Submitted:
07 April 2026
Posted:
09 April 2026
You are already at the latest version
Abstract
Endophytic bacteria are beneficial microbes that live within plant tissues and promote growth through nitrogen fixation, phosphate solubilization, and phytohormone production. Two endophytic isolates from bell pepper (Capsicum annuum L.) root were identified based on their morphology, and biochemistry properties using 16S rRNA gene sequencing. Winter barley (Hordeum vulgare L.) seeds were inoculated with two PGP (plant growth-promoting) bacterial strains (C-14 and C-27), previously characterized for IAA production and nitrogen fixation, and evaluated in a pot experiment with four treatments: A1(C-14), A2 (C-27), A3 (their consortium), and A4 (non-inoculated control). The results indicated that root and stem lengths increased in plants inoculated with bacteria compared to the uninoculated control. Among treatments, A2 produced the greatest root and shoot lengths (17.23 and 26.2 cm), while A3 showed the lowest (15.8 and 21.5 cm). SPAD values also increased by 6%, 10%, and 3.2% in A1, A2, and A3, respectively. This study clearly demonstrated that the endophytic isolates (C-14 and C-27) obtained from bell pepper roots significantly enhanced the growth and development of barley due to their abilities of IAA production and nitrogen fixation, thereby offering a promising alternate to chemical fertilizers.
Keywords:
barley
; endophytic bacteria
; Bacillus mycoides
; Bacillus tropicus
1. Introduction
Barley (Hordeum vulgare L.) is a fundamental cereal crop widely cultivated across the globe for its dual use as human food, beer production and its use as animal feed. Its high dietary fiber content, especially its abundance of β-glucan, has been associated with numerous health-promoting effects, including the prevention of degenerative diseases such as diabetes, obesity, hypertension, and colitis [1]. In addition to health benefits, barley is valued for its rich nutritional profile, serving as a cheapsource of high quality starch, vitamins, minerals, and proteins, that enhances its potential as a dietary supplement. Despite its agronomic and nutritional significance, barley production like many other crops is increasingly affected by various environmental and anthropogenic stressors [2]. The reduction in cultivable land, frequent crop losses, global warming, prolonged drought, and natural disasters have all contributed to substantial yield declines. In this context, achieving agricultural sustainability necessitates not only improvements in the physical and chemical characteristics of the soil but also the conservation and enhancement of its biological properties. Promoting soil health through environment friendly practices. The widespread use of synthetic fertilizers and pesticides poses significant risks to soil ecosystems Thus, the implementation of green production methods that prevent soil and water pollution has become a strategic priority everywhere [3].
Endophytes constitute a diverse group of microorganisms including primarily bacteria and fungi, along with archaea, algae, protozoa, and viruses that live on or close to various plant organs such as roots, stems, leaves, seeds, and fruits. The microorganisms must initially colonize the rhizosphere, followed by successful invasion of the plant endosphere to establish themselves as endophytes. This process relies on specific microbial traits, including motility, surface attachment, degradation of plant polymers, and the ability to evade plant defense mechanisms [4]. Endophytic bacteria colonise internal plant tissues and support plant growth by numerous methods such as nitrogen fixation, phosphate solubilisation, and the synthesis of siderophores and phytohormones. Additionally, these bacteria produce enzymes like ACC (1-aminocyclopropane-1-carboxylate) deaminase, which reduces ethylene levels in plants; thereby enhancing plant growth [5]. Although research on the plant growth-promoting effects of endophytic bacteria has increased in recent years, the studies on their impact on barley plants remain limited. Numerous studies involving endophytic bacteria suggest that the plant growth-promoting potential of local isolates should be investigated in different ecosystems and plant species [6,7].
This study aimed to assess the plant growth-promoting traits of endophytic bacteria isolated from bell pepper and to examine the effects of two bacterial strains (Bacillus mycoides C-14 and Bacillus tropicus C-27) on the growth and development of barley plants under controlled pot conditions.
2. Materials and Methods
2.1. Sample Collection
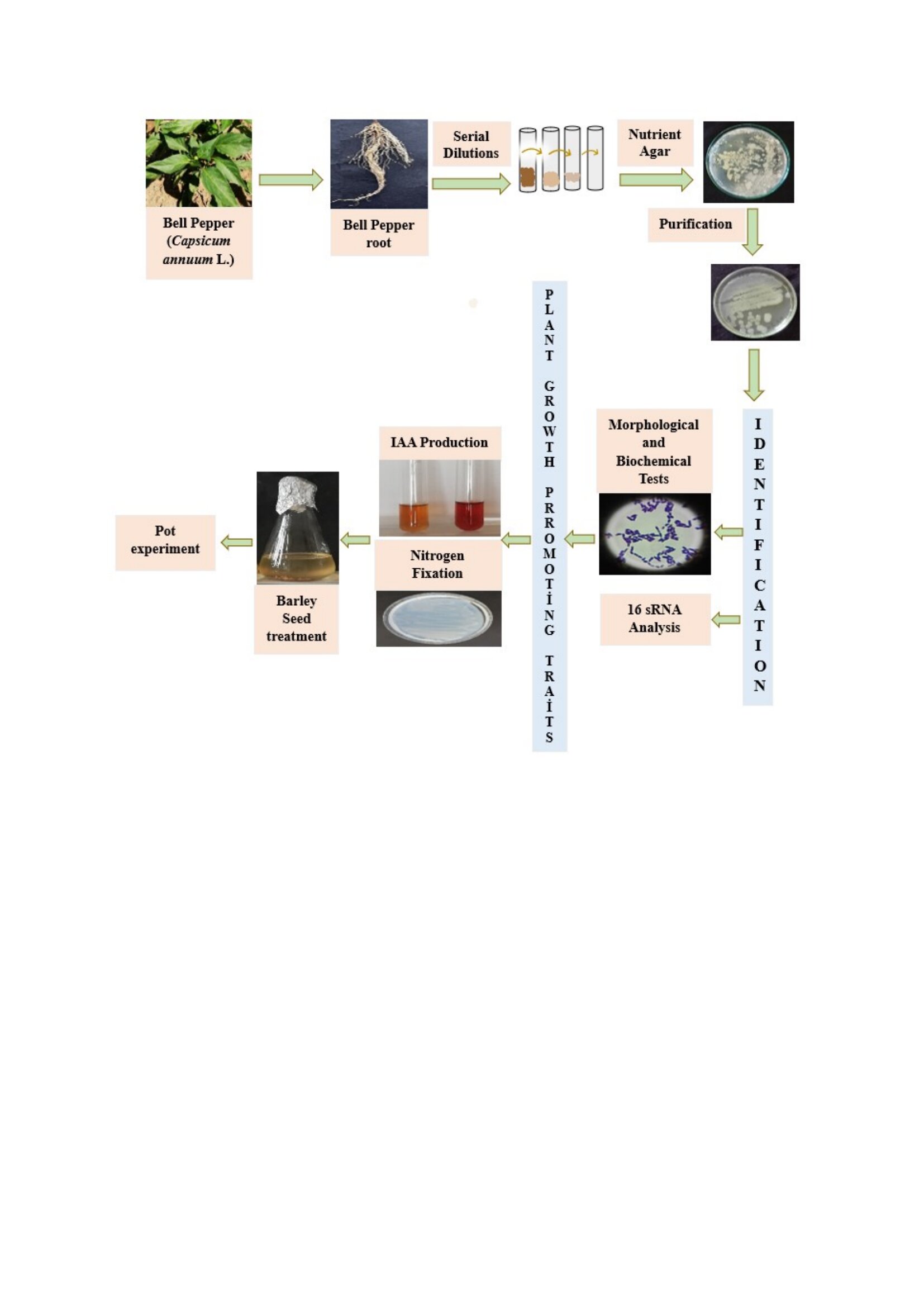
Bell pepper (Capsicum annuum L. var. grossum) were obtained from a bell pepper field in Çayağzı (Cemele) village (39°16'56.0‘’ N, 34°04'44.3‘’ E), Kırsehir, in May 2023. The pepper plants were properly labeled, sealed in sterile plastic bags, and transported to the bacteriology laboratory of the Department of Field Crops, Faculty of Agriculture, Ankara University under cold-chain conditions for subsequent isolation procedures (Figure 1).
2.2. Isolation of Endophytic Bacterial Isolates
Potential endophytic bacterial strains were isolated from the roots of three bell peppers following the method described by Amaresan et al.[8]. Bell pepper roots were washed thoroughly in tap water for 30 minutes to remove soil and similar particles. The roots were cut into 2-3 cm lengths, placed in a beaker and washed 3-4 times in sterile distilled water. After sterilization with 0.1% HgCl2 for 25 seconds, it was washed with 70 % ethanol for 5 minutes. Finally, it was rinsed 5-6 times with sterile distilled water. Surface disinfected root tissues were crushed and ground aseptically with homogenizers. The homogenized plant tissues were further macerated in sterile phosphate-buffered saline (0.2 g KCl, 8 g NaCl, 1.44 g Na₂HPO₄, 0.24 g KH₂PO₄, in 1 L of distilled water, pH 7.4) and subsequently subjected to serial dilution up to 10-³. From each dilution, 100 µl was spread on to Nutrient Agar (NA) plates and incubated at 30 °C for 48 hours. After incubation, bacteria were examined and selected based on desired morphological characteristics and then they were purified by the streak plate method. Purified bacteria were stored in 50 % (v/v) glycerol solution at -80 °C for future research.
Figure 1 illustrates the flowchart of the process used to identify endophytic bacteria isolated from bell pepper roots to determine their growth and development development on the barley.
2.3. Phenotypic/Biochemical Characterization and Molecular Identification
2.3.1. Morphological and Biochemical Characteristics
Physiological and biochemical characteristics, as well as Gram staining and spore formation of the bacterial isolates, were evaluated following the procedures outlined by Palleroni et al. [9].
2.3.2. Molecular Identification of Bacterial Endophytes
DNA Extraction
Genomic DNA was extracted from isolates C-14 and C-27 using the GeneMATRIX Bacterial & Yeast DNA Isolation Kit (EurX®, Poland) and following the manufacturer's instructions (https://eurx.com.pl/docs/manuals/en/e3580.pdf). The DNA quantity and purity were evaluated following extraction using a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA).
PCR Amplification of 16S rRNA and Sequencing
Amplification of the 16S rRNA gene was performed using universal primers BAC 27F (5' AGAGTTTGATCMTGGCTCAG 3') and BAC 1492R (5' TACGGYTACCTTGTTACGACTT 3') along with the FIREPol® DNA Polymerase Kit (Solis Biodyne, Estonia) in a 25 μL reaction volume [10]. PCR was conducted in a Kyratec thermocycler with the following conditions: 95 °C for 5 minutes for initial denaturation, followed by 30 cycles of denaturation at 95 °C for 45 seconds, annealing at 57 °C for 45 seconds, and extension at 72 °C for 60 seconds, with a final extension at 72 °C for 5 minutes. The amplified product, approximately 1470 bp, was run on a 1.5% agarose gel prepared with 1x TAE buffer.
Electrophoresis was performed at 100 volts for 90 minutes, and the results were visualized under UV light using ethidium bromide staining. Single-band PCR products were purified using the HighPrep™ PCR Clean-up System (AC-60005, MAGBIO) according to the manufacturer’s protocol. Purified 16S rDNA amplicons were sequenced using the Sanger dideoxy sequencing method with an ABI 3730XL Sanger sequencing instrument (Applied Biosystems, Foster City, CA) and the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA)
Bioinformatics Analysis
The resulting sequences, including two chromatograms for each bacterium (forward and reverse), were visualized, checked for quality, and analyzed using the Finch TV program version 1.4.0. They were then transformed into consensus sequences using ClustalW (http://www.ebi.ac.uk/Tools/msa/clustalo/). Similarities and identities of the query sequences were determined using the nucleotide Basic Local Alignment Search Tool (BLASTn; https://blast.ncbi.nlm.nih.gov/) and the rRNA/ITS GenBank database of NCBI. The sequences were submitted to the GenBank nucleotide database. For constructing the phylogenetic tree, highly similar sequences were subjected to multiple sequence alignment (MSA) using ClustalW2 in BioEdit software. The Maximum Likelihood (ML) phylogenetic tree was constructed using Kimura 2-parameter substitution model. The bootstrap method was applied with 1000 replicates. Evolutionary analyses were performed using Molecular Evolutionary Genetics Analysis Version 7.0 (MEGA7) software.
2.4. Characterization of Endophytic Bacteria
2.4.1. IAA Production Assay
The IAA production of endophytic strains cultured for four days in a nutrient medium supplemented with 0.2% L-tryptophan was determined spectrophotometrically at 535 nm using a SHIMADZU UVmini-1240 spectrophotometer, based on a standard curve generated from known IAA concentrations and using uninoculated NB medium as a control, following the method described by Gang et al. [11].
2.4.2. Nitrogen Fixation Assay
The nitrogen-fixation potential of the endophytic bacterial isolates was evaluated following the methodology proposed by Park et al. [12], with minor adjustments. Initially, the isolates were cultured on Nutrient Agar (NA) and incubated at 28 °C for 24 hours. After incubation, fresh colonies were transferred onto Petri plates containing solid nitrogen-free Burk’s medium composed of (per liter): 0.41 g KH₂PO₄, 10 g glucose, 0.52 g K₂HPO₄, 0.2 g CaCl₂, 0.05 g Na₂SO₄, 0.1 g MgSO₄·7H₂O, 0.005 g FeSO₄·7H₂O, 0.0025 g Na₂MoO₄·2H₂O, and 1.8 g agar, dissolved in 1 L of distilled water. The plates were then incubated at 28 ± 2 °C for four days to assess nitrogen-fixing activity. Isolates capable of growing in nitrogen-free media were regarded as positive.
2.5. Seed Preparation and Bacterial Inoculation
The Aydan Hanim winter barley seeds (Hordeum vulgare L.) used in this study were obtained from the Department of field crops of the Faculty of Agriculture, Ankara University (39°57'44.2"N, 32°51'36.7"E). Two endophytic isolates with plant growth promoting properties were used as barley seeds treatments. Barley seeds were placed in 10 % sodium hypochlorite (NaOCl) solution, for 15 minutes. It was followed by five times rinsing with distilled tap water to ensure sterilization. Then air-dried in laminar air flow. The pure cultures were grown on Nutrient Broth (NB) medium containing 4 % sugar (sucrose). The concentration of the prepared bacterial solutions (including 4 % sugar) was adjusted to 108 cfu ml-1 with sterile distilled water. The seeds were then treated with bacterial suspensions for 45 min under sterile conditions. Following the protocol of Heinonsalo et al. [13], control seeds underwent the same sterilization process but were treated with sterile distilled water instead of bacterial inoculum (Figure 2).
2.6. Greenhouse Pot Trials
A pot experiment was conducted to study the effect of two endophytic isolates (C-14 and C-27) with plant growth promoting properties on the development of barley plants. The plants were grown for 30 days in climate cabinets under controlled conditions of 20°C temperature, 65% relative humidity, a 10-hour fixed light photoperiod with light intensity (850 PAR), followed by 14 hours of complete darkness [14]. The experiment was conducted in Complete Randomized Design (CRD) with three replicates.
Growth Medium and Soil Properties
Pots (4.5 cm × 4.5 cm × 10 cm) were sterilized with potassium permanganate using garden soil: vermiculite: perlite as a growth medium (1:1:1 ratio). The garden soil samples to be used in pots were sterilized by autoclaving at 121 °C and 1.1 atm pressure. (Table 1). Single colonies of each isolate were individually inoculated into NB medium and cultured under shaking conditions at 28 °C with horizontal shaking at 200 rpm for 24 hours. The bacterial suspensions were subsequently adjusted to a final concentration of 1 × 10⁸ cfu ml-1 and utilized in further experiments. All treatments are described as following:
A1: Seeds bacterized with C-14,
A2: Seeds bacterized with C-27,
A3: Seeds bacterized with Consortium (C-14+C-27),
A4: Seeds of barley plants coated with 1% CMC (carboxymethylcelluloses) slurry was served as control.
A total of 5 seeds were sown in each of the 12 pots, with 3 replicates per treatment. The inoculated treatments included 20 ml of the bacterial suspension at the concentration of 1 × 108 cfu ml-1 into each pot. The uninoculated control treatments (A4) contained 20 ml of the sterile Luria Bertani (LB) liquid medium. Sterile water was slowly added to each pot every three days to maintain soil moisture.
Data Collection
After 30 days, the plants were harvested to evaluate plant fresh weight, dry weight, root length, stem length, and relative chlorophyll content (SPAD units). Total chlorophyll content in barley leaves was determined using a SPAD chlorophyll meter (SPAD-502, Minolta, Osaka, Japan). Measurements were taken from different leaves of the seedlings, with five replicates per treatment. The physicochemical characteristics of the soil employed in the experiment are presented in Table 1.
2.7. Statistical Analysis
Statistical analyses of the data was performed using JMP Pro 17.0 statistical software, based on three replicates. Dependant variables with normal distribution were shown as mean ± standard deviation (SD). Statistical analysis of data was done using Duncan’s test (p ≤ 0.05).
3. Results
3.1. Morphological and Biochemical Characterization
In the current study, Two endophytic bacteria were identified based on morphological observations and biochemical characterization. Physiological and biochemical characteristics, along with Gram staining of the bacterial isolates, were assessed according to the protocols described by Palleroni et al. [9]. Based on the phenotypic characterization of the two isolates, both were determined to be Gram-positive with the ability to form spores, exhibited motility, and tested positive for the oxidase test; however, isolate C-27 was found to be catalase-negative. Details are presented in Table 2.
3.2. Molecular Identification of Strains C-14 and C-27
The 16S rRNA gene sequences of isolates C-14 and C-27 were analyzed using in silico tools and have been deposited in the NCBI GenBank under accession numbers PQ610418 and PQ610419, respectively (Table 3). Isolate C-14 exhibited 99.93% identity with reference strains of Bacillus mycoides isolated from various countries (NR_113996, NR_036880, NR_024697, NR_115993, and NR_113990), while isolate C-27 showed 99.72% identity with Bacillus tropicus strain MCCC 1A01406 (NR_157736). The phylogenetic tree shows that C-14 clusters with B. mycoides strains, whereas C-27 forms a separate clade (Figure 3).
3.3. Evaluation of Plant Growth-Promoting Traits
Both the C-14 and C-27 isolates were determined to have the ability to produce indole-3-acetic acid (IAA) and to fix atmospheric nitrogen. Table 4, Figure 4 and Figure 5 present the plant growth-promoting traits of the isolates.
In the present study, the tested strains produced different levels of IAA depending on their growth in NB medium supplemented with L-tryptophan. After four days, based on spectrophotometric analysis, the highest IAA production was observed in isolate C-27 (103.7 µg ml⁻¹). This was followed by isolate C-14 (93.4 µg ml-1). Moreover, It was determined that IAA production increased from day 1 to day 4 in both isolates. The lowest IAA production was recorded on day 1 by isolate C-14 (45.8 4 µg ml-1), whereas the highest production was observed on day 4 by isolate C-27 (103.7 4 µg ml-1) (Figure 4).
3.4. Evaluation of Pot Experiments
A general enhancement in all measured plant growth parameters was evident in the pot trials. It was determined that both single-strain and consortium bacterial treatments significantly increased root and stem length, fresh and dry weight, and SPAD values. Plants treated with the A1 (C-14), A2 (C-27), and A3 (consortium) isolates had 5.8%, 9.5%, and 3.2% higher SPAD values, respectively, compared to the control plants in present study. The maximum SPAD value (47.2) was measured in barley seedlings treated with A2 treatment, while the lowest SPAD value was measured in A4 (Control) treatment (43.1) in present study.
The PGPR Isolates significantly affected the root and stem length of barley seedlings. According to the research results, the A2 treatment increased root length by 11.6% compared to the control treatment. The longest root (17.23 cm) was recorded in the A2 treatment. Although there was an increase in plant growth parameters in seeds treated with the consortium compared to the control, the longest stem (26.2 cm) and maximum fresh weight (60.03 mg) values were observed in the A2 treatment. Moreover, a significant increase in dry matter of barley seedlings was observed in response to PGPR isolates compared to the control. The maximum dry matter content was recorded in the A2 treatment (16.23 mg), while the minimum dry matter was observed in the A4 treatment (14.55 mg). It was determined that the A2 treatment exerted a more pronounced positive effect on plant growth compared to the other treatments. Details are presented in Table 5.
The results of pot study showed that treatment of barley seeds with bacterial strains showed a positive effect on root and stem length (Figure 6).
4. Discussion
The C-14 and C-27 isolates exhibited plant growth-promoting (PGP) traits such as nitrogen fixation, and production of indole-3-acetic acid (IAA), which are key indicators commonly associated with a strong potential for enhancing plant growth. These results indicate that the recovery of barley seedlings through PGPR treatment was likely achieved through the induction of IAA production, and nitrogen fixation (Figure 6).
Bacteria have the ability to promote plant growth through the production and secretion of phytohormones, particularly indole-3-acetic acid (IAA), which promotes root development by stimulating primary root elongation and the proliferation of lateral and adventitious roots. These alterations in root architecture positively influence water absorption and nutrient uptake [15]. Many researchers have reported that endophyte bacteria such as Bacillus spp. found in the roots of different plants are good producers of IAA and that the concentration of synthesised bacterial indole-3-acetic acid (IAA) has a significant effect on the development of the root system [16,17]. For instance, Solano-Alvarez et al. [18] reported that the application of phosphate-solubilizing and IAA-producing B. cereus isolated from the bean rhizosphere to tomato seeds increased root length and weight by 62% and 58%, respectively. Widowati et al. [19] observed that the inoculation of B. subtilis B11, an IAA-producing strain isolated from the roots, stems, and leaves of pepper, enhanced the stem length (19.75 cm) and root length (15.47 cm) of pepper seedlings. Similarly, there are also research studies focusing on the effects of inoculation with IAA-producing Bacillus strains on barley plants. For instance Baris et al. [20] reported that inoculation of barley seeds with Bacillus subtilis and B. megaterium, which have the potential to produce IAA, increased grain yield and total yield by 15.1-27.8% and 14.5-18.5%, respectively. These results were in agreement with those reported by Hafez et al. [21] in barley plants. The current study confirms a significant positive relationship between root-stem length and IAA production. In the current greenhouse experiments, root lengths increased by 10, 18, and 8%, while shoot lengths increased by 60, 7%, and 43% after A1, A2, and A3 treatments, respectively (Table 5). Additionally, The treatment of the consortium may have had a lesser effect on root development compared to single-strain treatments, possibly due to negative interactions between the isolates that did not contribute positively to barley root elongation. Ma et al. [22] reported that single inoculation of barley seeds with Bacillus mycoides PM35, a strain possessing plant growth-promoting traits, resulted in significant increases in shoot length, root length, number of leaves, leaf area, shoot fresh weight, root fresh weight, and shoot dry weight compared to consortium-based treatments. A similar finding was reported by Anušauskas et al. [23], who stated that inoculation of barley with Bacillus mycoides enhanced plant growth parameters and, consequently, improved crop yield. To the best of our knowledge, although B. tropicus has been reported to enhance yield in crops such as wheat [24], rice [25], mung bean and mustard [26], and pineapple [27]; this is the first study conducted on barley.
Nitrogen, an essential element for all living organisms, is a fundamental component of various biomolecules, including amino acids, vitamins, and nucleotides [28]. Numerous studies have shown that inoculation with plant growth-promoting rhizobacteria (PGPR) capable of nitrogen fixation leads to significant improvements in several growth parameters, including plant height as well as root and stem dry weights [29,30]. Additionaly, It has been reported that strains belonging to the Bacillus genus, which are abundant in the plant root, increase root development by producing nitrogen fixation and IAA, and thus play a role in plant development [31]. Negi et al. [32] reported that Bacillus tropicus EU-ARP-44, isolated from the rhizospheric soil of Aegilops kotschyi and possessing nitrogen-fixing potential, increased the fresh and dry weight of Aegilops kotschyi by 2.6-fold, stem length by 1.2-fold, and root length by 1.6-fold under in vitro conditions. Similarly, Shahzad et al. [33] reported that the nitrogen fixing endophytic bacterium B. mycoides isolated from Viburnum grandiflorum increased stem length, leaf width, protein content and sugar content by 57 %, 13 %. Similar findings were also reported by Nauanova et al. [34], who demonstrated that inoculation of barley seeds with Bacillus spp. possessing nitrogen-fixing and phosphate-solubilizing abilities significantly enhanced root and stem biomass, thereby improving overall crop yield, 20 %, and 34 %, respectively, compared to the control when inoculated onto wheat seeds. Our results indicated that the A2 treatment exhibited higher efficacy in promoting biomass accumulation and dry matter production compared to the A1 and A3 consortium treatments.
SPAD is an indirect measure of the chlorophyll content in plant leaves. The treatment of PGPR in barley plants can enhance nitrogen uptake, resulting in increased leaf chlorophyll content. Purwanto and Suharti, [35] reported a strong correlation between leaf chlorophyll content and nitrogen levels in leaves. According to Bagues et al. [36], bacterial inoculation significantly increases the growth of barley plants and SPAD values. For instance, Emami et al. [37] reported a significant increase in the SPAD value of barley plants inoculated with a salt-tolerant isolate possessing plant growth-promoting traits. Jamily et al. [38] stated that the inoculation of barley with Bacillus subtilis, which possesses the ability to produce indole-3-acetic acid (IAA), siderophores, and solubilize phosphate, increased the plant dry weight and SPAD value by approximately 10% to 20%, respectively. Similarly, Abideen et al. [39] reported that barley plants inoculated with endophytic Pseudomonas or Pantoea exhibited an approximately 18% increase in total chlorophyll content compared to the control plants.
In present study, both individual and consortium treatments led to an increase in total chlorophyll content in barley compared to the uninoculated control group (Table 5). Moreover, The results showed that both single-strain and consortium PGPR treatments increased the SPAD value by 19% compared to the control group. The observed increase in chlorophyll content may be associated with the nitrogen-fixing capacity of Bacillus mycoides C-14 and Bacillus tropicus C-27. Additionally, Consistent with our findings, Gul et al. [40] also reported enhanced chlorophyll content in barley plants inoculated with plant growth-promoting rhizobacteria (PGPR). In a recent study, Ferioun et al. [41] reported that inoculation of barley seeds with Pseudomonas amygdali, which possesses nitrogen fixation and phosphate solubilisation capabilities, increased the SPAD value by 12.9%. Consistent with these results, this study observed increases in root and stem length, root and stem dry weight, and chlorophyll content in plants treated with PGPR.
5. Conclusions
This study highlights the functional importance of endophytic Bacillus strains isolated from bell peppers (Capsicum annuum L.) in promoting barley plant growth. Molecular screening and phylogenetic analysis successfully identified the isolates as Bacillus mycoides C-14 and Bacillus tropicus C-27, each clustering within distinct, well-established taxonomic groups.
Comprehensive in vitro characterization revealed that these isolates exhibit a diverse suite of plant growth-promoting (PGP) traits, including nitrogen fixation, and the synthesis of indole-3-acetic acid (IAA). The transition from laboratory assays to greenhouse trials successfully validated the real-world efficacy of these strains. While individual applications notably enhanced seedling vigor, it was the C-27 treatment that drove the most substantial improvements in root and shoot architecture, biomass accumulation, and chlorophyll content. These findings underscore the fact that synergistic interactions within a microbial community can surpass the limitations of single-strain inoculants, offering a more robust and multifaceted approach to plant growth promotion. These two strains are considered promising candidates for use as microbial fertilizers in agricultural applications due to their ability to produce indole-3-acetic acid (IAA) and fix atmospheric nitrogen. However, further studies are needed to confirm the effectiveness of these endophytic bacteria in enhancing plant growth under actual field conditions.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Lahouar, L.; Ghrairi, F.; El Arem, A.; Medimagh, S.; El Felah, M.; Ben Salem, H.; Achour, L. BIOCHEMICAL COMPOSITION AND NUTRITIONAL EVALUATION OF BARLEY RIHANE (HORDEUM VULGARE L). Afr. J. Tradit. Complement. Altern. Med. 2016, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ullah, Z.; Ullah, R.; Kazi, M. RETRACTED: Barley a nutritional powerhouse for gut health and chronic disease defense. Heliyon 2024, 10, e38669. [Google Scholar] [CrossRef] [PubMed]
- Fasusi, O.A.; Cruz, C.; Babalola, O.O. Agricultural Sustainability: Microbial Biofertilizers in Rhizosphere Management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Kiselev, K.V. Interaction of Plants and Endophytic Microorganisms: Molecular Aspects, Biological Functions, Community Composition, and Practical Applications. Plants 2023, 12, 714. [Google Scholar] [CrossRef]
- da Silveira, A.P.D.; Iório, R.d.P.F.; Marcos, F.C.C.; Fernandes, A.O.; de Souza, S.A.C.D.; Kuramae, E.E.; Cipriano, M.A.P. Exploitation of new endophytic bacteria and their ability to promote sugarcane growth and nitrogen nutrition. Antonie van Leeuwenhoek 2018, 112, 283–295. [Google Scholar] [CrossRef]
- Walitang, D.I.; Kim, K.; Madhaiyan, M.; Kim, Y.K.; Kang, Y.; Sa, T. Characterizing endophytic competence and plant growth promotion of bacterial endophytes inhabiting the seed endosphere of Rice. BMC Microbiol. 2017, 17, 1–13. [Google Scholar] [CrossRef]
- Uçar, C.P.; Selem, E.; Tunçtürk, R.; Tunçtürk, M.; Akköprü, A. The Effect of Some Endophytic Bacteria on Seedling Growth and Physiological Properties of Salvia officinalis L. 2023, 33, 130–139. [Google Scholar] [CrossRef]
- Amaresan, N.; Jayakumar, V.; Thajuddin, N. Isolation and characterization of endophytic bacteria associated with chilli (Capsicum annuum) grown in coastal agricultural ecosystem. Indian Journal of Biotechnology 2014, 13(2), 247–255. [Google Scholar]
- Palleroni, N.J.; Krieg, N.R.; Holt, J.G. Bergey's manual of systematic bacteriology; The Willian and Wilkins. Co: Baltimore, 1984; pp. 141–219. [Google Scholar]
- Rahmoune, B.; Morsli, A.; Khelifi-Slaoui, M.; Khelifi, L.; Strueh, A.; Erban, A.; Kopka, J.; Prell, J.; van Dongen, J.T. Isolation and characterization of three new PGPR and their effects on the growth of Arabidopsis and Datura plants. J. Plant Interactions 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of indole-3-acetic acid (IAA) production in Klebsiella by LC-MS/MS and the Salkowski method. Bio-Protocol 2019, 9. [Google Scholar] [CrossRef]
- Park, M.; Kim, C.; Yang, J.; Lee, H.; Shin, W.; Kim, S.; Sa, T. Isolation and characterization of diazotrophic growth promoting bacteria from rhizosphere of agricultural crops of Korea. Microbiol. Res. 2005, 160, 127–133. [Google Scholar] [CrossRef]
- Heinonsalo, J.; Frey-Klett, P.; Pierrat, J.-C.; Churin, J.-L.; Vairelles, D.; Garbaye, J. Fate, tree growth effect and potential impact on soil microbial communities of mycorrhizal and bacterial inoculation in a forest plantation. Soil Biol. Biochem. 2004, 36, 211–216. [Google Scholar] [CrossRef]
- Zytynska, S.E.; Eicher, M.; Rothballer, M.; Weisser, W.W. Microbial-Mediated Plant Growth Promotion and Pest Suppression Varies Under Climate Change. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Slimani, A.; Raklami, A.; Oufdou, K.; Meddich, A. Isolation and Characterization of PGPR and Their Potenzial for Drought Alleviation in Barley Plants. Gesunde Pflanz. 2022, 75, 377–391. [Google Scholar] [CrossRef]
- Bhutani, N.; Maheshwari, R.; Negi, M.; Suneja, P. Optimization of IAA production by endophytic Bacillus spp. from Vigna radiata for their potential use as plant growth promoters. Isr. J. Plant Sci. 2018, 65, 83–96. [Google Scholar] [CrossRef]
- Khianngam, S.; Meetum, P.; Na Chiangmai, P.; Tanasupawat, S. Identification and Optimization of Indole-3-Acetic Acid Production of Endophytic Bacteria and Their Effects on Plant Growth. Trop. Life Sci. Res. 2023, 34, 219–239–219–239. [Google Scholar] [CrossRef] [PubMed]
- Solano-Alvarez, N.; Valencia-Hernández, J.A.; Rico-García, E.; Torres-Pacheco, I.; Ocampo-Velázquez, R.V.; Escamilla-Silva, E.M.; Romero-García, A.L.; Alpuche-Solís, Á.G.; Guevara-González, R.G. A Novel Isolate of Bacillus cereus Promotes Growth in Tomato and Inhibits Clavibacter michiganensis Infection under Greenhouse Conditions. Plants 2021, 10, 506. [Google Scholar] [CrossRef]
- Widowati, T.; Nuriyanah; Nurjanah, L.; Lekatompessy, S.J.R.; Simarmata, R. Bioproduction of indole acetic acid by endophytic bacteria of Bacillus strains isolated from chili (Capsicum annuum L.) and its potential for supporting the chili seedlings. In THE FIRST INTERNATIONAL CONFERENCE ON NEUROSCIENCE AND LEARNING TECHNOLOGY (ICONSATIN; LOCATION OF CONFERENCE, IndonesiaDATE OF CONFERENCE, 2021; p. 020018. [Google Scholar]
- Baris, O.; Sahin, F.; Turan, M.; Orhan, F.; Gulluce, M. Use of Plant-Growth-Promoting Rhizobacteria (PGPR) Seed Inoculation as Alternative Fertilizer Inputs in Wheat and Barley Production. Commun. Soil Sci. Plant Anal. 2014, 45, 2457–2467. [Google Scholar] [CrossRef]
- Hafez, Y.; Attia, K.; Alamery, S.; Ghazy, A.; Al-Doss, A.; Ibrahim, E.; Rashwan, E.; El-Maghraby, L.; Awad, A.; Abdelaal, K. Beneficial Effects of Biochar and Chitosan on Antioxidative Capacity, Osmolytes Accumulation, and Anatomical Characters of Water-Stressed Barley Plants. Agronomy 2020, 10, 630. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Chen, F.; Sun, Y.; Zhu, Y.; Wang, L. Bacillus mycoides PM35 in combination with titanium dioxide (TiO2)⎯nanoparticles enhanced morpho-physio-biochemical attributes in Barley (Hordeum vulgare L.) under cadmium stress. Chemosphere 2023, 323, 138224. [Google Scholar] [CrossRef]
- Anušauskas, J.; Steponavičius, D.; Romaneckas, K.; Lekavičienė, K.; Zaleckas, E.; Sendžikienė, E. The Influence of Bacteria-Inoculated Mineral Fertilizer on the Productivity and Profitability of Spring Barley Cultivation. Plants 2023, 12, 1227. [Google Scholar] [CrossRef]
- Malik, L.; Sanaullah, M.; Mahmood, F.; Hussain, S.; Shahzad, T. Co-application of biochar and salt tolerant PGPR to improve soil quality and wheat production in a naturally saline soil. Rhizosphere 2024, 29. [Google Scholar] [CrossRef]
- Gulzar, A.B.M.; Mazumder, P.B. Seed priming and subsequent inoculation of drought-tolerant plant growth-promoting rhizobacteria Bacillus tropicus KH90 improves rice (Oryza sativa L.) plant health under drought and non-drought conditions. Cereal Res. Commun. 2024, 53, 1051–1071. [Google Scholar] [CrossRef]
- Nandni; Rani, S.; Dhiman, I.; Wati, L. Biopriming with multifarious sulphur-oxidizing bacteria improve in vitro Vigna radiata L. (mung bean) and Brassica juncea L. (mustard) seed germination. Folia Microbiol. 2024, 70, 427–439. [Google Scholar] [CrossRef]
- Jaya, D.K.; Giyanto, G.; Nurhidayat, N.; Antonius, S. Isolation, identification, and detection of ACC deaminase gene-encoding rhizobacteria from rhizosphere of stressed pineapple. Indones. J. Biotechnol. 2019, 24, 17–25. [Google Scholar] [CrossRef]
- Güler, M. Identification of beneficial bacteria in Rosemary rhizospheres and determination of plant growth promoting (PGP) potential. J. Agric. Fac. Gaziosmanpasa Univ. 2024. [Google Scholar] [CrossRef]
- Ejaz, S.; Batool, S.; Anjum, M.A.; Naz, S.; Qayyum, M.F.; Naqqash, T.; Shah, K.H.; Ali, S. Effects of inoculation of root-associative Azospirillum and Agrobacterium strains on growth, yield and quality of pea (Pisum sativum L.) grown under different nitrogen and phosphorus regimes. Sci. Hortic. 2020, 270. [Google Scholar] [CrossRef]
- Raza, A.; Ejaz, S.; Saleem, M.S.; Hejnak, V.; Ahmad, F.; Ahmed, M.A.A.; Alotaibi, S.S.; El-Shehawi, A.M.; Alsubeie, M.S.; Zuan, A.T.K. Plant growth promoting rhizobacteria improve growth and yield related attributes of chili under low nitrogen availability. PLOS ONE 2021, 16, e0261468. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Vryzas, Z.; Papadopoulou-Mourkidou, E. Enhanced root uptake of acibenzolar-S-methyl (ASM) by tomato plants inoculated with selected Bacillus plant growth-promoting rhizobacteria (PGPR). Appl. Soil Ecol. 2014, 77, 26–33. [Google Scholar] [CrossRef]
- Negi, R.; Kaur, T.; Devi, R.; Kour, D.; Yadav, A.N. RETRACTED: Assessment of nitrogen-fixing endophytic and mineral solubilizing rhizospheric bacteria as multifunctional microbial consortium for growth promotion of wheat and wild wheat relative Aegilops kotschyi. Heliyon 2022, 8, e12579–e12579. [Google Scholar] [CrossRef]
- Shahzad, A.; Aslam, U.; Ferdous, S.; Qin, M.; Siddique, A.; Billah, M.; Naeem, M.; Mahmood, Z.; Kayani, S. Combined effect of endophytic Bacillus mycoides and rock phosphate on the amelioration of heavy metal stress in wheat plants. BMC Plant Biol. 2024, 24, 1–16. [Google Scholar] [CrossRef]
- Nauanova, A.; Shaikhin, S.; Ospanova, S.; Makenova, M.; Shumenova, N.; Bostubayeva, M. Enhancing Spring Barley Grain Yield with Local Biofertilizers in the Semi-Arid Steppe Zone of Northern Kazakhstan. Int. J. Des. Nat. Ecodynamics 2024, 19, 371–378. [Google Scholar] [CrossRef]
- Purwanto, P.; Suharti, W.S. Nutrient Uptake, Chlorophyll Content, and Yield of Rice (Oryza sativa L.) Under the Application of PGPR Consortium. Biosaintifika: J. Biol. Biol. Educ. 2021, 13, 336–344. [Google Scholar] [CrossRef]
- Bagues, M.; Neji, M.; Karbout, N.; Boussora, F.; Triki, T.; Guasmi, F.; Nagaz, K. Mitigating Salinity Stress in Barley (Hordeum vulgare L.) through Biochar and NPK Fertilizers: Impacts on Physio-Biochemical Behavior and Grain Yield. Agronomy 2024, 14, 317. [Google Scholar] [CrossRef]
- Emami, T.; Mirzaeiheydari, M.; Maleki, A.; Bazgir, M. Effect of native growth promoting bacteria and commercial biofertilizers on growth and yield of wheat (Triticum aestivum) and barley (Hordeum vulgare) under salinity stress conditions. Cell. Mol. Biol. 2019, 65, 22–27. [Google Scholar] [CrossRef]
- Jamily, A.S.; Koyama, Y.; Win, T.A.; Toyota, K.; Chikamatsu, S.; Shirai, T.; Uesugi, T.; Murakami, H.; Ishida, T.; Yasuhara, T. Effects of inoculation with a commercial microbial inoculant Bacillus subtilis C-3102 mixture on rice and barley growth and its possible mechanism in the plant growth stimulatory effect. J. Plant Prot. Res. 2023, 193-205–193-205. [Google Scholar] [CrossRef]
- Abideen, Z.; Cardinale, M.; Zulfiqar, F.; Koyro, H.-W.; Rasool, S.G.; Hessini, K.; Darbali, W.; Zhao, F.; Siddique, K.H. Seed Endophyte bacteria enhance drought stress tolerance in Hordeum vulgare by regulating, physiological characteristics, antioxidants and minerals uptake. Front. Plant Sci. 2022, 13, 980046. [Google Scholar] [CrossRef]
- Gul, F.; Khan, I.U.; Rutherford, S.; Dai, Z.-C.; Li, G.; Du, D.-L. Plant growth promoting rhizobacteria and biochar production from Parthenium hysterophorus enhance seed germination and productivity in barley under drought stress. Front. Plant Sci. 2023, 14, 1175097. [Google Scholar] [CrossRef]
- Ferioun, M.; Zouitane, I.; Bouhraoua, S.; Belahcen, D.; Srhiouar, N.; Louahlia, S.; El Ghachtouli, N. PGPR consortia promote soil quality and functioning in barley rhizosphere under different levels of drought stress. Ecol. Front. 2024, 45, 444–454. [Google Scholar] [CrossRef]
Figure 1.
Graphical abstract illustrating the effects of endophytic bacteria (C-14 and C-27), isolated from bell pepper, on the growth and development of barley plants.
Figure 1.
Graphical abstract illustrating the effects of endophytic bacteria (C-14 and C-27), isolated from bell pepper, on the growth and development of barley plants.

Figure 2.
Preparation of bacterial culture (C-14, C-27 and Consortium) in NB medium.

Figure 3.
Maximum likelihood phylogenetic tree of B. mycoides strain C-14 and B. tropicus strain C-27 based on the 16S rRNA gene.
Figure 3.
Maximum likelihood phylogenetic tree of B. mycoides strain C-14 and B. tropicus strain C-27 based on the 16S rRNA gene.

Figure 4.
IAA production by endophytic strains cultured in nutrient broth supplemented with 0.2% L-tryptophan over four days. All values are expressed as the mean of three replicates. Bars with different letters indicate statistically significant differences (p > 0.05). Error bars represent the standard deviation (SD) of the mean.
Figure 4.
IAA production by endophytic strains cultured in nutrient broth supplemented with 0.2% L-tryptophan over four days. All values are expressed as the mean of three replicates. Bars with different letters indicate statistically significant differences (p > 0.05). Error bars represent the standard deviation (SD) of the mean.

Figure 5.
PGPR test results of endophytic isolates (A) IAA production (B) Nitrogen fixation)

Figure 6.
The development of the barley plant in the pot after 30 days.


Table 1.
Physiochemical characteristics of the garden soil used in the experiment
| Soil characteristics | Value | Soil characteristics | Value |
| ECe* (dS m-1) | 2.6 | Total K (µg kg-1) | 18.2 |
| Soil pH | 5.3 | Total Fe (µg kg-1) | 2.33 |
| Total N (µg kg-1) | 13.1 | Total Zn (µg kg-1) | 665 |
| Total P (µg kg-1) | 3.98 | Total Cu (µg kg-1) | 160 |
| Total Mn (µg kg-1) | 2.15 | Total OM (%) | 0.84 |
*ECe, electrical conductivity of soil extract; OM, organic matter; N, nitrogen; P, phosphorus; K, potassium; Fe, iron; Mn, manganese; Zn, zinc; Cu, copper; OM, organic matter.
Table 2.
Morphological and biochemical properties of the isolated endophytes
| Isolate Code | Isolates |
Gram Characteristic |
Biochemical Characteristics |
Endospor forming | |||
|
Gram reaction |
KOH 3% |
Catalase | Oxidase | Motility | |||
| C-14 | Bacillus mycoides | + | - | + | + | + | + |
| C-27 | Bacillus tropicus | + | - | - | + | + | + |
Note: +, positive; –, negative
Table 3.
Molecular identification of endophytic bacterial isolates with their closely related species
Table 3.
Molecular identification of endophytic bacterial isolates with their closely related species
| Isolate ID | NCBI Accession ID | Taxonomic Closest relative | Percent Identity |
|---|---|---|---|
| C-14 | PQ610418 | Bacillus mycoides | 99.93 % |
| C-27 | PQ610419 | Bacillus tropicus | 99.72% |
Table 4.
Plant growth promoting traits of the endophytic isolates
| Isolate Code | Endophytic isolates | NF |
IAAP |
| C-14 | Bacillus mycoides | + | + |
| C-27 | Bacillus tropicus | + | + |
Note: +, positive; –, negative. NF: Nitrogen fixation. IAAP: Indole-3-acetic acid production.
Table 5.
Height, weight, and chlorophyll content (SPAD units) (C) of barley plants (H. vulgare L.) in response to treatments with A1 (C-14), A2 (C-27) and A3 Consortium (C-14+C-27) after 30 days
Table 5.
Height, weight, and chlorophyll content (SPAD units) (C) of barley plants (H. vulgare L.) in response to treatments with A1 (C-14), A2 (C-27) and A3 Consortium (C-14+C-27) after 30 days
| Isolates |
Root length (cm) (Mean± SD) |
Stem length (cm) (Mean± SD) |
Fresh Weight (mg) (Mean ± SD) |
Dry Weight (mg) (Mean ± SD) |
SPAD units | ||
| Root | Stem | Root | Stem | ||||
| A1(C-14) | 16.03±0.73ab |
24.1±0.91ab | 51.9±2.02b | 93.03±0.75a | 4.65±0.46b | 15.11±0.29ab | 45.6± 0.75b |
| A2 (C-27) | 17.23±1.02a |
26.2±1.47a | 60.03±1.45a | 95.0±1.30a | 6.36±0.42a | 16.23±0.62a |
47.2± 0.47a |
| A3Consortium(A1+A2) | 15.8± 0.62ab | 21.5±0.88b | 47.2±0.45c | 89.1±0.43b | 4.22±0.37b | 15.02±0.9b | 44.5±0.42a |
|
A4 (Control) |
14.6±0.75b* | 15.03±0.83c | 39.5±1.30d | 86.7±1.45b | 3.14±0.29c | 14.55±0.53b | 43.1± 0.4b |
Note: A1(C-14): Bacillus mycoides, A2 (C-27): Bacillus tropicus. A3 Consortium(C-14+C-27). Each value represents the mean of three replicates. *Values sharing the same letter within a column are not statistically different at the 0.05 level of confidence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



