Submitted:
04 April 2026
Posted:
07 April 2026
You are already at the latest version
Abstract
This study evaluated the effects of light spectral quality on shoot yield and essential oil of Tagetes erecta L. cultivated in controlled growth chambers. Plants were grown for up to 101 days under three LED lighting treatments with different red, blue, and white wavelength ratios and a constant 16 h photoperiod. The F2 treatment (5 red:1 blue) produced yields of fresh shoots, early blooming flowers, and oils of 271 ± 28 g/tray, 97.43 ± 13.14 g/tray, and 52.46 ± 5.41 mg/tray, respectively. These values were significantly higher (p < 0.05) than those of the F1 treatment (white:red-phosphor), and represented increases of 1.37-, 1.26-, and 1.38-fold, respectively. Gas chromatography identified three major oil constituents—(E)-β-ocimene (22.9–28.8%), (E)-myroxide (13.9–20.6%), and piperitone (7.3–9.6%)—among a total of 24—25 compounds. Essential oils inhibited from four to five of the seven tested microbial strains, with the notable activity against Escherichia coli and Candida albicans recorded in F2 and F1, respectively. These findings confirm that light spectral quality is a critical factor regulating flower, essential oil yield, and antimicrobial efficacy in T. erecta, and support the use of optimized LED spectra as a practical approach to improve plant’s yield and phytochemical quality.
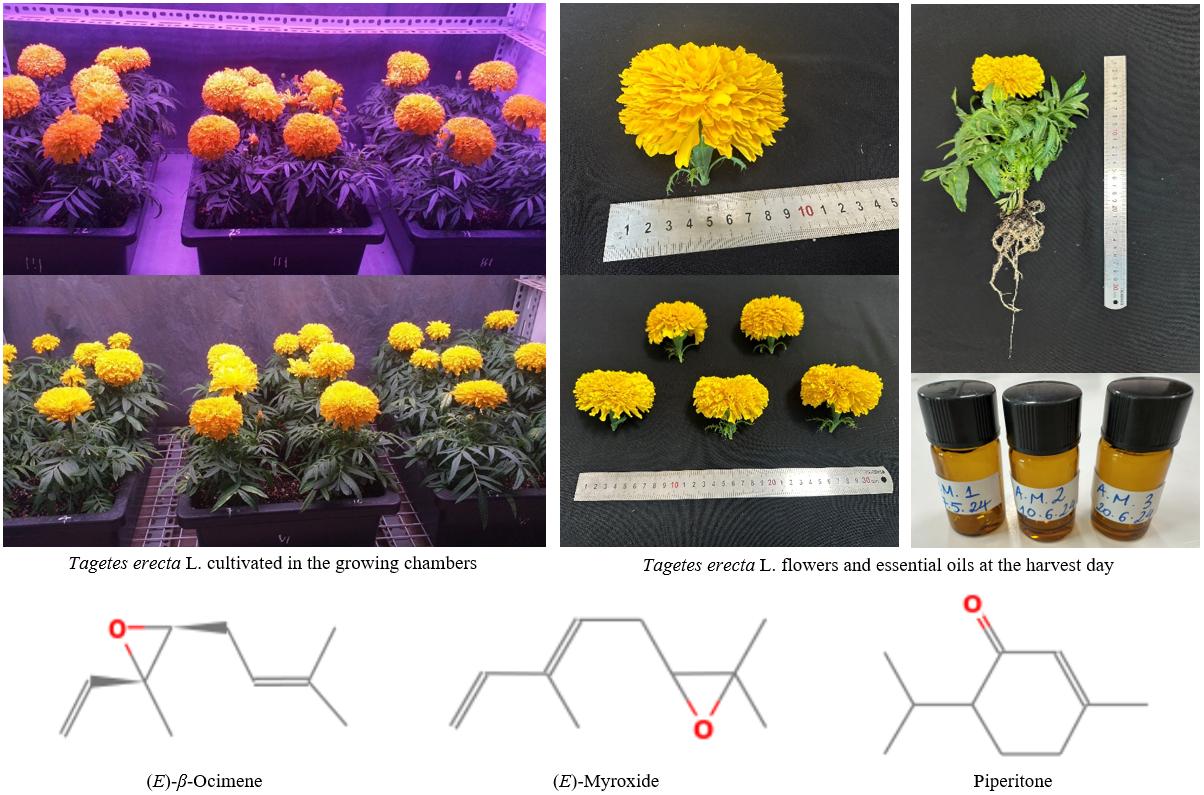
Keywords:
1. Introduction
2. Results and Discussions
2.1. The Effect of LED Light Conditions on Biomass and Essential Oil Yield of Tagetes Erecta
2.2. The Effect of LED Light Conditions on Essential Oil Composition of Tagetes Erecta
2.3. The Effect of LED Light Conditions on Antimicrobial Activity of the Essential Oil of Tagetes Erecta
3. Materials and Methods
3.1. Plant Materials and Lighting Conditions

| Treatments | F1 | F2 | F3 |
| LED ratio | White: Red-phosphor (WRp) | 5 Red : 1 Blue (5R1B) | 3 Red : 2 Blue : 1 White (3R2B1W) |
|
Spectral distribution (%) {(UV < 400 nm) : (B ~ 400-500 nm) : (G ~ 501-600 nm) : (R ~ 601-700nm) : (Fr >700 nm)} |
0.06 : 17.33 : 5.88 : 64.28 : 12.45 | 0.07 : 25.82 : 0.52 : 73.45 : 0.14 | 0.09 : 42.05 : 10.22 : 47.30 : 0.34 |
| Total of B & R (%) | 81.61 | 99.27 | 89.35 |
| The reduced ratio of R to B | 3.71 : 1 | 2.84 : 1 | 1.12 : 1 |
| Duration (h/day) | 16 | 16 | 16 |
| Lighting time | 6:00-22:00 | 6:00-22:00 | 6:00-22:00 |
| Light intensity (lux) | 2500 ± 100 | 2500 ± 100 | 2500 ± 100 |
| Light intensity (µmol·m-2·s-1) | 206 ± 8.2 | 208 ± 8.3 | 199 ± 8.0 |
| Total daily light (mol·m-2 ·d-1) | 11.866 ± 0.472 | 11.981 ± 0.478 | 11.462 ± 0.461 |
3.2. Essential Oil Isolation
3.3. Essential Oil GC-MS and GC-FID Analysis
3.4. Tested Microbial Strains
3.5. Screening of Antimicrobial Activity of Essential Oil
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| B | blue |
| DLI | daily light integral |
| Fr | far red |
| G | green |
| IC₅₀ | half-maximal inhibitory concentration |
| MIC | minimum inhibitory concentration |
| R | red |
| Rp | red-phosphor |
| Tr | trace |
| UV | ultra violet |
| W | white |
References
- Marotti, M.; Piccaglia, R.; Biavati, B.; Marotti, I. Characterization and Yield Evaluation of Essential Oils from Different Tagetes Species. J. Essent. Oil Res. 2004, 16, 440–444. [Google Scholar] [CrossRef]
- Cristina e Santos, P.; Granero, F.O.; Junior, J.L.B.; Pavarini, R.; Pavarini, G.M.P.; Chorilli, M.; Zambom, C.R.; Silva, L.P.; Silva, R.M.G. da Insecticidal Activity of Tagetes Erecta and Tagetes Patula Extracts and Fractions Free and Microencapsulated. Biocatal. Agric. Biotechnol. 2022, 45, 102511. [Google Scholar] [CrossRef]
- Sharma, G.; Rajhansa, K.C.; Sharma, P.; Singh, A.; Sharma, A.; Sahu, M.K.; Sharma, R.; Pandey, A.K. Marigold (Tagetes Spp.): A Diverse Crop with Multipurpose Value for Health and Environment: A Review. Agric. Rev. 2024, 45, 618–626. [Google Scholar] [CrossRef]
- Talukdar, N.; Kashyap, B.; Barman, I.; Gogoi, J.; Kalita, P.P. A Review on Tagetes Erecta (Marigold) with Reference to Its Pharmacologicalimportance. Indian J. Nat. Sci. 2023, 14, 56465–56472. [Google Scholar]
- Siddiqa, A.; Khaliq, A.; Mehmood, T.; Chughtai, M.F.J.; Sanchez-Migallon, A.M.; Ahsan, S.; Sabir, A.; Mohamed Ahmed, I.A. Phytochemical Profiling of Tagetes Erecta L. Flowers at Various Blooming Stages through Optimized Extraction of Bioactive Compounds for the Development of Functional Juice. Front. Sustain. Food Syst. 2025, 9. [Google Scholar] [CrossRef]
- Salehi, B.; Valussi, M.; Morais-Braga, M.F.; Carneiro, J.N.; Leal, A.L.; Coutinho, H.D.; Vitalini, S.; Kręgiel, D.; Antolak, H.; Sharifi-Rad, M.; et al. Tagetes Spp. Essential Oils and Other Extracts: Chemical Characterization and Biological Activity. Molecules 2018, 23, 2847. [Google Scholar] [CrossRef]
- Ravikumar, P. Chemical Examination and Insecticidal Properties of Tagetes Erecta and Tagetes Patula. Asian J. Biol. Sci. 2010, 5, 29–31. [Google Scholar]
- Gupta, P.; Vasudeva, N. A Potential Ornamental Plant Drug. Hamdard Med. 2012, 55, 45–59. [Google Scholar]
- Singh, P.; Krishna, A.; Kumar, V.; Krishna, S.; Singh, K.; Gupta, M.; Singh, S. Chemistry and Biology of Industrial Crop Tagetes Species: A Review. J. Essent. Oil Res. 2016, 28, 1–14. [Google Scholar] [CrossRef]
- Laosinwattana, C.; Wichittrakarn, P.; Teerarak, M. Chemical Composition and Herbicidal Action of Essential Oil from Tagetes Erecta L. Leaves. Ind. Crops Prod. 2018, 126, 129–134. [Google Scholar] [CrossRef]
- Kumar, A.; Gautam, R.D.; Kumar, A.; Singh, S.; Singh, S. Understanding the Effect of Different Abiotic Stresses on Wild Marigold (Tagetes Minuta L.) and Role of Breeding Strategies for Developing Tolerant Lines. Front. Plant Sci. 2022, Volume 12. [Google Scholar] [CrossRef]
- Ahmadpour, V.; Modarresi, M.; Eftekhari, M.; Saeedi, M.; Karimi, N.; Rasekhian, M. Chemical Composition of Essential and Fixed Oils of Tagetes Erecta Fruits (Iran) and Their Implications in Inhibition of Cancer Signaling. Sci. Rep. 2024, 14, 19667. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Lal, M.A. Plant Physiology, Development and Metabolism, 2nd ed.; Springer Singapore, 2023. [Google Scholar]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue Light Dose–Responses of Leaf Photosynthesis, Morphology, and Chemical Composition of Cucumis Sativus Grown under Different Combinations of Red and Blue Light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, L.; Zou, H.; Qiu, L.; Zheng, Y.; Yang, D.; Wang, Y. Effects of Light on Secondary Metabolite Biosynthesis in Medicinal Plants. Front. Plant Sci. 2021, Volume 12. [Google Scholar] [CrossRef] [PubMed]
- Gangadhar, B.H.; Mishra, R.K.; Pandian, G.; Park, S.W. Comparative Study of Color, Pungency, and Biochemical Composition in Chili Pepper (Capsicum Annuum) Under Different Light-Emitting Diode Treatments. HortScience 2012, 47, 1729–1735. [Google Scholar] [CrossRef]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal Red:Blue Ratio in Led Lighting for Nutraceutical Indoor Horticulture. Sci. Hortic. (Amsterdam). 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Flores-Pérez, S.; Castillo-González, A.M.; Valdez-Aguilar, L.A.; Avítia-García, E.; Flores-Pérez, S.; Castillo-González, A.M.; Valdez-Aguilar, L.A.; Avítia-García, E. Use of Different Proportions of Red and Blue LEDs to Improve the Growth of Lilium Spp. Rev. Mex. ciencias agrícolas 2021, 12, 835–847. [Google Scholar]
- Jeong, W.H.; Chun, W.L.; Kee, Y.P. Influence of Mixed LED Radiation on the Growth of Annual Plants. J. Plant Biol. 2006, 49, 286–290. [Google Scholar] [CrossRef]
- Ngcobo, B.L. Influence of LED Lights on Growth, Development, and Antioxidant Capacity of Marigold Plants in Protected Cultivation. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS), Leuven, Belgium, September 30 2025; pp. 119–124. [Google Scholar]
- Spall, C.E.; Lopez, R.G. Daily Light Integral and/or Photoperiod during the Young Plant and Finishing Stages Influence Floral Initiation and Quality of Witchgrass and Marigold Cut Flowers. Front. Plant Sci. 2022, 13, 956157. [Google Scholar] [CrossRef]
- Heo, J.W.; Kim, D.E.; Kang, K.K.; Park, S.H.; Chun, C. Growth and Flowering before and after Storage of African Marigold and Salvia Seedlings Stored under Different Light Conditions. Hortic. Sci. Technol. 2013, 31, 400–406. [Google Scholar] [CrossRef]
- Stutte, G.W. Controlled Environment Production of Medicinal and Aromatic Plants. In Medicinal and Aromatic Crops: Production, Phytochemistry, and Utilization; ACS Symposium Series; American Chemical Society, 2016; Vol. 1218, pp. 4–49. ISBN 9780841231276. [Google Scholar]
- Boyd, A.P.; Zankowski, P.; Wheeler, R.; Stokes-Draut, J.R.; Chudnovsky, Y.; Ingram, D.; Tijerina, M.; Mickens, M.; Steward, D.; Armstrong, K.; et al. Controlled Environment Agriculture: An Opportunity to Strengthen Interagency Research Collaboration in the US Government. PNAS Nexus 2025, 4, pgaf155. [Google Scholar] [CrossRef]
- Dsouza, A.; Dixon, M.; Shukla, M.; Graham, T. Harnessing Controlled-Environment Systems for Enhanced Production of Medicinal Plants. J. Exp. Bot. 2025, 76, 76–93. [Google Scholar] [CrossRef]
- Casal, J.J. Phytochromes, Cryptochromes, Phototropin: Photoreceptor Interactions in Plants. Photochem. Photobiol. 2000, 71, 1–11. [Google Scholar] [CrossRef]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.J.; Liu, D. Cryptochromes: Blue Light Receptors for Plants and Animals. Science 1999, 284, 760–765. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant Responses to Red and Far-Red Lights, Applications in Horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Gourrierec, J. Le; Morel, P.; Sakr, S.; Leduc, N. Light Signaling and Plant Responses to Blue and UV Radiations—Perspectives for Applications in Horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Nie, W.-F.; Li, Y.; Chen, Y.; Zhou, Y.; Yu, T.; Zhou, Y.; Yang, Y. Spectral Light Quality Regulates the Morphogenesis, Architecture, and Flowering in Pepper (Capsicum Annuum L.). J. Photochem. Photobiol. B. 2023, 241, 112673. [Google Scholar] [CrossRef] [PubMed]
- Cerdán, P.D.; Chory, J. Regulation of Flowering Time by Light Quality. Nature 2003, 129. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular Mechanisms Underlying Phytochrome-Controlled Morphogenesis in Plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef]
- C. Sager, J.; O. Smith, W.; L. Edwards, J.; L. Cyr, K. Photosynthetic Efficiency and Phytochrome Photoequilibria Determination Using Spectral Data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Reed, J.W.; Nagatani, A.; Elich, T.D.; Fagan, M.; Chory, J. Phytochrome A and Phytochrome B Have Overlapping but Distinct Functions in Arabidopsis Development. Plant Physiol. 1994, 104, 1139–1149. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. (Eds.) Plant Physiology and Development.; Sinauer Associates Incorporated, 2015; ISBN 9781605353531. [Google Scholar]
- Meng, Q.; Runkle, E.S. Regulation of Flowering by Green Light Depends on Its Photon Flux Density and Involves Cryptochromes. Physiol. Plant. 2019, 166, 762–771. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Diverse Flowering Response to Blue Light Manipulation: Application of Electric Lighting in Controlled-Environment Plant Production. Horticulturae 2024, 10, 578. [Google Scholar] [CrossRef]
- Hamamoto, H.; Shimaji, H.; Higashide, T. Budding and Bolting Responses of Horticultural Plants to Night-Break Treatments with LEDs of Various Colors. J. Agric. Meteorol. 2003, 59, 103–110. [Google Scholar] [CrossRef]
- Yang, J.; Song, J.; Jeong, B.R. Low-Intensity Blue Light Supplemented during Photoperiod in Controlled Environment Induces Flowering and Antioxidant Production in Kalanchoe. Antioxidants 2022, 11, 811. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Runkle, E.S. Low-Intensity Blue Light in Night-Interruption Lighting Does Not Influence Flowering of Herbaceous Ornamentals. Sci. Hortic. (Amsterdam). 2015, 186, 230–238. [Google Scholar] [CrossRef]
- Chory, J. Light Signal Transduction: An Infinite Spectrum of Possibilities. Plant J. 2010, 61, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth Responses of Marigold and Salvia Bedding Plants as Affected by Monochromic or Mixture Radiation Provided by a Light-Emitting Diode (LED). Plant Growth Regul. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Paucek, I.; Pennisi, G.; Pistillo, A.; Appolloni, E.; Crepaldi, A.; Calegari, B.; Spinelli, F.; Cellini, A.; Gabarrell, X.; Orsini, F.; et al. Supplementary LED Interlighting Improves Yield and Precocity of Greenhouse Tomatoes in the Mediterranean. Agronomy 2020, 10, 1002. [Google Scholar] [CrossRef]
- Avercheva, O. V; Berkovich, Y.A.; Erokhin, A.N.; Zhigalova, T. V; Pogosyan, S.I.; Smolyanina, S.O. Growth and Photosynthesis of Chinese Cabbage Plants Grown under Light-Emitting Diode-Based Light Source. Russ. J. Plant Physiol. 2009, 56, 14–21. [Google Scholar] [CrossRef]
- Cao, G.; Zhang, G.; Yu, J.; Ma, Y. Effects of Different Led Light Qualities on Cucumber Seedling Growth and Chlorophyll Fluorescence Parameters. Sci. Agric. Sin. 2013, 46, 1297–1304. [Google Scholar]
- Nanya, K.; Ishigami, Y.; Hikosaka, S.; Goto, E. Effects of Blue and Red Light on Stem Elongation and Flowering of Tomato Seedlings. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS), Leuven, Belgium, 14 October 2012; pp. 261–266. [Google Scholar]
- Deram, P.; Lefsrud, M.G.; Orsat, V. Supplemental Lighting Orientation and Red-to-Blue Ratio of Light-Emitting Diodes for Greenhouse Tomato Production. HortScience 2014, 49, 448–452. [Google Scholar] [CrossRef]
- Chen, Y.; Bian, Z.; Marcelis, L.F.M.; Heuvelink, E.; Yang, Q.; Kaiser, E. Green Light Is Similarly Effective in Promoting Plant Biomass as Red/Blue Light: A Meta-Analysis. J. Exp. Bot. 2024, 75, 5655–5666. [Google Scholar] [CrossRef]
- Sams, C.E.; Kopsell, D.; Morrow, R.C. Light Quality Impacts on Growth, Flowering, Mineral Uptake and Petal Pigmentation of Marigold. Acta Hortic. 2016, 1134, 139–145. [Google Scholar] [CrossRef]
- Keshavarz, M.; Matloobi, M.; Alizadeh-Salteh, S.; Rezghiyan, A. Optimizing Plant Growth and Pigment Profiles of African Marigold (Tagetes Erecta L.) under Different Light Spectra. BMC Plant Biol. 2025, 25, 1619. [Google Scholar] [CrossRef]
- Klein, R.M.; Edsall, P.C.; Gentile, A.C. Effects of Near Ultraviolet and Green Radiations on Plant Growth. Plant Physiol. 1965, 40, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Park, J.U.; An, S.K.; Kim, J. Far-Red Light Affects Stomatal Opening and Evapotranspiration of Sweet Basil. Horticulturae 2023, 9, 1095. [Google Scholar] [CrossRef]
- Taweesak, V.; Boonsong, E. Effects of Different Light-Emitting Diode (LED) Illumination on Growth and Flowering in Chrysanthemum. Int. J. Agric. Biol. 2025, 33, 330601. [Google Scholar] [CrossRef]
- Machado, M.I.L.; Silva, M.G. V; Matos, F.J.A.; Craveiro, A.A.; Alencar, J.W. The Presence of Indole as Minor Constituent of Tagetes Erecta Leaf Oil. J. Essent. Oil Res. 1994, 6, 203–205. [Google Scholar] [CrossRef]
- Heo, J.W.; Kang, D.H.; Bang, H.S.; Hong, S.G.; Chun, C.; Kang, K.K. Early Growth, Pigmentation, Protein Content, and Phenylalanine Ammonia-lyase Activity of Red Curled Lettuces Grown under Different Lighting Conditions. 2012, 30, 6–12. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Arzani, A.; Sabzalian, M.R. Thymol, Carvacrol, and Antioxidant Accumulation in Thymus Species in Response to Different Light Spectra Emitted by Light-Emitting Diodes. Food Chem. 2020, 307, 125521. [Google Scholar] [CrossRef]
- de Oliveira, P.F.; Alves, J.M.; Damasceno, J.L.; Oliveira, R.A.M.; Dias, H.J.; Crotti, A.E.M.; Tavares, D.C. Cytotoxicity Screening of Essential Oils in Cancer Cell Lines. Rev. Bras. Farmacogn. 2015, 25, 183–188. [Google Scholar] [CrossRef]
- Krishna, A.; Kumar, S.; Mallavarapu, G.R.; Ramesh, S. Composition of the Essential Oils of the Leaves and Flowers of Tagetes Erecta L. J. Essent. Oil Res. 2004, 16, 520–522. [Google Scholar] [CrossRef]
- Cerrón-Mercado, F.; Perez-Alvarez, J.A.; Nolazco-Cama, D.; Salva-Ruíz, B.; Tellez-Monzon, L.; Fernández-López, J.; Viuda-Martos, M. Chemical Composition, Antioxidant and Antibacterial Activities of Essential Oil Obtained from Chincho (Tagetes Elliptica Sm) Leaves Grown in the Peruvian Andes. Foods 2023, 12, 894. [Google Scholar] [CrossRef]
- Singh, G.; Singh, O.P.; De Lampasona, M.P.; Catalán, C.A.N. Studies on Essential Oils. Part 35: Chemical and Biocidal Investigations on Tagetes Erecta Leaf Volatile Oil. Flavour Fragr. J. 2003, 18, 62–65. [Google Scholar] [CrossRef]
- Martínez, R.; Diaz, B.; Vásquez, L.; Compagnone, R.S.; Tillett, S.; Canelón, D.J.; Torrico, F.; Suárez, A.I. Chemical Composition of Essential Oils and Toxicological Evaluation of Tagetes Erecta and Tagetes Patula from Venezuela. J. Essent. Oil Bear. Plants 2009, 12, 476–481. [Google Scholar] [CrossRef]
- Adebiyi, E.O.; Igberaese, P.O.; Ajoba, F.B.; Alao, A.A.; Aderibigbe, R.O.; Ayoola, D.O. Repeated-Dose Dermal Toxicity of Tagetes Erecta Essential Oil in Wistar Rats. African J. Adv. Sci. Technol. Res. 2026, 22, 1–10. [Google Scholar]
- Safar, A.A.; Ghafoor, A.O.; Dastan, D. Screening of Chemical Characterization, Antifungal and Cytotoxic Activities of Essential Oil Constituents of Tagetes Erecta L. from Erbil, Kurdistan Region-Iraq. Polish J. Environ. Stud. 2020, 29, 2317–2326. [Google Scholar] [CrossRef]
- Ptak, P.; Górecki, K.; Heleniak, J.; Orlikowski, M. Investigations of Electrical and Optical Parameters of Some LED Luminaires—A Study Case. Energies 2021, 14, 1612. [Google Scholar] [CrossRef]
- Ministry of Health Vietnamese Pharmacopoeia; Medical Publishing House: Hanoi, Vietnam, 2018.
- König, W.A.; Joulain, D.; Hochmuth, D.H. Terpenoids and Related Constituents of Essential Oils. Libr. MassFinder Hamburg, Ger. 2004, 2. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography, Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2017; ISBN 978-1-932633-21-4. [Google Scholar]
- NIST Chemistry Webbook, NIST Standard Reference Database Number 69; Linstrom, P.J., Mallard, W.G., Eds.; National Institute of Standards and Technology: Gaithersburg MD, 2021; p. 20899. [Google Scholar]
- Hadacek, F.; Greger, H. Testing of Antifungal Natural Products: Methodologies, Comparability of Results and Assay Choice. Phytochem. Anal. 2000, 11, 137–147. [Google Scholar] [CrossRef]
- Cos, P.; Vlietinck, A.J.; Berghe, D. Vanden; Maes, L. Anti-Infective Potential of Natural Products: How to Develop a Stronger in Vitro “Proof-of-Concept”. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef] [PubMed]


| Treatment | Fresh yield of shoot (g/tray) | Water content (%) | Dry yield of shoot (g/tray) | Essential oil content (% w/w, dry) |
Essential oil yield (mg/tray) |
|---|---|---|---|---|---|
| F1 | 198 ± 29b | 89.2 ±0.095c | 21.37 ± 3.15b | 0.1541 ± 0.0101c | 37.95 ± 5.59c |
| F2 | 271 ± 28a | 90.2 ± 0.124a | 26.57 ± 2.74a | 0.2285 ± 0.0155a | 52.46 ± 5.41a |
| F3 | 268 ± 21a | 90.0 ± 0.131b | 26.81 ± 2.10a | 0.1974 ± 0.0126b | 43.11 ± 3.38b |
| Compoundsa | RIb | F1c | F2c | F3c |
|---|---|---|---|---|
| α-Pinene | 940 | 0.2 | 0.2 | 0.2 |
| Sabinene | 980 | 0.4 | 0.4 | 0.4 |
| Myrcene | 993 | 3.3 | 2.3 | 2.5 |
| (3Z)-Hexenyl acetate | 1006 | 0.1 | 0.2 | 0.1 |
| Limonene | 1035 | 5.2 | 5.5 | 5.7 |
| (Z)-β-Ocimene | 1039 | 3.2 | 3.5 | 3.8 |
| (E)-β-Ocimene | 1050 | 22.9 | 27.3 | 28.8 |
| γ-Terpinene | 1064 | Tr | 0.1 | Tr |
| Terpinolene | 1095 | 4.5 | 5.5 | 5.1 |
| Linalool | 1102 | 0.8 | 0.6 | 0.5 |
| 1,3,8-p-Menthatriene | 1119 | 0.3 | 0.3 | 0.3 |
| (Z)-Myroxide | 1134 | 0.2 | 0.1 | 0.1 |
| (E)-Myroxide | 1145 | 20.6 | 15.0 | 13.9 |
| iso-Menthol | 1180 | 0.2 | 0.1 | 0.1 |
| Terpinen-4-ol | 1187 | 0.1 | 0.1 | 0.1 |
| (3Z)-Hexenyl butanoate | 1190 | 0.5 | 0.9 | 0.8 |
| p-Cymen-8-ol | 1192 | 0.2 | Tr | Tr |
| p-Methylacetophenone | 1193 | 0.2 | Tr | Tr |
| Octyl acetate | 1211 | Tr | Tr | 0.3 |
| 2-Phenylethyl acetate | 1263 | 2.7 | 2.3 | 2.3 |
| Piperitone | 1266 | 7.3 | 8.7 | 9.6 |
| 3-Thujyl acetate | 1296 | 4.0 | 4.8 | 5.1 |
| Indole | 1302 | 1.2 | 1.1 | 1.1 |
| Piperitenone | 1354 | 6.2 | 5.4 | 4.3 |
| Piperitenone oxide | 1378 | 2.6 | 1.7 | 2.1 |
| α-Santalene | 1433 | Tr | Tr | 0.2 |
| (E)-β-Caryophyllene | 1439 | 2.7 | 3.4 | 2.9 |
| (Z)-β-Farnesene | 1461 | 0.5 | 0.5 | 0.5 |
| Germacrene D | 1500 | 1.5 | 1.6 | 1.4 |
| Bicyclogermacrene | 1516 | 0.9 | 1.3 | 1.1 |
| (E)-Nerolidol | 1570 | 0.3 | 0.4 | 0.3 |
| Spathulenol | 1598 | 0.1 | 0.1 | Tr |
| Caryophyllene oxide | 1606 | 0.2 | 0.3 | 0.2 |
| Neophytadiene | 1842 | 0.6 | 0.8 | 0.9 |
| Total | 93.7 | 94.5 | 94.7 | |
| Monoterpene hydrocarbon | 40.2 | 45.1 | 46.8 | |
| Oxygenated monoterpene | 42.0 | 36.5 | 35.8 | |
| Sesquiterpene hydrocarbon | 5.6 | 6.8 | 6.1 | |
| Oxygenated sesquiterpene | 0.6 | 0.8 | 0.5 | |
| Diterpene hydrocarbon | 0.6 | 0.8 | 0.9 | |
| Benzenoids | 2.9 | 2.3 | 2.3 | |
| Oxylipins | 0.6 | 1.1 | 1.2 | |
| Others | 1.2 | 1.1 | 1.1 | |
| Number of compounds quantified | 25 | 24 | 24 |
| Treatments | Parameters | The concentration of essential oil inhibiting the tested microorganisms (µg/mL) | |||||||
| Gram (+) bacteria | Gram (-) bacteria | Yeast | |||||||
| Staphylococcus aureus | Bacillus subtilis | Lactobacillus fermentum | Salmonella enterica | Escherichia coli | Pseudomonas aeruginosa | Candida albicans | |||
| F1 | IC50 | 5134 ± 267 | >16,000 | >16,000 | 13,818 ± 610 | 6469 ± 320 | 6536 ± 341 | 4500 ± 201 | |
| MIC | 8000 | >16,000 | >16,000 | >16,000 | 16,000 | >16,000 | 8000 | ||
| F2 | IC50 | 5183 ± 278 | >16,000 | 12,000 ± 579 | >16,000 | 4774 ± 257 | 8453 ± 414 | 4758 ± 225 | |
| MIC | 8000 | >16,000 | >16,000 | >16,000 | 8000 | >16,000 | 8000 | ||
| F3 | IC50 | 9391 ± 476 | >16,000 | >16,000 | 16,000 ± 577 | 13,333 ± 569 | 7571 ± 377 | >16,000 | |
| MIC | >16,000 | >16,000 | >16,000 | >16,000 | >16,000 | >16,000 | >16,000 | ||
| Ampicillin | IC50 | 0.02 ± 0.005 | 3.62 ± 0.15 | 1.03 ± 0.07 | |||||
| MIC | 0.125 ± 0.0 | 32 ± 0.0 | 32 ± 0.0 | ||||||
| Cefotaxime | IC50 | 0.43 ± 0.05 | 0.007 ± 0.002 | 4.34 ± 0.15 | |||||
| MIC | 32 ± 0.0 | 0.5 ± 0.0 | 8 ± 0.0 | ||||||
| Nystatin | IC50 | 1.32 ± 0.05 | |||||||
| MIC | 8 ± 0.0 | ||||||||
| Nutrients | Components | Concentration (µM) |
|---|---|---|
| Macro-nutrients | Nitrogen | 298.95 |
| phosphorous | 62.00 | |
| Potassium | 155.95 | |
| Calcium | 17.00 | |
| Magnesium | 7.50 | |
| Sulfur | 13.55 | |
| Micro-nutrients | Iron | 0.50 |
| Manganese | 5.00 | |
| Zinc | 0.50 | |
| Boron | 0.50 | |
| Copper | 0.05 | |
| Molybdenum | 0.50 | |
| Cobalt | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



