Submitted:
19 February 2026
Posted:
27 February 2026
You are already at the latest version
Abstract
The invasive yellow-legged hornet Vespa velutina nigrithorax has spread across Europe following its accidental introduction into France in 2004. This species adversely affects biodiversity, apiculture, pomiculture and viticulture, and human health. Cur-rent management relies predominantly on nest destruction; however, manual removal is often logistically challenging and costly because nests are typically located high in trees (up to 30 m), frequently necessitating vehicle-mounted lifts. Ground-based ap-plication of biocides using long injection lances is comparatively rapid and inexpensive, but in many countries insecticides are not permitted because products are not specifi-cally authorized for hornet control. Consequently, alternative approaches are needed. Here, we evaluated the efficacy of activated charcoal for nest destruction in V. v. nig-rithorax. We injected 145 nests with 50–100 g of activated charcoal and subsequently destroyed the nests. One week later, we assessed worker survival and the establish-ment of new nests. Emergency nest construction by surviving workers was observed in 3 of 145 cases (2.1 %). This rate was comparable to that observed following insecticide treatment (2 of 136 cases; 1.5 %). Activated charcoal therefore appears to be similarly effective to insecticide-based control while offering advantages in terms of environ-mental compatibility, user safety, ease of handling, and legal applicability in Europe. Activated charcoal may represent a practical alternative to manual nest removal and unauthorized insecticide use.
Keywords:
invasive species
; biodiversity
; pest management
; allergies
; biocide
; bee keeping
; hornets
1. Introduction
Invasive species are recognized by IPBES as a major driver of biodiversity loss [1]. The yellow-legged hornet (Vespa velutina nigrithorax) is among the most rapidly expanding invasive hymenopterans of the past two decades and poses a particular challenge due to its high spread rate and impacts on biodiversity, human health, and the economy [2]. Following introduction to South Korea (2003) [3,4] and the first record in Southern Europe/France (2004), the species spread rapidly across Europe [5]. By 2025, occurrences have been documented in Asia, Europe, and North America [6]; additional records have been reported for North Africa (Algiers, Algeria) [7,8] and for Auckland, NZ [9]. Thus, the species has been recorded on four continents and in New Zealand.
V. velutina rapidly establishes large secondary nests, preys intensively on honey bees, and can locally impair pollination services and beekeeping. In Spain, approximately 5.1–20.5% of apiary revenue is spent on controlling V. velutina [10]. Under a scenario of maximal spread in France, annual losses of up to €30.8 million due to colony mortality alone have been projected [11]. Reported impacts also include damage in viticulture [12] and insufficient pollination associated with hornet presence [13]. For Germany, annual management costs under a maximal spread scenario have been modelled at ~€5 million, compared with ~€11.9 million in France [14], where projected beekeeping-related losses are substantially higher [15]. These estimates do not include potential damages to agriculture (wine and fruit production) or public-health and broader economic costs associated with stings [16]. Moreover, ecological impacts are likely considerable, although monetizing biodiversity remains challenging [17]. In Europe, management accounts for only about 6% of total costs attributed to invasive organisms, implying that prevented damages can greatly exceed control expenditures [18].
In Germany and parts of the EU, operational control is constrained by conservation and biocidal-product regulations. Trapping systems, including electric “harps” [19,20,21,22], are legally problematic because the capture of protected non-target species (e.g., Vespa crabro) — even as bycatch — is not permitted under nature conservation law. Given the repeatedly documented lack of selectivity of passive traps [23,24,25,26] and their limited contribution to sustained population suppression [27], their use is generally not recommended. Defensive measures at apiaries (e.g., entrance guards, nets [2,28]) may reduce colony risk but do not constitute population control [11]. Relocating colonies to hornet-free areas is often impractical in regions with widespread establishment and can impose additional costs that should be minimized under EU Regulation No. 1143/2014 [29]. Preventing spread to other Member States likewise requires management approaches that include legally viable nest elimination [29]. To date, systematic nest detection and removal remains the only consistently effective strategy [2].
A cost- and time-efficient method is the injection of insecticides using lance systems. However, in Germany and several other EU countries, there are no broadly authorized products for general use against V. velutina; available biocides are often restricted to use “in, on, or around” buildings. In addition, biocide use raises concerns about environmental residues and secondary poisoning of non-target organisms [30]. Similar concerns apply to “Trojan horse” baiting approaches, in which a biocide is applied to a worker in the expectation that it will be transported into the nest; environmental exposure, fate of treated individuals, and secondary poisoning risks are difficult to quantify, particularly when persistent and highly toxic substances (e.g., neonicotinoids) are involved. Purely mechanical approaches such as manual nest removal with vehicle-mounted lifts are limited by high operational costs, equipment logistics, terrain constraints, safety requirements, specialized personnel, and seasonal availability; accident risk is non-negligible. Shooting nests [28] is ineffective because nests are often repaired and reproductive interruption is not assured, and scattered workers may establish emergency nests that can increase public-health risk. CO₂-based immobilization is considered an unauthorized biocidal application under German biocidal-product provisions because biocides are legally defined by chemical interaction with the target organism [31]; CO₂ effects are mediated via reversible disruption of pH-dependent metabolic processes [32]. Steam treatment [33], while physical in principle, raises unresolved animal-welfare and ethical questions and imposes substantial technical and occupational-safety demands [34,35]. Overall, high equipment and maintenance costs, training requirements, and safe application constraints can be prohibitive for large-scale control under limited budgets.
For V. velutina nest control in public settings, no biocidal products are specifically authorized; transitional and case-by-case arrangements are heterogeneous and often impractical for area-wide programmes [36]. Diatomaceous earth has a physical mode of action but is classified as a biocide [37], shows weather-dependent performance [38] and is currently not authorized for invasive hornet control in Germany and Switzerland. In contrast, air-assisted lance systems provide a logistically efficient means of applying powdered agents with predictable one-time acquisition costs and high operational flexibility. In Rhineland-Palatinate (Germany), control of secondary nests in 2024 relied on such systems because they enabled flexible, cost-efficient operations with reported throughput of up to 15 nests per person per day, depending on nest location. Given that many nests occur in public areas, an applied substance must be legally unproblematic and either meet biocidal-product requirements or act outside the scope of biocidal regulation.
Accordingly, a biocide-free, legally robust, scalable, and effective method is required. Usability and clear animal-welfare compatibility are additional advantages, as they increase societal acceptance. Activated charcoal meets these criteria: it is chemically inert and acts via mechanical–physical processes. Physical dusts with low unit costs can support continued control of V. velutina, consistent with the obligation to continue management under Regulation (EU) No 1143/2014 [29]. The proposed mechanism (adsorption/surface effects) involves disruption of the cuticular lipid barrier, impairing water balance and leading to mortality [39]. The rapid functional impairment observed is also compatible with a suffocation component through mechanical occlusion of spiracles[40]. In laboratory and applied studies, activated charcoal showed effects comparable to or exceeding those of diatomaceous earth against the pharaoh ant, Monomorium pharaonis [39]. Because the action is mechanical, no biocidal authorization is required (subject to national competent-authority classification) [41]. Activated charcoal leaves no toxicological residues and is generally considered environmentally benign.
The aim of this study is to describe activated-charcoal nest control under field conditions and to evaluate its effectiveness using a standardized application protocol (including brood removal). We show that, when applied correctly, activated charcoal achieves effectiveness close to that of biocides.
2. Materials and Methods
2.1. Adult Mortality Assay (Flight Cage)
A complete colony (~400 adults, including a laying queen, young gynes, drones, and workers) was gently separated from the combs and transferred to a flight cage (38 × 38 × 60 cm³; four mesh sides, one PVC observation window, and a textile floor; QIODAZOO). Individuals were allowed to acclimate for approximately 30 minutes. Activated charcoal dust was then applied to the entire cage as “swarm dusting” using a Birchmeier DR5 powder sprayer (Birchmeier Sprühtechnik AG, Stetten, Switzerland). Three bursts were delivered within the first 5 minutes, followed by one additional burst at t = 20 minutes; burst duration was standardized to 2 seconds. Trials were conducted at 23 °C and 65% relative humidity. Primary endpoints were (i) sustained loss of flight capability (≥30 s) and (ii) mortality. Secondary observations included disorientation, loss of perching ability, impaired motor coordination, grooming behaviour, and visible nocifensive reactions.
2.2. Prospective Field Efficacy Study: Setting and Case Selection
A prospective field study was conducted during routine control operations in Rhineland-Palatinate, Germany, in 2024. Fifteen secondary nests of Vespa velutina were included if they were (i) reachable with an injection lance, (ii) readily observable, and (iii) associated with low immediate risk to unprotected bystanders.
For external confirmation, an independent dataset was collected in Switzerland using the same protocol (N = 25). Nest height was estimated from the number/length of extended lance segments up to the nest edge. Nest volume was estimated visually using prior calibration with rotation ellipsoids derived from reference nests. Nests where the surrounding area could not be adequately cordoned off were not included. No experiments were conducted during rain.
Activated charcoal was used as a black powder (S3 Handel und Dienstleistungen UG) with a particle size <40 µm, a BET surface area of approximately 900 m²/g, pH 9–12, water content ≤10%, and ash content ≤ 9% (CAS 7440-44-0; EC 231-153-3), at an estimated cost of ~€30 per kg. Field applications were performed with a “Buzz Busters” lance system (Buzzbusters, Guillerval, France) providing a basic reach of 20 m and extendable to ~30 m, operated with a battery-powered compressor (Einhell TE-AC 36/6/8 Li) and equipped with a moisture separator and emission-reducing application tips. For the flight-cage assay, activated charcoal was applied using a Birchmeier DR5 powder sprayer. Personal protective equipment consisted of a full hornet protective suit, gloves, and face protection/veil, along with cordoning equipment.
Field treatments followed a standardized application protocol. Activated charcoal was applied preferentially via the nest entrance; when the entrance was inaccessible, the nest envelope was punctured and the dust was introduced from below. Working pressure was adjusted to approximately 1 bar per 10 m of working height and increased stepwise as needed. Each nest received 50–100 g of activated charcoal delivered as an initial application followed by a second application 15 minutes later. Brood was removed by mechanically extracting combs using the “destroyer” attachment of the lance system; removed comb material was collected and disposed of. Where necessary, remaining envelope fragments were re-dusted to target adults still present. Operations were not conducted during rain, and deployments under strong wind were avoided.
Field outcomes were defined a priori: Primary outcomes were (i) acute functional impairment, recorded as flight incapacity and/or falling to the ground within the first hour after application, and (ii) nest inactivation, defined as the presence of no more than 1–2 adults during a 10-minute observation period on day 7 and the absence of flight activity by day 10. Secondary outcomes included whether the nest was repaired or continued operation (yes/no), the formation of emergency nests in the vicinity, and observable brood changes (e.g., discoloration or dust deposition). We recorded the applied dose (g), working time (min; from the beginning of setup to completion of teardown), and working height (m).
2.2.1. Data Collection and Quality Assurance
Observations were recorded using a standardized schedule with time stamps (0–60 min, day 1, day 7/10), with two observers whenever feasible. Fallen adults were not collected to avoid interfering with the course of effects. Equipment function (pressure build-up, nozzle fit) was checked before each deployment.
2.2.2. Preliminary Study on the Need of Mechanical Destruction
In order to test whether the application of activated charcoal alone without subsequent mechanical destruction of the nests would be sufficient, we left three nests undamaged after the application. We stopped this trial because in all three cases, the nests were still active 10 days after the treatment.
2.3. Independent Field Validation (Switzerland, 2025) and Insecticide Comparator
For external validation, two independent experimenters treated 108 secondary nests in the cantons of Basel-Stadt and Basel-Landschaft, Switzerland, in 2025 using the identical protocol (Supplemental Figure S1). Outcomes were documented in a standardized format; success control was performed at day 7 only (criterion: nest inactivation).
For contextual comparison, outcomes were contrasted with 136 nests treated in 2024 in Rhineland-Palatinate, Germany, with a biocide (Vespa; 0.5% cypermethrin; Armosa Tech SA) using an analogous operational approach (Supplemental Figure S2).
2.4. Sample Preparation for Microscopy (Mechanistic Assessment)
2.4.1. Spiracle Preparations
To assess morphological changes, thoraces were separated and embedded in UV-curing resin (MOCOBO, UV Resin, Hard Type) to improve stability and handling. Resin blocks were mechanically ground (Parkside, PTSG 140 C2) until spiracles and adjacent tissue remnants were exposed. Samples were incubated for 5 days in 1 M NaOH (S3 Chemicals) to macerate organic material and dissolve resin. After soft tissue dissolution, remaining chitinous thoracic structures were rinsed twice with demineralized water.
Specimens were bleached in 5% H₂O₂ (S3 Chemicals) for 30 min in a water bath at 80 °C and then left in solution for an additional 48 h until sufficient transparency for transmitted-light microscopy was achieved. As activated charcoal is chemically inert to H₂O₂, it remained visible as a distinct black material within tissues. Final mounting was performed in Kaiser’s glycerol gelatine (S3 Chemicals).
2.4.2. Tracheal Preparations
To minimize contamination during dissection, hornets were repeatedly washed in 0.1% SCS (sodium coco sulfate surfactant solution; Salandis GmbH) until no external charcoal residues were visible. Thoraces were then sagittally bisected. Primary tracheae and air sac membranes were dissected under water and mounted in Kaiser’s glycerol gelatine (S3 Chemicals).
3. Results
3.1. Adult Mortality Assay (Flight Cage; ~400 Adults)
Activated charcoal dusting caused rapid functional impairment followed by near-complete mortality (supplemental Figure S1). After the second to third dusting (t = 5 min), disorientation and flight disturbance (falls) were evident; at t = 5 min, ~25% of individuals were immobile or showed intensive grooming, and by t = 10 min ~50% were on the cage floor. After the additional application at t = 20 min, all individuals were flightless by t = 25 min. By t = 30 min, ~95% were on the floor with markedly reduced activity; between t = 40 and 60 min most individuals were moribund. At t = 18 h, mortality exceeded 99%, with only occasional very slow leg movements in isolated individuals. No conspicuous nocifensive behaviours (e.g., body curling or sudden aggression peaks) were observed; behaviour was dominated by grooming and crawling until immobilization.
3.2. Prospective Field Series (Germany 2024; n = 15 Secondary Nests)
During the acute observation period (0–60 min), many workers exited the nest immediately after application; within minutes, most individuals showed marked flight impairment and fell to the ground (Figure 1). Overall nest traffic declined sharply and was strongly reduced within approximately 1 hour. By day 1, numerous dead individuals had accumulated beneath treated nests. At follow-up, sporadic residual activity was occasionally observed on day 7 (~1–2 adults per 10 min), but by day 10 no flight activity was detected. Across all cases, no nest repair or continued operation was observed (15/15).
Operationally, 50–100 g of activated charcoal was applied per nest (mean 71 g). Total on-site time ranged from 40 to 70 minutes (mean 54 min, including setup and teardown). Mean working height was 9 m and mean estimated nest volume was 46 L, although treatments at heights of 20–30 m were feasible. No stings were recorded during operations, and no deployments were conducted during rain.
In all three cases where we purposefully did not destroy the nest mechanically after treatment with activated charcoal, activity at the nest persisted 10 days after treatment.
Larvae inside the combs were also affected by the activated carbon powder (Supplemental Figure S4) and turned yellowish.
Pupae were not visibly affected, as they are protected by a silk cocoon.
3.3. Cross-Site Operational Parameters (Germany 2024 vs Switzerland 2025; Subset Comparison)
A subset comparison (Germany: n = 15; Switzerland: n = 25) indicated that Swiss deployments targeted higher nests on average (18 ± 8.2 m vs 9.2 ± 3.8 m), whereas estimated nest volume was larger in the German sample (46 ± 5.8 L vs 38 ± 7.8 L). Despite smaller nests, the mean charcoal mass and working time were higher in Switzerland (96 ± 14 g; 86 ± 48 min) than in Germany (71 ± 12 g; 54 ± 9.1 min). The mean volume-specific dose was 2.6 ± 0.4 g/L in Switzerland vs 1.6 ± 0.3 g/L in Germany (+66%) (Table 1, Supplemental Table S1).
Within the observed autumnal range of temperature and relative humidity reported for both datasets, no efficacy differences were apparent in this subset (all nests inactive at day 7; no repairs or emergency nests observed). Because no failures occurred in this subset and environmental variability was limited, no weather- or dose-dependent thresholds could be inferred.
3.4. Field Efficacy of Activated Charcoal and Comparison with a Biocide Dust (Day-7 Endpoint)
After describing cross-site differences in deployment conditions and dosing intensity (Section 3.3), we assessed efficacy based on the day-7 terminal endpoint (nest inactivation) across the pooled protocol-adherent activated-charcoal dataset (Germany and Switzerland) and compared outcomes with an operational cypermethrin-based dust series. Across all protocol-adherent activated-charcoal treatments (total n = 145 secondary nests), 142/145 nests reached the terminal endpoint at day 7 (97.9%), whereas 3/145 nests remained active and/or showed repair (2.1%) (Table 2). The pooled dataset comprised the German prospective series (2024; n = 15; all inactive at day 7), a Swiss confirmation series (2025; n = 25 ; all inactive at day 7; operational-parameter data available for a subset reported in Section 3.3), and an independent Swiss field test (2025; n = 105). In the independent Swiss field test, 102/105 nests were inactive at day 7 (97.1%), while 3/105 showed persistent activity and/or repair (2.9%); these non-terminal outcomes were characterized by small repair structures despite otherwise protocol-adherent treatment. In a separate ancillary test evaluating whether brood removal could be omitted, three additional nests were treated with activated charcoal without brood removal (protocol deviation); all three remained active and exhibited repair at day 7 (0/3 inactivated), supporting brood removal as a required protocol element. For contextual comparison, a cypermethrin-based biocide dust applied via the same lance system in Germany (2024; n = 136) resulted in 134/136 (= 98.5%) nests being inactive at day 7, whereas 2/136 (= 1.5%) nests showed repair and/or emergency nest formation. This result (2 alive nests out of 136 alive) is not significantly different from the results of the activated charcoal trial (3 alive nests out of 145) (Fisher’s exact test, p = 1.000). Operationally, cessation of nest traffic occurred earlier than with activated charcoal, consistent with the expected rapid knockdown effect of the active ingredient (Table 2).
3.5. Microscopy
Transmitted-light microscopy of macerated and bleached thoracic sections revealed dense accumulations of activated charcoal particles within the spiracular atrium (Figure 2). Deposits persisted despite repeated washing and H₂O₂ treatment, and the atrial region appeared mechanically compromised by particle loading. We detected activated charcoal throughout the tracheal system, including partial to extensive occlusion of the main tracheae (Figure 3). Occasional particle adhesion was also observed on air sac membranes.
4. Discussion
4.1. Principal Findings
Under a standardized application protocol, activated charcoal rapidly incapacitated adult Vespa velutina and was followed by cessation of nest activity within 7days. Mortality in the flight-cage assay exceeded 99% after 18 h and aligned with the field observations.
4.2. Mode of Action, Welfare Considerations, and Safety
Across protocol-adherent field deployments, nests were reliably inactivated without behavioural patterns commonly interpreted as pronounced nocifensive responses (e.g., marked body curling, vigorous writhing on the ground, or abrupt aggression peaks). The observed sequence — disorientation, loss of flight capability, immobilization, and death supports a mechanical–physical mode of action. Two complementary mechanisms [42] are consistent with the data: (i) disruption of the cuticular lipid barrier with subsequent impairment of water balance (desiccation) (reviewed in [39]) and (ii) (partial) occlusion of spiracles and airways leading to respiratory compromise
Efficacy likely increases during phases of high worker activity. Nest disturbance induces elevated ventilation associated with flight, which may enhance particle intake from dust-saturated air and promote spiracle occlusion, accelerating functional impairment. Practically, this highlights the importance of personal protective equipment, cordoning, and weather-adapted deployments to optimize efficacy while minimizing risk to applicators and bystanders.
Effective secondary-nest control also requires consideration of colony relocation dynamics. If migration from the primary to the secondary nest is still ongoing at treatment, returning individuals may repopulate the treated nest, potentially resulting in repairs or small emergency nests. After queen loss, such emergency nests may become male-biased if workers begin laying unfertilized eggs.
4.3. Mechanistic Evidence from Microscopy
Microscopy supported uptake of activated charcoal via the respiratory system. Persistent particle loads in spiracular atria indicate mechanical impairment at the entry of the tracheal system, while charcoal in the main tracheae—including partial to extensive luminal occlusion—supports impaired gas exchange. Particle adhesion on air sac membranes further aligns with respiratory insufficiency as a relevant contributor to lethality. These findings match the phenotype observed in the mortality assay, particularly rapid loss of flight performance followed by immobilization. Tracheal obstruction [43] may represent a dominant pathway under the high airborne dust conditions generated during nest treatment [44].
Inhalation, albeit not obstruction, was also regarded as a prominent factor by [45]. By contrast, earlier studies in the much smaller (2-5 mm) bean weevil, Acanthoscelides obtectus, found no evidence for impaired respiration [44]. [46,47] excluded blockage of the trachea when using sodium fluoride (NaF) or magnesium carbonate (MgCO3), and [48] could also not find dust particles in the trachea. Possibly, insects in flight are more affected than crawling insects.
Cuticular disruption is also likely to contribute, particularly in individuals with lower respiratory uptake, suggesting a combined mechanism in which the relative importance of respiratory blockage versus desiccation depends on the balance between airway occlusion and whole-body coverage.
Sensory impairment may additionally contribute to early disorientation. Activated charcoal can coat compound eyes and antennal sensilla, potentially reducing visual and olfactory function. Dust-related interference with antennal olfaction has been demonstrated in flies [49] and may be particularly consequential in eusocial insects that rely on chemical communication for colony cohesion and coordinated defence.
4.4. Operational Feasibility and Cost Considerations
The approach is material-sparing and operationally efficient: treatments required 50–100 g activated charcoal per nest and mean on-site times of 54 min (Germany) and 86 min (Switzerland). Lance-based application to heights of 20–30 m reduces dependence on vehicle-mounted lifting equipment. The protocol is readily implemented as an standard operation procedure (SOP) (e.g., pressure scaling of ~1 bar per 10 m working height; second application after 15 min; brood removal), facilitating training, quality assurance, and scaling.
4.5. Contextualization Relative to Existing Approaches
Compared with synthetic insecticides, activated charcoal does not produce immediate knockdown but instead induces rapid functional impairment and a predictable colony collapse within a defined post-treatment window. Relative to diatomaceous earth, the activated-charcoal approach is intended as biocide-free in jurisdictions where mechanically acting dusts are not classified as biocidal products; it may also be less sensitive to weather-related limitations reported for some dust-based interventions. Laboratory and applied studies in other insect systems have reported comparable or superior performance of activated charcoal relative to diatomaceous earth [39,48,50,51].
A practical advantage is the absence of toxicological residues, minimizing concerns about secondary poisoning of non-target organisms [30]. Where appropriate, treated nest material may therefore be left in situ and could remain available to native scavengers. Finally, resistance evolution is expected to be less likely than with neurotoxic insecticides, as adaptation would require more complex morphological or behavioural changes [52] rather than target-site or metabolic resistance mechanisms [53,54,55,56].
4.7. Implications and Future Directions
Activated charcoal represents a legally robust, biocide-free, and scalable option for V. velutina nest management, particularly where authorized insecticides are unavailable or undesirable. Priority next steps include:
- Controlled comparative trials against alternative mineral/physical dusts (e.g., kaolin, zeolites) and, where legally permissible, benchmark insecticides, using standardized time-to-event endpoints.
- Optimization of dose and particle properties (size distribution, surface area, deposition).
- Stratified analyses across season and colony phase (nest stage, colony size, migration status).
- Targeted monitoring of non-target impacts and environmental pathways under operational conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Location of nests in Switzerland treated with activated charcoal;; Figure S2: Location of nests in Germany treated with biocide; Figure S3: Mortality and Loss of Flight after treatment with activated charcoal; Figure S4: Larvae inside combs are affected by the activated carbon powder and turned yellowish; Table S1: Data on nest height, nest volume, mass charcoal, duration, temperature, relative humidity; Table S2: Comparison of efficacy of treatments with activated charcoal and biocide.
Author Contributions
Conceptualization, A.P. and U.E.; Methodology, A.P. and U.E.; Validation, A.P. and U.E.; Formal Analysis, A.P. and U.E.; Investigation, A.P. and U.E.; Resources, A.P.; Data Curation, A.P. and U.E.; Writing – Original Draft Preparation, A.P. and U.E.; Writing – Review & Editing, A.P. and U.E.; Visualization, A.P. and U.E.; Supervision, A.P. and U.E.; Project Administration, A.P.; Funding Acquisition, A.P.
Funding
This research received no external funding.
Data Availability Statement
All data are presented within this publication.
Acknowledgments
The authors would like to acknowledge Maria Corpataux and Joost Oerlemans for their generous help in gathering data on nest destructions and emergency nest constructions. The authors also acknowledge the support of Dienstleistungszentrum Ländlicher Raum (DLR) Westerwald-Osteifel (Service Center for Rural Areas (DLR) Westerwald-Osteifel), Imkerverband Rheinland-Pfalz e.V. (Beekeepers’ Association Rhineland-Palatinate), and Ministerium für Klimaschutz, Umwelt, Energie und Mobilität Rheinland-Pfalz (Ministry for Climate Protection, Environment, Energy, and Mobility Rhineland-Palatinate).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| SD | Standard Deviation |
References
- Roy, H. E.; Pauchard, A.; Stoett, P.; Truong, T. Renard. “IPBES Invasive Alien Species Assessment: Full report”. [CrossRef]
- Lioy, S.; Bergamino, C.; Porporato, M. The invasive hornet Vespa velutina: distribution, impacts and management options. In CABI International; 01 Jan 2022. [Google Scholar] [CrossRef]
- Choi, M. B.; Martin, S. J.; Lee, J. W. Distribution, spread, and impact of the invasive hornet Vespa velutina in South Korea. J. Asia. Pac. Entomol. 2012, vol. 15(no. 3), 473–477. [Google Scholar] [CrossRef]
- Kim, J.; Choi, M.; Moon, T. Occurrence of Vespa velutina Lepeletier from Korea, and a revised key for Korean Vespa species (Hymenoptera: Vespidae). Entomol. Res. 2006, vol. 36(no. 2), 112–115. [Google Scholar] [CrossRef]
- Dillane, E.; Hayden, R.; O’Hanlon, A.; Butler, F.; Harrison, S. The first recorded occurrence of the Asian hornet (Vespa velutina) in Ireland, genetic evidence for a continued single invasion across Europe. J. Hymenopt. Res. 2022, vol. 93, 131–138. [Google Scholar] [CrossRef]
- Hoebeke, E. R.; Bartlett, L. J.; Evans, M.; Freeman, B. E.; Wares, J. P. First Records of Vespa velutina (Lepeletier) (Color form Nigrithorax) (Hymenoptera: Vespidae) in North America, an Invasive Pest of Domesticated Honeybees. Proc. Entomol. Soc. Wash. 2024, vol. 126(no. 2), 193–205. [Google Scholar] [CrossRef]
- Vespa velutina nigrithorax. von Kouba, DZ-AL, DZ am 17. August 2025 um 05:13 PM von Kevin Snair · iNaturalist. 21 Dec 2025. Available online: https://www.inaturalist.org/observations/307103550.
- Semmar, S. The First Record of Vespa velutina Lepeletier, 1836 in Algeria and Biocontrol Implications. Commagene J. Biol. 2025, vol. 9(no. 2), 243–244. [Google Scholar] [CrossRef]
- Public urged to look out for hornets in Auckland | NZ Government. 21 Dec 2025. Available online: https://www.mpi.govt.nz/news/media-releases/public-urged-to-look-out-for-hornets-in-auckland.
- Arias, A.-I.; Ferreira-Golpe, M.; Vázquez-González, I. V.; Nave, A.; Thiery, D.; Godinho, J. Economic costs and practices to control Vespa velutina nigrithorax in beekeeping: a survey in four regions in Europe 2023. [CrossRef]
- Requier, F.; Rome, Q.; Villemant, C.; Henry, M. A biodiversity-friendly method to mitigate the invasive Asian hornet’s impact on European honey bees. J. Pest Sci. (2004). 2020, vol. 93(no. 1), 1–9. [Google Scholar] [CrossRef]
- Lueje, Y. R.; Jácome, M. A.; Servia, M. J. New problems for old vineyards: Mitigating the impacts of yellow-legged hornets (Vespa velutina) in a historical wine-producing area. Agric. Ecosyst. Environ. 2024, vol. 367, 108969. [Google Scholar] [CrossRef]
- Rojas-Nossa, S. V.; Calviño-Cancela, M. The invasive hornet Vespa velutina affects pollination of a wild plant through changes in abundance and behaviour of floral visitors. Biol. Invasions 2020, vol. 22(no. 8), 2609–2618. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Salles, J.-M.; Courchamp, F. The economic cost of control of the invasive yellow-legged Asian hornet. NeoBiota 2020, vol. 55, 11–25. [Google Scholar] [CrossRef]
- Requier, F.; Fournier, A.; Pointeau, S.; Rome, Q.; Courchamp, F. Economic costs of the invasive Yellow-legged hornet on honey bees. Science of The Total Environment 2023, vol. 898, 165576. [Google Scholar] [CrossRef]
- Feás, X. Human Fatalities Caused by Hornet, Wasp and Bee Stings in Spain: Epidemiology at State and Sub-State Level from 1999 to 2018. Biology (Basel). 2021, vol. 10(no. 2), 73. [Google Scholar] [CrossRef]
- Pedersen, S. Broad ecological threats of an invasive hornet revealed through a deep sequencing approach. Science of The Total Environment vol. 970, 178978, 2025. [CrossRef]
- Haubrock, P. J. Economic costs of invasive alien species across europe. NeoBiota 2021, vol. 67, 153–190. [Google Scholar] [CrossRef]
- Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and selectiveness of traps and baits for catching the invasive hornet Vespa velutina. Insects 2020, vol. 11(no. 10), 1–13. [Google Scholar] [CrossRef]
- Teodoro Correia, A. Impact of Vespa velutina. monitoring baited traps on the entomofauna of three protected Portuguese areas. 09 Feb 2026. Available online: https://repositorio.ulisboa.pt/bitstream/10400.5/97363/1/TM_Alexandra_Correia.pdf.
- Pérez-Granados, B. Testing the selectiveness of electric harps: a mitigation method for reducing Asian hornet impact at beehives. J. Apic. Res. 2024, vol. 63(no. 2), 360–366. [Google Scholar] [CrossRef]
- Rojas-Nossa, S. V; Dasilva-Martins, D.; Mato, S.; Bartolomé, C.; Maside, X.; Garrido, J. Effectiveness of electric harps in reducing Vespa velutina predation pressure and consequences for honey bee colony development. Pest Manag. Sci. 2022, vol. 78(no. 12), 5142–5149. [Google Scholar] [CrossRef]
- Rein, Carolin. Bericht über die Versuche zu selektiven Fallen zum Fang der Asiatischen Hornisse (Vespa velutina nigrithorax. im Frühjahr 2025 in Baden-Württemberg, Hohenheim, Aug. 2025. 09 Feb 2026. Available online: https://bienenkunde.uni-hohenheim.de/fileadmin/einrichtungen/bienenkunde/PDF/Vespa_Velutina/Bericht_Selektive_Fallen_Fruehjahr_2025.pdf.
- Rojas-Nossa, S. V.; Mato, S.; Feijoo, P.; Lagoa, A.; Garrido, J. Comparison of Effectiveness and Selectiveness of Baited Traps for the Capture of the Invasive Hornet Vespa velutina. Animals 2024, vol. 14(no. 1). [Google Scholar] [CrossRef]
- Rojas-Nossa, S. V.; Novoa, N.; Serrano, A.; Calviño-Cancela, M. Performance of baited traps used as control tools for the invasive hornet Vespa velutina and their impact on non-target insects. Apidologie 2018, vol. 49(no. 6), 872–885. [Google Scholar] [CrossRef]
- Sánchez, O.; Arias, A. “All That Glitters Is Not Gold: The Other Insects That Fall into the Asian Yellow-Legged Hornet Vespa velutina ‘Specific’ Traps,”. Biology (Basel). 2021, vol. 10(no. 5), 448. [Google Scholar] [CrossRef]
- Monceau, K.; Thiéry, D. Vespa velutina nest distribution at a local scale: An 8-year survey of the invasive honeybee predator. Insect Sci. 2017, vol. 24(no. 4), 663–674. [Google Scholar] [CrossRef]
- Turchi, L.; Derijard, B. Options for the biological and physical control of Vespa velutina nigrithorax (Hym.: Vespidae) in Europe: A review. Journal of Applied Entomology 2018, vol. 142(no. 6), 553–562. [Google Scholar] [CrossRef]
- Commission. REGULATION (EU) No 1143/2014 OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species, Oct 2014.
- Guedes, R. N. C.; Berenbaum, M. R.; Biondi, A.; Desneux, N. The Side Effects of Pesticides on Nontarget Arthropods. Annu. Rev. Entomol. 2026, vol. 71(no. 1), 381–403. [Google Scholar] [CrossRef]
- Commission. REGULATION (EU) No 528/2012 OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL, May 2022.
- Badre, N. H.; Martin, M. E.; Cooper, R. L. The physiological and behavioral effects of carbon dioxide on Drosophila melanogaster larvae. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, vol. 140(no. 3), 363–376. [Google Scholar] [CrossRef]
- Ruiz-Cristi, I.; Berville, L.; Darrouzet, E. Characterizing thermal tolerance in the invasive yellow-legged hornet (Vespa velutina nigrithorax): The first step toward a green control method. PLoS One 2020, vol. 15(no. 10), e0239742. [Google Scholar] [CrossRef]
- Gibbons, M. Measuring Pain in Insects. Animal Behaviour and Welfare Cases 2025. [Google Scholar] [CrossRef]
- Gibbons, M.; Crump, A.; Barrett, M.; Sarlak, S.; Birch, J.; Chittka, L. Can insects feel pain? A review of the neural and behavioural evidence. In Advances in Insect Physiology; Academic Press Inc., 2022; vol. 63, pp. 155–229. [Google Scholar] [CrossRef]
- Baua ECHA Hilfen. Available online: https://www.reach-clp-biozid-helpdesk.de/DE/Helpdesk.
- Commission. COMMISSION IMPLEMENTING REGULATION (EU) 2017/ 794 - of 10 May 2017 - approving silicon dioxide Kieselguhr as an existing active substance for use in biocidal products of product-type 18.
- Ogreten, A. Insecticidal efficacy of native raw and commercial diatomaceous earths against Tribolium confusum DuVal (Coleoptera: Tenebrionidae) under different environmental conditions. J. King Saud Univ. Sci. 2023, vol. 35(no. 7), 102827. [Google Scholar] [CrossRef]
- Van Den Noortgate, H.; Lagrain, B.; Pulinthanathu Sree, S.; Kerkhofs, S.; Wenseleers, T.; Martens, J. A. Material properties determining the insecticidal activity of highly divided porous materials on the pharaoh ant (Monomorium pharaonis). Pest Manag. Sci. 2018, vol. 74(no. 6), 1374–1385. [Google Scholar] [CrossRef]
- Alexander, P.; Kitchener, J.; Briscoe, H. Inert dust insecticides: Part I. Mechanism of action. Annals of Applied Biology 2008, vol. 31, 143–149. [Google Scholar] [CrossRef]
- BfR Übersicht zu Biozidprodukten. Available online: https://www.bfr.bund.de/chemikaliensicherheit/biozidprodukte-und-behandelte-waren/.
- Mewis, I.; Ulrichs, C. Action of amorphous diatomaceous earth against different stages of the stored product pests Tribolium confusum, Tenebrio molitor, Sitophilus granarius and Plodia interpunctella. J. Stored Prod. Res. 2001, vol. 37(no. 2), 153–164. [Google Scholar] [CrossRef]
- Schneiderman, H. A. Discontinuous Respiration in Insects: Role of the Spiracles. Biol. Bull. 1960, vol. 119(no. 3), 494–528. [Google Scholar] [CrossRef]
- Chiu, S. F. Toxicity Studies of So-Called ‘Inert’ Materials With the Bean Weevil, Acanthoscelides obtectus (Say). J. Econ. Entomol. 1939, vol. 32(no. 2), 240–248. [Google Scholar] [CrossRef]
- Webb, E. The Penetration of Derris through the Spiracles and Cuticle of Melophagus ovinus, L. Bull. Entomol. Res. 1946, vol. 36(no. 1), 15–22. [Google Scholar] [CrossRef]
- Hochenyos, G. L. Effect of Dusts on the Oriental Roach. J. Econ. Entomol. 1933, vol. 26(no. 4), 792–794. [Google Scholar] [CrossRef]
- Hochenyos, G. L. The Mechanism of Absorption of Sodium Fluoride by Roaches. J. Econ. Entomol. 1933, vol. 26(no. 6), 1162–1169. [Google Scholar] [CrossRef]
- Alexander, P.; Kitchener, J. A.; Briscoe, H. V. A. Inert dust insecticides: Part I. Mechanism of action. Annals of Applied Biology 1944, vol. 31(no. 2), 143–149. [Google Scholar] [CrossRef]
- Wang, Q. Short-term particulate matter contamination severely compromises insect antennal olfactory perception. Nat. Commun. 2023, vol. 14(no. 1), 4112. [Google Scholar] [CrossRef]
- Ebeling, W.; Wagner, R. E. Rapid Desiccation of Drywood Termites with Inert Sorptive Dusts and Other Substances. J. Econ. Entomol. 1959, vol. 52(no. 2), 190–207. [Google Scholar] [CrossRef]
- Van Den Noortgate, H. Material properties determining insecticidal activity of activated carbon on the pharaoh ant (Monomorium pharaonis). J. Pest Sci.. 2019, vol. 92(no. 2), 643–652. [Google Scholar] [CrossRef]
- Zalucki, M.; Furlong, M. Behavior as a mechanism of insecticide resistance: evaluation of the evidence. Curr. Opin. Insect Sci. 2017, vol. 21, 19–25. [Google Scholar] [CrossRef]
- Soderlund, D. M.; Bloomquist, J. R. Molecular Mechanisms of Insecticide Resistance. In in Pesticide Resistance in Arthropods; Springer US: Boston, MA, 1990; pp. 58–96. [Google Scholar] [CrossRef]
- Al Naggar, Y. Mechanisms and Genetic Drivers of Resistance of Insect Pests to Insecticides and Approaches to Its Control. Toxics 2025, vol. 13(no. 8), 681. [Google Scholar] [CrossRef]
- ffrench-Constant, R. H. The Molecular Genetics of Insecticide Resistance. Genetics 2013, vol. 194(no. 4), 807–815. [Google Scholar] [CrossRef]
- Liang, J. Insect Resistance to Insecticides: Causes, Mechanisms, and Exploring Potential Solutions. Arch. Insect Biochem. Physiol. 2025, vol. 118(no. 2). [Google Scholar] [CrossRef]
Figure 1.
Individual Vespa velutina nigrithorax workers covered in activated charcoal. After application of activated charcoal, workers (and where present gynes and drones) leave the nest and fly or fall to the ground, where they often remain motionless within minutes.
Figure 1.
Individual Vespa velutina nigrithorax workers covered in activated charcoal. After application of activated charcoal, workers (and where present gynes and drones) leave the nest and fly or fall to the ground, where they often remain motionless within minutes.

Figure 2.
(a) Dense deposit of activated charcoal particles outside of a spiracle in the thorax. Seen from within the body, charcoal particles accumulate around the spiracle; (b) The schema drawing clarifies the location of the spiracle.
Figure 2.
(a) Dense deposit of activated charcoal particles outside of a spiracle in the thorax. Seen from within the body, charcoal particles accumulate around the spiracle; (b) The schema drawing clarifies the location of the spiracle.
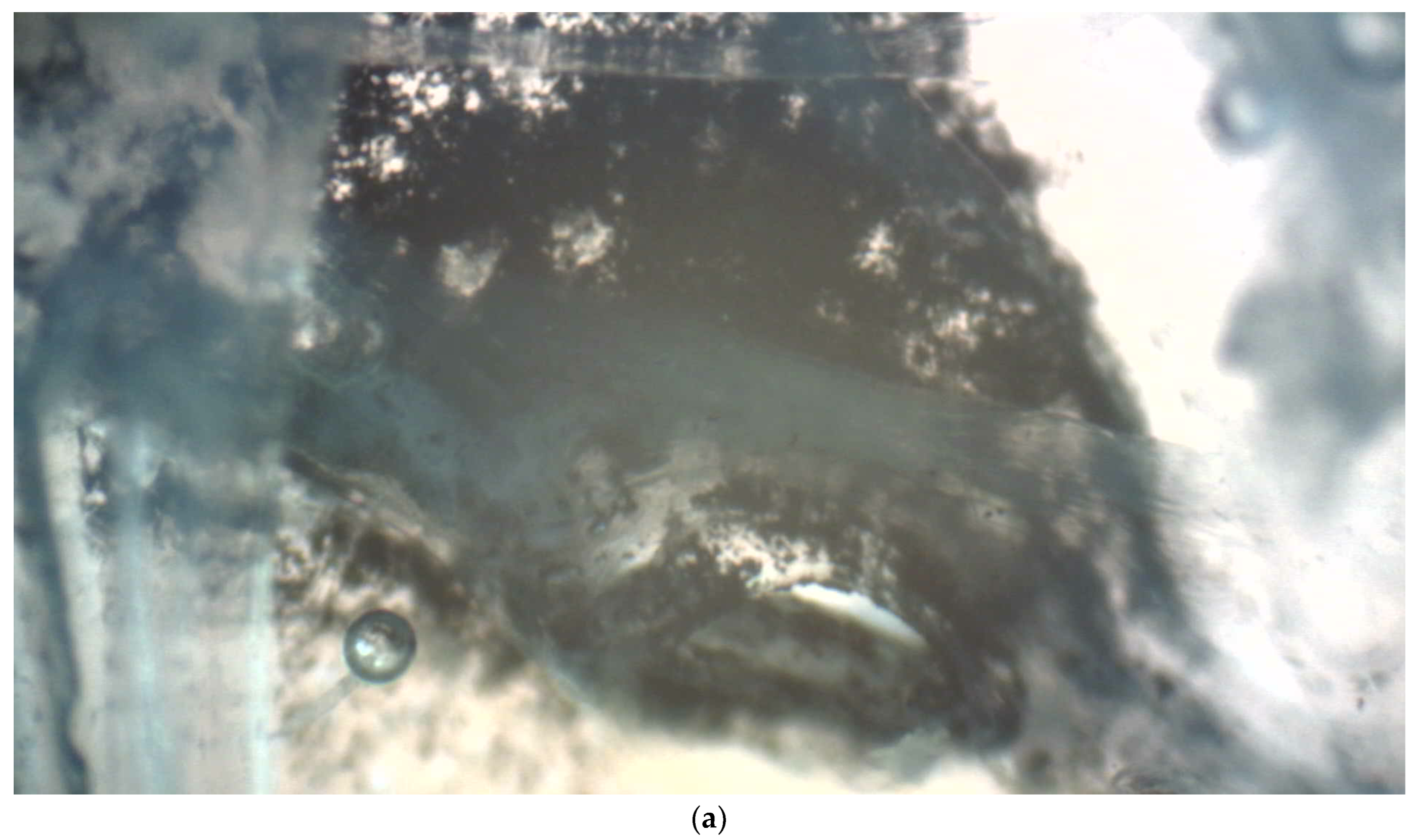
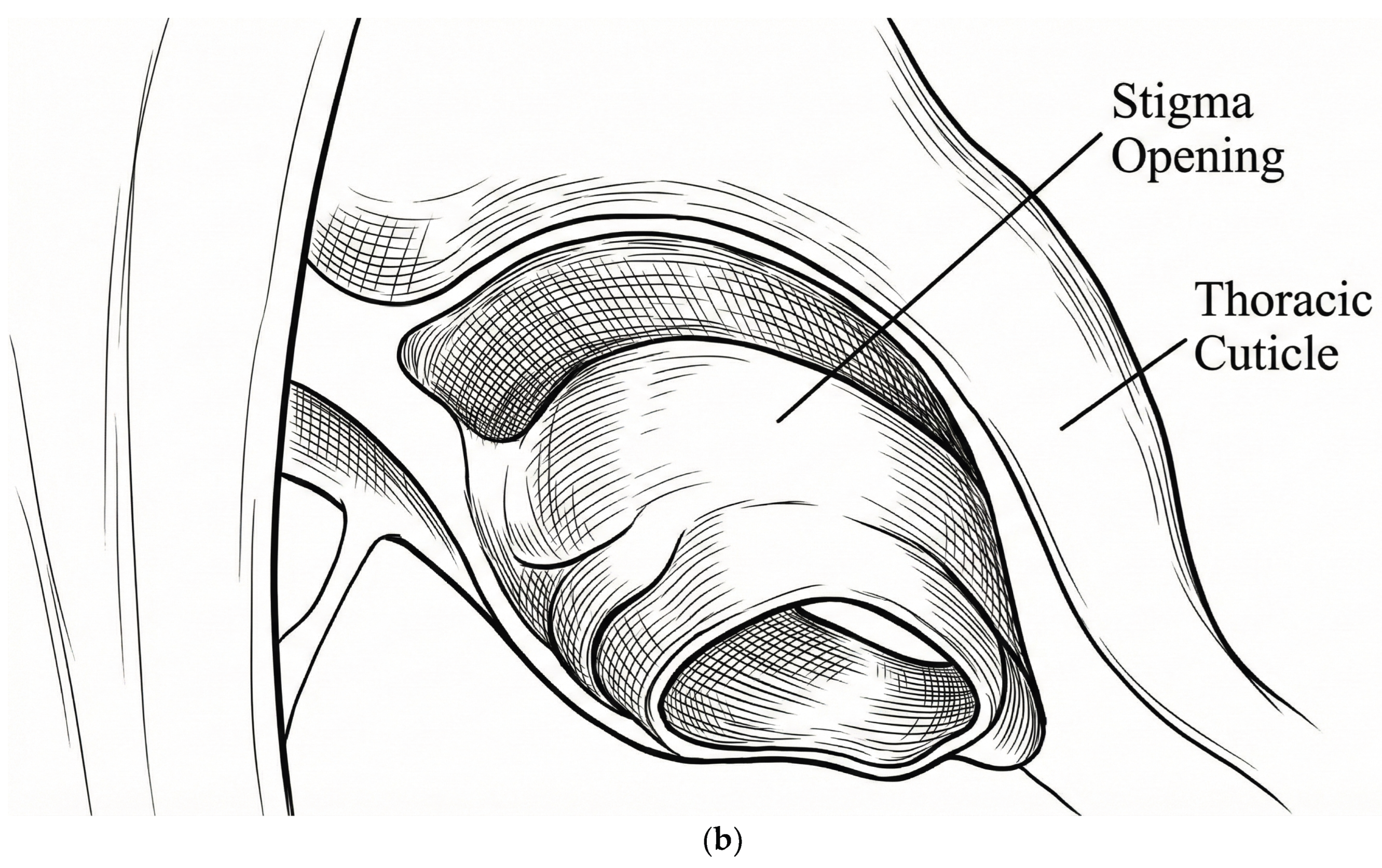
Figure 3.
Particles of activated charcoal entered the tracheal system. Shown is the main trachea in the thorax. Circular structures are artefacts (air bubbles). After treatment with activated charcoal, individuals died and were kept frozen till dissection. This trachea was taken from the thorax of a hornet that died after the treatment protocol. The thorax was dissected under water, and the trachea was detached from the stigma’s atrium using forceps and transferred into glycerol for light-microscopic examination.
Figure 3.
Particles of activated charcoal entered the tracheal system. Shown is the main trachea in the thorax. Circular structures are artefacts (air bubbles). After treatment with activated charcoal, individuals died and were kept frozen till dissection. This trachea was taken from the thorax of a hornet that died after the treatment protocol. The thorax was dissected under water, and the trachea was detached from the stigma’s atrium using forceps and transferred into glycerol for light-microscopic examination.
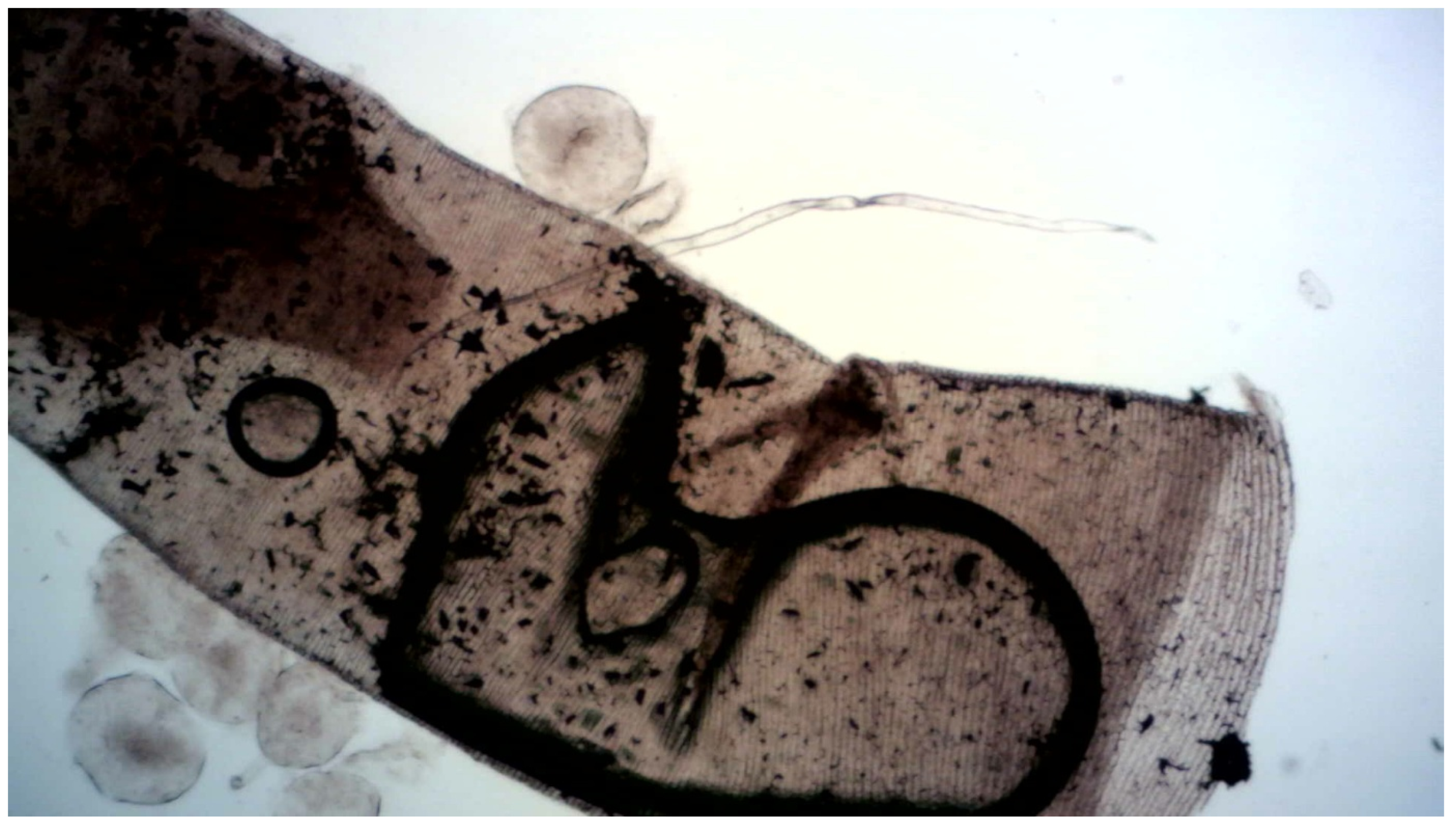

Table 1.
Operational parameters for activated-charcoal treatment (subset comparison).
| Parameter (mean ± SD) | Germany 2024 (n = 15) | Switzerland 2025 (n = 25) |
|---|---|---|
| Nest height (m) | 9.2 ± 3.8 | 18± 8.2 |
| Estimated nest volume (L) | 46 ± 5.8 | 38 ± 7.8 |
| Activated charcoal (g) | 71 ± 12 | 96 ± 14 |
| Dose per volume (g/L) | 1.6 ± 0.3 | 2.6 ± 0.4 |
| Working time (min) | 54 ± 9.1 | 86 ± 48 |
Table 2.
Table 2. Day-7 terminal endpoint (nest inactivation) after treatment with activated charcoal (by dataset; pooled protocol-adherent performance; and ancillary brood-removal omission test) and a cypermethrin-based biocide dust comparator.
Table 2.
Table 2. Day-7 terminal endpoint (nest inactivation) after treatment with activated charcoal (by dataset; pooled protocol-adherent performance; and ancillary brood-removal omission test) and a cypermethrin-based biocide dust comparator.
| Treatment / group | Location (year) | Protocol status | n | Inactive at day 7, n (%) | Active/repair at day 7, n (%) | Notes on non-terminal outcomes |
|---|---|---|---|---|---|---|
| Activated charcoal (prospective series) | Germany (2024) | Protocol-adherent (incl. brood removal) | 15 | 15 (100%) | 0 (0.0%) | — |
| Activated charcoal (confirmation series) | Switzerland (2024/2025) | Protocol-adherent (incl. brood removal) | 25 | 25 (100%) | 0 (0.0%) | — |
| Activated charcoal (independent field test) | Switzerland (2025) | Protocol-adherent (incl. brood removal) | 105 | 102 (97.1%) | 3 (2.9%) | Small repair structures |
| Activated charcoal (pooled) | Germany + Switzerland (2024–2025) | Pooled protocol-adherent datasets | 145 | 142 (97.9%) | 3 (2.1%) | Repair and/or persistent activity |
| Activated charcoal (ancillary test) | Switzerland (2025) | Brood removal omitted (protocol deviation) | 3 | 0 (0.0%) | 3 (100%) | Flight activity and repair |
| Biocide dust (0.5% cypermethrin) | Germany (2024) | Operational comparator | 136 | 134 (98.5%) | 2 (1.5%) | Repair and/or emergency nest formation |
Notes: The day-7 endpoint was recorded as a binary outcome (inactive vs. active/repair). In the German prospective series, nest inactivity was additionally corroborated by absence of flight activity at day 10.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



