Submitted:
07 March 2026
Posted:
09 March 2026
You are already at the latest version
Abstract
Pyrethroids, widely used insecticides, have raised concerns regarding their adverse effects on non-target organisms, particularly molluscs, which play vital roles in aquatic ecosystems. Pyrethroids disrupt immune responses in molluscs, impairing their ability to combat pathogens and environmental stressors. Behavioural changes, including altered feeding, locomotion, and mating patterns, are observed, which can affect population dynamics and ecosystem functioning. Additionally, pyrethroids influence molluscan metabolism by altering enzymatic pathways involved in detoxification, leading to potential metabolic overload and toxicity. These disruptions threaten molluscan biodiversity and ecological balance. This review summarize the impact of pyrethroid exposure on molluscan diversity, focusing on immunological, behavioural, and metabolic alterations. This review also explores potential mitigation strategies, including the use of environmentally safer alternatives, bioremediation techniques, and the development of resistance management approaches. Understanding the multifaceted impacts of pyrethroids on molluscs is crucial for safeguarding aquatic ecosystems and developing effective pest management strategies.
Keywords:
pyrethroids
; molluscan diversity
; metabolic pathways
; pest management
; bioremediation
Introduction
Aquatic ecosystems face growing threats from anthropogenic activities, including pesticide contamination. Pyrethroids are among the most commonly used insecticides in agriculture and public health. Pyrethroids, as widely used pesticides, often find their way into aquatic systems through agricultural runoff, improper disposal, and atmospheric deposition. While effective against target pests, their presence in aquatic ecosystems raises concerns due to their persistence and bio-accumulative potential (Iqbal et al., 2024). Molluscs, including bivalves and gastropods, are particularly vulnerable due to their filter-feeding habits and direct interaction with contaminated sediments and water (Vaughn et al., 2018). Their lipophilic nature enhances their affinity for organic matter in sediments, making benthic organisms like molluscs susceptible to chronic exposure (Landrum et al., 2020). Moreover, molluscs contribute to ecosystem health by acting as grazers, nutrient recyclers, and prey for higher trophic levels. Molluscs, including bivalves (clams, oysters, mussels), gastropods (snails, abalones), and cephalopods (squid, octopus), play a crucial role in global aquaculture and fisheries (Gökoğlu, 2021). Known for their rich nutritional profile, they provide high-quality proteins, essential fatty acids, and vital micronutrients, contributing significantly to human diets (Ngandjui et al., 2024). Disruption in their physiological or behavioural processes due to pyrethroid toxicity can result in cascading effects throughout the ecosystem. Molluscs, one of the most diverse animal phyla, exhibit a wide array of immune mechanisms, communication systems, and behavioural adaptations. Despite significant advancements in understanding these areas, critical research gaps remain. Molluscs are a highly diverse group, encompassing species ranging from bivalves and gastropods to cephalopods (Quiroz-Barroso et al., 2024). Their unique physiological, immune, and behavioural adaptations have made them subjects of increasing scientific interest. Expanding research in these areas will enhance our understanding of molluscan biology and their ecological and biomedical significance. This review synthesizes current knowledge on molluscan immunity, including haemocyte function, apoptotic mechanisms, and environmental stress impacts, alongside insights into their communication systems and behaviours. Special attention is given to the effects of pesticides on cellular communication and immune responses in molluscs. Identifying these gaps highlights opportunities for future research, particularly in underexplored species, molecular mechanisms, and responses to environmental change. Additionally, we discuss emerging molecular tools, potential remediation strategies, and sustainable pest management approaches to mitigate pyrethroid contamination.
Figure 1.
Graphical abstract.

Environmental Degradation of Pyrethroids
Pyrethroids are subject to both biotic and abiotic degradation processes in the environment (Méjanelle et al., 2020). Abiotic factors such as photodegradation and hydrolysis play significant roles in breaking down these compounds. Photodegradation, driven by sunlight exposure, can rapidly reduce pyrethroid concentrations on plant surfaces and in shallow water bodies (Katagi, 2004). Hydrolysis, the chemical breakdown due to reaction with water, also contributes to their degradation, with rates varying depending on the specific pyrethroid compound and environmental conditions (Méjanelle et al., 2020). Biotic degradation involves microbial activity, where bacteria and fungi metabolize pyrethroids into less harmful substances (Cycoń et al., 2016). The efficiency of microbial degradation depends on factors such as soil composition, temperature, pH, and the presence of suitable microbial communities (Cycoń et al., 2016). In soils rich in organic matter, pyrethroids tend to adsorb strongly to soil particles, which can reduce their bioavailability and slow down microbial degradation.
Microbial Degradation of Pyrethroids
One promising approach to controlling pyrethroid pollution is the use of pyrethroid-degrading microorganisms. Certain bacteria and fungi have been identified that can metabolize pyrethroids, breaking them down into less harmful substances. A comprehensive review by Cycoń et al. (2016) discusses various microorganisms isolated from pyrethroid-contaminated sites and their potential applications in bioremediation. The study emphasizes the role of microbial degradation in mitigating pyrethroid pollution in both soil and aquatic environments.
Factors Influencing Persistence of Pyrethroids
The persistence of pyrethroids in the environment is influenced by their chemical structure, environmental conditions, and application methods (Aznar-Alemany et al., 2020). For instance, pyrethroids with certain chemical modifications may exhibit increased resistance to photodegradation, leading to longer environmental half-lives (Samia et al., 2024). Environmental factors such as temperature, pH, and the presence of organic matter can also affect degradation rates. Higher temperatures and neutral to alkaline pH levels generally enhance degradation processes, while low temperatures and acidic conditions may slow them down (Jaggi et al., 2005).
Environmental Persistence and Bioavailability of Pyrethroids
Pyrethroids are known to adsorb strongly to sediments due to their hydrophobic nature, which can reduce their immediate bioavailability in the water column (Méjanelle et al., 2020). However, this sediment association poses a risk to benthic organisms, including many molluscs that inhabit or feed within sediment layers (Griscom et al., 2004). The persistence of pyrethroids in sediments can lead to prolonged exposure for these organisms, potentially affecting their health and the overall diversity of molluscan communities (Table 1).
Pyrethroid Toxicity to Molluscs
Research indicates that pyrethroids can be highly toxic to various aquatic invertebrates, including molluscs. A review by Farag et al. (2021) highlighted that crustaceans are generally more sensitive to pyrethroids than fish, with certain species exhibiting 96-hour LC50 values (lethal concentration for 50% of the population) below 0.02 μg/L for permethrin and below 0.01 μg/L for fenvalerate, cypermethrin, and flucythrinate (Kamrin, 1997). While specific data on molluscs were limited in this study, the high toxicity to related aquatic invertebrates suggests potential risks to molluscan species as well. Pyrethroids are widely used insecticides that have been found to have detrimental effects on non-target organisms, including aquatic molluscs (Saha et al., 2025). These effects are of particular concern because molluscs are essential components of aquatic ecosystems, playing key roles in nutrient cycling, sediment stability, and as a food source for higher trophic levels. Pyrethroids’ potential to disrupt the sex, age, and size of molluscs can have profound ecological consequences in molluscs (Major et al., 2020). Molluscs exhibit diverse reproductive strategies, and their sex determination systems can be hampered during exposure to chemicals like pyrethroids (Stemberg et al., 2010). The age and life span of molluscs can be directly influenced by exposure to pyrethroids, especially as these chemicals are known to cause physiological stress and impair growth (Werner et al., 2008). Pyrethroids, especially when encountered in early life stages, can delay the development of juvenile molluscs, leading to a longer time to reach sexual maturity (Werner et al., 2008). This delayed development may impact the timing of reproduction, with potential long-term consequences for population dynamics. Exposure to permethrin in freshwater snails (Lymnaea stagnalis) caused a delay in the development of juvenile snails, with slower shell growth and delayed reproductive capability (Leung et al., 2007). These effects can contribute to a shift in the age distribution of a population, with fewer older, reproductive individuals. Studies on mollusc longevity following pyrethroid exposure have shown conflicting results, but many report a reduction in life span (Major et al., 2020). Toxicological stress, such as oxidative stress caused by pyrethroid exposure, can accelerate aging processes in molluscs by damaging cellular structures like membranes and mitochondria (Saha et al., 2024). In Ruditapes philippinarum (a species of clam), exposure to pyrethroids like cypermethrin has led to increased mortality rates among older clams, thereby reducing the overall life span of exposed populations (Wu et al., 2023). The impaired ability to cope with oxidative damage results in early senescence, diminishing the population’s age structure. Molluscs are known to be sensitive to environmental stressors, and pyrethroid exposure can directly impact their growth, particularly in aquatic environments (Major et al., 2020). Pyrethroids can stunt growth by affecting feeding behaviour, metabolic processes, and overall health. For molluscs, the exposure to these insecticides can result in reduced shell growth, abnormal development of soft tissues, and smaller body size (Dhiman et al., 2021). In Mytilus edulis (blue mussel), exposure to pyrethroids like permethrin has been associated with a decrease in both shell length and body mass, especially in individuals exposed during the juvenile phase (Ait Ayad et al., 2011). The mechanism behind this is thought to involve interference with energy metabolism, as well as inhibition of feeding activity (Berthoud, 2004). Exposure to cypermethrin in freshwater gastropods like Gastropod planorbid has shown a significant reduction in the growth rate of juveniles (San Juan et al., 2020), attributed to reduced feeding rates and physiological stress induced by the pyrethroid. There is a direct relationship between body size and reproductive success in many molluscs. Reduced growth due to pyrethroid exposure can therefore result in decreased fecundity and reproductive success (Werner et al., 2008). In freshwater bivalves like the Asian clam (Corbicula fluminea), pyrethroid exposure leads to reduced body size and, consequently, reduced egg production (Shan et al., 2020). The smaller individuals have fewer gametes, which could affect population sustainability.
Absorption and Distribution of Pyrethroids in Molluscs
Absorption
Pyrethroids are typically absorbed through the respiratory and integumentary systems in molluscs, depending on the species and exposure route (waterborne or sediment-bound). Studies have shown that molluscs like snails and bivalves can absorb pyrethroids efficiently, with higher absorption rates typically observed in the gills, which are highly vascularized tissues (Langston et al., 2020).
Distribution
Once absorbed, pyrethroids distribute through the mollusc’s tissues. In aquatic molluscs, pyrethroids often accumulate in lipid-rich tissues such as the mantle, gills, and hepatopancreas (Katagi, 2009). This distribution is largely influenced by the lipophilicity of pyrethroids, which makes them prone to accumulate in fatty tissues and organs involved in detoxification.
Pyrethroid Metabolic Pathways in Molluscs
Pyrethroid metabolism in molluscs involves a series of enzymatic reactions that transform the parent compound into more water-soluble metabolites, facilitating its elimination. These metabolic processes can be broadly categorized into Phase I and Phase II reactions.
Phase I Metabolism
Phase I enzymes, particularly cytochrome P450 monooxygenases (CYPs), play a central role in pyrethroid metabolism in molluscs (Boutet et al., 2004). These enzymes catalyse the oxidation of pyrethroids, leading to the formation of more polar metabolites. Some pyrethroids, such as permethrin, undergo hydrolysis in molluscs, catalysed by esterases (carboxylesterases) (Sogorb et al., 2002). Hydrolysis of the ester bonds in pyrethroid molecules results in the formation of alcohol and acid metabolites, which are often less toxic than the parent compound. CYP450 enzymes oxidize the pyrethroid molecules, leading to the formation of hydroxylated metabolites (Kaneko, 2010). For instance, cypermethrin is oxidized to produce 3-phenoxybenzyl alcohol and 3-phenoxybenzoic acid.
Phase II Metabolism
Phase II enzymes, such as glutathione S-transferases (GSTs), UDP-glucuronosyltransferases (UGTs), and sulfotransferases, catalyse the conjugation of the polar metabolites produced in Phase I with endogenous molecules such as glutathione, sulphate, or glucuronic acid (Chen, 2011). This conjugation process increases the solubility of the metabolites, making them easier to excrete via the mollusc’s excretory systems. For instance, cypermethrin metabolites may undergo conjugation with glutathione via GSTs, resulting in less toxic and more water-soluble products. Understanding the metabolism of pyrethroids in molluscs is essential to predict their toxicological impact, assess environmental risks, and explore potential bioremediation strategies.
Life Stage Dependent Detoxification of Pyrethroids in Molluscs
Different mollusc species exhibit variability in their metabolic capabilities. For example, studies have shown that marine bivalves like mussels have higher CYP450 activity compared to freshwater gastropods like snails (Petushok et al., 2002). This difference in metabolic capacity can result in species-specific responses to pyrethroid exposure. The age and size of molluscs can influence their metabolic rate. Juvenile molluscs generally exhibit slower metabolic processes compared to adults (Bayne, 2004). This may affect their ability to detoxify and eliminate pyrethroids, making them more susceptible to toxic effects. In Lymnaea stagnalis, younger snails were shown to accumulate higher concentrations of pyrethroids than older individuals, likely due to slower metabolic rates in younger stages (Tripathi et al., 2004). The chemical formulation of pyrethroids, such as the presence of synergists (e.g., piperonyl butoxide), can influence their metabolism. Synergists enhance the effectiveness of pyrethroids by inhibiting the action of detoxification enzymes, thus altering the metabolism of pyrethroids in molluscs (Werner et al., 2008).
Effects of Pyrethroids on Molluscan Behaviours
Molluscs, one of the most diverse animal phyla, exhibit a remarkable array of behaviours that are influenced by environmental stimuli, neural circuitry, and genetic factors. The integration of behavioural ecology, neurobiology, and evolutionary biology provides insights into how molluscs interact with their environment, adapt to changing conditions, and exhibit complex neural plasticity (Di Cosmo, 2014). This review synthesizes findings on foraging, feeding, movement, homing, mating, and learning in molluscs, emphasizing the role of neuromodulation and genetic expression in shaping behaviour (Table 2).
Effects of Pyrethroids on Molluscsan Foraging and Feeding Behaviour
Foraging and feeding strategies in molluscs are highly adaptable, a feature that underscores their evolutionary success. Studies on gastropods like Helix aspersa reveal that food selection is not entirely random but follows a hierarchical preference based on prior experience and nutritional needs (Romano, 2022). The ability to modify foraging strategies suggests cognitive elements akin to associative learning in higher organisms. Feeding behaviour in molluscs is regulated by a dynamic motor control system that is continuously modulated by sensory inputs originating from both internal physiological signals and external environmental stimuli. The feeding process is primarily controlled by neural circuits, often referred to as central pattern generators, which coordinate rhythmic muscular movements involved in food detection, ingestion, and processing (Benjamin et al., 2020). The buccal ganglia, which control the radula and feeding appendages, exhibit a high degree of plasticity. Neuromodulators such as serotonin and dopamine play crucial roles in altering the rhythmicity and intensity of feeding behaviour (Miller, 2020). Carnivorous species, such as Pleurobranchaea californica, demonstrate prey-selective foraging behaviours controlled by neuromodulator circuits, highlighting a link between feeding strategy and neural organization (Gillette et al., 2015).
Effects of Pyrethroids on Molluscsan Mating Strategies and Evolutionary Implications
Molluscan reproductive strategies vary widely, particularly in hermaphroditic pulmonates. The relationship between shell morphology and mating behaviour is a key example of evolutionary adaptation. Low-spired species often engage in reciprocal sperm exchange, whereas high-spired species favour unilateral sperm transfer (Jordaens et al., 2009). These patterns are not rigid but show considerable intra-species variability, suggesting that behavioural plasticity plays a role in reproductive success. A particularly fascinating aspect of molluscan reproduction is dart-shooting behaviour observed in some land snails (Lodi et al., 2016). This phenomenon, wherein individuals pierce their mates with a calcareous dart, enhances sperm survival by introducing bioactive compounds that influence sperm storage. This form of sexual selection indicates a sophisticated evolutionary mechanism for increasing reproductive fitness. Pyrethroids primarily exert neurotoxic effects by interfering with voltage-gated sodium channels in neuronal membranes, resulting in prolonged nerve excitation and impaired neural signalling. Because molluscan courtship and mating behaviours depend on coordinated sensory perception, motor activity, and neural regulation, neurotoxic interference can alter the behavioural sequences necessary for successful reproduction. Individuals exposed to pyrethroids may exhibit reduced locomotory activity, delayed courtship initiation, or incomplete mating behaviour, which can decrease the probability of successful copulation (Jordaens et al., 2009).
In addition to behavioural disruption, pyrethroids may influence the physiological processes involved in gamete production and reproductive organ function. Oxidative stress induced by pyrethroid metabolism can damage cellular components within gonadal tissues, potentially affecting gametogenesis and sperm viability. Alterations in enzymatic activity and endocrine-like signalling pathways may further interfere with reproductive timing and hormonal regulation. Such disturbances can reduce fecundity, impair sperm transfer efficiency, and decrease fertilization success (Lodi et al., 2016). Behaviours such as dart-shooting and reciprocal sperm exchange require precise muscular coordination and accurate behavioural timing, both of which depend on intact neural function. Neurotoxic effects of pyrethroids may therefore disrupt these complex interactions, prevent successful dart deployment or alter the physiological responses that normally enhance sperm storage (Jordaens et al., 2009). Consequently, interference with these reproductive mechanisms may reduce reproductive success and ultimately influence population stability in contaminated environments.
Effects of Pyrethroids on Molluscsan Neurobiology and Behavioural Gene Expressions
Behavioural diversity in molluscs is closely linked to neuromodulation and gene expression. Serotonin receptor diversity is correlated with swimming behaviour in Pleurobranchaea californica and Tritonia diomedea, suggesting that receptor expression levels influence behavioural variability (Tamvacakis et al., 2018). Neurophysiological studies reveal that molluscs regulate synaptic strength through serotonin-dependent pathways, enabling adaptive changes in locomotion and decision-making (Bacqué-Cazenave et al., 2020). Gene expression studies provide further insight into how molluscs respond to environmental stressors (Li et al., 2013). Differential expression of genes related to oxidative stress, apoptosis, and neuromodulation has been observed in molluscs exposed to pollutants, indicating a direct link between gene activity and behavioural adaptation (Deidda et al., 2021). These findings have implications for understanding the role of epigenetics in molluscan behaviour and environmental resilience.
Effects of Pyrethroids on Molluscsan Memory and Learning Mechanisms
Molluscs are powerful models for studying memory formation due to their relatively simple nervous systems. Studies on Lymnaea stagnalis have demonstrated that long-term memory (LTM) formation is associated with specific neurons such as RPeD1 (Braun et al., 2012). Ablation of this neuron prevents LTM retention but does not affect short-term learning, suggesting that memory consolidation is localized to specific neural circuits (Sakakibara et al., 2008). Operant conditioning studies in Lymnaea show that learning induces changes in synaptic connectivity and gene expression (Benjamin et al., 2010). The modulation of calcium-dependent signalling pathways during memory consolidation parallels mechanisms observed in vertebrate learning models, providing a comparative framework for understanding the evolution of cognitive functions (Zha et al., 2022). Pyrethroids bind to voltage-gated sodium channels located in neuronal membranes and prolong their open state, resulting in sustained depolarization and abnormal nerve impulse transmission. Such disruption alters neuronal excitability and synaptic communication within neural circuits responsible for information processing and memory consolidation. In molluscs such as Lymnaea stagnalis, where memory formation depends on the precise activity of identifiable neurons such as RPeD1, disturbances in ion channel dynamics may impair the neuronal signalling required for long-term synaptic modification (Braun et al., 2012). The formation of long-term memory involves molecular events including calcium influx, activation of second-messenger systems, and regulation of gene transcription necessary for synaptic restructuring. Pyrethroid-induced neurotoxicity can disrupt these calcium-dependent signalling pathways by altering membrane excitability and ionic balance (Benjamin et al., 2010). Consequently, the intracellular cascades that normally support synaptic plasticity and memory consolidation may be weakened or interrupted. This interference can lead to reduced efficiency in neural communication between memory-related neurons and diminished capacity for long-term behavioural adaptation.

Table 2.
Impacts of Pyrethroids on molluscan immunology and behavioural changes in molluscs (↑ arrow indicates upregulation and ↓ indicates downregulation).
Table 2.
Impacts of Pyrethroids on molluscan immunology and behavioural changes in molluscs (↑ arrow indicates upregulation and ↓ indicates downregulation).
| Pyrethroid Type | Immunological Effects | Behavioral Changes | Mode of Action | Reference |
|---|---|---|---|---|
| Cypermethrin | ↓ Total hemocyte count (THC), ↑ lysosomal membrane damage, ↓ phenoloxidase activity, ↑ROS generation. | Lethargy, reduced mobility, frequent shell closure | Neurotoxic action via voltage-gated sodium channel disruption, oxidative stress-induced hemocyte apoptosis. | San Juan et al., 2020 |
| Deltamethrin | ↓ Phagocytic index, ↓ nitric oxide (NO) production, ↑lipid peroxidation (MDA levels). | Altered locomotion, reduced feeding rate | Disruption of immune signalling, oxidative stress, mitochondrial dysfunction. | Jiang et al., 2021 |
| Permethrin | ↓ Superoxide dismutase (SOD), ↓ catalase (CAT), ↑ glutathione-S-transferase (GST) activity. | Reduced surface activity, impaired muscular response | Generation of oxidative stress, reduced antioxidant defines. | Arslan, 2023 |
| Cyfluthrin | ↓ Hemocyanin level, ↑ pro-inflammatory cytokine expression. | Erratic movements, periodic inactivity | Alters calcium ion homeostasis, impairs neural signalling. | Arman., 2023 |
| Fenvalerate | ↑ Apoptosis of hemocytes, ↑ malondialdehyde (MDA), ↓ total protein content. | Spontaneous shell withdrawal, hyperreactivity to stimuli | Inhibits ATPase, induces ROS-mediated damage to immune cells. | Mukherjee, S., & Mandal, C. ,2023 |
| Esfenvalerate | DNA fragmentation in hemocytes, ↓ immune gene expression. | Disorientation, shell spinning | Genotoxic and epigenetic modulation of immune pathways. | Werner, I., & Moran, K., 2008 |
Molecular Mechanisms Behind Pyrethroid-Induced Changes in Molluscs
Neurotoxic Mechanisms
Pyrethroids exert their toxic effects primarily through interference with voltage-gated sodium channels (VGSCs), leading to overstimulation of nerve cells (Saha et al., 2025). This mechanism is not only lethal but also affects various physiological processes that influence size, age, and sex. Chronic exposure to sublethal concentrations of pyrethroids can result in persistent neuronal hyperactivity, disrupting feeding, growth, and reproductive behaviours in molluscs (Major et al., 2020; Kotsyuba et al., 2023).
Induce Oxidative Stress
Oxidative stress is another significant mechanism by which pyrethroids affect molluscs. The production of reactive oxygen species (ROS) increases in response to pyrethroid exposure, leading to cellular damage and impairments in growth and longevity (San Juan et al., 2020). Biomarkers of oxidative stress, such as glutathione peroxidase and superoxide dismutase, have been used to assess pyrethroid-induced damage in molluscs (Arsalan, 2023). Pyrethroids generate excessive ROS, leading to oxidative damage to cellular components (Dutta et al., 2014). Antioxidant enzyme activity, including catalase and superoxide dismutase, is often modulated, reflecting a physiological response to oxidative stress (Saha et al., 2025). In Chilina parchappii, sublethal exposure to cypermethrin increased glutathione-S-transferase (GST) and glutathione peroxidase (GPx) activity, suggesting a protective role against oxidative damage (San Juan et al., 2020). Prolonged oxidative stress may result in lipid peroxidation, protein denaturation, and DNA fragmentation, cumulatively impairing molluscan health (López-Pedrouso et al., 2022) (Figure 3).
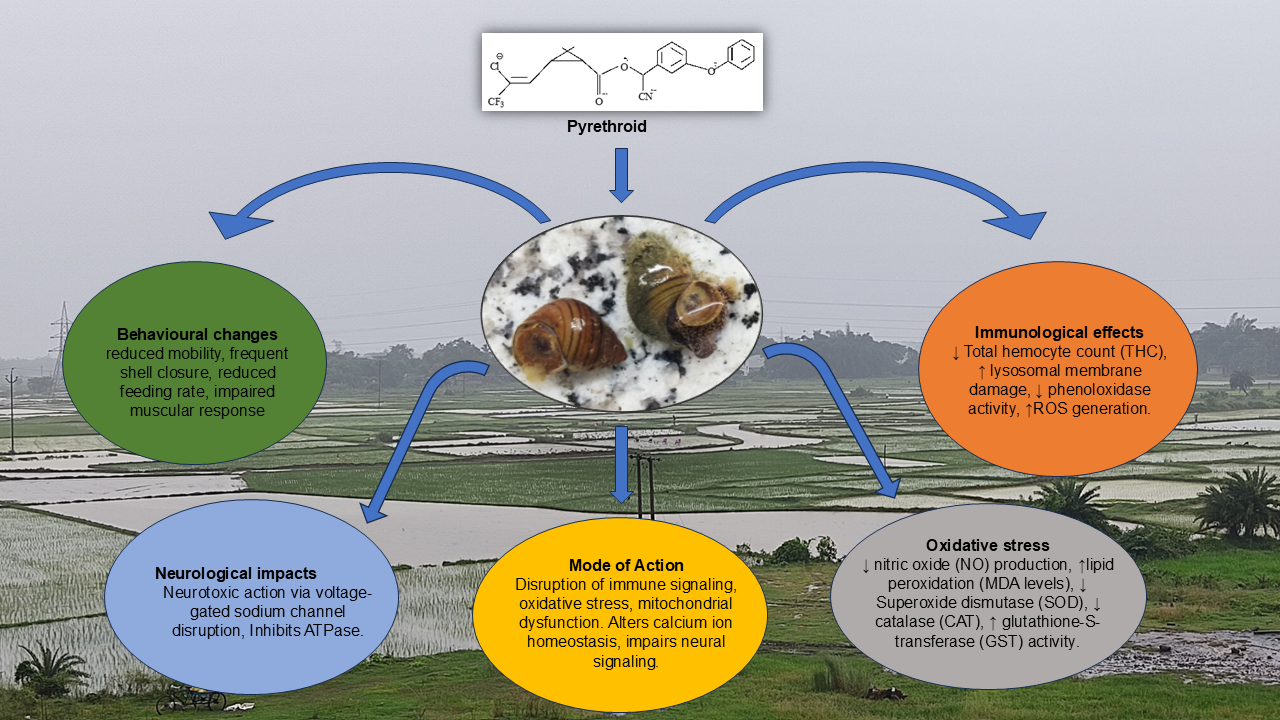
Figure 2.
Mode of action of pyrethroid on molluscs.

Figure 3.
Adverse effects of pyrethroid on molluscs.

Endocrine Disruption
Pyrethroids are known to interfere with endocrine systems in various species, and molluscs are no exception (Stemberg et al., 2010). The mechanism by which pyrethroids affect molluscan sex determination is thought to involve disruption of hormone systems, particularly those regulating sex differentiation and gametogenesis. Research has shown that exposure to cypermethrin can lead to skewed sex ratios in freshwater mussels (Lampsilis cardium), with more females than males being observed in treated populations (Hossain et al., 2023). In marine species such as Mytilus galloprovincialis (Mediterranean mussel), exposure to pyrethroids like deltamethrin has been associated with altered sexual maturation (Ait Ayad et al., 2011). Male mussels exposed to pyrethroids may exhibit delayed spermatogenesis, while females show abnormal oocyte development. Pyrethroid exposure can result in significant deviations from typical sexual dimorphism in molluscs (Major et al., 2020). In some species, exposure to pyrethroids can cause feminization of males, reduce reproductive success and lead to population imbalances. In addition to direct neurotoxicity, pyrethroids can disrupt hormonal systems involved in molluscan reproduction and growth. Alterations in estrogen and androgen signalling pathways can result in changes in sexual differentiation, maturation rates, and reproductive timing (Koene, 2010).
Pyrethroids and Molluscan Immunity
The immune system in molluscs relies on both cellular and humoral components. Hemocytes play a central role in phagocytosis, encapsulation, and production of reactive oxygen species (ROS), while humoral factors include antimicrobial peptides and enzymes such as phenol oxidase. Hemocytes play a pivotal role in molluscan immunity, engaging in phagocytosis, encapsulation, and pathogen elimination (Bergamini et al., 2023). Recent studies have highlighted, the diversity of hemocyte types and their functions across molluscan classes (De la Ballina et al., 2022). The role of cytokines, growth factors, and signalling pathways in hemocyte development.
Disrupt Haemocyte Function
Exposure to pyrethroids, such as permethrin and cypermethrin, reduces hemocyte viability and phagocytic activity (Impelliterri et al., 2022). Recent studies indicate that pyrethroids inhibit leukocyte proliferation and reduce immunoglobulin G (IgG) concentrations, impairing immune defences (Russo et al., 2020). Changes in hemocyte count and morphology have been observed, indicating immunosuppression. In Perna viridis, hemocytes exposed to deltamethrin showed altered lysosomal membrane stability, indicating cytotoxicity (Ray et al., 2013). Apoptosis, a critical mechanism in maintaining immune homeostasis, has been observed to regulate molluscan responses to pathogens and environmental stress (Romero et al., 2015). While its molecular pathways are partially understood, key regulators and signalling cascades remain poorly characterized. Altered expression of immune-related genes suggests disruption in signalling pathways critical for molluscan immunity. Reduced production of cytokines such as interleukin-2 (IL-2) and interferon-gamma (IFN-γ) has been documented, further highlighting immunosuppressive effects (Atta et al., 2024).
Histopathological Changes
Pyrethroid exposure can lead to damage in immune-related organs, such as the hepatopancreas and hemolymphatic tissues (Bernal-González et al., 2023). In bivalves like Crassostrea virginica, structural changes in hemocyte-producing tissues were observed after exposure to high pyrethroid concentrations (Ray et al., 2013). Environmental factors such as pollution and climate change significantly impact molluscan immunity (Morley, 2010). Studies reveal that pollutants impair hemocyte functionality and increase susceptibility to infections (Sun et al., 2023).
Ecological and Toxicological Implications
Impact on Molluscan Populations
The metabolism of pyrethroids in molluscs affects their survival and reproductive success (San Juan et al., 2020). Effective detoxification via metabolism can enhance the resilience of mollusc populations to pyrethroid exposure (Major et al., 2020). However, excessive or prolonged exposure can lead to metabolic overload, resulting in toxicity and population decline (Dhiman et al., 2021).
Delayed Growth and Reproduction
Studies have shown that the reproductive success of molluscs exposed to pyrethroids can be compromised, particularly if metabolic pathways are overwhelmed (Major et al., 2020). This may lead to smaller size and reduced fecundity in affected populations.
Implications for Molluscan Diversity
The sensitivity of molluscs to pyrethroid exposure, coupled with the environmental persistence of these compounds, suggests that pyrethroid contamination could lead to declines in molluscan diversity (Major et al., 2020). Given the ecological roles of molluscs, such as their contributions to nutrient cycling and as a food source for other wildlife, reductions in their populations could have cascading effects on aquatic ecosystems (Vaughn et al., 2018).
Impact on Molluscan Communication
Molluscs rely heavily on chemical cues for intra- and interspecific communication (Yu et al., 2023). Molluscan communication systems, including extracellular vesicles (e.g., exosomes) and tunnelling nanotubes (TNTs), are emerging areas of interest (Auguste et al., 2020). These systems are crucial for intercellular signalling and immune response coordination. Pesticides have been shown to disrupt these systems, impairing signal transmission and immune efficiency in molluscs (Bernal-González et al., 2023). Neurotransmitters such as serotonin and FMRFamide facilitate neuroimmune interactions, influencing molluscan cardiac and neural function (Li et al., 2019). Pesticides have been observed to interfere with these neurotransmitters, potentially impairing neuroimmune signalling.
Disruption of Pheromone Signalling
Pyrethroids can impair the synthesis and detection of pheromones, affecting mating behaviours and aggregation (San Juan et al., 2020). Disruption in pheromone communication in gastropods like Biomphalaria glabrata has led to decreased reproductive success (Jordaens et al., 2007). Altered chemical signalling due to pyrethroids may affect mating compatibility and hybridization rates in sympatric mollusc species (Major et al., 2020).
Interference with Alarm Cues
Exposure to sublethal concentrations of pyrethroids reduces molluscs’ ability to respond to predator-released chemical cues, increasing predation risk (Salice et al., 2013). In Lymnaea stagnalis, pyrethroid exposure resulted in impaired alarm response behaviour, suggesting sensory disruption (kowall et al., 2023). Alarm cue studies in Mytilus edulis suggest that pyrethroids may interfere with neural signal processing associated with predator detection (Ait Ayad et al., 2011).
Impairment of Olfactory Functions
Pyrethroids may affect the function of chemoreceptors, leading to altered sensory perception (Wei et al., 2021). Electrophysiological studies on Aplysia californica demonstrate that pyrethroids interfere with neural transmission in chemosensory pathways (Casida et al., 2013).
Pyrethroid Pollution Control
Pyrethroids, synthetic analogs of natural pyrethrins, are extensively utilized in agriculture and household pest control due to their high efficacy against insects and relatively low toxicity to mammals. However, their pervasive use has led to environmental contamination, necessitating effective strategies to mitigate pyrethroid pollution. Their persistence in the environment raises ecological concerns, particularly regarding their degradation and the control of their residual presence. To mitigate the environmental persistence of pyrethroids, several strategies can be employed. Implementing best management practices, such as precise application timing and methods, can minimize off-target deposition and reduce environmental contamination. Research into formulations that enhance the susceptibility of pyrethroids to environmental degradation can help reduce their persistence (Nkya et al., 2013). Utilizing plants capable of absorbing and metabolizing pyrethroids offers a potential method for decontaminating affected soils and water bodies (Cycoń et al., 2016). Enforcing regulations that limit pyrethroid usage in sensitive areas and promote the development of environmentally friendly alternatives can aid in controlling their environmental presence (Nkya et al., 2013). Regulatory agencies have implemented measures to address the ecological risks associated with pyrethroid use. The U.S. Environmental Protection Agency (EPA) has conducted systematic literature reviews to inform the registration and regulation of pyrethroid pesticides (Burns et al., 2018). Additionally, the EPA has developed ecological risk mitigation strategies for pyrethroids and pyrethrins. These strategies include label amendments to reduce runoff into water bodies, application restrictions to protect non-target organisms, and educational initiatives to promote best management practices among users. Implementing Integrated Pest Management (IPM) practices can also contribute to reducing pyrethroid pollution (Peshin et al., 2014). IPM emphasizes the use of non-chemical control methods, such as biological control agents, habitat manipulation, and cultural practices, to manage pest populations.
Discussion and Conclusions
Molluscs are a nutrient-dense, sustainable food source with substantial health, economic, ecological, and veterinary benefits. Their promotion can enhance global food security, support marine ecosystems, contribute essential nutrients to diverse diets, and play a critical role in monitoring and managing aquatic animal health (Fernández San Juan et al., 2020). Molluscs offer a unique perspective on the evolution of immune systems, communication networks, and behavioural adaptations. While significant progress has been made, substantial gaps remain in our understanding. Addressing these gaps, particularly the effects of pesticides on cellular communication, will not only advance molluscan biology but also provide broader insights into invertebrate ecology and evolution, with implications for conservation and biomedical research. Most studies focus on a few model species, neglecting the vast diversity within Mollusca. Expanding research to include lesser-studied groups can uncover novel adaptations. The molecular underpinnings of molluscan immune and communication systems remain incomplete. Advanced techniques like transcriptomics and proteomics could uncover key pathways and regulatory networks.
Pyrethroids pose significant risks to molluscan immunity, communication, and behaviour, with cascading effects on aquatic ecosystems. Recent findings underscore the multifaceted impacts of pyrethroids, including immunosuppression, oxidative stress, and histopathological changes (San Juan et al., 2020). Despite these advances, significant research gaps remain in understanding chronic and sublethal exposure effects, species-specific vulnerabilities, and ecosystem-level consequences. Limited data exist on the long-term impacts of chronic pyrethroid exposure on molluscan immunity. The interplay between immune suppression and increased susceptibility to pathogens under pyrethroid stress remains poorly understood. Species-specific variations in immune responses require further investigation to identify vulnerable taxa. The potential for epigenetic changes influencing immune functions remains unexplored. There is a lack of detailed studies on the molecular mechanisms underlying pyrethroid-induced disruptions in chemical signalling. Future research should prioritize these areas to inform risk assessment and mitigation strategies. Improved pesticide application practices and the development of less harmful alternatives are essential for safeguarding molluscan populations and their habitats. Pyrethroids exert their insecticidal action by targeting sodium channels and disrupting normal nerve function. Understanding these mechanisms is critical for optimizing their use, managing resistance, and minimizing non-target environmental impacts. While pyrethroids offer benefits in pest management, their potential adverse effects on non-target aquatic organisms, including molluscs, warrant careful consideration. Further research is needed to fully understand the extent of these impacts and to develop strategies that mitigate harm to molluscan diversity while maintaining effective pest control.
Molluscs are integral to the health and stability of aquatic ecosystems. Their roles in nutrient cycling, habitat formation, and food web dynamics underscore their ecological importance. However, mounting environmental pressures threaten their populations and, by extension, ecosystem resilience. The ecological consequences of impaired communication on population dynamics remain unexplored. More research is needed to assess the reversibility of communication impairments after cessation of pyrethroid exposure. Behavioural responses to combined stressors, including pyrethroids and other pollutants, are poorly documented. The thresholds for sublethal behavioural changes that lead to population-level consequences need further exploration. Studies on the potential recovery of normal behaviours post-exposure are limited. Comprehensive studies on how pyrethroid exposure impacts ecosystem-level processes are lacking. Long-term monitoring of molluscan populations in contaminated habitats is needed to assess recovery potential and resilience.
The synergistic effects of pyrethroid exposure and other environmental stressors such as heavy metals, temperature extremes, hypoxia, and food scarcity have profound implications for the health and survival of gastropod molluscs. These interactions amplify the toxic effects of pyrethroids, leading to oxidative stress, neurological dysfunction, and reproductive failure, which in turn affect behavioural responses, fitness, and population dynamics. As gastropods are key players in aquatic ecosystems, their behavioural and physiological impairments can disrupt food webs, nutrient cycling, and ecosystem services. Therefore, it is crucial to consider the combined effects of multiple environmental stressors when assessing the ecological risks posed by pyrethroids and other pollutants. Future research should focus on field-based studies and long-term ecological monitoring to better understand the real-world interactions between pyrethroids, environmental stressors, and gastropod populations. This will provide more accurate predictions of the cumulative impacts on aquatic ecosystems and guide the development of more sustainable pest management practices.
Effective management, conservation strategies, and pollution control are crucial to preserving molluscan biodiversity and their ecological functions. Addressing pyrethroid pollution requires a multifaceted approach that combines microbial bioremediation, stringent regulatory measures, and sustainable pest management practices. Ongoing research and collaboration among scientists, regulatory agencies, and stakeholders are essential to develop and implement effective strategies for controlling pyrethroid pollution and safeguarding environmental health. Understanding the degradation pathways and persistence factors of pyrethroids is crucial for developing effective strategies to control their environmental presence. Combining optimized application practices, innovative degradation-enhancing formulations, biological remediation techniques, and regulatory measures can collectively mitigate the ecological risks associated with pyrethroid persistence. Molluscs are vital to ecological resilience and human well-being. Their contributions to nutrient cycling, habitat, food web processes, and human economies emphasize their role in maintaining balance between nature and human society. Sustaining conservation and management is necessary to protect molluscan populations and the essential services they offer.
Funding
None.
Data and Materials Availability
Every dataset that was examined for this study is openly accessible to the public.
Conflicts of Interest
The writers say they have no competing interests.
Ethical Approval
Not applicable.
Code Availability
No software is used for writing this manuscript.
References
- Ait Ayad, M.; Ait Fdil, M.; Mouabad, A. Effects of cypermethrin (pyrethroid insecticide) on the valve activity behavior, byssal thread formation, and survival in air of the marine mussel Mytilus galloprovincialis. Archives of environmental contamination and toxicology 2011, 60(3), 462–470. [Google Scholar] [CrossRef]
- Atta, S. A.; Fahmy, Z. H.; Selim, E. A.; Aboushousha, T.; Mostafa, R. R. Effect of linex treatment on IFN-γ and IL-4 in mice infected with Trichinella. BMC Infectious Diseases 2024, 24(1), 1360. [Google Scholar] [CrossRef] [PubMed]
- Arman, S. Effects of Single and Combined Exposure to Environmentally Relevant Concentrations of Cyfluthrin and Copper on Digestive Gland Histology of Lymnaea stagnalis. Sakarya University Journal of Science 2023, 27(1), 39–48. [Google Scholar] [CrossRef]
- Arslan, P. Pyrethroid-induced oxidative stress and biochemical changes in the primary mussel cell cultures. Environmental Science and Pollution Research 2023, 30(16), 48484–48490. [Google Scholar] [CrossRef]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canesi, L. Conservation of cell communication systems in invertebrate host–defence mechanisms: possible role in immunity and disease. Biology 2020, 9(8), 234. [Google Scholar] [CrossRef]
- Australian Industrial Chemicals Introduction Scheme (AICIS). Available online: https://services.industrialchemicals.gov.au/search-inventory/.
- Aznar-Alemany, Ò.; Piotrowska-Seget, M.Z. Introduction to pyrethroid insecticides: chemical structures, properties, mode of action and use. In Pyrethroid insecticides (pp. 1-16). Cham: Springer International Publishing. https://doi.org/10.1007/698_2019_435 Cycoń, M.; Piotrowska-Seget, Z. (2016). Pyrethroiddegrading microorganisms and their potential for the bioremediation of contaminated soils: a review. Frontiers in microbiology 2020, 7, 1463. [Google Scholar] [CrossRef]
- Bacqué-Cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J. P.; Bouguiyoud, N.; Di Giovanni, G.; De Deurwaerdère, P. Serotonin in animal cognition and behavior. International journal of molecular sciences 2020, 21(5), 1649. [Google Scholar] [CrossRef]
- Bayne, B. L. Phenotypic flexibility and physiological tradeoffs in the feeding and growth of marine bivalve molluscs. Integrative and Comparative Biology 2004, 44(6), 425–432. [Google Scholar] [CrossRef]
- Benjamin, P. R.; Kemenes, G. Lymnaea learning and memory. Scholarpedia 2010, 5(8), 4247. [Google Scholar] [CrossRef]
- Benjamin, P.; Crossley, M. Gastropod feeding systems: Evolution of neural circuits that generate diverse behaviors. In Oxford Research Encyclopedia of Neuroscience; 2020. [Google Scholar] [CrossRef]
- Bergamini, G.; Sacchi, S.; Ferri, A.; Franchi, N.; Montanari, M.; Ahmad, M.; Malagoli, D. Clodronate liposome-mediated phagocytic hemocyte depletion affects the regeneration of the cephalic tentacle of the invasive snail, Pomacea canaliculata. Biology 2023, 12(7), 992. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H. R. Mind versus metabolism in the control of food intake and energy balance. Physiology & behavior 2004, 81(5), 781–793. [Google Scholar] [CrossRef]
- Bernal-González, K. G.; Covantes-Rosales, C. E.; Camacho-Pérez, M. R.; Mercado-Salgado, U.; Barajas-Carrillo, V. W.; Girón-Pérez, D. A.; Girón-Pérez, M. I. Organophosphate-pesticide-mediated immune response modulation in invertebrates and vertebrates. International Journal of Molecular Sciences 2023, 24(6), 5360. [Google Scholar] [CrossRef]
- Boutet, I.; Tanguy, A.; Moraga, D. Molecular identification and expression of two non-P450 enzymes, monoamine oxidase A and flavin-containing monooxygenase 2, involved in phase I of xenobiotic biotransformation in the Pacific oyster, Crassostrea gigas. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression 2004, 1679(1), 29–36. [Google Scholar] [CrossRef]
- Braun, M. H.; Lukowiak, K.; Karnik, V.; Lukowiak, K. Differences in neuronal activity explain differences in memory forming abilities of different populations of Lymnaea stagnalis. Neurobiology of Learning and Memory 2012, 97(1), 173–182. [Google Scholar] [CrossRef]
- Burns, C. J.; Pastoor, T. P. Pyrethroid epidemiology: a quality-based review. Critical reviews in toxicology 2018, 48(4), 297–311. [Google Scholar] [CrossRef]
- Casida, J. E.; Durkin, K. A. Neuroactive insecticides: targets, selectivity, resistance, and secondary effects. Annual review of entomology 2013, 58(1), 99–117. [Google Scholar] [CrossRef]
- Chen, C. H. Phase II Enzymes. In Activation and Detoxification Enzymes: Functions and Implications; Springer New York: New York, NY, 2011; pp. 37–48. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-degrading microorganisms and their potential for the bioremediation of contaminated soils: a review. Frontiers in microbiology 2016, 7, 1463. [Google Scholar] [CrossRef] [PubMed]
- Deidda, I.; Russo, R.; Bonaventura, R.; Costa, C.; Zito, F.; Lampiasi, N. Neurotoxicity in marine invertebrates: an update. Biology 2021, 10(2), 161. [Google Scholar] [CrossRef] [PubMed]
- De la Ballina, N. R.; Maresca, F.; Cao, A.; Villalba, A. Bivalve haemocyte subpopulations: a review. Frontiers in immunology 2022, 13, 826255. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, V.; Pant, D. Environmental biomonitoring by snails. Biomarkers 2021, 26(3), 221–239. [Google Scholar] [CrossRef]
- Di Cosmo, A.; Winlow, W.; Farag, M. R.; Alagawany, M.; Bilal, R. M.; Gewida, A. G.; Dhama, K.; Abdel-Latif, H. M.; Naiel, M. A.; Neuroecology and neuroethology in Molluscs The interface between behaviour and environment (pp. 1-239). NOVA PUBLISHER. An overview on the potential hazards of pyrethroid insecticides in fish, with special emphasis on cypermethrin toxicity. Animals 2014, 11(7), 1880. [Google Scholar] [CrossRef]
- Dutta, S. M.; Mustafi, S. B.; Raha, S.; Chakraborty, S. K. Assessment of thermal stress adaptation by monitoring Hsp70 and MnSOD in the freshwater gastropod, Bellamya bengalensis (Lamark 1882). Environmental monitoring and assessment 2014, 186(12), 8961–8967. [Google Scholar] [CrossRef]
- Fernández San Juan, M. R.; Cortelezzi, A.; Albornoz, C. B.; Landro, S. M.; Arrighetti, F.; Najle, R.; Lavarias, S. M. L. Ecotoxicology and environmental safety toxicity of pyrethroid cypermethrin on the freshwater snail Chilina parchappii: Lethal and sublethal effects. 2020. [Google Scholar] [CrossRef]
- Gillette, R.; Brown, J. W. The sea slug, Pleurobranchaea californica: a signpost species in the evolution of complex nervous systems and behavior. Integrative and Comparative Biology 2015, 55(6), 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Gökoğlu, N. Molluscan shellfish. In Shellfish processing and preservation; Springer International Publishing: Cham, 2021; pp. 129–250. [Google Scholar] [CrossRef]
- Griscom, S. B.; Fisher, N. S. Bioavailability of sediment-bound metals to marine bivalve molluscs: an overview. Estuaries 2004, 27(5), 826–838. [Google Scholar] [CrossRef]
- Hazardous Substances Data Bank (HSDB). Available online: https://pubchem.ncbi.nlm.nih.gov/source/hsdb/1511.
- Hossain, M. A.; Sarker, T. R.; Sutradhar, L.; Hussain, M.; Iqbal, M. M. Toxic effects of chlorpyrifos on the growth, hemocytes counts, and vital organ’s histopathology of freshwater mussel, Lamellidens marginalis. Journal of King Saud University-Science 2023, 35(2), 102482. [Google Scholar] [CrossRef]
- ILO-WHO International Chemical Safety Cards (ICSCs). Available online: https://www.ilo.org/dyn/icsc/showcard.display?p_version=2&p_card_id=0212.
- Impellitteri, F.; Curpăn, A. S.; Plăvan, G.; Ciobica, A.; Faggio, C. Hemocytes: a useful tool for assessing the toxicity of microplastics, heavy metals, and pesticides on aquatic invertebrates. International journal of environmental research and public health 2022, 19(24), 16830. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, H. H.; Qadir, A.; Ahmad, S. R.; Riaz, M. A.; Riaz, A.; Shahid, N.; Arslan, M. Residual assessment of emerging pesticides in aquatic sinks of Lahore, Pakistan. Sustainability 2024, 16(21), 9257. [Google Scholar] [CrossRef]
- Jaggi, R. C.; Aulakh, M. S.; Sharma, R. Impacts of elemental S applied under various temperature and moisture regimes on pH and available P in acidic, neutral and alkaline soils. Biology and Fertility of Soils 2005, 41(1), 52–58. [Google Scholar] [CrossRef]
- Jiang, Q.; Ao, S.; Ji, P.; Zhou, Y.; Tang, H.; Zhou, L.; Zhang, X. Assessment of deltamethrin toxicity in Macrobrachium nipponense based on histopathology, oxidative stress and immunity damage. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2021, 246, 109040. [Google Scholar] [CrossRef]
- Jordaens, K.; Dillen, L.; Backeljau, T. Effects of mating, breeding system and parasites on reproduction in hermaphrodites: pulmonate gastropods (Mollusca). Animal Biology 2007, 57(2), 137–195. [Google Scholar] [CrossRef]
- Jordaens, K.; Dillen, L.; Backeljau, T. Shell shape and mating behaviour in pulmonate gastropods (Mollusca). Biological Journal of the Linnean Society 2009, 96(2), 306–321. [Google Scholar] [CrossRef]
- Kamrin, M. A.; Katagi, T.; Pesticide profiles: toxicity; environmental impact; and fate. Photodegradation of pesticides on plant and soil surfaces. In Reviews of environmental contamination and toxicology: Continuation of residue reviews; CRC press, 1997; pp. 1–78. [Google Scholar] [CrossRef]
- Kaneko, H. Pyrethroid chemistry and metabolism. In Hayes’ handbook of pesticide toxicology; Academic press, 2010; pp. 1635–1663. [Google Scholar] [CrossRef]
- Katagi, T. Bioconcentration, bioaccumulation, and metabolism of pesticides in aquatic organisms. Reviews of environmental contamination and toxicology 2009, 1–132. [Google Scholar] [CrossRef]
- Kiss, T. Do terrestrial gastropods use olfactory cues to locate and select food actively? Invertebrate Neuroscience 2017, 17(3), 9. [Google Scholar] [CrossRef]
- Koene, J. M. Neuro-endocrine control of reproduction in hermaphroditic freshwater snails: mechanisms and evolution. Frontiers in behavioral neuroscience 2010, 4, 167. [Google Scholar] [CrossRef]
- Kotsyuba, E.; Dyachuk, V. Role of the neuroendocrine system of marine bivalves in their response to hypoxia. International Journal of Molecular Sciences 2023, 24(2), 1202. [Google Scholar] [CrossRef]
- Kowall, C. A.; Batabyal, A.; Lukowiak, K.; Phillips, I. D. Agricultural use of insecticides alters homeostatic behaviors and cognitive ability in Lymnaea stagnalis. Environmental Toxicology and Chemistry 2023, 42(11), 2466–2477. [Google Scholar] [CrossRef]
- Landrum, P. F.; Robbins, J. A. Bioavailability of sediment-associated contaminants to benthic invertebrates. In Sediments; CRC Press, 2020; pp. 237–263. [Google Scholar]
- Langston, W. J.; Pope, N. D. Molluscs. In Ecotoxicology of Antifouling biocides; Springer Japan: Tokyo, 2009; pp. 271–289. [Google Scholar] [CrossRef]
- Leung, K. M.; Grist, E. P.; Morley, N. J.; Morritt, D.; Crane, M. Chronic toxicity of tributyltin to development and reproduction of the European freshwater snail Lymnaea stagnalis (L.). Chemosphere 2007, 66(7), 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, X.; Kong, L.; Yu, H. J. I. S. J. Transcriptomic response to stress in marine bivalves. Invertebrate survival journal 2013, 10(1), 84–93. [Google Scholar]
- Li, M.; Wang, M.; Wang, W.; Wang, L.; Liu, Z.; Sun, J.; Song, L. The immunomodulatory function of invertebrate specific neuropeptide FMRFamide in oyster Crassostrea gigas. Fish & shellfish immunology 2019, 88, 480–488. [Google Scholar] [CrossRef]
- Lodi, M.; Koene, J. M. The love-darts of land snails: integrating physiology, morphology and behaviour. Journal of Molluscan Studies 2016, 82(1), 1–10. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Lorenzo, J. M.; Varela, Z.; Fernández, J. Á.; Franco, D. Finding biomarkers in antioxidant molecular mechanisms for ensuring food safety of bivalves threatened by marine pollution. Antioxidants 2022, 11(2), 369. [Google Scholar] [CrossRef]
- Major, K. M.; Brander, S. M. The ecological and evolutionary implications of pyrethroid exposure: a new perspective on aquatic ecotoxicity. In Pyrethroid insecticides; Springer International Publishing: Cham, 2020; pp. 109–148. [Google Scholar] [CrossRef]
- Matthews, R. W.; Matthews, J. R. Foraging and feeding. In Insect Behavior: 2nd Edition; Springer Netherlands: Dordrecht, 2009; pp. 131–184. [Google Scholar] [CrossRef]
- Méjanelle, L.; Jara, B.; Dachs, J. Fate of pyrethroids in freshwater and marine environments. In Pyrethroid insecticides; Springer International Publishing: Cham, 2020; pp. 81–107. [Google Scholar] [CrossRef]
- Middlebrooks, M. L.; Pierce, S. K.; Bell, S. S. Foraging behavior under starvation conditions is altered via photosynthesis by the marine gastropod, Elysia clarki. PLoS One 2011, 6(7), e22162. [Google Scholar] [CrossRef] [PubMed]
- Miller, M. W. Dopamine as a multifunctional neurotransmitter in gastropod molluscs: an evolutionary hypothesis. The Biological Bulletin 2020, 239(3), 189–208. [Google Scholar] [CrossRef] [PubMed]
- Morley, N. J. Interactive effects of infectious diseases and pollution in aquatic molluscs. Aquatic toxicology 2010, 96(1), 27–36. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mandal, C.; Ngandjui, Y. A. T.; Kereeditse, T. T.; Kamika, I.; Madikizela, L. M.; Msagati, T. A. M.; Phagocytic response and phenoloxidase activity of the hemocytes of Bellamya bengalensis exposed to synthetic fenvalerate. Nutraceutical and medicinal importance of marine molluscs. Journal of Stress Physiology & Biochemistry;Marine Drugs 2023, 19(3) 22(5), 143–151 201. [Google Scholar] [CrossRef]
- Ng, T. P.; Saltin, S. H.; Davies, M. S.; Johannesson, K.; Stafford, R.; Williams, G. A. Snails and their trails: the multiple functions of trail-following in gastropods. Biological Reviews 2013, 88(3), 683–700. [Google Scholar] [CrossRef]
- Nkya, T. E.; Akhouayri, I.; Kisinza, W.; David, J. P. Impact of environment on mosquito response to pyrethroid insecticides: facts, evidences and prospects. Insect biochemistry and molecular biology 2013, 43(4), 407–416. [Google Scholar] [CrossRef]
- Peshin, R.; Zhang, W. Integrated pest management and pesticide use. In Integrated Pest Management: Pesticide Problems; Springer Netherlands: Dordrecht, 2014; Vol. 3, pp. 1–46. [Google Scholar] [CrossRef]
- Petushok, N.; Gabryelak, T.; Pałecz, D.; Zavodnik, L.; Varga, I. S.; Deér, K. A. Comparative study of the xenobiotic metabolising system in the digestive gland of the bivalve molluscs in different aquatic ecosystems and in aquaria experiments. Aquatic toxicology 2002, 61(1-2), 65–72. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/.
- Quiroz-Barroso, S. A.; Guerrero-Arenas, R.; García-Barrera, P.; Sánchez-Beristain, F. Mollusca: Bivalvia and Gastropoda. In Past Environments of Mexico: Unveiling the Past Environments of a Megadiverse Country Through its Fossil Record; Springer Nature Switzerland: Cham, 2024; pp. 63–97. [Google Scholar] [CrossRef]
- Ray, M.; Bhunia, A. S.; Bhunia, N. S.; Ray, S. Density shift, morphological damage, lysosomal fragility and apoptosis of hemocytes of Indian molluscs exposed to pyrethroid pesticides. Fish & shellfish immunology 2013, 35(2), 499–512. [Google Scholar] [CrossRef]
- Romano, R. A. Feeding Biology of the Aeolid Nudibranch Flabellina verrucosa; University of New Hampshire, 2022. [Google Scholar]
- Romero, A.; Novoa, B.; Figueras, A. The complexity of apoptotic cell death in mollusks: An update. Fish & Shellfish Immunology 2015, 46(1), 79–87. [Google Scholar] [CrossRef]
- Russo, J.; Lagadic, L. Effects of parasitism and pesticide exposure on characteristics and functions of hemocyte populations in the freshwater snail Lymnaea palustris (Gastropoda, Pulmonata). Cell biology and toxicology 2000, 16(1), 15–30. [Google Scholar] [CrossRef]
- Saha, R.; Dutta, S. M. Pesticides’ mode of action on aquatic life. Toxicology Reports 2024, 13, 101780. [Google Scholar] [CrossRef]
- Saha, R.; Dutta, S. M. Pyrethroids have become a barrier to the daily existence of molluscs. Journal of Hazardous Materials Letters 2025, 100144. [Google Scholar] [CrossRef]
- Sakakibara, M. Cellular and molecular aspects of short-term and long-term memory from molluscan systems. In Novel Trends in Brain Science: Brain Imaging, Learning and Memory, Stress and Fear, and Pain; Springer Japan: Tokyo, 2008; pp. 131–148. [Google Scholar] [CrossRef]
- Salice, C. J.; Kimberly, D. A. Environmentally relevant concentrations of a common insecticide increase predation risk in a freshwater gastropod. Ecotoxicology 2013, 22(1), 42–49. [Google Scholar] [CrossRef] [PubMed]
- Samia, B.; Socorro, J.; Durand, A.; Quivet, E.; Wortham, H. Photolytic degradation of commonly used pesticides adsorbed on silica particles. Science of the Total Environment 2024, 949, 174964. [Google Scholar] [CrossRef]
- San Juan, M. F.; Cortelezzi, A.; Albornoz, C. B.; Landro, S. M.; Arrighetti, F.; Najle, R.; Lavarías, S. M. L. Toxicity of pyrethroid cypermethrin on the freshwater snail Chilina parchappii: Lethal and sublethal effects. Ecotoxicology and environmental safety 2020, 196, 110565. [Google Scholar] [CrossRef]
- Shan, Y.; Yan, S.; Hong, X.; Zha, J.; Qin, J. Effect of imidacloprid on the behavior, antioxidant system, multixenobiotic resistance, and histopathology of Asian freshwater clams (Corbicula fluminea). Aquatic Toxicology 2020, 218, 105333. [Google Scholar] [CrossRef] [PubMed]
- Sogorb, M. A.; Vilanova, E. Enzymes involved in the detoxification of organophosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicology letters 2002, 128(1-3), 215–228. [Google Scholar] [CrossRef]
- Sternberg, R. M.; Gooding, M. P.; Hotchkiss, A. K.; LeBlanc, G. A. Environmental-endocrine control of reproductive maturation in gastropods: implications for the mechanism of tributyltin-induced imposex in prosobranchs. Ecotoxicology 2010, 19(1), 4–23. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Teng, J.; Wang, D.; Zhao, J.; Shan, E.; Wang, Q. The adverse impact of microplastics and their attached pathogen on hemocyte function and antioxidative response in the mussel Mytilus galloprovincialis. Chemosphere 2023, 325, 138381. [Google Scholar] [CrossRef]
- Tamvacakis, A. N.; Senatore, A.; Katz, P. S. Single neuron serotonin receptor subtype gene expression correlates with behaviour within and across three molluscan species. Proceedings of the Royal Society B 2018, 285(1885), 20180791. [Google Scholar] [CrossRef]
- Tripathi, P. K.; Singh, A. Carbaryl induced alterations in the reproduction and metabolism of freshwater snail Lymnaea acuminata. Pesticide biochemistry and physiology 2004, 79(1), 1–9. [Google Scholar] [CrossRef]
- Toxin and Toxin Target Database (T3DB). Available online: http://www.t3db.ca/toxins/T3D1843.
- USEPA/Office of Pesticide Programs. Reregistration Eligibility Decision for Allethrins p.5 EPA 738-R-07001 (June 2007). 7 June 2008. Available online: https://www.epa.gov/pesticides/reregistration/status.htm.
- Vaughn, C. C.; Hoellein, T. J. Bivalve impacts in freshwater and marine ecosystems. Annual review of ecology, evolution, and systematics 2018, 49(1), 183–208. [Google Scholar] [CrossRef]
- Wei, S.; Xu, T.; Jiang, T.; Yin, D. Chemosensory dysfunction induced by environmental pollutants and its potential as a novel neurotoxicological indicator: a review. Environmental science & technology 2021, 55(16), 10911–10922. [Google Scholar] [CrossRef]
- Werner, I.; Moran, K. Effects of pyrethroid insecticides on aquatic organisms. Synthetic pyrethroids: Occurrence and behavior in aquatic environments 2008, 991, 310–335. [Google Scholar]
- Wu, M.; Tang, X.; Sun, C.; Miao, J.; Wang, Q.; Pan, L. Kinetics of uptake and depuration of synthetic pyrethroid insecticides in manila clam (Ruditapes philippinarum). Environmental Science and Pollution Research 2023, 30(30), 76246–76252. [Google Scholar] [CrossRef]
- Wyeth, R. C.; Woodward, O. M.; Willows, A. D. Orientation and navigation relative to water flow, prey, conspecifics, and predators by the nudibranch mollusc Tritonia diomedea. The Biological Bulletin 2006, 210(2), 97–108. [Google Scholar] [CrossRef]
- Yu, Z. L.; Yang, M. J.; Song, H.; Zhang, T.; Yuan, X. T. Gastropod chemoreception behaviors—Mechanisms underlying the perception and location of targets and implications for shellfish fishery development in aquatic environments. Frontiers in Marine Science 2023, 9, 1042962. [Google Scholar] [CrossRef]
- Zha, C.; Sossin, W. S. The molecular diversity of plasticity mechanisms underlying memory: An evolutionary perspective. Journal of neurochemistry 2022, 163(6), 444–460. [Google Scholar] [CrossRef]
Table 1.
Pearson correlation table for different type of pyrethroids. (Sources: USEPA https://www.epa.gov/pesticides/reregistration/status.htm; ILO-WHO International Chemical Safety Cards (ICSCs) https://www.ilo.org/dyn/icsc/showcard.display?p_version=2&p_card_id=0212; Hazardous Substances Data Bank (HSDB) https://pubchem.ncbi.nlm.nih.gov/source/hsdb/1511; Australian Industrial Chemicals Introduction Scheme (AICIS) https://services.industrialchemicals.gov.au/search-inventory/; Toxin and Toxin Target Database (T3DB) http://www.t3db.ca/toxins/T3D1843 and PubChem https://pubchem.ncbi.nlm.nih.gov/).
Table 1.
Pearson correlation table for different type of pyrethroids. (Sources: USEPA https://www.epa.gov/pesticides/reregistration/status.htm; ILO-WHO International Chemical Safety Cards (ICSCs) https://www.ilo.org/dyn/icsc/showcard.display?p_version=2&p_card_id=0212; Hazardous Substances Data Bank (HSDB) https://pubchem.ncbi.nlm.nih.gov/source/hsdb/1511; Australian Industrial Chemicals Introduction Scheme (AICIS) https://services.industrialchemicals.gov.au/search-inventory/; Toxin and Toxin Target Database (T3DB) http://www.t3db.ca/toxins/T3D1843 and PubChem https://pubchem.ncbi.nlm.nih.gov/).
| Pesticide | Molecular formula and molecular weight | xLogP3 | log Kow | Solubility and density | Applications | EPA Toxicity |
| Allethrin | C19H26O3 ; 302.4 g/mol |
4.8 | 4.78 | Solubility in water: none Relative density (water = 1): 1.01 |
Household and agricultural insecticide | LD50 Rat female oral 685 mg/kg; LD50 Mallard duck, oral <2000 mg/kg |
| Bifenthrin | C23H22ClF3O2 ; 422.9 g/mol |
6 | 6.00 | water <1.0X10-3 mg/L/ at 20 °C Density: 1.2 g/cu m at 125 °C |
Household and agricultural insecticide | LD50 Rat oral 54.5 mg/kg; LD50 Quail oral 18 mg/kg |
| Permethrin | C21H20Cl2O3 ; 391.3 g/mol | 6.5 | 6.5 | water, 6.00X10-3 mg/L (pH 7, 20 °C); cis-isomers 0.20 mg/L (25 °C); trans-isomers 0.13 mg/L (25 °C); density: 1.19–1.27 at 20 °C |
Household insecticide | LC50 Brook Trout 2.3; LC50 Channel Catfish 7.2 |
| Phenothrin | C23H26O3 ; 350.4 g/mol | 6.2 | 6.01 | Density: 1.06 at 20 °C; In methanol, >5.0 g/mL; hexane, >4.96 g/mL at 25 °C; In water, <9.7 ug/L at 25 °C |
Agricultural insecticide | LD50 Rat >10mg/kg; LD50 Quail >2150 mg/kg |
| Resmethrin | C22H26O3 ; 338.4 g/mol | 6.1 | 5.43 | Density:0.958-0.968 at 20 °C; In water, 0.0379 mg/L at 25 °C |
Veterinary pesticide and household insecticide | LD50: > 5000 mg/kg (Oral, Rat); |
| Tefluthrin | C17H14ClF7O2 ; 418.7 g/mol | 5.4 | 6.4 | Density: 1.48 g/mL (25 °C); In acetone, hexane, toluene, dichloromethane, ethylacetate: >500; methanol 263 (all in g/L, 21 °C); In water, 0.02 mg/L at 20 °C |
Agricultural insecticide | LD50: 22 mg/kg (Oral, Rat); LD50: Duck oral 4190 mg/kg |
| Tetramethrin | C19H25NO4 ; 331.4 g/mol | 4.7 | 4.73 | Density: 1.108 at 20 °C/20 °C; Methanol (53 g/kg), hexane (20 g/kg), xylene (1 g/kg), acetone, toluene. ; In water, 1.83 mg/L at 25 °C. |
Agricultural insecticide and household insecticide | LD50: 5000 mg/kg (Oral, Rat); |
| Metofluthrin | C18H20F4O3 ; 360.3 g/mol | 3.7 | 4.97-5.03 at 25 °C | Density: 1.21 at 20 °C; Solvent solubility (20 to 25 °C; g/L): acetone, methanol, ethyl acetate, toluene, n-hexanes, dichloromethane, n-octanol, isopropyl alcohol; In water, 0.73 mg/L at pH 7, 20 °C) |
Insecticides, mosquito control. | LD50 Rat oral > 2,000 mg/kg |
| Transfluthrin | C15H12Cl2F4O2 ; 371.2 g/mol | 5.0 | 5.46 at 20 °C | Density:1.51g/cm3(23C); In water is less than 1.13 mg/L at 20 °C. In organic solvents such as heptane, p-xylene, 1,2-dichloroethane,methanol, acetone, and ethyl acetate. |
Household insecticide | LC50 Rainbow Trout 0.71 PPB; LC50 Water flea 0.0686 PPB; LC50 Bob white quail 4909 PPM |
| Cyfluthrin | C22H18Cl2FNO3 ; 434.3 g/mol | 6.2 | 5.95 | >1000 g/L dichloromethane; In water, 3.0X10-3 mg/L at 20 °C; Density: 1.34 g/cu cm at 22 °C |
Agricultural insecticide | LD50: 869-1271 mg/kg (Oral, Rat); LD50: >5000 mg/kg (Quail) |
| Cyhalothrin | C23H19ClF3NO3 ; 449.8 g/mol | 6.1 | 6.8 | Acetone, dichloromethane, methanol, diethyl ether, ethyl acetate, hexane, toluene, In water, 5.0X10-3 mg/L, temp not specified; Density: 1.25 at 25 °C |
Agricultural insecticide | LD50: 144-243 mg/kg (Oral, Rat); LD50: >5000 mg/kg for Duck. |
| Cypermethrin | C22H19Cl2NO3 ; 416.3 g/mol | 6.0 | 6.60 | Acetone, chloroform, cyclohexanone, xylene, ethanol, hexane; In water, 4X10-3 mg/L at 20 °C; Density: 1.25 g/cu cm at 20 °C |
Agricultural insecticide and household insecticide | LD50: 250-300 mg/kg (Oral, Mouse); LD50: 400 mg/kg for hamster |
| Deltamethrin | C22H19Br2NO3 ; 505.2 g/mol | 6.2 | 6.20 | Cyclohexanone, dichloromethane, acetone, benzene, dimethyl sulphoxide, xylene, isopropanol; In water, <0.002 mg/L at 25 °C Relative density (water = 1): 0.5 |
Agricultural insecticide and household insecticide | LD50: 4123 mg/kg (Oral, Rat); >4640 mg/kg for Duck |
| Fenpropathrin | C22H23NO3 ; 349.4 g/mol | 5.7 | 6.0 at 20 °C | Xylene, cyclohexanone, Methanol; In water, 1.41X10-2 mg/l at 25 °C. Density: 1.15 at 25 °C |
Veterinary pesticides | LD50: 70.6 mg/kg (Oral, Rat); 1089 mg/kg for duck |
| Fenvalerate | C25H22ClNO3 ; 419.9 g/mol | 6.2 | 6.20 | Solubility at 20 °C (g/L): acetone >450; chloroform >450; methanol >450; hexane 77; In water, 2.4X10-2 mg/L at 22 °C (seawater); Density: 1.17 at 23 °C/4 °C |
Agricultural insecticide | LD50: 70.2 mg/kg (Oral, Rat); >4000 mg/kg for Quail. |
| Flumethrin | C28H22Cl2FNO3 ; 510.4 g/mol | 7.6 | 6.2 | solubility in pure water reported as 0.1 µg/L for one isomer and 0.2 µg/L for the sum of both isomers; in acetone, xylene, methanol, acetonitrile, ethylacetate, and dimethylsulfoxide; density: 1.342 g/cm³ |
Veterinary pesticide | LD50: > 100 mg/kg (Oral, Rat); |
| Fluvalinate | C26H22ClF3N2O3 ; 502.9 g/mol | 7.7 | 7.02 | Solubility in water, 0.012 mg/L at 25 °C ; density: 1.29 g/cu cm at 25 °C |
Agricultural insecticide and Veterinary pesticide | LD50: 261-282 mg/kg (Oral, Rat); >2510 mg/kg for quail |
| Tralomethrin | C22H19Br4NO3 ; 665.0 g/mol | 8.0 | approximately 5 at 25 °C | In water, 0.8 mg/L; In acetone, dichloromethane, toluene, xylene >1000; dimethyl sulfoxide >500; ethanol >180 (all in g/L); density: 1.70 at 20 °C |
Agricultural insecticide | LD50: 1250 mg/kg (Oral, Rat); >2150 mg/kg for quail |
| Esfenvalerate | C25H22ClNO3; 419.9 g/mol | 6.2 | 6.22 | xylene, acetone, chloroform, ethyl acetate, dimethylformamide, dimethyl sulfoxide, hexane, methanol; In water, 2.0X10-3 mg/L at 25 °C; density: 1.26 (4-26 °C) |
Agricultural insecticide and Veterinary pesticide | LD50: 70.2 mg/kg (Oral, Rat); |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



