Submitted:
13 March 2026
Posted:
16 March 2026
You are already at the latest version
Abstract
Genome editing is widely used and conceptually simple, yet in practice, it is hindered by laborious workflows and high costs. These challenges stem from the difficulty of identifying and isolating cells that contain the desired user‑defined modifications, a problem compounded by the wide variability in editing efficiencies across cell types. While homology-directed repair (HDR) provides a mechanism for precise genome modification following nuclease-induced double-strand breaks (DSBs), it is frequently outcompeted by the dominant mutagenic non-homologous end-joining (NHEJ) pathway in mammalian cells. Therefore, we developed a novel enrichment method, Essential HDRescue, to increase the frequency of HDR events at a target site by co-targeting an essential genomic locus. Using both intrinsic positive and negative selection at a common essential gene, we enabled enrichment of precise editing events at a second, unlinked target site. We demonstrated that co-targeting essential genes in cancer cell lines and iPSCs increased HDR rates without the need for an exogenous reporter or selective drug. Analysis of resulting clones revealed that Essential HDRescue produced up to a 6‑fold increase in single‑allele edits and almost a four‑fold increase in homozygous edits relative to single‑targeted controls. By harnessing the intrinsic cellular dependencies that arise from DSB repair at essential loci, Essential HDRescue offers a widely applicable method to improve precise genome editing outcomes in mammalian cells, leaving only a minimal, protein-silent scar at the essential gene.
Keywords:
HDR
; co-targeting
; cell line development
; CRISPR
; essential genes
; genome engineering
1. Introduction
CRISPR technologies play a crucial role in many biological applications, including cell modeling, gene and cell therapeutics, and functional genomics[1,2]. CRISPR can be used to engineer mutations at a target site by introducing programmed double-stranded breaks (DSBs) into the genome, which are generally repaired by one of two predominant DNA repair pathways[3] . The first and most active pathway in mammalian cells, non-homologous end joining (NHEJ), is considered error-prone because it frequently introduces insertion or deletion (indel) mutations and is commonly used to generate gene knockouts (KO)[4]. In contrast, the second pathway, homology-directed repair (HDR), can install precise, user-defined mutations, such as gene fragment insertions or single-nucleotide variants, at a target gene or locus of interest using a homologous DNA template[4,5,6]. However, HDR occurs less often than NHEJ in mammalian somatic cells, making the development of engineered cells for both basic and preclinical research more labor-intensive and costly. In the absence of a reporter, HDR-edited cells typically appear phenotypically indistinguishable from NHEJ- or non-edited cells, making targeted cell identification challenging and further limiting the practical application of genome engineering[7]. Numerous strategies can enhance HDR by using small molecules to either suppress NHEJ activity or arrest cells in G2/M phase, when homologous recombination genes are expressed[8]. However, some of these drugs cause chromosomal instability, necessitating further investigation of aberrant DSB repair by a site-directed nuclease[9]. Newer genome editing tools such as base editors and prime editors avoid generating DSBs, offering alternatives to HDR- and NHEJ-based editing altogether[10,11,12]. However, each technique has its own limitations. Base editors are restricted to making specific point mutations and can introduce unwanted bystander edits when nearby editable bases fall within the editing window[13]. Prime editors offer greater flexibility but often show low efficiencies across many cell types and require extensive optimization of prime editing guide RNA design, nicking strategies, and delivery conditions[14]. These constraints limit their broad applicability despite their potential advantages. Given the potential impact of CRISPR technologies across biological fields, it is vital to continue developing novel approaches that produce precisely edited cells while improving upon current efficiency rates.
A variety of selection strategies have been developed to increase the representation of edited cells within a larger population[15]. The most common and conceptually straightforward approach is to directly couple the desired edit to a genetic reporter, such as a fluorescent protein or antibiotic-resistance gene, enabling edited cells to be readily distinguished from unedited ones[16,17,18]. This approach generally introduces a transgene, which is not always desirable and can limit the number of possible edits if available selection markers are exhausted. In contrast, co-targeting represents a conceptually distinct strategy in which a gain-of-function mutation at one locus serves as the selectable event, thereby enriching for phenotypically silent edits at a second, unlinked site. Because editing outcomes at two independent loci are not statistically independent, simultaneous targeting can enrich for double-editing events[19,20,21,22]. Such co-targeting strategies have been adopted in model organisms such as Caenorhabditis elegans and Drosophila, as well as in mammalian cells[23,24,25,26,27]. Some co-targeting strategies use cytotoxic agents, such as drugs, toxins, or antibiotics, to impose positive or negative selection at the reporter locus[21,28,29,30,31,32,33]. Others require physical separation of cells using specialized equipment, such as fluorescence-activated cell sorting or magnetic separation[30,34,35]. Still other strategies have required the insertion of whole transgenes into an exon of a gene required for cell viability[36]. This approach restricts the knock-in (KI) to a single copy DNA, which is limiting when multiple variants or isoforms of a particular gene may exist naturally. It further eliminates endogenous expression control and disrupts other regulatory features of the native target locus. We therefore aimed to develop a universal co-selection method that: (1) does not require an exogenous selection marker, drug, or specialized equipment to enhance editing outcomes; (2) removes imprecisely edited cells in a cell-intrinsic manner; (3) introduces only synonymous changes at the selection locus, leaving the protein sequence unaltered; and (4) preserves natural regulatory control of the target locus.
To develop such a method, we turned to essential genes. Essential genes are required for cellular proliferation under normal environmental conditions and are considered intolerant to loss-of-function mutations. Among human cells, essential genes are indispensable for various cell-based processes, including cell cycle progression, nuclear trafficking, and metabolic processing[37]. A comprehensive list of pan-essential genes has been identified in both cancer cell lines and stem cells[38,39,40], which served as a basis for identifying candidate essential selection sites in our work.
Our co-selection strategy, hereafter referred to as HDRescue, uses essential genes as both positive and negative selection loci via HDR and NHEJ, respectively. Consequently, cell intrinsic selection for the primary, essential gene modification enriches the accompanying and desired passenger modification(s). Specifically, our approach installs synonymous blocking mutations within the protospacer and/or protospacer-adjacent motif (PAM) of the essential genes RAN and SF3B1 using CRISPR-Cas9 and an HDRescue donor. Negative selection occurs when NHEJ-mediated out-of-frame indel mutations at the essential site knock-out the gene, leading to cell death. However, cells that repair the DSB via HDR using the rescue donor undergo positive selection, leading to a substantial increase in editing at the second, desired target site.
Using HDRescue, we observed bulk KI efficiencies of up to 46% in induced pluripotent stem cells (iPSCs) and a substantial increase in the number of homozygously edited clonal lines. We further found that we can achieve a meaningful increase in KI mutations up to 800 bp. Additionally, we observed a minimal number of unwanted edits. Collectively, this work demonstrates that HDRescue is a robust, universal co-targeting strategy that will be widely applicable across cell lines to improve on-target editing while preserving the natural elements of the target locus.
2. Methods
2.1. Cell Culture and Reagents
BJFF.6 iPSCs (Cellosaurus, CVCL-VU02[41]), AN1.1 iPSCs (Washington University, St. Louis, MO, USA; T-019532) were cultured in StemFlex media (Gibco, Carlsbad, CA, USA), supplemented with 1% (vol/vol) antibiotic/antimycotic (Gibco) and 5% (vol/vol) CO2 in a humidified incubator at 37°C. Edited iPSC pools were generated using CRISPR-Cas9 technology and homologous DNA donors. One million iPSCs were nucleofected (4D-Nucleofector™ X-unit; Lonza, Basel, Switzerland) per the manufacturer’s recommended protocol with pre-complexed RNPs consisting of 100 pmol chemically modified sgRNA (Integrated DNA Technologies, Coralville, IA, USA) (Table S1) and 33 pmol 3XNLS SpCas9 protein (St. Jude Children’s Research Hospital [SJCRH] Protein Production Facility, Memphis, TN, USA) using P3 nucleofector solution and CA-137 program in 20-µL or 100-µL cuvettes. Control cell pools contained a non-targeting sgRNA and a non-targeting DNA donor to control for RNP and donor concentrations added to an HDRescue editing experiment. Unless otherwise noted, editing rates were assessed at day 10 post-transfection.
U2OS cells (ATCC, HTB-96) were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% (vol/vol) FBEssence, 1% (vol/vol) penicillin-streptomycin, and 1X GlutaMAX at 37°C, 5% vol/vol CO2 in a humidified incubator. U2OS cell pools were generated by nucleofecting 500,000 cells as described above using P3 nucleofector solution (Lonza) and the CM-104 program in 20-µL cuvettes. Unless otherwise notes, editing rates were assessed at day 28 post-transfection.
HeLa cells (ATCC, CRM-CCL-2) were cultured in DMEM supplemented with 10% (vol/vol) FBEssence, 1% (vol/vol) penicillin-streptomycin, and 1X GlutaMAX at 37°C, 5% vol/vol CO2 in a humidified incubator. HeLa cells pools were generated by nucleofecting 500,000 HeLa cells as described above using SE nucleofector solution (Lonza) and DS-150 program in 20-µL cuvettes. Unless otherwise noted, editing rates were assessed at day 16 post-transfection.
K-562 cells (ATCC, CCL-243) were cultured in Iscove’s Modified Dulbecco’s Medium supplemented with 10% (vol/vol) fetal bovine serum, 1% (vol/vol) penicillin-streptomycin, and 1X GlutaMAX at 37°C, 5% vol/vol CO2 in a humidified incubator. K-562 cells (ATCC CCL-243) pools were generated by nucleofecting one million K562 cells as described above using P3 nucleofector solution (Lonza) and FF-120 program in 20-µL cuvettes. Unless otherwise notes, editing rates were assessed at day 10 post-transfection.
2.2. sgRNAs and DNA Donors
Alt-R sgRNAs and ssODN donors were synthesized by Integrated DNA Technologies (Coralville, IA, USA). Plasmid constructs were generated by GenScript (Piscataway, NJ, USA). Sequences of sgRNAs and DNA templates can be found in SI Table 1 & 2, respectively.
2.3. Cell Sorting and Flow Cytometry
One million U2OS cells, AN1.1 or BJFF.6 iPSCs were centrifuged at 100 g for 4 min and resuspended in phosphate-buffered saline -Ca++, -Mg++, + 2.5 mM EDTA. Flow cytometry data were acquired on a Cytek Aurora (Cytek, Fremont, CA, USA), equipped with SpectroFlo (SJCRH Flow Cytometry and Cell Sorting Shared Resource), and subsequently analyzed using FlowJo v11.1.1 (BD Life Sciences, Franklin Lakes, NJ, USA).
2.4. Genomic DNA Isolation and DNA Sequencing
One hundred thousand cells were centrifuged at 800 g for 5 min and resuspended in 100 μL genomic DNA quick extraction buffer (10 mM Tris, 2 mM EDTA, 0.2% Triton X-100, Proteinase K), then processed at 65°C for 15 min and 95°C for 5 min. First-round PCR reactions were performed in a 10-μL reaction volume containing 0.25 μM forward and reverse primers with deep sequencing tags, 1 μL extracted genomic DNA, and 9 μL QuantaBio RepliQa HiFi ToughMix. The forward deep sequencing tag is “CTACACGACGCTCTTCCGATCT”. The reverse deep sequencing tag is “CAGACGTGTGCTCTTCCGATCT”. PCR conditions were as follows: 98°C for 10 s, followed by 25 cycles of 98°C for 10 s and 68°C for 10 s. Round 2 PCR was performed as previously described [42]. PCR reactions were pooled to make a sequencing library and spiked with PhiX DNA before running the sample on a MiSeq i100 (Illumina, San Diego, CA, USA) to generate paired 2x 250-bp reads, as per the manufacturer’s instructions. Primer sequences with deep sequencing tags for each amplicon are listed in Supplementary Table 3. Samples were de-multiplexed using Illumina DNA/RNA Unique Dual Indexes Set A (Illumina). FASTQ files were generated and analyzed using CRIS.py [42].
3. Results
3.1. HDRescue: Essential Gene co-Editing Enriched HDR at the Target Locus
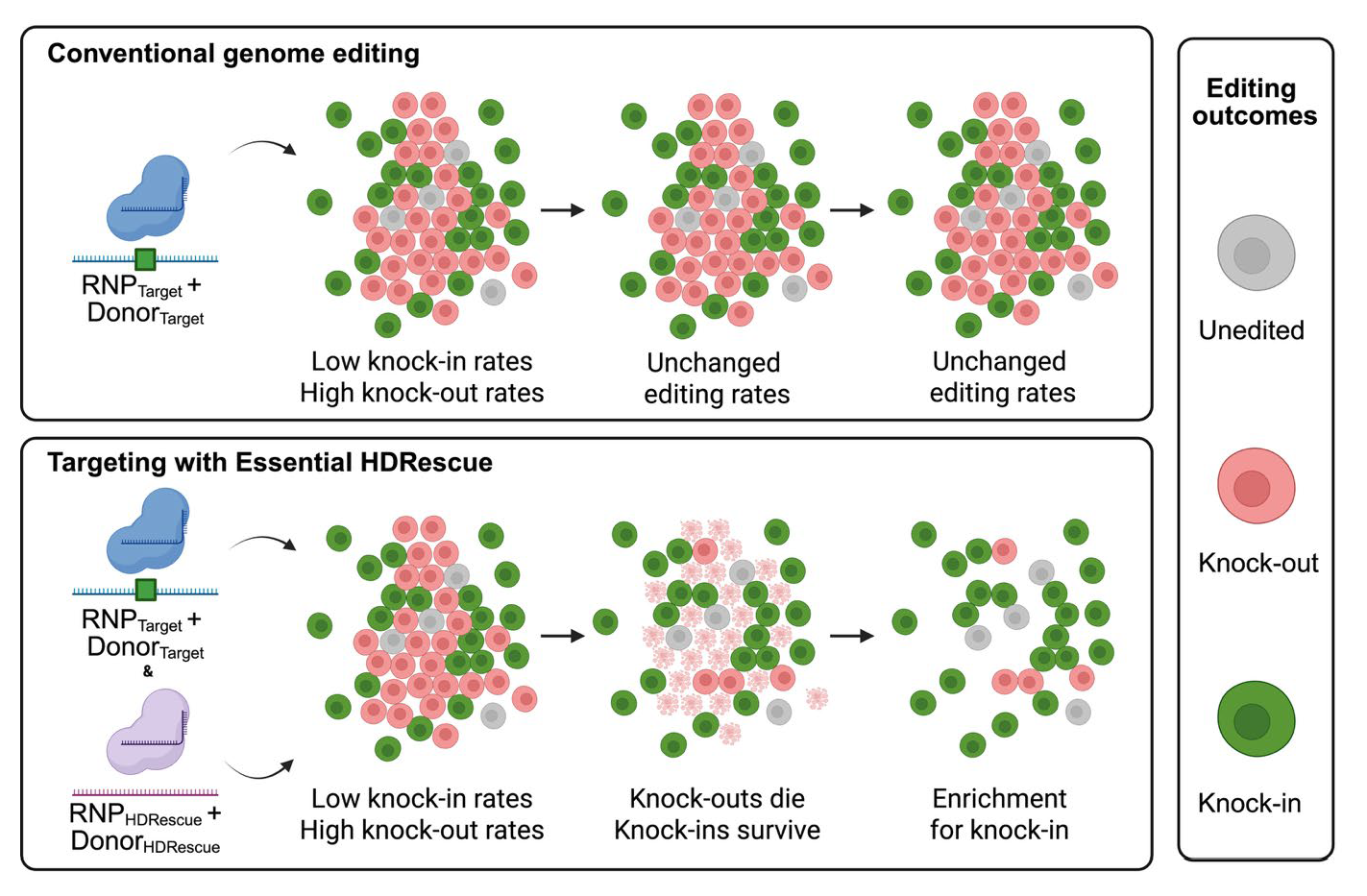
Our strategy to enhance HDR-mediated genome edits relies upon an essential gene as a co-selection locus. Unlike conventional HDR targeting, which typically uses one single guide RNA (sgRNA)-Cas ribonucleoprotein (RNP) complex and a single DNA donor template, HDRescue requires two distinct RNP complexes and two separate DNA donors (Figure 1a).
One RNP and donor pair are directed to the desired target site while the second RNP and donor pair target an essential gene, which is used as a co-selection locus for HDR-mediated editing events. We predicted that cells edited by NHEJ that acquire out-of-frame mutations in the essential gene would stop proliferating and die. To enrich for HDR-edited cells, the HDRescue donor incorporates a synonymous mutation that preserves the wild-type (WT) protein sequence of the essential gene while rendering the target site resistant to Cas9 re-cleavage. This enables positive selection for cells that successfully completed HDR. The essential gene RAN was chosen as a candidate co-selection locus because it is required for cellular proliferation and has been shown to be essential across thousands of cell types[43,44]. We designed a highly efficient sgRNA targeting the RAN coding sequence to induce a genetic KO via the NHEJ pathway and used a Cellular Fitness (CelFi) assay to measure fitness and monitor indel formation over time[45]. The CelFi assay showed that introducing a targeted DSB within the coding sequence of RAN resulted in a high ratio of out-of-frame indels on day 1 in U2OS cells, as measured by next-generation sequencing (NGS) (Figure 1b). However, we observed a sharp decline in out-of-frame indels by day 9 post-transfection, and by day 18, 77% of these indels were depleted following RAN targeting. In contrast, an intronic PPP1R12C locus, hereafter referred to as the AAVS1 safe harbor site, maintained a stable indel profile after day 3, when editing peaked. An HDRescue single-stranded oligodeoxynucleotide (ssODN) donor template was designed to KI two synonymous mutations in the protospacer sequence of the RAN sgRNA (Supplementary Information [SI] Figure 1). When installed, this HDRescue KI conferred a selective advantage to RAN-targeted cells repaired by HDR. Over time, out-of-frame indels at the RAN locus were depleted, and more than 52% of all RAN alleles in the surviving cells carried the HDRescue synonymous mutations (Figure 1b).
As a proof-of-concept, we next asked whether HDRescue increases simultaneous, precise editing at the second targeted locus. To investigate this, we transfected U2OS cells with the RAN HDRescue reagents, as well as an RNP and DNA donor template designed to introduce a 5-bp sequence at a second, unrelated target locus, AAVS1. By day 14 post-transfection, we observed a 2.4-fold increase in the rate of 5-bp insertions at AAVS1 with HDRescue compared to control cells, and this increase in HDR was maintained until the final day 28 timepoint (Figure 1c).
At the RAN locus, we initially observed NHEJ mutations occurring more frequently than those associated with HDRescue; however, this pattern reversed halfway through the experiment. On day 14, we observed the frequency of HDRescue mutations had reached ~70%, and this frequency was maintained through the final timepoint (Figure 1d). Overall, these results support the use of essential genes as co-selection loci with HDRescue reagents, show that the synonymous edits are stable, and demonstrate that HDRescue can enrich HDR-mediated edits.
3.2. HDRescue Enabled co-Selection of Desired Edits Across Multiple Cell Types Using Two Essential Gene Targets
To expand upon our proof-of-concept, we designed and validated a highly active sgRNA and cognate HDRescue ssODN donor targeting the essential splicing factor gene SF3B1 for use in our co-targeting studies alongside RAN. We evaluated the efficiency of installing a 5-bp sequence at the AAVS1 target site across five cell lines, including both cancer and iPSC lines, comparing conditions with and without HDRescue using RAN and SF3B1 as the essential genes. Although HDR efficiency varied across cell lines, the pan-essential nature of RAN and SF3B1 led us to predict that HDRescue would enhance targeting rates in all cell lines examined. In RAN HDRescue cells, we observed a 1.5- to 2.8-fold increase in HDR rates regardless of cell line compared to conventional genome targeting (Figure 2a). Similarly, in SF3B1 HDRescue cells, we observed a 1.4- to 2.9-fold increase in HDR rates regardless of cell line compared to single-targeted controls (Figure 2b). Notably, we achieved KI rates exceeding 20% in four of the five cell lines tested using HDRescue with both tested essential genes. These results show that HDRescue improves HDR targeting across multiple cell lines and types, regardless of whether RAN or SF3B1 HDRescue is used.
3.3. HDRescue Improved Multi-Allelic HDR Editing in Cell Clones
Given successful fold-enrichment of HDR edits observed in bulk cell pools, we generated single-cell–derived clones from BJFF.6 and AN1.1 iPSCs and from U2OS osteosarcoma cells and then assessed the percentage of alleles edited at both the target locus (AAVS1) and the essential gene (RAN or SF3B1). Compared with a single-targeted control, both RAN and SF3B1 HDRescue clones showed a higher proportion of cells with HDR-edited alleles and a lower proportion of cells with 100% indels at the AAVS1 target site (Figure 3, SI Table 4).
Additionally, we observed a substantial increase in the number of clones containing homozygous HDR edits installed at the target site (2.0- to 9.9-fold) compared to non-HDRescue cells, regardless of cell type. U2OS cells showed the smallest increase in homozygous HDR events, likely due to their polyploid nature. Except for a small subset of SF3B1 HDRescue clones, we found no clones to have 100% indels at the essential gene, indicating cells with NHEJ edits were lost due to essential gene dependency (SI Figure 2). Among the HDRescue clones that had 100% indels at SF3B1, each retained at least one in-frame allele, demonstrating that in-frame indels were tolerated at this locus (SI Figure 3). Notably, there were many permutations of editing outcomes across multiple alleles, even in diploid cells. We characterize desirable editing outcomes at essential RAN alleles as cells containing only HDR-mediated or non-edited alleles. However, cells with a combination of indel and WT or HDR alleles may be permissive for cellular proliferation (SI Figure 4). Out-of-frame indels on any allele at the RAN locus were always lethal and provided a means of predictable negative selection for cells edited imprecisely via NHEJ (SI Figure 4). These results suggest that HDRescue not only enhances total HDR editing rates but also increases the proportion of homozygous-targeted HDR edits across the three cell lines tested.
3.4. Detection and Removal of Aberrant Clones Improves HDRescue Accuracy
One risk of genome editing is off-target cleavage[46]. Because HDRescue uses two RNPs, there is an increased potential for off-target editing and translocations. Both the SF3B1 and RAN sgRNAs target unique sites in the human genome, with no predicted off-target sites containing fewer than two mismatches. In addition, each tested essential gene sgRNA has only a single predicted off-target site with two mismatches based on sequence homology (SI Table 5). To assess potential off-target cleavage, we screened each clone shown in Figure 3 for editing at the single predicted off-target site containing only two mismatches to the target sequence for each essential-gene sgRNA. We detected no off-target cutting in AN1.1 clones edited with SF3B1 sgRNAs, and less than 3% of all BJFF.6 and U2OS clones showed indels at the SF3B1 predicted off-target site, BDNF-AS (SI Figure 5, SI Table 4). In contrast, a higher proportion of clones edited with the RAN sgRNA showed indels at the predicted off-target site, the RAN pseudogene RANP6, across all cell lines tested (SI. Figure 5, SI Table 4). Despite this, we did not observe any change in indel ratios over time using the CelFi assay, suggesting that editing at the RANP6 locus did not affect cellular proliferation (SI Figure 5d).
Another major concern for co-targeting methods is the risk of translocations when two tandem DSBs occur in the same cell. To assess translocations after HDRescue, we developed PCR-based assays to detect translocations in both forward and reciprocal orientations. In theory, cells should not survive if a translocation disrupts an essential gene and no WT copies remain, particularly when using sgRNAs that cut early in the essential coding sequence of the essential gene. Not surprisingly, we detected no evidence of forward or reciprocal translocation products in the RAN HDRescue pools. In contrast, both forward and reciprocal translocations were observed in the SF3B1 HDRescue pools at early time points (SI Figure 6). However, when we screened individual clones for translocations between the target site and the corresponding essential gene, we found no positive clones out of more than 1,600 HDRescue clones screened (SI Table 4).
Finally, targeted DSBs can lead to larger chromosomal aberrations that might be missed by targeted amplicon sequencing approaches[47,48]. To investigate this further in our HDRescue clones and to reduce the risk of misclassifying clones as homozygous due to large deletions or complex rearrangements, we developed a PCR-based assay followed by NGS to detect a downstream heterozygous single-nucleotide polymorphism (SNP) located near, but outside, of the AAVS1 target site in BJFF.6 clones (SI Figure 7a). We found a low percentage (<4%) of clones that experienced loss of heterozygosity (LOH) at the tested SNP in all conditions. There was no significant increase in LOH between the control and HDRescue conditions, suggesting that HDRescue did not select for cells with LOH or for aberrant DSB resolution (SI Table 4, SI Figure 7b). Taken together, these data suggest that the off-target edits and larger chromosomal abnormalities observed in some HDRescue clones can be effectively detected and the affected clones excluded, enabling reliable identification of high-quality, correctly targeted clones.
3.5. HDRescue Improved HDR Editing Rates Across Multiple Loci and Different Donor Formats
Having demonstrated that we could generate HDR-modified cell pools and clones at higher yield using ssODNs at both the target site and essential gene, we next asked whether we could improve the yield of cells containing larger KI sequences, such as fluorescent protein tags, using dsDNA as the donor for the target gene. To test this, we first transfected U2OS cells with RNPs and a double-stranded DNA (dsDNA) donor to install a C-terminal enhanced green fluorescent protein (eGFP) tag at the target gene, G3BP1, with or without RAN HDRescue. By design, eGFP expression occurs only in correctly edited cells. In the absence of target-specific RNPs, no eGFP-positive cells were observed (data not shown). For HDRescue, we used two donor formats at the essential site: either an ssODN or a dsDNA plasmid. Using flow cytometry, we observed a 2.6-fold increase in the rate of eGFP-positive cells when dsDNA was used for the HDRescue donor and a 3.6-fold increase in eGFP-positive cells when an ssODN was used, compared to single-targeted control U2OS cells (Figure 4a).
Given the higher HDR rate when an ssODN donor was used as the HDRescue donor, we continued to use ssODNs for the essential gene in further studies. We transfected the G3BP1-eGFP tagging reagents with and without RAN or SF3B1 HDRescue into BJFF.6 iPSCs and found a significant 4.0-fold and 10.8-fold increase in eGFP-positive RAN and SF3B1 HDRescue cells, respectively, compared to single-targeted control cells (Figure 4b). Similarly, we found a significant 2.8- and 4.0-fold increase in the rate of eGFP targeting at the C-terminus of TARDBP when co-targeted with RAN or SF3B1 HDRescue, respectively (Figure 4c).
Finally, we assessed HDR rates across different loci using HDRescue. We knocked in SNPs in genes GATA1 and DHX9 and a 5-bp insertion in gene LIN28B, with and without RAN HDRescue, and observed significant increases in HDR across all three genes tested (Figure 4d). By measuring the rates of HDR across different cell lines and different target genes, we have highlighted the utility of HDRescue across donor modalities (ssODN or dsDNA) and demonstrate its loci-agnostic capability to improve HDR rates overall.
4. Discussion
The efficiency of HDR following a site-directed DSB varies markedly with the edit, target loci, and cell type, and is generally outcompeted by NHEJ[49]. Here, we demonstrate HDRescue, a novel CRISPR-based co-selection strategy that pairs NHEJ-mediated disruption and selective elimination of NHEJ-edited cells with HDR-mediated installation of rescuing synonymous SNPs at an essential locus. HDRescue enables strong enrichment of user-defined HDR edits at an unrelated target site, allowing efficient recovery of precisely engineered cells in a cell-intrinsic, marker-free manner. These results support previous findings that cells capable of completing one genomic manipulation are more likely to complete a second, independent manipulation when both rely on similar DNA repair mechanisms[21,30,31,32,33,34]. This co-selection is likely reliant on the cell cycle phase at the time of genome editing[49]. Cycling cells in late S or early G2 phases have a higher HDR frequency than those in other phases, and this has been used as a basis for increasing HDR frequency via cell cycle stalling[50,51]. However, our approach circumvents the need to chemically arrest cells to improve HDR rates.
HDRescue not only increases overall HDR editing in bulk cell populations but also increases the frequency of homozygous HDR mutations in clonal populations, enabling higher-throughput generation of engineered homozygous clones, which is particularly valuable in polyploid cell lines. HDRescue is a cost-, time-, and labor-saving method primarily suited for generating isogenic cell clones with precise HDR-engineered mutations, but it can also be used to create enriched pools of edited cells when downstream applications tolerate mixed editing outcomes.
Compared with other co-selection methods for HDR editing, HDRescue offers several key advantages. The use of pan-essential genes, such as RAN and SF3B1, as co-selection loci enables a universal co-selection method, as loss of pan-essential genes from out-of-frame indels repaired by NHEJ results in cell death. Our technique avoids cell cycle control and pharmacological inhibition of NHEJ to improve HDR rates, enabling the generation of HDR-enriched cell populations without the risk of triggering genomic instability or replication stress from the addition of cell cycle-modulating drugs[52,53]. HDRescue can enrich for small inserts and larger fluorescent tags without exogenous or episomal selection markers, specialized techniques, or equipment. Further, this strategy does not require a drug or toxin to induce selection and does not introduce bacterial elements into the selection locus, enabling the generation of relatively naïve cells for preclinical research and in vitro modeling. Importantly, HDRescue allows the target gene to remain under its own regulatory elements. Additionally, this technique stands out for its simplicity, since only one or two nucleotide changes are required to introduce synonymous mutations at the essential gene selection locus. We did not detect any clones with translocation products between the target and essential sites out of hundreds of screened clones. We predict that this translocation rate is lower than that for translocations between two non-essential loci, since cells lacking expression of an essential gene are not expected to proliferate.
Regardless of its advantages, each co-selection method comes with its own limitations. Despite cell-intrinsic positive and negative selection, WT and permissive-indel clones can remain because cells with at least one in-frame or WT allele can retain viability. For this reason, essential gene sgRNAs should be vetted for optimal, near-100% cutting efficiency with strong out-of-frame indel profiles to provide the most selective negative pressure. Furthermore, we tested two essential gene targets to provide options to enhance HDR across a range of targets and cell lines. While we observed HDR enrichment with both essential-gene targets, further empirical optimization may be necessary depending on the editing context, as factors such as DSB formation kinetics, NHEJ and HDR dynamics, cell-doubling time, essential gene expression, and protein half-life vary across cell types. Finally, we recommend that the essential gene should never be on the same chromosome as the targeted locus, as this could lead to a large deletion between the two concurrent DSBs.
This simple method can be broadly applied across cell lines and targeted sites to increase the frequency of genome-editing events in mammalian cells by harnessing cellular dependencies. HDRescue is easy to implement, as it only requires an additional RNP and a cognate ssODN to induce selection at the essential gene. Although we limited our studies to CRISPR-Cas9, it should be compatible with any nuclease that generates a site-directed DSB. Furthermore, the installation of HDRescue synonymous mutations could be common and naturally occurring SNPs within the coding sequences of essential genes, thus further reducing the risk of any unintended biological consequences from generating HDRescue-modified cells.
Collectively, our findings establish HDRescue as a powerful, cell-intrinsic co-selection approach that enhances HDR-mediated genome editing across multiple loci, cell types, and donor formats. HDRescue consistently improves HDR frequencies over 2-fold in both bulk pools and clonal populations, increases the recovery of homozygous edits, and maintains a low, manageable burden of off-target activity and structural variation. Importantly, this approach reduces the number of clones needing to be screened to identify correctly edited cells, providing significant savings in time, labor, and cost. This simple and robust technique delivers high-efficiency, precise genome editing, and we expect that HDRescue will help accelerate discoveries across a wide range of biological and biomedical research.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
J.F.S., J.P.C., and S.M.P-M conceived the study and designed experiments. J.F.S. performed the experiments and analyzed the data. J.F.S. and S.M.P-M wrote the manuscript. S.M.P-M super-vised the study. All authors read, reviewed, and approved of the final manuscript.
Funding
This study was funded by the American Lebanese and Syrian Associated Charities (ALSAC) and the St. Jude Graduate School of Biomedical Sciences. This research included experiments conducted by or reagents produced in the Center for Advanced Genome Engineering, Hartwell Center for Biotechnology, Protein Production Facility, and Flow Cytometry and Cell Sorting Shared Resource at SJCRH, which are supported in part by the National Cancer Institute grant P30 CA021765.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequencing data generated during this study are provided as fastq files. The SRA data generated in this study have been submitted to the NCBI BioProject database (https://www.ncbi.nlm.nih.gov/bioproject/) under accession number SUB16046062. Relevant materials are available upon request to shondra.miller@stjude.org.
Acknowledgments
We are grateful to Dr. Danny M. D’Amore (SJCRH) for her editing expertise and thoughtful feedback on the manuscript. We thank the Flow Cytometry & Cell Sorting Shared Resource and the CMB/CPNDR Microscopy Core at SJCRH for their assistance with flow cytometry and cell sorting. We thank the Center for Advanced Genome Engineering and the Hartwell Center for Biotechnology at SJCRH for assistance with next-generation sequencing library preparation and sequencing. We thank Caera Taylor for her assistance with LOH assay development.
Conflicts of Interest
The authors declare no competing interests.
References
- Cheng, H.; Jeong, E.; Cho, S.W. Applications of multiplexed CRISPR–Cas for genome engineering. Experimental & Molecular Medicine 2025, 57, 1373–1380. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, E.; Hochegger, H.; Saberi, A.; Taniguchi, Y.; Takeda, S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair (Amst) 2006, 5, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Jasin, M.; Haber, J.E. The democratization of gene editing: Insights from site-specific cleavage and double-strand break repair. DNA Repair 2016, 44, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Weber, T.; Wefers, B.; Wurst, W.; Sander, S.; Rajewsky, K.; Kühn, R. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nature Biotechnology 2015, 33, 543–548. [Google Scholar] [CrossRef]
- Jasin, M.; Rothstein, R. Repair of strand breaks by homologous recombination. Cold Spring Harb Perspect Biol 2013, 5, a012740. [Google Scholar] [CrossRef]
- Porter, S.N.; Levine, R.M.; Pruett-Miller, S.M. A Practical Guide to Genome Editing Using Targeted Nuclease Technologies. Compr Physiol 2019, 9, 665–714. [Google Scholar] [CrossRef]
- Haider, S.; Mussolino, C. Fine-Tuning Homology-Directed Repair (HDR) for Precision Genome Editing: Current Strategies and Future Directions. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Cullot, G.; Aird, E.J.; Schlapansky, M.F.; Yeh, C.D.; van de Venn, L.; Vykhlyantseva, I.; Kreutzer, S.; Mailander, D.; Lewkow, B.; Klermund, J.; et al. Genome editing with the HDR-enhancing DNA-PKcs inhibitor AZD7648 causes large-scale genomic alterations. Nat Biotechnol 2025, 43, 1778–1782. [Google Scholar] [CrossRef]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Lavrov, A.V.; Varenikov, G.G.; Skoblov, M.Y. Genome scale analysis of pathogenic variants targetable for single base editing. BMC Med Genomics 2020, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Doudna, J.A. CRISPR technology: A decade of genome editing is only the beginning. Science 2023, 379, eadd8643. [Google Scholar] [CrossRef]
- Ren, C.; Xu, K.; Segal, D.J.; Zhang, Z. Strategies for the Enrichment and Selection of Genetically Modified Cells. Trends Biotechnol 2019, 37, 56–71. [Google Scholar] [CrossRef]
- Mikkelsen, N.S.; Bak, R.O. Enrichment strategies to enhance genome editing. J Biomed Sci 2023, 30, 51. [Google Scholar] [CrossRef]
- Certo, M.T.; Ryu, B.Y.; Annis, J.E.; Garibov, M.; Jarjour, J.; Rawlings, D.J.; Scharenberg, A.M. Tracking genome engineering outcome at individual DNA breakpoints. Nat Methods 2011, 8, 671–676. [Google Scholar] [CrossRef]
- Brafman, D.A.; Wang, X.; Nguyen, T.; Schwarz, G.; Brookhouser, N.; Tekel, S.J.; Standage-Beier, K. A transient reporter for editing enrichment (TREE) in human cells. Nucleic Acids Research 2019, 47, e120–e120. [Google Scholar] [CrossRef]
- Reuven, N.; Adler, J.; Myers, N.; Shaul, Y. CRISPR Co-Editing Strategy for Scarless Homology-Directed Genome Editing. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Reuven, N.; Shaul, Y. Selecting for CRISPR-Edited Knock-In Cells. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Liao, S.; Tammaro, M.; Yan, H. Enriching CRISPR-Cas9 targeted cells by co-targeting the HPRT gene. Nucleic Acids Res 2015, 43, e134. [Google Scholar] [CrossRef]
- Kim, H.; Um, E.; Cho, S.-R.; Jung, C.; Kim, H.; Kim, J.-S. Surrogate reporters for enrichment of cells with nuclease-induced mutations. Nature Methods 2011, 8, 941–943. [Google Scholar] [CrossRef] [PubMed]
- Arribere, J.A.; Bell, R.T.; Fu, B.X.; Artiles, K.L.; Hartman, P.S.; Fire, A.Z. Efficient marker-free recovery of custom genetic modifications with CRISPR/Cas9 in Caenorhabditis elegans. Genetics 2014, 198, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.D. Rapid and precise engineering of the Caenorhabditis elegans genome with lethal mutation co-conversion and inactivation of NHEJ repair. Genetics 2015, 199, 363–377. [Google Scholar] [CrossRef]
- Kim, H.; Ishidate, T.; Ghanta, K.S.; Seth, M.; Conte, D., Jr.; Shirayama, M.; Mello, C.C. A co-CRISPR strategy for efficient genome editing in Caenorhabditis elegans. Genetics 2014, 197, 1069–1080. [Google Scholar] [CrossRef]
- Kane, N.S.; Vora, M.; Varre, K.J.; Padgett, R.W. Efficient Screening of CRISPR/Cas9-Induced Events in Drosophila Using a Co-CRISPR Strategy. G3 (Bethesda) 2017, 7, 87–93. [Google Scholar] [CrossRef]
- Ge, D.T.; Tipping, C.; Brodsky, M.H.; Zamore, P.D. Rapid Screening for CRISPR-Directed Editing of the Drosophila Genome Using white Coconversion. G3 (Bethesda) 2016, 6, 3197–3206. [Google Scholar] [CrossRef]
- Yan, N.; Sun, Y.; Fang, Y.; Deng, J.; Mu, L.; Xu, K.; Mymryk, J.S.; Zhang, Z. A Universal Surrogate Reporter for Efficient Enrichment of CRISPR/Cas9-Mediated Homology-Directed Repair in Mammalian Cells. Molecular Therapy - Nucleic Acids 2020, 19, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Akrap, N.; Cerboni, S.; Porritt, M.J.; Wimberger, S.; Lundin, A.; Möller, C.; Firth, M.; Gordon, E.; Lazovic, B.; et al. Universal toxin-based selection for precise genome engineering in human cells. Nature Communications 2021, 12, 497. [Google Scholar] [CrossRef]
- Agudelo, D.; Duringer, A.; Bozoyan, L.; Huard, C.C.; Carter, S.; Loehr, J.; Synodinou, D.; Drouin, M.; Salsman, J.; Dellaire, G.; et al. Marker-free coselection for CRISPR-driven genome editing in human cells. Nat Methods 2017, 14, 615–620. [Google Scholar] [CrossRef]
- Mitzelfelt, K.A.; McDermott-Roe, C.; Grzybowski, M.N.; Marquez, M.; Kuo, C.T.; Riedel, M.; Lai, S.; Choi, M.J.; Kolander, K.D.; Helbling, D.; et al. Efficient Precision Genome Editing in iPSCs via Genetic Co-targeting with Selection. Stem Cell Reports 2017, 8, 491–499. [Google Scholar] [CrossRef]
- Moriarity, B.S.; Rahrmann, E.P.; Beckmann, D.A.; Conboy, C.B.; Watson, A.L.; Carlson, D.F.; Olson, E.R.; Hyland, K.A.; Fahrenkrug, S.C.; McIvor, R.S.; et al. Simple and Efficient Methods for Enrichment and Isolation of Endonuclease Modified Cells. PLOS ONE 2014, 9, e96114. [Google Scholar] [CrossRef]
- Shy, B.R.; MacDougall, M.S.; Clarke, R.; Merrill, B.J. Co-incident insertion enables high efficiency genome engineering in mouse embryonic stem cells. Nucleic Acids Res 2016, 44, 7997–8010. [Google Scholar] [CrossRef]
- Li, S.; Akrap, N.; Cerboni, S.; Porritt, M.J.; Wimberger, S.; Lundin, A.; Moller, C.; Firth, M.; Gordon, E.; Lazovic, B.; et al. Universal toxin-based selection for precise genome engineering in human cells. Nat Commun 2021, 12, 497. [Google Scholar] [CrossRef]
- Liu, W.H.; Volse, K.; Senft, D.; Jeremias, I. A reporter system for enriching CRISPR/Cas9 knockout cells in technically challenging settings like patient models. Sci Rep 2021, 11, 12649. [Google Scholar] [CrossRef]
- Allen, A.G.; Khan, S.Q.; Margulies, C.M.; Viswanathan, R.; Lele, S.; Blaha, L.; Scott, S.N.; Izzo, K.M.; Gerew, A.; Pattali, R.; et al. A highly efficient transgene knock-in technology in clinically relevant cell types. Nat Biotechnol 2023. [Google Scholar] [CrossRef]
- Bartha, I.; di Iulio, J.; Venter, J.C.; Telenti, A. Human gene essentiality. Nat Rev Genet 2018, 19, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Lee, I.; Marcotte, E.M.; Hurles, M.E. Characterising and predicting haploinsufficiency in the human genome. PLoS Genet 2010, 6, e1001154. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Peretz, M.; Aharony, A.; Sagi, I.; Benvenisty, N. Defining essential genes for human pluripotent stem cells by CRISPR-Cas9 screening in haploid cells. Nat Cell Biol 2018, 20, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576 e516. [Google Scholar] [CrossRef]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J Biomol Tech 2018, 29, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Narina, S.; Connelly, J.P.; Pruett-Miller, S.M. Transgenesis: Methods and Protocols 2023, 155–182.
- Boudhraa, Z.; Carmona, E.; Provencher, D.; Mes-Masson, A.-M. Ran GTPase: A Key Player in Tumor Progression and Metastasis. Frontiers in Cell and Developmental Biology 2020, 8. [Google Scholar] [CrossRef]
- Institute, B. DepMap: The Cancer Dependency Map Project at Broad Institute. Available online: https://depmap.org/portal/.
- Loughran, A.J.; Narina, S.; Klein, J.; Siwak, J.F.; Connelly, J.P.; Pruett-Miller, S.M. Rapid and robust validation of pooled CRISPR knockout screens using CelFi. Sci Rep 2025, 15, 13358. [Google Scholar] [CrossRef]
- Guo, C.; Ma, X.; Gao, F.; Guo, Y. Off-target effects in CRISPR/Cas9 gene editing. Front Bioeng Biotechnol 2023, 11, 1143157. [Google Scholar] [CrossRef]
- Regan, S.B.; Medhi, D.; Xu, Y.; White, T.B.; Jiang, Y.Z.; Kim, J.E.; Wang, S.C.; Deng, Q.; Jia, S.; Baasan, D.; et al. Megabase-scale loss of heterozygosity provoked by CRISPR-Cas9 DNA double-strand breaks. Mol Cell 2025, 85, 4119–4137 e4110. [Google Scholar] [CrossRef]
- Kosicki, M.; Tomberg, K.; Bradley, A. Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements. Nat Biotechnol 2018, 36, 765–771. [Google Scholar] [CrossRef]
- Liao, H.; Wu, J.; VanDusen, N.J.; Li, Y.; Zheng, Y. CRISPR-Cas9-mediated homology-directed repair for precise gene editing. Mol Ther Nucleic Acids 2024, 35, 102344. [Google Scholar] [CrossRef]
- Li, G.; Yang, X.; Luo, X.; Wu, Z.; Yang, H. Modulation of cell cycle increases CRISPR-mediated homology-directed DNA repair. Cell Biosci 2023, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Kostyrko, K.; Bosshard, S.; Urban, Z.; Mermod, N. A role for homologous recombination proteins in cell cycle regulation. Cell Cycle 2015, 14, 2853–2861. [Google Scholar] [CrossRef] [PubMed]
- Aussel, C.; Cathomen, T.; Fuster-Garcia, C. The hidden risks of CRISPR/Cas: structural variations and genome integrity. Nat Commun 2025, 16, 7208. [Google Scholar] [CrossRef] [PubMed]
- Dibitetto, D.; Marshall, S.; Sanchi, A.; Liptay, M.; Badar, J.; Lopes, M.; Rottenberg, S.; Smolka, M.B. DNA-PKcs promotes fork reversal and chemoresistance. Mol Cell 2022, 82, 3932–3942 e3936. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Essential HDRescue: Essential gene co-editing enriches homology-directed repair (HDR) at the target locus. (A) Schematic diagram of the HDRescue co-selection strategy in comparison to conventional genome editing. HDRescue employs two CRISPR–Cas9 ribonucleoprotein (RNP) complexes to generate double-stranded DNA breaks, one at the target site and one at an essential locus, along with two matched DNA donors to enable HDR at both locations. One donor installs a user-defined genome modification at the target site, while the second donor, the HDRescue template, introduces synonymous substitutions at an essential gene to prevent Cas9 from re-cutting that locus and preserve the wild-type protein sequence. (B) Two targets were tested using a cellular fitness assay to assess dependencies of nonessential and essential loci: AAVS1, an intronic safe harbor site, and RAN, tested both with and without the HDRescue single-stranded oligodeoxynucleotide donor in U2OS cells. The stacked bar plots show the percentage of in-frame indels, out-of-frame indels, 0-bp alleles, and alleles incorporating the HDRescue synonymous mutations in the cell pool over time. (C) Non-homologous end-joining (NHEJ, squares) and HDR (circles) editing rates over time using conventional targeting (- HDRescue) and targeting with HDRescue (+ HDRescue) to install a 5-bp insertion at the target site, AAVS1, in U2OS cells. (D) NHEJ (red) and HDRescue (green) editing rates at the RAN locus over time in U2OS cells. Experiments were performed in triplicate with error bars representing the standard deviation. The schematic was generated using Biorender.com.
Figure 1.
Essential HDRescue: Essential gene co-editing enriches homology-directed repair (HDR) at the target locus. (A) Schematic diagram of the HDRescue co-selection strategy in comparison to conventional genome editing. HDRescue employs two CRISPR–Cas9 ribonucleoprotein (RNP) complexes to generate double-stranded DNA breaks, one at the target site and one at an essential locus, along with two matched DNA donors to enable HDR at both locations. One donor installs a user-defined genome modification at the target site, while the second donor, the HDRescue template, introduces synonymous substitutions at an essential gene to prevent Cas9 from re-cutting that locus and preserve the wild-type protein sequence. (B) Two targets were tested using a cellular fitness assay to assess dependencies of nonessential and essential loci: AAVS1, an intronic safe harbor site, and RAN, tested both with and without the HDRescue single-stranded oligodeoxynucleotide donor in U2OS cells. The stacked bar plots show the percentage of in-frame indels, out-of-frame indels, 0-bp alleles, and alleles incorporating the HDRescue synonymous mutations in the cell pool over time. (C) Non-homologous end-joining (NHEJ, squares) and HDR (circles) editing rates over time using conventional targeting (- HDRescue) and targeting with HDRescue (+ HDRescue) to install a 5-bp insertion at the target site, AAVS1, in U2OS cells. (D) NHEJ (red) and HDRescue (green) editing rates at the RAN locus over time in U2OS cells. Experiments were performed in triplicate with error bars representing the standard deviation. The schematic was generated using Biorender.com.

Figure 2.
HDRescue enables co-selection of desired edits across multiple cell types using two essential gene targets. Knock-in of a 5-bp insertion at the AAVS1 locus in BJFF.6 and AN1.1 iPSCs, HeLa, K562 and U2OS cells under conditions with and without (a) RAN or (b) SF3B1 HDRescue. n=3 biological replicates per experiment; error bars represent mean +/- s.d.; one-way Student’s t-test, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
Figure 2.
HDRescue enables co-selection of desired edits across multiple cell types using two essential gene targets. Knock-in of a 5-bp insertion at the AAVS1 locus in BJFF.6 and AN1.1 iPSCs, HeLa, K562 and U2OS cells under conditions with and without (a) RAN or (b) SF3B1 HDRescue. n=3 biological replicates per experiment; error bars represent mean +/- s.d.; one-way Student’s t-test, *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.

Figure 3.
HDRescue improves multi-allelic homology-directed repair editing in cell clones. Genotypes of clones at the target site, AAVS1, with and without RAN or SF3B1 HDRescue in (a) BJFF.6 iPSCs, (b) AN1.1 iPSCs, and (c) U2OS cells. Data plotted is a percentage of total viable clones from one biological replicate per condition. Abbreviations: WT – wild-type, het. - heterozygous, hom.- homozygous.
Figure 3.
HDRescue improves multi-allelic homology-directed repair editing in cell clones. Genotypes of clones at the target site, AAVS1, with and without RAN or SF3B1 HDRescue in (a) BJFF.6 iPSCs, (b) AN1.1 iPSCs, and (c) U2OS cells. Data plotted is a percentage of total viable clones from one biological replicate per condition. Abbreviations: WT – wild-type, het. - heterozygous, hom.- homozygous.

Figure 4.
HDRescue improves knock-in editing rates across multiple loci and different donor formats. (a) Frequency of homology-directed repair (HDR) at target site G3BP1 for a C-terminus enhanced GFP (eGFP) tag measured by flow cytometry with two different RAN HDRescue DNA templates, either double-stranded DNA or single-stranded oligodeoxynucleotide, in U2OS cells. (b) Frequency of HDR at target site G3BP1 for a C-terminus eGFP tag as measured by flow cytometry in BJFF.6 iPSCs with and without RAN or SF3B1 HDRescue. (c) Frequency of HDR at target site TARDBP for a C-terminus eGFP tag as measured by flow cytometry in BJFF.6 iPSCs with and without RAN or SF3B1 HDRescue. (d) HDR efficiency of three different targets using BJFF.6 iPSCs with and without RAN HDRescue. n=3 biological replicates; error bars represent mean +/- s.d.; Student’s t-test, *P<0.05, **P<0.01, **P<0.0001.
Figure 4.
HDRescue improves knock-in editing rates across multiple loci and different donor formats. (a) Frequency of homology-directed repair (HDR) at target site G3BP1 for a C-terminus enhanced GFP (eGFP) tag measured by flow cytometry with two different RAN HDRescue DNA templates, either double-stranded DNA or single-stranded oligodeoxynucleotide, in U2OS cells. (b) Frequency of HDR at target site G3BP1 for a C-terminus eGFP tag as measured by flow cytometry in BJFF.6 iPSCs with and without RAN or SF3B1 HDRescue. (c) Frequency of HDR at target site TARDBP for a C-terminus eGFP tag as measured by flow cytometry in BJFF.6 iPSCs with and without RAN or SF3B1 HDRescue. (d) HDR efficiency of three different targets using BJFF.6 iPSCs with and without RAN HDRescue. n=3 biological replicates; error bars represent mean +/- s.d.; Student’s t-test, *P<0.05, **P<0.01, **P<0.0001.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



