
Submitted:
28 February 2026
Posted:
04 March 2026
You are already at the latest version
Abstract
Background/Objectives: Kawasaki’s disease (KD) is a leading cause of heart disease in children. The multisystem inflammatory syndrome (MIS) associated with the SARS-CoV-2 virus is similar to KD. The etiologies of KD and MIS are unknown. Both diseases are associated with pathogens and immunizations. Methods: The Vaccine Adverse Event Reporting System (VAERS) was retrospectively examined for etiology insights into both KD and MIS. Results: Immediate-onset KD-associated safety signals were detected for specific vaccines and coadministered combinations of these vaccines for young infants; a subset of these associations have a male sex bias, whereas others appear to be sex neutral. These specific vaccines may contain manufacturing contaminants (e.g., endotoxins) that are activating mast cells via Toll-like receptors. MIS-V cases were enriched in children of all ages. Conclusions: Both KD and MIS appear to involve two activation pathways. The first pathway involves high titers of immune complexes that activate Fc receptors on mast cells, platelets, and other immune cells. Immune complex titers higher than primary immune response levels are predicted to be required to activate low-affinity IgG FcgR2α receptors on immune cells and platelets. IVIG treatment is predicted to directly compete with immune complex binding to FcgR2α receptors. The second pathway appears to directly activate mast cells and other immune cells without involving immune complexes or Fc receptors. Cardiac adverse events, coronary artery aneurysms (CAAs), myocarditis, transient left ventricular dysfunction, and acquired heart disease are predicted to result from pressure induced lesions indirectly caused by cardiac capillary vasoconstrictions. Second, mast cell activation (e.g., endotoxin contaminants) or persistent infections are likely causes of IVIG nonresponders for KD and MIS. MIS appears to be KD associated with the SARS-CoV-2 virus or the COVID-19 spike protein (MIS-V).
Keywords:
Kawasaki disease
; multisystem inflammatory syndrome
; MIS
; vasculitis disease
; mast cells
; histamine
; vaccines
; immunization
; manufacturing contaminant
; endotoxin
1. Introduction
Kawasaki disease (KD) (also known as mucocutaneous lymph node syndrome) is a form of vasculitis in which medium-sized blood vessels become inflamed throughout the body. KD primarily affects children under 5 years of age. Symptoms include fever, rash, conjunctivitis (red eye), oral changes (red, dry, cracked, or fissured lips, “strawberry tongue”, and inflamed oral mucosa), palmar and plantar erythema (redness of hands and feet), cervical adenopathy (enlarged lymph nodes of the neck), coronary artery aneurysms (CAAs) or lesions (CALs) (~25%), and peripheral artery aneurysms [1]. KD fever typically lasts for more than five days and is unresponsive to paracetamol (acetaminophen) or ibuprofen. KD is the leading cause of acquired heart disease (including myocarditis and CAA) in children. The skin from the hands and feet may peel after patient recovery. The etiology of KD is currently unknown. Atypical (or incomplete) KD patients do not fulfill the complete diagnostic criteria for KD but are also at risk for developing coronary artery abnormalities [2]; treatment of KD patients and atypical KD patients with intravenous immunoglobulin (IVIG) and aspirin greatly reduces the incidence of CALs in patients (overview [2]). Notably, aspirin is normally contraindicated for children because of the possible risk of Reye’s syndrome [3]. Up to 20% of IVIG-treated patients develop recurrent or persistent fever (IVIG-resistant) (overview [2]). Kawasaki disease shock/toxic-shock syndrome (KDSS) is an acute phase of KD [4].
Associations between KD and multiple viruses [5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26] and bacterial pathogens [14,27,28,29,30,31,32] have been reported. KD cases can frequently occur several weeks after pathogen outbreaks [22,33,34]. Pyroptosis is a form of inflammatory, programmed, and lytic cell death triggered by infections or other signals were cells rupture and release some proinflammatory molecules. Infection triggered pyroptosis [35] and endothelial cell pyroptosis may play a role in some KD patients [36]. Seasonal exposure patterns are associated with some KD patients aged 3 years or older but not younger [37]. During the COVID-19 pandemic, the incidence of KD cases decreased and remained low during the period of masking and school closures for older children more than for infants [38,39]. KD has also been reported as a rare adverse event associated with individual vaccines and concomitant vaccine combinations [40,41,42,43,44,45,46,47,48,49,50,51,52,53,54]. Patients with KD can also have altered gastrointestinal microbiota [55,56]. KD cases also temporally cluster [57]. Environmental exposures may be triggering some KD cases [58]. Cumulative prenatal and postnatal air pollution exposure to carbon monoxide (CO), nitric oxide (NO), nitric dioxide (NO2), and nitrogen oxide (NOx) but not ozone (O3) exposure has a dose-dependent effect on increasing KD incidence [59]. O3, but not CO, NO2, particulate matter with an aerodynamic diameter < 10 μm (PM10), and SO2 were not found to be associated with each other in a different study [60]. A study of CO, NO2, SO2, O3, PM2.5, and PM10 reported positive associations for only SO2 and PM2.5 for KD [61]. An exposure dosage relationship between PM2.5 and KD has been reported [62]. Additionally, increases in the monthly mean temperature and dry season were associated with increased KD in the Philippines [63]. PM2.5, PM10, SO2 (warm season), and temperature associations have been detected [64]. A meta-analysis revealed both prenatal and postnatal associations between ambient air pollution and KD [65]. KD associations include pathogens, vaccines, air pollution, and increased temperature.
Individuals with COVID-19 can develop multisystem inflammatory syndrome (MIS) in children (MIS-C), adults (MIS-A) [66], and neonates (MIS-N), with significant similarities to KD or KD toxic-shock syndrome [67]. MIS-C has also been named Pediatric Inflammatory Syndrome temporally associated with SARS-CoV-2 infection (PIMS-TS) [68]. MIS is thought to be distinct from KD because of differences in patient age; gastrointestinal and cardiovascular system involvement (including myocarditis, transient left ventricular dysfunction, and depressed cardiac output); and laboratory findings [67,68,69,70,71]. Elevated troponin and elevated B-type natriuretic peptide are key laboratory findings of MIS compared with KD [72]. KD and KDSS are associated with coronary artery pathologic changes and long-term cardiovascular sequelae [73,74,75]. KD and MIS symptoms overlap with those of mast cell activation syndromes (MCAS) [76]. Note that temperature changes and air pollution are known to trigger MCAS.
Hypothesis 1: Multisystem inflammatory syndrome is Kawasaki disease associated with the SARS-CoV-2 pathogen, with differences associated with specific infectious pathogen (e.g., SARS-CoV-2). Similarly, MIS-V is KD-V [77] associated with a COVID-19 (spike protein) vaccine. The differences between KD and MIS are likely due to the SARS-CoV-2 virus associated symptoms (MIS-C, MIS-A, and MIS-N).
Platelet activation plays an important role in KD pathogenesis [78]; monocyte‒platelet aggregates (MPAs) (markers of platelet activation) are significantly elevated in the acute stages of KD [78]. Platelet count and plateletcrit (PCT) were found to be diagnostic markers for KD [79]. Thrombocytopenia has been reported in a KD patient [80]. Thrombocytopenia or thrombocytosis can be associated with KD [81]. In a murine model of KD vasculitis, platelets exacerbated cardiovascular inflammation [82]. A KD etiology model in which activated mast cells and platelets are important KD pathogenic characteristics has been proposed [83]. In KD, platelets and activated monocytes can result in Kawasaki disease complicated with macrophage activation syndrome (KD-MAS) [84].
In this study, the Vaccine Adverse Event Reporting System (VAERS) was retrospectively examined to obtain additional insights into both KD pathogenesis and MIS pathogenesis. Previously proposed KD and MIS etiology models are refined, linking pathogen infections, immunization, and environmental triggers with activated mast cells. Candidate manufacturing contaminants were identified as likely causative agents triggering KD and MIS post immunization.
2. Methods
This is a retrospective analysis of the VAERS database [85] from January 1, 1990, until January 30, 2026. VAERS was used to search for Kawasaki’s disease, Multisystem inflammatory syndrome, Multisystem inflammatory syndrome in children, Multisystem inflammatory syndrome in adults, and Death AEs. The Ruby program vaers_slice5.rb [86] was used for retrospective analysis of the VAERS data files VAERSDATA, VAERSSYMPTOMS, and VAERSVAX for the years 1990--2026 and NonDomestic.
For the vaccine (Vname) and each adverse event (X) in VAERS, normalized AE frequencies per P=100,000 VAERS reports per category of AEs can be calculated with Equation (1).
3. Results
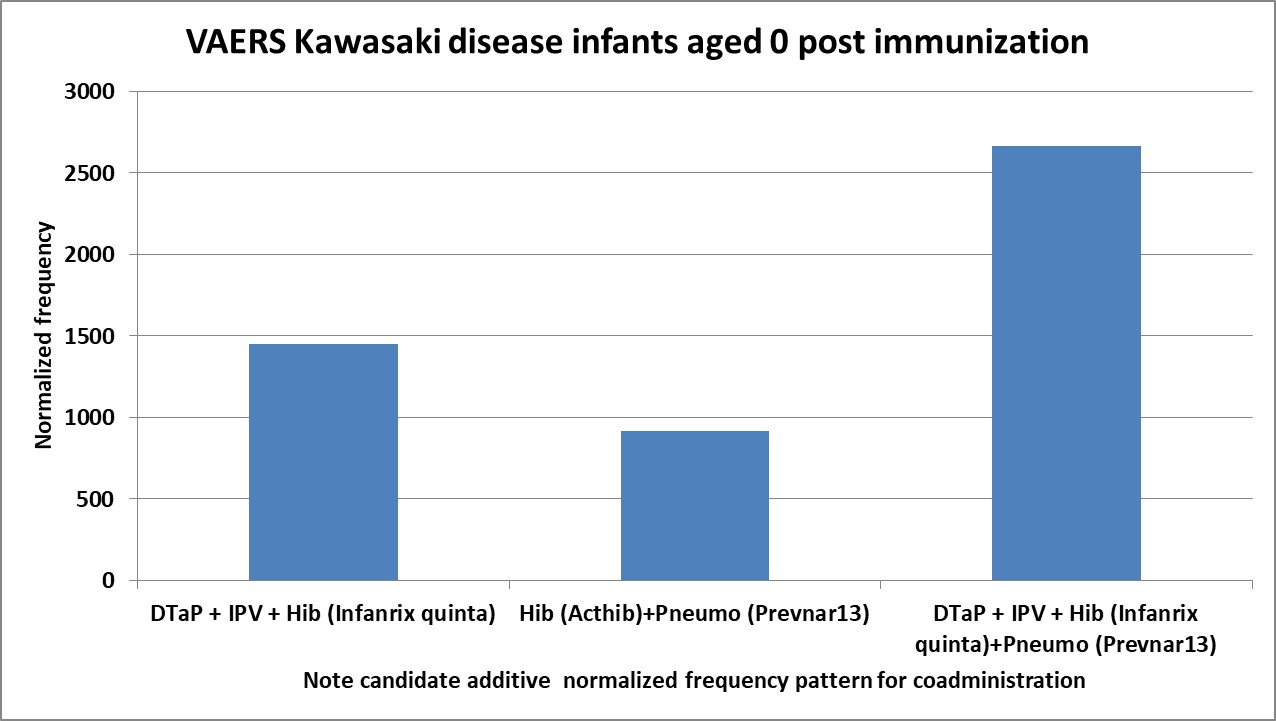
In VAERS, KD is nonrandomly associated with multiple specific vaccines with different normalization frequencies for children aged 0, 1, and 2 years (Figure 1); a subset of these vaccines are associated with elevated normalized frequencies for Infants aged 0 year (Figure 2A). Associations of KD with immunization (Figure 1) may account for the lack of seasonal exposure patterns for some KD patients aged younger than 3 years [37]. The frequency of vaccine names in Figure 2A is plotted in Figure 2B; these same vaccines have similar patterns of normalized frequencies for AE death, Pearson r=0.66 (Figure 2C). The vaccines and concurrent vaccine combinations with higher normalized frequencies (Figure 1 & 2) are predicted to contain unknown manufacturing contaminant(s), most likely endotoxins. Endotoxin exposure activates mast cells [87]. The normalized frequencies for MIS-V associated with the COVID-19 Pfizer-BioNTech vaccine are shown in Figure 3. The four vaccine lots with the most MIS reports include lot FK5127 with a normalized frequency of 696 (24 of 3,448 VAERS reports), FK5618 at 782 (20 of 2,255), FL00007 at 1,173 (21 of 1,790), and FN4072 at 1,775 (6 of 338) (Figure 4). While adverse events reported to VAERS are subject to reporting bias with fewer reports with increased time since immunization, the highest reports for KD and MIS are 1--2 days, with possible small increases associated with antibody immune responses (Figure 5). Note that the day of onset patterns post immunization for KD and MIS correlate with Pearson r=0.90 (Figure 5). An increased male sex bias is known for KD; normalized frequencies for AE KD by sex in VAERS are illustrated in Figure 6. Only four vaccines have imbalances between normalized frequencies for males versus females: DTaP+IPV+Hib (Infanrix quinta), DTaP+IPV+HepB+Hib (Infanrix hexa), Measles+Mumps+Rubella (Priorix), and Meningococcal B (Bexsero) (Figure 6), whereas the other vaccines have roughly equivalent normalized frequencies (Figure 6). Note that Measles+Mumps+Rubella (MMR II) has low normalized frequencies similar to Measles+Mumps+Rubella+Varicella (Proquad) but discordant from Measles+Mumps+Rubella (Priorix) with higher normalized frequencies for females and much higher normalized frequencies for males (Figure 6). Surprisingly, there are specific immunization doses associated with KD reports (Figure 7). For MIS, the majority of the reports are associated with the second COVID-19 shot. For COVID-19 Pfizer-BioNTech, the normalized frequency is higher for the second shot (169 reports) versus the first shot (155 reports). The symptoms reported in the VAERS for KD and MIS patients are summarized in Table S1.
For Kawasaki’s disease, five vaccines have manufacturing lots with three reported Kawasaki’s disease cases: DTaP+HepB+IPV (Pediarix): AC21B248CA, Hib (Acthib): T1E12, Pneumo (Prevnar13): EG8873 and CS7258, Rotavirus (Rotarix): RT014 and RT018, and Rotavirus (RotaTeq): 0324X. For MIS, seven COVID-19 (Pfizer-BioNTech) manufacturing lots have four or more MIS cases: EW0179: 4 reports, FE7051: 4 reports, FG6273: 4 reports, FK5127: 24 reports, FK618: 20 reports, FL0007: 21 reports, and FN4072: 6 reports (Figure 4). The elevated MIS normalized frequencies for manufacturing lots are not associated with high-risk recipient groups (none are known) or background occurrences.
4. Discussion
The initial etiology model for KD and MIS is for high titers of IgG antibodies in immune complexes binding to low-affinity FcγR2α receptors activating mast cells, platelets, and other immune cells [83]. These retrospective VAERS results support expanding this etiology model to also include activation of mast cells from predicted manufacturing contaminant(s) (e.g., endotoxins, etc.) for specific vaccines (Figure 1 and Figure 2, and 4). The limulus amebocyte lysate (LAL)-based assays may miss endotoxins (e.g., low endotoxin recovery (LER)) due to a “masking effect” caused by chelators or detergents commonly used in buffer formulations [88]. Note that the SARS-CoV-2 spike protein bind to bacterial LPS (endotoxin) and boots proinflammatory activity [89,90]. By design, vaccines stimulate innate and humeral immune responses. The Toll-like receptor (TLR4) are activated by LPS (endotoxin) of Gram-negative bacteria [[91]]. TLRs recognize pathogen-associated molecular patterns (PAMPS). KD is associated with pathogen-associated molecular patterns (PAMPS) [92] and microbe-associated molecular patterns (MAMPS) [32]. The immediate onset of KD and MIS associated with immunization (Figure 5) are predicted to result from manufacturing contaminant(s) activating mast cells via TLR4. Increased TLR2 and TLR4 expression in peripheral neutrophils has been detected in some KD patients [93]. Some KD cases may be associated with endotoxins and elevated soluble CD14 (sCD14) [94,95]. Polyclonal expansion of TCRBV2- and TCRBV6-bearing T cells occurs in KD patients (likely associated with endotoxin exposure) [31]. Low-level endotoxin induces potent inflammatory activation of human blood vessels [96].
Multiple patients with KD or MIS also have associated gastrointestinal (GI) symptoms/intestinal involvement [97,98]; this may include intestinal dysbiosis and sometimes disruption of gut barrier [99]. Disruption of the gut barrier is likely associated with the presence of a superantigen (perhaps endotoxin) [100]. Elevated Vβ2 T cells expansion in some KD patients is consistent with the superantigen model [101]. KD patients with abdominal manifestations (symptoms) are more likely to be IVIG-resistant (p<0.005) and have CAA (p=0.007) [102].
4.1. KD and MIS Etiology Model
Etiology Model: KD and MIS are associated with activated mast cells and platelets.
Pathogen-associated reports are predicted to be associated with elevated immune complexes IgG antibody levels above primary immune response levels, activating low-affinity IgG FcγR2α receptors on platelets, mast cells, and additional immune cells (note the risk of persistent infections) (Table 1) [103,104]. Elevated histamine and likely serotonin levels are likely associated with most of the KD and MIS symptoms [83]. For KD and MIS patients with high IgG antibody titers, IVIG treatment is predicted to directly compete with immune complex binding to FcγR2α receptors, resulting in reduced activation of mast cells and platelets resulting in relief of associated symptoms. Immunization and environmental exposures can activate mast cells, immune cells, and likely platelets without (likely IVIG resistant) or sometimes with FcγR2α receptor binding (e.g., humeral responses post immunization) (Table 1). Gastrointestinal symptoms are reported in the majority of MIS-C patients [105,106,107,108,109]. A MIS-A patient with profound gastrointestinal symptoms have been reported [110]. SARS-CoV-2 virus or spike protein (COVID-19 vaccines) can induce additional gastrointestinal and cardiac symptoms in MIS and MIS-V patients, respectively. The spike protein also activates mast cells via TLR4 and angiotensin-converting enzyme 2 (ACE2) receptors [111]. For KD and MIS associated with onset within a few days of immunization, endotoxin manufacturing contaminant(s) (or Spike protein binding) is predicted to activate mast cells via TLR4; this activation pathway does not involve FcγR2α receptors and these patients are anticipated to be resistant to IVIG treatment (Table 1).
Notably, overall immune activation is increased in KD [112]. It is unknown whether histamine intolerance (HIT) plays a role in KD or MIS. Multiple factors can influence an individual’s tolerance threshold for histamine; including drugs [113]; foods (cocoa, spinach, tomatoes, wine, beer, cheeses, yogurt, meat, soy, fermented foods, etc.) [113,114]; the gastrointestinal microbiome [113]; and the stage of the menstrual cycle [114].
4.2. Age-Related Risk Patterns
The proposed KD and MIS etiology model proposes the activation of mast cells, platelets, and immune cells by Fc receptor binding to immune complexes or via direct activation of immune cells. Maternally transferred antibodies (matAbs) may play a role in KD-N and MIS-N in neonates with neonate antibody responses combined with matAbs to reach the higher levels of IgG antibodies in immune complexes needed to trigger disease [115]. For the 0–5 year age group, it appears that specific vaccines predicted to be contain the endotoxin manufacturing contaminant(s) are associated with KD (KD-V) (Figure 1) and also MIS-V (Figure 3). The normalized frequencies observed for COVID-19 (Pfizer-BioNTech) may approximate MIS (both MIS-C and MIS-V) risk levels in children (Figure 3).
4.3. Cardiac Adverse Events and Acquired Heart Disease
This model also proposes that aneurysms are pressure induced by contracted cardiac capillary pericyte vasoconstrictions [104]; notably, serotonin released from activated platelets is also associated with vasoconstrictions [116,117]. Induced cardiac capillary pericyte contractions are predicted to be associated with anoxia and possibly pressure-induced CAA and peripheral artery aneurysms [104,118]. Untreated patients with ongoing ischemia are predicted to experience cardiac myocyte anoxia, which may account for KD-associated acquired heart disease; this also explains the vascular dysfunction in patients who do not have echocardiographic evidence of coronary artery abnormalities in the acute phase of KD. An increased proportion of KD patients with CAA also have the plasma fibrinogen (FG) alpha genotype Thr312Ala [119]. Sex differences in cardiac mast cells activation have also been observed [120]; this may be associated with the KD male sex bias for specific vaccines (Figure 6).
The differences between MIS-related cardiac symptoms and KD-related symptoms (myocarditis, transient left ventricular dysfunction, and depressed cardiac output) may be directly due to the SARS-CoV-2 virus or the SARS-CoV-2 vaccine spike protein. For COVID-19 mRNA vaccines, circulating spike proteins are observed in vaccinees with myocarditis [121], along with elevated cardiac troponin levels [122]. For COVID-19 vaccines, the Spike protein disrupts cardiac pericytes through cluster of differentiation 147 (CD147) receptor-mediated signaling and another unknown mechanism [123]. The spike protein also activates mast cells via TLR4 and angiotensin-converting enzyme 2 (ACE2) receptors [111]. These spike protein interactions may account for the increased risk for myocarditis and transient left ventricular dysfunction observed in MIS compared with KD [124]. Note that the spike protein interactions cannot account for MIS normalized frequency disparities for COVID-19 Pfizer-BioNTech manufacturing lots (Figure 4).
4.4. KD and MIS Delayed Onset
Clusters of KD and MIS (Figure S1) reports are frequently observed with delayed disease onset (approximately 1 month or more) following various pathogen [22,33,34] and SARS-CoV-2 outbreaks [125], respectively. For this delayed disease onset pattern, the proposed etiology model requires IgG antibody levels to be higher than primary immune response levels to trigger disease [115]. One scenario includes persistent infections (e.g., gastrointestinal infections), which may occur in some KD and MIS patients [115]. Elevated SARS-CoV-2 antibody titers [105,126,127,128], current SARS-CoV-2 infections, or prior SARS-CoV-2 infections or exposures [129,130,131,132,133,134] are observed in MIS patients. For MIS-C, sustained levels of inflammatory macrophage-activating, Fc receptor-binding antibodies are selectively maintained in severe disease [135]. MIS-C develops in some children with COVID-19 and persistent SARS-CoV-2 infections [136].
4.5. KD Genetics
Genetic variants are predicted to increase or decrease associated KD and MIS risks. Confirmed KD genetic variants include inositol 1,4,5-trisphosphate 3-kinase C (ITPKC) (calcineurin, a nuclear factor of the activated T-cell pathway—calcium signaling pathway) [137,138], caspase-3 (CASP3) [138,139], toll like receptor 6 (TLR6) [140], and the low-affinity IgG receptor gene FcγR2α (encoding FcγRIIa) [141,142,143,144]. The FcγR2α rs1801274 C allele encodes arginine (R) (low binding to IgG2 and IgG3), and the T allele encodes histidine (H) (high binding to IgG2 and IgG3) [145]. The FcγR2α pHis167Arg is associated with KD risk in males [146]. Candidate KD associated genes are associated with immune system, calcium signaling, KD susceptibility, IVIG resistance, and aneurysm formation (reviewed [147,148]); identified gene variants include B-cell lymphoid tyrosine kinase (BLK) [143,149,150,151], CD40 (tumor necrosis factor receptor superfamily member 5) [149,150], FcγRIIA-131H variant [152], FcγR2B [153], FcγR2C gene copy number [154], FcγR2C-ORF [155], FcγR3B gene copy number [154], FcγRIIIB neutrophil antigen 1 (NA1) variant overexpression in IVIG nonresponders [152], immunoglobulin heavy variable gene (IGHV) [156], human leukocyte antigen (HLA) [150], calcium release-activated calcium modulator 1 (ORAI1) (involved in calcium influx in immune cells) [157], potassium calcium-activated channel subfamily N member 2 (KCNN2) (associated with CAA) [158], TGF-beta receptor II (TGFBR2) [159,160], Mothers against decapentaplegic homolog 3 (SMAD3) (TGF-beta signaling pathway) [159], Mothers against decapentaplegic homolog 5 (SMAD5) (TGF-beta signaling pathway) [161]. Note that CASP3 is released by activated mast cells [162]. Mast cells express CD40 ligand (CD40L) that interacts with CD40 on B cells [163]. Differentially expressed candidate KD genes have also been characterized via transcriptome analysis [164,165]. Whole-exome sequencing revealed that KD candidates include myosin heavy chain 14 (MYH14) and retinol-binding protein 3 (RBP3) [166]. While no association with the FcγR2α rs1801274 polymorphism was found, MIS-C patients with the homozygous FcγR2α rs1801274 gene polymorphism developed severe cardiac dysfunction [167]. Individual genetics alter KD and MIS risks. Genetic variants in T helper cell pathways may contribute to immune dysregulation in KD [168]. Identified genetic variants associated with KD play roles in immune cells, including mast cells, activation or signaling.
4.6. IVIG Treatment and IVIG Resistance
The model of high IgG antibody activation of FcγR2α receptor binding represents a novel form of antibody-dependent enhancement (ADE) for both KD and MIS [103]. IVIG treatment is predicted to compete with pathogen IgG antibodies for FcγR2α receptor binding with a possible increased risk for IVIG resistance; note that TLR4 (non-FcγR2α receptor) activated mast cells are likely IVIG resistant (due to different activation pathway). This model explains the unpredictable ineffectiveness of current therapy and the observed IVIG resistance in both KD and MIS patients.
4.7. MIS Differences from KD
KD and MIS reports not associated with recent vaccinations may be associated with persistent (perhaps gastrointestinal in some reports) infections. The greater number of KDs at ages 0--5 may be associated with immunizations. Resilience against the development of pressure-induced CAA may reduce incidence rates as the age of patients increases. SARS-CoV-2 infection or spike protein interactions may account for cardiac differences between MIS and KD. Otherwise, MIS appears to be KD associated with SARS-CoV-2 virus or COVID-19 immunization (MIS-V).
4.8. Candidate Adjunctive Treatments
If the proposed KD and MIS etiology model is correct, then additional adjunctive treatments, including mast cell stabilizers, antihistamines, and possibly serotonin antagonists, may reduce symptoms; institutional review board (IRB)-approved targeted clinical studies (e.g., report series) are suggested (perhaps targeting IVIG nonresponders).
4.9. Study Limitations
The VAERS database includes only a small subset of adverse events experienced by vaccinees. Any reporting biases or exclusion of adverse events would perturb the accuracy of VAERS, which represents the population.
4.10. Study Recommendations
This study proposes that mast cell and platelet activation drive the etiology of both KD and MIS. Many of the disease symptoms are consistent with predicted elevated levels of histamine and/or serotonin. Evaluations of adjunctive treatments targeting elevated histamine or serotonin levels are candidates for evaluation in approved clinical studies. Early treatments may reduce the risk of CALs and acquired heart disease in KD patients and ventricular dysfunction and cardiac adverse events in MIS patients. KD and MIS reports not associated with immunizations may have undiagnosed persistent infections for which appropriate treatments should be considered. Elimination or reduction of predicted endotoxin manufacturing contamination level in identified vaccines with higher KD and MIS normalized frequencies is predicted to reduce AEs KD, MIS, and death in children.
5. Conclusions
The etiology of both KD and MIS are both likely novel MCAS. An etiology model is proposed that can account for the etiology of both KD and MIS. For pathogen-associated infections, high-titer immune complexes are required to activate low IgG -affinity FcγR2α receptors; this results in delayed disease onset clusters following pathogen outbreaks. Air pollution and increased temperature can also activate mast cells, triggering KD. Immediate onset KD and MIS post immunization is likely caused by predicted endotoxin manufacturing contaminants in specific vaccines activating mast cells directly via TLR4; these patients are predicted to be IVIG nonresponders. KD-related male sex bias may be partially due to specific vaccine associations and sex differences between cardiac mast cells. CAA, myocarditis, myocarditis, transient left ventricular dysfunction, and acquired heart disease are predicted to result from cardiac capillary vasoconstrictions, and aneurysms are predicted to be pressure induced. While appearing clinically distinct, MIS appears to be KD associated with the SARS-CoV-2 virus or COVID-19 spike protein (MIS-V).
Author Contributions
Conceptualization, D.R.; Methodology, D.R.; Software, D.R.; Validation, D.R.; Formal Analysis, D.R..; Investigation, D.R..; Data Curation, D.R.; Writing—Original Draft Preparation, D.R.; Writing—Review & Editing, D.R.
Funding
This research received no external funding.
Data Availability Statement
The datasets generated for this study from VAERS are available as open data at https://doi.org/10.7910/DVN/QRBEQT, Harvard Dataverse, V2.
Acknowledgments
None.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ACE2 | Angiotensin-converting enzyme 2 |
| ADE | Antibody-dependent enhancement |
| AE | Adverse event |
| B-cell | Immune B lymphocyte |
| BLK | B-cell lymphoid tyrosine kinase |
| CAA | Coronary artery aneurysm |
| CALs | Coronary artery lesion |
| CASP3 | Caspase 3 |
| CD14 | Cluster of differentiation 14 |
| CD147 | Cluster of differentiation 147, also known as EMMPRIN (Extracellular Matrix Metalloproteinase Inducer), or Basigin |
| CD40 | Cluster of differentiation 40 |
| CD40L | Cluster of differentiation 40 ligand |
| CO | Carbon monoxide |
| COVID-19 | Coronavirus disease 2019 |
| DTaP | Diphtheria, tetanus, and pertussis (whooping cough) vaccine |
| Fc | fragment crystallizable region of antibody |
| FcγR | Fc gamma receptor |
| FG | plasma fibrinogen |
| GI | gastrointestinal |
| Hep B | hepatitis B |
| Hib | Haemophilus influenzae type b vaccine |
| HIT | histamine intolerance |
| HLA | human leukocyte antigen |
| IgG | immunoglobulin G |
| IGHV | immunoglobulin heavy variable gene |
| IPV | inactivated poliovirus vaccine |
| ITPKC | inositol 1,4,5-trisphosphate 3-kinase C |
| IVIG | intravenous immunoglobulin |
| KCNN2 | Potassium Calcium-Activated Channel Subfamily N Member 2 |
| KD | Kawasaki’s disease |
| KD-MAS | Kawasaki disease complicated with macrophage activation syndrome |
| KD-N | Kawasaki’s disease in neonates |
| KDSS | Kawasaki disease shock/toxic-shock syndrome |
| KD-V | Kawasaki’s disease associated with vaccination |
| LAL | Limulus Amebocyte Lysate |
| LER | Low endotoxin recovery |
| LPS | Lipopolysaccharide |
| MAMPS | Microbe-associated molecular patterns |
| matAbs | Maternally transferred antibodies |
| MCAS | Mast cell activation syndromes |
| MIS | Multisystem inflammatory syndrome |
| MIS-A | Multisystem inflammatory syndrome in adults |
| MIS-C | Multisystem inflammatory syndrome in children |
| MIS-N | Multisystem inflammatory syndrome in neonates |
| MIS-V | Multisystem inflammatory syndrome after COVID-19 vaccination |
| MMR | Measles, mumps, and rubella vaccine |
| MPAs | Monocyte‒platelet aggregates |
| mRNA | Messenger ribonucleic acid |
| MYH14 | Myosin heavy chain 14 |
| NA1 | Neutrophil antigen 1 |
| NO | Nitric oxide |
| NO2 | Nitric dioxide |
| NOx | Nitrogen oxide |
| O3 | Ozone |
| ORAI1 | calcium release-activated calcium modulator 1 |
| ORF | Open reading frame |
| PAMPS | pathogen-associated molecular patterns |
| PCT | Plateletcrit |
| PIMS-TS | Pediatric Inflammatory Syndrome temporally associated with SARS-CoV-2 infection |
| PM10 | Inhalable particulate matter 10 micrometers or smaller |
| PM25 | fine inhalable particles less than or equal to 2.5 micrometers in diameter |
| RBP3 | retinol-binding protein 3 |
| SAE | Serious adverse event |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| sCD14 | Soluble CD14 protein |
| SMAD3 | Mothers against decapentaplegic homolog 3 |
| SMAD5 | Mothers against decapentaplegic homolog 5 |
| SO2 | Sulfur dioxide |
| T-cell | T lymphocyte |
| TCRBV2 | T-cell receptor Beta-chain V2 |
| TCRBV6 | T-cell receptor Beta-chain V6 |
| TGF-beta | Transforming growth factor-beta |
| TGFBR2 | Transforming growth factor-beta receptor type 2 |
| TLR | Toll-like receptor |
| VAERS | Vaccine Adverse Event Reporting System |
References
- McCrindle, B.W.; Rowley, A.H.; Newburger, J.W.; Burns, J.C.; Bolger, A.F.; Gewitz, M.; Baker, A.L.; Jackson, M.A.; Takahashi, M.; Shah, P.B.; et al. Diagnosis, Treatment, and Long-Term Management of Kawasaki Disease: A Scientific Statement for Health Professionals from the American Heart Association. Circulation 2017, 135, e927–e999. [Google Scholar] [CrossRef]
- Rife, E.; Gedalia, A. Kawasaki Disease: an Update. Curr. Rheumatol. Rep. 2020, 22, 1–10. [Google Scholar] [CrossRef]
- Bayer. 2025. [PubMed]
- Lamrani, L.; Manlhiot, C.; Elias, M.D.; Choueiter, N.F.; Dionne, A.; Harahsheh, A.S.; Portman, M.A.; McCrindle, B.W.; Dahdah, N. Kawasaki Disease Shock Syndrome vs Classical Kawasaki Disease: A Meta-analysis and Comparison With SARS-CoV-2 Multisystem Inflammatory Syndrome. Can. J. Cardiol. 2021, 37, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Embil, J.A.; McFarlane, E.S.; Murphy, D.M.; Krause, V.W.; Stwart, H.B. Adenovirus type 2 isolated from a patient with fatal Kawasaki disease. Can. Med. Assoc. J. 1985, 132, 1400. [Google Scholar]
- Chang, L.-Y.; Lu, C.-Y.; Shao, P.-L.; Lee, P.-I.; Lin, M.-T.; Fan, T.-Y.; Cheng, A.-L.; Lee, W.-L.; Hu, J.-J.; Yeh, S.-J.; et al. Viral infections associated with Kawasaki disease. J. Formos. Med. Assoc. 2014, 113, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Catalano-Pons, C.; Giraud, C.; Rozenberg, F.; Meritet, J.-F.; Lebon, P.; Gendrel, D. Detection of human bocavirus in children with Kawasaki disease. Clin. Microbiol. Infect. 2007, 13, 1220–1222. [Google Scholar] [CrossRef]
- Shirato, K.; Imada, Y.; Kawase, M.; Nakagaki, K.; Matsuyama, S.; Taguchi, F. Possible involvement of infection with human coronavirus 229E, but not NL63, in Kawasaki disease. J. Med Virol. 2014, 86, 2146–2153. [Google Scholar] [CrossRef]
- Esper, F.; Weibel, C.; Ferguson, D.; Landry, M.L.; Kahn, J.S. Evidence of a Novel Human Coronavirus That Is Associated with Respiratory Tract Disease in Infants and Young Children. J. Infect. Dis. 2005, 191, 492–498. [Google Scholar] [CrossRef]
- Catalano-Pons, C.; Quartier, P.; Leruez-Ville, M.; Kaguelidou, F.; Gendrel, D.; Lenoir, G.; Casanova, J.-L.; Bonnet, D. Primary Cytomegalovirus Infection, Atypical Kawasaki Disease, and Coronary Aneurysms in 2 Infants. Clin. Infect. Dis. 2005, 41, e53–e56. [Google Scholar] [CrossRef]
- Jagadeesh, A.; Krishnamurthy, S.; Mahadevan, S. Kawasaki Disease in a 2-year-old Child with Dengue Fever. Indian J. Pediatr. 2015, 83, 602–603. [Google Scholar] [CrossRef]
- Sopontammarak, S.; Promphan, W.; Roymanee, S.; Phetpisan, S. Positive Serology for Dengue Viral Infection in Pediatric Patients With Kawasaki Disease in Southern Thailand. Circ. J. 2008, 72, 1492–1494. [Google Scholar] [CrossRef]
- Weng, K.-P.; Wei, J.C.-C.; Hung, Y.-M.; Huang, S.-H.; Chien, K.-J.; Lin, C.-C.; Huang, S.-M.; Lin, C.-L.; Cheng, M.-F. Enterovirus Infection and Subsequent Risk of Kawasaki Disease: A Population-based Cohort Study. Pediatr. Infect. Dis. J. 2018, 37, 310–315. [Google Scholar] [CrossRef]
- Kikuta, H.; Nakanishi, M.; Ishikawa, N.; Konno, M.; Matsumoto, S. Detection of Epstein-Barr virus sequences in patients with Kawasaki disease by means of the polymerase chain reaction. Intervirology 1992, 33, 1–5. [Google Scholar] [CrossRef]
- Okano, M.; Luka, J.; Thiele, G.M.; Sakiyama, Y.; Matsumoto, S.; Purtilo, D.T. Human herpesvirus 6 infection and Kawasaki disease. J. Clin. Microbiol. 1989, 27, 2379–2380. [Google Scholar] [CrossRef]
- Okano, M. Kawasaki Disease and Human Lymphotropic Virus Infection. Curr. Med Res. Opin. 1999, 15, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.V.; Jones, K.D.; Buckley, A.; E Coren, M.; Kampmann, B. Kawasaki disease coincident with influenza A H1N1/09 infection. Pediatr. Int. 2011, 53, e1–e2. [Google Scholar] [CrossRef] [PubMed]
- Whitby, D.; Hoad, J.G.; Tizard, E.J.; Dillon, M.J.; Weber, J.N.; A Weiss, R.; Schulz, T.F. Isolation of measles virus from child with Kawasaki disease. Lancet 1991, 338, 1215–1215. [Google Scholar] [CrossRef]
- Holm, J.M.; Hansen, L.K.; Oxhøj, H. Kawasaki disease associated with parvovirus B19 infection. Eur. J. Pediatr. 1995, 154, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Nigro, G.; Krzysztofiak, A.; Porcaro, M.; Mango, T.; Zerbini, M.; Gentilomi, G.; Musiani, M. Active or recent parvovirus B19 infection in children with Kawasaki disease. Lancet 1994, 343, 1260–1261. [Google Scholar] [CrossRef]
- Keim, D.; Keller, E.; Hirsch, M. MUCOCUTANEOUS LYMPH-NODE SYNDROME AND PARAINFLUENZA 2 VIRUS INFECTION. Lancet 1977, 310, 303. [Google Scholar] [CrossRef]
- Kim, G.B.; Park, S.; Kwon, B.S.; Han, J.W.; Park, Y.W.; Hong, Y.M. Evaluation of the Temporal Association between Kawasaki Disease and Viral Infections in South Korea. Korean Circ. J. 2014, 44, 250–254. [Google Scholar] [CrossRef]
- Matsuno, S.; Utagawa, E.; Sugiura, A. Association of Rotavirus Infection with Kawasaki Syndrome. J. Infect. Dis. 1983, 148, 177–177. [Google Scholar] [CrossRef] [PubMed]
- Ogboli, M.; Parslew, R.; Verbov, J.; Smyth, R. Kawasaki disease associated with varicella: a rare association. Br. J. Dermatol. 1999, 141, 1136–1152. [Google Scholar] [CrossRef] [PubMed]
- Kossiva, L.; Papadopoulos, M.; Lagona, E.; Papadopoulos, G.; Athanassaki, C. Myocardial infarction in a 35-day-old infant with incomplete Kawasaki disease and chicken pox. Cardiol. Young- 2010, 20, 567–570. [Google Scholar] [CrossRef]
- Thissen, J.B.; Isshiki, M.; Jaing, C.; Nagao, Y.; Aldea, D.L.; E Allen, J.; Izui, M.; Slezak, T.R.; Ishida, T.; Sano, T. A novel variant of torque teno virus 7 identified in patients with Kawasaki disease. PLOS ONE 2018, 13, e0209683. [Google Scholar] [CrossRef]
- Hall, M.; Schlievert, P.M.; Hoyt, L.; Ferrieri, P.; Jenson, H.B. Kawasaki Syndrome-Like Illness Associated with Infection Caused by Enterotoxin B-Secreting Staphylococcus aureus. Clin. Infect. Dis. 1999, 29, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Shinomiya, N.; Takeda, T.; Kuratsuji, T.; Takagi, K.; Kosaka, T.; Tatsuzawa, O.; Tsurumizu, T.; Hashimoto, T.; Kobayashi, N. Variant Streptococcus sanguis as an etiological agent of Kawasaki disease. Prog. Clin. Biol. Res. 1987, 250, 571–2. [Google Scholar]
- Leung, D.; Kotzin, B.; Meissner, H.; Fulton, R.; Murray, D.; Schlievert, P. Toxic shock syndrome toxin-secreting Staphylococcus aureus in Kawasaki syndrome. Lancet 1993, 342, 1385–1388. [Google Scholar] [CrossRef]
- Matsubara, K.; Fukaya, T.; Miwa, K.; Shibayama, N.; Nigami, H.; Harigaya, H.; Nozaki, H.; Hirata, T.; Baba, K.; Suzuki, T.; et al. Development of serum IgM antibodies against superantigens ofStaphylococcus aureusandStreptococcus pyogenesin Kawasaki disease. Clin. Exp. Immunol. 2006, 143, 427–434. [Google Scholar] [CrossRef]
- Yoshioka, T.; Matsutani, T.; Iwagami, S.; Maeda, T.; Yutsudo, T.; Tsuruta, Y.; Suzuki, H.; Uemura, S.; Takeuchi, T.; Koike, M. Polyclonal expansion of TCRBV2- and TCRBV6-bearing T cells in patients with Kawasaki disease. Immunology 1999, 96, 465–472. [Google Scholar] [CrossRef]
- Kusuda, T.; Nakashima, Y.; Murata, K.; Kanno, S.; Nishio, H.; Saito, M.; Tanaka, T.; Yamamura, K.; Sakai, Y.; Takada, H.; et al. Kawasaki Disease-Specific Molecules in the Sera Are Linked to Microbe-Associated Molecular Patterns in the Biofilms. PLOS ONE 2014, 9, e113054–e113054. [Google Scholar] [CrossRef]
- Kang, J.-M.; Jung, J.; Kim, Y.-E.; Huh, K.; Hong, J.; Kim, D.W.; Kim, M.Y.; Jung, S.Y.; Kim, J.-H.; Ahn, J.G. Temporal Correlation Between Kawasaki Disease and Infectious Diseases in South Korea. JAMA Netw. Open 2022, 5, e2147363–e2147363. [Google Scholar] [CrossRef]
- Dean, A.G.; Melish, M.E.; Hicks, R.; Palumbo, N.E. An epidemic of Kawasaki syndrome in Hawaii. J. Pediatr. 1982, 100, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Han, X.-Y.; Qi, H.-R. Pyroptosis in Kawasaki disease: from mechanisms to targeted interventions. Front. Immunol. 2025, 16, 1566985. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhang, J.; Chen, H.; Zhuge, Y.; Chen, H.; Qian, F.; Zhou, K.; Niu, C.; Wang, F.; Qiu, H.; et al. Endothelial cell pyroptosis plays an important role in Kawasaki disease via HMGB1/RAGE/cathespin B signaling pathway and NLRP3 inflammasome activation. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- DeHaan, L.L.; Copeland, C.D.; Burney, J.A.; Nakamura, Y.; Yashiro, M.; Shimizu, C.; Miyata, K.; Burns, J.C.; Cayan, D.R. Age-Dependent Variations in Kawasaki Disease Incidence in Japan. JAMA Netw. Open 2024, 7, e2355001–e2355001. [Google Scholar] [CrossRef]
- Ae, R.; Shibata, Y.; Kosami, K.; Nakamura, Y.; Hamada, H. Kawasaki Disease and Pediatric Infectious Diseases During the Coronavirus Disease 2019 Pandemic. J. Pediatr. 2021, 239, 50–58.e2. [Google Scholar] [CrossRef]
- Burney, J.A.; Roberts, S.C.; DeHaan, L.L.; Shimizu, C.; Bainto, E.V.; Newburger, J.W.; Dominguez, S.; Jone, P.-N.; Jaggi, P.; Szmuszkovicz, J.R.; et al. Epidemiological and Clinical Features of Kawasaki Disease During the COVID-19 Pandemic in the United States. JAMA Netw. Open 2022, 5, e2217436–e2217436. [Google Scholar] [CrossRef]
- Schmöeller, D.; Keiserman, M.W.; Staub, H.L.M.; Velho, F.P.; Grohe, M.d.F. Yellow Fever Vaccination and Kawasaki Disease. Pediatr. Infect. Dis. J. 2009, 28, 1037–1038. [Google Scholar] [CrossRef]
- Ece, I; Akbayram, S; Demiroren, K; et al. Is Kawasaki Disease a Side Effect of Vaccination as Well? J Vaccines Vaccin n.d., 5, 234. [Google Scholar]
- Banday, A.Z.; Patra, P.K.; Jindal, A.K. Kawasaki disease – when Bacillus Calmette–Guérin (BCG) lymphadenitis blooms again and the vaccination site peels off! Int. J. Dermatol. 2020, 60, E233–E234. [Google Scholar] [CrossRef]
- Alsager, K.; Vadlamudi, N.K.; Jadavji, T.; Bettinger, J.A.; Constantinescu, C.; Vaudry, W.; Tan, B.; Sauvé, L.; Sadarangani, M.; Halperin, S.A.; et al. Kawasaki disease following immunization reported to the Canadian Immunization Monitoring Program ACTive (IMPACT) from 2013 to 2018. Hum. Vaccines Immunother. 2022, 18, 2088215. [Google Scholar] [CrossRef]
- Hall, G.C.; Tulloh, R.M.; E Tulloh, L. The incidence of Kawasaki disease after vaccination within the UK pre-school National Immunisation Programme: an observational THIN database study. Pharmacoepidemiol. Drug Saf. 2016, 25, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Islam, S. Kawasaki disease and vasculitis associated with immunization. Pediatr. Int. 2018, 60, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Miron, D.; Fink, D.; Hashkes, P.J. Kawasaki disease in an infant following immunisation with hepatitis B vaccine. Clin. Rheumatol. 2003, 22, 461–463. [Google Scholar] [CrossRef]
- Jeong, S.W.; Kim, D.H.; Han, M.Y.; Cha, S.; Yoon, K.L. An infant presenting with Kawasaki disease following immunization for influenza: A case report. Biomed. Rep. 2018, 8, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Kraszewska-Głomba, B.; Kuchar, E.; Szenborn, L. Three episodes of Kawasaki disease including one after the Pneumo 23 vaccine in a child with a family history of Kawasaki disease. J. Formos. Med Assoc. 2016, 115, 885–886. [Google Scholar] [CrossRef]
- Shimada, S.; Watanabe, T.; Sato, S. A Patient with Kawasaki Disease Following Influenza Vaccinations. Pediatr. Infect. Dis. J. 2015, 34, 913. [Google Scholar] [CrossRef]
- Yin, S.; Liubao, P.; Chongqing, T.; Xiaomin, W. The first case of Kawasaki disease in a 20-month old baby following immunization with rotavirus vaccine and hepatitis A vaccine in China: A case report. Hum. Vaccines Immunother. 2015, 11, 2740–2743. [Google Scholar] [CrossRef]
- Matsubara, D.; Minami, T.; Seki, M.; Tamura, D.; Yamagata, T. Occurrence of Kawasaki disease after simultaneous immunization. Pediatr. Int. 2019, 61, 1171–1173. [Google Scholar] [CrossRef]
- Huang, W.-T.; Juan, Y.-C.; Liu, C.-H.; Yang, Y.-Y.; Chan, K.A. Intussusception and Kawasaki disease after rotavirus vaccination in Taiwanese infants. Vaccine 2020, 38, 6299–6303. [Google Scholar] [CrossRef]
- Showers, C.R.; Maurer, J.M.; Khakshour, D.; Shukla, M. Case of adult-onset Kawasaki disease and multisystem inflammatory syndrome following SARS-CoV-2 vaccination. BMJ Case Rep. 2022, 15, e249094. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Amaro, A.L.; Tejada-Ruiz, M.I.; Rivera-Alvarado, K.L.; Cobos-Quevedo, O.d.J.; Romero-Hernández, P.; Macías-Arroyo, W.; Avendaño-Ponce, A.; Hurtado-Díaz, J.; Vera-Lastra, O.; Lucas-Hernández, A. Atypical Kawasaki Disease after COVID-19 Vaccination: A New Form of Adverse Event Following Immunization. Vaccines 2022, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Kinumaki, A.; Sekizuka, T.; Hamada, H.; Kato, K.; Yamashita, A.; Kuroda, M. Characterization of the gut microbiota of Kawasaki disease patients by metagenomic analysis. Front. Microbiol. 2015, 6, 824. [Google Scholar] [CrossRef]
- Esposito, S.; Polinori, I.; Rigante, D. The Gut Microbiota-Host Partnership as a Potential Driver of Kawasaki Syndrome. Front. Pediatr. 2019, 7, 124. [Google Scholar] [CrossRef]
- Burney, J.A.; DeHaan, L.L.; Shimizu, C.; Bainto, E.V.; Newburger, J.W.; DeBiasi, R.L.; Dominguez, S.R.; Portman, M.A.; Melish, M.; Bratincsak, A.; et al. Temporal clustering of Kawasaki disease cases around the world. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Rodó, X.; Curcoll, R.; Robinson, M.; Ballester, J.; Burns, J.C.; Cayan, D.R.; Lipkin, W.I.; Williams, B.L.; Couto-Rodriguez, M.; Nakamura, Y.; et al. Tropospheric winds from northeastern China carry the etiologic agent of Kawasaki disease from its source to Japan. Proc. Natl. Acad. Sci. USA 2014, 111, 7952–7957. [Google Scholar] [CrossRef]
- Kuo, N.-C.; Lin, C.-H.; Lin, M.-C. Prenatal and early life exposure to air pollution and the incidence of Kawasaki disease. Sci. Rep. 2022, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.-R.; Chen, W.-T.; Lin, Y.-T.; Hwang, B.-F. Ambient Air Pollutant Exposures and Hospitalization for Kawasaki Disease in Taiwan: A Case-Crossover Study (2000–2010). Environ. Health Perspect. 2017, 125, 670–676. [Google Scholar] [CrossRef]
- Kwon, D.; Choe, Y.J.; Kim, S.; Chun, B.C.; Choe, S. Ambient Air Pollution and Kawasaki Disease in Korean Children: A Study of the National Health Insurance Claim Data. J. Am. Hear. Assoc. 2022, 11, e024092. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, K.; Shinjo, D.; Takahashi, N.; Fushimi, K. Spatiotemporal analysis of the association between Kawasaki disease incidence and PM2.5exposure: a nationwide database study in Japan. BMJ Paediatr. Open 2024, 8, e002887. [Google Scholar] [CrossRef]
- Seposo, A.K.C.; Madaniyazi, L.; Yoshida, L.M.; Toizumi, M. Incidence and Seasonality of Kawasaki Disease in Children in the Philippines, and its Association with Ambient Air Temperature. Isee Conf. Abstr. 2023. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Zheng, F.; Guo, Y.; Wang, X.; Meng, H.; Min, L.; Hu, X. Research on the Influence of Air Pollutants and Meteorological Factors on Kawasaki Disease. Aerosol Air Qual. Res. 2025, 25. [Google Scholar] [CrossRef]
- Yang, P.; Zhang, J.; Zhang, K.; Zhang, D.; Liu, Y.; Wu, J.; Wei, Y.; Feng, S.; Yi, Q. Prenatal and Postnatal Ambient Air Pollution and Kawasaki Disease. JACC: Adv. 2025, 4, 101651. [Google Scholar] [CrossRef] [PubMed]
- Elouardi, Y.; Rebahi, H.; Zarrouki, Y.; Ziadi, A.; Younous, S.; Samkaoui, M. COVID-19 associated Kawasaki-like multisystem inflammatory syndrome in an adult. Rev. Esp. Anestesiol. Reanim. Engl. Ed. 2022, 69, 43–47. [Google Scholar] [CrossRef]
- Sharma, C.; Ganigara, M.; Galeotti, C.; Burns, J.; Berganza, F.M.; Hayes, D.A.; Singh-Grewal, D.; Bharath, S.; Sajjan, S.; Bayry, J. Multisystem inflammatory syndrome in children and Kawasaki disease: a critical comparison. Nat. Rev. Rheumatol. 2021, 17, 731–748. [Google Scholar] [CrossRef]
- on behalf of the Rheumatology Study Group of the Italian Pediatric Society; Cattalini, M.; Della Paolera, S.; Zunica, F.; Bracaglia, C.; Giangreco, M.; Verdoni, L.; Meini, A.; Sottile, R.; Caorsi, R.; et al. Defining Kawasaki disease and pediatric inflammatory multisystem syndrome-temporally associated to SARS-CoV-2 infection during SARS-CoV-2 epidemic in Italy: results from a national, multicenter survey. Pediatr. Rheumatol. 2021, 19, 1–11. [Google Scholar] [CrossRef]
- Loke, Y.-H.; Berul, C.I.; Harahsheh, A.S. Multisystem inflammatory syndrome in children: Is there a linkage to Kawasaki disease? Trends Cardiovasc. Med. 2020, 30, 389–396. [Google Scholar] [CrossRef]
- Darby, J.B.; Jackson, J.M. Kawasaki Disease and Multisystem Inflammatory Syndrome in Children: An Overview and Comparison. 2021, 104, 244–252. [Google Scholar]
- Cannon, L.; Campbell, M.J.; Wu, E.Y. Multisystem Inflammatory Syndrome in Children and Kawasaki Disease: Parallels in Pathogenesis and Treatment. Curr. Allergy Asthma Rep. 2023, 23, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.; Raghuveer, G.; Harahsheh, A.; Portman, M.A.; Lee, S.; Khoury, M.; Dahdah, N.; Fabi, M.; Dionne, A.; Harris, T.H.; et al. Cardiac Biomarkers Aid in Differentiation of Kawasaki Disease from Multisystem Inflammatory Syndrome in Children Associated with COVID-19. Pediatr. Cardiol. 2023, 46, 116–126. [Google Scholar] [CrossRef]
- Rivas, M.N.; Arditi, M. Kawasaki Disease and Multisystem Inflammatory Syndrome in Children. Rheum. Dis. Clin. North Am. 2023, 49, 647–659. [Google Scholar] [CrossRef]
- Suzuki, J.; Abe, K.; Matsui, T.; Honda, T.; Yasukawa, K.; Takanashi, J.-I.; Hamada, H. Kawasaki Disease Shock Syndrome in Japan and Comparison With Multisystem Inflammatory Syndrome in Children in European countries. Front. Pediatr. 2021, 9, 625456. [Google Scholar] [CrossRef]
- Lee, S.; Harahsheh, A.S.; Raghuveer, G.; Portman, M.A.; Sabati, A.A.; Khoury, M.; Dahdah, N.; Fabi, M.; Jain, S.S.; Dionne, A.; et al. Spectrum of Coronary Artery Involvement With Multisystem Inflammatory Syndrome in Children Versus Kawasaki Disease. J. Am. Hear. Assoc. 2025, 14, e037761. [Google Scholar] [CrossRef]
- Weiler, C.R.; Austen, K.F.; Akin, C.; Barkoff, M.S.; Bernstein, J.A.; Bonadonna, P.; Butterfield, J.H.; Carter, M.; Fox, C.C.; Maitland, A.; et al. AAAAI Mast Cell Disorders Committee Work Group Report: Mast cell activation syndrome (MCAS) diagnosis and management. J. Allergy Clin. Immunol. 2019, 144, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Ricke, D.O. Vaccine-associated Kawasaki disease in children. Microbes Immun. 2025, 025200044. [Google Scholar] [CrossRef]
- Vignesh, P.; Rawat, A.; Shandilya, J.K.; Sachdeva, M.U.S.; Ahluwalia, J.; Singh, S. Monocyte platelet aggregates in children with Kawasaki disease- a preliminary study from a tertiary care centre in North-West India. Pediatr. Rheumatol. 2021, 19, 1–7. [Google Scholar] [CrossRef]
- Kim, S.H.; Hwang, I.J.; Cho, Y.K. Platelet Indices as Diagnostic Marker for Kawasaki Disease. Chonnam Med J. 2022, 58, 110–118. [Google Scholar] [CrossRef]
- Souni, G.; Ayad, G.; Elouali, A.; Rkain, M.; Babakhouya, A. Severe Kawasaki Disease and Thrombocytopenia: A Case Report. Cureus 2023, 15, e42916. [Google Scholar] [CrossRef]
- Beken, B.; Ünal, Ş.; Çetin, M.; Gümrük, F. The Relationship Between Hematological Findings and Coronary Artery Aneurysm in Kawasaki Disease. Turk. J. Hematol. 2014, 31, 199–200. [Google Scholar] [CrossRef] [PubMed]
- Kocatürk, B.; Lee, Y.; Nosaka, N.; Abe, M.; Martinon, D.; Lane, M.E.; Moreira, D.; Chen, S.; Fishbein, M.C.; Porritt, R.A.; et al. Platelets exacerbate cardiovascular inflammation in a murine model of Kawasaki disease vasculitis. J. Clin. Investig. 2023, 8. [Google Scholar] [CrossRef]
- Ricke, D.O.; Smith, N. VAERS Vasculitis Adverse Events Retrospective Study: Etiology Model of Immune Complexes Activating Fc Receptors in Kawasaki Disease and Multisystem Inflammatory Syndromes. Life 2024, 14, 353. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Xiao, M.; Zhou, D.; Yan, F.; Zhang, Y. Platelet and ferritin as early predictive factors for the development of macrophage activation syndrome in children with Kawasaki disease: A retrospective case-control study. Front. Pediatr. 2023, 11, 1088525. [Google Scholar] [CrossRef]
- VAERS. Vaccine Adverse Event Reporting System. U.S. Department of Health & Human Services, 2025. [Google Scholar]
- Ricke, DO. VAERS-Tools 2025.
- Gupta, K.; Subramanian, H.; Ali, H. Modulation of host defense peptide-mediated human mast cell activation by LPS. J. Endotoxin Res. 2015, 22, 21–30. [Google Scholar] [CrossRef]
- Schwarz, H.; Gornicec, J.; Neuper, T.; Parigiani, M.A.; Wallner, M.; Duschl, A.; Horejs-Hoeck, J. Biological Activity of Masked Endotoxin. Sci. Rep. 2017, 7, srep44750. [Google Scholar] [CrossRef]
- Petruk, G.; Puthia, M.; Petrlova, J.; Samsudin, F.; Strömdahl, A.-C.; Cerps, S.; Uller, L.; Kjellström, S.; Bond, P.J.; Schmidtchen, A. SARS-CoV-2 Spike protein binds to bacterial lipopolysaccharide and boosts proinflammatory activity. J. Mol. Cell Biol. 2020, 12, 916–932. [Google Scholar] [CrossRef]
- Samsudin, F.; Raghuvamsi, P.; Petruk, G.; Puthia, M.; Petrlova, J.; MacAry, P.; Anand, G.S.; Bond, P.J.; Schmidtchen, A. SARS-CoV-2 spike protein as a bacterial lipopolysaccharide delivery system in an overzealous inflammatory cascade. J. Mol. Cell Biol. 2022, 14, 1–14. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Ikeda, K.; Hamaoka, K. Aetiological Significance of Infectious Stimuli in Kawasaki Disease. Front. Pediatr. 2019, 7, 244. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, K.; Yusa, T.; Miyazaki, S.; Ohara, A.; Saji, T. Increased TLR2 and TLR4 Expression in Peripheral Neutrophils Isolated from Kawasaki Disease. Pediatr. Allergy, Immunol. Pulmonol. 2014, 27, 24–29. [Google Scholar] [CrossRef]
- Takeshita, S.; Nakatani, K.; Tsujimoto, H.; Kawamura, Y.; Kawase, H.; Sekine, I. Increased levels of circulating soluble CD14 in Kawasaki disease. Clin. Exp. Immunol. 2000, 119, 376–381. [Google Scholar] [CrossRef]
- Takeshita, S.; Tsujimoto, H.; Kawase, H.; Kawamura, Y.; Sekine, I. Increased Levels of Lipopolysaccharide Binding Protein in Plasma in Children with Kawasaki Disease. Clin. Vaccine Immunol. 2002, 9, 205–6. [Google Scholar] [CrossRef]
- Rice, J.B.; Stoll, L.L.; Li, W.-G.; Denning, G.M.; Weydert, J.; Charipar, E.; Richenbacher, W.E.; Miller, F.J.; Weintraub, N.L. Low-Level Endotoxin Induces Potent Inflammatory Activation of Human Blood Vessels. Arter. Thromb. Vasc. Biol. 2003, 23, 1576–1582. [Google Scholar] [CrossRef]
- Baker, A.L.; Lu, M.; Minich, L.L.; Atz, A.M.; Klein, G.L.; Korsin, R.; Lambert, L.; Li, J.S.; Mason, W.; Radojewski, E.; et al. Associated Symptoms in the Ten Days Before Diagnosis of Kawasaki Disease. J. Pediatr. 2009, 154, 592–595.e2. [Google Scholar] [CrossRef]
- Colomba, C.; La Placa, S.; Saporito, L.; Corsello, G.; Ciccia, F.; Medaglia, A.; Romanin, B.; Serra, N.; Di Carlo, P.; Cascio, A. Intestinal Involvement in Kawasaki Disease. J. Pediatr. 2018, 202, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Tao, E.; Lang, D. Unraveling the gut: the pivotal role of intestinal mechanisms in Kawasaki disease pathogenesis. Front. Immunol. 2024, 15, 1496293. [Google Scholar] [CrossRef]
- Curtis, N.; Zheng, R.; Lamb, J.R.; Levin, M. Evidence for a superantigen mediated process in Kawasaki disease. Arch. Dis. Child. 1995, 72, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Akagawa, S.; Akagawa, Y.; Kimata, T.; Tsuji, S. Our Evolving Understanding of Kawasaki Disease Pathogenesis: Role of the Gut Microbiota. Front. Immunol. 2020, 11, 1616. [Google Scholar] [CrossRef]
- Fabi, M.; Corinaldesi, E.; Pierantoni, L.; Mazzoni, E.; Landini, C.; Bigucci, B.; Ancora, G.; Malaigia, L.; Bodnar, T.; Di Fazzio, G.; et al. Gastrointestinal presentation of Kawasaki disease: A red flag for severe disease? PLOS ONE 2018, 13, e0202658. [Google Scholar] [CrossRef]
- Ricke, D.O. Two Different Antibody-Dependent Enhancement (ADE) Risks for SARS-CoV-2 Antibodies. Front. Immunol. 2021, 12, 640093. [Google Scholar] [CrossRef]
- Ricke, D.; Gherlone, N.; Fremont-Smith, P.; Tisdall, P.; Fremont-Smith, M. Kawasaki Disease, Multisystem Inflammatory Syndrome in Children: Antibody-Induced Mast Cell Activation Hypothesis. J. Pediatr. Pediatr. Med. 2020, 4, 1–7. [Google Scholar] [CrossRef]
- Riollano-Cruz, M.; Akkoyun, E.; Briceno-Brito, E.; Kowalsky, S.; Reed, J.; Posada, R.; Sordillo, E.M.; Tosi, M.; Trachtman, R.; Paniz-Mondolfi, A. Multisystem inflammatory syndrome in children related to COVID-19: A New York City experience. J. Med Virol. 2020, 93, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.F.B.; Litvinov, N.; Farhat, S.C.L.; Eisencraft, A.P.; Gibelli, M.A.B.C.; de Carvalho, W.B.; Fernandes, V.R.; Fink, T.d.T.; Framil, J.V.d.S.; Galleti, K.V.; et al. Severe clinical spectrum with high mortality in pediatric patients with COVID-19 and multisystem inflammatory syndrome. Clinics 2020, 75, e2209. [Google Scholar] [CrossRef]
- Torres, J.P.; Izquierdo, G.; Acuña, M.; Pavez, D.; Reyes, F.; Fritis, A.; González, R.; Rivacoba, C.; Contardo, V.; Tapia, L.I. Multisystem inflammatory syndrome in children (MIS-C): Report of the clinical and epidemiological characteristics of cases in Santiago de Chile during the SARS-CoV-2 pandemic. Int. J. Infect. Dis. 2020, 100, 75–81. [Google Scholar] [CrossRef]
- Sahn, B.; Eze, O.P.; Edelman, M.C.; Chougar, C.E.; Thomas, R.M.; Schleien, C.L.M.; Weinstein, T. Features of Intestinal Disease Associated With COVID-Related Multisystem Inflammatory Syndrome in Children. J. Pediatr. Gastroenterol. Nutr. 2020, 72, 384–387. [Google Scholar] [CrossRef]
- Miller, J.; Cantor, A.; Zachariah, P.; Ahn, D.; Martinez, M.; Margolis, K.G. Gastrointestinal Symptoms as a Major Presentation Component of a Novel Multisystem Inflammatory Syndrome in Children That Is Related to Coronavirus Disease 2019: A Single Center Experience of 44 Cases. Gastroenterology 2020, 159, 1571–1574.e2. [Google Scholar] [CrossRef] [PubMed]
- Kofman, A.D.; Sizemore, E.K.; Detelich, J.F.; Albrecht, B.; Piantadosi, A.L. A young adult with COVID-19 and multisystem inflammatory syndrome in children (MIS-C)-like illness: a case report. BMC Infect. Dis. 2020, 20, 716. [Google Scholar] [CrossRef]
- Tsilioni, I.; Theoharides, T.C. Recombinant SARS-CoV-2 Spike Protein Stimulates Secretion of Chymase, Tryptase, and IL-1β from Human Mast Cells, Augmented by IL-33. Int. J. Mol. Sci. 2023, 24, 9487. [Google Scholar] [CrossRef]
- Chang, L.-S.; Guo, M.M.-H.; Lo, M.-H.; Kuo, H.-C. Identification of increased expression of activating Fc receptors and novel findings regarding distinct IgE and IgM receptors in Kawasaki disease. Pediatr. Res. 2019, 89, 191–197. [Google Scholar] [CrossRef]
- Hrubisko, M.; Danis, R.; Huorka, M.; Wawruch, M. Histamine Intolerance—The More We Know the Less We Know. A Review. Nutrients 2021, 13, 2228. [Google Scholar] [CrossRef] [PubMed]
- Kovacova-Hanuskova, E.; Buday, T.; Gavliakova, S.; Plevkova, J. Histamine, histamine intoxication and intolerance. Allergol. et Immunopathol. 2015, 43, 498–506. [Google Scholar] [CrossRef]
- Ricke, DO. Etiology Scenarios for Multisystem Inflammatory Syndrome in Children and Adults Associated with SARS-CoV-2. J Integr Pediatr Heal n.d., 3, 1–5. [Google Scholar]
- Vikenes, K.; Farstad, M.; Nordrehaug, J.E. Serotonin Is Associated with Coronary Artery Disease and Cardiac Events. Circulation 1999, 100, 483–489. [Google Scholar] [CrossRef]
- Golino, P.; Piscione, F.; Willerson, J.T.; Cappelli-Bigazzi, M.; Focaccio, A.; Villari, B.; Indolfi, C.; Russolillo, E.; Condorelli, M.; Chiariello, M. Divergent Effects of Serotonin on Coronary-Artery Dimensions and Blood Flow in Patients with Coronary Atherosclerosis and Control Patients. New Engl. J. Med. 1991, 324, 641–648. [Google Scholar] [CrossRef]
- Fremont-Smith, M.; Gherlone, N.; Smith, N.; Tisdall, P.; Ricke, D.O. Models for COVID-19 Early Cardiac Pathology Following SARS-CoV-2 Infection. Int. J. Infect. Dis. 2021, 113, 331–335. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y.; Yang, Y.; Su, Z.; Wang, F.; Zhanghuang, C.; Wu, Y.; Zhang, X. Association between FGA gene polymorphisms and coronary artery lesion in Kawasaki disease. Front. Med. 2023, 10, 1193303. [Google Scholar] [CrossRef]
- Levick, S.P.; Meléndez, G.C.; Plante, E.; McLarty, J.L.; Brower, G.L.; Janicki, J.S. Cardiac mast cells: the centrepiece in adverse myocardial remodelling. Cardiovasc. Res. 2010, 89, 12–19. [Google Scholar] [CrossRef]
- Yonker, L.M.; Swank, Z.; Bartsch, Y.C.; Burns, M.D.; Kane, A.; Boribong, B.P.; Davis, J.P.; Loiselle, M.; Novak, T.; Senussi, Y.; et al. Circulating Spike Protein Detected in Post–COVID-19 mRNA Vaccine Myocarditis. Circulation 2023, 147, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Kamat, I.; Hotez, P.J. Myocarditis With COVID-19 mRNA Vaccines. Circulation 2021, 144, 471–484. [Google Scholar] [CrossRef]
- Avolio, E.; Carrabba, M.; Milligan, R.; Williamson, M.K.; Beltrami, A.P.; Gupta, K.; Elvers, K.T.; Gamez, M.; Foster, R.R.; Gillespie, K.; et al. The SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: a potential non-infective mechanism of COVID-19 microvascular disease. Clin. Sci. 2021, 135, 2667–2689. [Google Scholar] [CrossRef]
- Ricke, D.O. Cardiac adverse events post-vaccination. Brain Hear. 2025, 3, 5747. [Google Scholar] [CrossRef]
- Shulman, S.T. Pediatric Coronavirus Disease-2019–Associated Multisystem Inflammatory Syndrome. J. Pediatr. Infect. Dis. Soc. 2020, 9, 285–286. [Google Scholar] [CrossRef]
- Vella, L.A.; Rowley, A.H. Current Insights Into the Pathophysiology of Multisystem Inflammatory Syndrome in Children. Curr. Pediatr. Rep. 2021, 9, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Rostad, C.A.; Chahroudi, A.; Mantus, G.; Lapp, S.A.; Teherani, M.; Macoy, L.; Tarquinio, K.M.; Basu, R.K.; Kao, C.; Linam, W.M.; et al. Quantitative SARS-CoV-2 Serology in Children With Multisystem Inflammatory Syndrome (MIS-C). Pediatrics 2020, 146. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Diorio, C.; Goodwin, E.C.; O McNerney, K.; E Weirick, M.; Gouma, S.; Bolton, M.J.; Arevalo, C.P.; Chase, J.; Hicks, P.; et al. Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2) Antibody Responses in Children With Multisystem Inflammatory Syndrome in Children (MIS-C) and Mild and Severe Coronavirus Disease 2019 (COVID-19). J. Pediatr. Infect. Dis. Soc. 2020, 10, 669–673. [Google Scholar] [CrossRef]
- Gruber, C.N.; Patel, R.S.; Trachtman, R.; Lepow, L.; Amanat, F.; Krammer, F.; Wilson, K.M.; Onel, K.; Geanon, D.; Tuballes, K.; et al. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (MIS-C). Cell 2020, 183, 982–995.e14. [Google Scholar] [CrossRef]
- Moraleda, C.; Serna-Pascual, M.; Soriano-Arandes, A.; Simó, S.; Epalza, C.; Santos, M.; Grasa, C.; Rodríguez, M.; Soto, B.; Gallego, N.; et al. Multi-inflammatory Syndrome in Children Related to Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in Spain. Clin. Infect. Dis. 2020, 72, e397–e401. [Google Scholar] [CrossRef]
- Grazioli, S.; Tavaglione, F.; Torriani, G.; Wagner, N.; Rohr, M.; L’hUillier, A.G.; Leclercq, C.; Perrin, A.; Bordessoule, A.; Beghetti, M.; et al. Immunological Assessment of Pediatric Multisystem Inflammatory Syndrome Related to Coronavirus Disease 2019. J. Pediatr. Infect. Dis. Soc. 2020, 10, 706–713. [Google Scholar] [CrossRef]
- Cirks, B.T.; Rowe, S.J.; Jiang, S.Y.; Brooks, R.M.; Mulreany, M.P.; Hoffner, W.; Jones, O.Y.; Hickey, P.W. Sixteen Weeks Later: Expanding the Risk Period for Multisystem Inflammatory Syndrome in Children. J. Pediatr. Infect. Dis. Soc. 2021, 10, 686–690. [Google Scholar] [CrossRef]
- Al Ameer, H.H.; AlKadhem, S.M.; Busaleh, F.; AlKhwaitm, S.; Llaguno, M.B.B. Multisystem Inflammatory Syndrome in Children Temporally Related to COVID-19: A Case Report From Saudi Arabia. Cureus 2020, 12, e10589. [Google Scholar] [CrossRef]
- Abbas, M.; Törnhage, C.-J. Family Transmission of COVID-19 Including a Child with MIS-C and Acute Pancreatitis. Int. Med Case Rep. J. 2021, ume 14, 55–65. [Google Scholar] [CrossRef]
- Bartsch, Y.C.; Wang, C.; Zohar, T.; Fischinger, S.; Atyeo, C.; Burke, J.S.; Kang, J.; Edlow, A.G.; Fasano, A.; Baden, L.R.; et al. Humoral signatures of protective and pathological SARS-CoV-2 infection in children. Nat. Med. 2021, 27, 454–462. [Google Scholar] [CrossRef]
- Clouser, K.M.; Baer, A.; Bhavsar, S.; Gadhavi, J.M.; Li, S.M.; Schnall, J.; Weiss, J.E. MIS-C After ARDS Associated With SARS-CoV-2. Pediatr. Infect. Dis. J. 2020, 39, e363–e365. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, Y.; Gunji, T.; Burns, J.C.; Shimizu, C.; Newburger, J.W.; Yashiro, M.; Nakamura, Y.; Yanagawa, H.; Wakui, K.; Fukushima, Y.; et al. ITPKC functional polymorphism associated with Kawasaki disease susceptibility and formation of coronary artery aneurysms. Nat. Genet. 2007, 40, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-C.; Hsu, Y.-W.; Wu, C.-M.; Chen, S.H.-Y.; Hung, K.-S.; Chang, W.-P.; Yang, K.D.; Hsieh, K.-S.; Chen, W.-C.; Onouchi, Y. A Replication Study for Association of ITPKC and CASP3 Two-Locus Analysis in IVIG Unresponsiveness and Coronary Artery Lesion in Kawasaki Disease. PLOS ONE 2013, 8, e69685. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, Y.; Ozaki, K.; Buns, J.C.; Shimizu, C.; Hamada, H.; Honda, T.; Terai, M.; Honda, A.; Takeuchi, T.; Shibuta, S.; et al. Common variants in CASP3 confer susceptibility to Kawasaki disease. Hum. Mol. Genet. 2010, 19, 2898–2906. [Google Scholar] [CrossRef]
- Kim, J.; Shimizu, C.; Kingsmore, S.F.; Veeraraghavan, N.; Levy, E.; Dos Santos, A.M.R.; Yang, H.; Flatley, J.; Hoang, L.T.; Hibberd, M.L.; et al. Whole genome sequencing of an African American family highlights toll like receptor 6 variants in Kawasaki disease susceptibility. PLOS ONE 2017, 12, e0170977. [Google Scholar] [CrossRef]
- Khor, C.C.; Davila, S.; Breunis, W.B.; Lee, Y.-C.; et al.; Hong Kong–Shanghai Kawasaki Disease Genetics Consortium; Korean Kawasaki Disease Genetics Consortium; Taiwan Kawasaki Disease Genetics Consortium; International Kawasaki Disease Genetics Consortium; US Kawasaki Disease Genetics Consortium; Blue Mountains Eye Study Genome-wide association study identifies FCGR2A as a susceptibility locus for Kawasaki disease. Nat. Genet. 2011, 43, 1241–1246. [Google Scholar] [CrossRef]
- Duan, J.; Lou, J.; Zhang, Q.; Ke, J.; Qi, Y.; Shen, N.; Zhu, B.; Zhong, R.; Wang, Z.; Liu, L.; et al. A Genetic Variant rs1801274 in FCGR2A as a Potential Risk Marker for Kawasaki Disease: A Case-Control Study and Meta-Analysis. PLOS ONE 2014, 9, e103329. [Google Scholar] [CrossRef] [PubMed]
- Sim, B.K.; Park, H.; Kim, J.-J.; Yun, S.W.; Yu, J.J.; Yoon, K.L.; Lee, K.-Y.; Kil, H.-R.; Kim, G.B.; Han, M.-K.; et al. Assessment of the Clinical Heterogeneity of Kawasaki Disease Using Genetic Variants of BLK and FCGR2A. Korean Circ. J. 2019, 49, 99–108. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Hsu, Y.-W.; Wu, M.-S.; Woon, P.Y.; Wong, H.S.-C.; Tsai, L.-J.; Lin, R.-K.; Klahan, S.; Hsieh, K.-S.; Chang, W.-C. FCGR2A Promoter Methylation and Risks for Intravenous Immunoglobulin Treatment Responses in Kawasaki Disease. Mediat. Inflamm. 2015, 2015, 564625. [Google Scholar] [CrossRef]
- Cariaso, M.; Lennon, G. SNPedia: a wiki supporting personal genome annotation, interpretation and analysis. Nucleic Acids Res. 2011, 40, D1308–D1312. [Google Scholar] [CrossRef]
- Kwon, Y.-C.; Kim, J.-J.; Yun, S.W.; Yu, J.J.; Yoon, K.L.; Lee, K.-Y.; Kil, H.-R.; Kim, G.B.; Han, M.-K.; Song, M.S.; et al. Male-specific association of the FCGR2A His167Arg polymorphism with Kawasaki disease. PLOS ONE 2017, 12, e0184248. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, Y. Molecular Genetics of Kawasaki Disease. Pediatr. Res. 2009, 65, 46R–54R. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.C. The etiologies of Kawasaki disease. J. Clin. Investig. 2024, 134. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Kuo, H.-C.; Chang, J.-S.; Chang, L.-Y.; Huang, L.-M.; Chen, M.-R.; Liang, C.-D.; Chi, H.; Huang, F.-Y.; Lee, M.-L.; et al. Two new susceptibility loci for Kawasaki disease identified through genome-wide association analysis. Nat. Genet. 2012, 44, 522–525. [Google Scholar] [CrossRef]
- Onouchi, Y.; Ozaki, K.; Burns, J.C.; Shimizu, C.; Terai, M.; Hamada, H.; Honda, T.; Suzuki, H.; et al.; Japan Kawasaki Disease Genome Consortium; US Kawasaki Disease Genetics Consortium A genome-wide association study identifies three new risk loci for Kawasaki disease. Nat. Genet. 2012, 44, 517–521. [Google Scholar] [CrossRef]
- Chang, C.-J.; Kuo, H.-C.; Chang, J.-S.; Lee, J.-K.; Tsai, F.-J.; Khor, C.C.; Chang, L.-C.; Chen, S.-P.; Ko, T.-M.; Liu, Y.-M.; et al. Replication and Meta-Analysis of GWAS Identified Susceptibility Loci in Kawasaki Disease Confirm the Importance of B Lymphoid Tyrosine Kinase (BLK) in Disease Susceptibility. PLOS ONE 2013, 8, e72037. [Google Scholar] [CrossRef]
- Shrestha, S.; Wiener, H.; Shendre, A.; Kaslow, R.A.; Wu, J.; Olson, A.; Bowles, N.E.; Patel, H.; Edberg, J.C.; Portman, M.A. Role of Activating Fc γ R Gene Polymorphisms in Kawasaki Disease Susceptibility and Intravenous Immunoglobulin Response. Circ. Cardiovasc. Genet. 2012, 5, 309–316. [Google Scholar] [CrossRef]
- Shrestha, S.; Wiener, H.W.; Olson, A.K.; Edberg, J.C.; Bowles, N.E.; Patel, H.; Portman, M.A. Functional FCGR2B gene variants influence intravenous immunoglobulin response in patients with Kawasaki disease. J. Allergy Clin. Immunol. 2011, 128, 677–680.e1. [Google Scholar] [CrossRef] [PubMed]
- Makowsky, R.; Wiener, H.W.; Ptacek, T.S.; Silva, M.; Shendre, A.; Edberg, J.C.; Portman, M.A.; Shrestha, S. FcγR gene copy number in Kawasaki disease and intravenous immunoglobulin treatment response. Pharmacogenetics Genom. 2013, 23, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, S.Q.; Schmidt, D.E.; de Haas, M.; Kuijpers, T.W. Genetic Variation in Low-To-Medium-Affinity Fcγ Receptors: Functional Consequences, Disease Associations, and Opportunities for Personalized Medicine. Front. Immunol. 2019, 10, 2237. [Google Scholar] [CrossRef]
- Johnson, T.A.; Mashimo, Y.; Wu, J.-Y.; Yoon, D.; Hata, A.; Kubo, M.; Takahashi, A.; Tsunoda, T.; Ozaki, K.; Tanaka, T.; et al. Association of an IGHV3-66 gene variant with Kawasaki disease. J. Hum. Genet. 2020, 66, 475–489. [Google Scholar] [CrossRef]
- Onouchi, Y.; Fukazawa, R.; Yamamura, K.; Suzuki, H.; Kakimoto, N.; Suenaga, T.; Takeuchi, T.; Hamada, H.; Honda, T.; Yasukawa, K.; et al. Variations in ORAI1 Gene Associated with Kawasaki Disease. PLOS ONE 2016, 11, e0145486–e0145486. [Google Scholar] [CrossRef]
- Kim, J.-J.; Park, Y.-M.; Yoon, D.; Lee, K.-Y.; Song, M.S.; Lee, H.D.; Kim, K.-J.; Park, I.-S.; Nam, H.-K.; et al.; Korean Kawasaki Disease Genetics Consortium Identification of KCNN2 as a susceptibility locus for coronary artery aneurysms in Kawasaki disease using genome-wide association analysis. J. Hum. Genet. 2013, 58, 521–525. [Google Scholar] [CrossRef]
- Shimizu, C.; Jain, S.; Davila, S.; Hibberd, M.L.; Lin, K.O.; Molkara, D.; Frazer, J.R.; Sun, S.; Baker, A.L.; Newburger, J.W.; et al. Transforming Growth Factor-β Signaling Pathway in Patients With Kawasaki Disease. Circ. Cardiovasc. Genet. 2011, 4, 16–25. [Google Scholar] [CrossRef]
- Choi, Y.M.; Shim, K.S.; Yoon, K.L.; Han, M.Y.; Cha, S.H.; Kim, S.K.; Jung, J.H. Transforming growth factor beta receptor II polymorphisms are associated with Kawasaki disease. Korean J. Pediatr. 2012, 55, 18–23. [Google Scholar] [CrossRef]
- Cho, J.H.; Han, M.Y.; Cha, S.H.; Jung, J.H.; Yoon, K.L. Genetic Polymorphism of SMAD5 is Associated With Kawasaki Disease. Pediatr. Cardiol. 2013, 35, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Faroldi, G.; Melo, F.R.; Rönnberg, E.; Grujic, M.; Pejler, G. Active Caspase-3 Is Stored within Secretory Compartments of Viable Mast Cells. J. Immunol. 2013, 191, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.M.; Hanes, M.R.; Marshall, J.S. Mast Cell Modulation of B Cell Responses: An Under-Appreciated Partnership in Host Defence. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Hu, W. Identifying differentially expressed genes and miRNAs in Kawasaki disease by bioinformatics analysis. Sci. Rep. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Gu, W.; Mirsaidi-Madjdabadi, S.; Ramirez, F.; Simonson, T.S.; Makino, A.; Gu, \. Transcriptome meta-analysis of Kawasaki disease in humans and mice. Front. Pediatr. 2024, 12, 1423958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, Y.; Meng, L.; Ye, C.; Han, H.; Zhang, T.; Feng, Y.; Li, J.; Duan, L.; Chen, Y. Whole-exome sequencing analysis identifies novel variants associated with Kawasaki disease susceptibility. Pediatr. Rheumatol. 2023, 21, 1–7. [Google Scholar] [CrossRef]
- Yeşiltepe, E.; Duman, D.; Kuyucu, N.; Bozdoğan, S.T.; Çıtırık, L.; Yeşil, E.; Karpuz, D. FCGR2A Gene Polymorphism Association in Children with Multisystem Inflammatory Syndrome. Indian Pediatr. 2025, 62, 372–377. [Google Scholar] [CrossRef]
- Jia, S.; Li, C.; Wang, G.; Yang, J.; Zu, Y. The T helper type 17/regulatory T cell imbalance in patients with acute Kawasaki disease. Clin. Exp. Immunol. 2010, 162, 131–137. [Google Scholar] [CrossRef]
- CDC. Weekly U.S. MIS-C Cases and COVID-19 Percent Positivity Reported to CDC 2025.
- Centers for Disease Control and Prevention. COVID data tracker - Wastewater Suveillance (2021). Available online: https://covid.cdc.gov/covid-data-tracker/#wastewdatatracker-home (accessed on 11 April 2022).
Figure 1.
Kawasaki disease normalized frequency by year of age 0, 1, and 2.

Figure 2.
Kawasaki disease normalized frequency for infants aged 0 (A) concomitantly administered vaccines, (B) number of occurrences in concomitantly administered vaccines, and (C) with AE death (Pearson r=0.66).
Figure 2.
Kawasaki disease normalized frequency for infants aged 0 (A) concomitantly administered vaccines, (B) number of occurrences in concomitantly administered vaccines, and (C) with AE death (Pearson r=0.66).



Figure 3.
MIS normalized frequency by age for COVID-19 (Pfizer-BioNTech).

Figure 4.
MIS normalized frequency by COVID-19 (Pfizer-BioNTech) manufacturing lot.

Figure 5.
Kawasaki disease and MIS onset day (Pearson r=0.90).

Figure 6.
Kawasaki disease normalized frequency by gender.

Figure 7.
Kawasaki disease normalized frequency according to vaccine dose.


Table 1.
Kawasaki and MIS etiologsy model disease factors, mast cell activators, and predicted IVIG resistance.
Table 1.
Kawasaki and MIS etiologsy model disease factors, mast cell activators, and predicted IVIG resistance.
| Primary factor | Additional factor | Mast cell activator(s) | Likely IVIG resistance |
| neonate pathogen infection | maternally transferred antibodies (MatAbs) | high Ab titers | very low |
| pathogen infection | elevated Ab titers (ongoing, prior infections, …) | high Ab titers | very low |
| immunization | elevated Ab titers | high Ab titers | very low |
| immunization | manufacturing contaminant(s)—endotoxin | endotoxin or bacterial components | high |
| immunization | live attenuated vaccine GI pathogen (e.g., rotavirus)—possible disruption of gut barrier | possible high Ab titers and/or bacterial components | variable depending upon mast cell activators |
| GI infection | disruption of gut barrier [99] | endotoxin or bacterial components | high |
| environmental exposures including increased temperature | genetic risk factor | direct mast cell activation | high |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



