 82 days: a control diet without insect meal and diets containing 100 g kg⁻¹ dry matter of T. molitor, H. illucens, or a 1:1 mixture. Growth performance and somatic indices were not affected by dietary treatment. Lipid digestibility remained high and similar among diets, whereas protein digestibility differed among diets and insect ingredients. Erythrocyte and total leukocyte counts were unchanged, but neutrophil and lymphocyte proportions differed among treatments. After intraperitoneal challenge with S. agalactiae, cumulative mortality was numerically lower in fish fed T. molitor or H. illucens than in the control group, although survival curves did not differ significantly. These findings indicate that both insect meals can be included at 10% in juvenile Nile tilapia diets without impairing growth, while influencing protein digestibility and leukocyte distribution.
Keywords:
Nile tilapia
; Tenebrio molitor
; Hermetia illucens
; apparent digestibility
; haematological profile
; Streptococcus agalactiae
1. Introduction
Nile tilapia (Oreochromis niloticus) is one of the most widely farmed freshwater fish species worldwide and a cornerstone of affordable animal protein, particularly in low- and middle-income countries. Recent reports from the Food and Agriculture Organization indicate that global aquaculture continues to expand and tilapia ranks among the main contributors to freshwater finfish production, with increasing relevance in emerging producer countries such as Brazil. [1,2]. This rapid growth intensifies the demand for nutritionally adequate, cost-effective and environmentally sustainable feeds, while conventional protein sources such as fishmeal and soybean meal are constrained by price volatility, land and water use, and concerns about environmental footprints. [3,4]. In this context, insect meals have attracted increasing attention as alternative feed ingredients for aquaculture. Several recent reviews highlight that insect larvae can convert low-value organic substrates into high-quality biomass rich in protein, lipids and micronutrients, with a more favorable environmental profile than many conventional ingredients. [4,5,6]. Among the species considered, yellow mealworm (Tenebrio molitor) and black soldier fly (Hermetia illucens) have emerged as leading candidates due to their scalable production, balanced amino acid profile and promising performance in several farmed fish [4,7].
Recent experimental and review studies suggest that T. molitor meal can be used in fish diets without compromising growth, nutrient utilisation or fillet quality when included at appropriate levels [7,8]. In Nile tilapia, dietary inclusion of T. molitor larvae meal has been associated with improved fillet quality traits and modulation of muscle metabolomic profiles, indicating potential benefits beyond simple protein replacement [9]. In addition, data from other species show that T. molitor meal can sustain growth and feed efficiency while affecting tissue lipid composition and antioxidant status, supporting its consideration as a versatile ingredient for aquafeeds [7,8]. The nutritional potential of H. illucens larvae for fish feeding has been even more extensively investigated. Meta-analyses and feeding trials indicate that black soldier fly meal can partially or totally replace fishmeal in diets for several freshwater and marine fish without impairing growth performance when formulations are properly balanced [3,4,7]. In Nile tilapia, replacement of fishmeal by H. illucens larvae meal has been shown to maintain or improve growth, feed utilisation and body condition, while also influencing haematological parameters and mucosal responses [10,11,12]. More recently, studies using defatted H. illucens meal reported improved gut morphology, modulation of intestinal microbiota and enhanced resistance to pathogenic challenge in Nile tilapia, underscoring the relevance of insect meals for fish health as well as productivity [13,14].
Bacterial diseases remain a major bottleneck for sustainable tilapia aquaculture, and streptococcosis caused by Streptococcus agalactiae is among the most impactful conditions worldwide [15,16]. Outbreaks of S. agalactiae are associated with high cumulative mortalities, substantial economic losses and increasing concern about antimicrobial resistance due to the frequent use of antibiotics for disease control [15,17]. While vaccination and biosecurity measures are central to prevention, there is growing interest in nutritional approaches that support innate and adaptive defences and improve survival under experimental challenge. In this regard, insect meals have been shown to influence immune-related biomarkers and survival in fish exposed to bacterial pathogens, including tilapia challenged with Streptococcus iniae [13,14,18].
Although the literature on insect meals in tilapia diets has expanded rapidly, most studies focus on a single insect species, use relatively short feeding periods, or assess only growth-related endpoints. Comparative information on diets containing T. molitor and H. illucens meals under a common experimental design, particularly including digestibility measurements, haematological profiling and survival after S. agalactiae challenge, is still limited. [4,5,8]. Addressing this gap is important to clarify how different insect meals affect nutrient utilisation, health status and disease outcomes in Nile tilapia, thereby guiding their rational inclusion in commercial feeds. Therefore, this study evaluated the effects of diets containing meals derived from T. molitor and H. illucens on the growth performance, nutrient digestibility, haematological profile and survival of Nile tilapia juveniles subjected to experimental S. agalactiae challenge. The findings aim to contribute to the evidence base for the safe and effective use of insect-based ingredients in tilapia aquaculture.
2. Materials and Methods
2.1. Experimental Facilities and Fish
The feeding trial with Nile tilapia (Oreochromis niloticus) was carried out at the Laboratory of Nutrition and Aquaculture of the Fishing Institute (São José do Rio Preto, São Paulo, Brazil). The experimental system comprised 12 cylindrical tanks (310 L each), supplied individually by a recirculating water system equipped with a mechanical filter free of phyto- and zooplankton, a biological filter, thermostatic temperature control (26–28 °C) and continuous aeration, maintaining dissolved oxygen at ≥ 5 mg L⁻¹. Throughout the experimental period, water temperature and dissolved oxygen were monitored daily (Figure 1), whereas pH (6.35 ± 0.19), total ammonia (0.79 ± 0.96 mg L⁻¹) and nitrite (0.69 ± 0.40 mg L⁻¹) were monitored weekly. Filters of the recirculating system were cleaned weekly, and a partial water exchange (10%) was performed to maintain water quality.
Juvenile tilapia (20 g) were acquired from a commercial fish farm; fish were sex-reversed (♂) and not vaccinated, and were acclimated for approximately 20 days before selection for the feeding trial. Each experimental unit was stocked with 20 juvenile tilapia (initial body weight 28.79 ± 2.14 g at stocking). Before stocking, fish were anesthetized (eugenol, 60 mg L⁻¹), previously classified to obtain homogeneous groups, counted, individually weighed and measured, and then randomly allocated to the tanks. The feeding trial with insect-based diets lasted 82 days. An overview of the experimental design and sampling schedule is provided in Figure 2.
All procedures involving fish were approved by the Animal Use Ethics Committee of the Instituto de Pesca (CEUA-IP, São José do Rio Preto, SP, Brazil) and complied with CONCEA guidelines (approval no. CEUA-IP 002/2022).
2.2. Experimental Diets
2.2.1. Chemical Characterization of Insect Meals
The chemical characterization of insect meals was performed at the Food Technology Institute (ITAL, Campinas - SP, Brazil), following standardized protocols under ABNT NBR ISO 9001 certification [19] . Crude protein was determined from total nitrogen according to AOAC Official Method 2001.11 [20], using a nitrogen-to-protein conversion factor of 6.25 (N × 6.25) to report crude protein for comparability with conventional feed-ingredient datasets. Total lipids were quantified by solvent extraction following the AOAC Official Method 920.39 [21], and results were expressed on a dry-matter basis. For fatty acid profiling, extracted lipids were converted to fatty acid methyl esters (FAME) following the AOAC Official Method 969.33 [22]. Amino acid composition was determined after hydrolysis and chromatographic quantification following the AOAC Official Method 982.30 for amino acid profiling [23]. Sulfur amino acids (cystine and methionine) were determined after oxidation/acid hydrolysis according to the AOAC Official Method 994.12 [24], and tryptophan was determined after alkaline hydrolysis according to the AOAC Official Method 988.15 [25]. Amino acids were reported as g 100 g⁻¹ protein, as appropriate for diet formulation and ingredient evaluation.

Table 1.
Crude protein and total lipids (dry matter basis, per 100 g) of insect larvae meals.
| Identification* |
Crude protein (%) (mean ± SD**) |
p |
Total lipids (%) (mean ± SD**) |
p |
| TM | 49.9 ± 0.9 | NR | 35.5 ± 0.2 | NR |
| HI | 57.9 ± 0.2 | NR | 7.7 ± 0.2 | NR |
*TM: Tenebrio molitor larvae meal; HI: Hermetia illucens larvae meal. **SD: standard deviation. NR: not reported (no statistical test performed).
Table 2.
Total fatty acid classes (dry matter basis, g/100 g) of insect larvae meals.
| Identification* | Saturated | Monounsaturated | ω-3 | ω-6 |
| TM | 7.97 | 16.63 | 0.26 | 8.49 |
| HI | 4.87 | 1.38 | 0.02 | 0.71 |
*TM: Tenebrio molitor larvae meal; HI: Hermetia illucens larvae meal.
Table 3.
Amino acid composition (dry matter basis, g/100 g protein) of insect larvae meals.
| Amino acid | TM* | HI* |
| Aspartic acid | 4.15 | 5.56 |
| Glutamic acid | 5.72 | 6.51 |
| Serine | 2.28 | 2.41 |
| Glycine | 3.13 | 3.86 |
| Histidine | 1.50 | 1.83 |
| Arginine | 2.81 | 3.11 |
| Threonine | 2.04 | 2.25 |
| Alanine | 3.59 | 3.50 |
| Proline | 2.85 | 3.03 |
| Tyrosine | 3.79 | 3.78 |
| Valine | 3.24 | 3.56 |
| Methionine | 0.80 | 1.12 |
| Cystine | 0.29 | 0.24 |
| Isoleucine | 2.20 | 2.40 |
| Leucine | 3.68 | 3.91 |
| Phenylalanine | 1.93 | 2.53 |
| Lysine | 2.71 | 3.51 |
| Tryptophan | 0.22 ± 0.00** | 0.42 ± 0.00** |
*TM: Tenebrio molitor larvae meal; HI: Hermetia illucens larvae meal **As reported in the source table.
2.2.2. Diet Formulation and Processing
Four experimental diets were formulated to be isonitrogenous (31.5% crude protein) and isoenergetic (3176–3244 kcal kg⁻¹ digestible energy) to meet the nutritional requirements of the species [26], differing in the inclusion of insect larvae meals at 100 g kg⁻¹ diet (dry matter basis): a control diet without insect ingredients (CON), a diet containing Tenebrio molitor larvae meal (TM), a diet containing Hermetia illucens larvae meal (HI), and a diet containing a 50:50 blend of T. molitor and H. illucens larvae meals (TMHI) (Table 4). The diets were formulated using conventional plant and rendered animal ingredients (e.g., soybean meal, poultry by-product meal, meat and bone meal, and blood meal) and common cereal energy sources (e.g., corn, broken rice, wheat bran), with ingredient proportions adjusted across treatments to maintain the target nutrient levels (Table 4).
To approximate field-like conditions, ingredients were sourced from commercial feed manufacturing supply chains; diets were ground through a 0.7-mm screen (hammer mill) and extruded as 3-mm pellets in a commercial extruder (Ferraz® 62-e; ~80 kg h⁻¹).
2.2.3. Growth Performance and Somatic Indices
At the beginning and end of the feeding period, fish from each tank were anesthetized (eugenol, 60 mg L⁻¹), individually weighed and measured. The following growth performance indicators were calculated at the tank level using standard equations for fish growth studies: feed intake, weight gain (WG), specific growth rate (SGR), feed conversion ratio (FCR), protein efficiency ratio (PER) and survival. Fish were fed four times daily to apparent satiation (ad libitum), avoiding feed leftovers in the water. Feed intake per tank was quantified by weighing feed containers when full and when leftovers were observed after a few days of feeding, followed by refilling with the experimental diets and reweighing. For the determination of somatic indices, three fish per replicate were euthanized at the end of the growth trial. Euthanasia was performed by eugenol overdose (200 mg L⁻¹). Fish were dissected and viscera, liver and visceral fat were weighed for calculation of:
2.2.4. Apparent Digestibility
A digestibility trial was conducted in parallel with the growth trial in a separate set of tanks. While insect meals were included at 100 g kg⁻¹ (dry matter) in the growth trial, ingredient digestibility was estimated using the substitution method, in which test diets contained 30% (dry matter) of each insect ingredient and 70% of a reference diet [27] (Bureau et al., 1999). A total of 360 juvenile Nile tilapia (mean body weight: 47.81 ± 7.13 g) were anesthetized with eugenol (60 mg L⁻¹), sorted to obtain a homogeneous group, weighed, counted, and randomly distributed into twelve 300-L tanks (30 fish per tank). Water temperature (27.91 ± 1.32 °C) and dissolved oxygen (4.86 ± 0.64 mg L⁻¹) were monitored daily using a YSI 55 probe.
The reference diet used the same formulation as the control diet (Table 4). Test diets consisted of 70% of the reference diet and 30% of the test insect ingredient (TM, HI, or TMHI; dry-matter basis). All diets contained 0.5% chromium(III) oxide (Cr₂O₃) as an inert marker for the indirect digestibility method, replacing corn in the formulation.
After a 7-day feeding period with the experimental diets, fish were transferred to feces-collection aquaria. Feces were repeatedly collected at 1-h intervals to minimize nutrient leaching and stored frozen until analysis. Feces were collected using a modified Guelph system consisting of cylindrical, conical-bottom tanks (120 L) fitted with a lower ball valve coupled to Falcon-type tubes for sedimented feces recovery [28]. Samples were dried in a forced-air oven at 55 °C for 48 h, ground with a mortar and pestle, separated from scales, and submitted for laboratory analyses.
After determining Cr₂O₃, crude protein and ether extract in the reference diet, test diets and feces, apparent digestibility coefficients (ADC) of the diets were calculated according to Nose [29]:
where and are chromium oxide concentrations in diet and feces, respectively, and and are the concentrations of the nutrient of interest (crude protein or ether extract) in diet and feces.
Digestibility of the test insect ingredients was calculated using the equation described by Bureau et al. [27]:
where ADCI is the ADC of the test ingredient; ADCT is the ADC of the test diet; ADCR is the ADC of the reference diet; DR is the % nutrient of the reference diet mash; DI is the % nutrient of the test ingredient; 0.3 is the proportion of the test ingredient in the test diet mash; and 0.7 is the proportion of the reference diet mash in the test diet mash.
2.2.5. Haematological Analyses
Haematological sampling was performed immediately after the growth trial. Two fish were randomly selected from each tank (six fish per diet). Blood was collected by caudal vessel puncture using 3-mL disposable syringes, and aliquots were transferred to 500-µL microtubes containing EDTA as anticoagulant. Erythrocyte counts, haematocrit, total leukocyte and thrombocyte estimates, and differential leukocyte counts (neutrophils, lymphocytes, monocytes, eosinophils and basophils) were determined. Red blood cells were counted in a Neubauer chamber, and haematocrit was determined using the microhaematocrit method described by Goldenfarb et al. [30].
Blood smears were prepared, air-dried, stained, and examined under light microscopy for differential leukocyte counts, with at least 200 leukocytes counted per slide. Total leukocyte counts were estimated using an indirect smear-based method by counting approximately 2,000 erythrocytes per smear and deriving leukocyte numbers from the leukocyte:erythrocyte ratio. Differential leukocyte counts were performed on smears stained with Rosenfeld stain.
2.2.6. Streptococcus Agalactiae Challenge
Before the main challenge test, a preliminary trial was conducted to determine the median lethal dose (LD₅₀) of a pathogenic Streptococcus agalactiae strain in Nile tilapia through intraperitoneal injection of 100 µL bacterial suspensions at graded concentrations. For the bacterial preparation, the isolate was plated on Brain Heart Infusion (BHI) agar supplemented with 5% blood (30 °C, 24 h); colonies were suspended in sterile 0.85% saline to an optical density of 0.330 at 600 nm (stock suspension), followed by 10-fold serial dilutions and CFU confirmation by plate counts.
Fish were anesthetized in buckets with eugenol (15 mg L⁻¹) before injection; 100 µL of inoculum was injected intraperitoneally using insulin syringes, and fish were returned to tanks until full recovery of normal activity. Mortality was recorded daily until three consecutive days without deaths were observed, and LD₅₀ was defined as the dose producing ~50% cumulative mortality. The LD₅₀ was determined as 1.65 × 10⁵ CFU.
In the main challenge assay, fish previously fed the experimental diets were injected intraperitoneally with 100 µL of the S. agalactiae suspension at the LD₅₀ dose determined in the preliminary test (1.65 × 10⁵ CFU). Two independent challenge blocks were conducted under the same protocol on different dates (Infection 1 and Infection 2). A negative control group (CN; derived from the CON treatment) received sterile 0.85% saline injection, and an infected positive control group (CP) was included to verify challenge validity. Mortality was monitored daily throughout the observation period, and cumulative mortality (%) was calculated as (cumulative deaths/total fish) × 100. The infection course typically ended around day 10 post-inoculation, defined as three consecutive days without mortality. Potential block effects were assessed by comparing final mortality proportions between Infection 1 and Infection 2; as no evidence of block differences was detected, data were pooled for presentation and analysis (Supplementary Table S1).
2.2.7. Statistical Analysis
Data on growth performance, somatic indices and apparent digestibility were analysed by one-way analysis of variance (ANOVA) with dietary treatment as the fixed factor, using the tank as the experimental unit. When significant differences were detected (P < 0.05), means were compared using Tukey’s multiple comparison test. Haematological variables were analysed using a mixed model in which dietary treatment was included as a fixed effect and tank (nested within diet) as a random effect, with fish treated as subsamples within tank. When significant effects were detected (P < 0.05), means were compared using Tukey’s test. Statistical analyses were performed using SAS software, version 9 (SAS Institute Inc., Cary, NC, USA).
Post-challenge survival was analysed using Kaplan–Meier time-to-event curves based on daily mortality records, and differences among dietary treatments were tested using the log-rank test (P < 0.05). Potential challenge-block effects (Infection 1 vs Infection 2) were assessed by comparing final mortality proportions between blocks; as no evidence of block differences was detected, data were pooled for presentation and analysis (Supplementary Table S1). Cumulative mortality was additionally summarized descriptively as the proportion of dead fish per treatment over the 13-day observation period.
3. Results
3.1. Growth Performance
Growth performance data for Nile tilapia are summarized in Table 5. No statistically significant differences among diets were detected for initial body weight (P = 0.2802), final body weight (P = 0.7388), weight gain (P = 0.7464), specific growth rate (SGR; P = 0.7110) or feed intake (P = 0.3043). Initial body weight ranged from 28.28 ± 0.28 g (TM) to 29.22 ± 0.32 g (TMHI), whereas final body weight ranged from 248.31 ± 13.31 g (CON) to 260.36 ± 6.35 g (TMHI) (Table 5).
Feed conversion ratio (FCR) did not differ among diets (P = 0.0525), although fish fed the H. illucens diet (HI) showed a numerically higher (i.e., poorer) FCR (1.25 ± 0.04) than T. molitor diet (TM) (1.15 ± 0.05) and TMHI (1.17 ± 0.02), while CON presented intermediate values (1.20 ± 0.03) (Table 5). Overall, partial replacement of the control diet by Tenebrio molitor and/or Hermetia illucens larvae meals at 100 g kg⁻¹ diet did not impair growth performance of Nile tilapia juveniles under the present experimental conditions.
3.2. Survival and Somatic Indices
Survival and somatic indices are presented in Table 6. Survival was high across treatments (93.3 ± 5.8% to 100.0 ± 0.0%) and did not differ among diets (P = 0.7797). Condition factor (P = 0.3832), viscerosomatic index (VSI; P = 0.8487) and hepatosomatic index (HSI; P = 0.5478) were also not affected by dietary treatment (P > 0.05), with comparable mean values among CON, TM, HI and TMHI (Table 6).
Liposomatic index (LSI) did not differ among diets (P = 0.0544), although fish fed the mixed-insect diet (TMHI) showed a numerically higher LSI (2.40 ± 0.06) than fish fed HI (1.72 ± 0.49), while CON (1.95 ± 0.34) and TM (2.23 ± 0.32) presented intermediate values (Table 6).
3.3. Apparent Digestibility Coefficients
Apparent digestibility coefficients for crude protein (ADC-CP) and ether extract (ADC-EE) of the experimental diets and insect ingredients are shown in Table 7. Ingredient digestibility values were estimated using the substitution method (test diets containing 30% insect ingredient and 70% reference diet). Dietary treatment significantly affected diet ADC-CP (P = 0.0134), with fish fed TM showing a lower ADC-CP (83.60 ± 1.87%) than the other treatments, whereas CON (86.95 ± 0.71%), HI (87.31 ± 0.87%) and TMHI (85.71 ± 0.42%) did not differ from each other (Table 7). In contrast, diet ADC-EE did not differ among treatments (P = 0.9132), ranging from 86.76 ± 1.66% (HI) to 88.95 ± 0.91% (TMHI) (Table 7).
At the ingredient level, ADC-CP differed among insect meals (P = 0.0265). H. illucens meal showed the highest value (88.16 ± 2.90%), T. molitor meal the lowest (75.79 ± 6.22%), and the mixed insect meal (TMHI) intermediate digestibility (82.83 ± 1.41%) (Table 7). Ingredient ADC-EE did not differ among insect meals (P = 0.8343), with values of 88.70 ± 4.73% (TM), 85.69 ± 5.52% (HI) and 93.48 ± 2.93% (TMHI) (Table 7). Overall, lipid digestibility was high across insect meals, whereas protein digestibility ranged from 75.79 to 88.16% under the present experimental conditions.
3.4. Haematological Parameters
Red blood cell (erythrocytes) and total leukocyte counts did not differ among dietary treatments (P = 0.3332 and P = 0.3616, respectively; Table 8). However, differential leukocyte counts showed a dietary effect on the relative proportions of neutrophils (P = 0.0247) and lymphocytes (P = 0.0172) (Table 9). Fish fed the control diet exhibited a higher percentage of neutrophils (32.33 ± 5.75%) and a lower percentage of lymphocytes (55.33 ± 3.62%) compared with fish fed HI (22.50 ± 5.57% neutrophils; 66.33 ± 6.41% lymphocytes) and TMHI (21.83 ± 8.11% neutrophils; 65.00 ± 7.77% lymphocytes), whereas TM showed intermediate values (Table 9). No significant differences were detected for monocytes (P = 0.5101), eosinophils (P = 0.8832) or basophils (P = 0.0628) (Table 9).
3.5. Streptococcus Agalactiae Challenge and Mortality
Cumulative mortality after experimental challenge with Streptococcus agalactiae is shown in Figure 2 and summarized in Table 10 and Table 11. Mortality data are presented as pooled outcomes from two independent challenge blocks (Infection 1: n = 24 fish per diet; Infection 2: n = 22 fish per diet; total n = 46 fish per diet) conducted under the same protocol. Final mortality proportions were comparable between Infection 1 and Infection 2 (P > 0.05), supporting pooling of both challenge blocks (Supplementary Table S1). The observation period lasted 13 days post-challenge (Table 10). Kaplan–Meier survival curves derived from daily mortality records did not detect differences among dietary treatments (log-rank test, P = 0.592), and thus between-group differences in cumulative mortality are interpreted descriptively under the present experimental conditions. The negative control (CN) showed 4/46 deaths (8.7%), whereas the positive control (CP) showed 30/46 deaths (65.2%) over the observation period (Supplementary Table S1). Cumulative mortality was highest in fish fed the control diet (CON: 65.2%, 30/46; 95% CI: 50.8–77.3), corresponding to a survival of 34.8% (Table 10). Among the insect-based diets, cumulative mortality was numerically lower in HI (50.0%, 23/46; 95% CI: 36.1–63.9) and TM (52.2%, 24/46; 95% CI: 38.1–65.9), whereas TMHI showed an intermediate value (60.9%, 28/46; 95% CI: 46.5–73.6) (Table 10). The temporal pattern of deaths is provided in Table 11. Most mortality occurred during the early post-challenge phase, particularly between days 3 and 5, after which deaths became sporadic and cumulative mortality curves approached a plateau toward the end of the observation period (Figure 3; Table 11).
4. Discussion
4.1. Growth Performance and Somatic Responses
The present study showed that partial replacement of the control diet by Tenebrio molitor and/or Hermetia illucens larvae meals at 100 g kg⁻¹ diet did not impair growth performance of Nile tilapia juveniles under the present experimental conditions. No statistically significant differences among diets were detected for final body weight, weight gain, specific growth rate (SGR), or feed intake (P > 0.05) (Table 5). Feed conversion ratio (FCR) also did not differ among diets (P = 0.0525), although fish fed HI showed a numerically higher (poorer) FCR than fish fed TM and TMHI, while CON presented intermediate values (Table 5). Taken together, these results indicate that including TM and/or HI at 10% of diet dry matter can maintain productive performance when diets are properly formulated, as broadly reported for insect-derived ingredients in aquafeeds [31,32].
It is also noteworthy that overall zootechnical performance was strong across all treatments (SGR approximately 2.76–2.82% day⁻¹; FCR approximately 1.15–1.25; survival 93.3–100.0%). This pattern is consistent with the broader literature showing that insect meals—particularly T. molitor and H. illucens—can be incorporated into tilapia feeds without compromising growth and feed utilization at appropriate inclusion levels and balanced formulations [4,10,33,34,35]. In Nile tilapia, for instance, H. illucens larvae meal has been tested as a fishmeal replacer without detrimental effects on growth and feed utilization in controlled feeding trials [10,33], and graded substitution designs further support feasibility when diets are nutritionally balanced [34]. Likewise, studies using T. molitor in tilapia diets (including biofloc systems) support the feasibility of inclusion without impairing productive performance when nutritional constraints are addressed and formulations remain adequate [36], while digestibility-focused trials in tilapia fingerlings indicate that T. molitor can present high apparent digestibility among insect meals [37]. Beyond productive performance, dietary T. molitor has also been associated with improvements in tilapia meat quality and metabolomic signatures in a supplementation context, suggesting that insect ingredients may modulate product quality traits depending on formulation strategy and inclusion level [9]. Overall, the absence of clear performance effects in the present trial suggests that, at 100 g kg⁻¹ inclusion, TM and HI functioned primarily as nutritionally compatible substitutes rather than as performance-enhancing additives [4], consistent with broader syntheses of insect meal use in fish feeds [35,38].
Somatic indices further support the conclusion that insect meal inclusion at 100 g kg⁻¹ did not induce major alterations in organ allocation or energy reserves. Survival, condition factor, viscerosomatic index (VSI), and hepatosomatic index (HSI) were not affected by dietary treatment (P > 0.05), indicating broadly comparable somatic responses among diets (Table 6), which is consistent with tilapia trials using H. illucens meal and reporting stable body condition under practical formulations [10,33]. Liposomatic index (LSI) did not differ among diets (P = 0.0544), although TMHI showed a numerically higher mean LSI than HI, with CON and TM presenting intermediate values (Table 6). Because the experimental diets were formulated to be isoenergetic but were not strictly isolipidic, variation in LSI may reflect differences in dietary lipid supply and lipid source characteristics (e.g., insect lipid profiles and the fatty acid composition of added oils) (Table 1; Table 2). Insect-derived lipids and added oils can differ markedly in fatty acid profile (e.g., saturation degree and medium-chain fatty acids), which may influence lipid deposition patterns and visceral fat partitioning depending on inclusion level and the ingredients replaced [31,32,39]. Under this context, the small LSI shift observed here should be interpreted cautiously and remains descriptive, as it did not meet the significance threshold at α = 0.05.
Overall, the growth and somatic responses observed here add to evidence that TM and HI meals are technically feasible ingredients in Nile tilapia feeds at low-to-moderate inclusion levels, provided that diets are formulated to meet nutrient requirements and maintain adequate amino acid balance and digestible energy [4,10,34,36].
4.2. Nutrient Digestibility of Tenebrio Molitor and Hermetia Illucens Meals
The apparent digestibility coefficients (ADC) obtained in the present study indicate that both insect meals supported high nutrient utilization by Nile tilapia. Protein digestibility showed greater sensitivity to diet/ingredient differences than lipid digestibility, a pattern frequently reported when comparing protein-rich ingredients that differ in non-protein fractions and processing history [37,40]. Notably, insect meals were included at 100 g kg⁻¹ in the growth trial, whereas ingredient ADC values were estimated using the substitution method with test diets containing 30% insect ingredient (70:30 reference:test) [27]. At the diet level, ADC-CP differed among treatments (P = 0.0134), with the TM diet showing lower protein digestibility (83.60 ± 1.87%) than CON (86.95 ± 0.71%), HI (87.31 ± 0.87%) and TMHI (85.71 ± 0.42%), which did not differ from each other. In contrast, diet ADC-EE did not differ among treatments (P = 0.9132) and remained high (86.76 ± 1.66% to 88.95 ± 0.91%). These results suggest that Nile tilapia maintained a consistent capacity to digest dietary lipids across formulations, consistent with the generally high lipid digestibility reported for fish and with studies showing strong utilization of dietary lipids across contrasting energy sources in tilapia [41,42]. From a practical standpoint, the lower ADC-CP observed for the TM diet did not translate into impaired growth performance at the 10% inclusion level, suggesting that—under adequate formulation—overall digestible nutrient supply remained sufficient to sustain productive outcomes despite modest differences in apparent protein utilization (Table 1) [10,43].
Across treatments, ADC-EE values were generally comparable to, and often numerically higher than, ADC-CP, consistent with the high digestibility of lipids in fish [40] (Santinha et al., 1996) and with tilapia studies evaluating insect-derived ingredients, including H. illucens meal inclusion [10,43]. In the HI diet, ADC-CP (87.31 ± 0.87%) was slightly higher than ADC-EE (86.76 ± 1.66%), but both coefficients remained within ranges reported for practical aquafeeds, including diets containing insect meals [10,37].
When digestibility was estimated at the ingredient level (substitution method), ingredient ADC-CP also differed among insect meals (P = 0.0265). The HI meal showed the highest ADC-CP (88.16 ± 2.90%), exceeding TM (75.79 ± 6.22%), while TMHI was intermediate (82.83 ± 1.41%) and did not differ from either HI or TM. In contrast, ingredient ADC-EE did not differ among insect meals (P = 0.8343) and remained high, with TMHI showing the highest numerical value (93.48 ± 2.93%, compared with 88.70 ± 4.73% for TM and 85.69 ± 5.52% for HI). Together, these findings reinforce that protein digestibility is a key axis of variability among insect ingredients, whereas lipid utilization may remain consistently high across insect sources when formulations are balanced, as previously observed in tilapia digestibility screening assays [36] (Fontes et al., 2019) and in feeding trials including H. illucens meal [10,43]. The intermediate ingredient ADC-CP for TMHI suggests that blending the two insect sources may buffer differences in protein digestibility, which may be relevant for formulation strategies aiming at consistent nutrient utilization when ingredient quality varies across batches and processing routes [40].
Variation in insect-meal protein digestibility has been linked to differences in composition and processing, including non-protein fractions such as chitin, degree of defatting, and physicochemical properties that can alter nutrient accessibility and enzyme–substrate interactions [44,45]. In Nile tilapia, chitin can be digested to some extent but may also reduce nutrient digestibility depending on inclusion level and how it is presented, with evidence of negative relationships between chitin-related fractions and nutrient digestibility under some conditions [45]. From a practical standpoint, the present TM values remain compatible with reports that mealworm-based ingredients can provide moderate-to-high protein digestibility in fish, often exceeding 70% depending on processing and formulation [36,46]. Therefore, the lower ADC-CP observed for TM in the present trial does not preclude its application, but it underscores the importance of ingredient characterization and formulation adjustments (e.g., amino acid balancing (Table 3) and consideration of processing effects) when using specific insect meals, as recommended in ingredient-evaluation frameworks for aquaculture feeds [40].. Finally, alongside broader sustainability perspectives, the consistently high lipid digestibility and overall favorable nutrient utilization observed here support continued evaluation of insect meals as sustainable ingredients in tilapia feeds, while acknowledging that environmental outcomes depend on production assumptions and supply-chain parameters [47].
4.3. Haematological Responses and Immunomodulation
Haematological profiles are widely used as practical indicators of fish health and can reflect nutritional adequacy as well as aspects of immune status [48]. In the present study, erythrocyte counts and total leukocyte counts were not affected by dietary treatment (P > 0.05; Table 8), indicating that inclusion of TM and HI meals at 100 g kg⁻¹ did not compromise basic haematological homeostasis in Nile tilapia juveniles. Comparable outcomes have been reported for Nile tilapia fed T. molitor meal under intensive rearing conditions, where baseline haematological parameters remained within physiological ranges at similar inclusion levels [36]. Similar patterns in core haematological variables have also been described in tilapia trials using black soldier fly meal under balanced formulations [10].
Although total leukocyte counts did not differ among treatments (Table 8), the differential leukocyte profile (Table 9) showed diet-related differences in the relative proportions of neutrophils and lymphocytes. Neutrophil percentage differed among diets (P = 0.0247), with CON presenting higher values than HI and TMHI, while TM was intermediate and did not differ from the other groups. Conversely, lymphocyte percentage also differed among diets (P = 0.0172), with HI and TMHI presenting higher values than CON, and TM again showing an intermediate profile. No significant differences were observed for monocytes or eosinophils (P > 0.05). Basophil percentage also did not differ among diets (P = 0.0628) (Table 9).
Importantly, the diet-associated differences in neutrophils and lymphocytes occurred without changes in total leukocyte counts, suggesting a redistribution of leukocyte subsets rather than generalized leukocytosis or leukopenia. In practical terms, this pattern is more consistent with immunomodulation under basal conditions than with overt haematological disruption. This interpretation aligns with syntheses indicating that insect-derived ingredients can influence immune-related endpoints without necessarily causing adverse shifts in baseline haematological variables [4,49]. In tilapia, insect-based diets have been associated with changes in mucosal innate immunity (e.g., skin mucus indicators) alongside otherwise stable baseline haematology, supporting the view that insect meals may modulate immune function through coordinated systemic and mucosal pathways [5,10].
Such patterns are compatible with the broader literature proposing that insect meals may act as mild immunomodulators, partly due to non-protein fractions and bioactive components (e.g., chitin/chitosan-related fractions) that can interact with immune signaling and mucosal physiology [4,44,49,50]. In addition, insect meals may contain other bioactive fractions, and black soldier fly-derived ingredients typically provide higher lauric acid than many conventional feed ingredients, which could plausibly influence host–microbe interactions and immune dynamics depending on processing and inclusion level [5,49,51].
Nevertheless, because the present study did not include functional immune assays (e.g., lysozyme, complement, respiratory burst), oxidative-stress biomarkers, cytokine/gene-expression profiling, or microbiota analyses, the current evidence should be interpreted as haematological support for possible immune modulation rather than definitive proof of improved immune competence [5,49]. Moreover, differential leukocyte percentages do not necessarily translate into functional protection against pathogens and are best interpreted alongside challenge outcomes and mechanistic endpoints. Future work combining differential leukocyte profiles with functional and molecular assays would help clarify the mechanisms and biological relevance of the leukocyte shifts observed here [4,44], including integrated assessment of mucosal immunity and microbiota responses [5].
4.4. Resistance to Streptococcus Agalactiae
Streptococcosis caused by Streptococcus agalactiae is among the most relevant bacterial diseases affecting Nile tilapia production, being frequently associated with acute outbreaks, high mortality and substantial economic losses to producers[52,53]. Beyond pathogen factors, host outcomes are strongly modulated by farming conditions (e.g., temperature and stocking density), pathogen strain/virulence and the baseline immune status of the fish, which helps explain why mortality can vary considerably among studies and production settings [52,54]. Consequently, nutritional strategies that may support robustness and post-challenge survival are of practical interest as complementary tools alongside vaccination, biosecurity and husbandry adjustments [53,55].
In the present study, pre-challenge feeding with insect-based diets (TM, HI, TMHI) was associated with numerically lower cumulative mortality after S. agalactiae challenge compared with the control diet (CON). However, Kaplan–Meier analysis did not detect statistically significant differences among treatments (log-rank test, P = 0.592); therefore, the observed between-group differences should be interpreted descriptively under the present experimental conditions. Still, the magnitude of the numerical differences—approximately 13–15 percentage points lower cumulative mortality in HI and TM relative to CON (65.2% in CON versus 50.0% in HI and 52.2% in TM), with TMHI intermediate (60.9%)—may warrant confirmation in adequately powered trials. From an applied perspective, even modest reductions in cumulative mortality can translate into meaningful gains in harvestable biomass and lower unit production costs, especially in intensive systems where disease events can rapidly erode margins [53,56,57]. Farm-level economic appraisals of streptococcosis control strategies explicitly model profitability under different mortality scenarios and illustrate how shifts in cumulative mortality can materially change economic outcomes at the production-cycle level [53,56].
The descriptive pattern observed here is directionally consistent with reports that insect meals can influence health-related endpoints and, in some contexts, challenge outcomes. For example, defatted HI meal has been associated with modulation of hemato-immunological, antioxidant and inflammatory-related endpoints in experimental streptococcosis models, including gene-expression shifts consistent with immunophysiological support during infection [14]. Although that work used Streptococcus iniae rather than S. agalactiae, both agents cause systemic streptococcosis in tilapia and share overlapping host-response pathways, supporting cautious mechanistic parallels [52].
In addition, evidence across fish models indicates that T. molitor meals can modulate innate immune effectors and antibacterial activities that may be relevant to bacterial-disease outcomes. Feeding European sea bass diets containing T. molitor larvae meal increased lysozyme antibacterial activity and altered humoral inflammatory-related markers (e.g., myeloperoxidase, nitric oxide), consistent with mild immunostimulation rather than immunosuppression [58]. Likewise, in juvenile yellow catfish, graded inclusion of T. molitor meal enhanced multiple immune and antioxidant indicators and was evaluated in the context of bacterial challenge responses, supporting the concept that mealworm-based formulations can influence host defense readiness under infection pressure [59]. In other fish, T. molitor inclusion has also been associated with changes in lysozyme activity and antioxidant enzyme activity (e.g., glutathione peroxidase), aligning with an “immune-support” interpretation that could be compatible with the numerically improved survival observed for TM in the present work [60].
More broadly, nutritional interventions (e.g., organic acids, probiotics/synbiotics, phytogenics) have been explored as non-antibiotic strategies to mitigate streptococcosis outcomes in Nile tilapia, reinforcing that diet can influence post-challenge survival even when effects are moderate [55,61]. In this context, the numerical mortality reduction observed in TM and HI relative to CON, together with the diet-associated leukocyte redistribution observed pre-challenge (higher lymphocyte proportions and lower neutrophil proportions in some insect-fed groups), is consistent with the broader idea that insect meals can modulate baseline immune status [4,49]. However, because the present study did not measure lysozyme/complement activity, oxidative stress, cytokine expression, microbiota composition, pathogen load, or tissue-level pathology, any mechanistic links between diet composition, leukocyte distribution and survival outcomes remain hypothetical.
Mechanistically, the observed pattern could reflect combined effects of: (i) high nutrient digestibility and adequate digestible nutrient supply supporting immune competence; and (ii) immunomodulatory components present in insect meals (e.g., chitin/chitosan-related fractions and other non-protein components) that may interact with mucosal immunity and inflammatory pathways [4,44]. In addition, HI-derived ingredients typically contain higher lauric acid than many conventional feed ingredients, and lauric acid has documented antimicrobial activity that may influence gut microbial ecology and immune–microbe interactions depending on processing and inclusion level [62]. Experimental evidence also indicates that lipids extracted from HI can show antimicrobial activity, supporting the plausibility that HI-associated lipid fractions could contribute to antimicrobial pressure in the intestinal environment [63].
A key observation in the present dataset is that TMHI showed an intermediate outcome (60.9% mortality), numerically higher than TM and HI alone. This does not imply antagonism, because the experiment was not powered to resolve small-to-moderate differences among insect diets, and the survival curves did not differ statistically (P = 0.592). Nevertheless, several non-mutually exclusive hypotheses may be considered. First, blending two insect sources at a fixed inclusion level could yield an intermediate profile if functional fractions do not scale linearly across blends. For example, if HI-associated lipid fractions (including lauric acid) contribute to antimicrobial pressure in the gut environment [62,63], a 50:50 blend could attenuate that specific signal relative to HI alone. Second, matrix effects in the gut (microbiota and mucosa) may differ for blends versus single-source meals. HI contains bioactive compounds (including lauric acid and chitin) that can affect fish gut microbiota and intestinal responses, while insect-meal–driven shifts in immune and barrier endpoints can be context-dependent (e.g., defatting level, chitin fraction, and background diet) [44,49,62]. A blend could therefore generate a different non-protein substrate profile and a different microbiota trajectory than either insect alone, which might be neutral, beneficial, or less favorable depending on background diet and processing. Third, recent work in other fish species has tested mixtures of HI and TM meals and indicates that responses can vary with inclusion level and endpoints measured, supporting the general point that synergy should not be assumed [64].
Accordingly, the present evidence supports a descriptive association between insect feeding and mortality patterns, but not a definitive causal improvement in resistance to S. agalactiae, nor a definitive explanation for the intermediate TMHI response. Future work should therefore combine challenge trials with integrated immune and microbiological endpoints (e.g., lysozyme/complement activity, mucosal markers, gut histology, microbiota profiling and pathogen load), and—given the economic relevance of mortality—should also consider powering experiments to detect smaller but commercially meaningful differences in survival [53,56].
5. Conclusions
This study demonstrates that replacing an insect-free control diet with Tenebrio molitor and/or Hermetia illucens larvae meals at 100 g kg⁻¹ (dry matter) is nutritionally feasible for juvenile Nile tilapia. At this inclusion level, growth performance and major somatic indices were maintained. Feed conversion ratio and liposomatic index did not differ among diets (P > 0.05), although numerical variation was observed. Digestibility results further supported the technical suitability of both ingredients, showing consistently high lipid digestibility, while protein digestibility differed among diets and insect ingredients; nevertheless, overall digestible nutrient supply remained sufficient to sustain growth under the present experimental conditions. Haematological profiles were largely stable, and post-challenge outcomes should be interpreted descriptively, with numerically lower cumulative mortality in TM and HI than in the control.
It is important to notice that the observed shifts in leukocyte distribution in insect-fed fish—occurring without changes in total leukocyte counts—are consistent with mild immunomodulation rather than haematological disruption. Such modulation can be relevant in intensive, large-scale production, where fish are continuously exposed to fluctuating environmental and microbial pressures, and small changes in baseline immune readiness may influence population-level robustness. However, the biological meaning of these shifts and their relationship with disease outcomes requires confirmation through adequately powered trials integrating functional immune markers, oxidative-stress endpoints, gut histology, microbiota profiling and pathogen-load measurements.
Finally, both T. molitor and H. illucens meals appear to be strong candidates as ingredients for tilapia feeds. The choice between them has to be guided less by “which insect is better” and more by business-specific feasibility. In large-scale production, supply-chain availability plays an important role, as do local regulations, environmental constraints, and economic feasibility (e.g., the costs of energy, water, and rearing substrates, including organic residues, and processing infrastructure). From this perspective, the use of insect meals has to be evaluated as a regionalised solution within aquafeed systems. Consequently, future work should couple biological performance with techno-economic and life-cycle assessments to support realistic implementation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis; methodology, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis.; software, Maria Lucia Cocato Eduardo Gianini Abimorad and Pietro Ragozzino-Paulino; validation, Eduardo Gianini Abimorad and Pietro Ragozzino-Paulino.; formal analysis, Maria Lucia Cocato; Eduardo Gianini Abimorad, Leandro Lopes Borges and Daniela Castellani; investigation, Maria Lucia Cocato, Eduardo Gianini Abimorad, Leandro Lopes Borges and Daniela Castellani; writing—original draft preparation, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis; writing—review and editing, Pietro Ragozzino-Paulino and Eduardo Gianini Abimorad; visualization, Pietro Ragozzino-Paulino; supervision, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis.; project administration, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis; funding acquisition, Maria Lucia Cocato, Eduardo Gianini Abimorad and Jorge Eduardo de Souza Sarkis. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the São Paulo Research Foundation (FAPESP) through the PIPE program (Research Project PIPE I—Grant No. 2018/01038-5; Research Project PIPE II—Grant No.).
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors gratefully acknowledge Metamorphosis Biotechnology Pesquisas Científicas Ltda. for kindly providing Tenebrio molitor larvae used in the experimental diets. The authors also acknowledge the São Paulo Research Foundation (FAPESP) for financial support through the PIPE program (Research Project PIPE I—Grant No. 2018/01038-5; Research Project PIPE II—Grant No. 2019/16802-5). During the preparation of this manuscript, the authors used ChatGPT (OpenAI) for the purpose of assisting in the drafting and refinement of some figure layouts and graphical elements. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest..
Abbreviations
The following abbreviations are used in this manuscript:
| ADC | Apparent Digestibility Coefficient |
| ADC-CP | Apparent Digestibility Coefficient of Crude Protein |
| ADC-EE | Apparent Digestibility Coefficient of Ether Extract |
| AOAC | Association of Official Analytical Chemists |
| BHI | Brain Heart Infusion |
| CFU | Colony-Forming Units |
| CON | Control Diet |
| Cr₂O₃ | Chromium(III) Oxide |
| EDTA | Ethylenediaminetetraacetic Acid |
| FAME | Fatty Acid Methyl Esters |
| FCR | Feed Conversion Ratio |
| HI | Hermetia illucens Larvae Meal Diet |
| HSI | Hepatosomatic Index |
| LD₅₀ | Median Lethal Dose |
| LSI | Liposomatic Index |
| PER | Protein Efficiency Ratio |
| SGR | Specific Growth Rate |
| TM | Tenebrio molitor Larvae Meal Diet |
| TMHI | Diet Containing a 1:1 Mixture of Tenebrio molitor and Hermetia illucens Larvae Meals |
| VSI | Viscerosomatic Index |
References
- Food and Agriculture Organization of the United Nations (FAO). The State of World Fisheries and Aquaculture 2024: Blue Transformation in Action; FAO: Rome, 2024; Available online: https://www.fao.org/publications/sofia-2024.
- Paredes-Trujillo, A.I.; Mendoza-Carranza, M. Streptococcosis in Nile tilapia farming: a severe yet overlooked health challenge in Mexico, a study case in Campeche, Mexico. Bull. Eur. Assoc. Fish. Pathol. Available from. 2024, 45, 1–17. [Google Scholar] [CrossRef]
- Hua, K. A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture Available from. 2021, 530, 735732. [Google Scholar] [CrossRef]
- Maulu, S.; Langi, S.; et al. Recent advances in the utilization of insects as an ingredient in aquafeeds: a review. 2022. Available online: https://pubmed.ncbi.nlm.nih.gov/36329686/.
- Islam, S.M.M.; Amrin, A.A.H.; Yousef, A.; Abol-Munafi, K.; Dayal, A.; Abdul-Rahim, A.M.K. Insect-based aquafeeds and fish mucosal immune responses: a systematic review. Rev. Aquac. 2024, 16, 1359–1390. [Google Scholar] [CrossRef]
- Hasan, M.M.; Alam, M.S.; Rayhan, N.; Rahman, M.M. Insects as alternative protein sources in fish nutrition: a review. Fishes 2023, 8, 291. [Google Scholar] [CrossRef]
- Ragozzino-Paulino, P.; Cocato, M.L.; de Souza Sarkis, J.E. Nutritional Potential of Edible Insects as Alternative Ingredients in Fish Feed: A Path to Modern Aquaculture. Aquac. Nutr. 2025, 7009004, 30. [Google Scholar] [CrossRef]
- Ihuț, A.; Răducu, C.; Uiuiu, P.; Munteanu, C. From gut to fillet: comprehensive effects of Tenebrio molitor in fish nutrition. Fishes Available from. 2025, 10, 468. [Google Scholar] [CrossRef]
- Liu, Q.; Li, J.; Liu, J.; Cai, Y.; Luo, Y.; Sangzhu, T.; et al. Enhanced Nile tilapia meat quality by the metabolomic effects of Tenebrio molitor larval meal dietary supplement. Appl. Food Res. 2025, 5, 101117. [Google Scholar] [CrossRef]
- Tippayadara, N.; Dawood, M.A.O.; Krutmuang, P.; Hoseinifar, S.H.; Doan, H.V.; Paolucci, M. Replacement of fish meal by black soldier fly (Hermetia illucens) larvae meal: effects on growth, haematology, and skin mucus immunity of Nile tilapia, Oreochromis niloticus. Animals 2021, 11, 193. [Google Scholar] [CrossRef]
- Shati, S.; Öz, M.; Erteken, A. Hermetia illucens in the feeding of Nile tilapia larvae. Aquat. Res. 2022, 5, 207–218. Available online: https://aquatres.scientificwebjournals.com/en/pub/issue/69220/1108366.
- Oliveira, C.G.; Araújo, R.D.; Prado, V.G.L.; Guilherme, H.O.; Paulino, R.R.; Costa, L.S. Impact of replacing fish meal with black soldier fly (Hermetia illucens) meal on diet acceptability in juvenile Nile tilapia: palatability and nutritional and health considerations for dietary preference. Aquac. Res. Available from. 2024, 2024, 3409955. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Y.Z.; Bao, M.Y.; Qin, X.P.; Guan, G.X. Defatted Hermetia illucens meal improves gut health and disease resistance in Nile tilapia under bacterial challenge. Fish Shellfish. Immunol. 2025, 110242. [Google Scholar] [CrossRef]
- Abd El-Gawad, E.A.; Zahran, E.; Youssuf, H.; Shehab, A.; Matter, A.F. Defatted black soldier fly (Hermetia illucens) diets improved hemato-immunological responses, biochemical parameters, and antioxidant activities in Streptococcus iniae-infected Nile tilapia (Oreochromis niloticus). BMC Vet. Res. 2025, 21, 104. [Google Scholar] [CrossRef]
- Haenen, O.L.M.; Dalsgaard, I.; Vendrell, D.; Peters, R.; et al. Bacterial diseases of tilapia, their zoonotic potential and risk management. Rev. Aquac. 2023, 15, e12743. Available online: https://onlinelibrary.wiley.com/doi/10.1111/raq.12743. [CrossRef]
- Abdallah, A.; Elbestawy, A.; Radwan, I.; Eltahan, A.; Soliman, W.S.; Nasr-Elsayed, S.; et al. Update on Streptococcus agalactiae infection in tilapia: epidemiology, pathogenesis, diagnosis and control. Biology. 2024. Available online: https://pubmed.ncbi.nlm.nih.gov/39472379/.
- Owatari, M.S.; Mouriño, J.L.P.; Martins, M.L. Antimicrobial resistance: a concern related to streptococcosis in tilapia farming. Mar Fish Sci (MAFIS) 2024, 37, 337–352. [Google Scholar] [CrossRef]
- Alves, A.P.C.; Paulino, R.R.; Pereira, R.T.; Costa, D.V.; Rosa, P.V. Nile tilapia fed insect meal: growth and innate immune response in different times under lipopolysaccharide challenge. Aquac. Res. Available from. 2021, 52, 529–540. [Google Scholar] [CrossRef]
- ABNT. NBR ISO 9001:2015. Sistemas de gestão da qualidade – requisitos. ABNT: Rio de Janeiro, 2015.
- AOAC. Official Methods of Analysis of AOAC International. In Method 2001.11, 17th edn; AOAC International: Gaithersburg, MD, 2002. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International. In Method 920.39, 22nd edn; AOAC International: Rockville, MD, 2023. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International. In Method 969.33, 22nd edn; AOAC International: Rockville, MD, 2023. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, Method 982.30, 22nd edn; AOAC International: Rockville, MD, 2023. [Google Scholar]
- Rockville. AOAC; 2023 Official Methods of Analysis of AOACInternational 22nd edn Method 99412 AOACInternational.
- AOAC. Official Methods of Analysis of AOAC International, Method 988.15, 22nd edn; AOAC International: Rockville, MD, 2023. [Google Scholar]
- National Research Council (NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; p. 376 pp. [Google Scholar]
- Bureau, D.P.; Harris, A.M.; Cho, C.Y. Apparent digestibility of rendered animal protein ingredients for rainbow trout (Oncorhynchus mykiss). Aquaculture 1999, 180, 345–358. [Google Scholar] [CrossRef]
- Abimorad, E.G.; Carneiro, D.J. Fecal collection methods and determination of crude protein and gross energy digestibility coefficients of feedstuffs for pacu, Piaractus mesopotamicus (Holmberg, 1887). Braz. J. Anim. Sci. 2004, 33, 1101–1109. [Google Scholar] [CrossRef]
- Nose, T. On the digestion of food protein by goldfish and rainbow trout. Bull. Freshw. Fish. Res. Lab. 1960, 10, 11–22. [Google Scholar]
- Goldenfarb, P. B.; Bowyer, F. P.; Hall, E.; Brosious, E. Reproducibility in the hematology laboratory: the microhematocrit determination. Am. J. Clin. Pathol. 1971, 56, 35–39. [Google Scholar] [CrossRef]
- Tran, H.Q.; Van Doan, H.; Stejskal, V. Environmental consequences of using insect meal as an ingredient in aquafeeds: A systematic view. Rev. Aquac. 2021, 14, 237–251. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: a review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Devic, E.; Leschen, W.; Murray, F.; Little, D.C. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing Black Soldier Fly (Hermetia illucens) larvae meal. Aquacult Nutr. 2018, 24, 416–423. [Google Scholar] [CrossRef]
- Dietz, C.; Liebert, F. Does graded substitution of soy protein concentrate by an insect meal respond on growth and N-utilization in Nile tilapia (Oreochromis niloticus)? Aquac. Rep. 2018, 12, 43–48. [Google Scholar] [CrossRef]
- Goldenfarb, P.B.; Bowyer, F.P.; Hall, E.; Brosious, E. Reproducibility in the hematology laboratory: the microhematocrit determination. Am. J. Clin. Pathol. 1971, 56, 35–39. [Google Scholar] [CrossRef]
- Tubin, J.S.B.; Paiano, D.; Hashimoto, G.S.O.; Furtado, W.E.; Martins, M.L.; Durigon, E.; et al. Tenebrio molitor meal in diets for Nile tilapia juveniles reared in biofloc system. Aquaculture 2020, 519, 734763. [Google Scholar] [CrossRef]
- Fontes, T.V.; de Oliveira, K.R.B.; Gomes Almeida, I.L.; Maria Orlando, T.; Rodrigues, P.B.; Costa, D.V.d.; Rosa, P.V.e. Digestibility of insect meals for Nile tilapia fingerlings. Animals 2019, 9, 181. [Google Scholar] [CrossRef]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a feed ingredient for fish culture: Status and trends. Aquac. Fish. 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Glencross, B.D. A feed is still only as good as its ingredients: An update on the nutritional research strategies for the optimal evaluation of ingredients for aquaculture feeds. Aquacult Nutr. 2020, 26, 1871–1883. [Google Scholar] [CrossRef]
- Santinha, P.J.M.; Gomes, E.F.S.; Coimbra, J.O. Effects of protein level of the diet on digestibility and growth of gilthead sea bream, Sparus auratus L. Aquacult Nutr. 1996, 2, 81–87. [Google Scholar] [CrossRef]
- Schrama, J.W.; Saravanan, S.; Geurden, I.; Heinsbroek, L.T.N.; Kaushik, S.J.; Verreth, J.A.J. Dietary nutrient composition affects digestible energy utilisation for growth: A study on Nile tilapia (Oreochromis niloticus) and a literature comparison across fish species. Br. J. Nutr. 2012, 108, 277–289. [Google Scholar] [CrossRef]
- Muin, H.; Taufek, N.M. Evaluation of growth performance, feed efficiency and nutrient digestibility of red hybrid tilapia fed dietary inclusion of black soldier fly larvae (Hermetia illucens). Aquac. Fish. 2024, 9, 46–51. [Google Scholar] [CrossRef]
- Hasan, I.; Gai, F.; Cirrincione, S.; Rimoldi, S.; Saroglia, G.; Terova, G. Chitinase and insect meal in aquaculture nutrition: A comprehensive overview of the latest achievements. Fishes 2023, 8, 607. [Google Scholar] [CrossRef]
- Eggink, K.M.; Pedersen, P.B.; Lund, I.; Dalsgaard, J. Chitin digestibility and intestinal exochitinase activity in Nile tilapia and rainbow trout fed different black soldier fly larvae meal size fractions. Aquac. Res. 2022, 53, 5536–5546. [Google Scholar] [CrossRef]
- Ebling, M.L.P.; Taschetto, E.A.; Pelizari, A.; Pietro, B.C.; Loureiro, B.B.; Lazzari, R.; et al. Tenebrio molitor larvae meal and evaluation of nutritional composition and apparent digestibility coefficient in silver catfish (Rhamdia quelen) feed. Cienc. Rural. 2025, 55, e20240408. [Google Scholar] [CrossRef]
- Quang Tran, H.; Woynarovich, A.; Thongvam, T.; Phuong, N.T. Environmental consequences of using insect meal as an alternative protein source in aquafeed: a review. Rev. Aquac. 2022, 14, 1474–1494. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquac. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Hameed, A.; Majeed, A.; Maqsood, S.; Hamed, M.; Mahmood, A.; et al. Success of aquaculture industry with new insights of using insects as fish feed: a review. Fishes 2022, 7, 395. Available online: https://www.mdpi.com/2410-3888/7/6/395. [CrossRef]
- Patyra, E.; Kwiatek, K. Insect meals and insect antimicrobial peptides as an alternative for antibiotics and growth promoters in livestock production. Pathogens 2023, 12, 854. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, E.S.H.; Metwally, W.G.M.; Abdel-Rahman, M.A.M.; Albano, M.; Mahmoud, M.M. Streptococcus agalactiae infection in Nile tilapia (Oreochromis niloticus): A review. Biology 2024, 13, 914. [Google Scholar] [CrossRef] [PubMed]
- Delphino, M.K.V.C.; Leal, C.A.G.; Gardner, I.A.; Assis, G.C.; Jr, W.S.; Inoue, L.A.K.A.; et al. Economic appraisal of vaccination against Streptococcus agalactiae in Nile tilapia farms in Brazil. Aquaculture 2019, 518, 734881. Available online: https://pubmed.ncbi.nlm.nih.gov/30621892/.
- Fyrand, K.; Xu, C.; Evensen, Ø. Characterization of Streptococcus agalactiae 1a isolated from farmed Nile tilapia (Oreochromis niloticus) in North America, Central America, and Southeast Asia. Fish. Shellfish. Immunol. 2024, 154, 109919. [Google Scholar] [CrossRef]
- Owatari, M.S.; Mouriño, J.L.P.; Martins, M.L. Antimicrobial resistance: a concern related to streptococcosis in tilapia farming. Mar Fish Sci (MAFIS) 2024, 37, 337–352. [Google Scholar] [CrossRef]
- Delphino, M.; Joshi, R.; Alvarez, A.T. Economic appraisal of using genetics to control Streptococcus agalactiae in Nile tilapia under cage and pond farming system in Malaysia. Sci. Rep. 2022, 12, 8754. Available online: https://pmc.ncbi.nlm.nih.gov/articles/PMC9130118/. [CrossRef]
- Pau, E.J.J.N.; Yong, C.C. Input–output analysis of Streptococcus disease impact on Malaysian tilapia production and exports. Aquacult Int. 2025, 33, 494. [Google Scholar] [CrossRef]
- Henry, M.A.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish. Shellfish. Immunol. 2018, 83, 308–313. [Google Scholar] [CrossRef]
- Su, J.; Gong, Y.; Cao, S.; Lu, F.; Han, D.; Liu, H.; et al. Effects of dietary Tenebrio molitor meal on the growth performance, immune response and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish. Shellfish. Immunol. 2017, 69, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sankian, Z.; Khosravi, S.; Kim, Y.O.; Lee, S.M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, and immune responses of mandarin fish (Siniperca scherzeri). Aquaculture 2018, 496, 79–87. [Google Scholar] [CrossRef]
- Libanori, M.C.M.; Santos, G.G.; Pereira, S.A.; Lopes, G.R.; Owatari, M.S.; Soligo, T.A.; Yamashita, E.; Pereira, U.P.; Martins, M.L.; Mouriño, J.L.P. Dietary supplementation with benzoic organic acid improves the growth performance and survival of Nile tilapia (Oreochromis niloticus) after challenge with Streptococcus agalactiae (Group B). Aquaculture 2021, 545, 737204. [Google Scholar] [CrossRef]
- Rimoldi, S.; Di Rosa, A.R.; Oteri, M.; Chiofalo, B.; Hasan, I.; Saroglia, M.; Terova, G. The impact of diets containing Hermetia illucens meal on the growth, intestinal health, and microbiota of gilthead seabream (Sparus aurata). Fish. Physiol. Biochem. 2024, 50, 1003–1024. [Google Scholar] [CrossRef]
- Franco, A.; Scieuzo, C.; Salvia, R.; Pucciarelli, V.; Borrelli, L.; Addeo, N.F.; Bovera, F.; Laginestra, A.; Schmitt, E.; Falabella, P. Antimicrobial activity of lipids extracted from Hermetia illucens reared on different substrates. Appl. Microbiol. Biotechnol. 2024, 108, 167. [Google Scholar] [CrossRef]
- Costa, R.S.; Basto, A.; Monteiro, M.; Pinho, B.; Sá, T.; Santos, M.V.; Murta, D.; Schrama, J.W.; Valente, L.M.P. Combining Hermetia illucens and Tenebrio molitor meals in diets for European seabass: Effects on growth, nutrient utilisation, intestinal morphology and muscle quality. Aquaculture 2026, 610, 742899. [Google Scholar] [CrossRef]
Figure 1.
Water temperature and dissolved oxygen variation during the tilapia growth trial.

Figure 2.
Experimental design and methodological workflow.

Figure 3.
Cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled Infection 1 and Infection 2). Cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled data from two independent challenge blocks: Infection 1, n = 24 fish per diet; Infection 2, n = 22 fish per diet; total n = 46 fish per diet). Fish were previously fed the experimental diets for 82 days. Cumulative mortality was recorded daily for 13 days post-challenge and calculated as the proportion of dead fish per treatment. Diets: CON, control diet without insect ingredients; TM, diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI, diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI, diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Survival curves did not differ among diets (log-rank test, P = 0.592).
Figure 3.
Cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled Infection 1 and Infection 2). Cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled data from two independent challenge blocks: Infection 1, n = 24 fish per diet; Infection 2, n = 22 fish per diet; total n = 46 fish per diet). Fish were previously fed the experimental diets for 82 days. Cumulative mortality was recorded daily for 13 days post-challenge and calculated as the proportion of dead fish per treatment. Diets: CON, control diet without insect ingredients; TM, diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI, diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI, diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Survival curves did not differ among diets (log-rank test, P = 0.592).

Table 4.
Ingredient composition and calculated nutritional composition of the experimental diets for Nile tilapia juveniles.
Table 4.
Ingredient composition and calculated nutritional composition of the experimental diets for Nile tilapia juveniles.
| Diet | CON | TM | HI | TMHI |
| Ingredient composition (g kg⁻¹) | ||||
| Insect meal¹ | 0 | 100 | 100 | 100 |
| Poultry by-product meal | 109 | 38.6 | 40 | 42.5 |
| Meat and bone meal | 70 | 70 | 70 | 70 |
| Blood meal | 50 | 33.5 | 32.8 | 30 |
| Soybean meal | 310 | 320 | 303.7 | 314.1 |
| Wheat bran | 100 | 100 | 100 | 100 |
| Broken rice | 80 | 80 | 80 | 80 |
| Corn | 200 | 200 | 190 | 191 |
| Corn starch | 40 | 40 | 40 | 40 |
| Soybean oil | 26.5 | 0 | 28.6 | 17.3 |
| Dicalcium phosphate | 8.4 | 10 | 9.2 | 9.3 |
| DL-methionine | 1 | 2 | 1 | 0.7 |
| Vitamin–mineral premix² | 5 | 5 | 5 | 5 |
| Calculated composition | ||||
| Crude protein (%) | 31.5 | 31.5 | 31.5 | 31.5 |
| Digestible energy (kcal kg⁻¹) | 3176 | 3234 | 3244 | 3243 |
| Ether extract (%) | 6.7 | 6.7 | 6.7 | 6.7 |
| Ash (%) | 7.7 | 7.1 | 7.7 | 7.3 |
| Crude fibre (%) | 3.4 | 3.4 | 3.2 | 3.3 |
| Nitrogen-free extract (%) | 40.6 | 41 | 40.8 | 41.3 |
| Calcium (%) | 1.4 | 1.33 | 1.37 | 1.32 |
| Available phosphorus (%) | 0.81 | 0.83 | 0.84 | 0.82 |
| Lysine (%) | 1.72 | 1.76 | 1.79 | 1.79 |
| Methionine (%) | 0.54 | 0.65 | 0.54 | 0.56 |
¹Insect meal was included at 100 g kg⁻¹ diet on a dry matter basis. CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. ²Vitamin–mineral premix composition (per kg of premix): choline 100 g; vitamin A 1,750,000 IU; vitamin D₃ 375,000 IU; vitamin E 20,000 IU; vitamin K₃ 500 mg; vitamin B₁ 2,000 mg; vitamin B₂ 2,500 mg; vitamin B₆ 2,500 mg; vitamin B₁₂ 5,000 µg; niacin 8,750 mg; pantothenic acid 7,500 mg; folic acid 625 mg; biotin 50 mg; vitamin C 37.5 g; inositol 12.5 g; iron 15 g; copper 1,250 mg; manganese 3,750 mg; zinc 17.5 g; cobalt 50 mg; iodine 100 mg; selenium 75 mg.
Table 5.
The Ca and P content median ± U (expanded uncertainty with K=2 and 95% confidence interval) (n = 3) and recovery (%) of Ca and P in NIST SRM 1548a (Total Diet). Growth performance of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
Table 5.
The Ca and P content median ± U (expanded uncertainty with K=2 and 95% confidence interval) (n = 3) and recovery (%) of Ca and P in NIST SRM 1548a (Total Diet). Growth performance of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
| Diet | Initial body weight (g) | Final body weight (g) |
Weight gain (g) |
SGR (% day⁻¹) |
Feed intake (g) |
FCR |
| CON | 28.83 ± 0.64 | 248.31 ± 13.31 | 219.48 ± 13.23 | 2.76 ± 0.07 | 262.34 ± 15.74 | 1.20 ± 0.03 |
| TM | 28.28 ± 0.28 | 254.99 ± 10.77 | 226.71 ± 10.50 | 2.82 ± 0.04 | 260.58 ± 3.94 | 1.15 ± 0.05 |
| HI | 28.75 ± 0.75 | 252.97 ± 19.14 | 224.21 ± 19.05 | 2.79 ± 0.10 | 276.24 ± 13.38 | 1.25 ± 0.04 |
| TMHI | 29.22 ± 0.32 | 260.36 ± 6.35 | 231.14 ± 6.05 | 2.80 ± 0.02 | 270.63 ± 2.71 | 1.17 ± 0.02 |
| P-value | 0.2802 | 0.7388 | 0.7464 | 0.7110 | 0.3043 | 0.0525 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. SGR: specific growth rate; FCR: feed conversion ratio.
Table 6.
Survival and somatic indices of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
Table 6.
Survival and somatic indices of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
| Diet | Survival (%) | Condition factor | VSI (%) | HSI (%) | LSI (%) |
| CON | 96.7 ± 5.8 | 2.05 ± 0.12 | 10.36 ± 0.24 | 2.38 ± 0.17 | 1.95 ± 0.34 |
| TM | 100.0 ± 0.0 | 2.19 ± 0.05 | 10.88 ± 0.51 | 1.99 ± 0.43 | 2.23 ± 0.32 |
| HI | 93.3 ± 5.8 | 2.09 ± 0.11 | 10.34 ± 2.01 | 2.14 ± 0.83 | 1.72 ± 0.49 |
| TMHI | 100.0 ± 0.0 | 2.10 ± 0.06 | 10.96 ± 0.74 | 1.81 ± 0.14 | 2.40 ± 0.06 |
| P-value | 0.7797 | 0.3832 | 0.8487 | 0.5478 | 0.0544 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. VSI: viscerosomatic index; HSI: hepatosomatic index; LSI: liposomatic index. Values are means (n = 3 tanks per treatment) ± standard deviation.
Table 7.
Apparent digestibility coefficients (ADC, %) of crude protein and ether extract of the experimental diets and test ingredients (insect meals).
Table 7.
Apparent digestibility coefficients (ADC, %) of crude protein and ether extract of the experimental diets and test ingredients (insect meals).
| Diet | Diets – test diets | Ingredients – insect meals | ||
| ADC-CP (%) | ADC-EE (%) | ADC-CP (%) | ADC-EE (%) | |
| CON | 86.95 ± 0.71ᵃ | 87.22 ± 1.57 | — | — |
| TM | 83.60 ± 1.87ᵇ | 87.66 ± 1.42 | 75.79 ± 6.22ᵇ | 88.70 ± 4.73 |
| HI | 87.31 ± 0.87ᵃ | 86.76 ± 1.66 | 88.16 ± 2.90ᵃ | 85.69 ± 5.52 |
| TMHI | 85.71 ± 0.42ᵃ | 88.95 ± 0.91 | 82.83 ± 1.41ᵃᵇ | 93.48 ± 2.93 |
| P-value | 0.0134** | 0.9132 | 0.0265* | 0.8343 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. ADC-CP: apparent digestibility coefficient of crude protein; ADC-EE: apparent digestibility coefficient of ether extract. Values are means (n = 3 tanks per treatment) ± standard deviation. Different superscript letters within the same column indicate significant differences (P < 0.05, Tukey’s test).
Table 8.
Red blood cell (erythrocytes) and total leukocyte counts of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
Table 8.
Red blood cell (erythrocytes) and total leukocyte counts of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
| Diet | Erythrocytes (×10³ µL⁻¹) | Leukocytes (×10³ µL⁻¹) |
| CON | 2461.6 ± 216.5 | 26.58 ± 3.28 |
| TM | 2536.7 ± 270.3 | 25.71 ± 3.70 |
| HI | 2307.5 ± 284.8 | 26.37 ± 4.73 |
| TMHI | 2612.5 ± 369.3 | 31.18 ± 9.31 |
| P-value | 0.3332 | 0.3616 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Values are means ± SD of six fish per diet (two fish per tank; three tanks). Statistical comparisons accounted for tank as the experimental unit (tank nested within diet).
Table 9.
Differential leukocyte counts (%) of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
Table 9.
Differential leukocyte counts (%) of Nile tilapia juveniles fed diets containing Tenebrio molitor and Hermetia illucens larvae meals.
| Diet | Neutrophils (%) | Lymphocytes (%) | Monocytes (%) | Eosinophils (%) | Basophils (%) |
| CON | 32.33 ± 5.75ᵃ | 55.33 ± 3.62ᵇ | 12.00 ± 4.20 | 0.17 ± 0.41 | 0.17 ± 0.41 |
| TM | 26.33 ± 3.88ᵃᵇ | 61.83 ± 4.49ᵃᵇ | 11.67 ± 2.34 | 0.17 ± 0.41 | 0.00 ± 0.00 |
| HI | 22.50 ± 5.57ᵇ | 66.33 ± 6.41ᵃ | 10.33 ± 3.01 | 0.33 ± 0.52 | 0.50 ± 0.55 |
| TMHI | 21.83 ± 8.11ᵇ | 65.00 ± 7.77ᵃ | 13.00 ± 2.10 | 0.16 ± 0.41 | 0.00 ± 0.00 |
| P-value | 0.0247* | 0.0172* | 0.5101 | 0.8832 | 0.0628 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Values are means ± SD of six fish per diet (two fish per tank; three tanks). Statistical comparisons accounted for tank as the experimental unit (tank nested within diet). Different superscript letters within the same column indicate significant differences (P < 0.05, Tukey’s test).
Table 10.
Cumulative mortality and survival of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled data from Infection 1 and Infection 2).
Table 10.
Cumulative mortality and survival of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled data from Infection 1 and Infection 2).
| Diet | n (total) | Dead (n) | Survivors (n) | Cumulative mortality (%) [95% CI] | Survival (%) | Observation period (days) |
| CON | 46 | 30 | 16 | 65.2 [50.8–77.3] | 34.8 | 13 |
| TM | 46 | 24 | 22 | 52.2 [38.1–65.9] | 47.8 | 13 |
| HI | 46 | 23 | 23 | 50.0 [36.1–63.9] | 50.0 | 13 |
| TMHI | 46 | 28 | 18 | 60.9 [46.5–73.6] | 39.1 | 13 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Values are pooled outcomes from two independent challenge blocks (Infection 1: n = 24 fish per diet; Infection 2: n = 22 fish per diet; total n = 46 per diet). Percentages are based on counts (Dead/n). 95% Confidence Interval (CI) for cumulative mortality was calculated using the Wilson method. Kaplan–Meier curves were derived from daily deaths (Table 11); log-rank test among diets: χ²(3)=1.907, P=0.592.
Table 11.
Daily deaths (n) and cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled Infection 1 + Infection 2; n = 46 per diet).
Table 11.
Daily deaths (n) and cumulative mortality (%) of Nile tilapia juveniles after intraperitoneal challenge with Streptococcus agalactiae (pooled Infection 1 + Infection 2; n = 46 per diet).
| Day post-challenge |
CON (dead n) |
CON (cum. mort. %) |
TM (dead n) |
TM (cum. mort. %) |
HI (dead n) |
HI (cum. mort. %) |
TMHI (dead n) |
TMHI (cum. mort. %) |
| 0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 2 | 0 | 0.0 | 1 | 2.2 | 2 | 4.3 | 1 | 2.2 |
| 3 | 10 | 21.7 | 8 | 19.6 | 9 | 23.9 | 8 | 19.6 |
| 4 | 9 | 41.3 | 7 | 34.8 | 6 | 37.0 | 8 | 37.0 |
| 5 | 3 | 47.8 | 3 | 41.3 | 2 | 41.3 | 7 | 52.2 |
| 6 | 1 | 50.0 | 2 | 45.7 | 1 | 43.5 | 1 | 54.3 |
| 7 | 1 | 52.2 | 1 | 47.8 | 1 | 45.7 | 1 | 56.5 |
| 8 | 2 | 56.5 | 0 | 47.8 | 0 | 45.7 | 0 | 56.5 |
| 9 | 0 | 56.5 | 1 | 50.0 | 0 | 45.7 | 0 | 56.5 |
| 10 | 1 | 58.7 | 0 | 50.0 | 1 | 47.8 | 1 | 58.7 |
| 11 | 2 | 63.0 | 0 | 50.0 | 1 | 50.0 | 1 | 60.9 |
| 12 | 1 | 65.2 | 1 | 52.2 | 0 | 50.0 | 0 | 60.9 |
| 13 | 0 | 65.2 | 0 | 52.2 | 0 | 50.0 | 0 | 60.9 |
CON: diet without insect ingredients; TM: diet containing 100 g kg⁻¹ Tenebrio molitor larvae meal; HI: diet containing 100 g kg⁻¹ Hermetia illucens larvae meal; TMHI: diet containing 50 g kg⁻¹ T. molitor + 50 g kg⁻¹ H. illucens larvae meals. Daily values are based on pooled deaths from two independent challenge blocks (Infection 1 + Infection 2), total n = 46 fish per diet. “cum. mort. (%)” was calculated as (cumulative deaths / 46) × 100.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



