Submitted:
01 April 2026
Posted:
03 April 2026
You are already at the latest version
Abstract
The current use of artificial light during natural dark phase had been acquired contaminant dimensions, which is named “light pollution”. It is well known that the exposure to dim light at night (dLAN) during the postnatal period severely impair the immune system and related organs, but few reports have demonstrated the effect of dim light when exposed during foetal periods. That is why this report ask does dim light at night in two different stages of development (i.e., prenatal vs. postnatal exposure) generate a long-lasting dysregulation of circadian rhythms that modifies the circadian immune organization and responses of the spleen in the early adulthood? To answer this question, we exposed two groups of C57BL/6J male mice to dim night light at gestational and postnatal period and compared to control groups where the mice were exposured to light-dark conditions (12 h each, LD). Parametric and non-parametric activity/rest values were analyzed with circular statistics. Compared to their controls, we found differences in alpha, onset, offset, M10 and L5 startime in dLAN groups. We also assessed the transcript levels of clock genes and genes that mediate inflammation in spleen tissue and found a dampening daily variation in mRNA expression in both experimental groups. Finally, we use an ovalbumin (OVA) allergy challenge to test the B-cell response in the spleen and found a significant higher cell recruitment to the spleen and more anti-OVA IgE. Together, these results clearly show that dLAN, affects the peripheral molecular clocks and responses from the spleen and these effects are independent of period of life exposure of dim light at night.
Keywords:
dim light at night
; circadian disruption
; developmental programming
; splenic immune response
; inflammation
1. Introduction
Currently, exposure to light at night has reached unprecedented levels in human history, with estimates indicating that more than 80% of the world’s population now lives under night skies polluted by artificial light [1,2]. This environmental change has altered the natural photoperiod and disrupts the expression of genes that regulate the biological clock, a condition known as chronodisruption [3]. Chronodisruption has been associated with a wide range of adverse effects on human and animal health, particularly those related to the development of chronic diseases [4,5,6,7,8,9]. The widespread use of LED-based devices has resulted in pervasive exposure to dim light during the dark phase of the natural cycle, occurring throughout virtually all stages of ontogeny [10,11,12,13], even impacting the foetal period [14,15,16,17,18]. Lately, many studies had been associating dim light pollution and impaired immune system function [19,20,21,22,23,24,25,26,27,28,29,30,31]. Indeed, this significant change in ambient lighting has occurred concurrently with the marked increase in non-communicable diseases (NCDs) in the human population, many of which appear to involve meta-inflammation or chronic low-grade inflammation as a key component of their pathophysiology [32,33,34].
Furthermore, the well-known inflammatory component of these pathologies raises the question of how exposure to dim light during the dark phase of the daily cycle affects immune system plasticity and how these effects relate to alterations in the biological clock[5,35,36,37,38,39].
In this regard, extensive epidemiological and experimental studies in animal models support the hypothesis that exposure to environmental alterations during early stages of development can condition the risk of developing pathophysiological processes associated with chronic NCDs [40,41,42,43,44], which can be related with chronodisruption [8,12,20,31,42,45,46,47].
The immune effects generated by chronodisruption in the fetal stage are related with a pro-inflammatory environment, defective vascularization, reduced uterine CD161+ NK and impaired maternal-fetal exchange [48].
The spleen, the largest secondary lymphoid organ, performs a wide variety of immune functions [49,50] and has increasingly emerged as an intriguing target for studying inflammation-mediated diseases [51,52,53]. In addition, its well-documented sympathetic innervation [54,55,56,57,58,59] and the presence of endogenous circadian rhythms in its resident cells [60,61,62,63,64,65] make it a particularly interesting focus for investigating the circadian and immune effects caused by light pollution during the dark phase of the day. In this context, the relationship between chronodisruption, inflammation, immune system plasticity, and light pollution across different developmental stages, particularly gestational and postnatal periods, has not yet thoroughly studied. Therefore, the main aim of this study is to determine the effects of dim light at night during gestation (GdLAN) or the postnatal period (PdLAN) on activity-rest rhythms and on the spleen as a peripheral organ of the immune system.
In the present study, we exposed two groups of male mice to dim night light: GdLAN and PdLAN, both compared with a control group maintained under a standard light–dark (LD) cycle. Activity-rest rhythms were evaluated using both parametric and non-parametric measures derived from the actigraphy data and analyzed using appropriate circular statistical approaches. In addition, we assessed the expression levels of clock genes and inflammation-related genes in the spleen. Following an ovalbumin (OVA) allergy challenge, we quantified total splenocytes measured OVA-specific IgG- and IgE-producing B cells from the spleen. Our results indicate that dLAN induces chronodisruption in male mice, altering rest/activity patterns and affecting peripheral molecular clocks in the spleen. Importantly, these effects appear to occur regardless of whether exposure to dim light at night takes place during gestation or in the postnatal period.
2. Results
2.1. Running Wheel Activity in PdLAN and GdLAN vs. Control Groups
We evaluated voluntary locomotor activity rhythms to determine whether exposure to dim light during the dark phase induces circadian disruption. Visual comparison of the actograms across experimental conditions revealed clear evidence supporting dim light at night–induced chronodisruption (Figure 2A-C). A representative actogram from the LD control group is shown in Figure 2A. As expected, activity began immediately when the lights were switched off at ZT12 and ceased when the lights were turned on at ZT0, a pattern that remained consistent across all analyzed segments. Under constant darkness (DD; segments S5 and S6), animals exhibited the characteristic day-by-day phase advance in activity onset previously reported for C57BL/6 mice [77].
Figure 2B. show a representative actogram from a mouse in the PdLAN group. During the habituation period (S1), the animal displayed most of its activity during the dark phase (ZT12-ZT0) and minimal activity during the light phase (ZT0-ZT12). Interestingly, after the introduction of dim light at night at the beginning of segment S2, a delay in activity onset became evident. In addition, total activity decreased (around ZT5 ± 2 ZT) over the following days until approximately day 70, when the onset of activity advanced toward ZT11. A similar delay in activity onset has been observed in previously published actograms [78], although these alterations were not explicitly discussed by the authors. In contrast, the reduction in overall activity persisted throughout the experiment. During the DD segments (S5 and S6), we also observed a noticeable fragmentation of activity.
A representative actogram from the GdLAN group is shown in Figure 2C. In this case, the offset of activity was progressively delayed, extending to approximately ZT2 (± 2 ZT) and even continuing into the light phase. Complete actograms for all animals are provided in Supplementary Figure 3, Figure 4 and Figure 5. Because individual changes in daily locomotor activity were observed in the actograms, we next examined whether the overall behavioral activity profiles differed between groups. To address this, the percentage of daily activity was averaged across all animals within each group, and the resulting activity profiles were plotted in Figure 2D. Data were normalized to facilitate comparison of the activity distribution within each segment. For segments S5 and S6, data are expressed as circadian time (CT) because recordings were obtained under constant darkness (DD) conditions.
In the PdLAN group, segments S2-S5 showed a delay in the peak of activity, whereas a different pattern was observed in the GdLAN group. To statistically evaluate the effects of dim light at night on activity profile patterns within each segment, PdLAN and GdLAN groups were compared with the LD control group using two-way ANOVA with the factors group (GdLAN, PdLAN) and Zeitgeber time (ZT 0–24), aggregating all activity samples within each ZT (each ZT consisted of 1,440 samples).
Segment S1 was considered a habituation period, and therefore differences observed during this phase were not further analyzed because they likely reflected individual adaptation to the running wheel. In segment S2, after the habituation phase, a significant interaction between group and ZT was detected for the average activity profile [F(46,414)=2.765,p<0.0001]. Post hoc analysis revealed that the PdLAN group showed reduced activity at ZT12 (p = 0.0202), ZT22 (p = 0.0398), and ZT23 (p = 0.0175) (Figure 2F). In contrast, the GdLAN group displayed increased activity at ZT13 (p = 0.0291) (Figure 2F).
Additionally, in segment S4, at ZT14, when both LD and GdLAN groups began to reduce their activity, the PdLAN group exhibited the highest average activity levels. This difference was supported by a significant group x ZT interaction [F(46,414)=2.142, p<0.0001], with post hoc significance at ZT14 (p=0,0467) (Figure 2H).
We next examined whether dim light at night affected endogenous rest–activity rhythms under free-running conditions, achieved by transferring animals to constant darkness (DD). During segment S5, a group × circadian time (CT) interaction was detected for the average activity profile[F(46,414)=1.350, p=0.07]. Post hoc analysis indicated that the PdLAN group exhibited increased activity at CT21 (p = 0.0453) and CT22 (p = 0.0156) (Figure 2I). However, in the subsequent segment S6, no significant differences between groups were detected (Figure 2J).
The daily activity profiles observed in the LD control group, under both LD and DD conditions, were consistent with previously reported patterns in C57BL/6J male mice [40].
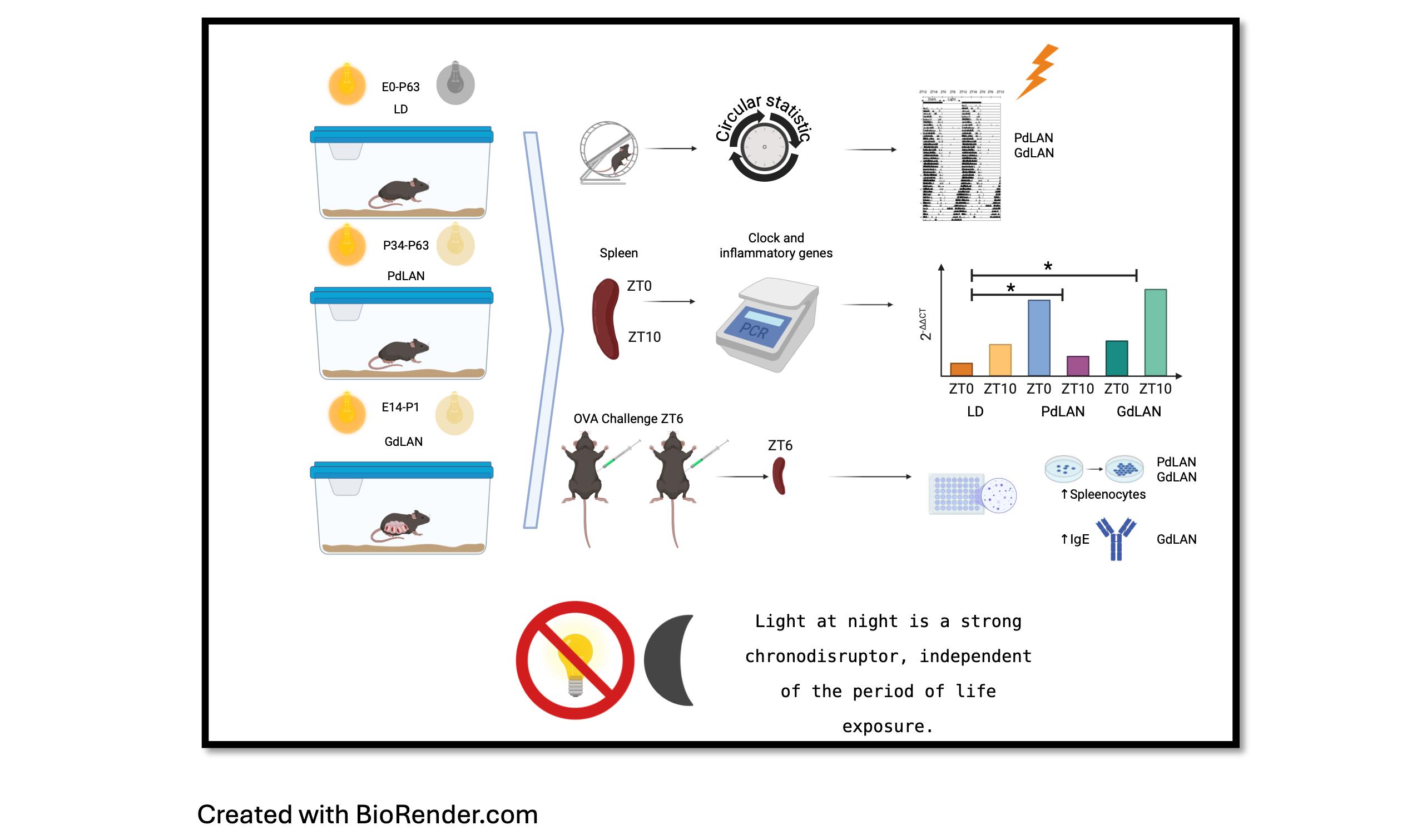
Figure 1.
Experimental design and illumination protocols. A. Schematic representation of the illumination protocols. At gestational day 14, dams were assigned to either standard 12 h light:12 h dark (12:12 LD) conditions for the control (LD) and postnatal dim light at night (PdLAN) groups, or to 12 h light:12 h dim light at night (L/Dim) conditions for the gestational dim light at night (GdLAN) group. At postnatal day 21 (P21), litters were sexed and weaned. In the GdLAN group, immediately after birth dams and litters were transferred to LD conditions. In the PdLAN group, male mice were exposed to L/Dim conditions beginning at P34 until sample collection, typically at P65. Illuminance was maintained at 150 ± 10 lux during the light phase, 0 lux during the dark phase, and 5 ± 1 lux during the dim phase. B. For locomotor activity experiments, three independent cohorts from the LD, PdLAN, and GdLAN groups were housed in running wheels from P28 to P69. At P56, lights were switched off and animals were maintained in constant darkness (DD) until P69. n = 7 per group. C. Additional cohorts from the LD, PdLAN, and GdLAN groups were used for mRNA expression analysis. At P65, spleens were collected at two Zeitgeber times (ZT0 and ZT10) and processed for real-time PCR quantification. n = 9 per ZT from independent experimental cohorts. D. Separate cohorts from the LD, PdLAN, and GdLAN groups were used for an allergic-like challenge and ELISpot assay. At P53 (ZT6) mice received an intraperitoneal injection of 100 μg ovalbumin (OVA) (day 0). A second injection was administered 7 days later as a challenge. Samples were collected at P65.
Figure 1.
Experimental design and illumination protocols. A. Schematic representation of the illumination protocols. At gestational day 14, dams were assigned to either standard 12 h light:12 h dark (12:12 LD) conditions for the control (LD) and postnatal dim light at night (PdLAN) groups, or to 12 h light:12 h dim light at night (L/Dim) conditions for the gestational dim light at night (GdLAN) group. At postnatal day 21 (P21), litters were sexed and weaned. In the GdLAN group, immediately after birth dams and litters were transferred to LD conditions. In the PdLAN group, male mice were exposed to L/Dim conditions beginning at P34 until sample collection, typically at P65. Illuminance was maintained at 150 ± 10 lux during the light phase, 0 lux during the dark phase, and 5 ± 1 lux during the dim phase. B. For locomotor activity experiments, three independent cohorts from the LD, PdLAN, and GdLAN groups were housed in running wheels from P28 to P69. At P56, lights were switched off and animals were maintained in constant darkness (DD) until P69. n = 7 per group. C. Additional cohorts from the LD, PdLAN, and GdLAN groups were used for mRNA expression analysis. At P65, spleens were collected at two Zeitgeber times (ZT0 and ZT10) and processed for real-time PCR quantification. n = 9 per ZT from independent experimental cohorts. D. Separate cohorts from the LD, PdLAN, and GdLAN groups were used for an allergic-like challenge and ELISpot assay. At P53 (ZT6) mice received an intraperitoneal injection of 100 μg ovalbumin (OVA) (day 0). A second injection was administered 7 days later as a challenge. Samples were collected at P65.

Figure 2.
Effect of dLAN on Locomotor activity. Locomotor activity was recorded individually for at least 41 consecutive days and grouped into six temporal segments (S1–S6), with n = 7 animals per group. Recording days are displayed vertically starting from day 1 and marked every 5 days. The horizontal axis indicates Zeitgeber time (ZT), where ZT0 corresponds to lights on and ZT12 to lights off. Activity records are double-plotted to facilitate visualization of circadian patterns. A. Representative actogram from a control animal maintained under light–dark (LD) conditions. B. Representative actogram from a postnatal dim light at night (PdLAN) animal. Periods of dim light exposure are indicated by grey shading. C. Representative actogram from a gestational dim light at night (GdLAN) animal. D. Percentage of daily locomotor activity averaged across animals from the LD, PdLAN, and GdLAN groups (n=7) per group). Data were normalized to illustrate activity distribution across each segment. During S5 and S6, animals were maintained in constant darkness (DD) and activity is expressed in circadian time (CT). The onset of activity was defined as CT12, representing the beginning of the subjective night, which extends for half of the circadian cycle; the remaining period corresponds to the subjective day. E-J. Segment-specific analysis of the percentage of daily activity across LD, PdLAN, and GdLAN groups (n = 7). LD is shown as a black solid line, PdLAN as a grey dashed line, and GdLAN as a black dotted line. Values represent mean ± SEM. Differences between PdLAN vs LD in grey asterisk and GdLAN vs LD in black asterisk. Two-way ANOVA and Sidak’s multiple comparisons posthoc test. *p<0.05 and ** p<0.01 asterisk in grey when the differences are between PdLAN vs LD control group and asterisk in black when the differences are between GdLAN vs LD control group.
Figure 2.
Effect of dLAN on Locomotor activity. Locomotor activity was recorded individually for at least 41 consecutive days and grouped into six temporal segments (S1–S6), with n = 7 animals per group. Recording days are displayed vertically starting from day 1 and marked every 5 days. The horizontal axis indicates Zeitgeber time (ZT), where ZT0 corresponds to lights on and ZT12 to lights off. Activity records are double-plotted to facilitate visualization of circadian patterns. A. Representative actogram from a control animal maintained under light–dark (LD) conditions. B. Representative actogram from a postnatal dim light at night (PdLAN) animal. Periods of dim light exposure are indicated by grey shading. C. Representative actogram from a gestational dim light at night (GdLAN) animal. D. Percentage of daily locomotor activity averaged across animals from the LD, PdLAN, and GdLAN groups (n=7) per group). Data were normalized to illustrate activity distribution across each segment. During S5 and S6, animals were maintained in constant darkness (DD) and activity is expressed in circadian time (CT). The onset of activity was defined as CT12, representing the beginning of the subjective night, which extends for half of the circadian cycle; the remaining period corresponds to the subjective day. E-J. Segment-specific analysis of the percentage of daily activity across LD, PdLAN, and GdLAN groups (n = 7). LD is shown as a black solid line, PdLAN as a grey dashed line, and GdLAN as a black dotted line. Values represent mean ± SEM. Differences between PdLAN vs LD in grey asterisk and GdLAN vs LD in black asterisk. Two-way ANOVA and Sidak’s multiple comparisons posthoc test. *p<0.05 and ** p<0.01 asterisk in grey when the differences are between PdLAN vs LD control group and asterisk in black when the differences are between GdLAN vs LD control group.

2.1.1. Circadian Locomotor Activity: Parametric and Non-Parametric Variables Analyzed with Linear Statistics
Because differences between groups were observed in both actograms and average activity profiles, we next compared parametric and non-parametric variables of the rest–activity rhythm between groups, as well as intragroup variations across experimental segments. We first describe the results obtained for linear parametric variables.
Analysis of the circadian period (τ) revealed no significant group × segment interaction (Figure 3A). In contrast, a significant group × segment interaction was detected for the activity interval (α) ([F(10,90)=7.210, p<0.0001]). Post host analysis showed that the PdLAN group displayed significantly lower α values in segments S2, S3, and S4 compared with the LD group (p=0.0007; p=0.0042 and p=0.0011, respectively; Figure 3B). The α values observed in the LD group were consistent with previously reported data for male C57BL/6 mice [40,79].
To further explore intragroup differences, we analyzed segment-to-segment variations within each group. As expected, in the LD group, α values were lower during segments S1–S4 compared with segments S5 and S6, reflecting the transition to constant darkness conditions. A similar pattern was observed in the PdLAN group. Interestingly, in the GdLAN group, no significant differences in α were detected across segments (Supplementary Figure 6A; p-values reported in Table 1). This result may reflect a reduced ability to entrain or an impaired increase in locomotor activity in response to constant darkness conditions.
When examining linear non-parametric variables, specifically interdaily stability (IS), we observed that the PdLAN group exhibited significantly lower IS values compared with the LD group in segments S3 and S5 (p = 0.0478 and p = 0.0384, respectively; Figure 3C). The IS values observed in the LD group were also consistent with previously reported data [40].
The other linear non-parametric variables, including interdaily variability (IV), M10, L5, and relative amplitude (RA) did not show significant differences either between groups or within groups across segments (Supplementary Figure 6B-E).
Figure 3.
Circadian locomotor activity variables. Locomotor activity was recorded individually for at least 41 days and grouped into six temporal segments (S1–S6). S1 corresponds to the first six days of recording and was considered a habituation period. S2 includes the subsequent seven days and coincides with the onset of dim light at night exposure in the PdLAN group. S3 represents the following seven days, and S4 the final week before the free-running period under constant darkness (DD). S5 corresponds to the first seven days in DD, and S6 to the subsequent seven days in DD. Circadian parameters were analyzed separately for each segment, including (A) circadian period, (B) activity duration (α), and (C) interdaily stability (IS). Data are presented as boxand-whisker plots (minimum to maximum) with n = 7 animals per group. Symbols. represent experimental groups as follows: LD (black circles), PdLAN (grey triangles), and GdLAN (black stars). Statistical comparisons were performed using two-way ANOVA followed by Dunnett’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001 asterisk in grey when the differences are between PdLAN vs LD control group and asterisk in black when the differences are between GdLAN vs LD control group.
Figure 3.
Circadian locomotor activity variables. Locomotor activity was recorded individually for at least 41 days and grouped into six temporal segments (S1–S6). S1 corresponds to the first six days of recording and was considered a habituation period. S2 includes the subsequent seven days and coincides with the onset of dim light at night exposure in the PdLAN group. S3 represents the following seven days, and S4 the final week before the free-running period under constant darkness (DD). S5 corresponds to the first seven days in DD, and S6 to the subsequent seven days in DD. Circadian parameters were analyzed separately for each segment, including (A) circadian period, (B) activity duration (α), and (C) interdaily stability (IS). Data are presented as boxand-whisker plots (minimum to maximum) with n = 7 animals per group. Symbols. represent experimental groups as follows: LD (black circles), PdLAN (grey triangles), and GdLAN (black stars). Statistical comparisons were performed using two-way ANOVA followed by Dunnett’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001 asterisk in grey when the differences are between PdLAN vs LD control group and asterisk in black when the differences are between GdLAN vs LD control group.

2.1.2. Circadian Locomotor Activity: Parametric and Non-Parametric Variables Analyzed with Circular Statistics
Although conventional statistical approaches are traditionally applied to analyze circadian locomotor variables, several parameters correspond to specific clock times or Zeitgeber time points, which are inherently circular rather than linear variables. For example, the time points 23:59 and 00:01 are separated by only two minutes, yet linear statistics would treat them as being nearly 24 h apart. For this reason, circular statistical methods were used to analyze these variables, with Zeitgeber time (ZT) as the unit of measurement.
Segments S5 and S6, corresponding to constant darkness (DD) conditions, were also evaluated using ZT units rather than circadian time (CT), maintaining the same cumulative time scale used under LD conditions. This approach allowed direct comparison of onset, offset, M10 start time, and L5 start time relative to the LD control group.
Analysis of acrophase revealed that, across all segments, subjects from the three groups were distributed with overall circular uniformity, and comparisons with the LD group did no revealed statistically significant differences. Nevertheless, the suggested a trend toward lower acrophase values in the PdLAN group under dim light conditions (Supplementary Figure 7).
We next examined whether dim light at night altered the onset of activity, evaluated using the parametric variable onset and the non-parametric variable M10 start time, which represents the beginning of the 10 consecutive hours with the highest activity. Interestingly, in the GdLAN group, segments S1 and S2 showed a significant advance in activity onset compared with the LD group (LD S1 and S2 circular mean = 12.09 and 12.18; GdLAN S1 and S2 circular mean = 11.54 and 11.58, respectively). In contrast, the PdLAN group exhibited a significant delay in activity onset during segment S2, coinciding with the introduction of dim light at night (LD and PdLAN S2 circular mean = 12.18 and 13.80, respectively). This delay persisted through segments S3 and S4, although statistical significance was detected only in S4 (LD S3 and S4 circular mean = 12.10 and 12.03; PdLAN S3 and S4 circular mean = 12.82 and 12.34, respectively). Under DD conditions, as expected, the LD group exhibited a phase advance in activity onset; however, during segment S5, the PdLAN group displayed an even larger advance compared with the LD group (LD and PdLAN S5 circular mean = 10.58 and 8.47, respectively). A summary of onset values is presented in Figure 4. Analysis of activity offset is summarized in Figure 5. In segment S1, the GdLAN group showed a significant delay relative to the LD group (LD and GdLAN S1 circular mean = 0.42 and 1.07, respectively). In contrast, the PdLAN group displayed a significant advance in activity offset during segments S2–S4, coinciding with the period of dim light exposure (LD S2–S4 circular mean = 0.47, 0.61, and 0.24; PdLAN S2–S4 circular mean = 21.96, 22.16, and 20.92, respectively). These results correspond with the reduced α values observed in the PdLAN group (Figure 3B). Interestingly, under DD conditions, this advance in offset observed in the PdLAN group disappeared, and values became comparable to those of the LD group (LD S5–S6 circular mean = 0.27 and 23.32; PdLAN S5–S6 circular mean = 23.17 and 22.67, respectively).
Figure 4.
Clock waypoints and summary of activity onset parameters obtained by circular statistics. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 to lights off, or to dim light onset in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to the constant darkness (DD) condition. Each point represents an individual activity onset value, while arrows indicate the circular mean vector for each experimental group. Group colors are indicated as follows: LD (control; light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red (n = 7 per group). Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk.(*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.
Figure 4.
Clock waypoints and summary of activity onset parameters obtained by circular statistics. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 to lights off, or to dim light onset in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to the constant darkness (DD) condition. Each point represents an individual activity onset value, while arrows indicate the circular mean vector for each experimental group. Group colors are indicated as follows: LD (control; light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red (n = 7 per group). Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk.(*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.


When analyzing the non-parametric variable M10 start time (Figure 6), the PdLAN group exhibited a significant advance beginning in segment S2 compared with the LD group (LD and PdLAN S2 circular mean = 12.53 and 11.67, respectively). This advance persisted through segment S6, although statistically significant differences relative to LD were detected only in segments S4 and S6 (LD S4 and S6 circular mean = 11.91 and 21.03; PdLAN S4 and S6 circular mean = 11.03 and 17.07, respectively). Notably, this effect persisted under DD conditions, like the pattern observed for activity onset. Together, these findings suggest that chronic exposure to dim light at night induces persistent chronodisruption. In contrast, the GdLAN group did not show significant differences compared with the LD group (Figure 6).
Figure 5.
Clock waypoints and summary of activity offset parameters obtained by circular statistics. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 corresponds to lights off, or to the onset of dim light in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to constant darkness (DD) conditions. Each point represents an individual value, and arrows indicate the circular mean vector for each group. Experimental groups are represented as follows: LD (control, light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red. Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk (*) in statistic significative differences between PdLAN and LD and Red asterisk (*) in statistic significative differences between GdLAN and LD.
Figure 5.
Clock waypoints and summary of activity offset parameters obtained by circular statistics. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 corresponds to lights off, or to the onset of dim light in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to constant darkness (DD) conditions. Each point represents an individual value, and arrows indicate the circular mean vector for each group. Experimental groups are represented as follows: LD (control, light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red. Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk (*) in statistic significative differences between PdLAN and LD and Red asterisk (*) in statistic significative differences between GdLAN and LD.


Finally, we examined L5 start time (Figure 7), which represents the onset of the five consecutive hours with the lowest activity. In the GdLAN group, subjects did not exhibit circular uniformity in segments S2–S4, indicating greater variability in the timing of minimal activity. Under DD conditions, however, the group displayed circular uniformity in segments S5 and S6, reflecting the emergence of endogenous rhythms. Despite this, significant circular differences relative to the LD group were detected in segments S5 and S6 (LD S5 and S6 circular mean = 1.34 and 15.42; GdLAN S5 and S6 circular mean = 4.44 and 4.18, respectively). When considered together with the onset and offset results, these findings suggest a more erratic activity pattern in animals exposed to dim light during gestation, possibly reflecting impaired entrainment to light–dark transitions. We can rule out defects in the development of the intrinsically photosensitive retinal ganglion cells, since they begin to form around day 2 after birth [80] and the maturation of this retina-hypothalamic system is complete around postnatal day 21 after weaning [81]. Notably, the only parameter in the LD control group that did not exhibit circular uniformity was L5 start time in segment S6. The PdLAN group also showed a lack of circular uniformity in this segment, but its circular mean did not differ significantly from the LD group (LD and PdLAN S6 circular mean = 15.42 and 14.69, respectively). A summary of L5 start-time values is provided in Figure 7.
Figure 6.
Clock waypoints and summary of M10 start-time parameters obtained by circular statistics. Clock way individual points and summary of M10 start-time’s values of circular mean, test statistic and p-values obtained by circular statistic. Clock in zeitgebers (ZT), ZT 0/24 when lights turn on and ZT12 when lights turn off or dim (since S2) in PdLAN group. Segments S5 and S6 correspond to dark-dark conditions. Each value is displayed individually, arrow represented circular mean, from each group. LD in black (Control group, Light and dark), PdLAN (Postnatal dim light at night) in blue, GdLAN (Gestational dim light at night) in red. Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk (*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.
Figure 6.
Clock waypoints and summary of M10 start-time parameters obtained by circular statistics. Clock way individual points and summary of M10 start-time’s values of circular mean, test statistic and p-values obtained by circular statistic. Clock in zeitgebers (ZT), ZT 0/24 when lights turn on and ZT12 when lights turn off or dim (since S2) in PdLAN group. Segments S5 and S6 correspond to dark-dark conditions. Each value is displayed individually, arrow represented circular mean, from each group. LD in black (Control group, Light and dark), PdLAN (Postnatal dim light at night) in blue, GdLAN (Gestational dim light at night) in red. Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7. Blue asterisk (*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.


2.2. Spleen Clock Genes Expression and Inflammatory Gene Expression After Immune Challenge
Having confirmed chronodisruption through alterations un circadian locomotor activity, we next examined whether peripheral immune function, specifically in the spleen, was also affected by dim at night exposure. To assess gene expression at time points distinct from the period of light-induced disruption (which occurs during the activity phase/dark period), tissue sampling was performed during the rest phase/light period at two specific Zeitgeber times: ZT0, corresponding to the beginning of the rest phase, and ZT10, approximately two hours before the expected onset of activity, when some aspect of the circadian physiology and immune functions begin to anticipate the upcoming active phase [35,38,39], for example start to increase corticosterone levels and decrease the proinflammatory cytokines.
Figure 7.
Clock way individual points and summary of L5 start-time’s values of circular mean, test statistic and p-values obtained by circular statistic. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 corresponds to lights off, or to the onset of dim light in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to constant darkness (DD) conditions. Each point represents an individual value, while arrows indicate the circular mean vector for each experimental group. Groups are represented as follows: LD (control, light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red. Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7.Blue asterisk.(*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.
Figure 7.
Clock way individual points and summary of L5 start-time’s values of circular mean, test statistic and p-values obtained by circular statistic. Clock time is expressed in Zeitgeber time (ZT), where ZT0/24 corresponds to lights on and ZT12 corresponds to lights off, or to the onset of dim light in the PdLAN group beginning at segment S2. Segments S5 and S6 correspond to constant darkness (DD) conditions. Each point represents an individual value, while arrows indicate the circular mean vector for each experimental group. Groups are represented as follows: LD (control, light–dark cycle) in black, PdLAN (postnatal dim light at night) in blue, and GdLAN (gestational dim light at night) in red. Circular data were analyzed using the Rayleigh test of uniformity for each group and Watson’s Two-Sample test of homogeneity versus LD control group. N=7.Blue asterisk.(*) in statistic significative differences between PdLAN and LD and red asterisk (*) in statistic significative differences between GdLAN and LD.


2.2.1. Spleen Clock Gene Expression
Two types of analyses were performed to evaluate the mRNA expression of clock genes in the spleen. First, we compared the two sampling time points (ZT0 and ZT10) to determine whether daily expression differences occurred within each group. Subsequently, we compared the expression levels at each ZT in the experimental groups (PdLAN and GdLAN) with those of the LD control group. As shown in Figure 8A, Bmal1 mRNA expression exhibited a significant intragroup × Zeitgeber time interaction [F(1,31)=9.080, p=0.0051]. Post hoc analysis revealed that, in the LD group, Bmal1 mRNA expression was significantly lower at ZT10 (p = 0.0325). This daily variation is consistent with previous reports in peritoneal macrophages [72,82]. Interestingly, this daily variation between both ZT was absent in both PdLAN and GdLAN groups. Moreover, a group x zeitgeber interaction was found ([F(2,31)=8.024, p=0.0016, with post hoc analysis showing that the GdLAN group displayed significantly lower Bmal1 mRNA expression at ZT10 compared with the LD group (p = 0.0009).
Analysis of Clock mRNA expression also revealed a significant interaction [F(2,46)=9.630, p=0.0003]. In the LD group, a daily difference between ZT0 and ZT10 was detected (p = 0.0023), with lower expression at ZT0. In the PdLAN group, a daily variation was also observed, but in contrast to the LD group, Clock mRNA expression was higher at ZT0. As observed for Bmal1, the GdLAN group showed a dampened daily variation in Clock mRNA expression between the two time points. When comparing between groups, both PdLAN and GdLAN groups exhibited significantly lower Clock mRNA expression at ZT10 (p = 0.001 and p = 0.0045, respectively) (Figure 8B).
The mRNA expression of Per2 showed an interaction with Zeitgeber time [F(1,48)=7.219, p=0.0099]. Post hoc analysis revealed a daily difference in the LD group (p=0.0069), with lower expression at ZT0. As expected, Per2 expression was in antiphase with Bmal1, a pattern previously reported in spleen tissue [82]. However, this daily variation was absent in both PdLAN and GdLAN groups. Additionally, a difference between LD and GdLAN groups was observed at ZT0 (p = 0.0058) (Figure 8C).The results for Nr1d1 are shown in Figure 8D. Expression of this gene displayed significant interactions with zeitgeber time and experimental protocol [F(1,48)=249, p<0.0001] and [F(2,48)=5.132, p=0.0096], respectively. Post hoc analysis revealed that all three groups (LD, PdLAN and GdLAN) exhibited daily variation, with ZT0 representing the time point with the lowest mRNA expression (p < 0.0001). However, the ZT10 expression levels were significantly lower in both PdLAN and GdLAN groups compared with the LD group (p = 0.0071 and p = 0.0001, respectively).
Finally, Rora mRNA expression also showed significant interactions with Zeitgeber time and experimental protocol [F(1,32)=12.13, p=0.0015] and[F(2,32)=6.716, p=0.0037], respectively. In the LD group, a significant daily variation was detected, with ZT0 showing lower expression levels, consistent with previous observations in peritoneal macrophages [72]. However, this daily oscillation was lost in both experimental groups. When comparing between groups, Rora expression at ZT10 was significantly reduced in both PdLAN and GdLAN groups (PdLAN p = 0.0003; GdLAN p = 0.0054) (Figure 8E).
2.2.2. Spleen Anti-Inflammatory and Pro-Inflammatory Gene Expression
The same analytical approach used for clock gene expression was applied to evaluate the mRNA expression of inflammatory genes. The results for Mcp-1 are shown in Figure 9A. Mcp-1 mRNA expression displayed significant interactions with Zeitgeber time and experimental protocol [F(2,47)=3.275, p=0.0466] and [F(2,47)=12.24, p<0.0001], respectively. Post hoc analysis revealed daily variation in the LD group, with higher expression at ZT0 compared with ZT10 (p = 0.0161). As observed for clock genes, this daily oscillation was dampened in both experimental groups (PdLAN and GdLAN). Additionally, Mcp-1 mRNA expression at ZT10 was significantly lower in both PdLAN and GdLAN groups compared with the LD control group, as revealed by post hoc analysis (PdLAN p < 0.0001; GdLAN p = 0.0309).
Analysis of Tnfa mRNA expression did not reveal daily differences in either the LD or GdLAN groups (Figure 9B). However, in the PdLAN group, a significant interaction was detected [F(2,47)=4.916, p=0.0115]). Post hoc analysis indicated a daily variation, with higher Tnfa mRNA expression at ZT0 (p = 0.0190). In addition, a significant difference was observed at ZT10 compared with the LD group (p = 0.0257).
The mRNA expression of Tgfb1 did not show significant interactions, daily variation, or differences between groups at either Zeitgeber time point (Figure 9C).For Il-6 mRNA expression, no daily differences were observed in either the LD or PdLAN groups. However, in the GdLAN group, a significant zeitgeber time interaction was detected [F(1,48)=10.02, p=0.0027]. Post hoc analysis revealed daily variation, with lower Il-6 mRNA expression at ZT0 (p < 0.0001) (Figure 9D).
Finally, analysis of Il-10 mRNA expression revealed significant interactions with Zeitgeber time and experimental protocol [F(2,44)=4.262, p=0.0203] and [F(2,44)=8.007, p=0.0011], respectively. Post hoc analysis revealed a daily variation in the LD control group, with higher Il-10 mRNA expression at ZT10 (p = 0.0063). As observed for other genes, this daily oscillation was absent in both PdLAN and GdLAN groups. Furthermore, Il-10 mRNA expression at ZT10 was significantly lower in both experimental groups compared with the LD control group (PdLAN p = 0.0048; GdLAN p < 0.0001) (Figure 9E).
2.3. Allergic-Like Ovalbumin (OVA) Challenge
After confirming that dim light at night affected both the daily variation and expression levels of clock and inflammatory genes in the spleen, we next investigated whether immune responses following an allergic-like challenge with ovalbumin (OVA) were also altered by dLAN exposure. Following the protocol illustrated in Figure 1, we observed a significantly higher number of splenocytes recruited in the experimental groups after the OVA challenge. The mean number of splenocytes in the LD control group was1.687 × 108, whereas the PdLAN group showed 2.5611 × 108 cells (p=0.0007) and the GdLAN group showed 2.7114 × 108 cells (p<0.001) (Figure 10A).
When analyzing the number of anti-OVA IgG–secreting B cells, measured as ELISpot spots, no significant differences were detected between groups (Figure 10B). However, analysis of anti-OVA IgE–secreting B cells revealed a significant increase in the GdLAN group compared with the LD control group (p = 0.0050) (Figure 10C).
3. Discussion
The physiological consequences of dim light at night (dLAN) have been widely investigated in adult animals that retain normal melatonin production19, 23, 24, 26-29, 83-87. In contrast, relatively few studies have examined the effects of light-at-night exposure during early developmental stages, and most of these investigations have also relied on melatonin-proficient models14, 15, 20, 21. Moreover, recent evidence suggests that the effects observed in transgenerational models may arise from mechanisms that extend beyond classical melatonin signaling 88.
In the present study, we investigated the circadian and immune consequences of dLAN exposure using a melatonin-deficient mouse model 89, 90. Two independent exposure paradigms were evaluated: postnatal dim light at night (PdLAN), representing continuous exposure during early adulthood, and gestational dim light at night (GdLAN), in which exposure occurred during prenatal development but produced persistent effects detectable in adulthood. Across these models we identified three major outcomes: (i) alterations in circadian locomotor activity and associated parametric and non-parametric variables, (ii) disruption of clock-gene and inflammatory-gene expression in the spleen, and (iii) an enhanced cellular recruitment response following an allergic-like ovalbumin (OVA) challenge. In addition, PdLAN, but not GdLAN, animals displayed increased body weight, consistent with previous reports30, suggesting that gain could serve as a useful phenotypic marker for validating PdLAN exposure in future studies using C57BL/6 male mice (see supplementary Figure 2). In animals exposed to PdLAN, the most prominent behavioral changes were a delay in activity onset and an advance in activity offset, resulting in a reduction of the active phase (alpha) and a more irregular pattern of locomotor activity compared with control LD conditions. Interestingly, the reduction in alpha was less pronounced than that reported in constant light protocols40. The application of circular statistics allowed a more robust comparison of actogram variables and revealed additional alterations in non-parametric parameters such as the start of the most active 10-h period (M10ST), which also displayed an advance even under free-running conditions. Previous studies using dim light at night have rarely explored these non-parametric variables, despite their value for describing circadian behavior. Although earlier reports using similar lighting conditions have described changes in voluntary activity in rodents26, 86, only one study clearly illustrated a delayed onset of activity in actograms, and this observation was not extensively analyzed because the focus of that work was sleep regulation 78. The disturbances observed in our study likely reflect impaired entertainability of the circadian system, in which dim nocturnal light may extend the perception of the subjective day and delay the transition into the active phase. Notably, similar alterations were detected in free-running conditions, suggesting that the disruption is not solely driven by external cues, also involves endogenous oscillators controlled by molecular clock genes91, 92. Just onset of activity exhibited discrepancies in DD conditions versus LD group. This phenomenon could be elucidated, yet the alteration of the other parameter was only observed under dim light at night masking, and when the animals were placed in DD showed its endogens rhythm. Collectively, these findings support the idea that PdLAN acts as a potent disruptor of rest–activity rhythms by altering the temporal alignment between activity onset and offset affecting the resting and active phase.
In GdLAN we found differences in the onset and offset, especially in the first-time segment of activity (S1), this period we named as a habituation period. Nevertheless, the differences in the onset appear in S2 segment too. These differences are principally due an advance in the onset, and in the behavior of activity especially because this group displayed a prominent anticipatory activity since ZT10 which change the pattern and the acrophase of the percentage of activity in segments S1 and S2. In GdLAN group the offset in S1 showed a delayed, also displayed a highest average of alpha respect to LD control group (13.60; 12.34, respectively) counter wise as expected because the activity starts early. This could be explained as a lack of entertainability due to phase change. This is reinforced by the absence of alpha differences between LD and DD period in the GdLAN group, differences that are present in the LD group (see Table 1). This subtle changes in activity entertainability are poor studied and reported in gestational models, nevertheless Delorme, in 2021 found significatively changes in the activity pattern, total count of activity and alpha in males’ offspring when the pregnant dams were insulted with a mimic of virus infection. The above supports the theory that insults in the pregnant mother impact circadian rhythms of the offspring40 and in our case was simply dim light at night. One factor that may explain this lack entrainment could be maternal training, specially because some authors have reported that offspring from suprachiasmatic nucleus lesioned (SCN) mothers started to free-run earlier than those from SCN-intact mothers relate with an impairment corticosterone rhythms in the pups93. They suggest that presence of maternal entrainment and the beginning of circadian oscillation at as early as E1094. After birth, in rats, maternal rhythms reset the circadian clock of the pups, the entertainability of which depends on the strength of maternal care, which in turn depends on factors such as a small litter size95, 96and the timing of maternal care97. We did not find difference in the size of de litters (we had an average of 6 pups per litter), mortality and weight of birth, consentient with other reports15. Unfortunately, we did not evaluate the maternal care.
The most alterations that we noticed in the GdLAN actogram’s occurred individually (see supplementary Figure 5) but was not a group characteristic. This is like what happens in variety of studies when the insults befell during pregnancy15, 20, 98, 99. Even in humans with the developmental origins of health and disease (DOHaD) theories which propose that what happens during pregnancy can be considered a predisposing factor to different diseases, but it is not always a predominant factor when diseases occur 41, 100. The multifactorial nature of NCDs further supports the idea that chronodisruption during gestation may represent one of several interacting risk factors101-103.
Analysis of splenic mRNA gene expression revealed substantial alterations in the molecular clock and inflammatory signaling pathways, although we know that the judgments and conclusions we can draw are limited when we use only two time points during the day to monitor circadian events. In control LD animals, all evaluated clock genes exhibited robust daily differences in this two time points consistent with previous reports describing circadian regulation in peripheral immune tissues62, 71, 73. In contrast, these differences were largely lost in both PdLAN and GdLAN groups. An exception was the mRNA Clock gene expression in PdLAN animals, which displayed an altered daily expression profile compared with controls. Both experimental groups also showed reduced the mRNA expression of Nr1d1 and Rora at ZT10. The reduction in Rora mRNA expression (fold change ~0.7 in PdLAN and ~0.5 in GdLAN) is particularly notable because this nuclear receptor is considered an anti-inflammatory component of the circadian clock and plays a key role in regulating cytokine production 104, 105. Consistent with this change, we also observed reduced the mRNA expression of the inflammatory mediators Mcp-1 and Il-10 in both experimental groups. Because Il-10 functions as a major anti-inflammatory cytokine that counteracts the actions of pro-inflammatory mediators such as Il-1, Tnf-α and Il-12106-108, the combined dysregulation of mRNA expression of Rora, Il-10, and Mcp-1 suggests a disturbance in immune homeostasis that could modify susceptibility to infections or allergic responses in this time points. These findings are consistent with previous work showing that chronic circadian misalignment can accelerate immune senescence and promote inflammatory phenotypes in peripheral organs such as the liver and kidney6.
Additional evidence of immune dysregulation emerged from the altered mRNA expression of Il-6 in GdLAN animals. In this group, Il-6 mRNA expression exhibited significant daily variation, with a marked increase at ZT10, coinciding with reduced mRNA levels of Il-10 and N1rd1. Because N1rd1 acts as a transcriptional repressor of inflammatory genes and negatively regulates the NLRP3 inflammasome37, 109, 110, its decreased expression may contribute to the pro-inflammatory environment suggested by the elevated Il-6 mRNA levels. Together, these observations point to the emergence of a pro-inflammatory state immediately before the onset of the active phase. However, given the limitations of sampling at two time points, we cannot determine whether the desynchronization in the rhythms is due to a phase advance, delay, resetting, or changes in amplitude.
In PdLAN animals, we detected a reduction in Tnfa mRNA expression at ZT10, a finding consistent with observations reported by Cisse et al. in offspring of parents exposed to dim nocturnal light. TNF-α has been shown to modulate clock-gene expression111, 112, suggesting the possibility of bidirectional regulation between inflammatory signaling and the circadian clock. The observed changes in Tnfa mRNA expression may therefore reflect epigenetic mechanisms triggered by nocturnal light exposure rather than direct effects mediated by clock-gene oscillations. Interestingly, the Clock gene itself has been implicated in both pro- and anti-inflammatory processes depending on its interacting partners and transcriptional context46 37. In macrophages, CLOCK activity has been associated with pro-inflammatory responses113, which may help explain the temporal changes in Tnfa mRNA expression observed in our PdLAN model.
The functional relevance of these molecular alterations became evident in the immune challenge experiments. Following OVA sensitization, both experimental groups displayed a marked increase in splenocyte numbers (approximately 1.5-fold in PdLAN and 1.7-fold in GdLAN). Because the antigen was administered intraperitoneally, the spleen is expected to serve as the primary lymphoid organ orchestrating the immune response. The elevated splenocyte counts therefore suggest enhanced immune cell recruitment or expansion. This interpretation is supported by the increased number of B cells producing OVA-specific IgE, which reached statistical significance in the GdLAN group. The altered immune response may also be related to the loss of rhythmic Bmal1 mRNA expression observed in both experimental conditions, as BMAL1 has been implicated in the regulation of B-cell trafficking and homing to lymphoid tissues 36.
To date, only a limited number of studies have explored the immune consequences of parental exposure to dim light at night. Previous reports have shown reduced T-cell-mediated responses and altered antibody production against novel antigens such as KLH in offspring of mothers exposed to nocturnal light20, 21. Interestingly, these effects appear to be transmitted primarily through maternal exposure, suggesting the involvement of transgenerational epigenetic mechanisms.
Our study has several limitations, Sampling for gene expression was limited to specific two time points rather than covering the full circadian cycle, the immune challenge was restricted to a single OVA exposure, and we use only male’s subjects, indeed we are working to include a sexual dimorphism for next publication. Now we want to characterize a baseline inflammatory tendency in naïve animals rather than fully stimulated immune responses. Nevertheless, the study also has notable strengths, including the use of both linear and circular statistical approaches to evaluate circadian behavioral data and the parallel analysis of two independent dLAN exposure paradigms.
In summary, our findings demonstrate that exposure to dim light at night—either during early adulthood or during gestation—induces significant chronodisruption in melatonin-deficient male mice. This disruption manifests as altered rest–activity rhythms, differences of clock-gene expression in peripheral immune tissue, and enhanced immune cell recruitment following antigenic challenge. Importantly, these effects occur regardless of the life stage at which light exposure takes place, highlighting the pervasive impact of nocturnal light pollution on circadian and immune physiology.
4. Materials and Methods
4.1. Animals
Animal handling and care followed the Guide for the Care and Use of Laboratory Animals of the Institute for Laboratory Animal Research of the National Research Council. C57BL/6J mice were kindly donated from Dr. Carlos Flores (Centro de Estudios Científicos) which originally were obtained from the Jackson Laboratories (product number: 000664). Mice were placed in ventilated cages and housed in our animal facility at the Universidad Austral de Chile. All animals used in this study had ad libitum access to water and food. Animals were fed with a standard diet (Prolab® RMH 3000, Labdiet). The protocols were approved by the Bioethics Commission of the Universidad Austral de Chile (# 328/218). This study incorporated a sample exclusively of male subjects, consistent with the findings of a preliminary experiment. The results of that experiment indicated that male subjects exhibited an increase in body mass under conditions of dim nighttime illumination (Supplementary Figure 1). The current study is ongoing, and the next publication will include a sample of female subjects.
4.2. Lighting Conditions
Animals were maintained under three different lighting conditions. The control group was housed under a standard 12 h light:12 h dark cycle (12:12 LD), whereas the experimental dim light groups—gestational dim light at night (GdLAN) or postnatal dim light at night (PdLAN)—were maintained under a 12 h light:12 h dim cycle (12:12 dim). An additional condition of constant darkness (DD) consisted of 24 h of uninterrupted darkness.
Illumination was provided using white LED light sources and was calibrated with a luxmeter (HI 95500, Hanna®). Light intensity during the light phase was set to 150 ± 10 lux, while the dark phase corresponded to 0 lux, and dim conditions were maintained at 5 ± 1 lux. Lights were programmed to turn on at 8:00 AM (GMT−4), corresponding to Zeitgeber Time 0 (ZT0), and to turn off at 8:00 PM (ZT12), following standard chronobiological conventions. Lighting schedules were controlled using an external timer.
The spectral characteristics of the light sources at 150 lux and 5 lux have been previously described (see Figure S3 in Sarmiento et al., 2022). To prevent external light contamination, animals were housed in isolated light-proof cabinets throughout the experiments.
4.3. Gestational (GdLAN) and Postnatal (PdLAN) Dim at Night Protocols
Pregnant dams were received at 14 days of gestation since vaginal plug detection (designated embryonic day 0.5, E0.5), and body weight gain was monitored thereafter. Upon arrival, dams were randomly placed in individual cages and assigned to either dim light at night conditions (for the GdLAN group) or standard LD conditions (for the PdLAN and control groups). A total of eight litters were utilized for the purpose of generating the samples within the LD and PdLAN groups. The animals were distributed in a balanced manner across both protocols, with each litter contributing an equal number of subjects. Five distinct litters were utilized for the GdLAN group. To circumvent the potential for the litter effect, it was imperative that experiments were not conducted using animals from the same litter in more than one experiment. Rather, the n was completed for each experiment with a minimum of three litters. To determine the exact timing and light phase of birth, pregnant dams were monitored beginning at E18, with inspections conducted three times during the dark/dim phase and twice during the light phase. After birth, dams and their offspring were maintained under LD conditions. At postnatal day 21 (P21), male pups were weaned, tagged, randomly allocated into cages of three individuals, and weighed weekly until reaching adulthood (P65 ± 3 days), when experimental testing was conducted.
A total of 34 male mice per condition (LD, GdLAN or PdLAN) were used across three different cohorts: n=7 for wheel-running assays, n=18 for mRNA expression analyses, and n=9 for the OVA challenge experiments. Dams were euthanized after weaning.
For the PdLAN model, male pups gestated under LD conditions were weighed, tagged, and housed in groups of three at P21 under LD conditions. At P35, these mice were transferred to dim light at night conditions, where they remained until adulthood (P65 ± 3 days), with body weight monitored weekly. A schematic representation of the experimental protocol is shown in Figure 1A.
4.4. Wheel-Running Activity Behavior (Actograms)
Animals were housed individually in cages equipped with running wheels beginning at postnatal day 28 (P28). In all experimental conditions, the period from P28 to P34 was considered a trained phase, while P35 to P55 corresponded to the experimental sampling period. Subsequently, animals were transferred to constant darkness (DD) conditions from P56 to P69. A schematic representation of this protocol is shown in Figure 1B. Wheel-running activity was recorded using a handmade electro-magnetic device connected to a BioPac MP100 data acquisition system. Raw data were processed using Acknowledge 3.9 software (BioPac Systems, Inc., Aero Camino Goleta, CA). Running wheels used were Respironics® Mini mitter®, 4.5” (ref. # 610-0004-00).
Circadian locomotor activity was quantified using both parametric and non-parametric variables as previously described [66]. Parametric parameters included circadian period (τ) calculated using a chi-square periodogram, acrophase (Φ), defined as the time interval during which peak activity occurs, and the times of activity onset and offset. Actograms and the calculation of these variables were obtained using the ActogramJ plugin in ImageJ2 (v2.3.0/1.53q) [67]. The activity interval (α) was calculated as the number of hours between activity onset and offset, and daily activity was expressed as a percentage of total daily activity.
Non-parametric circadian variables, including interdaily stability (IS) that quantifies the stability of rest-activity rhythms or the invariability of the rhythm between different days, interdaily variability (IV) that quantifies the fragmentation of a rest-activity pattern, L5 that is the averaged activity of the five consecutive hours with minimal activity, L5 onset, M10 that is the averaged activity of the ten consecutive hours with maximal activity, M10 onset, amplitude and relative amplitude calculated from the M10 and L5 values, were calculated in R (v. 4.1.21) using the nParACT package [68]. Variables representing non-linear 24-h periodic events expressed as Zeitgeber time (ZT) (acrophase, onset ZT, offset ZT, L5 start-time and M10 start-time) were analyzed using circular statistical methods implemented in R through the bpnreg and circular packages [69].
Onset variability was calculated as the difference between the activity onset and the time of lights-off, following the procedure described by Delorme (2021). Under DD conditions, onset variability was determined as the difference between the observed activity onset and the average onset of the final seven days preceding DD exposure.
To minimize potential masking effects [70], actogram data were analyzed in sequential segments corresponding to different phases of the experimental protocol. These segments were defined as follows: Segment 1 (S1), P29–P34; S2, P35–P41; S3, P42–P48; S4, P49–P55; S5, P56–P62; and S6, P63–P69. Segments S5 and S6 corresponded to the DD condition and in S5 we discard the first 3 days of record according with Jud, et al. 2005 [70], indication. Segment S1 represented the initial six days of sampling and served as the habituation period. Segment S2 comprised the following seven days and coincided with the introduction of dim light at night in the PdLAN group. Segment S3 corresponded to the subsequent week of recording, while S4 represented the final week prior to the free-running period in DD. Segments S5 and S6 corresponded to the first and second weeks under DD conditions, respectively. Under DD conditions, the beginning of the subjective night (circadian time 12, CT12) was defined as the time of activity onset, extending for half of the circadian period, while the remaining half of the cycle was defined as the subjective day.
4.5. RNA Extraction and Gene Expression
The experimental procedure is summarized in Figure 1.C. At postnatal day 65 (P65), mice were anesthetized with isoflurane and euthanized by cervical dislocation. The spleen was rapidly excised and immediately immersed in 1mL of RNAlater® (Sigma-Aldrich, Cat. #R0901) for 24h at 4°C, after which the samples were stored in liquid nitrogen until further processing.
Mice were euthanized at two circadian time points, ZT0 and ZT10 (n=9 per ZT). In these time points occurred the maximal and minimal expression levels reported for several circadian and inflammation-related genes in the spleen [61,71,72]. Additionally, we ensuring that sampling occurred during the light phase or resting period not in the dLAN intervention in PdLAN group. Moreover, sampling at two Zeitgeber times has been considered adequate to detect temporal differences in immune-related assays [73] but we are aware of the limitations of this sample.
Total RNA was extracted from 30mg of spleen tissue using Ambion TRIzol Reagent (Sigma-Aldrich, Cat. #66115) according to the manufacturer’s instruction and eluted in 50 μL of RNase free water. RNA concentration and integrity were assessed using a MaestroNano Micro-Volume Spectrophotometer (Maestrogen, Cat. #MN-913,) and confirmed by agarose gel electrophoresis. Prior to cDNA synthesis, 2 µg of total RNA were treated with RQ1 RNase-free DNase (Promega, Cat. ·M610A) following manufacturer’s instructions.
First-strand cDNA was synthesized using random primers (Promega® Cat. # C1181), a dNTP mix (dATP, dCTP, dGTP, dTTP Promega® Cat# U1330), and M-MLV reverse transcriptase kit (Promega® Cat. #M1701; Cat. #M5313), according to the manufacturer’s protocols.
The mRNA expression of five circadian clock genes and five inflammation-related genes was quantified using gene-specific primers (Supplementary Table 1) and SYBR® Green Supermix (Biorad® Cat. # 1725270) according to the manufacturer’s standard protocol. Amplification reactions were performed on a Rotor Gene Q 5Plex HRM (Qiagen, Hilden, Germany) using 5 μL cDNA template. β-actin (β-Actin) was used as the reference gene for normalization. Relative gene expression levels were calculated using the comparative CT method (2−ΔΔCT) [74,75].
4.6. Immune Antigen Like Allergic Challenge with Ovalbumin (OVA) and ELISpot Assay
The experimental protocol is summarized in Figure 1D. At ZT6, male mice at P53 were anesthetized with isoflurane and sensitized with 100 µg of OVA administered intraperitoneally (i.p.) on day 0 (P53) and day 7 (P60) (Sigma Cat. #A5503-5G). OVA was emulsified in aluminum hydroxide (Alum Imject, Thermo Scientific Cat. #77161) at an equal volume (250 µL each), resulting in a final injection volume of 500 µL, following a previously described protocol with minor modifications [76].
Five days after the second sensitization (P65), mice were anesthetized with isoflurane and euthanized by cervical dislocation. The spleen was immediately excised and mechanically dissociated in a laminar flow cabinet using ice-cold dishes containing 5mL of RPMI 1640 medium (Gibco Cat. #31800-02) supplemented with 10% fetal bovine serum (Biowest Cat. #S1810-500) and 1% penicillin-streptomycin (Gibco Cat. #151140-122). The resulting cell suspension was filtered through a 40µM cell strainer, and splenocytes were counted using a Neubauer hemocytometer.
For ELISpot assays, cells were plated inMultiScreen® filter plates (Merck® Cat. #MSIPS4510). For IgG detection, 2 x 105 cells per plate were seeded, whereas 5 x 105 cells per plate were used for IgE detection. All samples were plated in triplicate. OVA-specific antibody-secreting cells were detected using the Mouse IgG ELISpotBASIC (HRP) KIT (Mabtech® Cat. #3825-2h) and the Mouse IgE ELISpotBASIC (HRP) KIT (Mabtech® Cat. #3815-2h); with TMB substrate for ELISpot (Mabtech®, Cat. #3651-10), following the manufacturer’s instructions.
ELISpot plates were analyzed using an AID iSpot Reader ELR08IFL (Autoimmun Diagnostika GmbH).
4.7. Statistical Analysis
Statistical power was estimated based on a pilot study in which sex differences in body weight gain were observed as an effect of the PdLAN model (see Supplementary Figure 1). From this analysis, a sample size of n=9 per group was calculated to achieve a power of 80-90% with α= 0,05, using the StatMate 2.00 extension of GraphPad Prism (version 7.00 for Mac).
Data and analysis and graphical representations were performed using GraphPad Prism (version 7.00 for Mac), except for actograms and circadian parameters such as period, acrophase, activity onset and offset, which were obtained using the ActogramJ plugin in ImageJ2 (v2.3.0/1.53q). Non-parametric circadian variables, including interdaily stability (IS), interdaily variability (IV), L5, L5 start-time, M10, M10 start-time, amplitude, and relative amplitude, were calculated in R (v. 4.1.21) using the nParACT package.
Circular statistical analyses were performed to graph and analyze non-linear 24h periodic variables expressed as Zeitgeber time (ZT), including acrophase, onset ZT, offset ZT, L5 start-time ZT and M10 start-time ZT). These analyses were conducted in R (v. 4.1.21) using the circular package. Group comparisons were performed using Watson’s two-sample test of homogeneity, while Rayleigh’s test of uniformity was used to assess significant unimodal orientation within groups. In circular plots, individual points represent data from individual mice, and arrows indicate the circular mean. A p-value < 0.05 was considered statistically significant.
For gene expression, linear circadian variables, and OVA challenge experiments, comparisons intragroup by two time points were performed using an independent samples t-test and between groups a two-way ANOVA were performed. In mRNA gene expression analyses, intragroup differences between ZT0 and ZT10 were evaluated, and intergroup differences at each ZT were compared using the corresponding LD group as control. Data normality was assessed using the Shapiro-Wilk normality test, and datasets that did not meet normality were analyzed using the Mann-Whitney U test and represented as box and whisker plots.
In all graphs, individual points represent individual mice, and bars indicate mean ± SEM. Statistical significance was defined as p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1.
Author Contributions
Conceptualization, Carlos A Trujillo and Jose Sarmiento; Methodology, Fernando Miranda and Jose Sarmiento; Validation, Carlos A Trujillo and Fernando Miranda; Formal analysis, Fernando Miranda and Jose Sarmiento; Investigation, Carlos A Trujillo and Jose Sarmiento; Resources, Carlos A Trujillo and Jose Sarmiento; Data curation, Fernando Miranda; Writing—original draft, Carlos A Trujillo; Writing—review & editing, Carlos A Trujillo; Supervision, Jose Sarmiento; Project administration, Jose Sarmiento; Funding acquisition, Carlos A Trujillo and Jose Sarmiento. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Proyecto Fondecyt 1150789 (JS) of the Fondo Nacional de Ciencia y Tecnología (Chile). CAT is supported by an ANID BECAS/DOCTORADO NACIONAL PhD fellowship (##21192192).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of NAME OF INSTITUTE (protocol code XXX and date of approval).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors want to thank Dr. Marcelo A. Catalan and Mr. Felipe Tribiños for valuable discussions and grammar revision.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cinzano, P.; Falchi, F.; Elvidge, C. D. The first World Atlas of the artificial night sky brightness. Monthly Notices of the Royal Astronomical Society 2001, 328 (3), 689-707. [CrossRef]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C. C.; Elvidge, C. D.; Baugh, K.; Portnov, B. A.; Rybnikova, N. A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci Adv 2016, 2 (6), e1600377. [CrossRef]
- Erren, T. C.; Reiter, R. J. Defining chronodisruption. J Pineal Res 2009, 46 (3), 245-247. [CrossRef]
- Breymaier, S. O. B. AMA Adopts New Public Health Policies at Annual Meeting. AMA American Medical Association, 2012. http://news.cision.com/american-medical-association/r/ama-adopts-new-public-health-policies-at-annual-meeting,c9275938 (accessed 2012 june 20).
- Haspel, J. A.; Anafi, R.; Brown, M. K.; Cermakian, N.; Depner, C.; Desplats, P.; Gelman, A. E.; Haack, M.; Jelic, S.; Kim, B. S.; et al. Perfect timing: circadian rhythms, sleep, and immunity—an NIH workshop summary. JCI Insight 2020, 5 (1). [CrossRef]
- Inokawa, H.; Umemura, Y.; Shimba, A.; Kawakami, E.; Koike, N.; Tsuchiya, Y.; Ohashi, M.; Minami, Y.; Cui, G.; Asahi, T.; et al. Chronic circadian misalignment accelerates immune senescence and abbreviates lifespan in mice. Sci Rep 2020, 10 (1), 2569. [CrossRef]
- Lunn, R. M.; Blask, D. E.; Coogan, A. N.; Figueiro, M.; Gorman, M. R.; Hall, J. E.; Hansen, J.; Nelson, R. J.; Panda, S.; Smolensky, M. H.; et al. Health consequences of electric lighting practices in the modern world: A report on the National Toxicology Program’s workshop on shift work at night, artificial light at night, and circadian disruption. Science of the Total Environment 2017, 607, 1073-1084. [CrossRef]
- Russart, K. L. G.; Nelson, R. J. Light at night as an environmental endocrine disruptor. Physiol Behav 2017. [CrossRef]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sciences 2017, 173, 94-106. [CrossRef]
- Chang, A. M.; Aeschbach, D.; Duffy, J. F.; Czeisler, C. A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc Natl Acad Sci U S A 2015, 112 (4), 1232-1237. [CrossRef]
- Chen, R.; Weitzner, A. S.; McKennon, L. A.; Fonken, L. K. Light at night during development in mice has modest effects on adulthood behavior and neuroimmune activation. Behav Brain Res 2021, 405, 113171. [CrossRef]
- Kyba, C. C. M.; Tong, K. P.; Bennie, J.; Birriel, I.; Birriel, J. J.; Cool, A.; Danielsen, A.; Davies, T. W.; den Outer, P. N.; Edwards, W.; et al. Worldwide variations in artificial skyglow. Scientific Reports 2015, 5, ARTN 8409. [CrossRef]
- Phillips, D. J.; Savenkova, M. I.; Karatsoreos, I. N. Environmental disruption of the circadian clock leads to altered sleep and immune responses in mouse. Brain Behavior and Immunity 2015, 47, 14-23. [CrossRef]
- Carmona, P.; Perez, B.; Trujillo, C.; Espinosa, G.; Miranda, F.; Mendez, N.; Torres-Farfan, C.; Richter, H. G.; Vergara, K.; Brebi, P.; et al. Long-Term Effects of Altered Photoperiod During Pregnancy on Liver Gene Expression of the Progeny. Front Physiol 2019, 10, 1377. [CrossRef]
- Mendez, N.; Halabi, D.; Spichiger, C.; Salazar, E. R.; Vergara, K.; Alonso-Vasquez, P.; Carmona, P.; Sarmiento, J. M.; Richter, H. G.; Seron-Ferre, M.; et al. Gestational Chronodisruption Impairs Circadian Physiology in Rat Male Offspring, Increasing the Risk of Chronic Disease. Endocrinology 2016, 157 (12), 4654-4668. [CrossRef]
- Mendez, N.; Torres-Farfan, C.; Salazar, E.; Bascur, P.; Bastidas, C.; Vergara, K.; Spichiger, C.; Halabi, D.; Vio, C. P.; Richter, H. G. Fetal Programming of Renal Dysfunction and High Blood Pressure by Chronodisruption. Frontiers in Endocrinology 2019, 10, 362, 10.3389/fendo.2019.00362.
- Salazar, E. R.; Richter, H. G.; Spichiger, C.; Mendez, N.; Halabi, D.; Vergara, K.; Alonso, I. P.; Corvalán, F. A.; Azpeleta, C.; Seron-Ferre, M.; et al. Gestational chronodisruption leads to persistent changes in the rat fetal and adult adrenal clock and function. The Journal of Physiology 2018, 596 (23), 5839-5857. (accessed 2020/09/22). [CrossRef]
- Torres-Farfan, C.; Mendez, N.; Ehrenfeld, P.; Seron-Ferre, M. In utero circadian changes; facing light pollution. Current Opinion in Physiology 2020, 13, 128-134. [CrossRef]
- Bedrosian, T. A.; Fonken, L. K.; Walton, J. C.; Nelson, R. J. Chronic exposure to dim light at night suppresses immune responses in Siberian hamsters. Biology Letters 2011, 7 (3), 468-471. [CrossRef]
- Cisse, Y. M.; Russart, K. L.; Nelson, R. J. Parental Exposure to Dim Light at Night Prior to Mating Alters Offspring Adaptive Immunity. Sci Rep 2017, 7, 45497. [CrossRef]
- Cisse, Y. M.; Russart, K.; Nelson, R. J. Exposure to dim light at night prior to conception attenuates offspring innate immune responses. PLoS One 2020, 15 (4), e0231140. [CrossRef]
- Cisse, Y. M.; Peng, J.; Nelson, R. J. Effects of Dim Light at Night on Food Intake and Body Mass in Developing Mice. Front Neurosci 2017, 11, 294. [CrossRef]
- Fonken, L. K.; Frank, M. G.; Kitt, M. M.; Barrientos, R. M.; Watkins, L. R.; Maier, S. F. Microglia inflammatory responses are controlled by an intrinsic circadian clock. Brain Behav Immun 2015, 45, 171-179. [CrossRef]
- Fonken, L. K.; Weil, Z. M.; Nelson, R. J. Mice exposed to dim light at night exaggerate inflammatory responses to lipopolysaccharide. Brain Behavior and Immunity 2013, 34, 159-163. [CrossRef]
- Fonken, L. K.; Nelson, R. J. Effects of light exposure at night during development. Current Opinion in Behavioral Sciences 2016, 7, 33-39. [CrossRef]
- Fonken, L. K.; Haim, A.; Nelson, R. J. Dim light at night increases immune function in Nile grass rats, a diurnal rodent. Chronobiol Int 2012, 29 (1), 26-34. [CrossRef]
- Fonken, L. K.; Lieberman, R. A.; Weil, Z. M.; Nelson, R. J. Dim light at night exaggerates weight gain and inflammation associated with a high-fat diet in male mice. Endocrinology 2013, 154 (10), 3817-3825. [CrossRef]
- Fonken, L. K.; Aubrecht, T. G.; Melendez-Fernandez, O. H.; Weil, Z. M.; Nelson, R. J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J Biol Rhythms 2013, 28 (4), 262-271. [CrossRef]
- Fonken, L. K.; Weil, Z. M.; Nelson, R. J. Dark Nights Reverse Metabolic Disruption Caused by Dim Light at Night. Obesity 2013, 21 (6), 1159-1164. [CrossRef]
- Sarmiento, J.; Pulgar, R.; Mandakovic, D.; Porras, O.; Flores, C. A.; Luco, D.; Trujillo, C. A.; Díaz-Esquivel, B.; Alvarez, C.; Acevedo, A.; et al. Nocturnal Light Pollution Induces Weight Gain in Mice and Reshapes the Structure, Functions, and Interactions of Their Colonic Microbiota. Int J Mol Sci 2022, 23 (3). [CrossRef]
- Shuboni, D.; Yan, L. Nighttime dim light exposure alters the responses of the circadian system. Neuroscience 2010, 170 (4), 1172-1178. [CrossRef]
- Bakkar, N. Z.; Dwaib, H. S.; Fares, S.; Eid, A. H.; Al-Dhaheri, Y.; El-Yazbi, A. F. Cardiac Autonomic Neuropathy: A Progressive Consequence of Chronic Low-Grade Inflammation in Type 2 Diabetes and Related Metabolic Disorders. Int J Mol Sci 2020, 21 (23). [CrossRef]
- Chen, J.; Yang, Y.; Kong, W. Cross Talk between Inflammation and Metabolic Disorders. Mediators Inflamm 2022, 2022, 9821506. [CrossRef]
- Hotamisligil, G. Inflammation and metabolic disorders. Nature 2006, 444 (7121), 860-867, Article. [CrossRef]
- Cermakian, N.; Lange, T.; Golombek, D.; Sarkar, D.; Nakao, A.; Shibata, S.; Mazzoccoli, G. Crosstalk between the circadian clock circuitry and the immune system. Chronobiol Int 2013, 30 (7), 870-888. [CrossRef]
- Druzd, D.; Matveeva, O.; Ince, L.; Harrison, U.; He, W.; Schmal, C.; Herzel, H.; Tsang, A. H.; Kawakami, N.; Leliavski, A.; et al. Lymphocyte Circadian Clocks Control Lymph Node Trafficking and Adaptive Immune Responses. Immunity 2017, 46 (1), 120-132. [CrossRef]
- Hergenhan, S.; Holtkamp, S.; Scheiermann, C. Molecular Interactions Between Components of the Circadian Clock and the Immune System. J Mol Biol 2020. [CrossRef]
- Scheiermann, C.; Gibbs, J.; Ince, L.; Loudon, A. Clocking in to immunity. Nat Rev Immunol 2018, 18 (7), 423-437. [CrossRef]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P. S. Circadian control of the immune system. Nat Rev Immunol 2013, 13 (3), 190-198. [CrossRef]
- Delorme, T. C.; Srivastava, L. K.; Cermakian, N. Altered circadian rhythms in a mouse model of neurodevelopmental disorders based on prenatal maternal immune activation. Brain Behav Immun 2021, 93, 119-131. [CrossRef]
- Hanson, M. A.; Gluckman, P. D. Early developmental conditioning of later health and disease: physiology or pathophysiology? Physiol Rev 2014, 94 (4), 1027-1076. [CrossRef]
- Russart, K. L. G.; Chbeir, S. A.; Nelson, R. J.; Magalang, U. J. Light at night exacerbates metabolic dysfunction in a polygenic mouse model of type 2 diabetes mellitus. Life Sci 2019, 231, 116574. [CrossRef]
- Varcoe, T. J.; Gatford, K. L.; Kennaway, D. J. Maternal circadian rhythms and the programming of adult health and disease. Am J Physiol Regul Integr Comp Physiol 2018, 314 (2), R231-R241. [CrossRef]
- Varcoe, T. J.; Voultsios, A.; Gatford, K. L.; Kennaway, D. J. The impact of prenatal circadian rhythm disruption on pregnancy outcomes and long-term metabolic health of mice progeny. Chronobiol Int 2016, 33 (9), 1171-1181. [CrossRef]
- Acosta-Rodríguez, V.; Rijo-Ferreira, F.; Izumo, M.; Xu, P.; Wight-Carter, M.; Green, C. B.; Takahashi, J. S. Circadian alignment of early onset caloric restriction promotes longevity in male C57BL/6J mice. Science 2022, 376 (6598), 1192-1202. [CrossRef]
- Carter, S. J.; Durrington, H. J.; Gibbs, J. E.; Blaikley, J.; Loudon, A. S.; Ray, D. W.; Sabroe, I. A matter of time: study of circadian clocks and their role in inflammation. J Leukoc Biol 2016, 99 (4), 549-560. [CrossRef]
- Patterson, R. E.; Laughlin, G. A.; LaCroix, A. Z.; Hartman, S. J.; Natarajan, L.; Senger, C. M.; Martinez, M. E.; Villasenor, A.; Sears, D. D.; Marinac, C. R.; et al. Intermittent Fasting and Human Metabolic Health. J Acad Nutr Diet 2015, 115 (8), 1203-1212. [CrossRef]
- Li, X.; Rothämel, P.; Nussbaum, C.; Sperandio, M.; Scheiermann, C. Development of a circadian immune system. Trends Immunol 2025, 46 (9), 614-623. [CrossRef]
- Lewis, S. M.; Williams, A.; Eisenbarth, S. C. Structure and function of the immune system in the spleen. Sci Immunol 2019, 4 (33). [CrossRef]
- Wluka, A.; Olszewski, W. L. Innate and adaptive processes in the spleen. Ann Transplant 2006, 11 (4), 22-29.
- Bellinger, D. L.; Lorton, D. Sympathetic Nerve Hyperactivity in the Spleen: Causal for Nonpathogenic-Driven Chronic Immune-Mediated Inflammatory Diseases (IMIDs)? Int J Mol Sci 2018, 19 (4). [CrossRef]
- Lori, A.; Perrotta, M.; Lembo, G.; Carnevale, D. The Spleen: A Hub Connecting Nervous and Immune Systems in Cardiovascular and Metabolic Diseases. Int J Mol Sci 2017, 18 (6). [CrossRef]
- Tarantino, G.; Savastano, S.; Capone, D.; Colao, A. Spleen: A new role for an old player? World J Gastroenterol 2011, 17 (33), 3776-3784. [CrossRef]
- Ackerman, K. D.; Felten, S. Y.; Bellinger, D. L.; Felten, D. L. Noradrenergic sympathetic innervation of the spleen: III. Development of innervation in the rat spleen. J Neurosci Res 1987, 18 (1), 49-54, 123-125. [CrossRef]
- Bellinger, D. L.; Felten, S. Y.; Collier, T. J.; Felten, D. L. Noradrenergic sympathetic innervation of the spleen: IV. Morphometric analysis in adult and aged F344 rats. J Neurosci Res 1987, 18 (1), 55-63, 126-129. [CrossRef]
- Bellinger, D. L.; Felten, S. Y.; Lorton, D.; Felten, D. L. Origin of noradrenergic innervation of the spleen in rats. Brain Behav Immun 1989, 3 (4), 291-311. [CrossRef]
- Carlson, S. L.; Felten, D. L.; Livnat, S.; Felten, S. Y. Noradrenergic sympathetic innervation of the spleen: V. Acute drug-induced depletion of lymphocytes in the target fields of innervation results in redistribution of noradrenergic fibers but maintenance of compartmentation. J Neurosci Res 1987, 18 (1), 64-69, 130-131. [CrossRef]
- Felten, D. L.; Ackerman, K. D.; Wiegand, S. J.; Felten, S. Y. Noradrenergic sympathetic innervation of the spleen: I. Nerve fibers associate with lymphocytes and macrophages in specific compartments of the splenic white pulp. J Neurosci Res 1987, 18 (1), 28-36, 118-121. [CrossRef]
- Felten, S. Y.; Olschowka, J. Noradrenergic sympathetic innervation of the spleen: II. Tyrosine hydroxylase (TH)-positive nerve terminals form synapticlike contacts on lymphocytes in the splenic white pulp. J Neurosci Res 1987, 18 (1), 37-48. [CrossRef]
- Ding, J.; Chen, P.; Qi, C. Circadian rhythm regulation in the immune system. Immunology 2024, 171 (4), 525-533 (accessed 2024/09/03). [CrossRef]
- Arjona, A.; Boyadjieva, N.; Sarkar, D. K. Circadian rhythms of granzyme B, perforin, IFN-gamma, and NK cell cytolytic activity in the spleen: effects of chronic ethanol. J Immunol 2004, 172 (5), 2811-2817.
- Arjona, A.; Sarkar, D. K. Circadian oscillations of clock genes, cytolytic factors, and cytokines in rat NK cells. J Immunol 2005, 174 (12), 7618-7624.
- Arjona, A.; Sarkar, D. K. The circadian gene mPer2 regulates the daily rhythm of IFN-gamma. J Interferon Cytokine Res 2006, 26 (9), 645-649. [CrossRef]
- Silver, A. C.; Arjona, A.; Walker, W. E.; Fikrig, E. The circadian clock controls toll-like receptor 9-mediated innate and adaptive immunity. Immunity 2012, 36 (2), 251-261. [CrossRef]
- Cordoba-Moreno, M. O.; de Souza, E. D. S.; Quiles, C. L.; Dos Santos-Silva, D.; Kinker, G. S.; Muxel, S. M.; Markus, R. P.; Fernandes, P. A. Rhythmic expression of the melatonergic biosynthetic pathway and its differential modulation in vitro by LPS and IL10 in bone marrow and spleen. Sci Rep 2020, 10 (1), 4799. [CrossRef]
- Calogiuri, G.; Weydahl, A.; Carandente, F. Methodological issues for studying the rest-activity cycle and sleep disturbances: a chronobiological approach using actigraphy data. Biol Res Nurs 2013, 15 (1), 5-12. [CrossRef]
- Schmid, B.; Helfrich-Förster, C.; Yoshii, T. A new ImageJ plug-in “ActogramJ” for chronobiological analyses. J Biol Rhythms 2011, 26 (5), 464-467. [CrossRef]
- Blume, C.; Santhi, N.; Schabus, M. ‘nparACT’ package for R: A free software tool for the non-parametric analysis of actigraphy data. MethodsX 2016, 3, 430-435. [CrossRef]
- Cremers, J.; Klugkist, I. One Direction? A Tutorial for Circular Data Analysis Using R With Examples in Cognitive Psychology. Frontiers in Psychology 2018, 9, Methods. [CrossRef]
- Jud, C.; Schmutz, I.; Hampp, G.; Oster, H.; Albrecht, U. A guideline for analyzing circadian wheel-running behavior in rodents under different lighting conditions. Biol Proced Online 2005, 7, 101-116. [CrossRef]
- Silver, A. C.; Arjona, A.; Hughes, M. E.; Nitabach, M. N.; Fikrig, E. Circadian expression of clock genes in mouse macrophages, dendritic cells, and B cells. Brain Behav Immun 2012, 26 (3), 407-413. [CrossRef]
- Hayashi, M.; Shimba, S.; Tezuka, M. Characterization of the molecular clock in mouse peritoneal macrophages. Biol Pharm Bull 2007, 30 (4), 621-626.
- Kiessling, S.; Dubeau-Laramee, G.; Ohm, H.; Labrecque, N.; Olivier, M.; Cermakian, N. The circadian clock in immune cells controls the magnitude of Leishmania parasite infection. Sci Rep 2017, 7 (1), 10892. [CrossRef]
- Livak, K. J.; Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25 (4), 402-408. [CrossRef]
- Schmittgen, T. D.; Livak, K. J. Analyzing real-time PCR data by the comparative CT method. Nature Protocols 2008, 3 (6), 1101-1108. [CrossRef]
- Czerkinsky, C. C.; Nilsson, L. A.; Nygren, H.; Ouchterlony, O.; Tarkowski, A. A solid-phase enzyme-linked immunospot (ELISPOT) assay for enumeration of specific antibody-secreting cells. J Immunol Methods 1983, 65 (1-2), 109-121.
- Valentinuzzi, V. S.; Scarbrough, K.; Takahashi, J. S.; Turek, F. W. Effects of aging on the circadian rhythm of wheel-running activity in C57BL/6 mice. Am J Physiol 1997, 273 (6), R1957-1964. [CrossRef]
- Panagiotou, M.; Deboer, T. Effects of Chronic Dim-light-at-night Exposure on Sleep in Young and Aged Mice. Neuroscience 2020, 426, 154-167. [CrossRef]
- Stowie, A. C.; Glass, J. D. Longitudinal Study of Changes in Daily Activity Rhythms over the Lifespan in Individual Male and Female C57BL/6J Mice. Journal of Biological Rhythms 2015, 30 (6), 563-568. [CrossRef]
- Sekaran, S.; Lupi, D.; Jones, S.; Sheely, C.; Hattar, S.; Yau, K.-W.; Lucas, R.; Foster, R.; Hankins, M. Melanopsin-dependent photoreception provides earliest light detection in the mammalian retina. Current Biology 2005, 15 (12), 1099-1107.
- Schmidt, T. M.; Taniguchi, K.; Kofuji, P. Intrinsic and extrinsic light responses in melanopsin-expressing ganglion cells during mouse development. J Neurophysiol 2008, 100 (1), 371-384. [CrossRef]
- Keller, M.; Mazuch, J.; Abraham, U.; Eom, G. D.; Herzog, E. D.; Volk, H. D.; Kramer, A.; Maier, B. A circadian clock in macrophages controls inflammatory immune responses. Proc Natl Acad Sci U S A 2009, 106 (50), 21407-21412. [CrossRef]
- Bedrosian, T. A.; Aubrecht, T. G.; Kaugars, K. E.; Weil, Z. M.; Nelson, R. J. Artificial light at night alters delayed-type hypersensitivity reaction in response to acute stress in Siberian hamsters. Brain Behavior and Immunity 2013, 34, 39-42. [CrossRef]
- Bedrosian, T. A.; Fonken, L. K.; Nelson, R. J. Endocrine Effects of Circadian Disruption. Annu Rev Physiol 2016, 78, 109-131. [CrossRef]
- Bedrosian, T. A.; Galan, A.; Vaughn, C. A.; Weil, Z. M.; Nelson, R. J. Light at Night Alters Daily Patterns of Cortisol and Clock Proteins in Female Siberian Hamsters. Journal of Neuroendocrinology 2013, 25 (6), 590-596. [CrossRef]
- Fonken, L. K.; Kitsmiller, E.; Smale, L.; Nelson, R. J. Dim nighttime light impairs cognition and provokes depressive-like responses in a diurnal rodent. J Biol Rhythms 2012, 27 (4), 319-327. [CrossRef]
- Fonken, L. K.; Weber, M. D.; Daut, R. A.; Kitt, M. M.; Frank, M. G.; Watkins, L. R.; Maier, S. F. Stress-induced neuroinflammatory priming is time of day dependent. Psychoneuroendocrinology 2016, 66, 82-90. [CrossRef]
- Belpiede, L. T.; Gomes, P. R. L.; Gomes, G.; Mendes, C.; da Cunha, F. M.; Macedo, F.; Motta, L. C.; Teixeira, S. S. J.; Amaral, F. G.; Neto, J. C. Transgenerational effects of maternal melatonin deficiency and melatonin replacement in rats during pregnancy and lactation on the energy metabolism and thermoregulation in the offspring subjected to a high-fat diet.
- Kennaway, D. J.; Voultsios, A.; Varcoe, T. J.; Moyer, R. W. Melatonin in mice: rhythms, response to light, adrenergic stimulation, and metabolism. Am J Physiol Regul Integr Comp Physiol 2002, 282 (2), R358-365. [CrossRef]
- Roseboom, P. H.; Namboodiri, M. A.; Zimonjic, D. B.; Popescu, N. C.; Rodriguez, I. R.; Gastel, J. A.; Klein, D. C. Natural melatonin ‘knockdown’ in C57BL/6J mice: rare mechanism truncates serotonin N-acetyltransferase. Brain Res Mol Brain Res 1998, 63 (1), 189-197.
- Aschoff, J. Freerunning and Entrained Circadian Rhythms. In Biological Rhythms, Aschoff, J. Ed.; Springer US, 1981; pp 81-93.
- Takahashi, J. S.; Zatz, M. Regulation of circadian rhythmicity. Science 1982, 217 (4565), 1104-1111.
- Honma, S. Development of the mammalian circadian clock. Eur J Neurosci 2020, 51 (1), 182-193. [CrossRef]
- HONMA, S.; HONMA, K.-I.; Shirakawa, T.; HIROSHIGE, T. Maternal phase setting of fetal circadian oscillation underlying the plasma corticosterone rhythm in rats. Endocrinology 1984, 114 (5), 1791-1796.
- Honma, K.; Honma, S.; Shirakawa, T.; Hiroshige, T. Phase setting of circadian locomotor rhythm of infant rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1987, 252 (2), R256-R261.
- Takahashi, K.; Deguchi, T. Entrainment of the circadian rhythms of blinded infant rats by nursing mothers. Physiology & behavior 1983, 31 (3), 373-378.
- Honma, S.; Honma, K.; Hiroshige, T. Restricted daily feeding during nursing period resets circadian locomotor rhythm of infant rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1987, 252 (2), R262-R268.
- Spichiger, C.; Torres-Farfan, C.; Galdames, H. A.; Mendez, N.; Alonso-Vazquez, P.; Richter, H. G. Gestation under chronic constant light leads to extensive gene expression changes in the fetal rat liver. Physiol Genomics 2015, 47 (12), 621-633. [CrossRef]
- Méndez, N.; Corvalan, F.; Halabi, D.; Vasquez, A.; Vergara, K.; Noriega, H.; Ehrenfeld, P.; Sanhueza, K.; Seron-Ferre, M.; Valenzuela, G. J.; et al. Sex-Specific Metabolic Effects of Gestational Chronodisruption and Maternal Melatonin Supplementation in Rat Offspring. Journal of Pineal Research 2024, 76 (8), e70015. [CrossRef]
- Pennell, C. Approaches to evaluating gene-environment interactions underlying DOHaD. Early Human Development 2007, 83, S36-S36. [CrossRef]
- Cornélissen, G. Circadian disruption and metabolic disease risk. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases, Elsevier, 2022; pp 509-516.
- Curioni, C. C.; Silva, A. C. F. d.; Silva Pereira, A. d.; Mocellin, M. C. The Role of Dietary Habits on Development and Progress of Risk Factors of Chronic Non-communicable Diseases. In Healthy Lifestyle, Springer, 2022; pp 105-129.
- Gillman, M. W. Developmental origins of health and disease. The New England journal of medicine 2005, 353 (17), 1848.
- Delerive, P.; Monte, D.; Dubois, G.; Trottein, F.; Fruchart-Najib, J.; Mariani, J.; Fruchart, J. C.; Staels, B. The orphan nuclear receptor ROR alpha is a negative regulator of the inflammatory response. Embo Reports 2001, 2 (1), 42-48. [CrossRef]
- Kopmels, B.; Mariani, J.; Delhaye-Bouchaud, N.; Audibert, F.; Fradelizi, D.; Wollman, E. E. Evidence for a hyperexcitability state of staggerer mutant mice macrophages. J Neurochem 1992, 58 (1), 192-199. [CrossRef]
- Downton, P.; Early, J. O.; Gibbs, J. E. Circadian rhythms in adaptive immunity. Immunology 2019. [CrossRef]
- Hotamisligil, G. S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542 (7640), 177-185. [CrossRef]
- Hotamisligil, G. S. Inflammation and metabolic disorders. Nature 2006, 444 (7121), 860-867. [CrossRef]
- Gibbs, J. E.; Blaikley, J.; Beesley, S.; Matthews, L.; Simpson, K. D.; Boyce, S. H.; Farrow, S. N.; Else, K. J.; Singh, D.; Ray, D. W.; et al. The nuclear receptor REV-ERBα mediates circadian regulation of innate immunity through selective regulation of inflammatory cytokines. Proceedings of the National Academy of Sciences 2012, 109 (2), 582-587.
- Sato, S.; Sakurai, T.; Ogasawara, J.; Takahashi, M.; Izawa, T.; Imaizumi, K.; Taniguchi, N.; Ohno, H.; Kizaki, T. A circadian clock gene, Rev-erbalpha, modulates the inflammatory function of macrophages through the negative regulation of Ccl2 expression. J Immunol 2014, 192 (1), 407-417. [CrossRef]
- Curtis, A. M.; Bellet, M. M.; Sassone-Corsi, P.; O’Neill, L. A. Circadian clock proteins and immunity. Immunity 2014, 40 (2), 178-186. [CrossRef]
- Abreu, M.; Basti, A.; Genov, N.; Mazzoccoli, G.; Relógio, A. The reciprocal interplay between TNFα and the circadian clock impacts on cell proliferation and migration in Hodgkin lymphoma cells. Scientific Reports 2018, 8 (1), 11474.
- Bellet, M. M.; Deriu, E.; Liu, J. Z.; Grimaldi, B.; Blaschitz, C.; Zeller, M.; Edwards, R. A.; Sahar, S.; Dandekar, S.; Baldi, P.; et al. Circadian clock regulates the host response to Salmonella. Proc Natl Acad Sci U S A 2013, 110 (24), 9897-9902. [CrossRef]
Figure 8.
Relative mRNA expression levels of clock genes in the spleen. Relative mRNA expression levels of clock genes in the spleen were determined by qPCR and are presented as box-and-whisker plots (minimum to maximum). Panels show the expression of (A) Bmal1, (B) Clock, (C) Per2, (D) Nr1d1, and (E) Rorα. Experimental groups are represented as follows: LD (control, light–dark cycle) as black circles, PdLAN (postnatal dim light at night) as grey triangles, and GdLAN (gestational dim light at night) as black stars. Intra-group daily differences between Zeitgeber times (ZT) are indicated by connecting lines: dashed black lines for LD, grey dashed lines for PdLAN, and black dotted lines for GdLAN. Intergroup differences are indicated by solid lines and asterisks, with grey denoting comparisons between PdLAN vs. LD and black denoting comparisons between GdLAN vs. LD. Relative expression levels at each ZT were calculated using the 2−ΔΔCT method. The average of ZT0 of the LD group was used as the reference time point (Livak and Schmittgen, 2001; Schmittgen and Livak, 2008). n=9, except for Bmal1 and Rorα, where n=5-9. Values are in box and whiskers min to max. Two-way ANOVA and Sidak’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001.
Figure 8.
Relative mRNA expression levels of clock genes in the spleen. Relative mRNA expression levels of clock genes in the spleen were determined by qPCR and are presented as box-and-whisker plots (minimum to maximum). Panels show the expression of (A) Bmal1, (B) Clock, (C) Per2, (D) Nr1d1, and (E) Rorα. Experimental groups are represented as follows: LD (control, light–dark cycle) as black circles, PdLAN (postnatal dim light at night) as grey triangles, and GdLAN (gestational dim light at night) as black stars. Intra-group daily differences between Zeitgeber times (ZT) are indicated by connecting lines: dashed black lines for LD, grey dashed lines for PdLAN, and black dotted lines for GdLAN. Intergroup differences are indicated by solid lines and asterisks, with grey denoting comparisons between PdLAN vs. LD and black denoting comparisons between GdLAN vs. LD. Relative expression levels at each ZT were calculated using the 2−ΔΔCT method. The average of ZT0 of the LD group was used as the reference time point (Livak and Schmittgen, 2001; Schmittgen and Livak, 2008). n=9, except for Bmal1 and Rorα, where n=5-9. Values are in box and whiskers min to max. Two-way ANOVA and Sidak’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001.

Figure 9.
Relative mRNA expression levels of inflammatory genes in the spleen. Relative mRNA expression levels of inflammatory genes in the spleen were determined by qPCR and are presented as box-and-whisker plots (minimum to maximum). Panels show the expression of (A) Mcp-1, (B) Tnf-α, (C) Tgf-β, (D) Il-6, and (E) Il-10. Experimental groups are represented as follows: LD (control, light–dark cycle) as black circles, PdLAN (postnatal dim light at night) as grey triangles, and GdLAN (gestational dim light at night) as black stars. Intra-group daily differences between ZT are indicated by connecting lines: dashed black lines for LD, grey dashed lines for PdLAN, and black dotted lines for GdLAN. Intergroup differences are indicated by solid lines and asterisks, with grey denoting comparisons between PdLAN vs. LD and black denoting comparisons between GdLAN vs. LD. Relative expression levels at each ZT were calculated using the 2-ΔΔCT method. The average of ZT0 of the LD group was used as the reference time point (Livak and Schmittgen, 2001; Schmittgen and Livak, 2008). Sample size was n = 9 per group, except for Il-10, where n = 7–9. Two-way ANOVA and Sidak’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001.
Figure 9.
Relative mRNA expression levels of inflammatory genes in the spleen. Relative mRNA expression levels of inflammatory genes in the spleen were determined by qPCR and are presented as box-and-whisker plots (minimum to maximum). Panels show the expression of (A) Mcp-1, (B) Tnf-α, (C) Tgf-β, (D) Il-6, and (E) Il-10. Experimental groups are represented as follows: LD (control, light–dark cycle) as black circles, PdLAN (postnatal dim light at night) as grey triangles, and GdLAN (gestational dim light at night) as black stars. Intra-group daily differences between ZT are indicated by connecting lines: dashed black lines for LD, grey dashed lines for PdLAN, and black dotted lines for GdLAN. Intergroup differences are indicated by solid lines and asterisks, with grey denoting comparisons between PdLAN vs. LD and black denoting comparisons between GdLAN vs. LD. Relative expression levels at each ZT were calculated using the 2-ΔΔCT method. The average of ZT0 of the LD group was used as the reference time point (Livak and Schmittgen, 2001; Schmittgen and Livak, 2008). Sample size was n = 9 per group, except for Il-10, where n = 7–9. Two-way ANOVA and Sidak’s multiple comparisons post-hoc test. *p<0.05 and ** p<0.01 ***p<0.001.

Figure 10.
Effect of dLAN on splenocyte recruitment and anti-OVA IgG/IgE responses following OVA challenge. Mice were challenged with ovalbumin (OVA) to evaluate immune responses under different lighting conditions. At postnatal day 53 (P53) and ZT6, mice received an intraperitoneal injection of 100 μg OVA (day 0). After 7 days, animals received a second injection as a challenge. Five days later, samples were collected for analysis. A. Total number of splenocytes recovered from the spleen. Both PdLAN and GdLAN groups showed increased cell numbers compared with the LD control group. Cell counts were determined using a Neubauer chamber (n = 9). B. ELISpot assay for anti-OVA IgG–secreting cells (n = 9). C. ELISpot assay for anti-OVA IgE–secreting cells, presented as mean ± SEM (n = 7–9). A two-tailed unpaired t test was used vs LD control group. Asterisk in grey when the differences are between PdLAN vs LD control group. Asterisk in black when the differences are between GdLAN and LD. *p<0.05, ** p<0.01, ***p<0.001.
Figure 10.
Effect of dLAN on splenocyte recruitment and anti-OVA IgG/IgE responses following OVA challenge. Mice were challenged with ovalbumin (OVA) to evaluate immune responses under different lighting conditions. At postnatal day 53 (P53) and ZT6, mice received an intraperitoneal injection of 100 μg OVA (day 0). After 7 days, animals received a second injection as a challenge. Five days later, samples were collected for analysis. A. Total number of splenocytes recovered from the spleen. Both PdLAN and GdLAN groups showed increased cell numbers compared with the LD control group. Cell counts were determined using a Neubauer chamber (n = 9). B. ELISpot assay for anti-OVA IgG–secreting cells (n = 9). C. ELISpot assay for anti-OVA IgE–secreting cells, presented as mean ± SEM (n = 7–9). A two-tailed unpaired t test was used vs LD control group. Asterisk in grey when the differences are between PdLAN vs LD control group. Asterisk in black when the differences are between GdLAN and LD. *p<0.05, ** p<0.01, ***p<0.001.


Table 1.
Summary of alpha p-values from intra-group comparison.
|
Multiple Comparison |
LD | PdLAN | GdLAN |
| S1 VS S2 | ns | 0.0179* | ns |
| S1 VS S3 | ns | ns | ns |
| S1 VS S4 | ns | 0.0097** | ns |
| S1 VS S5 | ns | 0.0313* | ns |
| S1 VS S6 | 0.0195* | ns | ns |
| S2 VS S3 | ns | ns | ns |
| S2 VS S4 | ns | ns | ns |
| S2 VS S5 | 0.0276* | 0.0042** | ns |
| S2 VS S6 | 0.0088** | 0.0072** | ns |
| S3 VS S4 | ns | ns | ns |
| S3 VS S5 | 0.0372* | 0.0355* | ns |
| S3 VS S6 | 0.0104* | ns | ns |
| S4 VS S5 | 0.0025** | 0.0077** | ns |
| S4 VS S6 | 0.0033** | 0.0318* | ns |
| S5 VS S6 | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



