Submitted:
04 April 2026
Posted:
07 April 2026
You are already at the latest version
Abstract
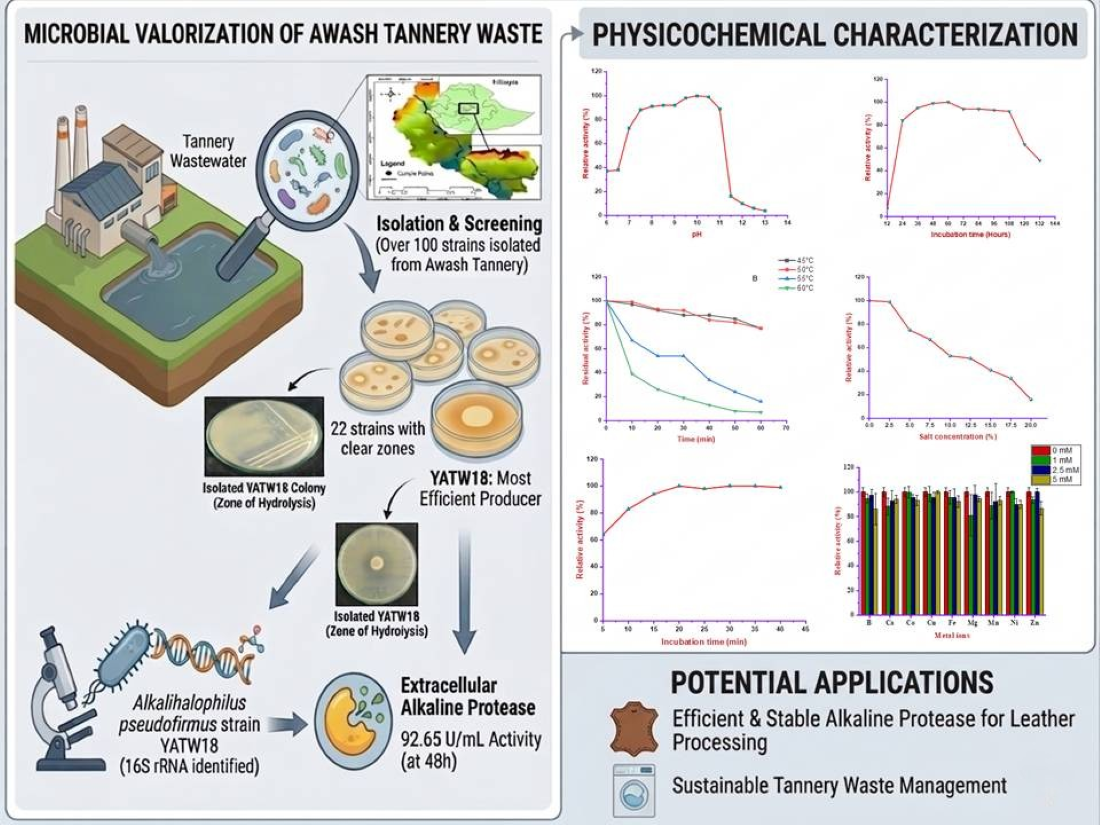
The global leather industry generates significant volumes of tannery wastes, creating conducive niche for the proliferation of proteolytic microbes. The study investigated microbial valorization of tannery waste through production and physicochemical characterization of a highly stable alkaline protease. Over 100 bacterial strains were isolated from the aqueous waste of Awash Tannery. Among these, 22 isolates were exhibited visible distinct clear zones around the colonies. Out of 22 bacterial strains, YATW18 was identified as the most efficient producer of extracellular alkaline protease; with 92.65 U/mL Activity at 48h. Molecular identification based on 16S rRNA gene sequence identified the strain YATW18 as Alkalihalophilus pseudofirmus. Physicochemical characterization of alkaline protease showed optimum activity at 55℃ and pH 10. The enzyme reached its saturation level within 20 min of incubation time and then linear activity was observed until 45 min. The enzyme activity of alkaline protease demonstrated a notable tolerance to increasing salt concentrations. Most metal ions had minimal influence on protease activity across concentrations, with the relative activity close to 100%. Overall, this study showed microbial valorization of tannery waste and its huge potential to be used as a key source of efficient and stable alkaline protease.
Keywords:
alkaline protease
; microbial valorization
; tannery waste
; strain YATW18
1. Introduction
Wastewater generated from the leather industry is typically alkaline, with a high pH and substantial organic load, making it a promising source for isolating alkaline enzyme producing microorganisms. In recent years, alkaline enzymes have gained considerable biotechnological importance due to their wide-ranging applications in industries such as leather processing, detergents, waste treatment, and food production. Tannery wastewater, in particular, offers significant potential for the isolation of alkaline protease–producing microbes, as large amounts of proteinaceous materials are released during the alkaline processing of hides and skins for leather production [1,2]. However, improper treatment of these proteins and associated chemicals can lead to serious environmental pollution and health hazards. Therefore, the development of eco-friendly and sustainable technologies is urgently required to mitigate the environmental and public health risks associated with the leather industry.
Recently, bacterial alkaline proteases have attracted considerable attention as environmentally friendly alternatives to inorganic catalysts. Alkaline protease producing bacteria are capable of growing under extreme pH conditions, such as those found in tannery liquid waste, which contains high levels of protein-based pollutants [3]. These bacteria degrade residual proteins through the secretion of extracellular alkaline proteases and subsequently assimilate the resulting hydrolysis products for growth and metabolism [4]. Therefore, the present study focuses on the microbial valorization of tannery waste through production and physicochemical characterization of an alkaline protease.
2. Materials and Methods
2.1. Sample Area Description
Tannery waste samples were collected from Awash leather industry. Awash Tannery is located in Nifas Silk Lafto Subcity, Addis Ababa, Ethiopia at 8°56′33′′ N and 38°45′41′′ E (Figure 1). The factory is located in an area of 123,000 m2. It is specialized in the manufacture of refined and quality leather products. The Awash tannery is capable of processing 3,600,000 sheep and goat skins as well as 396,000 hides per year. It generates 1000–5000 cubic meters of liquid waste daily [5].
2.2. Sample Collection
Approximately 3 L of tannery wastewater was collected from Awash Leather Industry using sterile bottles. Samples were randomly obtained in triplicate from the aerobic digestion unit, bulk wastewater, homogenization unit, and secondary treatment plant to ensure reliability, after which they were pooled into a composite sample. During sampling, temperature, pH, and salinity were measured on site. Total dissolved solids (TDS), total suspended solids (TSS), chemical oxygen demand (COD), and biochemical oxygen demand (BOD) were analyzed offsite. COD was determined using the dichromate method [6], while BOD was measured using the iodometric or Winkler method [7]. The samples were then transported to the Microbial Biotechnology Laboratory, Institute of Biotechnology, Addis Ababa University, and stored at 4 °C until further analysis.
2.3. Enrichment of the Samples
One milliliter of the wastewater sample was enriched in a selective medium containing 1% (w/v) skim milk powder (as the sole carbon and nitrogen source), 0.02% MgSO4·7H2O, 0.1% K2HPO4, 0.03% CaCl2·2H2O, and 1% NaCl [8]. The medium pH was adjusted to 10 using separately autoclaved 25% (w/v) Na2CO3 [9]. The inoculated broth was incubated in an orbital shaker incubator at 30 °C and 120 rpm for seven days.
2.4. Isolation and Screening of Alkaline Protease Producing Bacteria
Alkaline protease producing bacteria were isolated and screened following the method of Gessesse et al. [9] with minor modifications. Skim milk agar medium was used for screening and contained (w/v): skim milk (1%), NaCl (1%), yeast extract (0.05%), MgSO4·7H2O (0.02%), CaCl2·2H2O (0.03%), and K2HPO4 (0.1%). The medium components were mixed and heated until completely dissolved, after which the pH was adjusted to 7.5–8.0 using 1 M NaOH. Agar (2%, w/v) was added, and the medium was boiled and autoclaved at 121 °C for 15 min. A 25% (w/v) Na2CO3 solution was prepared and autoclaved separately, and 1% (v/v) was added aseptically to the medium to raise the final pH to 10.0.
Aliquots (0.1 mL) from appropriate dilutions of the enriched samples were spread onto skim milk agar plates using a sterile glass spreader and incubated at 30 °C for 48 h in an incubator (Electrical Heat Thermostatic Incubator, DH5000B). Colonies showing clear zones were selected, and more than 100 morphologically distinct isolates were purified by repeated streaking on skim milk agar and incubated at 30 °C for 48 h until uniform colonies were obtained. Extracellular protease activity was evaluated by calculating the ratio of the diameter of the clear zone to that of the bacterial colony. Isolates exhibiting larger clear zones were selected for further studies.
2.5. Production of Alkaline Protease
To select the strain exhibiting the highest extracellular alkaline protease activity under submerged fermentation, a loopful of freshly grown culture was inoculated into 100 mL of skim milk broth in a 500 mL Erlenmeyer flask containing the same medium composition used for isolation and screening, except for the absence of agar. This culture served as the starter. Subsequently, 3 mL of the 48-h-old starter culture was transferred into 100 mL of fresh medium, and the flasks were incubated in an orbital shaker incubator at 30 °C and 121 rpm for 12–120 h. At 12-h intervals, 1.5 mL of the fermented broth was aseptically withdrawn and centrifuged at 10,000 rpm for 5 min. The resulting cell-free supernatant was collected and used as the crude enzyme preparation.
2.6. Molecular Identification
2.6.1. Genomic DNA Extraction, PCR Amplification, Restriction Digestion and Sequencing
Genomic DNA from the selected alkaliphilic protease-producing bacterium was extracted using the cetyltrimethylammonium bromide (CTAB) method as described by Surzycki [10], with minor modifications. Briefly, 1.5 mL of an overnight bacterial culture was centrifuged at 10,000 rpm for 5 min, and the supernatant was discarded. The resulting cell pellet was resuspended in 567 µL of TE buffer. Cell lysis was achieved by adding 30 µL of 10% SDS and 3 µL of Proteinase K (20 mg/mL), followed by incubation at 37 °C for 1 h. Subsequently, 100 µL of 5 M NaCl was added and mixed thoroughly, after which 80 µL of CTAB/NaCl solution was added and the mixture incubated at 65 °C for 10 min.
The lysate was extracted with an equal volume of phenol:chloroform:isoamyl alcohol (25:24:1, v/v/v), vortexed for 30 s, and centrifuged at 10,000 rpm for 10 min. The upper aqueous phase was carefully transferred to a new tube and further purified using an equal volume of chloroform:isoamyl alcohol (24:1), followed by vortexing and centrifugation under the same conditions. DNA was precipitated by adding 360 µL of isopropanol, and the DNA pellet was collected by centrifugation at 12,000 rpm for 10 min. The pellet was washed with 1 mL of 70% ethanol, air-dried for 15 min, and resuspended in 50 µL of TE buffer. The extracted genomic DNA was stored at 4 °C for short-term use. Sterile techniques were employed throughout the procedure, and phenol was handled with appropriate safety precautions.
The yield and purity of genomic DNA were assessed both quantitatively and qualitatively using spectrophotometric analysis and agarose gel electrophoresis [11]. DNA concentration and purity were determined using a NanoDrop spectrophotometer by measuring absorbance at 260 and 280 nm. Agarose gel electrophoresis was performed using a 0.8% (w/v) agarose gel stained with ethidium bromide. Genomic DNA (2 µL) was mixed with loading dye, loaded onto the gel, and visualized under UV illumination following electrophoresis.
For molecular identification, the 16S rRNA gene was amplified by PCR using universal primers FD1 (5′-AGAGTTTGATCCTGGCTCAG-3′) and RD1 (5′-AAGGAGGTGATCCAGCC-3′), which amplify approximately 1,500 bp of the 16S rRNA gene [12]. The PCR reaction mixture (25 µL total volume) consisted of 19.5 µL nuclease-free water, 2.5 µL PCR buffer, 0.25 µL MgCl2, 0.25 µL dNTPs, 0.25 µL Taq DNA polymerase, 0.125 µL of each primer, and 2 µL of template DNA. PCR amplification was performed in a thermal cycler (Bio-Rad CFX96) under the following conditions: initial denaturation at 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 1 min, and extension at 72 °C for 30 s; followed by a final extension at 72 °C for 5 min [13].
The amplified PCR products were purified and sequenced bi-directionally using the same primer sets on an ABI 3730XL Genetic Analyzer. PCR purification and sequencing were outsourced to MRC Holland Company, Amsterdam, Netherlands.
2.6.2. Sequence Analysis
Sequences were edited manually using BioEdit software 7.4. Reference 16S rRNA sequences were retrieved from NCBI GenBank database available at http://www.ncbi.nlm.nih.gov using BLAST (Basic Local Alignment Search Tool) analysis to identify the sequence [14]. Phylogenetic relationship of sequences to closest matches in public database based on 16S rRNA gene sequences was constructed by using Neighbor-Joining method using distances calculated with Jukes-Cantor correction [15] in MEGA 11 software [16]. The stability and reliability of the relationships of lineages on the inferred trees was tested by bootstrap analysis for 1000 replicates [17]. The 16S rRNA gene sequences were submitted to NCBI public database with accession number PP942174.
2.7. Physicochemical Characterization of Alkaline Protease
2.7.1. Enzyme Activity
Protease activity was determined using casein substrate according to the method adopted by Gessesse et al., [9]. A 1% casein and 50 mM Glycine-NaOH buffer solution was prepared by mixing equal volume of solutions and adjusting at pH 10.0. Then, 450 µL of this substrate solution was transferred to four clean Eppendorf tubes, three for the enzyme and one for the enzyme blank. One reagent blank was also prepared by adding 50 µL of distilled water instead of crude enzyme. Then, 50 µL of crude enzyme was added to the Eppendorf tubes labeled as enzyme and incubated at 50 °C for 30 min in a water bath. After incubation, 500 µL of 10% TCA was added to stop the reaction. The solution was left at room temperature for 10-15 min to allow the undigested casein to precipitate. The reaction mixtures were centrifuged at 10,000 rpm for 5 min. Clear supernatant (500 µL) from each reaction mixture was transferred to the respective labeled clean test tubes, and 2.5 mL of 0.5 M sodium carbonate solution was added to each tubes. Folin-Ciocalteau’s phenol reagent (1:5; 500 µL) was added, thoroughly mixed by vortex, and then incubated in a dark room for 30 min. Finally, optical density (OD) was measured using a spectrophotometer (JENWAY 6300 UV/Vis) at 660 nm. Enzyme activity was measured against enzyme blank and reagent blank. One unit of alkaline protease activity was defined as the amount of enzyme that liberates one micromole of tyrosine per ml per min under standard experimental conditions [18].
Residual activity was calculated as the percentage enzyme activity in the sample compared to the activity of the control (untreated enzyme).
2.7.2. Tyrosine Standard Curve Preparation
First, a 200 μg/mL stock solution of tyrosine was made and diluted proportionately to obtain the required concentration of tyrosine (Figure S1). Then, 0.5M Na2CO3 was added in all the test tubes, including the blank. Next, 500 μL of the tyrosine solution was added to each test tube and the test tubes were vortexed and left to incubate at room temperature for 10 min. Finally, 500 μL of 2N Folin Ciocalteau’s phenol reagent was added to every test tube. The solution was vortexed immediately and left at room temperature for 30 min. Finally, optical density was measured at 660 nm using a spectrophotometer (JENWAY 6300 UV/Vis), and a standard curve was plotted using Origin Pro 2021 software (Figure S1).
2.7.3. pH Profile
The effect of pH on alkaline protease activity was evaluated by assaying the enzyme in various buffer systems. The buffers used were: KH2PO4–K2HPO4 (pH 6.0–7.5), Tris–HCl (pH 7.5–9.0), glycine–NaOH (pH 9.5–11.0), Na2PO4–NaOH (pH 11.0–12.0), and KCl (pH 12.5–13.0), all at a concentration of 50 mM and in 0.5 pH unit intervals. Assays were performed at 50 °C, and enzyme activity was measured according to the standard protease assay described in subsection 2.7.1, including the calculation of relative enzyme activity [9].
2.7.4. Temperature Profile and Stability
To determine the effect of temperature on the alkaliphilic protease activity the standard protease was performed at different temperature values ranging from 30 °C to 80 °C, in 5 °C intervals, in glycine-NaOH buffer, pH 10.0. To determine the thermal stability of the enzyme, the method described by Gessesse et al. [9] was followed. The enzyme was diluted in 0.1 M Glycine-NaOH buffer (pH 10.0) with and without 5 mM CaCl2 and incubated at temperatures ranging 45 °C to 60 °C for 1 h in a water bath. After incubation, the enzyme samples were withdrawn in every 10 min and stored at 4 °C. Residual activity was measured followed the standard protease assay protocol, with the untreated enzyme as 100%.
2.7.5. Salt Profile
The ionic strength tolerance of the alkaline protease was assessed by adding NaCl to the reaction mixture at concentrations ranging from 0% to 20% in 2.5% intervals (except for the initial 0% concentration). A standard protease assay was then conducted using 1% casein in 50 mM Glycine-NaOH buffer as a substrate at a pH of 10.0. Finally, the relative activity was calculated by setting the concentration with the highest activity as 100%.
2.7.6. Incubation Time
The effect of incubation time on enzyme activity was investigated using the reaction mixture prepared as described in Section 2.7.1. The mixture was incubated at 55 °C, and samples were withdrawn ranging 5 to 40 min, with 5 min intervals, followed by a standard protease assay. The incubation time profile was used to determine the optimal incubation time for maximum enzyme activity.
2.7.7. Metal Ions Profile
The effect of various metal ions (B3+, Zn2+, Mn2+, Ni2+, Cu2+, Fe2+, Co2+, Mg2+, and Ca2+) on enzyme activity was evaluated by pre-incubating the enzyme in 50 mM glycine–NaOH buffer (pH 10.0) at room temperature with each metal ion added to final concentrations of 1, 2.5, and 5 mM for 1 h. Following incubation, the residual protease activity was determined using the standard protease assay and compared with a control lacking added metal ions.
3. Results
3.1. Physicochemical Characteristics of the Tannery Waste Water
The temperature, pH, and salinity of the wastewater were 28 °C, 8.1, and 1%, respectively (Table 1). The total dissolved solids (TDS) concentration was 9.63 ppt. In contrast, the total suspended solids (TSS), chemical oxygen demand (COD), and biochemical oxygen demand (BOD) were 1044.7 mg/L, 1276 mg/L, and 375.4 mg/L, respectively (Table 1).
3.2. Isolation and Screening of Alkaline Protease Producing Bacteria
3.2.1. Isolation and Primary Screening
More than 100 bacterial strains were isolated from the characterized tannery wastewater. The presence of visible distinct clear zones around the bacterial colonies on skim milk agar plates indicated skim milk hydrolysis and extracellular protease production (Figure 2). Accordingly 22 isolates exhibit visible distinct clear zones around their colonies (Table 2). The proteolytic index measured ranged 2.5 mm (YATW5) to 7.9 mm (YATW9). Isolates with large proteolytic index, greater than 2.9 mm, were selected and subjected for secondary screening (Table 2).
3.2.2. Secondary Screening
Among the 20 isolates with a proteolytic index of greater than 2.9 mm that subjected for secondary screening, isolate YATW18 was identified as the most efficient producer of extracellular alkaline protease (Figure 3). In the submerged fermentation, isolate YATW18 produced a fairly high enzyme yield within 36-60 h of incubation at 30 °C and 120 rpm (Figure 3). The enzyme production had reached the highest value of about 92.65 U/mL (100% relative activity) at its optimum incubation period of 48 h. Therefore, isolate YATW18 was selected for further study.
3.3. Molecular Identification
The phylogenetic relationship of the isolates is presented in Figure 4. The sequences of the isolate YATW18 showed ≥ 99% 16S rRNA gene sequence similarity to Alkalihalophilus pseudofirmus strain DSM 8715 (NR_026139) [19]. Accordingly the isolate was named as Alkalihalophilus pseudofirmus strain YATW18.
3.4. Physicochemical Characterization of Alkaline Protease
3.4.1. Effect of pH in Alkaliphilic Protease Activity
The enzyme showed a high activity within a broad pH range of 7.5 to 11.0, with relative activity of more than 65%, reaching its optimum at pH 10.0 (relative activity of 100%). Activity slightly declined beyond pH 10.0 and dropped sharply after pH 11.0 (Figure 5).
3.4.2. Effect of Temperature on Alkaliphilic Protease Activity and Stability
As shown in Figure 6A, the temperature profile analysis showed that the enzyme exhibited its lowest activity at 30 °C (approximately 30% relative activity). Relative activity increased to 60% at 40 °C and 80% at 50 °C, reaching a maximum of 100% at 55 °C which was identified as the optimal temperature for alkaline protease activity under the study conditions. Figure 6B shows the stability of the enzyme at four distinct temperatures (45 °C, 50 °C, 55 °C, and 60 °C) without calcium over 60 min. The enzyme remains stable, retaining more than 80% of its original activity at 45 °C and 50 °C, while drooped sharply at 55 °C and above. Figure 7C demonstrates the stability of the enzyme at different temperatures in the presence of 5 mM calcium, in 10 min intervals. On the contrary, the enzyme was stable even at 55 °C, retaining more than 80% of its original activity in the presence of 5 mM calcium (Figure 6C).
3.4.3. Effect of Salt Concentration on Alkaliphilic Protease Activity
As shown in Figure 7 the enzyme demonstrated a notable tolerance to high salt concentrations. It exhibited optimal enzyme activity at low salt contraptions 0% to 2.5% salt and retained more than 80% of its original; activity at 5% salt. At 10% and 15% salt, the enzyme maintained 60% and 40% of its original activity, respectively. Even at 20% salt, it retained 20% activity, demonstrating tolerance to moderately high salt levels.
3.4.4. Effect of Incubation Time in Alkaliphilic Protease Activity
The activity of the alkaliphilic protease in response to the incubation time is presented in Figure 8. The enzyme activity reached an optimum level at 20 min of incubation (Figure 8). At first, the relative activity increased rapidly, reaching out 60% at 5 min then increased to 85% at 10 min. This increment was sustained, and it was peaked 100% at 20 min. After this point, the relative activity remained relatively near this peak while oscillating slightly between 20 and 40 min in the process (Figure 8).
3.4.5. Effect of Metal Ions in Alkaliphilic Protease Activity
Most metal ions exhibited minimal effects on enzyme activity across all tested concentrations, with relative activity remaining close to or slightly below 100% (Figure 9). The enzyme showed high stability in the presence of boron (B3+), calcium (Ca2+), and magnesium (Mg2+) ions. In contrast, a slight reduction in activity was observed with nickel (Ni2+) and zinc (Zn2+) at 5 mM. Manganese (Mn2+) caused a more pronounced decline in enzyme activity with increasing concentration. Overall, the enzyme demonstrated good stability in the presence of B3+, Ca2+, and Mg2+, whereas higher concentrations of Ni2+, Zn2+, and Mn2+ resulted in mild inhibitory effects.
4. Discussion
High proteolytic index values for proteolytic potential were recorded in several microbial isolates, indicating a strong capacity for extracellular protease production. This potential was confirmed by their ability to hydrolyze skim milk agar, forming distinct clear zones around their colonies [20]. The presence of clear zones is considered a valid indicator of proteolytic activity, confirming the ability of these isolates to hydrolyze casein, the primary protein substrate in the skim milk agar medium (Figure 2). The proteolytic index values obtained for the alkaline protease producing bacterial strains varied, reflecting differences in enzyme production capacity, which are likely influenced by genetic factors and variations in metabolic rates. Accordingly, isolates producing larger clear zones were selected for secondary screening, as these large clear zones correlate with higher protease production. Consequently, such isolates are considered promising candidates for industrial applications [21].
The results of the present study demonstrate that Alkalihalophilus pseudofirmus strain YATW18 exhibits high efficiency in producing extracellular protease under submerged fermentation [22]. The high enzyme levels achieved by this strain within a relatively short incubation period underscore its potential for commercial protease production. These findings are consistent with reports from previous studies in the area [23] However, some differences were observed in terms of the time required to reach peak enzyme production and the maximum activity levels reported. In comparison to the study by Kumar et al. [24], this reported peak enzyme production after 72 hrs. The YATW18 strain in the present study achieved maximum production at just 48 h of incubation (Figure 3). This efficiency suggests that YATW18 would have a shorter enzyme production time, making it particularly advantageous for industrial applications [25]. The reduced incubation time observed is promising, as it could enhance the cost-effectiveness of protease production at large scale. Furthermore, this implies that strain YATW18 would have a shorter generation time and a higher metabolic rate compared to the other 22 alkaline protease producing bacterial strains evaluated during submerged fermentation.
The alkaline protease produced by Alkalihalophilus pseudofirmus strain YATW18 demonstrated effective activity across a pH range of 8 to 11, with optimal activity observed at pH 10 (Figure 5). This pH profile fits well with the requirements of the detergent industry, where enzymes, particularly proteases, are commonly used as additives to enhance cleaning efficiency. The ability of the YATW18 derived enzyme to function in alkaline conditions corresponds to the typical pH range (8–11) found in laundry detergents, making it a suitable candidate for industrial detergent formulations [26]. This pH positioning indicates that the enzyme is capable of effectively degrading and removing proteinaceous materials without being inactivated by the high pH conditions typically encountered during use. Its high activity across a broad pH range enhances its versatility, making it suitable for incorporation into various detergent formulations, including liquid, powder, and tablet forms. Beyond detergents, the leather industry also makes extensive use of proteolytic enzymes in key processing stages such as dehairing, bating, and soaking, where proteases facilitate the breakdown of non-collagenous proteins [27].
The protease from Alkalihalophilus pseudofirmus strain YATW18 was found to be highly efficient, and stable, qualities that make it particularly advantageous for tannery applications. Dehairing is typically performed using alkaline chemicals or enzymes to remove hair and epidermal materials. This protease functions optimally within a pH range of 8–11, with peak activity at pH 10, aligning well with the alkaline conditions commonly used in dehairing processes [28]. Additionally, the enzyme is potentially valuable in the bating process, where it helps remove unwanted proteins from hides, resulting in softer and more pliable leather. Its high activity in alkaline environments enhances the removal of non-collagenous proteins, ultimately improving the overall quality of the leather.
The temperature profile of the enzyme demonstrated a progressive increase in activity with rising temperatures from 30 °C to 55 °C, with optimal activity at 55 °C (Figure 6A). At 30 °C, the enzyme activity was relatively low, would be due to limited intermolecular interactions and reduced kinetic energy [29]. Activity increased to 60% at 40 °C and reached 80% at 50 °C, suggesting enhanced molecular motion and greater enzyme flexibility at elevated temperatures [30]. These results indicate that the enzyme performs more efficiently under moderately high temperatures, which is advantageous for various industrial applications requiring thermostable proteases. The enzyme exhibited maximum activity at 55 °C, which aligns with the optimal temperature range commonly reported for proteolytic enzymes [31]. The steady increase in activity from 40 °C to 55 °C indicates that the enzyme is well-suited for warm and hot wash cycles, which generally range between 40 °C and 60 °C. This temperature stability ensures the enzyme remains active during laundering, effectively breaking down protein-based stains. Furthermore, the enzyme’s partial activity at lower temperatures is relevant to the trend in modern detergent formulations, which are increasingly designed to perform efficiently at reduced temperatures (30 °C to 40 °C) to conserve energy [32]. The enzyme’s ability to retain 60% of its activity at 40 °C suggests that it can still play a significant role in protein degradation within energy-efficient, cold-water detergent formulations (Figure 7B). Its optimal performance at 55 °C aligns well with conditions commonly employed in industrial dishwashing and heavy-duty laundry applications [33]. High enzymatic activity at elevated temperatures also reduces the need for large enzyme dosages, thereby potentially lowering overall formulation costs. In tannery processes such as dehairing, temperatures are typically maintained between 40 °C and 55 °C to facilitate the breakdown of keratin and other non-collagenous proteins, making this protease particularly suitable for such applications. The enzyme’s peak activity at 55 °C indicates its ability to efficiently accelerate processes such as dehairing and bating, thereby reducing processing time and enhancing the overall efficiency of leather production. Given that leather processing often involves fluctuating temperatures, the enzyme’s broad temperature activity range ensures consistent performance across multiple stages of the workflow [34]. This thermal stability minimizes the need for strict temperature regulation, simplifying operational requirements and contributing to reduced energy consumption and lower processing costs.
The alkaline protease produced by Alkalihalophilus pseudofirmus strain YATW18 demonstrated notable tolerance to increasing salt concentrations, retaining significant enzymatic activity even in the presence of elevated salt levels (Figure 7). This characteristic is particularly valuable, as many household and industrial detergents are often compromised in areas with hard water or high mineral content, which can hinder enzyme performance [35]. The salt tolerance of this enzyme enhances its potential for use in detergent formulations designed for such challenging water conditions. A key advantage of this protease is its ability to maintain high activity in the presence of 5–10% salt concentrations, indicating strong stability in hard water and enhancing the performance of detergent formulations. Salt is commonly used in leather processing, particularly during the soaking and curing stages, for preservation and to inhibit microbial growth [36]. Remarkably, this enzyme remains active even at salt concentrations as high as 15%, allowing it to function effectively during these stages by facilitating the breakdown of non-collagenous proteins without being inhibited by the saline environment. This salt tolerance also supports its potential application in the early stages of leather processing, where raw hides are preserved in high-salt conditions. By reducing unwanted proteins and impurities under such conditions, the enzyme contributes to improving the quality and uniformity of the final leather product.
The enzyme activity profile shows a gradual increase in reaction rate during the initial incubation, followed by a sharp rise between the 5–10 min intervals (Figure 8). This suggests that the enzyme requires a short period to reach optimal catalytic efficiency, likely due to factors such as substrate binding, enzyme activation, or the establishment of reaction equilibrium [37]. Maximum activity, with 100% relative activity, was achieved at 20 min. At this point, nearly all active sites of the enzyme are fully occupied, and the reaction rate stabilizes, showing minimal further change over time [38]. The observed pattern of alkaline protease activity, an initial increase followed by stabilization, corresponds with classical enzyme kinetics, where the reaction rate accelerates until it reaches maximum velocity. This behavior reflects both the efficiency and stability of the enzyme over time, demonstrating its ability to sustain consistent activity once optimal conditions are established [39]. Such performance is particularly beneficial for industrial processes that demand precise timing or prolonged enzymatic action. The enzyme’s rapid attainment and maintenance of peak activity make it especially suitable for applications requiring both efficiency and reliability [40].
The effect of various metal ions, boron, zinc, nickel, cobalt, magnesium, copper, and calcium, on the activity of the alkaline protease was assessed, and the enzyme demonstrated stable performance across a range of concentrations, with no significant reduction in activity (Figure 9). These findings suggest that most of the tested metal ions do not substantially inhibit the enzyme’s activity [41]. The absence of major inhibition across all concentrations indicates that the system operates at or near baseline relative activity, exhibiting resilience to metal ion interference. This resistance is a valuable trait, particularly in industrial applications where metal ion fluctuations are common [42]. In the detergent industry, metal ions may be present either as functional additives or as residuals from raw materials used in production [43]. The enzyme’s ability to maintain activity under such chemically diverse conditions ensures consistent stain degradation, even complex formulations [44]. However, manganese was found to inhibit enzyme activity across all tested concentrations, indicating a potential inhibitory effect that may require consideration in specific industrial settings.
5. Conclusions
This study has successfully demonstrated a sustainable way of microorganisms intermediated tannery effluents transformation through the production and characterization of highly stable alkaline protease from Alkalihalophilus pseudofirmus strain YATW18.The enzyme exhibited high stability across a broad pH and temperature range, salt tolerance, and metal ions resistance, indicating its potential as a reliable and sustainable alternative for diverse industrial applications.
Supplementary Materials
Figure S1: Tyrosine standard curve.
Author Contributions
Conceptualization, Y. S. and A. S.; Methodology, Y. S.; Formal Analysis, Y. S.; Resources, A. S.; Data Curation, Y. S.; Writing—Original Draft Preparation, Y. S.; Writing—Review and Editing, A. S; Visualization, Y. S.; and Supervision, A.S.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data in this publication can be made available upon request to the first author.
Acknowledgments
The authors acknowledge Mr. Solomon Enquahone and Mrs. Tirunesh Shiferaw from AAU for their technical support during sample collection, microbial, and molecular experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahsan, S.; Liaqat, A.; Khaliq, A.; Iqbal, R.; Chughtai, M.; Mehmood, T.; Sameed, N. Current Trends and Prospects of Transforming Animal Waste into Food. In Climate Changes Mitigation and Sustainable Bioenergy Harvest through Animal Waste; Springer Nature: Cham, Switzerland, 2023; pp. 469–503.
- Mohamed, A.G.; El Mehalawy, A.A.; Mohamed, S.S. Loofah immobilized with Cladosporium cladosporioides CEL14 is a potential bio-remediating agent for hexavalent chromium in tannery wastewater. Malays. J. Microbiol. 2021, 17, 5.
- Nadeem, M.; Qazi, J.; Syed, Q.; Gulsher, M. Purification and characterization of an alkaline protease from Bacillus licheniformis UV-9 for detergent formulations. Songklanakarin J. Sci. Technol. 2013, 35, 2.
- Khan, Z.; Shafique, M.; Jabeen, N.; Naz, S.; Yasmeen, K.; Ejaz, U.; Sohail, M. Protease from Bacillus subtilis ZMS-2: Evaluation of production dynamics through Response Surface Methodology and application in leather tannery. J. King Saud Univ. Sci. 2023, 35, 102643.
- Amde, A. Environmental impact of Awash leather industry and tannery in Ethiopia. Ethiop. J. Environ. Stud. Manag. 2019, 12, 95–104.
- Gnanavelu, A.; Shanmuganathan, T.; Deepesh, V.; Suresh, S. Validation of a Modified Procedure for the determination of Chemical Oxygen Demand using standard dichromate method in industrial wastewater samples with high calcium chloride content. Indian J. Sci. Technol. 2021, 14, 2391–2399.
- Bennett, V.; MarkManuel, D. Winkler Titration Assessment of Biochemical Oxygen Demand (BOD5) in Oka Creek, Toru-Orua, Bayelsa State. Asian J. Appl. Chem. Res. 2024, 15, 147–156.
- Tiwari, N.; Singh, R.; Gupta, N. Enrichment and isolation of protease-producing alkaliphiles. Biotechnol. J. 2015, 14, 412–417.
- Gessesse, A.; Hatti-Kaul, R.; Gashe, B. Novel alkaline proteases from alkaliphilic bacteria: Purification and characterization. Enzyme Microb. Technol. 2003, 32, 589–594.
- Surzycki, S. Extraction and purification of nucleic acids using CTAB method. In Basic Techniques in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 163–168.
- Cropper, A. Quantitative assessment of DNA: A spectrophotometric and electrophoretic approach. J. Genet. Res. 2019, 42, 155–162.
- Brukner, I.; Paliouras, M.; Trifiro, M.; Bohbot, M.; Shamir, D.; Kirk, A. Assessing Different PCR Master Mixes for Ultrarapid DNA Amplification: Important Analytical Parameters. Diagnostics 2024, 14, 477.
- Bartlett, J.; Stirling, D. PCR Protocols, 2nd ed.; Humana Press: Totowa, NJ, USA, 2008.
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410.
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. Mamm. Protein Metab. 1969, 3, 132.
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549.
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791.
- Rawski, R.; Sanecki, P.; Kalembkiewicz, J. Units and Methods of Proteolytic Activity Determination. Curr. Pharm. Anal. 2020, 16, 661–670.
- Nielsen, P.; Fritze, D.; Priest, F.G. Phenetic diversity of alkaliphilic Bacillus strains: proposal for Alkalihalophilus pseudofirmus sp. nov. Nord. Microbiol. 1995, 45, 120–127.
- Ullah, N.; Mujaddad-ur-Rehman, M.; Sarwar, A.; Nadeem, M.; Nelofer, R.; Irfan, M.; Aziz, T. Effect of bioprocess parameters on alkaline protease production by locally isolated Bacillus cereus AUST-7 using tannery waste in submerged fermentation. Biomass Convers. Biorefin. 2023, 14, 22977–22987.
- Masi, C.; Gemechu, G.; Tafesse, M. Isolation, screening, characterization, and identification of alkaline protease-producing bacteria from leather industry effluent. Ann. Microbiol. 2021, 71, 1–11.
- Singh, K.; Kumari, M.; Prasad, K. Tannery effluents: current practices, environmental consequences, human health risks, and treatment options. CLEAN–Soil Air Water 2023, 51, 2200303.
- Jiang, X.; Xu, X.; Yang, M.; Zhang, M. Emerging applications of solid-state fermentation in bioengineering and biopharmaceuticals. Curr. Opin. Biotechnol. 2021, 70, 54–59.
- Kumar, R.; Tiwari, P.; Jany, K. Comparative production of proteases in Bacillus sp. under submerged fermentation. J. Environ. Biol. 2012, 33, 203–207.
- Gupta, R.; Beg, Q.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32.
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial proteases applications. Front. Bioeng. Biotechnol. 2019, 7, 110.
- Wanyonyi, W.W.; Mulaa, F.J. Alkaline proteases: Advances and applications in the leather industry. Enzyme Eng. 2019, 8, 1–10.
- Sharma, R.; Bhatt, M.; Agrawal, A. A comparative study of chemical and enzyme-assisted leather dehairing. Environ. Technol. Innov. 2018, 9, 235–241.
- Taylor, R. Identifying intermolecular atom-atom interactions that are not just bonding but also competitive. CrystEngComm 2020, 22, 7145–7151.
- Govindasamy, V.; Kumar, M. Thermostable alkaline protease production from Bacillus licheniformis under solid-state fermentation: Purification and characterization. J. Genet. Eng. Biotechnol. 2021, 19, 1–10.
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules 2021, 26, 1858.
- Cortez, D.; Bekke, M.; Liang, Z.; Stamminger, R. The impact of detergent performance on sustainable consumer laundry behavior: a socio-technical challenge. Tenside Surfactants Deterg. 2024, 61, 203–215.
- Stoner, M.R. Mechanistic studies of enzyme degradation in liquid detergent. PhD Thesis, University of Colorado at Boulder, USA, 2004.
- Sivakumar, V. Towards environmental protection and process safety in leather processing–A comprehensive analysis and review. Process Saf. Environ. Prot. 2022, 163, 703–726.
- Galante, Y.; Formantici, C. Enzyme applications in detergency and in manufacturing industries. Curr. Org. Chem. 2003, 7, 1399–1422.
- Samidurai, S.; Khambhaty, Y.; Alagamuthu, T. Bio-preservation of raw hides/skins: A review on greener substitute to conventional salt curing. Environ. Sci. Pollut. Res. 2022, 29, 64513–64535.
- Punekar, N.S. Enzymes: Catalysis, Kinetics and Mechanisms; Springer: Mumbai, India, 2018.
- Romero, E.; Gómez Castellanos, J.; Gadda, G.; Fraaije, M.; Mattevi, A. Same substrate, many reactions: oxygen activation in flavoenzymes. Chem. Rev. 2018, 118, 1742–1769.
- Callender, R.; Dyer, R. The dynamical nature of enzymatic catalysis. Acc. Chem. Res. 2015, 48, 407–413.
- Chapman, J.; Ismail, A.; Dinu, C. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238.
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.; Thornton, J. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2012, 17, 1091–1098.
- Shahid, M.; Noman, M.; Ahmed, T. Enzyme stabilization: Stabilization of proteases for industrial applications. Crit. Rev. Biotechnol. 2017, 37, 309–322.
- Baldiris, R.; Pulido, P.; Pulido, H.; Montes, J.; Cardozo, A.; Marrugo-Negrete, J. Effects of manganese on oxidative stress in the brain. Neurotox. Res. 2019, 35, 664–670.
- Jakubowski, M.; Kuciel, R.; Chelstowska, A. Heavy metals and enzyme activity: Influence on enzymes and their removal from industrial wastewater. Water Environ. Res. 2011, 83, 278–289.
Figure 1.
Sample area description of Awash tannery with respect to sample points in treatment plant; thus, S1 (aerobic digestion), S2 (bulk), S3 (homogenization), and S4 (secondary treatment).
Figure 1.
Sample area description of Awash tannery with respect to sample points in treatment plant; thus, S1 (aerobic digestion), S2 (bulk), S3 (homogenization), and S4 (secondary treatment).

Figure 2.
Isolation and purification of strain YATW18 through further streaking

Figure 3.
Enzyme activity of strain YATW18 at 12-hour intervals during the incubation period

Figure 4.
Neighbor-joining phylogenetic tree construction showing the relationship among the alkaline protease producing bacterial strains studied and close matches; on the bases of 16S rRNA gene sequence. Numbers at nodes indicate percentages of occurrence in 1000 bootstrapped trees; only values greater than 50% are shown. Scale bar 0.05 number of substitutions per nucleotide position. Pseudomonas aeruginosa NO2 (FJ972530) was used as an out-group. Alkalihalophilus pseudofirmus strain YATW18 was isolated in this study.
Figure 4.
Neighbor-joining phylogenetic tree construction showing the relationship among the alkaline protease producing bacterial strains studied and close matches; on the bases of 16S rRNA gene sequence. Numbers at nodes indicate percentages of occurrence in 1000 bootstrapped trees; only values greater than 50% are shown. Scale bar 0.05 number of substitutions per nucleotide position. Pseudomonas aeruginosa NO2 (FJ972530) was used as an out-group. Alkalihalophilus pseudofirmus strain YATW18 was isolated in this study.

Figure 5.
Effect of pH in alkaline protease activity.

Figure 6.
Effect of temperature on the activity and stability of alkaline protease (A) alkaline protease activity (B) alkaline protease stability without calcium (C) alkaline protease stability with 5mM calcium.
Figure 6.
Effect of temperature on the activity and stability of alkaline protease (A) alkaline protease activity (B) alkaline protease stability without calcium (C) alkaline protease stability with 5mM calcium.

Figure 7.
Effect of salt concentration in alkaline protease activity

Figure 8.
Effect of incubation time in alkaline protease activity

Figure 9.
Effect of some metals on alkaline protease activity


Table 1.
Characterization of tannery wastewater samples concerning different parameters.
| No. | Parameters | Measured value | Site of measurement |
| 1. | pH | 8.1 | On-site measurement |
| 2. | Temperature | 28 °C | |
| 3. | Salinity | 1% | |
| 4. | TDS | 9.63 ppt | Off-site measurement |
| 5. | TSS | 1044.7 mg/L | |
| 6. | COD | 1276 mg/L | |
| 7. | BOD | 375.4 mg/L |
Table 2.
Clear zone comparison of bacterial isolates
| No. | Bacterial isolates | Colony diameter (mm) | Clear zone diameter (mm) | Protolytic index (ratio) |
| 1. | YATW1 | 3.7 | 18.7 | 5.1 |
| 2. | YATW2 | 3 | 19.7 | 6.6 |
| 3. | YATW3 | 3 | 17.3 | 5.8 |
| 4. | YATW4 | 3.3 | 21.3 | 6.5 |
| 5. | YATW5 | 3.7 | 9.3 | 2.5 |
| 6. | YATW6 | 3 | 22 | 7.3 |
| 7. | YATW7 | 2.7 | 20.7 | 7.7 |
| 8. | YATW8 | 3.3 | 20.7 | 6.3 |
| 9. | YATW9 | 2.7 | 21.3 | 7.9 |
| 10. | YATW10 | 2.7 | 17.3 | 6.4 |
| 11. | YATW11 | 3.3 | 22 | 6.7 |
| 12. | YATW12 | 3.7 | 22 | 5.9 |
| 13. | YATW13 | 2.7 | 21.3 | 7.1 |
| 14. | YATW14 | 3 | 8.7 | 2.9 |
| 15. | YATW15 | 3.3 | 21.3 | 6.5 |
| 16. | YATW16 | 2.7 | 8 | 3 |
| 17. | YATW17 | 3 | 9.3 | 3.1 |
| 18. | YATW18 | 3 | 20.7 | 6.9 |
| 19. | YATW19 | 3.7 | 22 | 5.9 |
| 20. | YATW20 | 4 | 20.7 | 5.2 |
| 21. | YATW21 | 2.7 | 20 | 7.4 |
| 22. | YATW22 | 3.3 | 17.3 | 5.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



