Submitted:
25 February 2026
Posted:
25 February 2026
Read the latest preprint version here
Abstract
Extremophilic bacteria survive salt, temperature, and pH extremes by coordinating stress-induced protein networks that preserve macromolecules, sustain energetics, and repair damage. This review integrates recent proteomics with functional genomics to resolve both network state and causality across halophiles, thermophiles, acidophiles, and alkaliphiles, with targeted contrasts from psychrophiles and radiation-resistant bacteria. Quantitative proteomics maps condition-specific induction of chaperones, proteases, ion transporters, osmolyte pathways, DNA repair proteins, antioxidants, and envelope remodelling enzymes. Complementary perturbation genetics/functional genomics and transcriptomics help identify essential nodes and regulatory circuits underlying stress tolerance. In halophiles, compatible solute synthesis and Na+/H+ exchange couple to protein quality control and central metabolism. Thermophiles rely on heat-shock systems, ATP-dependent proteolysis, membrane adjustments, and redox balancing. Acidophiles maintain near-neutral cytosol via proton export and low-permeability membranes while linking iron handling to oxidative defence. Alkaliphiles use Na+-based bioenergetics, multi-subunit antiporters, and cell wall modifications to retain protons. Psychrophiles emphasize cold-shock RNA chaperones, flexible enzymes, and cryoprotectants, whereas radiophiles combine exceptional DNA repair with strong antioxidant capacity. Across clades, oxidative stress forms a cross-cutting axis that explains extensive regulon overlap and cross-protection. We synthesize network architecture, highlight conserved modules and lineage-specific solutions, and outline open questions in stress sensing, multi-stress integration, and functions of uncharacterized proteins. These insights provide a framework for engineering robust biocatalysts and organisms for biotechnology and environmental applications.
Keywords:
extremophilic bacteria
; functional genomics
; osmoadaptation
; oxidative stress
; proteomics
; stress-induced protein networks
1. Introduction
Extremophilic bacteria thrive under conditions lethal to most life, from hypersaline brines and hot springs to acidic mine drainage and alkali lakes. These environments impose severe stress on cellular macromolecules, causing protein misfolding, membrane damage, and DNA lesions. To survive, extremophiles have evolved robust stress response networks that sense environmental changes and rapidly reprogram cellular physiology [1]. Proteomics studies reveal that under stress, extremophiles broadly upregulate chaperones, DNA repair enzymes, transporters, and antioxidant proteins, suggesting a coordinated protective response. In parallel, perturbation-based functional genomics (e.g., targeted knockouts/knockdowns and pooled genetic screens such as transposon mutagenesis with sequencing readout) can test causality by identifying genes required for stress tolerance, while transcriptomics provides complementary expression context for the same stress response networks [2]. Nucleoid-associated proteins (NAPs) can also contribute to stress adaptation by modulating chromosome architecture and global transcription programs. For example, H-NS couples nucleoid structuring with transcriptional silencing and is responsive to environmental cues such as temperature, while HU and IHF-family proteins influence DNA topology and promoter architecture. In addition, DNA-binding protective proteins such as Dps can compact and shield DNA and limit iron-driven oxidative damage during stress. In radiation-resistant Deinococcus, stress-triggered nucleoid remodelling has been linked to changes in HU abundance and dynamics, supporting a model in which chromosome organization is one regulatory layer that interfaces with repair, redox control, and proteostasis [3]. Integrating proteomic “snapshots” of the stress-adjusted proteome with functional genomic evidence is therefore critical for deciphering how these networks operate mechanistically. In this review, we distinguish between expression-based omics (transcriptomics and proteomics), which quantify condition-dependent RNA and protein abundance and thereby describe network “state”, and perturbation-based functional genomics, which links genotype to phenotype through genetic perturbations and screening (e.g., gene deletions, knockdowns, or genome-wide insertion/CRISPR-based libraries). Expression profiling is primarily correlative, whereas perturbation-based functional genomics provides direct evidence for causal contributions of specific genes and pathways to stress tolerance.
Notably, many extremophiles face multiple concurrent stresses. For example, acidophilic bacteria in acid mine drainage endure low pH and high heavy metal levels, while haloalkaliphiles tolerate both high salinity and high pH. Such conditions demand cross-protection strategies [4,5]. Common modules – protein quality control systems, compatible solute pathways, redox-balancing enzymes, and membrane transporters – recur across different stress conditions. Cross-talk is evident in cases like alkaliphilic Bacillus marmarensis, which at pH 10 showed minimal induction of classical heat or salt shock proteins but did upregulate oxidative stress defenses, suggesting that high pH primarily imposes an oxidative challenge [6,7]. Likewise, the famously radiation-resistant Deinococcus radiodurans simultaneously combats desiccation, oxidative damage, and DNA breaks by mobilizing potent antioxidants (e.g. manganese complexes) and unique DNA repair proteins (PprA, Ddr proteins) that are strongly induced upon irradiation [8,9]. These examples underscore that extremophiles leverage a flexible, overlapping stress response toolkit. In the following sections, we examine how proteomic and functional genomic studies have illuminated stress-induced protein networks in major classes of extremophilic bacteria halophiles, thermophiles, acidophiles, and alkaliphiles emphasizing the integration of protein-level insights with genetic and transcriptomic evidence to understand adaptive mechanisms [10].
2. Osmotic Stress in Halophilic Bacteria
Halophilic and halotolerant bacteria endure high salinity by rigorously managing osmotic balance and protein stability. Proteomic analyses show that when exposed to hyper-saline conditions, these cells markedly increase proteins involved in synthesizing or importing compatible solutes (osmoprotectants) and in expelling excess ions [11]. For instance, under salt up-shock, Halomonas spp. and other halophiles induce enzymes for ectoine and trehalose biosynthesis as well as transporters for glycine betaine and proline uptake, which collectively raise the internal osmolyte pool and counteract water efflux [1]. Concurrently, membrane-bound Na+/H+ antiporters and ATP-driven pumps are upregulated to extrude Na+ and maintain ionic homeostasis. Proteogenomic studies on a halotolerant Tistlia bacterium captured this pattern: at high salt, Tistlia’s exoproteome became enriched in osmolyte transporters and osmosensing proteins, alongside flagellar and chemotaxis proteins that were not seen at optimal salinity [1]. Induction of motility under stress suggests cells may actively seek microenvironments with more favorable osmotic conditions. Conversely, under sudden low-salinity (dilution) stress, halophiles activate mechanosensitive channel proteins that act as emergency valves to rapidly release internal solutes and prevent lysis a response confirmed by genetic studies of osmotic down-shock survival in multiple bacteria [12]. Proteomics has also revealed less obvious osmotic adaptations; for example, Tistlia showed elevated levels of a hopanoid biosynthesis enzyme (HpnM) at low external salt, hinting that reinforcing the membrane with hopanoids may reduce water influx when osmotic pressure drops [1]. This novel insight, later supported by mutational evidence from other bacteria that hopanoid-lacking strains are osmosensitive, exemplifies how proteomics can discover unanticipated stress defenses [13].
Functional genomics corroborates the essential roles of these osmo adaptive proteins. Transcriptomic profiling of an alkaliphilic halotolerant Bacillus identified dozens of salt-responsive genes, including those for ectoine synthesis, ion transporters, and universal stress proteins [14]. Knockout and transposon mutant studies in several halophiles have shown that disrupting major osmo protectant pathways (such as ectoine or trehalose biosynthesis) or mechanosensitive channel genes dramatically impairs growth at high salinity, establishing causality for the proteomic observations [15]. Moreover, comparative genomics reveals that halophilic bacteria often possess redundant or uniquely modified systems for osmotic stress: multiple transporters for K+ uptake (to counterbalance Na+) and expanded families of ion channels and solute-binding proteins [16,17]. These expansions likely ensure robustness in fluctuating salinity. In summary, an integrated picture emerges in which proteomic network shifts (increased osmolyte production, ion pumping, chaperones, and cell envelope modifications) form the immediate response to salt stress, while genetic validation / perturbation-based functional genomic evidence pinpoints the specific proteins (e.g. Ectoine synthase, Na+/H+ antiporters, mechanosensitive channels) that are critical for halophile survival under osmotic duress. Together, these approaches highlight a highly coordinated osmotic stress network centered on maintaining protein structure and turgor pressure in extreme salt [18].
3. Thermal Stress in Thermophilic Bacteria
Thermophilic bacteria, adapted to high temperatures (often 60–100 °C), must prevent heat-induced protein unfolding and membrane destabilization. Proteomic surveys show that upon sudden heat shock or when pushed to upper temperature limits, thermophiles strongly induce classic heat-shock proteins (molecular chaperones like GroEL/GroES, DnaK/DnaJ, and small heat shock proteins) that refold or sequester misfolded proteins [19,20]. Simultaneously, ATP-dependent proteases (Lon, ClpPX) are upregulated to degrade irreversibly damaged proteins. Comparative genomics of thermophiles indicates they often harbor multiple copies of these proteostasis genes, ensuring a robust folding/refolding capacity [21]. In extreme acidothermophiles, for example, an abundance of folding chaperones and protease subunits is maintained genomically, with many genes present in duplicates or clusters for coordinated regulation [21]. Proteomic data confirm these proteins surge during heat stress, underscoring their central role in maintaining proteome integrity at high temperature.
Beyond chaperones, thermophilic bacteria remodel their metabolism and cell envelope in response to heat. An integrated proteomic-transcriptomic analysis of Alicyclobacillus acidoterrestris (a thermoacidophile) revealed that heat exposure provoked changes in cell envelope architecture: proteins for fatty acid synthesis and cell wall remodeling were regulated to adjust membrane fluidity and peptidoglycan cross-linking, thereby maintaining membrane stability at elevated temperatures [22]. Notably, many heat-induced changes in A. acidoterrestris occurred at the protein level without corresponding mRNA changes, implying extensive post-transcriptional control to rapidly fine-tune the proteome [22]. This highlights the value of integrating proteomics with transcriptomics: transcript levels alone can miss major protein-level regulation, and perturbation-based functional genomics is then needed to test which components are causal for heat tolerance. The heat resistance of A. acidoterrestris, like many thermophiles, ultimately relied on fortifying the cell envelope and preserving protein function rather than novel heat-specific proteins [22]. Proteomic time-course studies in other thermophiles have similarly shown a transient burst of glycolytic and energy metabolism enzymes, thought to fuel the energy-intensive processes of protein refolding and repair during acute heat stress.
Thermal stress often triggers secondary stresses, and proteogenomic data capture this interplay. In a thermophilic Geobacillus, heat shock not only upregulated expected chaperones but also disrupted electron transport processes, leading to an accumulation of NADH and a shortfall in oxygen utilization [23]. The cell responded by down-regulating several NADH-dependent dehydrogenases and up-regulating alternative dehydrogenases that use FADH2, a shift that inadvertently increased reactive oxygen species (ROS) production [23]. Consistent with this, proteomics detected induction of superoxide dismutase and suppression of iron uptake proteins (to limit Fenton chemistry), while transcripts indicated a temporary switch to anaerobic respiration pathways [23]. These findings illustrate that thermophiles coordinate their stress responses holistically: heat shock proteins address protein folding, whereas metabolic and redox adjustments mitigate oxidative stress caused by high-temperature metabolism. Genetic validation comes from studies showing that mutants lacking key antioxidant enzymes or heat-shock chaperones in thermophiles suffer reduced heat tolerance, confirming that both arms – proteostasis and redox balance are vital for surviving thermal extremes [24].
4. pH Extremes: Acidophiles and Alkaliphiles
Bacteria inhabiting extremely low or high pH environments deploy distinctive yet analogous strategies to maintain intracellular homeostasis. Acidophilic bacteria (optimal pH ≤ 3) face an onslaught of protons and often dissolved metals. Proteomic and genomic analyses indicate that extreme acidophiles possess highly redundant protein homeostasis systems to cope with protein stress [25]. A recent comparative genomics survey of iron- and sulfur-oxidizing acidophiles found an enrichment and redundancy of genes encoding chaperones (e.g. multiple small Hsp20 holdases, extra copies of GroEL/GroES) and proteases (ClpP/ClpX, Lon) compared to neutrophiles, with many of these genes clustered in regulons. This suggests acidophiles have an especially large “buffer” capacity for refolding or removing damaged proteins, which is consistent with the constant protein-denaturing conditions at low pH [21]. Proteomic studies confirm that acid-challenged cells markedly elevate their chaperones and proteases, keeping proteins correctly folded even as protons and metal ions destabilize structures. DNA-binding protective proteins, including some NAPs, may contribute to acid/alkali adaptation by promoting stress-associated DNA protection/compaction and by influencing global transcription programs; however, direct causal links to proton-homeostasis modules remain limited in most extremophiles and should be interpreted cautiously without perturbation-based evidence [3]. Additionally, acidophiles reinforce their cell membranes – high levels of hopanoids, lipid modifications, and membrane spanning proteins are observed – to minimize proton leakage into the cytoplasm. They also rely on robust proton extrusion systems. Although membrane proteins can be under-represented in some proteomic assays, transcriptomic data in model acidophiles (e.g. Acidithiobacillus spp.) show strong upregulation of H+-ATPases and Na+/H+ antiporters under low pH, indicating active pumping of H+ out and cation influx to maintain a near-neutral cytosol [26]. Mutational evidence supports this: strains of acidophiles with defective proton pumps or membrane lipid synthesis have dramatically impaired growth below pH 3, underscoring the importance of membrane-level defenses alongside proteostasis.
Alkaliphilic bacteria, thriving at pH 10–11, conversely must retain protons and exclude excess hydroxide. Proteomic and transcriptomic profiling of the alkaliphile Bacillus marmarensis under high pH revealed a strategy centered on cell envelope remodeling and ion homeostasis [6]. As external pH rose, B. marmarensis altered its peptidoglycan peptide composition and membrane lipid saturation to reduce membrane fluidity and proton leak, effectively tightening the cell boundary against proton loss [6]. The bacterium also induced a suite of cation transporters, notably not only the expected Na+ transporters (common in alkaliphiles using Na+ in place of H+ for bioenergetics) but also K+ transport systems, indicating a broader ionic response to alkali stress [6]. Intriguingly, proteomics showed little increase in general stress proteins at high pH heat shock proteins and nutrient stress responders were unchanged implying that B. marmarensis does not perceive alkaline conditions as a classic “stress” in the way a non-adapted organism would [6]. Instead, the main detected response was an oxidative stress signature: genes encoding catalases, peroxiredoxins, and other ROS-detoxifying enzymes were upregulated, hinting that alkalinity may indirectly provoke oxidative stress, perhaps through altered redox balance or metal availability [7]. This finding aligns with the concept of cross-stress modules: even in an alkaliphile well-adapted to pH 11, any residual physiological strain manifests as oxidative stress, which is countered by antioxidant proteins. Functional genomics reinforces the importance of the observed adaptations. Deletion of genes involved in cell wall teichoic acid synthesis or fatty acid cyclopropanation (processes that acidify or rigidify the cell surface) renders moderate alkaliphiles far less pH-tolerant, and mutants in Na+/H+ antiport systems likewise fail to grow at high pH, demonstrating that these proteome shifts are essential, not incidental [7]. Alkaliphiles also concentrate cytoplasmic polyamines and negatively charged polymers at the cell surface to attract protons a feature supported by proteomic detection of amino-acid decarboxylases and secreted acidic polymer-binding proteins in some species.
5. Comparative Synthesis of Stress Networks Across Extremophiles
Extremophilic bacteria display a remarkable convergence of stress response strategies despite the diversity of extreme environments they inhabit. Comparative analyses reveal recurring molecular modules that safeguard cellular integrity, including protein chaperones, antioxidant enzymes, DNA repair systems, osmolyte synthesis pathways, and dedicated regulatory circuits [27,28]. These core defenses are mobilized in different combinations: for example, thermophiles, halophiles, acidophiles, and alkaliphiles all induce proteostasis and DNA protection mechanisms tailored to their specific stresses [29]. Integrative proteomics and genomics have been instrumental in mapping these networks, showing that many extremophiles share a “toolkit” of stress proteins even as the exact components and expression levels vary with habitat [27]. Community metagenomes from extreme habitats often encode diverse NAP-like genes, suggesting that variation in chromosome-organization factors may contribute to differences in stress-responsive regulation across lineages; however, the functional roles of specific variants under combined stresses require targeted experimental validation [30]. Interestingly, in some cases a few highly specialized proteins can effectively neutralize a dominant environmental stressor – for instance, a single robust enzyme or pigment may suffice to counter extreme heat, salinity, or radiation if it directly targets the key damage pathway [29]. Such findings underscore that extreme habitats exert strong selective pressure for efficient, minimalist solutions, leading to both common and unique adaptations across extremophilic taxa.
Figure 1.
Comparative stress-adaptation profiles across extremophile groups. Bar plots summarize the relative contribution of key functional modules (proteostasis, antioxidants, DNA repair, osmolytes, and ion transport) in halophiles, thermophiles, acidophiles, alkaliphiles, psychrophiles, and radiophiles. Colors indicate qualitative level: low (1), medium (2), and high (3).
Figure 1.
Comparative stress-adaptation profiles across extremophile groups. Bar plots summarize the relative contribution of key functional modules (proteostasis, antioxidants, DNA repair, osmolytes, and ion transport) in halophiles, thermophiles, acidophiles, alkaliphiles, psychrophiles, and radiophiles. Colors indicate qualitative level: low (1), medium (2), and high (3).
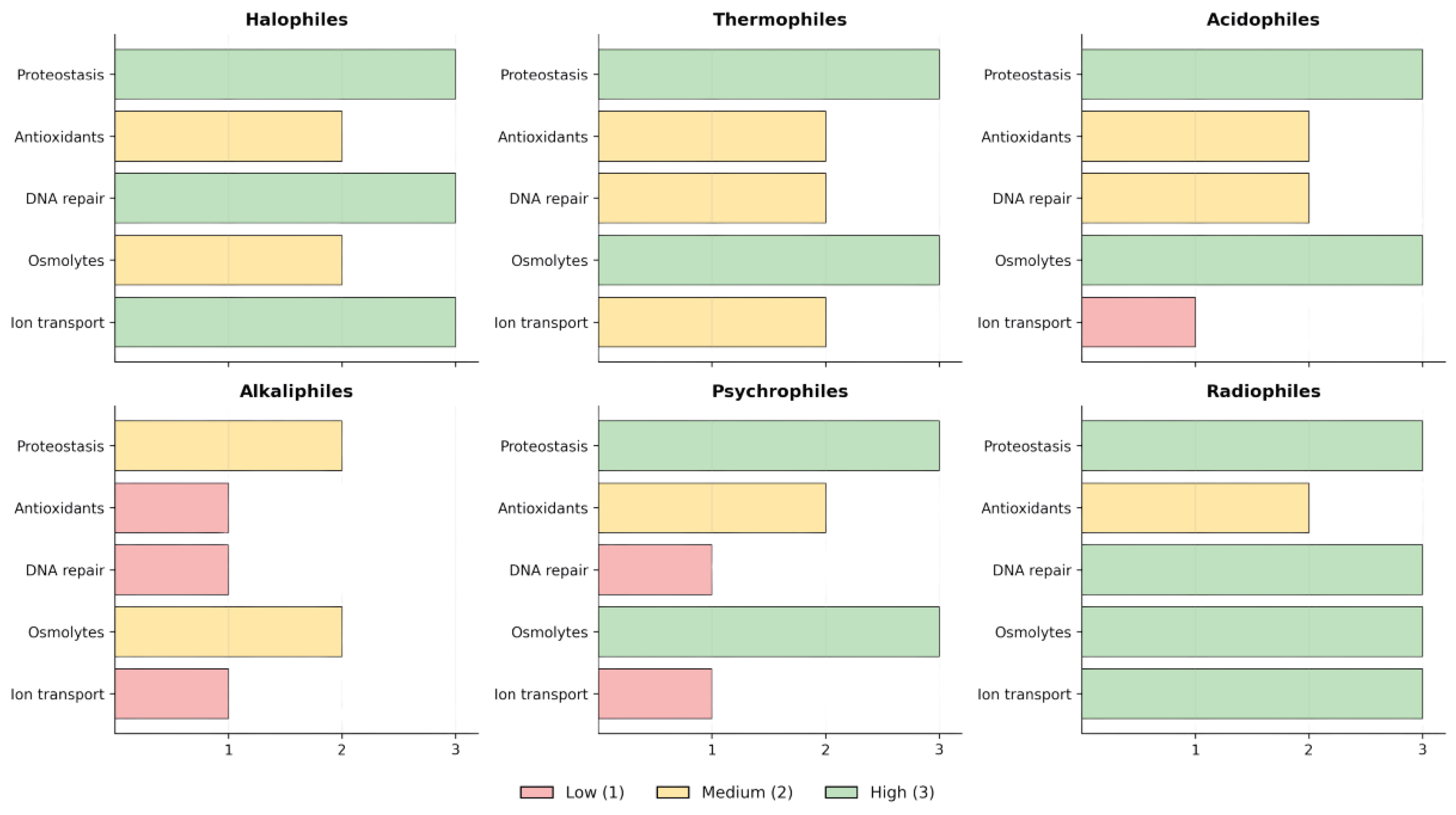
A universal theme is the reliance on molecular chaperones to maintain proteome stability under harsh conditions. Heat-shock proteins (HSPs) such as Hsp70 (DnaK) and Hsp90 are induced not only in thermophiles facing acute thermal stress but also in other extremophiles where misfolded proteins accumulate. These chaperones assist in refolding or degrading damaged proteins, preventing toxic aggregation and preserving enzymatic functions at temperature, pH, or salt extremes [29]. Notably, small heat-shock proteins are found across virtually all classes of extremophiles (Table 1), from acidophilic bacteria to hyperthermophilic archaea, acting as holdases that stabilize unfolding proteins [31]. Proteomic surveys frequently identify HSPs among the most upregulated proteins upon stress exposure, highlighting their central role in extremophile viability. Functional genomics studies further validate this role: deletion or inhibition of key chaperone genes often cripples stress tolerance, confirming that robust chaperone networks are indispensable for life in extreme settings [27]. In sum, maintaining proteostasis via chaperone machinery is a common strategy threading through the biology of otherwise disparate extremophiles.
Figure 2.
The overlap of environmental stressors in polyextremophiles highlighting microorganisms that tolerate multiple extreme conditions (polyextremophiles).
Figure 2.
The overlap of environmental stressors in polyextremophiles highlighting microorganisms that tolerate multiple extreme conditions (polyextremophiles).
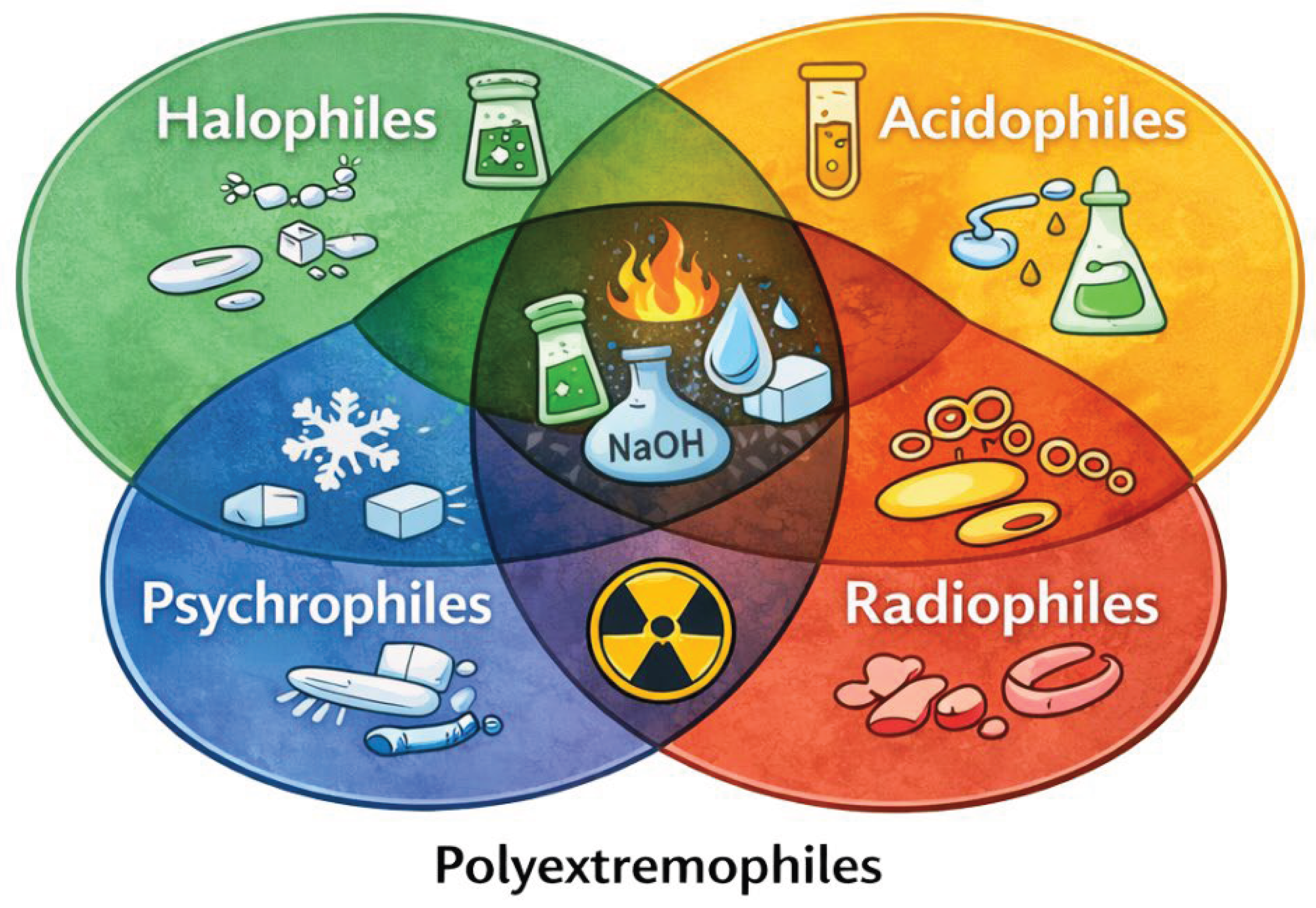
Another recurring pillar is the deployment of potent antioxidant defenses. Diverse extreme conditions high temperature, high salinity, acidic pH, radiation, or desiccation—ultimately funnel into oxidative stress by generating reactive oxygen species (ROS) that damage proteins, DNA, and lipids. Accordingly, extremophiles universally enhance their antioxidant enzyme repertoire to detoxify ROS [21]. Hyperthermophiles and acidophiles commonly express elevated levels of superoxide dismutases, peroxidases, and catalases to scavenge oxygen radicals and peroxides formed as by-products of thermal or chemical stress [38]. Halophilic microbes inhabiting solar salterns or alkaline lakes constitutively produce carotenoid pigments and enzymes that quench singlet oxygen and peroxides generated by intense UV and osmotic stress [39]. Even radiation-tolerant bacteria leverage robust redox control systems for example, Deinococcus cells are rich in catalases, peroxiredoxins, and thioredoxin-dependent enzymes that preserve the reduced state of the proteome under barrage of ionizing radiation [40]. Proteomic profiles across extremophiles consistently show antioxidant proteins among the top induced factors during stress challenges, illustrating that controlling ROS is a unifying priority in extreme environments.
Extremophiles also converge on strategies to protect genetic information and repair macromolecular damage. Efficient DNA repair pathways are critical in habitats where damage to DNA is frequent – whether due to UV radiation in high-altitude deserts, desiccation in saline flats, or acid-induced hydrolytic damage in mine drainages [28]. Comparative genomics reveals expansions or enhancements of DNA repair genes in many extremophiles, including multiple DNA ligases, excision repair enzymes, recombinases (RecA and RecBCD systems), and photolyases for UV lesions [41]. High-performance DNA repair is perhaps most celebrated in radiation-resistant bacteria, but even thermophiles and acidophiles rely on faithful repair to counteract spontaneous damage accelerated by heat or low pH [41]. Proteomic studies often detect upregulation of DNA repair proteins (e.g. UvrABC excision nucleases, RecA recombinase, DNA glycosylases) when extremophiles are stressed, indicating activation of genome maintenance modules as part of the adaptive response [40]. Moreover, many extremophiles possess DNA-protective proteins that shield chromosomes from damage – for example, DPS family DNA-binding proteins that sequester iron and prevent hydroxyl radical formation, or unique nucleoid-associated proteins that compact DNA during stress. Moreover, many extremophiles possess DNA-protective proteins that can reduce lesion formation and complement repair. For example, Dps-family proteins can sequester iron and bind DNA, helping limit hydroxyl radical chemistry while promoting stress-associated DNA protection and compaction [42]. Consistent with this idea, live-cell studies in Deinococcus show that UV or irradiation can trigger rapid nucleoid remodeling linked to altered HU dynamics, indicating that stress-induced chromosome reorganization can accompany genome maintenance. We therefore refer to NAPs and related DNA-binding protective factors as contributors to genome protection and regulation during stress (Table 2), while emphasizing that causal roles should be supported by perturbation-based evidence when available [43,44].
Modulation of cellular chemistry through osmolytes and membrane adaptations is another shared theme. Extremophilic bacteria frequently accumulate compatible solutes small molecules like trehalose, glycine betaine, ectoine, or diglycerol phosphate to counteract osmotic stress and stabilize proteins without interfering with metabolism [31]. Halophiles are a classic example, either concentrating KCl to molar levels or synthesizing organic osmolytes to balance the external salt, and proteomics confirms high expression of osmolyte transporters and synthesis enzymes in salt-stressed cells [39]. Psychrophiles and desiccation-tolerant microbes likewise produce sugars (glucose, trehalose) and polyols (glycerol, mannitol) as cryoprotectants, which depress the freezing point and also act as chaperone-like stabilizers for proteins and membranes [31]. Membrane lipid remodeling works in concert with osmolytes: extremophiles adjust fatty acid saturation, hopanoid content, or ether lipid chemistry to maintain membrane fluidity and integrity at extreme temperature or pH [27]. The genes for these adaptive traits (e.g. desaturases, cyclopropane synthases, cardiolipin synthases) are often identified in extremophile genomes and show elevated transcript/protein levels under stress conditions. Together, these biochemical adjustments create an intracellular milieu and membrane state permissive for life under otherwise chaotropic or denaturing external conditions [21]. In cold soils, desiccation–rehydration and mineral interfaces further bias the balance between osmolyte and antioxidant modules [45].
Figure 3.
Overview of bacterial stress response pathways. Core stress regulators (σ factors, IrrE/PprI, and two-component systems) control key protective functions, including proteostasis, redox/antioxidant defense, DNA repair, osmolyte synthesis, and ion transport. Lineage-specific adaptations modulate these responses.
Figure 3.
Overview of bacterial stress response pathways. Core stress regulators (σ factors, IrrE/PprI, and two-component systems) control key protective functions, including proteostasis, redox/antioxidant defense, DNA repair, osmolyte synthesis, and ion transport. Lineage-specific adaptations modulate these responses.
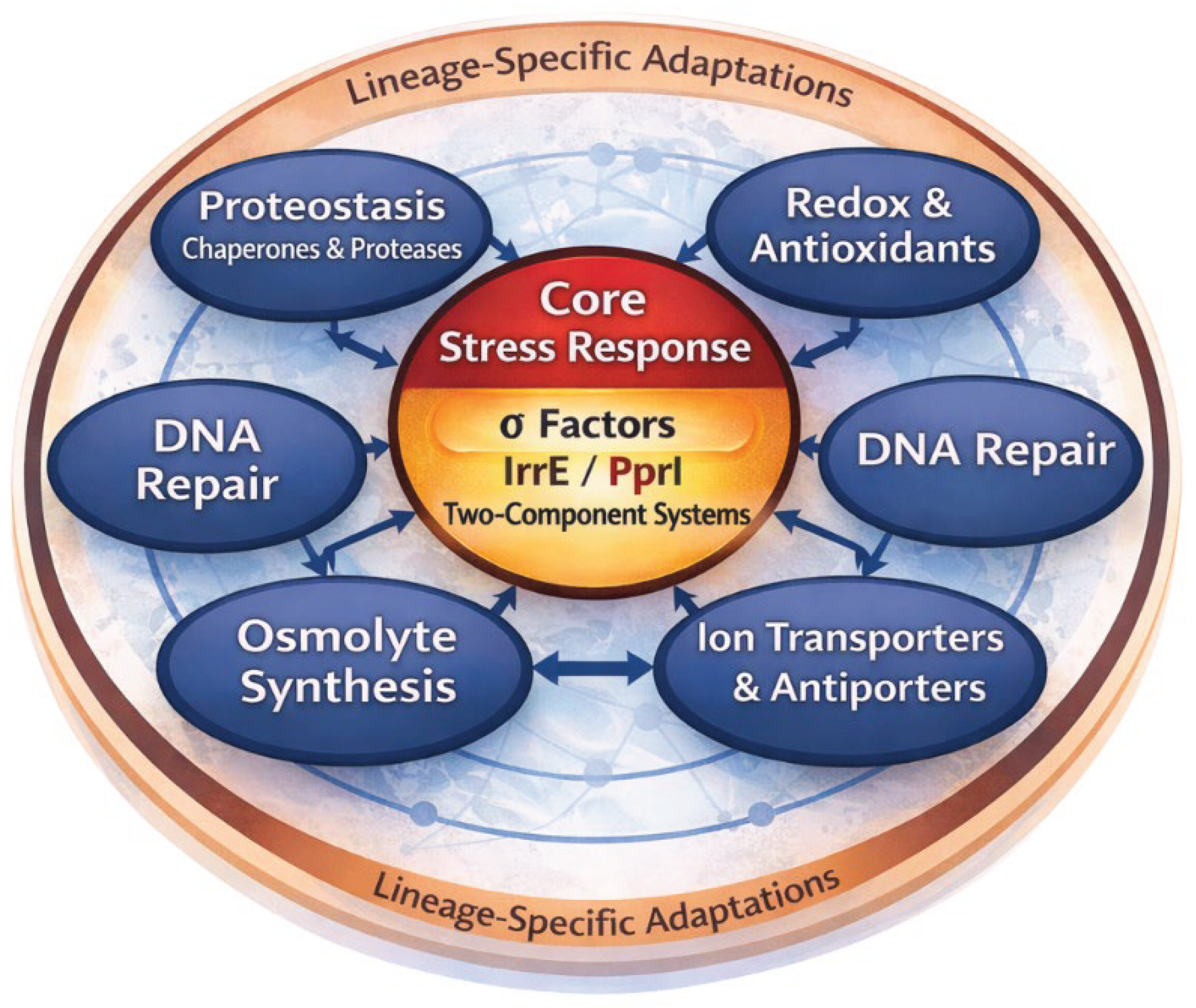
Finally, sophisticated regulatory networks tie these modules together, enabling a coordinated response to multiple stresses. Extremophilic bacteria have evolved stress-sensing two-component systems and alternative sigma factors that rapidly reprogram gene expression when conditions change [8]. For example, many Gram-negative extremophiles utilize the RpoS sigma factor or its analogs to induce a general stress response encompassing antioxidants, osmolyte transporters, and repair enzymes during nutrient starvation or osmotic shock. In Deinococcus, a unique regulatory protein (PprI/IrrE) serves as a master switch that, upon radiation damage, upregulates dozens of genes including recA (DNA repair), catalases, and other survival proteins [41]. Transcriptomic and proteomic profiling show that these global regulators activate overlapping sets of stress genes, illustrating cross-protection – a heat shock may trigger antioxidant defenses, or osmotic shock may induce DNA repair enzymes, preparing the cell for secondary stresses [40]. The integration of proteomics with functional genomics (e.g. mutant analyses) has been key to mapping these regulatory hierarchies, confirming that deletions of stress regulators abolish the coordinated induction of defense proteins. In sum, extremophiles manage survival not through isolated pathways but via interconnected networks, dynamically regulated to sense stresses and mount a rapid, multifaceted protective response.
6. Oxidative Stress as a Cross-Cutting Axis
Amid the diversity of extreme environments, oxidative stress emerges as a unifying threat that nearly all extremophiles must overcome. Many extreme conditions impose oxidative damage either directly or indirectly, making reactive oxygen species (ROS) detoxification a central axis of extremophile stress networks. High radiation, for example, causes radiolysis of water and generates a burst of ROS that can be more lethal than the initial DNA breaks; likewise, desiccation and rehydration cycles foster oxidative bursts that damage cellular components [8,41]. Even thermal extremes and pH extremes accelerate metabolic reactions that leak ROS or mobilize redox-active metals (as in acidic, iron-rich environments), heightening the risk of oxidative injury. Comparative studies underscore that when extremophilic cells are subjected to their respective stresses, a significant portion of the induced proteome consists of antioxidant and redox-maintenance proteins [40]. This indicates that no matter the primary stressor heat, salt, acid, cold, or radiation – the downstream challenge of ROS must be addressed for the cell to survive.
A hallmark of this cross-cutting oxidative stress axis is the ubiquitous presence of enzymatic antioxidants in extremophiles. Catalases and peroxidases, which decompose hydrogen peroxide into water and oxygen, are broadly expressed at high levels in extremophiles from hypersaline lakes to hydrothermal vents [40]. Superoxide dismutases (SODs) that convert superoxide radicals into safer molecules are similarly widespread; for instance, thermophilic bacteria often have multiple SOD isoforms or a thermally stable superoxide reductase to cope with elevated oxidative metabolism at high temperatures [29]. In acidophiles and metal-tolerant extremophiles, enzymes like cytochrome c peroxidases and glutathione peroxidases play key roles in neutralizing peroxide stress arising from Fenton reactions with heavy metals [27]. Radiation-resistant bacteria provide perhaps the clearest example: Deinococcus radiodurans constitutively expresses powerful antioxidant enzymes, and upon radiation exposure it further elevates proteins like catalase, peroxiredoxin, and DNA repair-linked helicases that specifically counter ROS-mediated damage [40]. The universality of these enzymatic defenses across extremophiles highlights that controlling ROS is a fundamental requirement, deeply embedded in their genetic and proteomic architecture (Table 3).
Figure 4.
Mechanisms of oxidative stress-induced damage and integrated cellular defence strategies in extremophiles.
Figure 4.
Mechanisms of oxidative stress-induced damage and integrated cellular defence strategies in extremophiles.
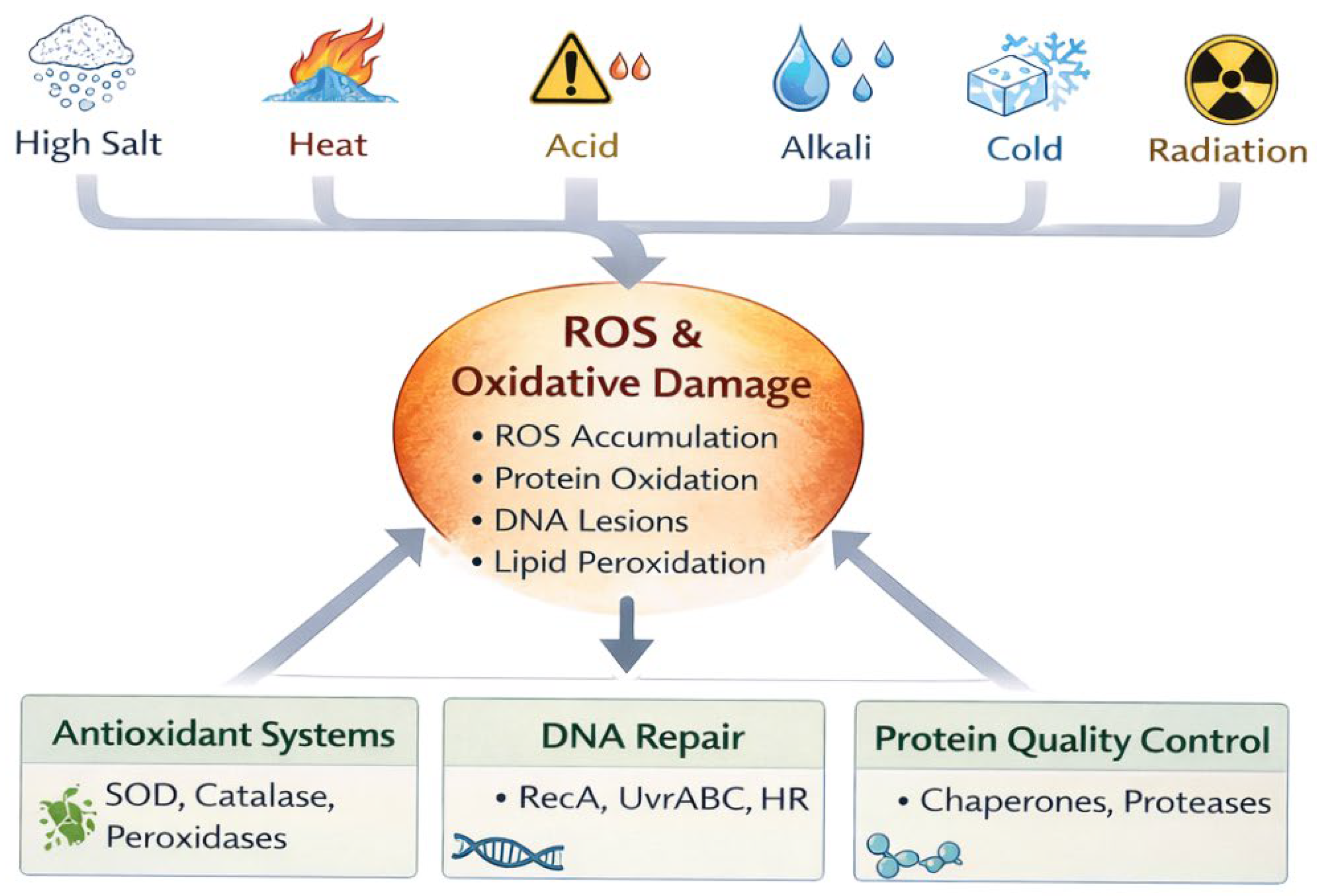
Besides enzymes, extremophiles also accumulate diverse small-molecule antioxidants and protective pigments as part of convergent evolution against ROS. Haloarchaea and many desert bacteria synthesize carotenoid pigments (e.g. bacterioruberin, deinoxanthin) that impart vivid orange or pink colors and efficiently scavenge free radicals and singlet oxygen. In fact, the haloarchaeal carotenoid bacterioruberin has extraordinary antioxidant potency and is thought to shield cells from both high solar UV and oxidative radicals in hypersaline pools [39]. Likewise, Antarctic psychrophiles and alpine microbes commonly produce carotenoids or flavonoid pigments, which have been correlated with survival under high UV/oxidative conditions in cold, thin-atmosphere environments. Some extremophiles accumulate compatible solutes that double as ROS scavengers: for instance, trehalose and mannitol can quench hydroxyl radicals and stabilize proteins against oxidative denaturation, aiding both osmotic balance and antioxidant defense [31]. Unusual metal-ion strategies also exist – D. radiodurans concentrates manganese ions and small peptides to form ROS-scavenging complexes that protect proteins from oxidative damage in lieu of a classical glutathione system [40]. Such chemical defenses often work in synergy with enzymes, creating multi-layered antioxidant fortifications. The prevalence of pigments and extremolytes across unrelated extremophiles is a testament to oxidative stress being a common denominator: evolution has repeatedly selected for molecules that absorb UV and neutralize radicals as life’s molecular sunscreen and antioxidant shield [52].
Crucially, the oxidative stress axis links together various stress-response pathways, explaining why extremophiles often exhibit cross-protection between different stresses. Exposure to one extreme can preemptively bolster ROS defenses, thereby increasing tolerance to other stresses that cause oxidative harm. For example, heat-shocked cells of a thermophilic bacterium show elevated catalase and SOD activity, which incidentally also improves their survival under subsequent exposure to peroxide or radiation [40]. In the halophilic archaeon Halobacterium, osmotic shock triggers the production of bacterioruberin and DNA repair enzymes, providing enhanced resistance to UV irradiation by mitigating oxidative DNA lesions [39]. These observations align with global transcriptome analyses where the regulons of heat shock, osmotic stress, and oxidative stress substantially overlap in extremophiles, all converging on a set of core protective genes. Master regulators like OxyR, PerR, or more specialized ones in extremophiles, sense redox imbalances and activate multiple defense genes, from antioxidants to chaperones and repair factors, in one concerted response. Thus, oxidative stress is not an isolated challenge but a central axis around which extremophiles have built a coordinated response network. By prioritizing ROS mitigation, extremophilic bacteria ensure that their other adaptive strategies be it protein folding, osmolyte accumulation, or DNA repair can operate in an intracellular environment not corrupted by oxidative damage. This integrated approach exemplifies how life at the limit’s copes with a universal hazard of reactive oxygen, turning a potential Achilles’ heel into a well-defended front [8,41,53].
7. Mechanistic Insights from Psychrophiles and Radiophiles
Extremely cold-adapted bacteria (psychrophiles) and radiation-tolerant bacteria (radiophiles) offer illuminating case studies that expand our understanding of stress-induced protein networks. These organisms thrive at the edges of temperature and radiation tolerance, respectively, and have evolved particularly novel solutions that complement the general extremophile strategies discussed above [19,28]. Integrative omics studies of psychrophiles and radiophiles highlight how fundamental modules chaperones, antioxidants, DNA repair are customized and amplified to meet their unusual challenges, yielding insights applicable across all extremophiles. While not exhaustive reviews of each field, the following contrasts emphasize key mechanisms by which psychrophilic and radiophilic bacteria push the boundaries of stress adaptation [19]. Metagenome-resolved analyses extend these insights to uncultured lineages and generate testable hypotheses about the contribution of chromosome-organization proteins (including NAPs) to stress adaptation, which can be examined using targeted genetics where tractable [30].
Psychrophile Adaptations (Low-Temperature Extremophiles): Psychrophilic bacteria flourish in permanently cold environments such as polar oceans, high-altitude clouds, or permafrost soils, where temperatures remain at 0–5 °C or below. Cold terrestrial matrices add oligotrophy and freeze–thaw cycles that shape stress-network deployment in soil psychrophiles [45]. A primary challenge at such low temperatures is preserving biochemical reaction rates and membrane fluidity in the face of sluggish kinetics and rigidified structures. Psychrophiles have converged on several strategies to mitigate these effects [54]. First, their proteins tend to be more flexible and unstable at moderate temperatures, an adaptation that ensures sufficient conformational mobility and enzymatic activity in the cold. Comparative proteomic analyses show that psychrophiles often express specific “cold shock” proteins (CSPs) and RNA chaperones that assist in transcription/translation at low temperature by preventing mRNA secondary structure formation. For example, after an abrupt drop in temperature, group I cold shock proteins like CspA in many bacteria are massively induced to help the cell acclimate, whereas group II proteins (including certain helicases and nucleases) sustain longer-term adaptation [31]. Alongside CSPs, psychrophiles deploy classic chaperones (e.g. GroEL/ES, DnaK) which, intriguingly, are required not just for heat stress but also to fold proteins at low temperatures where folding can stall in kinetically trapped states. This overlap in chaperone usage underlies a broader principle: maintaining proteostasis is as critical in extreme cold as in extreme heat, and many heat-shock proteins are also cold-essential when appropriately tuned [29].
Figure 5.
Systems Biology Framework for the Engineering of Robust Strains and Enzymes via Multi-Omics Integration.
Figure 5.
Systems Biology Framework for the Engineering of Robust Strains and Enzymes via Multi-Omics Integration.
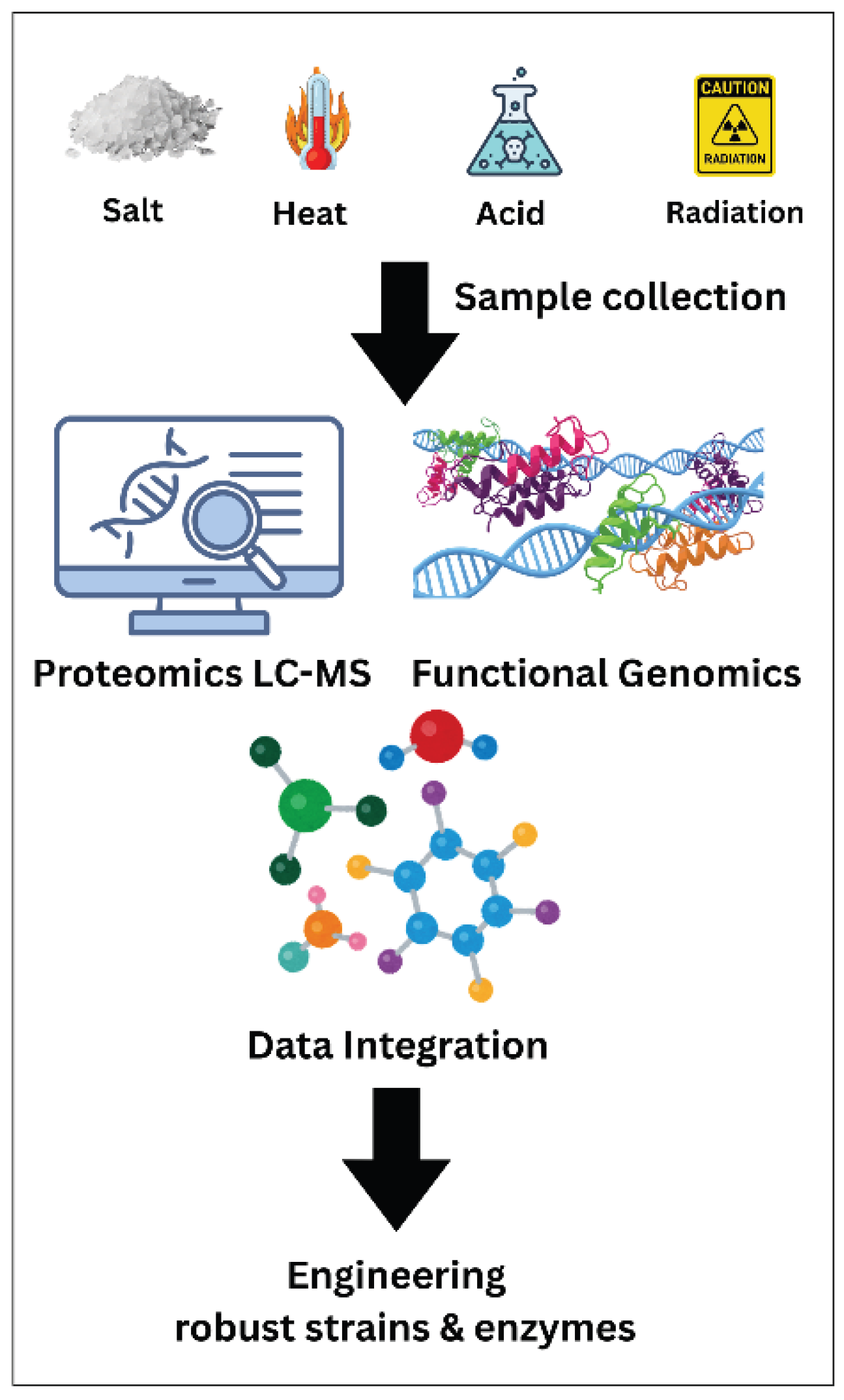
Psychrophiles also adjust their cellular chemistry to function in the cold. Their cell membranes are enriched in polyunsaturated and branched fatty acids, which remain fluid at low temperatures, preventing the membrane from freezing into a gel-like state [27]. They often accumulate cryoprotectant osmolytes for instance, Exiguobacterium and Arthrobacter isolates from Antarctic soils build up trehalose, glycine betaine, and glycerol, which lower the freezing point and protect proteins from cold-denaturation. Proteomic studies confirm that enzymes for synthesizing or importing these cryoprotectants are upregulated during cold acclimation, and mutants deficient in cryoprotectant production frequently show impaired freeze-thaw survival. Another striking feature is the frequent production of pigments and extracellular polymers: many psychrophiles are vividly pigmented (yellow, orange, red), a trait linked to carotenoid antioxidants that guard against the oxidative stress of high oxygen solubility and UV exposure in snow or ice [31]. These carotenoids, as noted, can protect cells from UV-induced ROS, suggesting an adaptation to cold environments that are often also high-UV (thin atmosphere at poles) – thus psychrophiles illustrate how multiple stressors (cold and radiation) can elicit an integrated response [28]. Genomic and functional studies reinforce that psychrophiles have expanded repertoires of oxidative stress genes: for example, Antarctic Pseudoalteromonas and Colwellia species encode extra catalase and peroxidase enzymes, which are produced at low temperatures to counteract ROS generated during sporadic exposure to light or when metabolic activity spikes during rare warm periods [27,55].
Radiophile Adaptations (Radiation-Resistant Extremophiles)
Radiophilic bacteria such as Deinococcus radiodurans have the astonishing capacity to survive and recover from doses of ionizing radiation (gamma or UV) that shatter genomes and generate acute oxidative stress [8,41,56]. This level of tolerance represents a pinnacle of stress network efficacy, and radiophile mechanisms have become a reference point for understanding cellular defenses against DNA damage and oxidative assault. A defining feature of D. radiodurans and its radiotolerant relatives is an extraordinarily effective DNA repair system [57]. These bacteria can reassemble a genome fragmented by dozens of double-strand breaks, thanks to a high copy number of the genome (providing multiple templates) and a very efficient homologous recombination machinery. Proteomic and transcriptomic analyses after irradiation show rapid and massive induction of DNA repair proteins, including RecA and numerous end-processing enzymes, along with unique DNA repair factors like PprA that help stabilize broken DNA ends. Within hours of exposure, Deinococcus cells execute an orchestrated “radiation recovery” program termed the DNA damage response (or “RDR” akin to a radiation-induced SOS response), which is regulated by the irrE (pprI) gene product acting as a global transcriptional activator. Mutational studies confirm that D. radiodurans lacking irrE cannot properly induce its repair and antioxidant genes and becomes radiation-sensitive, highlighting how a single regulator coordinates a network of survival proteins. In essence, radiophiles have amplified the DNA repair module common to many bacteria into a hyperactive, tightly regulated system that can reverse extreme genomic damage [8,41].
Equally important, radiophiles fortify their cells against the collateral oxidative damage that accompanies radiation. Ionizing radiation in aqueous cells produces a storm of ROS (especially hydroxyl radicals) that can oxidize proteins and lipids indiscriminately. D. radiodurans preempts this by constitutively accumulating large reserves of antioxidants and by maintaining a cytoplasm that favors protective reactions. Proteomic profiling has revealed that unstressed D. radiodurans already expresses high baseline levels of oxidative stress proteins, such as robust catalases, peroxiredoxins, and Mn-dependent superoxide scavengers, compared to radiation-sensitive bacteria [38]. Upon radiation exposure, these antioxidant enzymes are further upregulated: for example, Sudharsan et al. [40] showed that a 4 kGy gamma dose triggered increased expression of catalase, an OsmC peroxidase, and other oxidoreductases, which collectively mitigate the burst of peroxides and radicals generated by radiation. Remarkably, D. radiodurans also uses non-enzymatic strategies—its cytosol contains millimolar levels of manganese complexes with metabolites (orthophosphate, peptides) that act as radical scavengers to protect proteins from oxidative inactivation [8,41]. This metabolic innovation means that even if some macromolecules are damaged by radiation, critical enzymes are preserved in an active state, ready to restore cell function post-irradiation.
8. Conclusions and Future Perspectives
Extremophilic bacteria have emerged as model systems for understanding how life can robustly adapt to and thrive under conditions once thought prohibitive for cellular function. By combining high-throughput proteomics and transcriptomics (expression profiling) to capture network state with perturbation-based functional genomics to test causality, researchers are beginning to resolve how extremophiles assemble stress-responsive protein networks that enable survival under otherwise lethal conditions. The comparative approach taken in this review underscores that, despite the superficially disparate challenges of salt, heat, acid, cold, or radiation, there are unifying themes in extremophile survival. All rely on a set of core protective functions protein folding maintenance, oxidative damage control, DNA repair, and metabolic homeostasis but each extremophile lineage has tuned and augmented these functions to suit its niche. By examining proteomic expression patterns side-by-side with genomic content, researchers can link specific molecular adaptations (such as a novel chaperone or DNA repair enzyme) to organismal stress tolerance, and then validate their roles via gene knockout or heterologous expression. This integrative paradigm has already led to the discovery of new stress resilience factors (for example, unique cryoprotectant enzymes and radical-scavenging metabolites) and revealed how canonical pathways (like the heat shock response or SOS response) are repurposed in extreme contexts. Looking ahead, tighter integration of proteomics and transcriptomics (expression profiling) with comparative genomics and perturbation-based functional genomics should enable more causal, systems-level models of extremophile stress tolerance and accelerate translation to strain and enzyme engineering.
Despite significant advances, many questions remain about the intricacies of stress-induced protein networks in extremophiles. One major challenge is to map the higher-order regulation and cross-talk between different stress responses. While key regulators (e.g. RpoS, IrrE, HspR) have been identified in various systems, the full circuitry of signal transduction that allows a cell to sense an extreme condition and proportionally adjust dozens of pathways is not yet fully delineated. Future research will likely leverage systems biology approaches, using time-resolved proteomics and phosphoproteomics to capture how regulatory networks dynamically turn proteins on or off during stress acclimation. Another frontier is understanding the structural biology of extremophile proteins: high-throughput proteomics tells us what changes, but detailed structural and biophysical studies are needed to explain how certain proteins remain functional under extreme conditions. For instance, what specific amino acid changes confer salt tolerance to enzymes, or how do DNA repair complexes in radiophiles withstand massive damage loads? Answering these questions will benefit from combining genomics (to identify genetic determinants), proteomics (to see expression and modifications), and experimental evolution or protein engineering (to test causality of specific adaptations). In parallel, the role of cellular context the contribution of membranes, cytosolic solute milieu, and compartmentalization is an area ripe for exploration via metabolomics and imaging, complementing the proteogenomic data to complete the picture of extremophile cell physiology.
The study of extremophilic stress networks is not only of fundamental interest but also holds promise for practical applications. As we decode how extremophiles protect and repair their biomolecules, we unlock potential tools for biotechnology and medicine. Extremophile-derived chaperones, osmolytes, and antioxidants are already being investigated for stabilizing industrial enzymes or therapeutic proteins against stress. DNA repair enzymes from hyperthermophiles and radiophiles may inform new strategies for improving radiation therapies or developing robust biosensors that operate in harsh environments. Moreover, understanding extremophiles broadens our perspective in astrobiology – it defines biochemical and proteomic benchmarks for life under Martian-level radiation or Europan ocean pressure, guiding the search for life beyond Earth. Future research, therefore, will likely see interdisciplinary efforts that not only delve into extremophile biology with finer resolution but also transpose those insights into engineered systems. For instance, one can envision synthetic biology projects that implant extremophile stress networks into model organisms or cell-free systems to confer resilience, effectively “rewiring” tolerance to heat, desiccation, or oxidative stress using nature’s blueprints. Achieving this will require the comprehensive catalogs of genes and proteins that integrative omics is providing, as well as an intimate functional understanding of each component.
Author Contributions
Conceptualization, H.V.P.; Methodology, H.V.P.; Resources (literature collection), H.V.P.; Data curation (reference organization), H.V.P.; Writing - original draft preparation, H.V.P.; Visualization, V.P.; Writing - review and editing, M.C., H.V.P., and V.P.; Supervision, M.C. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rubiano-Labrador, C.; Bland, C.; Miotello, G.; Armengaud, J.; Baena, S. Salt Stress Induced Changes in the Exoproteome of the Halotolerant Bacterium Tistlia consotensis Deciphered by Proteogenomics. PloS one 2015, 10, e0135065. [Google Scholar] [CrossRef]
- Remonsellez, F.; Castro-Severyn, J.; Pardo-Esté, C.; Aguilar, P.; Fortt, J.; Salinas, C.; Barahona, S.; León, J.; Fuentes, B.; Areche, C.; Hernández, K. L.; Aguayo, D.; Saavedra, C. P. Characterization and Salt Response in Recurrent Halotolerant Exiguobacterium sp. SH31 Isolated From Sediments of Salar de Huasco, Chilean Altiplano. Frontiers in microbiology 2018, 9, 2228. [Google Scholar] [CrossRef]
- Purohit, H. V. Nucleoid-associated proteins: molecular mechanisms in microbial adaptation. World journal of microbiology & biotechnology 2025, 41, 277. [Google Scholar] [CrossRef]
- Valdez-Nuñez, L. F.; Kappler, A.; Ayala-Muñoz, D.; Chávez, I. J.; Mansor, M. Acidophilic sulphate-reducing bacteria: Diversity, ecophysiology, and applications. Environmental microbiology reports 2024, 16, e70019. [Google Scholar] [CrossRef]
- Matarredona, L.; Zafrilla, B.; Camacho, M.; Bonete, M. J.; Esclapez, J. Understanding the tolerance of halophilic archaea to stress landscapes. Environmental microbiology reports 2024, 16, e70039. [Google Scholar] [CrossRef]
- Altinisik Kaya, F. E.; Avci, F. G.; Sayar, N. A.; Kazan, D.; Sayar, A. A.; Sariyar Akbulut, B. What Are the Multi-Omics Mechanisms for Adaptation by Microorganisms to High Alkalinity? A Transcriptomic and Proteomic Study of a Bacillus Strain with Industrial Potential. Omics: a journal of integrative biology 2018, 22, 717–732. [Google Scholar] [CrossRef] [PubMed]
- Atakav, Y.; Pinar, O.; Kazan, D. Investigation of the Physiology of the Obligate Alkaliphilic Bacillus marmarensis GMBE 72T Considering Its Alkaline Adaptation Mechanism for Poly(3-hydroxybutyrate) Synthesis. Microorganisms 2021, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, F.; Chen, H.; Yang, Z.; Ning, Y.; Chang, C.; Yang, D. New Insights into Radio-Resistance Mechanism Revealed by (Phospho)Proteome Analysis of Deinococcus Radiodurans after Heavy Ion Irradiation. International journal of molecular sciences 2023, 24, 14817. [Google Scholar] [CrossRef]
- Han, J. M.; Mwiti, G.; Yeom, S. J.; Lim, J.; Kim, W. S.; Lim, S.; Lim, S. T.; Byun, E. B. Radiation-Resistant Bacteria Deinococcus radiodurans-Derived Extracellular Vesicles as Potential Radioprotectors. Advanced healthcare materials 2025, 14, e2403192. [Google Scholar] [CrossRef]
- Fongaro, G.; Maia, G. A.; Rogovski, P.; Cadamuro, R. D.; Lopes, J. C.; Moreira, R. S.; Camargo, A. F.; Scapini, T.; Stefanski, F. S.; Bonatto, C.; Marques Souza, D. S.; Stoco, P. H.; Duarte, R. T. D.; Cabral da Cruz, A. C.; Wagner, G.; Treichel, H. Extremophile Microbial Communities and Enzymes for Bioenergetic Application Based on Multi-Omics Tools. Current genomics 2020, 21, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Nie, T.; Wang, L.; Liu, Y.; Fu, S.; Wang, J.; Cui, K.; Wang, L. A Halophilic Bacterium for Bioremediation of Saline-Alkali Land: The Triadic and Synergetic Response Mechanism of Oceanobacillus picturae DY09 to Salt Stress. Microorganisms 2025, 13, 1474. [Google Scholar] [CrossRef]
- Kung, C.; Martinac, B.; Sukharev, S. Mechanosensitive channels in microbes. Annual review of microbiology 2010, 64, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Belin, B. J.; Busset, N.; Giraud, E.; Molinaro, A.; Silipo, A.; Newman, D. K. Hopanoid lipids: from membranes to plant-bacteria interactions. Nature reviews. Microbiology 2018, 16, 304–315. [Google Scholar] [CrossRef]
- Yin, L.; Xue, Y.; Ma, Y. Global Microarray Analysis of Alkaliphilic Halotolerant Bacterium Bacillus sp. N16-5 Salt Stress Adaptation. PloS one 2015, 10, e0128649. [Google Scholar] [CrossRef]
- Vandrich, J.; Pfeiffer, F.; Alfaro-Espinoza, G.; Kunte, H. J. Contribution of mechanosensitive channels to osmoadaptation and ectoine excretion in Halomonas elongata. Extremophiles: life under extreme conditions 2020, 24, 421–432. [Google Scholar] [CrossRef]
- Oren, A. Microbial life at high salt concentrations: phylogenetic and metabolic diversity. Saline systems 2008, 4, 2. [Google Scholar] [CrossRef]
- Enuh, B. M.; Aytar Çelik, P. Genome Analysis of Halomonas elongata Strain 153B and Insights Into Polyhydroxyalkanoate Synthesis and Adaptive Mechanisms to High Saline Environments. Current microbiology 2022, 80, 18. [Google Scholar] [CrossRef]
- Dildar, T.; Cui, W.; Ikhwanuddin, M.; amp; Ma, H. Aquatic Organisms in Response to Salinity Stress: Ecological Impacts, Adaptive Mechanisms, and Resilience Strategies. Biology 2025, 14, 667. [Google Scholar] [CrossRef]
- Chan, Y. F.; Chen, C. Y.; Lu, C. Y.; Tu, Y. C.; Tandon, K.; Shikina, S.; Tang, S. L. A first insight into the heat-induced changes in proteomic profiles of the coral symbiotic bacterium Endozoicomonas montiporae. Frontiers in Marine Science 2022, 9, 808132. [Google Scholar] [CrossRef]
- Sato, Y.; Okano, K.; Honda, K. Effects of small heat shock proteins from thermotolerant bacteria on the stress resistance of Escherichia coli to temperature, pH, and hyperosmolarity. Extremophiles: life under extreme conditions 2024, 28, 12. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Fiallo, K.; Muñoz-Villagrán, C.; Orellana, O.; Sjoberg, R.; Levicán, G. Comparative genomics of the proteostasis network in extreme acidophiles. PloS one 2023, 18, e0291164. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Jiao, L.; Xu, J.; Zhang, J.; Qi, Y.; Qiu, M.; Wei, X.; Fan, M. Integrated transcriptomic and proteomic analysis reveals the response mechanisms of Alicyclobacillus acidoterrestris to heat stress. Food research international (Ottawa, Ont.) 2022, 151, 110859. [Google Scholar] [CrossRef]
- Shih, T. W.; Pan, T. M. Stress responses of thermophilic Geobacillus sp. NTU 03 caused by heat and heat-induced stress. Microbiological research 2011, 166, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Ranawat, P.; amp; Rawat, S. Stress response physiology of thermophiles. Archives of microbiology 2017, 199, 391–414. [Google Scholar] [CrossRef]
- Belnap, C. P.; Pan, C.; VerBerkmoes, N. C.; Power, M. E.; Samatova, N. F.; Carver, R. L.; Hettich, R. L.; Banfield, J. F. Cultivation and quantitative proteomic analyses of acidophilic microbial communities. The ISME journal 2010, 4, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Cortez, D.; Neira, G.; González, C.; Vergara, E.; Holmes, D. S. A Large-Scale Genome-Based Survey of Acidophilic Bacteria Suggests That Genome Streamlining Is an Adaption for Life at Low pH. Frontiers in microbiology 2022, 13, 803241. [Google Scholar] [CrossRef]
- Somayaji, A.; Dhanjal, C. R.; Lingamsetty, R.; Vinayagam, R.; Selvaraj, R.; Varadavenkatesan, T.; Govarthanan, M. An insight into the mechanisms of homeostasis in extremophiles. Microbiological research 2022, 263, 127115. [Google Scholar] [CrossRef]
- Huanca-Juarez, J.; Nascimento-Silva, E. A.; Silva, N. H.; Silva-Rocha, R.; Guazzaroni, M. E. Identification and functional analysis of novel protein-encoding sequences related to stress-resistance. Frontiers in microbiology 2023, 14, 1268315. [Google Scholar] [CrossRef]
- Marzban, G.; Tesei, D. The Extremophiles: Adaptation Mechanisms and Biotechnological Applications. Biology 2025, 14, 412. [Google Scholar] [CrossRef]
- Purohit, H. V.; Chakraborty, J. Metagenomic approaches for studying ubiquitous yet diverse nucleoid associated proteins in microbial communities: challenges and advances. World journal of microbiology & biotechnology 2025, 41, 383. [Google Scholar] [CrossRef]
- Baraúna, R. A.; Freitas, D. Y.; Pinheiro, J. C.; Folador, A. R.; Silva, A. A Proteomic Perspective on the Bacterial Adaptation to Cold: Integrating OMICs Data of the Psychrotrophic Bacterium Exiguobacterium antarcticum B7. Proteomes 2017, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Boas Lichty, K. E.; Gregory, G. J.; Boyd, E. F. NhaR, LeuO, and H-NS are part of an expanded regulatory network for ectoine biosynthesis expression. Applied and Environmental Microbiology 2023, 89, e00479-23. [Google Scholar] [CrossRef]
- García-Descalzo, L.; García-López, E.; Cid, C. Comparative proteomic analysis of psychrophilic vs. mesophilic bacterial species reveals different strategies to achieve temperature adaptation. Frontiers in Microbiology 2022, 13, 841359. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends in microbiology 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T. A. Alkaline pH homeostasis in bacteria: new insights. Biochimica et biophysica acta (BBA)-biomembranes 2005, 1717, 67–88. [Google Scholar] [CrossRef]
- Kumar, S.; Suyal, D. C.; Yadav, A.; Shouche, Y.; Goel, R. Psychrophilic Pseudomonas helmanticensis proteome under simulated cold stress. Cell Stress and Chaperones 2020, 25, 1025–1032. [Google Scholar] [CrossRef]
- Qi, H. Z.; Wang, W. Z.; He, J. Y.; Ma, Y.; Xiao, F. Z.; He, S. Y. Antioxidative system of Deinococcus radiodurans. Research in microbiology 2020, 171, 45–54. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S. Y.; Alwasel, S. H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Archives of toxicology 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Martínez-Espinosa, R. M. Halocins and C50 Carotenoids from Haloarchaea: Potential Natural Tools against Cancer. Marine drugs 2024, 22, 448. [Google Scholar] [CrossRef] [PubMed]
- M, S.; R. P., N; Chakraborty, A.; Rajendrasozhan, S. Proteomic profiling of Deinococcus radiodurans with response to thioredoxin reductase inhibitor and ionizing radiation treatment. Journal of proteomics 2022, 267, 104697. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, N.; Zhang, Y. The radioresistant and survival mechanisms of Deinococcus radiodurans. Radiation Medicine and Protection 2023, 4, 70–79. [Google Scholar] [CrossRef]
- Orban, K.; Finkel, S. E. Dps Is a Universally Conserved Dual-Action DNA-Binding and Ferritin Protein. Journal of bacteriology 2022, 204, e0003622. [Google Scholar] [CrossRef]
- Vauclare, P.; Wulffelé, J.; Lacroix, F.; Servant, P.; Confalonieri, F.; Kleman, J. P.; Bourgeois, D.; Timmins, J. Stress-induced nucleoid remodeling in Deinococcus radiodurans is associated with major changes in Heat Unstable (HU) protein dynamics. Nucleic acids research 2024, 52, 6406–6423. [Google Scholar] [CrossRef]
- Ge, P.; Rashid, F. M.; Dame, R. T. The role of nucleoid-associated proteins in mediating responses to environmental changes. Current opinion in microbiology 2025, 87, 102628. [Google Scholar] [CrossRef] [PubMed]
- Purohit, HV; Kanojia, H; Pandya, V; Nalla, Y; Raval, KY; Kapadiya, KM; Kamdar, JH. Soil as a host to the biotic community. In Soil Microbiome of the cold habitats: trends and applications; Gupta, P, Shahnawaz, M, Eds.; CRC, 2023; pp. pp 17–30. [Google Scholar] [CrossRef]
- Mogk, A.; Tomoyasu, T.; Goloubinoff, P.; Rüdiger, S.; Röder, D.; Langen, H.; Bukau, B. Identification of thermolabile Escherichia coli proteins: prevention and reversion of aggregation by DnaK and ClpB. The EMBO journal 1999. [Google Scholar] [CrossRef] [PubMed]
- Figaj, D. The role of heat shock protein (Hsp) chaperones in environmental stress adaptation and virulence of plant pathogenic bacteria. International Journal of Molecular Sciences 2025, 26, 528. [Google Scholar] [CrossRef]
- Deng, M.; Xu, X.; Yang, Z.; Lu, K. CSP15 attenuates chlorpyrifos toxicity through sequestration and bioactivation suppression in Nilaparvata lugens. Insect Biochemistry and Molecular Biology 2025, 104453. [Google Scholar] [CrossRef]
- Lu, H.; Hua, Y. PprI: the key protein in response to DNA damage in Deinococcus. Frontiers in Cell and Developmental Biology 2021, 8, 609714. [Google Scholar] [CrossRef]
- Fatima, K.; Naqvi, F.; amp; Younas, H. A review: Molecular chaperone-mediated folding,unfolding and disaggregation of expressed recombinant proteins. Cell Biochemistry and Biophysics 2021, 79, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Weinisch, L.; Kühner, S.; Roth, R.; Grimm, M.; Roth, T.; Netz, D. J.; Filker, S. Identification of osmoadaptive strategies in the halophile, heterotrophic ciliate Schmidingerothrix salinarum. PLoS Biology 2018, 16, e2003892. [Google Scholar] [CrossRef]
- Silva, R.; Gonçalves, T.; Morone, J.; Moreira, G. A.; Morais, J.; Hentschke, G. S.; Lopes, G.; amp. Pigments profile and antioxidant potential of extremophile cyanobacteria isolated from the Mexican Volcanic Lake Chichonal. Algal Research 2024, 81, 103578. [Google Scholar] [CrossRef]
- Kanekar, P.P.; Kanekar, S.P. Radiophilic, Radioresistant, and Radiotolerant Microorganisms. In Diversity and Biotechnology of Extremophilic Microorganisms from India. Microorganisms for Sustainability; Springer: Singapore, 2022. [Google Scholar] [CrossRef]
- Yusof, N. A.; Hashim, N. H. F.; Bharudin, I. Cold Adaptation Strategies and the Potential of Psychrophilic Enzymes from the Antarctic Yeast, Glaciozyma antarctica PI12. Journal of fungi (Basel, Switzerland) 2021, 7, 528. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Min, J.; Shin, Y.; Park, W. Morphological and physiological adaptations of psychrophilic Pseudarthrobacter psychrotolerans YJ56 under temperature stress. Scientific reports 2023, 13, 14970. [Google Scholar] [CrossRef]
- Basu, B. The radiophiles of Deinococcaceae family: Resourceful microbes for innovative biotechnological applications. Current research in microbial sciences 2022, 3, 100153. [Google Scholar] [CrossRef]
- Minton, K. W. DNA repair in the extremely radioresistant bacterium Deinococcus radiodurans. Molecular microbiology 1994, 13, 9–15. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Major Extremophilic Groups, Representative Species, and Primary Stressors.
| Extremophile type | Typical conditions | Representative species | Primary stressors / notes | References |
| Halophiles | High salt (≈2–5 M NaCl) | Halobacterium salinarum, Halomonas spp. | Ionic/osmotic stress; reliance on compatible solutes and specialized Na⁺ transport. | [32] |
| Thermophiles / Hyperthermophiles | 60–120 °C | Thermus thermophilus, Pyrococcus spp. | Protein thermostability, membrane adaptations, heat-shock systems and ATP-dependent proteases. | [33] |
| Acidophiles | pH ~0–3 | Acidithiobacillus ferrooxidans, Ferroplasma | Proton toxicity, metal stress; low-permeability membranes and proton export systems. | [34] |
| Alkaliphiles | pH ≈9–12 | Bacillus halodurans, Halomonas alkaliphila | Proton scarcity → Na⁺-based bioenergetics and multi-subunit antiporters (Mrp/Mnh). | [35] |
| Psychrophiles | ~−5 to 10 °C | Psychromonas, Antarctic Pseudomonas spp. | Cold-adapted enzymes, RNA chaperones (Csps), membrane fluidity and cryoprotectants. | [36] |
| Radiophiles / Desiccation-resistant | High ionizing radiation / desiccation | Deinococcus radiodurans | Exceptional DNA-repair systems, PprI/DdrO regulatory circuit, strong antioxidant defenses. | [37] |
Table 2.
Integrated Proteomics and Functional Genomics Findings Across Extremophiles.
| Extremophile group | Proteomics signature (selected) | Functional genomics / genetic evidence | Takeaway (essential network components) | References |
| Halophiles | Strong induction of ectoine pathway enzymes; Na⁺ transporters | Knockouts or regulatory mutants in ectoine pathways or antiporters alter salt tolerance and ectoine production. | Compatible-solute synthesis + Na⁺ antiporters tightly coupled to proteostasis and metabolism. | [32] |
| Thermophiles | High levels of HSPs (DnaK/GroEL) and ATP-dependent proteases | Deletion or depletion of groESL, clpB, or lon reduces thermotolerance. | Proteostasis network (folding + targeted proteolysis) is essential for high-T survival. | [46,47] |
| Acidophiles | Upregulated proton exporters and low-permeability membrane components | Mutational studies show loss of pH tolerance with impaired proton export / membrane integrity genes. | Proton homeostasis + iron handling + ROS defenses shape acid tolerance. | [34] |
| Alkaliphiles | Elevated Na⁺-coupled ATP synthase subunits and Mrp/Mnh antiporters | Genetic disruption of multi-subunit antiporters compromises growth at high pH. | Na⁺-bioenergetics + antiport complexes are core to alkaline adaptation. | [35] |
| Psychrophiles | Upregulation of Csps and flexible metabolic enzymes | csp mutants show impaired cold growth; proteomics shows increased chaperones & desaturases. | RNA chaperones + membrane fluidity adjustments are central cold strategies. | [36,48] |
| Radiophiles | Enriched DNA-repair proteins & antioxidants | PprI / DdrO and RecA systems are essential; mutants show radiation sensitivity. | Robust DNA repair + antioxidant networks underpin extreme radio/desiccation resistance. | [49] |
Table 3.
Core Stress-Induced Protein Classes Identified in Proteomic Studies.
| Protein class / module | Functional role | Extremophiles with strong proteomic induction | Typical examples (proteins) | References |
| Molecular chaperones | Prevent protein misfolding & refold damaged proteins | Thermophiles, halophiles, psychrophiles | DnaK (Hsp70), GroEL/GroES, small HSPs. | [33,50] |
| ATP-dependent proteases / proteolysis | Remove irreversibly damaged proteins; proteostasis | Thermophiles, radiophiles | Lon, ClpP/ClpX, FtsH. | [46] |
| Compatible solute synthesis & uptake | Osmotic balance / protein stabilisation | Halophiles, some psychrophiles | Ectoine biosynthetic enzymes, glycine-betaine transporters. | [32] |
| Ion transporters / antiporters | pH & ionic homeostasis | Halophiles, alkaliphiles, acidophiles | Na⁺/H⁺ antiporters (Mrp/Mnh), K⁺ transporters. | [51] |
| DNA-repair & genome maintenance | Repair double-strand breaks, base damage | Radiophiles, thermophiles | RecA, UvrABC, PprA / PprI regulatory elements. | [49] |
| Antioxidants & redox enzymes | Detoxify ROS; maintain redox balance | Radiophiles, acidophiles | Superoxide dismutase (SOD), catalase, peroxiredoxins. | [37] |
| Membrane remodeling enzymes | Preserve membrane integrity/fluidity | Thermophiles, psychrophiles, alkaliphiles | Fatty-acid desaturases, cardiolipin synthases, hopanoid biosynthesis. | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



