Submitted:
01 April 2026
Posted:
03 April 2026
You are already at the latest version
Abstract
The ability to precisely edit genetic characteristics with a CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas (CRISPR-associated) immunity complex is a revolutionary advance in science. Originally discovered in bacteria as part of a natural defense mechanism against viruses, CRISPR/Cas provides a precise, efficient, and relatively simple method for editing genes in microbes, plants, animals, and humans. The process relies on the Cas protein, an enzyme that cleaves and unwinds DNA at targeted locations. This process is guided by RNA sequences complementary to the DNA or RNA sequence of interest, allowing for changes to the genome through innate non-homologous end joining (NHEJ) and homology-directed repair (HDR). The potential applications of CRISPR/Cas are immense and in agriculture, is facilitating crop development with resistance to abiotic, biotic, and agronomic characteristics that improve yield, quality, and food security. Gene editing also facilitates the relatively rapid modification of regulatory and complex pathways that enable studies to advance our understanding of gene function. This review provides an update of the fast-evolving CRISPR/Cas modification of important crops to address emerging global population, environmental and climate challenges.
Keywords:
immunity
; RNA
; gene editing
; crop improvement
Introduction
Traditional plant breeding and transgenic/cisgenic genetically modified plants/organisms (GMOs) capabilities to improve crop production have been restricted by relatively long timelines, lack of precision, and acceptance limitations. Precise genetic engineering methods have emerged, but application of zinc finger nucleases (ZFN) and Tal effector nucleases (TALEN) have not been widely applied because of the complexity [1,2]. A relatively simple method for site-specific gene editing was recently discovered in prokaryote adaptive immunity involving clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated (Cas) nuclease [3]. Remarkably, a non-coding single guide RNA (sgRNA) may target nucleic acid and direct a Cas endonuclease to cleave nucleic acid with specificity, and the genomic site is subsequently repaired by innate cellular repair mechanisms.
CRISPR was initially observed as an unusual, interrupted repeat of sequences in Escherichia coli [4]. Subsequently, the term CRISPR was introduced to describe the increasing reports describing the occurrence of similar sequences [5,6]. Several breakthroughs indicated spacers matched viral sequences indicating an adaptive immunity mechanism, identified the Cas9 sequence and adjacent Protospacer Adjacent Motif (PAM), and confirmed bacterial immunity against phage infection [7,8,9,10,11,12,13]. A key advance in our understanding of CRISPR was the observation that the clustered repeats were associated with CRISPR-associated (Cas) proteins with helicase and nuclease motifs [14]. Further characterization identified essential non-coding CRISPR RNA (crRNA) as the guiding sequence and trans-activating CRISPR (tracrRNA) that complexes with crRNA to secure Cas protein and target DNA for cleavage. In Streptococcus pyogenes the CRISPR Cas9 complex includes a trans-activating CRISPR RNA combined into a single guide RNA (sgRNA) and the crRNA may be synthesized to target key sequences for editing [3]. A simplified engineered two-part CRISPR-Cas9 in vitro system was developed to facilitate gene editing of genomes in human and animal cells that allows precise changes across a wide range of organisms.
Crop Applications
Initial plant CRISPR-Cas genome editing exploited DNA double-strand breaks (DSB) created by the Cas enzyme, guided by the single-stranded guide RNA (sgRNA), that cleaved the DNA at the specific crRNA target sequence 3 nucleotides 5’ of the PAM site [15,16,17]. A sgRNA specificity was conferred by the 5’ sequence and is usually 20 nucleotides in length N20NGG (Figure 1). Plant RNA polymerase III promoters such as U6 are required for a defined transcription start nucleotide of the sgRNA. Delivery of the sgRNA may involve protoplasts, callus, or axillary meristems through direct delivery with Cas sequence or transformation using Agrobacterium tumefaciens [18]. Repairs may be precise using homology-directed repair (HDR) or unpredictable by non-homologous end joining (NHEJ) producing mutations, insertions, or deletions. Introduction of missense, nonsense, or frameshift sequence usually produces knockout mutations or functional changes that can alter trait characteristics. In plants that are polyploid, the mutation may be epistatic and masked if recessive. An insertion is also possible through HDR by the addition of an epigenetic template for repair. Edits using a Cas9 substitute such as Cas12a (Cpf1), a cytosine base editor that does not require a double-strand DNA cleavage or Cas13 that recognizes single-stranded RNA (ssRNA), increases the flexibility and precision of the sequence modifications [19,20].
RNA Adaptive Immunity in Plants
Plant genetic research has been transformed by two remarkable molecular innate mechanisms, CRISPR gene editing and RNA silencing [3,21]. Both technologies exploit RNA activated immunity and allow for the modulation of gene expression, but operate via distinct mechanisms and have different applications in plant biology and biotechnology. Understanding their similarities and differences is essential for selecting the appropriate method for specific research or crop improvement objectives. The CRISPR-Cas enables precise and stable modifications of plant genomes using sgRNA to direct a Cas nuclease to a specific DNA sequence. The cleaved nucleic acid repair often introduces small insertions/deletions, allowing for gene modifications [3]. RNA silencing involves RNA interference (RNAi) via the RNA-induced silencing complex (RISC), a post-transcriptional gene regulation mechanism that generate small RNA molecules (sRNA), such as small interfering RNAs (siRNAs) or microRNAs (miRNAs). These siRNAs or miRNAs, respectively bind to virus double-stranded RNA (dsRNA) or complementary messenger RNA (mRNA) to mediate their degradation or inhibit translation [21]. This process does not alter the genomic DNA but rather downregulates replication and gene expression transiently or stably through transgenic constructs. Applications of antisense RNA have grown rapidly since the initial reports of capabilities to alter phenotype and obtain high levels of disease resistance [22,23]
Mechanisms of adaptive RNA immunity differ in that CRISPR-Cas gene editing is a DNA-level modification via nuclease activity and silencing is an RNA-level suppression via small RNA-mediated degradation by the RISC [21]. Whereas CRISPR-Cas are usually permanent genomic changes, RNAi may be transient or semi-stable depending on the delivery method. Precision of CRISPR-Cas and RNAi is high, but off-target mutations or unexpected expression downregulation may occur. Regulatory agency evaluation is similar for RNA immunity products, but consumer acceptance is often influenced by delivery method and novelty of the plant-trait combination. RNAi is often effective for transient applications of exogenously applied double-stranded RNA (dsRNA) to the targets with nanoparticle protection and optimized delivery [24,25]. While RNAi is ideal for temporary gene silencing and functional genomics studies, CRISPR is often applied in genome editing, gene therapy, and studying gene function [3]. CRISPR-Cas is particularly well-suited for creating knockouts, correcting alleles, and engineering cis-regulatory elements and offers precision breeding opportunities without necessarily introducing foreign DNA. In summary, RNAi and CRISPR are powerful adaptive immunity tools for gene manipulation, but mechanisms, targets, and applications differ significantly.
CRISPR-Cas Characterization
Agroinfiltration is considered a transient sequence delivery procedure that utilizes Agrobacterium tumefaciens strains, such as GV3101 or EHA105, to deliver foreign DNA into plant tissues [26]. Frequently used in gene expression studies, agroinfiltration has been widely adapted for functional genomics and genome editing research notably with CRSPR Cas-systems. A relatively simple, cost-effective, rapid, and efficient procedure has made agroinfiltration an invaluable method for characterization of CRISPR targets and constructs (Figure 2). Screening of vector and construct efficiency in generating gene edits may be determined by reporter genes such as luciferase, fluorescent proteins, or phytoene desaturase (PDS) [27,28,29]. Delivery into protoplasts or leaves and axillary meristems through agroinfiltration facilitates the relatively rapid determination and optimization of the expression vectors and regulatory elements. For example, expression of sgRNA and Cas may be elevated with the use of an enhancer such as the duplicated CaMV 35S promoter [30]. Combination of the enhancer and a promoter with a defined transcription start nucleotide to produce sgRNA can increase efficiency of gene editing (Figure 2). Observation of tissue culture or soil propagated plant editing occurs within weeks and allows for relatively fast widescale screening of sgRNA vectors and production of gene edited tissues.
CRISPR-Cas Delivery
Several procedures are available for stable plant genome knockout or gain of function editing (Figure 3). Protoplasts allow direct delivery of sgRNA and Cas but are relatively slow, requiring the regrowth of cell walls and use of phytohormones that can cause undesirable somaclonal off-target mutant phenotypes [31]. Production of callus from wounded tissues with A. tumefaciens delivery of sgRNA and Cas reduces exposure to phytohormones and plant production timelines. While axillary meristems produce gene edited adventitious shoots within weeks in culture-free propagation with minimal need of phytohormones and developmental regulators but often produces chimeric tissues [18,32].
Abiotic Stress Resilience: Single Genes to Network Modulation
Abiotic stresses, including drought, salinity, extreme temperatures, and oxidative stress, remain major constraints on crop productivity under changing climates and intensifying production pressures. Although traditional breeding has improved stress tolerance, progress is often slow because these traits are typically quantitative, polygenic, and strongly influenced by environmental conditions. Transgenic strategies have introduced beneficial alleles, but regulatory barriers and public concerns have limited widespread adoption. The emergence of CRISPR–Cas genome editing has shifted this paradigm by enabling precise modification of endogenous genes and regulatory elements, allowing targeted rewiring of stress-response networks without necessarily introducing foreign DNA. One of the most significant advances is the capacity to modulate hormone signaling pathways that integrate environmental sensing with growth regulation (Table 1). For example, editing ABA-related regulators such as AREB1 genes enhances drought responsiveness by fine-tuning stomatal closure and water-use efficiency [36], while modifications to OST2 alter guard cell dynamics [35]. Importantly, these examples illustrate a broader shift in engineering strategy; rather than constitutive overexpression of stress-response genes, that often imposes growth penalties, CRISPR enables adjustment of signaling sensitivity, thereby balancing stress tolerance with yield stability.
Applications targeting salinity and temperature tolerance further demonstrate how genome editing can refine key regulatory nodes in plant stress physiology. Editing ion transporters such as ZmHKT2 improves sodium exclusion or compartmentalization, enhancing ionic homeostasis under saline conditions [60], while disruption of negative regulators such as HyPRP1 in tomato increases survival under high salinity [56]. However, these interventions also highlight an important limitation; modification of ion transport or hormone signaling pathways can generate pleiotropic effects on development and nutrient balance. Consequently, emerging approaches emphasize multiplex editing and promoter engineering to modulate the spatial or temporal expression of target genes rather than relying solely on gene knockouts. A similar transition toward regulatory network engineering is evident in temperature stress responses. CRISPR-mediated manipulation of heat shock factors (HSFs), heat shock proteins (HSPs), and upstream regulators such as SlAGL6, ZmTMS5, and SlMAPK3 enhances thermotolerance by stabilizing proteins and reproductive tissues during heat stress [68,70,72], while editing targets including OsMYB, VInv, and proline-rich proteins improves cold tolerance through increased membrane stability and osmolyte accumulation [74,77,85]. Herbicide resistance was similarly obtained by editing the acetolactate synthase OsALS , that is involved in amino acid synthesis [95], and the polyamine uptake transporters OsPUT, which improved paraquat resistance without yield loss [97]. Collectively, these studies underscore that CRISPR’s greatest potential lies not simply in gene disruption but in reshaping transcriptional and metabolic pathways that coordinate stress adaptation. Because abiotic tolerance is rarely governed by single loci and often reflects quantitative trait loci (QTL) architecture, future progress will depend on multiplex editing of pathway components, integration with genomic selection, and the use of transcriptomic and epigenomic data to identify central regulatory hubs rather than peripheral stress markers.
Elevating Biotic Resilience: Immunity and Ecological Interactions
CRISPR-Cas genome editing, derived from a prokaryotic adaptive immune system, has emerged as a powerful platform for engineering plant responses to biotic stress. Its applications extend across resistance to pathogens, insect pests, and nematodes, as well as the modulation of beneficial plant–microbe interactions (Table 2). Although pesticides, crop rotation, and conventional breeding have historically mitigated biotic stress, these strategies often lack genetic precision, require lengthy breeding cycles, or raise environmental and health concerns. Moreover, resistance breeding is frequently constrained by available germplasm and linkage drag, limiting the speed and specificity with which desirable traits can be deployed. In contrast, CRISPR–Cas enables targeted editing of genes controlling immunity, defense signaling, and host–pathogen compatibility [3], thereby facilitating both incremental improvements and the creation of novel resistance phenotypes through precise modification of coding or regulatory sequences (Table 2). A prominent strategy involves disruption of plant susceptibility S genes, host factors exploited by pathogens during infection. For example, knockout of MLO alleles across multiple crops confers durable resistance to powdery mildew [101,102,103], while CRISPR-mediated mutation of OsSWEET genes in rice blocks pathogen-induced activation of sugar transporters by Xanthomonas oryzae, enhancing resistance to bacterial blight without detectable developmental penalties [115]. These cases demonstrate how loss-of-function edits can generate broad-spectrum disease resistance while minimizing the growth–defense trade-offs often associated with constitutive activation of immune pathways. Similarly, CRISPR has been used to engineer resistance to plant viruses and viroids either by directly targeting viral genomes or by modifying host factors required for viral replication [123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139]. Although these strategies demonstrate considerable versatility, their long-term durability remains influenced by viral mutation rates and the potential emergence of escape variants.
Beyond pathogens, CRISPR–Cas provides opportunities to strengthen endogenous defenses against herbivores and nematodes while also enabling the rational engineering of beneficial plant–microbe interactions. Unlike transgenic approaches that introduce exogenous toxins, genome editing can enhance native defense pathways, potentially improving regulatory acceptance and ecological compatibility. For instance, modification of cytokinin oxidase/dehydrogenase OsCKX genes in rice influences jasmonic acid–mediated insect defense signaling, increasing resistance to chewing and sucking pests [140], while knockout of GmGUT genes in soybean alters flavonoid biosynthesis and enhances resistance to chewing insects through relatively minor genomic changes [142]. Resistance to nematodes has likewise been achieved through editing of susceptibility genes such as HPP04 in rice and SNAP02 in soybean [143,144], reinforcing a broader conceptual framework in which CRISPR-mediated resistance often arises from removal of host compatibility factors or modulation of defense metabolism rather than introduction of novel resistance genes. Increasing attention is also being directed toward engineering beneficial symbioses with organisms such as arbuscular mycorrhizal fungi and nitrogen-fixing bacteria. Because these associations rely on finely regulated signaling networks, precise genome edits can be particularly advantageous. For example, modification of genes in the common symbiosis signaling pathway, including SYMRK, CCaMK, and CYCLOPS, may enhance mycorrhizal colonization efficiency [146], while manipulation of the flavone biosynthetic pathway in rice promoted bacterial biofilm formation and improved biological nitrogen fixation, resulting in increased seed yield [147]. Collectively, these studies reflect a shift from purely resistance-based paradigms toward optimization of plant holobiont function. As understanding of plant–pathogen–pest–microbe interactions expands, CRISPR–Cas will remain an essential tool for refining immune recognition, regulating defense networks, and improving crop resilience within next-generation crop improvement strategies [148,149,150].
Phenotype and Agronomic Performance Improvement
CRISPR-Cas9 genome editing has been successfully applied in a range of crop plants to enhance yield, improve quality traits, and boost agronomic performance (Table 3). These advances are increasingly important for meeting the demands of a growing global population while supporting food security and the development of more resilient and efficient agricultural systems. By enabling precise modification of genes controlling plant growth, metabolism, and environmental responses, CRISPR not only accelerates crop improvement but also expands the range of traits that can be modified beyond the constraints of traditional breeding. Yield improvement efforts have largely focused on genes regulating plant architecture, reproductive development, and resource allocation. For example, CRISPR-mediated editing of the rice grain number gene Gn1a, that controls cytokinin degradation, has generated loss-of-function alleles that increase cytokinin levels and enhance grain number and yield [165]. Similarly, editing of PYL genes involved in abscisic acid signaling has produced rice plants with optimized architecture and improved grain production [167]. In tomato, targeted modification of SlIAA, a regulator of parthenocarpy, and the polygalacturonase gene PG has resulted in seedless fruit and delayed softening, respectively, improving both productivity and postharvest shelf life [172,173]. These examples highlight how CRISPR enables direct manipulation of developmental pathways that were previously difficult to modify with precision.
Beyond yield, CRISPR-based potato varieties with modified granule-bound starch synthase StGBSS influences composition [175], while disruption of the inositol tetrakisphosphate kinase gene BnITPK in oilseed crops reduces phytic acid content, improving mineral bioavailability and nutritional quality [151]. In wheat, modification of TaGW2, a negative regulator of grain weight, increases grain size and mass, contributing to improved baking quality and market value [186]. Genome editing has also been used to improve crop performance under diverse environmental conditions. In soybean, editing of flowering-time genes GmFT2a and GmFT5a has enabled the development of varieties better adapted to different latitudes and seasonal conditions, supporting broader cultivation and improved yield stability [155]. Collectively, these studies illustrate CRISPR’s versatility in refining developmental, metabolic, and adaptive pathways to simultaneously enhance yield potential, nutritional quality, and environmental resilience. As plant genomic and transcriptomic resources continue to expand, the identification of new gene and regulatory targets will further increase the precision and scope of genome editing strategies, positioning CRISPR as a central tool for developing crop varieties capable of meeting evolving agronomic, industrial, and societal demands.
Future Applications of Gene Editing in Plants
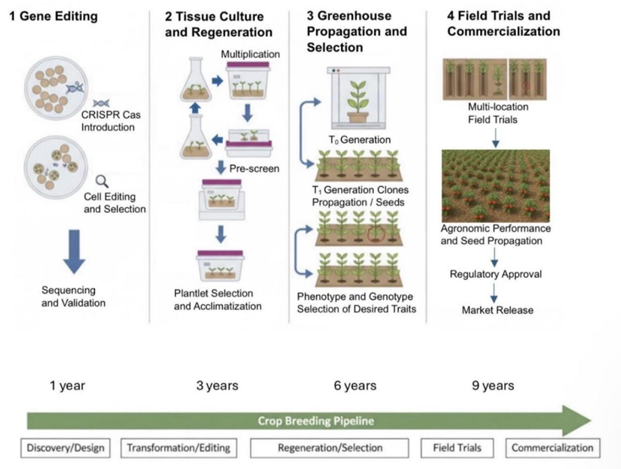
CRISPR–Cas systems have transformed plant genome engineering by enabling precise, multiplexed, and increasingly transgene-free modification of agronomic traits with substantially greater speed and predictability than conventional breeding or earlier nuclease platforms. Following their adaptation for genome editing, CRISPR tools were rapidly deployed in crops to generate targeted knockouts, allele replacements, and regulatory modifications affecting yield, disease resistance, and abiotic stress tolerance (Figure 4). Compared with previous genome repair and modification, CRISPR offers simpler design, lower cost, and scalable multiplexing, features particularly valuable for editing plants recalcitrant to tissue culture propagation, redundant gene families and polyploid genomes. As gene editing methodologies continue to evolve, their applications are expected to expand significantly in the future, especially when combined with other emerging technologies such as deep sequencing, epigenetics, and artificial intelligence [191,192,193,194,195,196,197,198,199,200,201,202,203]. Combining multiple sgRNA is providing an accelerated genetic editing strategy to accelerate crop improvement, especially when pathways are identified that confer value-added characteristics [204]. However, technical and biological constraints remain significant; PAM requirements restrict targetable loci, efficient homology-directed repair is rare in most somatic plant tissues, delivery and regeneration are genotype-dependent and often recalcitrant, and polyploidy complicates complete allele modification. Off-target activity and regulatory heterogeneity across jurisdictions further hinder application. Thus, while CRISPR–Cas platforms provide unparalleled precision and versatility for crop improvement, their agronomic impact ultimately depends on advances in delivery systems, tissue culture independence, and deeper understanding of plant DNA repair and genome complexity.
Emerging computational and technological innovation is further expanding the scope and precision of plant genome editing. This may be especially important for crops that lack a diverse genetic gene pool and are especially susceptible to attack by emerging pathogens and vulnerable to extreme growing conditions [205,206,207,208,209,210,211,212,213,214,215,216,217]. The AI-assisted analysis of large genomic datasets has started to improve guide RNA design and reduce off-target activity, a persistent challenge resulting from the inherent tolerance of CRISPR systems to guide–target mismatches [5,6,7,8,9,10,11,12,13,14]. Such unintended cleavage events can generate genomic rearrangements, including deletions, inversions, or translocations, that may activate stress-response pathways, highlighting the need for improved predictive algorithms and enhanced editor specificity. AI-driven modeling and structural prediction platforms, such as SWISS-Model and AlphaFold2, also facilitate the prediction of protein-level consequences of genomic edits, thereby accelerating the identification of functional targets [218,219]. At the technological level, advances in delivery systems, including viral vectors, nanoparticles, agroinfiltration, biolistic methods, and meristematic injection, are expanding the diversity of crops amenable to genome editing while allowing greater spatial or temporal control of CRISPR activity [24,220,221,222,223].
CRISPR platforms consistently outperform earlier genome-editing platforms in terms of design simplicity, multiplexing capacity, and editing efficiency, with Cas9 frequently achieving mutation rates of 50–80% in protoplasts and 10–70% in stable transformants [34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190]. As these technologies continue to evolve, they are enabling new strategies such as accelerated domestication of wild plant species, engineering of metabolic pathways for pharmaceutical or biofuel production, and optimization of crop traits through regulatory and epigenetic modification. The emergence of base and prime editing that combines Cas9 with a reverse transcriptase to program a prime editing guide RNA (pegRNA) that specifies the target site and encodes the desired edit, further extends precision by enabling predictable nucleotide substitutions without double-strand DNA breaks [224]. However, the deployment of genome-edited crops remains shaped by regulatory frameworks, intellectual property considerations, and public acceptance, that vary considerably among jurisdictions [19,20,225,226]. The commercialization of genome-edited tomatoes with elevated γ-aminobutyric acid (GABA) levels in Japan illustrates the growing transition of CRISPR technologies from experimental platforms to agricultural products [227], while also highlighting the complex legal and societal landscape that continues to influence their global adoption.
Conclusions
The future of gene editing in plants promises to revolutionize global agriculture and food systems. By enabling targeted modifications with high specificity and efficiency by endogenous repair mechanisms, this technology empowers researchers to address pressing issues with unprecedented speed and possibilities, from food security and population growth to environmental sustainability during climate and market needs. Continued interdisciplinary collaboration and responsible innovation exploiting emerging technologies and advances will be key to realizing the full potential of plant gene editing. Innate RNA adaptive immunity represents exciting tools for plant genetic research and biotechnology improving product development. CRISPR’s precision and permanence contrast with the flexibility and reversibility of RNAi-based approaches and complements existing breeding technologies. The choice of technology to address crop challenges should be decided by experimental objectives, regulatory context, and desired transient or stable outcome. For optimal results, integrated approaches leveraging various technologies will offer synergistic advantages in functional genomics and multidisciplinary crop improvement programs for industry and global populations.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This work was funded by Solanum International Inc. Project #45402 and Natural Sciences and Engineering Research Council of Canada (NSERC) Alliance Program Project #44842.
References
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Klug, A.; Rhodes, D. Zinc fingers: a novel protein fold for nucleic acid recognition. Cold Spring Harb. Symp. Quant. Biol. 1987, 52, 473–482. [Google Scholar] [CrossRef]
- Boch, J. TALEs of genome targeting. Nat. Biotechnol. 2011, 29, 135–136. [Google Scholar] [CrossRef]
- Mojica, F.J.; Montoliu, L. On the origin of CRISPR-Cas technology: from prokaryotes to mammals. Trends Microbiol. 2016, 24, 811–820. [Google Scholar] [CrossRef]
- Mojica, F.J.; Rodriguez-Valera, F. The discovery of CRISPR in archaea and bacteria. FEBS J. 2016, 283, 3162–3169. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Erdmann, S.; Mojica, F.J.; Garrett, R.A. Protospacer recognition motifs: mixed identities and functional diversity. RNA Biol. 2013, 10, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Mojica, F. J.; Díez-Villaseñor, C.; Soria, E.; Juez, G. Biological significance of a family of regularly spaced repeats in the genomes of archaea, bacteria and mitochondria. Mol. Microbiol. 2000, 36, 244–246. [Google Scholar] [CrossRef]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Ehrlich, S.D. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiol. 2005, 151, 2551–2561. [Google Scholar] [CrossRef]
- Mojica, F. J.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiol. 2005, 151, 653–663. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D. A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Brouns, S. J. J.; Jore, M. M.; Lundgren, M.; Westra, E. R.; Slijkhuis, R. J. H.; Snijders, A. P. L.; Dickman, M. J.; Makarova, K. S.; Koonin, E. V.; van der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef]
- Jansen, R.; van Embden, J.D.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, B.; Ding, W.; Liu, X.; Yang, D.-L.; Wei, P.; Cao, F.; Zhu, S.; Zhang, F.; Mao, Y.; Zhu, J.-K. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qui, J.L.; Gao, C. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheem, J. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; Severinov, K.; Regev, A.; Lander, E.S.; Koonin, E.V.; Zhang, F. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double- stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a chimeric chalcone synthase gene into Petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef]
- Kawchuk, L.M.; Martin, R.R.; McPherson, J. Sense and antisense RNA-mediated resistance to potato leafroll virus in Russet Burbank potato plants. Mol. Plant Microbe Interact. 1991, 4, 247–253. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.M.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef] [PubMed]
- Koeppe, S.; Kawchuk, L.; Kalischuk, M. RNA interference past and future applications in plants. Int. J. Mol. Sci. 2023, 24, 9755. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Ow, D.W.; Wood, K.V.; Deluca, M.; De Wet, J.R.; Helinski, D.R.; Howell, S.H. Transient and stable expression of the firefly luciferase gene in plant cells and transgenic plants. Science 1986, 234, 856–859. [Google Scholar] [CrossRef]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef]
- Qin, G.; Gu, H.; Ma, L.; Peng, Y.; Deng, X.W.; Chen, Z.; Qu, L.-J. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Res. 2007, 17, 471–482. [Google Scholar] [CrossRef]
- Kay, R.; Chan, A.; Daly, M.; McPherson, J. Duplication of CaMV 35S promoter sequences creates a strong enhancer for plant genes. Science 1987, 236, 1299–1302. [Google Scholar] [CrossRef]
- Kawchuk, L.M.; Lynch, D.R.; Martin, R.R.; Kozub, G.C.; Farries, B. Field resistance to the potato leafroll luteovirus in transgenic and somaclone potato plants reduces tuber disease symptoms. Can. J. Plant Path. 1997, 19, 260–266. [Google Scholar] [CrossRef]
- Kawchuk, L.M.; Nielsen, J. Improved bunt inoculation of wheat utilizing adventitious shoots. Can. J. Botany 1987, 65, 1284–1285. [Google Scholar] [CrossRef]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S. A modular cloning system for standardized assembly of multigene constructs. PLoS One 2011, 6, e16765. [Google Scholar] [CrossRef] [PubMed]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Nekrasov, V. Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013, 9, 39. [Google Scholar] [CrossRef]
- Osakabe, Y.; Watanabe, T.; Sugano, S.S.; Ueta, R.; Ishihara, R.; Shinozaki, K.; Osakabe, K. Optimization of CRISPR/Cas9 genome editing to modify abiotic stress responses in plants. Sci. Rep. 2016, 6, 26685. [Google Scholar] [CrossRef]
- Paixão, J.F.R.; Gillet, F.-X.; Ribeiro, T.P.; Bournaud, C.; Lourenço-Tessutti, I.T.; Noriega, D.D.; Melo, B.P.; de Almeida-Engler, J.; Grossi-de-Sa, M.F. Improved drought stress tolerance in Arabidopsis by CRISPR/dCas9 fusion with a Histone AcetylTransferase. Sci Rep. 2019, 9, 8080. [Google Scholar] [CrossRef]
- Du, Y.-T.; Zhao, M.-J.; Wang, C.-T.; Gao, Y.; Wang, Y.-X.; Liu, Y.-W.; Chen, M.; Chen, J.; Zhou, Y.-B.; Xu, Z.-S.; Ma, Y.-Z. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18, 320. [Google Scholar] [CrossRef]
- Ogata, T.; Ishizaki, T.; Fujita, M.; Fujita, Y. CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under non-stressed conditions in rice. PLoS ONE 2020, 15, e0234156. [Google Scholar] [CrossRef]
- Kim, M.-S.; Ko, S.-R.; Jung, Y.J.; Kang, K.-K.; Lee, Y.-J.; Cho, Y.-G. Knockout mutants of OsPUB7 generated using CRISPR/Cas9 revealed abiotic stress tolerance in rice. Int. J. Mol. Sci. 2023, 24, 5338. [Google Scholar] [CrossRef]
- Zhao, J.; Yao, B.; Peng, Z.; Yang, X.; Li, K.; Zhang, X.; Zhu, H.; Zhou, X.; Wang, M.; Jiang, L.; He, X.; Liang, Y.; Zhan, X.; Wang, X.; Dai, Y.; Yang, Y.; Yang, A.; Dong, M.; Shi, S.; Lu, M.; Du, Y. Splicing defect of StDRO2 intron 1 promotes potato root growth by disturbing auxin transport to adapt to drought stress. Hortic. Plant J. 2025, 11, 706–720. [Google Scholar] [CrossRef]
- Wang, X.; Guo, Y.; Wang, Y.; Peng, Y.; Zhang, H.; Zheng, J. ZmHDT103 negatively regulates drought stress tolerance in maize seedlings. Agronomy 2024, 14, 134. [Google Scholar] [CrossRef]
- Li, P.; Li, X.; Jiang, M. CRISPR/Cas9-mediated mutagenesis of WRKY3 and WRKY4 function decreases salt and Me-JA stress tolerance in Arabidopsis thaliana. Mol. Biol. Rep. 2021, 48, 5821–5832. [Google Scholar] [CrossRef]
- Kim, S.T.; Choi, M.; Bae, S.J.; Kim, J.S. The functional association of Acqos/Victr with salt stress resistance in Arabidopsis thaliana was confirmed by CRISPR-mediated mutagenesis. Int. J. Mol. Sci. 2021, 22, 11389. [Google Scholar] [CrossRef]
- Curtin, S.J.; Xiong, Y.; Michno, J.M.; Campbell, B.W.; Stec, A.O.; Cermak, T.; Starker, C.; Voytas, D.F.; Eamens, A.L.; Stupar, R.M. CRISPR/Cas9 and TALENs generate heritable mutations for genes involved in small RNA processing of Glycine max and Medicago truncatula. Plant Biotechnol. J. 2018, 16, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xun, H.; Wang, W.; Ding, X.; Tian, H.; Hussain, S.; Dong, Q.; Li, Y.; Cheng, Y.; Wang, C.; Lin, R.; Li, G.; Qian, X.; Pang, J.; Feng, X.; Dong, Y.; Liu, B.; Wang, S. Mutation of GmAITR genes by CRISPR/Cas9 genome editing results in enhanced salinity stress tolerance in soybean. Front. Plant Sci. 2021, 12, 779598. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Zhang, Q.; Sun, T.; Wang, P. Cloning of the soybean GmNHL1 gene and functional analysis under salt stress. Plants 2023, 12, 3869. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Ma, Y.; Yang, Z.; Pu, Y.; Liu, M.; Du, J.; Xu, Z.; Xu, Z.; Zhang, S.; Zhang, H.; Zhang, W.; Yu, D.; Kan, G. Knockdown of β-conglycinin α′ and α subunits alters seed protein composition and improves salt tolerance in soybean. Plant J. 2024, 120, 1488–1507. [Google Scholar] [CrossRef]
- Kloc, Y.; Dmochowska-Boguta, M.; Zebrowska-Rozanska, P.; Laczmanski, L.; Nadolska-Orczyk, A.; Orczyk, W. HvGSK1.1 controls salt tolerance and yield through the brassinosteroid signaling pathway in barley. Int. J. Mol. Sci. 2024, 25, 998. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.J.; Xiong, Y.; Michno, J.M.; Campbell, B.W.; Stec, A.O.; Čermák, T.; Starker, C.; Voytas, D.F.; Eamens, A.L.; Stupar, R.M. CRISPR/Cas9 and TALENs generate heritable mutations for genes involved in small RNA processing of Glycine max and Medicago truncatula. Plant Biotechnol. J. 2017, 16, 1125–1137. [Google Scholar] [CrossRef]
- Kumar, V.V.S.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.V.; Chinnusamy, V. CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiol. Mol. Biol. Plants 2020, 26, 1099–1110. [Google Scholar] [CrossRef]
- Duan, Y.B.; Li, J.; Qin, R.Y.; et al. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, Y.; Wang, F.; et al. Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene. Mol. Breed. 2019, 39, 47. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. OsNAC45 is involved in ABA response and salt tolerance in rice. Rice 2020, 13, 79. [Google Scholar] [CrossRef]
- Lan, T.; Zheng, Y.; Su, Z.; Yu, S.; Song, H.; Zheng, X.; Lin, G.; Wu, W. OsSPL10, an SBP-box gene, plays a dual role in salt tolerance and trichome formation in rice (Oryza sativa L.). G3: Genes Genomes Genet. 2019, 9, 4107–4114. [Google Scholar] [CrossRef]
- Ye, N.; Wang, Y.; Yu, H.; Qin, Z.; Zhang, J.; Duan, M.; Liu, L. Abscisic acid enhances trehalose content via OsTPP3 to improve salt tolerance in rice seedlings. Plants 2023, 12, 2665. [Google Scholar] [CrossRef]
- Tran, M.T.; Doan, D.T.H.; Kim, J.; Song, Y.J.; Sung, Y.W.; Das, S.; Kim, E.J.; Son, G.H.; Kim, S.H.; Vu, T.V.; Kim, J.Y. CRISPR/Cas9-based precise excision of SlHyPRP1 domain(s) to obtain salt stress-tolerant tomato. Plant Cell Rep. 2021, 40, 999–1011. [Google Scholar] [CrossRef]
- Xiang, Y.; Jimenez-Gomez, J.M. SlHAK20: a new player in plant salt tolerance. EMBO J. 2020, 39, e104997. [Google Scholar] [CrossRef] [PubMed]
- Makhotenko, A.V.; Khromov, A.V.; Snigir, E.A.; Makarova, S.S.; Makarov, V.V.; Suprunova, T.P.; Kalinina, N.O.; Taliansky, M.E. Functional analysis of coilin in virus resistance and stress tolerance of potato (Solanum tuberosum) using CRISPR-Cas9 editing. Dokl. Biochem. Biophys. 2019, 484, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Lin, J.; Liu, X.; Chu, W.; Li, J.; Gao, Y.; An, K.; Song, W.; Xin, M.; Yao, Y.; Peng, H.; Ni, Z.; Sun, Q.; Hu, Z. Histone acetyltransferase TaHAG1 acts as a crucial regulator to strengthen salt tolerance of hexaploid wheat. Plant Physiol. 2021, 186, 1951–1969. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liang, X.; Yin, P.; Zhang, M.; Jiang, C. A domestication-associated reduction in K⁺-preferring HKT transporter activity underlies maize shoot K⁺ accumulation and salt tolerance. New Phytol. 2018, 222, 301–317. [Google Scholar] [CrossRef]
- Li, R.; Liu, C.; Zhao, R.; Wang, L.; Chen, L.; Yu, W.; Zhang, S.; Sheng, J.; Shen, L. CRISPR/Cas9-mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 2019, 19, 38. [Google Scholar] [CrossRef]
- Bertier, L.D.; Ron, M.; Huo, H.; Bradford, K.J.; Britt, A.B.; Michelmore, R.W. High-resolution analysis of the efficiency, heritability, and editing outcomes of CRISPR/Cas9-induced modifications of NCED4 in lettuce (Lactuca sativa). G3 (Bethesda) 2018, 8, 1513–1521. [Google Scholar] [CrossRef]
- Nandy, S.; Pathak, B.; Zhao, S.; Srivastava, V. Heat-shock-inducible CRISPR/Cas9 system generates heritable mutations in rice. Plant Direct 2019, 3, e00145. [Google Scholar] [CrossRef]
- Malzahn, A.A.; Tang, X.; Lee, K.; Ren, Q.; Sretenovic, S.; Zhang, Y.; Chen, H.; Kang, M.; Bao, Y.; Zheng, X.; Deng, K.; Zhang, T.; Salcedo, V.; Wang, K.; Zhang, Y.; Qi, Y. Application of CRISPR-Cas12a temperature sensitivity for improved genome editing in rice, maize, and Arabidopsis. BMC Biol. 2019, 17, 9. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Pham, D.T.; Nguyen, N.H.; Do, P.T.; To, H.T.M. The germin-like protein gene OsGER4 is involved in heat stress response in rice root development. J. Biosci. 2023, 48, 1201. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. U.S.A. 2018, 115, 6058–6063. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; He, M.; Li, J.; Chen, L.; Huang, Z.; Zheng, S.; Zhu, L.; Ni, E.; Jiang, D.; Zhao, B.; Zhuang, C. Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediated TMS5 editing system. Sci. Rep. 2016, 6, 37395. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Yeshayahou, E.; Bolger, A.M.; Arazi, T.; Gupta, S.K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato facultative parthenocarpy results from SlAGAMOUS-LIKE 6 loss of function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef]
- Hu, Z.; Li, J.; Ding, S.; Cheng, F.; Li, X.; Jiang, Y.; Yu, J.; Foyer, C.H.; Shi, K. The protein kinase CPK28 phosphorylates ascorbate peroxidase and enhances thermotolerance in tomato. Plant Physiol. 2021, 186, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Wang, L.; Zhao, R.; Sheng, J.; Zhang, S.; Li, R.; Shen, L. Knockout of SlMAPK3 enhances tolerance to heat stress involving ROS homeostasis in tomato plants. BMC Plant Biol. 2019, 19, 354. [Google Scholar] [CrossRef]
- Wang, H.; Feng, M.; Jiang, Y.; Du, D.; Dong, C.; Zhang, Z.; Wang, W.; Liu, J.; Liu, X.; Li, S. Thermosensitive SUMOylation of TaHsfA1 defines a dynamic ON/OFF molecular switch for the heat stress response in wheat. Plant Cell 2023, 35, 3889–3910. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Si, X.; Tian, Y.; Chen, K.; Liu, J.; Chen, H.; Gao, C. Generation of thermosensitive male-sterile maize by targeted knockout of the ZmTMS5 gene. J. Genet. Genomics 2017, 44, 465–468. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z.; Ji, Y.; Wang, C.; Wang, S.; Shi, Y.; Le, J.; Zhang, M. The heat shock factor 20–HSF4–cellulose synthase A2 module regulates heat stress tolerance in maize. Plant Cell 2024, 36, 2652–2667. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, G.; Han, Y.; Usman, B.; Liu, F.; Qin, B.; Li, R. Knockout of OsPRP1, a gene encoding proline-rich protein, confers enhanced cold sensitivity in rice (Oryza sativa L.) at the seedling stage. 3 Biotech 2019, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiang, Z.; Li, J.; Wang, S.; Chen, Y.; Liu, Y.; Mao, D.; Luan, S.; Chen, L. bHLH57 confers chilling tolerance and grain yield improvement in rice. Plant Cell Environ. 2023, 46, 1402–1418. [Google Scholar] [CrossRef]
- Li, R.; Zhang, L.; Wang, L.; Chen, L.; Zhao, R.; Sheng, J.; Shen, L. Reduction of tomato-plant chilling tolerance by CRISPR-Cas9-mediated SlCBF1 mutagenesis. J. Agric. Food Chem. 2018, 66, 9042–9051. [Google Scholar] [CrossRef]
- Yasmeen, A.; Shakoor, S.; Azam, S.; Bakhsh, A.; Shahid, N.; Latif, A.; Shahid, A.A.; Husnain, T.; Rao, A.Q. CRISPR/Cas-mediated knockdown of vacuolar invertase gene expression lowers the cold-induced sweetening in potatoes. Planta 2022, 256, 107. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, S.; Zhao, S.; Chen, D.; Tian, H.; Li, J.; Zhang, L.; Li, S.; Liu, L.; Shi, C.; Yu, X.; Ren, Y.; Chen, F. Global crotonylatome and GWAS revealed a TaSRT1–TaPGK model regulating wheat cold tolerance through mediating pyruvate. Sci. Adv. 2023, 9, eadg1012. [Google Scholar] [CrossRef]
- Li, X.; Cai, Q.; Yu, T.; Li, S.; Li, S.; Li, Y.; Sun, Y.; Ren, H.; Zhang, J.; Zhao, Y.; Zhang, J.; Zuo, Y. ZmG6PDH1 in glucose-6-phosphate dehydrogenase family enhances cold stress tolerance in maize. Front. Plant Sci. 2023, 14, 1116237. [Google Scholar] [CrossRef]
- Baeg, G.J.; Kim, S.H.; Choi, D.M.; Tripathi, S. CRISPR/Cas9-mediated mutation of 5-oxoprolinase gene confers resistance to sulfonamide compounds in Arabidopsis. Plant Biotechnol. Rep. 2021, 15, 803–812. [Google Scholar] [CrossRef]
- Wang, F.Z.; Chen, M.X.; Yu, L.J.; Xie, L.J.; Yuan, L.B.; Qi, H.; Xiao, M.; Guo, W.; Chen, Z.; Yi, K.; Zhang, J.; Qiu, R.; Shu, W.; Xiao, S.; Chen, Q.F. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Mohamed, S.; Tanoi, K.; Kobayashi, N.I.; Takagi, K.; Vernet, A.; Guiderdoni, E.; Périn, C.; Sentenac, H.; Véry, A.A. Production of low-Cs⁺ rice plants by inactivation of the K⁺ transporter OsHAK1 with the CRISPR-Cas system. Plant J. 2017, 92, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ye, R.; Liang, Y.; Zhang, S.; Liu, X.; Sun, C.; Li, F.; Yi, J. Generation of low-cadmium rice germplasms via knockout of OsLCD using CRISPR/Cas9. J. Environ. Sci. 2023, 126, 138–152. [Google Scholar] [CrossRef]
- Lu, H.P.; Liu, S.M.; Xu, S.L.; Chen, W.Y.; Zhou, X.; Tan, Y.Y.; Huang, J.Z.; Shu, Q.Y. CRISPR-S: an active interference element for a rapid and inexpensive selection of genome-edited, transgene-free rice plants. Plant Biotechnol. J. 2017, 15, 1371–1373. [Google Scholar] [CrossRef]
- Ding, J.; Ji, C.; Yu, L.; Wang, C.; Ding, G.; Wang, S.; Shi, L.; Xu, F.; Cai, H. OsMYB84, a transcriptional regulator of OsCOPT2 and OsHMA5, modulates copper uptake and transport and yield production in rice. The Crop J. 2024, 12, 456–469. [Google Scholar] [CrossRef]
- Sun, S.-K.; Chen, Y.; Che, J.; Konishi, N.; Tang, Z.; Miller, A.J.; Ma, J.F.; Zhao, F.-J. Decreasing arsenic accumulation in rice by overexpressing OsNIP1;1 and OsNIP3;3 through disrupting arsenite radial transport in roots. New Phytol 2018, 219, 641–653. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; Pan, Y.; Hu, Y.; Peng, Y.; Fu, X.; Li, H.; Xia, S.; Zhao, B. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Chu, C.; Huang, R.; Liu, L.; Tang, G.; Xiao, J.; Yoo, H.; Yuan, M. The rice heavy-metal transporter OsNRAMP1 regulates disease resistance by modulating ROS homoeostasis. Plant Cell Environ. 2022, 45, 1109–1126. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Qu, M.; Zhu, Y.; Peng, C.; Wang, J.; Gao, D.; Chen, C. ZINC TRANSPORTER5 and ZINC TRANSPORTER9 function synergistically in zinc/cadmium uptake. Plant Physiol. 2020, 183, 1235–1249. [Google Scholar] [CrossRef]
- Mao, X.; Zheng, Y.; Xiao, K.; Wei, Y.; Zhu, Y.; Cai, Q.; Chen, L.; Xie, H.; Zhang, J. OsPRX2 contributes to stomatal closure and improves potassium deficiency tolerance in rice. Biochem. Biophys. Res. Commun. 2018, 495, 461–467. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Zhao, F.J.; Sun, C.; Jin, C.; Shi, Y.; Sun, Y.; Li, Y.; Yang, M.; Jing, X.; Luo, J.; Lian, X. OsATX1 interacts with heavy metal P1B-type ATPases and affects copper transport and distribution. Plant Physiol. 2018, 178, 329–344. [Google Scholar] [CrossRef]
- Ibrahim, S.; Saleem, B.; Rehman, N.; Zafar, S.A.; Naeem, M.K.; Khan, M.R. CRISPR/Cas9-mediated disruption of Inositol Pentakisphosphate 2-Kinase 1 (TaIPK1) reduces phytic acid and improves iron and zinc accumulation in wheat grains. J. Adv. Res. 2022, 37, 33–41. [Google Scholar] [CrossRef]
- Wu, J.; Chen, C.; Xian, G.; Liu, D.; Lin, L.; Yin, S.; Sun, Q.; Fang, Y.; Zhang, H.; Wang, Y. Engineering herbicide-resistant oilseed rape by CRISPR/Cas9-mediated cytosine base-editing. Plant Biotechnol. J. 2020, 18, 1857–1859. [Google Scholar] [CrossRef]
- Hummel, A.W.; Chauhan, R.D.; Cermak, T.; Mutka, A.M.; Vijayaraghavan, A.; Boyher, A.; Starker, C.G.; Bart, R.; Voytas, D.F.; Taylor, N.J. Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant Biotechnol. J. 2018, 16, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, X.; Wu, C.; He, Y.; Ma, Y.; Hou, H.; Guo, X.; Du, W.; Zhao, Y.; Xia, L. Engineering herbicide-resistant rice plants through CRISPR/Cas9-mediated homologous recombination of Acetolactate Synthase. Mol. Plant 2016, 9, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.; Rao, G.S.; Sedeek, K.; Aman, R.; Kamel, R.; Mahfouz, M. Engineering herbicide resistance via prime editing in rice. Plant Biotechnol. J. 2020, 18, 2370–2372. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.S.; Cao, L.M.; Huang, W.Q.; Liu, J.X.; Lu, H.P. Disruption of three polyamine uptake transporter genes in rice by CRISPR/Cas9 gene editing confers tolerance to the herbicide paraquat. aBIOTECH 2022, 3, 140–145. [Google Scholar] [CrossRef]
- Liu, H.; Hu, M.; Wang, Q.; Cheng, L.; Zhang, Z. Role of papain-like cysteine proteases in plant development. Front. Plant Sci. 2018, 9, 1717. [Google Scholar] [CrossRef]
- Hu, S.; Chen, Y.; Qian, C.; Ren, H.; Liang, X.; Tao, W.; Chen, Y.; Wang, J.; Dong, Y.; Han, J.; Ouyang, X.; Huang, X. Nuclear accumulation of rice UV-B photoreceptors is UV-B- and OsCOP1-independent for UV-B responses. Nat. Commun. 2024, 15, 6396. [Google Scholar] [CrossRef]
- Liu, C.; Lin, J.-Z.; Wang, Y.; Tian, Y.; Zheng, H.-P.; Zhou, Z.-K.; Zhou, Y.-B.; Tang, X.-D.; Zhao, X.-H.; Wu, T.; Xu, S.L.; Tang, D.-Y.; Zuo, Z.C.; He, H.; Bai, L.-Y.; Yang, Y.-Z.; Liu, X.-M. The protein phosphatase PC1 dephosphorylates and deactivates CatC to negatively regulate H2O2 homeostasis and salt tolerance in rice. The Plant Cell 2023, 35, 3604–3625. [Google Scholar] [CrossRef] [PubMed]
- Shnaider, Y.; Elad, Y.; Rav-David, D.; Pashkovsky, E.; Leibman, D.; Kravchik, M.; Shtarkman-Cohen, M.; Gal-On, A.; Spiegelman, Z. Development of powdery mildew resistance in cucumber using CRISPR/Cas9-mediated mutagenesis of CsaMLO8. Phytopathology 2023, 113, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Naing, A.H.; Kang, H.; Lee, S.Y.; Li, W.; Chung, M.Y.; Kim, C.K. CRISPR/Cas9-mediated editing of PhMLO1 confers powdery mildew resistance in petunia. Plant Biotechnol. Rep. 2023, 17, 767–775. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Wan, D.Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hortic. Res. 2020, 7, 116. [Google Scholar] [CrossRef]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.M.A.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Hong, Y.; Meng, J.; He, X.; Zhang, Y.; Liu, Y.; Zhang, C.; Qi, H.; Luan, Y. Editing miR482b and miR482c simultaneously by CRISPR/Cas9 enhanced tomato resistance to Phytophthora infestans. Phytopathology 2021, 111, 1008–1016. [Google Scholar] [CrossRef]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Petersen, B.L.; Andreasson, E. Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci. Rep. 2021, 11, 4487. [Google Scholar] [CrossRef]
- Hegde, N.; Joshi, S.; Soni, N.; Kushalappa, A.C. The caffeoyl-CoA O-methyltransferase gene SNP replacement in Russet Burbank potato variety enhances late blight resistance through cell wall reinforcement. Plant Cell Rep. 2021, 40, 237–254. [Google Scholar] [CrossRef]
- Lin, X.; Jia, Y.; Heal, R.; Prokchorchik, M.; Sindalovskaya, M.; Olave-Achury, A.; Makechemu, M.; Fairhead, S.; Noureen, A.; Heo, J.; Witek, K.; Smoker, M.; Taylor, J.; Shrestha, R.K.; Lee, Y.; Zhang, C.; Park, S.J.; Sohn, K.H.; Huang, S.; Jones, J.D.G. Solanum americanum genome-assisted discovery of immune receptors that detect potato late blight pathogen effectors. Nat. Genet. 2023, 55, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.J.; van den Abeele, C.; Ortega-Salazar, I.; Papin, V.; Adaskaveg, J.A.; Wang, D.; Casteel, C.L.; Seymour, G.B.; Blanco-Ulate, B. Host susceptibility factors render ripe tomato fruit vulnerable to fungal disease despite active immune responses. J. Exp. Bot. 2021, 72, 2696–2709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Blahut-Beatty, L.; Zheng, S.; Clough, S.J.; Simmonds, D.H. The role of a soybean 14-3-3 gene (Glyma05g29080) on white mold resistance and nodulation investigations using CRISPR-Cas9 editing and RNA silencing. Mol. Plant Microbe Interact. 2023, 36, 159–164. [Google Scholar] [CrossRef]
- He, F.; Wang, C.; Sun, H.; Tian, S.; Zhao, G.; Liu, C.; Wan, C.; Guo, J.; Huang, X.; Zhan, G.; et al. Simultaneous Editing of Three Homoeologues of TaCIPK14 Confers Broad-spectrum Resistance to Stripe Rust in Wheat. Plant Biotechnol. J. 2022, 21, 354–368. [Google Scholar] [CrossRef]
- Dai, Z.; Yang, Q.; Chen, D.; Li, B.; Que, J.; Hu, L.; Zhang, B.; Zhang, Z.; Chen, K.; Zhang, S.; et al. ZmAGO18b negatively regulates maize resistance against southern leaf blight. Theor. Appl. Genet. 2023, 136, 158. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, G.; Usman, B.; Peng, H.; Zhao, N.; Yuan, R.; Liu, Y.; Li, R. Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-based proteomic analysis of mutants revealed new insights into M. oryzae resistance in elite rice line. Genes 2020, 11, 735. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.-S.; Li, C.; Nguyen, H.; Liu, B.; Auguy, F.; Sciallano, C.; Luu, V.T.; Dossa, G.S.; Cunnac, S.; Schmidt, S.M.; Slamet-Loedin, I.H.; Cruz, C.V.; Szurek, B.; Frommer, W.B.; White, F.F.; Yang, B. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef]
- Kim, M.-S.; Le, V.T.; Jung, Y.J.; Kang, K.-K.; Cho, Y.-G. OsPUB9 gene edited by CRISPR/Cas9 enhanced resistance to bacterial leaf blight in rice (Oryza sativa L.). Int. J. Mol. Sci. 2024, 25, 7145. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Ortigosa, A.; Gimenez-Ibanez, S.; Leonhardt, N.; Solano, R. Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant Biotechnol. J. 2019, 17, 665–673. [Google Scholar] [CrossRef]
- Ortega, A.; Seong, K.; Schultink, A.; de Toledo Thomazella, D.P.; Seo, E.; Zhang, E.; Pham, J.; Cho, M.-J.; Dahlbeck, D.; Warren, J.; Minsavage, G.V.; Jones, J.B.; Sierra-Orozco, E.; Hutton, S.F.; Staskawicz, B. CRISPR/Cas9-Mediated Editing of Bs5 and Bs5L in Tomato Leads to Resistance against Xanthomonas. Plant Biotech. J. 2024, 22, 2785–2787. [Google Scholar] [CrossRef]
- Yang, J.; Fang, Y.; Wu, H.; Zhao, N.; Guo, X.; Mackon, E.; Peng, H.; Huang, S.; He, Y.; Qin, B.; Liu, Y.; Liu, F.; Chen, S.; Li, R. Improvement of resistance to rice blast and bacterial leaf streak by CRISPR/Cas9-mediated mutagenesis of Pi21 and OsSULTR3;6 in rice (Oryza sativa L.). Front. Plant Sci. 2023, 14, 1209384. [Google Scholar] [CrossRef]
- Saikia, B.; S, R.; Debbarma, J.; Maharana, J.; Sastry, G.N.; Chikkaputtaiah, C. CRISPR/Cas9-based genome editing and functional analysis of SlHyPRP1 and SlDEA1 genes of Solanum lycopersicum L. in imparting genetic tolerance to multiple stress factors. Front. Plant Sci. 2024, 15. [Google Scholar] [CrossRef]
- Zhao, A.; Li, Q.; Meng, P.; Liu, P.; Wu, S.; Lang, Z.; Song, Y.; Macho, A.P. Reduced content of gamma-aminobutyric acid enhances resistance to bacterial wilt disease in tomato. Plant Biotech. J. 2025, 23, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. conferring resistance to geminiviruses with the CRISPR-Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like immune system conferring dna virus resistance in plants. Nature Plants 2015, 1, 15144. [Google Scholar] [CrossRef]
- Ali, Z.; Abulfaraj, A.; Idris, A.; Ali, S.; Tashkandi, M.; Mahfouz, M.M. CRISPR/Cas9-mediated viral interference in plants. Genome Biol. 2015, 16, 238. [Google Scholar] [CrossRef]
- Tashkandi, M.; Ali, Z.; Aljedaani, F.; Shami, A.; Mahfouz, M.M. Engineering resistance against tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato. Plant Signal Behav. 2018, 13, e1525996. [Google Scholar] [CrossRef]
- Yin, K.; Han, T.; Xie, K.; Zhao, J.; Song, J.; Liu, Y. Engineer complete resistance to cotton leaf curl multan virus by the CRISPR/Cas9 system in Nicotiana benthamiana. Phytopath. Res. 2019, 1, 9. [Google Scholar] [CrossRef]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating highly efficient resistance against wheat dwarf virus in barley by employing CRISPR/Cas9 system. Plant Biotech. J. 2019, 17, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Mahmood, M.S.; Rahman, S.U.; Rizvi, F.; Ahmad, A. Evaluation of the CRISPR/Cas9 system for the development of resistance against cotton leaf curl virus in model plants. Plant Protect. Sci. 2020, 56, 154–162. [Google Scholar] [CrossRef]
- Liu, H.; Soyars, C.L.; Li, J.; Fei, Q.; He, G.; Peterson, B.A.; Meyers, B.C.; Nimchuk, Z.L.; Wang, X. CRISPR/Cas9-mediated resistance to cauliflower mosaic virus. Plant Direct 2018, 2, e00047. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Commun. Biol. 2019, 2, 46. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Zhai, Y.; Ortiz, J.; Neff, M.; Mandal, B.; Mukherjee, S.K.; Pappu, H.R. multiplexed editing of a begomovirus genome restricts escape mutant formation and disease development. PLoS One 2019, 14, e0223765. [Google Scholar] [CrossRef]
- Pramanik, D.; Shelake, R.M.; Park, J.; Kim, M.J.; Hwang, I.; Park, Y.; Kim, J.-Y. CRISPR/Cas9-mediated generation of pathogen-resistant tomato against tomato yellow leaf curl virus and powdery mildew. Int. J. Mol. Sci. 2021, 22, 1878. [Google Scholar] [CrossRef]
- Zhang, Z.; Ge, X.; Luo, X.; Wang, P.; Fan, Q.; Hu, G.; Xiao, J.; Li, F.; Wu, J. Simultaneous editing of two copies of Gh14-3-3d confers enhanced transgene-clean plant defense against Verticillium dahliae in allotetraploid upland cotton. Front. Plant Sci. 2018, 9, 842. [Google Scholar] [CrossRef]
- Zhan, X.; Zhang, F.; Zhong, Z.; Chen, R.; Wang, Y.; Chang, L.; Bock, R.; Nie, B.; Zhang, J. Generation of virus-resistant potato plants by RNA genome targeting. Plant Biotechnol. J. 2019, 17, 1814–1822. [Google Scholar] [CrossRef]
- Mehta, D.; Stürchler, A.; Anjanappa, R.B.; Zaidi, S.S.-A.; Hirsch-Hoffmann, M.; Gruissem, W.; Vanderschuren, H. Linking CRISPR-cas9 interference in cassava to the evolution of editing-resistant Geminiviruses. Genome Biol. 2019, 20, 80. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, X.; Xu, H.; Zhan, B.; Zhou, C.; Li, S.; Zhang, Z. Knockout of SlDCL2b attenuates the resistance of tomato to potato spindle tuber viroid infection. Mol. Plant Pathol. 2024, 25, e13441. [Google Scholar] [CrossRef]
- Liu, C.; Kong, M.; Yang, F.; Zhu, J.; Qi, X.; Weng, J.; Di, D.; Xie, C. Targeted generation of null mutants in ZmGDIα confers resistance against maize rough dwarf disease without agronomic penalty. Plant Biotechnol. J. 2022, 20, 803–805. [Google Scholar] [CrossRef]
- Arra, Y.; Auguy, F.; Stiebner, M.; Chéron, S.; Wudick, M.M.; Miras, M.; Schepler-Luu, V.; Köhler, S.; Cunnac, S.; Frommer, W.B.; Albar, L. Rice yellow mottle virus resistance by genome editing of the Oryza sativa L. ssp. japonica nucleoporin gene OsCPR5.1 but not OsCPR5.2. Plant Biotechnol. J. 2024, 22, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, D.; Gao, D.; Zhao, W.; Du, H.; Qiu, Z.; Huang, J.; Wen, P.; Wang, Y.; Li, Q.; et al. Cytokinin confers brown planthopper resistance by elevating jasmonic acid pathway in rice. Int. J. Mol. Sci. 2022, 23, 5946. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Alariqi, M.; Wang, Y.; Wang, Q.; Xu, Z.; Zafar, M.N.; Yang, G.; Jia, R.; Hussain, A.; Chen, Y.; et al. Construction of host plant insect-resistance mutant library by high-throughput CRISPR/Cas9 system and identification of a broad-spectrum insect resistance gene. Advanced Science 2024, 11, 2306157. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Chen, L.; Shen, X.; Yang, H.; Fang, Y.; Ouyang, W.; Mai, S.; Chen, H.; Chen, S.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmUGT enhanced soybean resistance against leaf-chewing insects through flavonoids biosynthesis. Front. Plant Sci. 2022, 13, 802716. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Lin, B.; Cao, Y.; Zhang, Y.; Song, H.; Huang, C.; Sun, T.; Long, C.; Liao, J.; Zhuo, K. CRISPR/Cas9-mediated mutagenesis of the susceptibility gene OsHPP04 in rice confers enhanced resistance to rice root-knot nematode. Front. Plant Sci. 2023, 14, 1134653. [Google Scholar] [CrossRef]
- Usovsky, M.; Gamage, V.A.; Meinhardt, C.G.; Dietz, N.; Triller, M.; Basnet, P.; Gillman, J.D.; Bilyeu, K.D.; Song, Q.; Dhital, B.; et al. Loss-of-function of an α-SNAP gene confers resistance to soybean cyst nematode. Nat. Commun. 2023, 14, 7629. [Google Scholar] [CrossRef]
- Hao, J.; Yang, Y.; Futrell, S.; Kelly, E.A.; Lorts, C.M.; Nebie, B.; Runo, S.; Yang, J.; Alvarez, S.; Lasky, J.R.; et al. CRISPR/Cas9-mediated mutagenesis of carotenoid cleavage dioxygenase (CCD) genes in sorghum alters strigolactone biosynthesis and plant biotic interactions. Phytobiomes J. 2023, 7, 339–351. [Google Scholar] [CrossRef]
- Vernié, T.; Rich, M.; Pellen, T.; Teyssier, E.; Garrigues, V.; Chauderon, L.; Medioni, L.; van Beveren, F.; Libourel, C.; Keller, J.; et al. Conservation of symbiotic signaling since the most recent common ancestor of land plants. Proc. Natl. Acad. Sci. U.S.A. 2025, 122, e2408539121. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Tajima, H.; Cline, L.C.; Fong, R.Y.; Ottaviani, J.I.; Shapiro, H.Y.; Blumwald, E. Genetic modification of flavone biosynthesis in rice enhances biofilm formation of soil diazotrophic bacteria and biological nitrogen fixation. Plant Biotechnol. J. 2022, 20, 2135–2148. [Google Scholar] [CrossRef]
- Kourelis, J.; Marchal, C.; Posbeyikian, A.; Harant, A.; Kamoun, S. NLR immune receptor–nanobody fusions confer plant disease resistance. Science 2023, 379, 934–939. [Google Scholar] [CrossRef]
- Kalischuk, M.; Müller, B.; Fusaro, A.F.; Wijekoon, C.P.; Waterhouse, P.M.; Prüfer, D.; Kawchuk, L. Amplification of cell signaling and disease resistance by an immunity receptor Ve1Ve2 heterocomplex in plants. Commun. Biol. 2022, 5, 497. [Google Scholar] [CrossRef] [PubMed]
- Guayazán-Palacios, N.; Steinbrenner, A.D. Plant cell surface receptors at the forefront of the growth-defense trade-off. Dev. Cell. 2025, 491–492. [Google Scholar] [CrossRef]
- Sashidhar, N.; Harloff, H.J.; Potgieter, L.; Jung, C. Gene editing of three BnITPK genes in tetraploid oilseed rape leads to significant reduction of phytic acid in seeds. Plant Biotechnol. J. 2020, 18, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kobayashi, A.; Endo, M.; Sage-Ono, K.; Toki, S.; Ono, M. CRISPR/Cas9-mediated mutagenesis of the dihydroflavonol-4-reductase-B (DFR-B) locus in the Japanese morning glory ipomoea (Pharbitis) Nil. Sci. Rep. 2017, 7, 10028. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, L.; Fu, Q.; Xu, Z.F. Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas. Peer J. 2018, 6, e4812. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, L.; Gao, Q.; Zhang, J.; Zong, M.; Zhang, H.; Ren, Y.; Guo, S.; Gong, G.; Liu, F.; et al. Efficient CRISPR/Cas9-based gene knockout in watermelon. Plant Cell Rep. 2017, 36, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, L.; Chen, L.; Wu, T.; Liu, L.; Sun, S.; Wu, C.; Yao, W.; Jiang, B.; Yuan, S.; et al. Mutagenesis of GmFT2a and GmFT5a mediated by CRISPR/Cas9 contributes for expanding the regional adaptability of soybean. Plant Biotechnol. J. 2019, 18, 298–309. [Google Scholar] [CrossRef]
- Kis, A.; Polgári, D.; Dalmadi, Á.; Ahmad, I.; Rakszegi, M.; Sági, L.; Csorba, T.; Havelda, Z. Targeted Mutations in the GW2.1 Gene modulate grain traits and induce yield loss in barley. Plant Science 2024, 340, 111968. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, J.; Liu, B.; Cao, D.; Zong, Y.; Chang, Y.; Li, Y. Novel hina alleles created by genome editing increase grain hardness and reduce grain width in barley. Transgenic Res. 2022, 31, 637–645. [Google Scholar] [CrossRef]
- Zeng, Z.; Han, N.; Liu, C.; Buerte, B.; Zhou, C.; Chen, J.; Wang, M.; Zhang, Y.; Tang, Y.; Zhu, M.; et al. Functional dissection of HGGT and HPT in barley vitamin E biosynthesis via CRISPR/Cas9-enabled genome editing. Ann. Bot. 2020, 126, 929–942. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Lu, X.; Li, R.; Liu, J.; Fu, S.P.; Hu, X.; Jianchun, G.; Yao, Y. Construction and verification of CRISPR/Cas9 gene editing vector for cassava MeSSIII gene. Mol. Plant Breed. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Juma, B.S.; Mukami, A.; Mweu, C.; Ngugi, M.P.; Mbinda, W. Targeted mutagenesis of the CYP79D1 gene via CRISPR/Cas9-mediated genome editing results in lower levels of cyanide in cassava. Front. Plant Sci. 2022, 13, 1009860. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Hu, H.; Kai, J.; Traw, M.B.; Yang, S.; Zhang, X. Different knockout genotypes of OsIAA23 in rice using CRISPR/Cas9 generating different phenotypes. Plant Mol. Biol. 2019, 100, 467–479. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, S.; Liang, Y.; Zhang, R.; Liu, L.; Qin, P.; Zhang, Z.; Wang, Y.; Zhou, J.; Tang, X.; et al. Loss-function mutants of OsCKX gene family based on CRISPR-Cas systems revealed their diversified roles in rice. The Plant Genome 2023, 16, e20283. [Google Scholar] [CrossRef]
- Iwamoto, M. In-frame editing of transcription factor gene RDD1 to suppress miR166 recognition influences nutrient uptake, photosynthesis, and grain quality in rice. Sc.i Rep. 2022, 12, 10795. [Google Scholar] [CrossRef]
- Liu, K.; Sakuraba, Y.; Ohtsuki, N.; Yang, M.; Ueda, Y.; Yanagisawa, S. CRISPR/Cas9-mediated elimination of OsHHO3, a transcriptional repressor of three AMMONIUM TRANSPORTER1 genes, improves nitrogen use efficiency in rice. Plant Biotechnol. J. 2023, 21, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Zhou, Z.; Wu, P.; Fang, M.; Pan, X.; Lin, Q.; Luo, W.; Wu, G.; Li, H. Reassessment of the four yield-related genes Gn1a, DEP1, GS3, and IPA1 in rice using a CRISPR/Cas9 System. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, S.; Jaganathan, D.; Ramanathan, V.; Rahman, H.; Palaniswamy, R.; Kambale, R.; Muthurajan, R. Creation of novel alleles of fragrance gene OsBADH2 in rice through CRISPR/Cas9 mediated gene editing. PLOS ONE 2020, 15, e0237018. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. U.S.A. 2018, 115, 6058–6063. [Google Scholar] [CrossRef]
- Hu, X.; Cui, Y.; Dong, G.; Feng, A.; Wang, D.; Zhao, C.; Zhang, Y.; Hu, J.; Zeng, D.; Guo, L.; et al. Using CRISPR-Cas9 to generate semi-dwarf rice lines in elite landraces. Sci. Rep. 2019, 9, 19096. [Google Scholar] [CrossRef]
- Zhu, Y.; Lin, Y.; Chen, S.; Liu, H.; Chen, Z.; Fan, M.; Hu, T.; Mei, F.; Chen, J.; Chen, L.; Wang, F. CRISPR/Cas9-mediated functional recovery of the recessive rc allele to develop red rice. Plant Biotechnol. J. 2019, 17, 2096–2105. [Google Scholar] [CrossRef]
- Xu, J.; Kang, B.-C.; Naing, A.H.; Bae, S.-J.; Kim, J.-S.; Kim, H.; Kim, C.K. CRISPR/Cas9-mediated editing of 1-aminocyclopropane-1-carboxylate oxidase1 enhances Petunia flower longevity. Plant Biotechnol. J. 2019, 18, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, X.; Yang, C.; Li, M.; Guo, Y. Exploiting the CRISPR/Cas9 system for targeted genome mutagenesis in Petunia. Sci. Rep. 2016, 6, 20315. [Google Scholar] [CrossRef]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez Fons, L.; et al. Characterization of CRISPR mutants targeting genes modulating pectin degradation in ripening tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar] [PubMed]
- Ueta, R.; Abe, C.; Watanabe, T.; Sugano, S.S.; Ishihara, R.; Ezura, H.; Osakabe, Y.; Osakabe, K. Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Sci. Rep. 2017, 7, 507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zha, M.; Huang, J.; Li, L.; Imran, M.; Zhang, C. StMYB44 negatively regulates phosphate transport by suppressing expression of PHOSPHATE1 in potato. J. Exp. Bot. 2017, 68, 1265–1281. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.-S.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiol. Plantarum 2018, 164, 378–384. [Google Scholar] [CrossRef]
- Sevestre, F.; Facon, M.; Wattebled, F.; Szydlowski, N. Facilitating gene editing in potato: A single-nucleotide polymorphism (SNP) map of the Solanum tuberosum L. Cv. Desiree genome. Sci. Rep. 2020, 10, 2045. [Google Scholar] [CrossRef]
- Tuncel, A.; Corbin, K.R.; Ahn-Jarvis, J.; Harris, S.; Hawkins, E.; Smedley, M.A.; Harwood, W.; Warren, F.J.; Patron, N.J.; Smith, A.M. Cas9-mediated mutagenesis of potato starch-branching enzymes generates a range of tuber starch phenotypes. Plant Biotech. J. 2019, 17, 2259–2271. [Google Scholar] [CrossRef]
- Khromov, A.V.; Gushchin, V.A.; Timerbaev, V.I.; Kalinina, N.O.; Taliansky, M.E.; Makarov, V.V. Guide RNA design for CRISPR/Cas9-mediated potato genome editing. Dokl. Biochem. Biophys. 2018, 479, 90–94. [Google Scholar] [CrossRef]
- Nakayasu, M.; Umemoto, N.; Ohyama, K.; Fujimoto, Y.; Lee, H.J.; Watanabe, B.; Muranaka, T.; Saito, K.; Sugimoto, Y.; Mizutani, M. A dioxygenase catalyzes steroid 16α-hydroxylation in steroidal glycoalkaloid biosynthesis. Plant Physiol. 2017, 175, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Ye, G.; Zhou, Y.; Pu, X.; Su, W.; Wang, J. Editing sterol side chain reductase 2 gene (StSSR2) via CRISPR/cas9 reduces the total steroidal glycoalkaloids in potato. All Life 2021, 14, 401–413. [Google Scholar] [CrossRef]
- González, M.N.; Massa, G.A.; Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.-S.; Storani, L.; Décima Oneto, C.A.; Hofvander, P.; Feingold, S.E. Reduced enzymatic browning in potato tubers by specific editing of a polyphenol oxidase gene via ribonucleoprotein complexes delivery of the CRISPR/Cas9 system. Front. Plant Sci. 2019, 10, 1649. [Google Scholar] [CrossRef]
- Pfotenhauer, A.C.; Occhialini, A.; Harbison, S.A.; Li, L.; Piatek, A.A.; Luckett, C.R.; Yang, Y.; Stewart, C.N.; Lenaghan, S.C. Genome-editing of FtsZ1 for alteration of starch granule size in potato tubers. Plants 2023, 12, 1878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, H.; Li, S.; Li, J.; Yan, L.; Xia, L. Increasing yield potential through manipulating of an ARE1 ortholog related to nitrogen use efficiency in wheat by CRISPR/Cas9. J. Integr. Plant Biol. 2021, 63, 1649–1663. [Google Scholar] [CrossRef]
- Abdul Rahim, A.; Uzair, M.; Rehman, N.; Fiaz, S.; Attia, K.A.; Abushady, A.M.; Hwan Yang, S.; Ramzan Khan, M. CRISPR/Cas9 mediated TaRPK1 root architecture gene mutagenesis confers enhanced wheat yield. J. King Saud Univ. Sci. 2024, 36, 103063. [Google Scholar] [CrossRef]
- Ibrahim, S.; Saleem, B.; Naeem, M.K.; Arain, S.M.; Khan, M.R. Next-generation technologies for iron and zinc biofortification and bioavailability in cereal grains. Crop Pasture Sci. 2021. [Google Scholar] [CrossRef]
- Wang, W.; Simmonds, J.; Pan, Q.; Davidson, D.; He, F.; Battal, A.; Akhunova, A.; Trick, H.N.; Uauy, C.; Akhunov, E. Gene editing and mutagenesis reveal inter-cultivar differences and additivity in the contribution [i]of TaGW2 homoeologues to grain size and weight in wheat. Theor. Appl. Genet. 2018, 131, 2463–2475. [Google Scholar] [CrossRef]
- Dong, O.X.; Yu, S.; Jain, R.; Zhang, N.; Duong, P.Q.; Butler, C.; Li, Y.; Lipzen, A.; Martin, J.A.; Barry, K.W.; et al. Marker-free carotenoid-enriched rice generated through targeted gene insertion using CRISPR-Cas9. Nat. Commun. 2020, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Figueredo, E.F.; da Cruz, T.A.; de Almeida, J.R.; Batista, B.D.; Marcon, J.; de Andrade, P.A.M.; Hayashibara, C.A. de A.; Rosa, M.S.; Azevedo, J.L.; Quecine, M.C. The Key Role of indole-3-acetic acid biosynthesis by Bacillus Thuringiensis RZ2MS9 in promoting maize growth revealed by the IpdC gene knockout mediated by the CRISPR-Cas9 system. Microbiol. Res. 2023, 266, 127. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, S.; Du, W.; Ding, Y.; Li, W.; Chen, Y.; Zheng, Z.; Wang, Y. Sugar transporter ZmSWEET1b is responsible for assimilate allocation and salt stress response in maize. Funct. Integr. Genom. 2023, 23, 137. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Zheng, X.; Wang, W.; Yin, X.; Liu, H.; Ma, C.; Niu, X.; Zhu, J.-K.; Wang, F. Creation of aromatic maize by CRISPR/Cas. J. Integr. Plant Biol. 2021, 63, 1664–1670. [Google Scholar] [CrossRef]
- Kalischuk, M.L.; Johnson, D.; Kawchuk, L.M. Priming with a double-stranded DNA virus alters Brassica rapa seed architecture and facilitates a defense response. Gene 2015, 557, 130–137. [Google Scholar] [CrossRef]
- Kalischuk, M.L.; Fusaro, A.F.; Waterhouse, P.M.; Pappu, H.R.; Kawchuk, L.M. Complete genomic sequence of a Rubus Yellow Net Virus isolate and detection of genome-wide pararetrovirus-derived small RNAs. Virus Res. 2013, 178, 306–313. [Google Scholar] [CrossRef]
- Kalischuk, M.; Hachey, J.; Kawchuk, L. Complete genome sequence of phytopathogenic Pectobacterium atrosepticum bacteriophage Peat1. Genome Announc. 2015, 3. [Google Scholar] [CrossRef]
- Kalischuk, M.; Hachey, J.; Thomas, D.; Johnson, D.; Kawchuk, L. Complete genome sequence of phytopathogenic Pectobacterium atrosepticum lytic bacteriophage Peat2. Am. J. Potato Res. 2019, 96, 614–616. [Google Scholar] [CrossRef]
- Kathiria, P.; Sidler, C.; Golubov, A.; Kalischuk, M.; Kawchuk, L.M.; Kovalchuk, I. Tobacco mosaic virus infection results in an increase in recombination frequency and resistance to viral, bacterial, and fungal pathogens in the progeny of infected tobacco plants. Plant Physiol. 2010, 153, 1859–1870. [Google Scholar] [CrossRef]
- Kalischuk, M.; Paret, M.L.; Freeman, J.H.; Raj, D.; Da Silva, S.; Eubanks, S.; Wiggins, D.J.; Lollar, M.; Marois, J.J.; Mellinger, H.C.; et al. An improved crop scouting technique incorporating unmanned aerial vehicle-assisted multispectral crop imaging into conventional scouting practice for gummy stem blight in watermelon. Plant Dis. 2019, 103, 1642–1650. [Google Scholar] [CrossRef]
- Kawchuk, L.M.; Lynch, D.R.; Kozub, G.C.; Nelson, G.A.; Kulcsar, F.; Fujimoto, D.K. Multi-year evaluation of Clavibacter michiganensis subsp. sepedonicus disease symptoms in cultivated potato genotypes. Am. J. Potato Res. 1998, 75, 235–243. [Google Scholar] [CrossRef]
- Hill, B.D.; Kalischuk, M.; Waterer, D.R.; Bizimungu, B.; Howard, R.; Kawchuk, L.M. An environmental model predicting bacterial ring rot symptom expression. Am. J. Potato Res. 2011, 88, 294–301. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Kawchuk, L.M.; Armstrong, J.D.; Lynch, D.R.; Knowles, N.R. Potatoes having improved quality characteristics and methods for their production. U.S. Patent, 1999, 5,998,701.
- Kawchuk, L.M.; Hachey, J.; Lynch, D.R.; Kulcsar, F.; van Rooijen, G.; Waterer, D.R.; Robertson, A.; Kokko, E.; Byers, R.; Howard, R.J.; Fischer, R.; Prüfer, D. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 6511–6515. [Google Scholar] [CrossRef] [PubMed]
- Kawchuk, L.M.; Lynch, D.R.; Hachey, J.; Kulcsar, F. Tomato nucleic acid sequences that confer resistance to Verticillium and plants transformed therewith. U.S. Patent, 2003, 6,608,245.
- Lynch, D.R.; Kozub, G.C.; Kawchuk, L.M. The relationship between yield, mainstem number, and tuber number in five maincrop and two early-maturing cultivars. Am. J. Potato Res. 2001, 78, 83–90. [Google Scholar] [CrossRef]
- Berman, A.; Su, N.; Li, Z.; Landau, U.; Chakraborty, J.; Gerbi, N.; Liu, J.; Qin, Y.; Yuan, B.; Wei, W.; Yanai, O.; Mayrose, I.; Zhang, Y. Construction of multi-targeted CRISPR libraries in tomato to overcome functional redundancy at genome-scale level. Nat. Commun. 2025, 16, 4111. [Google Scholar] [CrossRef]
- Khadhair, A.H.; Kawchuk, L.M.; Taillon, R.C.; Botar, G. Detection and molecular characterization of an aster yellows phytoplasma in parsley. Can. J. Plant Path. 1998, 20, 55–61. [Google Scholar] [CrossRef]
- Kalischuk, M.L.; Kawchuk, L.M.; Leggett, F. First report of Rubus yellow net virus on Rubus idaeus in Alberta, Canada. Plant Dis. 2008, 92, 974. [Google Scholar] [CrossRef] [PubMed]
- Henrickson, A.; Kalischuk, M.; Lynn, J.; Meers, S.; Johnson, D.; Kawchuk, L. First report of zebra chip on potato in Canada. Plant Dis. 2019, 1016. [Google Scholar] [CrossRef]
- Kalischuk, M.L.; Al-Mughrabi, K.I.; Peters, R.D.; Howard, R.J.; Platt, H.W.; Kawchuk, L.M. Genetic composition of Phytophthora infestans in Canada reveals migration and increased diversity. Plant Dis. 2012, 96, 1729–1735. [Google Scholar] [CrossRef]
- Devendran, R.; Kavalappara, S.R.; Simmons, A.M.; Bag, S. Whitefly-transmitted viruses of cucurbits in the Southern United States. Viruses 2023, 15, 2278. [Google Scholar] [CrossRef]
- Wijekoon, C.P.; Kalischuk, M.L.; Brunelle, P.; Howard, R.J.; Kawchuk, L.M. Characterization of bronze leaf disease in western Canadian aspen and poplar trees. Can. J. Plant Sci. 2021, 102, 11–19. [Google Scholar] [CrossRef]
- Babu, B.; Paret, M.L.; Martini, X.; Knox, G.W.; Riddle, B.; Ritchie, L.; Aldrich, J.; Kalischuk, M.; Da Silva, S. Impact of foliar application of acibenzolar-S-methyl on rose rosette disease and rose plant quality. Plant Dis. 2022, 106, 818–827. [Google Scholar] [CrossRef]
- Kalischuk, M.; Lynn, J.; Kawchuk, L. First report of Potato mop-top virus infecting potatoes in Alberta. Plant Dis. 2016, 100, 2544. [Google Scholar] [CrossRef]
- McNally, J.; Prapagar, K.; Goldenhar, K.; Pate, E.; Shan, S.; Kalischuk, M. First report of an aggressive species of Neopestalotiopsis affecting strawberry in Canada. New Dis. Rep. 2023, 48, e12210. [Google Scholar] [CrossRef]
- Kalischuk, M.; Hendricks, K.; Hochmuth, R.; Freeman, J.; Roberts, P.D.; Paret, M. A portable recombinase polymerase amplification assay for the rapid detection of cucurbit leaf crumple virus in watermelon leaves and fruits. J. Plant Pathol. 2022, 104, 215–224. [Google Scholar] [CrossRef]
- Luckew, A.; Meru, G.; Wang, Y.Y.; Mwatuwa, R.; Paret, M.; Carvalho, R.; Kalischuk, M.; da Silva, A.L.; Candian, J.; Dutta, B.; Srinivasan, R.; Kavalappara, S.R.; RRD, N.C.K.; Bag, S.; McGregor, C. Field evaluation of Cucurbita germplasm for resistance to whiteflies and whitefly-transmitted viruses. HortScience 2022, 57, 337–344. [Google Scholar] [CrossRef]
- Kalischuk, M.L.; Roberts, P.D.; Paret, M.L. A rapid fluorescence-based real-time isothermal assay for the detection of Cucurbit yellow stunting disorder virus in squash and watermelon plants. Mol. Cell. Probes 2020, 53, 101613. [Google Scholar] [CrossRef]
- Kalischuk, M.; Baek, J.; Koeppe, S.; Shan, S.; Pate, E.; Goldenhar, K. Neopestalotiopsis disease of strawberry in Ontario – 2020 to 2022. Can. Plant Dis. Surv. 2025, 105, 229–233. [Google Scholar]
- Bairoch, A.; Boeckmann, B. The SWISS-PROT protein sequence data bank. Nucleic Acids Res. 1992, 20 Suppl., 2019–2022. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Tim Green, Michael Figurnov, Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; Bridgland, A.; Meyer, C.; Kohl, S.A.A.; Ballard, A.J.; Cowie, A. Highly accurate protein structure prediction with AlphaFold. Nature 596(2021), 583–589. [CrossRef] [PubMed]
- Kawchuk, L.; Jaag, H.M.; Toohey, K.; Martin, R.; Rohde, W.; Prüfer, D. In planta agroinfection by Canadian and German Potato leafroll virus full-length cDNAs. Can. J. Plant Pathol. 210(24), 239–243.
- Ma, W.J.; Wang, C.; Kothandapani, J.; Luzentales-Simpson, M.; Menzies, S.C.; Bescucci, D.M.; Lange, M.E.; Fraser, A.S.C.; Gusse, J.F.; House, K.E.; Moote, P.E.; Xing, X.; Grondin, J.M.; Hui, B.W.; Clarke, S.T.; Shelton, T.G.; Haskey, N.; Gibson, D.L.; Martens, E.C.; Abbott, D.W.; Inglis, G.D.; Sly, L.M.; Brumer, H. Bespoke plant glycoconjugates for gut microbiota-mediated drug targeting. Science 2025, 388, 1410–1416. [Google Scholar] [CrossRef]
- Prüfer, D.; Kawchuk, L.M.; Rohde, W. Polerovirus ORF0 genes induce a host-specific response resembling viral infection. Can. J. Plant Pathol. 2006, 28, 302–309. [Google Scholar] [CrossRef]
- Fusaro, A.F.; Barton, D.A.; Nakasugi, K.; Jackson, C.; Kalischuk, M.L.; Kawchuk, L.M.; Vaslin, M.F.; Correa, R.L.; Waterhouse, P.M. The luteovirus P4 movement protein is a suppressor of systemic RNA silencing. Viruses 2017, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; Liu, D.R. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Tuncel, A.; Pan, C.; Clem, J.S.; Lui, D.; Qi, Y. CRISPR-Cas applications in agriculture and plant research. Nat. Rev. Mol. Cell Biol. 2025, 26, 419–441. [Google Scholar] [CrossRef]
- Bai, S.; Cao, X; Hu, L.; Hu, D.; Li, D.; Sun, Y. Engineering an optimized hypercompact CRISPR/Cas12j-8 system for efficient genome editing in plants. Plant Biotechnol. J. 2025, 23, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Waltz, E. GABA-enriched tomato is first CRISPR-edited food to enter market. Nat. Biotech. 2022, 40, 9–11. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A schematic illustration of the Clustered Regularly Interspaced Palindromic Repeats (CRISPR) single guide RNA (sgRNA)and Cas endonuclease. The sgRNA consists of the N20 CRISPR RNA (crRNA) guide sequence and the trans-activating RNA (tracrRNA) that acts as a scaffold for Cas recognition. Restriction of the targeted DNA occurs 3 nucleotides 5’ to NGG Protospacer Adjacent Motif (PAM) sequence and sgRNA transcript starting with 5’G transcription initiation nucleotide using the U6 promoter.
Figure 1.
A schematic illustration of the Clustered Regularly Interspaced Palindromic Repeats (CRISPR) single guide RNA (sgRNA)and Cas endonuclease. The sgRNA consists of the N20 CRISPR RNA (crRNA) guide sequence and the trans-activating RNA (tracrRNA) that acts as a scaffold for Cas recognition. Restriction of the targeted DNA occurs 3 nucleotides 5’ to NGG Protospacer Adjacent Motif (PAM) sequence and sgRNA transcript starting with 5’G transcription initiation nucleotide using the U6 promoter.

Figure 2.
Application CRISPR-Cas to facilitate vector development and determine editing efficiency. A) Illustration of crRNA sequence for phytoene desaturase (PDS) in Nicotiana benthamiana in a sgRNA Cas complex [18,26]. B) Schematic diagram of DNA constructs used for expression and characterization of impact with the PDS visual reporter crRNA and tracrRNA [26,33,34]. C) Agroinfiltration of N. benthamiana leaves 3 weeks post-infiltration showing impact of the duplicated CaMV 35S promoter-enhancer as previously described [18,26,30,34].
Figure 2.
Application CRISPR-Cas to facilitate vector development and determine editing efficiency. A) Illustration of crRNA sequence for phytoene desaturase (PDS) in Nicotiana benthamiana in a sgRNA Cas complex [18,26]. B) Schematic diagram of DNA constructs used for expression and characterization of impact with the PDS visual reporter crRNA and tracrRNA [26,33,34]. C) Agroinfiltration of N. benthamiana leaves 3 weeks post-infiltration showing impact of the duplicated CaMV 35S promoter-enhancer as previously described [18,26,30,34].

Figure 3.
Introduction of CRISPR-Cas into plants for sequence editing. A) Binary vectors for Agrobacterium tumefaciens, simultaneously expressing sgRNA with desired crRNA target sequence [33,34]. B) Methods for transitional transformation include protoplasts (left), wounded tissue and callus formation (center), and axillary meristems (right) producing adventitious shoots (right) [18]. (C) Sequence of phytoene desaturase (PDS) editing detected following agroinfiltration of Nicotiana benthamiana leaves 3 weeks post-infiltration identifies nucleotide insertions and deletions (gold) as described by previous studies [18,26].
Figure 3.
Introduction of CRISPR-Cas into plants for sequence editing. A) Binary vectors for Agrobacterium tumefaciens, simultaneously expressing sgRNA with desired crRNA target sequence [33,34]. B) Methods for transitional transformation include protoplasts (left), wounded tissue and callus formation (center), and axillary meristems (right) producing adventitious shoots (right) [18]. (C) Sequence of phytoene desaturase (PDS) editing detected following agroinfiltration of Nicotiana benthamiana leaves 3 weeks post-infiltration identifies nucleotide insertions and deletions (gold) as described by previous studies [18,26].

Figure 4.
Schematic overview of key considerations and limitations associated with CRISPR–Cas–mediated crop breeding. A) Gene editing of chromosomes (colored) in polyploid plants can generate dominant, codominant, incompletely dominant, additive, or recessive alleles (edits indicated by black and white zigzag arrows), with reported editing efficiencies of up to 89% in dicots and 91% in monocots [34]. B) Edited tissues may be multiplied through subculture and vegetative propagation or through sexual crosses and seed production, followed by molecular screening to determine genotypic sequences, the presence of transgenic nucleic acids, and levels of heterozygosity. Current CRISPR–Cas applications are largely limited to well-characterized, simply inherited traits, whereas complex quantitative trait loci remain challenging targets. C) Phenotypic screening of germplasm is essential to confirm trait expression, detect deleterious off-target edits, and ensure agronomic performance. While a known single locus edit may be achieved within months for trait modification, the timeline for variety selection and release still involves many steps over several years.
Figure 4.
Schematic overview of key considerations and limitations associated with CRISPR–Cas–mediated crop breeding. A) Gene editing of chromosomes (colored) in polyploid plants can generate dominant, codominant, incompletely dominant, additive, or recessive alleles (edits indicated by black and white zigzag arrows), with reported editing efficiencies of up to 89% in dicots and 91% in monocots [34]. B) Edited tissues may be multiplied through subculture and vegetative propagation or through sexual crosses and seed production, followed by molecular screening to determine genotypic sequences, the presence of transgenic nucleic acids, and levels of heterozygosity. Current CRISPR–Cas applications are largely limited to well-characterized, simply inherited traits, whereas complex quantitative trait loci remain challenging targets. C) Phenotypic screening of germplasm is essential to confirm trait expression, detect deleterious off-target edits, and ensure agronomic performance. While a known single locus edit may be achieved within months for trait modification, the timeline for variety selection and release still involves many steps over several years.


Table 1.
Abiotic CRISPR-Cas improvements.
| Trait | Plant | Locus | Reference |
|---|---|---|---|
| Drought | Arabidopsis | AtOST2 | 35 |
| AtAREB1 | 36 | ||
| Glycine max | GMMYB118 | 37 | |
| Orza sativa | OsERA1 | 38 | |
| OsPUB7 | 39 | ||
| StDRO2 | 40 | ||
| Zea mays | ZmHDT103 | 41 | |
| Salt | Arabidopsis | AtWRKY3, AtWRKY4 | 42 |
| AtACQOS | 43 | ||
| G. max | GmDrb2a, GmDrb2b | 44 | |
| GmAITR | 45 | ||
| GmNHL1 | 46 | ||
| GmCG | 47 | ||
| Hordeum vulgare | HvGSK1.1 | 48 | |
| Medicago truncatula | MtHEN1 | 49 | |
| O. sativa | OsDST | 50 | |
| OsRAV2 | 51 | ||
| OsRP22 | 52 | ||
| OsNAC45 | 53 | ||
| OsSPL10 | 54 | ||
| OsTPP3 | 55 | ||
| Solanum lycopersium | SIHyPRP1 | 56 | |
| SIHAK20 | 57 | ||
| Solanum tuberosum | Stcoilin | 58 | |
| Triticum aestivum | TaHAG1 | 59 | |
| Zea mays | ZmHKT2 | 60 | |
| Heat | Gossypium hirsutum | GhPGF, GhCLA1 | 61 |
| Lactuca sativa | LsNCED4 | 62 | |
| O. sativa | GmHSP17.5Ep | 63 | |
| OsDEP1, OsROC5 | 64 | ||
| OsGER4 | 65 | ||
| OsPYL | 66 | ||
| OsTMS5 | 67 | ||
| S. lycopersicum | SIAGL6 | 68 | |
| SICPK28 | 69 | ||
| SIMAPK3 | 70 | ||
| T. aestivum | TaHsfA1 | 71 | |
| Z. mays | ZmTMS5 | 72 | |
| ZmHSPs | 73 | ||
| Cold | O. sativa | OsPRP1 | 74 |
| OsbHLH57 | 75 | ||
| S. lycopersicum | SICBF1 | 76 | |
| S. tuberosum | StVInv | 77 | |
| T. aestivum | TaPGK | 78 | |
| Z. mays | ZmG6PDH1 | 79 | |
| Metal | Arabidopsis | Atoxp1 | 80 |
| O. sativa | OsARM1 | 81 | |
| OsHAK1 | 82 | ||
| OsLCD | 83 | ||
| OsLCT1 | 84 | ||
| OsMYB84 | 85 | ||
| OsNIP3 | 86 | ||
| OsNramp5 | 87 | ||
| OsNRAMP1 | 88 | ||
| OsZIP5, OsZIP9 | 89 | ||
| OsPRX2 | 90 | ||
| OsATX1 | 91 | ||
| T. aestivum | TalPK1 | 92 | |
| Herbicide | Brassica napus | BnALS | 93 |
| Manihot esculenta | MeEPSPS | 94 | |
| O. sativa | OsALS | 95 | |
| OsTB1 | 96 | ||
| OsPUT1/2/3 | 97 | ||
| OsACC | 98 | ||
| UV Radiation | O, sativa | OsCOP1 | 99 |
| Oxidation | O. sativa | OsCAT2 | 100 |
Table 2.
Biotic CRISPR-Cas improvements.
| Biotic Stress Type | Pathogen/Pest | Plant | Locus | Reference |
|---|---|---|---|---|
| Fungi/ Fungus-like |
Powdery mildew | Cucumis sativus | CsMLO8 | 101 |
| Petunia x hybrida | PhMLO | 102 | ||
| Solanum lycopersicum | SIMLO | 103 | ||
| S. lycopersicum | SIPMR4 | 104 | ||
| Vitis vinifera | VvMOL3 | 105 | ||
| Late blight | S. lycopersicum | miR482b/c | 106 | |
| Solanum tuberosum | S-genes | 107 | ||
| CCoAOMT | 108 | |||
| Solanum americanum | SaNRL1 | 109 | ||
| Gray mold | S. lycopersicum | SIPL | 110 | |
| White mold | Glycine max | Gm5g29080 | 111 | |
| Stripe rust | Triticum aestivum | TaCIPK14 | 112 | |
| Southern late blight | Zea mays | ZmAGO18b | 113 | |
| Rice blast | Oryza sativa | OsPi21 | 114 | |
| Bacteria | Bacterial blight | O. sativa | OsSWEET115 | 115 |
| OsPUB9 | 116 | |||
| Citrus sinensis | CsLOB1 | 117 | ||
| Bacterial speck | S. lycopersicum | SIJAZ2 | 118 | |
| Bacterial spot | S. lycopersicum | SIBs5 | 119 | |
| Bacterial leaf streak | O. sativa | OsSULTR | 120 | |
| Bacterial wilt | S. lycopersicum | SIPRP1/DEA1 | 121 | |
| SIGAD2 | 122 | |||
| Virus | Bean yellow dwarf | Nicotiana benthamiana | LIR | 123 |
| Beet severe curly top | N. benthamiana | IR | 124 | |
| Tomato yellow leaf curl | N. benthamiana | IR | 125 | |
| Tomato yellow leaf curl | S. lycopersicum | CP/Rep | 126 | |
| Cotton leaf curl | N. benthamiana | IR | 127 | |
| Wheat dwarf | Hordeum vulgare | MP/CP/IP | 128 | |
| Cotton leaf curl | N. benthamiana | Rep | 129 | |
| Cauliflower mosaic | A. thaliana | CP | 130 | |
| Banana streak | Musa spp. | ORF1, 2, 3 | 131 | |
| Chili leaf curl | N. benthamiana | C + V | 132 | |
| Tomato yellow leaf curl | S. lycopersicum | SIPelo | 133 | |
| Cucumber mosaic | N. benthamiana | ORF1a, 3a | 134 | |
| Potato virus Y, S, and A | S. tuberosum | P3, CI, CP | 135 | |
| Geminiviruses | Manihot esculenta | IR, ORFs | 136 | |
| Potato spindle viroid | S. lycopersicon | SIDCL2b | 137 | |
| Maize rough dwarf | Z. mays | ZmGDla | 138 | |
| Yellow mottle | O. sativa | OsCPR5.1 | 139 | |
| Pest | Plant hopper | O. sativa | OsCKX | 140 |
| Insects | Gossypium hirsutum | GhMLP423 | 141 | |
| Chewing insects | G. max | GmUGT | 142 | |
| Root-knot nematode | O. sativa | OsHPP04 | 143 | |
| Cyst nematode | G. max | GmSNAP02 | 144 | |
| Parasitic weeds | Sorghum bicolor | CCD | 145 | |
| Beneficial | Mycorrhizal | Marchantia paleacea | CCaMK | 146 |
| Nitrogen fixation | O. sativa | CYP75 | 147 |
Table 3.
Phenotypic agronomic CRISPR-Cas characteristic improvements.
| Plant | Locus | Trait | Reference |
|---|---|---|---|
| Brassica napus | BnITPK | Phytic acid | 151 |
| Ipomoea nil | DFR-B | Flower colour | 152 |
| Jatropha curcas | JcCYP735A, JcCKX | Growth | 153 |
| Citrullus lanatus | PDS gene | Colour | 154 |
| Glycine max | GmFT2a | Flowering | 155 |
| Hordeum vulgare | GW2.1 | Seed set | 156 |
| Hina | Grain hardiness | 157 | |
| HvHGGT, HvHPT | Vitamin E | 158 | |
| Manihot esculenta | MeMSIII | Starch synthesis | 159 |
| MeCYP79D1 | Cyanide | 160 | |
| Oryza sativa | OsIAA23 | Development | 161 |
| OsCKX | Growth and quality | 162 | |
| OsRDD1 | Photosynthesis | 163 | |
| OsHHO3 | Nitrogen uptake | 164 | |
| OsGn1a, OsGL3 | Grain number/size | 165 | |
| OsBADH2 | Fragrance | 166 | |
| OsPYL9 | Yield | 167 | |
| OsSD1 | Lodging | 168 | |
| OsRc, OsRd | Red rice | 169 | |
| Petunia hybrid | PhACO | Longevity | 170 |
| PhPDS | Albino | 171 | |
| Solanum lycopersicon | PI, PG2a, TBG4 | Fruit shelf-life | 172 |
| SIIAA9 | Parthenocarpy | 173 | |
| Solanum tuberosum | StMYB44 | Phosphate transport | 174 |
| StGBBS | Starch quality | 175 | |
| StSS6 | Starch quantity | 176 | |
| StSBE | Starch quality | 177 | |
| STPDS | Carotenoids | 178 | |
| St16DOX | Glycoalkaloids | 179 | |
| StSSR2 | Glycoalkaloids | 180 | |
| StPPO2 | Enzymatic browning | 181 | |
| FtsZ1 | Starch granule size | 182 | |
| Triticum aestivum | TaARE1 | Nitrogen use | 183 |
| TaRPK1 | Yield | 184 | |
| Zn and Fe uptake | 185 | ||
| TaGW2 | Seed size and weight | 186 | |
| Zea mays | SSU-crt1, ZmPSY | Carotenoid increase | 187 |
| ipdC | Improved growth | 188 | |
| ZmSWEET1b | Sugar transport | 189 | |
| Zmbadh2 | Aroma | 190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



