
Submitted:
10 February 2026
Posted:
11 February 2026
You are already at the latest version
Abstract
The growing demand for high-quality gluten-free bread requires innovative technological and functional approaches. This study investigates, for the first time, the use of Lacticaseibacillus paracasei SP5, in free and immobilized form on traditional Greek trahanas, for producing GF sourdough bread based on rice and buckwheat flour. Sourdough breads produced with free and immobilized cells displayed greater performance than the control sourdough bread in microbial stability and physicochemical properties, with the immobilized form showing the best results. In particular the application of immobilized L. paracasei SP5 led to gluten free sourdough bread with enhanced acidification (pH 4.55; TTA 12.9 mL) and remarkable lactic (2.20 g/kg) and acetic acid (0.76 g/kg) levels, extending the shelf-life to 7.0 days against mold and 7.5 days against rope spoilage, statistically significantly higher than the other two gluten free sourdough samples. It also showed the strongest antifungal activity and improved technological characteristics, including higher loaf volume (2.84 mL/g), greater height (5.50 cm), and lower baking loss (17.42%). Total phenolic content (87.3 mg GAE/100 g) and antioxidant capacity were also statistically significantly increased. Overall, immobilized L. paracasei SP5 on trahanas appears to be a promising clean-label strategy to improve quality, shelf-life, and functional value of GF bread.
Keywords:
Lacticaseibacillus paracasei SP5
; immobilization
; trahanas
; gluten free
; sourdough
; antifungal activity
1. Introduction
The demand for gluten-free bakery products has significantly increased in recent years, driven by the growing prevalence of celiac disease, non-celiac gluten sensitivity, and consumer interest in health-conscious diets [1]. However, gluten free products on the market are frequently nutritionally inadequate (lower micronutrients, protein, fiber) compared with their gluten-containing counterparts [2]. Gluten free breads face important technological challenges because the absence of gluten causes inferior dough rheology and structure, leading to reduced loaf volume, denser crumb, higher firmness and faster staling than wheat breads. These effects are linked to the high starch content and the lack of a viscoelastic protein network in gluten free doughs [3]. Likewise, incorporation of nutrient-dense flours (e.g., wholegrain pseudocereals) and other non-traditional functional ingredients has been widely explored, in order to tackle both nutritional and technological shortcomings [4]. Pseudocereal flours such as buckwheat and quinoa supply higher protein quality, dietary fiber, minerals, vitamins and antioxidant compounds, improving the nutritional and functional profile of gluten free breads; other novel ingredients (hemp, carob, seaweeds, fruit/vegetable flours) have also been evaluated. [5]. In addition, technological approaches that modify starch retrogradation and water binding are important to extend crumb softness and shelf life, because gluten free breads (especially starch-rich formulas) are susceptible to staling.
Sourdough fermentation has emerged as a key strategy to enhance GF bread quality, as it has the potential to improve dough handling, flavor and nutritional properties, increase protein digestibility and mineral bioavailability (via phytate hydrolysis) and generate exopolysaccharides and volatile compounds that positively affect texture, sensory profile and microbial stability [6]. Sourdoughs can be produced either spontaneously (native microbiota) or through inoculation with selected well defined starter cultures mainly lactic acid bacteria [7]. Spontaneous fermentations offer complex, authentic flavors but are less predictable and slower; inoculated/selected starters enable controlled, reproducible fermentation and lower contamination risk. However, starter cultures developed for wheat breads may not be optimal for gluten free pseudocereal flours and the selection of effective lactic acid bacteria adapted to gluten free matrices is important. [8]. Selected lactic acid bacteria strains (including strains of the Lacticaseibacillus paracasei group) have shown beneficial technological and functional traits for bread applications (acidification, enzyme activities, exopolysaccharides production, antifungal metabolites and flavor formation) [9]. On the other hand, immobilization of lactic acid bacteria on food matrices or solid supports is an established technique to enhance cell stability, viability and controlled metabolite release during processing and storage. Immobilized cells often show improved functional activity and protection compared with free cells [10]. Traditional food matrices and other supports have been used successfully as immobilization carriers [11]. Combining nutrient-rich flours (e.g., rice and buckwheat/quinoa) with sourdough fermentation using free or immobilized lactic acid bacteria may offer a practical route to overcome gluten free bread limitations [12]. Recently, a novel Lacticaseibacillus paracasei SP5 strain was successfully applied in wheat sourdough bread production in free and immobilized form in a starch gluten matrix (trahanas). The outcome showed nutritional improvements, upgraded sensorial features and higher resistance versus spoilage of the produced breads [13]. Therefore, the employment of this novel biocatalyst in gluten free bread production could be very interesting.
The aim of the present study was the assessment of free and immobilized L. paracaseiSP5 in gluten free sourdough breads formulated from rice and buckwheat flours. Various quality features of the produced sourdough breads were evaluated, and the feasibility of the proposed technology is addressed and discussed.
2. Materials and Methods
2.1. Raw Materials
Gluten free breads were prepared using commercially available rice flour (Oryza sativa L.) and buckwheat flour (Fagopyrum esculentum Moench) as the main cereal components. In particular, rice flour (white, medium-grain) and buckwheat flour (whole meal) were used as the primary flours for gluten free bread production. Rice flour contained approximately 80% starch, 6% protein, and 1.5% fiber, while buckwheat flour contained 72% starch, 11% protein, and 5.5% fiber. Water, salt (NaCl), and commercial baker’s yeast (Saccharomyces cerevisiae, Lesaffre, France) were used in all formulations. Trahanas, a traditional Greek fermented wheat-dairy product, was utilized as an immobilization matrix for L. paracasei SP5 cells. It ws produced by mixing 70% hard wheat flour with sour sheep’s milk, boiling and soaking in fresh sour milk. Afterwards it was dried and left to matured for 4 approximately 4 days at 30 °C. Then it was cut into ~1 cm3 cubes and sterilized prior to use to ensure the absence of contaminating microorganisms as described before [11].
2.2. Microbial Strain and Culture Conditions
The lactic acid bacteria L. paracasei SP5 was maintained at –80 °C in MRS broth supplemented with 20% glycerol. Prior to use, cultures were propagated twice in MRS broth at 37 °C for 18 h under anaerobic conditions. For the preparation of free-cell sourdough, the activated culture was harvested by centrifugation (5000×g, 10 min, 4 °C), washed twice with sterile saline (0.85% NaCl), and resuspended to a final concentration of 109 CFU/mL. Immobilization of cells took palace by mixing L. paracasei SP5 suspensions with trahanas powder at a ratio of 1:5 (w/w), followed by freeze-drying to obtain a stable immobilized starter as described before [11]. Specifically, freeze drying of either free cell of L. paracasei SP5 or immobilized on trahanas L. paracasei SP5 was conducted by freezing at −44 °C (5 °C/min) and drying for 48 h (at 5–15 mbar and −45 °C) with a FreeZone 4.5 Freeze-Drying System (Labconco, Kansas City, MO, USA)
2.3. Sourdough Preparation
Three gluten free sourdoughs were prepared by mixing rice and buckwheat flours. In particular, in all sourdough samples the following recipe was followed: 350g rice flour, 150g, buckwheat flour, and 90ml water. The two flours were combined in a 70:30 ratio (rice:buckwheat) to ensure sufficient starch for crumb softness and gas retention, while providing protein and fiber to improve dough structure and water retention. Free or immobilized L. paracasei SP5 were inoculated at 108 CFU/g of dough. The doughs were incubated at 30 °C for 24 h to allow fermentation. Sourdoughs were subsequently incorporated at 30% w/w into final bread formulations. For comparison, control dough without lactic acid bacteria inoculation were prepared under similar conditions and recipe.
2.4. Bread Preparation
The three sourdoughs were mixed in a concentration of 50% w/w with the remaining rice and buckwheat flour (proportion of 70/30 and total weight 100g), approximately 100ml water, salt (2% w/w), and baker’s yeast (1% w/w). Doughs were mixed for 10 min, proofed at 30 °C for 60 min, and baked at 180 °C for 35 min in a convection oven. Likewise, there sourdough gluten free breads were produced: containing: (i) free L. paracasei SP5 (SDGF1), (ii) immobilized L. paracasei SP5 (SDGF2) and (iii) without addition of L. paracasei SP5 (SDGF3). All sourdough bread samples were cooled at room temperature for 2 h prior to analysis.
2.5. Analytical Methods
2.5.1. Determination of Antimicrobial Activity
Antifungal activity of L. paracasei SP5 was assessed using the double-layer agar diffusion method. Overnight LAB cultures were centrifuged at 5000×g for 5 min, and the resulting cell-free supernatants (CFS) were neutralized to pH 6 with 0.1% NaOH to avoid inhibitory effects due to acidity. The CFS were also heat-treated at 120 °C for 15 min to evaluate the presence of heat-stable bacteriocins. Antifungal activity was tested against common bread spoilage molds, including Aspergillus niger, Penicillium roqueforti, and Fusarium oxysporum, by measuring inhibition zones after 48 h of incubation at 25 °C.
2.5.2. pH and Total Titratable Acidity (TTA)
The pH of all the gluten free sourdough breads was obtained by mixing 10 g sample with 90 ml distilled water and measuring the pH with a standard pH meter. Total titratable acidity values were obtained as described before [11].
2.5.3. Baking Loss
The baking loss of all gluten free sourdough breads was obtained from weight measurements taken before and after baking, according to the following formula:
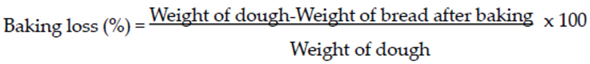
2.5.4. Specific Loaf Volume and Height
The specific volume of all the gluten free sourdough breads was measured using the rapeseed displacement method performed one hour after baking. The loaves were weighed and their specific volume was determined from the volume/mass ratio (mL/g). A pair of digital calipers was used to estimate loaf height [11].
2.5.6. Moisture Content
Moisture contents of breadcrumb of all the gluten free sourdough breads (crust and central crumb) were determined according to the AOAC Method 925.09, by heating at 98–100 °C in a partial vacuum, and reporting loss in weight as moisture [14].
2.5.7. Total Phenolic Content (TPC)
After baking, the 3 sourdough breads were left at room temperature for 3 h. to cool Afterwards, crumb samples were freeze-dried for 48 h using a FreeZone 4.5 lyophilizer (Labconco, Kansas City, MO, USA). Then the extraction procedure was followed according to:1 g of the freeze-dried crumb was suspended in 20 mL of phosphate-buffered saline (PBS, pH 7.4) and incubated under continuous shaking at 37 °C for 1 h. Subsequently, a second extraction was performed to the residue with further 20 mL of PBS. The supernatants were collected and stored at −20 °C until the determination of total phenolic content (TPC) and antioxidant capacity (AC).
Total phenolic content (TPC) was measured using the Folin–Ciocalteu reagent method as described previously [13]. Briefly, 200 μL of each bread crumb extract was mixed with 800 μL of Folin–Ciocalteu reagent and allowed to react in the dark for 2 min. Then, 2 mL of 7.5% (v/v) sodium carbonate solution were added, and the volume of the reaction was set to 10 mL with distilled water. The mixture was incubated for 60 min in the dark at room temperature. Finally, the absorbance was measured at 765 nm using a spectrophotometer. Standard gallic acid (GA) solutions and blanks were also prepared. TPC values were expressed as milligrams of gallic acid equivalents per 100 g of dried sample (mg GAE/100 g).
2.5.8. Antioxidant Capacity (AC)
The antioxidant capacity of the sourdough bread extracts was measured using 2 methods: (i) the ABTS [2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid)] radical cation assay, expressed as Trolox Equivalent Antioxidant Capacity (TEAC), and (ii) the DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging assay. Regarding the ABTS assay, the ABTS•⁺ stock solution was produced by mixing equal volumes of 7.4 mM ABTS and 2.6 mM potassium persulfate in water, followed by incubation for 12 h at room temperature. Afterwards, the solution was diluted with distilled water, in order to to achieve an absorbance of approximately 0.70 at 734 nm. Then, 100 μL of bread extract was mixed with 3.9 mL of the diluted ABTS•⁺ solution, incubated for 4 min. Finally, the absorbance was measured at 734 nm against a blank. Concerning the DPPH assay, the radical scavenging activity was determined by reacting the extracts with a DPPH solution, and the decrease in absorbance at 517 nm was recorded. Standard curves were prepared using Trolox and the results were expressed as milligrams of Trolox equivalents per 100 g of dried sample (mg TE/100 g) [13].
2.5.9. Mould and Rope Spoilage
Bread samples were stored under controlled conditions and evaluated for spoilage appearance macroscopically. Evaluation criteria for rope formation included the detection of ripe cantaloupe aroma and typical sticky crumb textures. Mould stability was assessed by monitoring the samples for visible surface colonization, with the shelf-life recorded as the duration (in days) preceding the first appearance of either bacterial or mould contamination. [13].
2.5.10. Statistical Analysis
Analysis of Variance (ANOVA) followed by Duncan’s post hoc multiple range test was applied to extract the specific differences between the various treatments. The analysis was performed using the SPSS Statistics 20.0 (IBM Corp., Armonk, NY, USA) software at an alpha level of 5%. randomly to prevent bias, and panelists were instructed to cleanse their palate with water between samples.
3. Results and Discussion
3.1. Antifungal Activity
The immobilized L. paracasei SP5 cells in trahanas exhibited significantly larger inhibition zones against Aspergillus niger, Penicillium roqueforti, and Fusarium culmorum compared with the free cell L. paracasei SP5 and none cell formulation (Table 1). These results indicate that immobilization not only enhances the viability and metabolic activity of the culture, but it is likely to increase the production or release of antifungal metabolites during fermentation and subsequent storage. The physical protection afforded by the trahanas matrix likely reduces stress on the cells, promoting more consistent synthesis of inhibitory compounds throughout the growth cycle.
This trend aligns with previous sourdough studies in which L. paracasei strains showed potent antagonism toward Fusarium and other spoilage fungi, with antifungal effects ascribed to the accumulation of organic acids and diverse low-molecular-weight metabolites including phenyllactic acid (PLA) and lactic acid [15]. Similar enhancement of bread shelf life were reported in sourdough prepared with specific lactic acid bacteria exhibiting antifungal properties, especially against A. niger and F. culmorum, when such cultures were used to partially replace calcium propionate [16]. In accordance to these observations, various lactic acid bacteria with antifungal properties were used in sourdough preparations and have been proposed to partially substitute calcium propionate in bread, extending shelf life and maintaining overall quality [16]. The enhanced antifungal activity reported in immobilized systems is also consistent with reports that such cultures produce a broader spectrum of volatile and non-volatile antifungal compounds such as cyclic dipeptides, medium-chain fatty acids, and specific fatty acid derivatives, which act synergistically to inhibit spore germination and hyphal growth in molds like A. niger and F. culmorum [17]. Recently, the application of immobilized L. paracasei SP5 in wheat sourdough breads showed that immobilization in prebiotic matrices prolongs resistance to mould and rope spoilage beyond 10 days, supporting the present findings [13]. This sustained bioprotective effect likely stems from improved cell activity and continuous release of antifungal metabolites from the immobilization carrier. This outcome is particularly valuable for gluten free breads, which are more susceptible to microbial spoilage, due to their higher water activity and starch content. Likewise, the significantly higher inhibition zones observed in the case of immobilized L. paracasei SP5 strongly suggest that trahanas-supported cultures can act as an effective natural bioprotective strategy, reducing reliance on synthetic preservatives and at the same time contributing to extended shelf life and product safety in gluten free bread applications. Such functional starter systems respond directly to increasing consumer demand for clean-label, additive-free products, which is an important consideration for both artisanal and industrial bakery markets.
3.2. pH and Total Titratable Acidity (TTA)
Th results obtained regarding the levels of pH and TTA of the produced gluten free sourdough breads are presented in Table 2. The noticeable decrease in pH and increase in total titratable acidity observed for sourdoughs fermented with L. paracasei SP5, particularly with immobilized cells, indicate a more intense and sustained acidification than the unfermented control, which is critical for both structure formation and microbial stability in gluten free bread systems. Similar ranges of final pH (4.5–5.0) and elevated total titratable acidity have been reported for gluten-free sourdoughs fermented with selected lactic acid bacteria, where stronger acidification correlated with improved crumb texture and delayed staling [18].
The incorporation of sourdough fermented with L. paracasei SP5 significantly optimized the chemical and functional parameters of the gluten free bread, with the immobilized cell system (SDGF2) demonstrating the most efficient metabolic activity. This enhanced performance is directly evidenced by the acidification parameters, where SDGF2 gluten free sourdough bread achieved the most significant decrease in pH to 4.55 and the highest total titratable acidity of 12.9 mL Sufficiently low pH and high total titratable acidity is essential to compensate for the lack of gluten by optimizing starch gelatinization, protein hydration and gas-holding capacity [19]. More recent gluten free models confirm that acidification in mixed cereal systems through lactic acid fermentation by lactic acid bacteria (e.g., teff- or rice-based sourdoughs) improves dough rheology and crumb softness over storage [20]. In rice-based gluten free breads, sourdough fermentation has been shown to lower pH, increase total titratable acidity and in parallel enhance specific volume and sensory attributes, underlining the central role of controlled LAB acidification for high-quality gluten free products [21]. The lower pH and higher total titratable acidity of sourdough breads fermented with immobilized L. paracasei SP5 (SDGF2) align with literature and probably support both the improved mould resistance and the technological performance of the resulting gluten free breads. Regarding the organic acid concentrations, SDGF2 gluten free sourdough bread reached the highest lactic acid level of 2.20 g/kg and acetic acid level of 0.76 g/kg, both significantly higher (p<0.05) than SDGF1 gluten free sourdough bread and control gluten free sourdough bread (SDGF3). Τhe concentration of lactic acid is technologically critical as it exerts a softening effect on the buckwheat protein matrix and promotes controlled starch swelling [22]. Simultaneously, the acetic acid concentration of 0.76 g/kg in SDGF2 plays a disproportionate role in biopreservation. Although present in lower quantities than lactic acid, acetic acid is the primary reason of the antifungal activity observed against spoilage organisms like Aspergillus niger [23]. At low pH levels, acetic acid penetrates fungal cell membranes more effectively, disrupting internal homeostasis and extending the mold-free shelf life of the bread. The results suggest that the immobilization of L. paracasei SP5 on the trahanas matrix provided a protective microenvironment that favored intense acid production. This biotechnological approach addresses the inherent challenges of buckwheat-based matrices, such as high buffering capacity and mineral-bound phytic acid. In particular, organic acids facilitate phytic acid breakdown, leading to greater bioavailability of essential mineral [24]. Overall, these results corroborate international studies demonstrating that, in gluten free formulations, targeted organic acid production through controlled lactic acid fermentation can successfully replace synthetic improvers and preservative [25].
3.3. Bread Height, Specific Volume and Baking Loss
The use of L. paracasei SP5, either free or immobilized in trahanas in gluten free sourdough bread production showed a significant impact on both the height and specific volume of gluten free rice–buckwheat sourdough breads (Table 3).
Among the treatments, sourdough bread fermented with immobilized L. paracasei SP5 showed the highest values of both height (5.50 cm) and specific volume (2.84 mL/g), whereas the control sourdough bread exhibited the lowest values (4.88 cm; 2.51 mL/g). Sourdough breads fermented with free L. paracasei SP5 showed intermediate values, indicating that immobilization in trahanas enhances the gas-holding capacity and structural stability of the dough.
These findings align with Alper (2024), who reported that sourdough addition softens gluten free doughs, promoting gas bubble expansion during fermentation and increasing loaf specific volume [26]. The efficient performance of sourdough produced with immobilized cells of L. paracasei SP5 may be attributed to enhanced metabolic activity of immobilized L. paracasei SP5, including exopolysaccharide production [11], which improves the viscosity and elasticity of the gluten free matrix. Moreover, the contribution of trahanas as an immobilization matrix can be attributed to its composition, which offers additional amino acids and fermentable substrates that further promote gas retention and loaf expansion[27]. The level of baking loss, was also significantly affected by the starter culture (Table 3). The lowest baking loss was observed in SDGF2 gluten free sourdough bread (17.42%), followed by SDGF1 gluten free sourdough bread (17.90%), while the control gluten free sourdough bread (SDGF3) exhibited the highest loss (18.55%). The reduced baking loss in sourdough breads, particularly those produced with free or immobilized L. paracasei SP5, can be attributed to improved moisture retention and matrix structure, consistent with previous reports in sourdough-based gluten free breads [28,29].
3.4. Moisture Content
The moisture content of both crumb and crust was significantly influenced by the starter culture and storage time (Figure 1). Immediately after baking, SDGF2 gluten free sourdough bread retained the highest crumb moisture, followed by SDGF1 gluten free sourdough bread and finally the control gluten free sourdough bread (SDGF3) suggesting improved water retention associated with immobilized L. paracasei SP5.
This behavior is consistent with previous observations in gluten free sourdough breads. Specifically, exopolysaccharides produced by specific lactic acid bacteria are known to bind water, delaying moisture loss and contributing to softer crumbs during storage [30]. SDGF1 gluten free sourdough bread also retained more moisture than the control, but less than the immobilized counterpart, indicating that immobilization enhances the functional activity of the strain in maintaining water content over time. These results are in agreement with Agirbasli et al. (2024), who reported that immobilized lactic acid bacteria improve hydration properties in gluten free bread matrices [31].
3.5. TPC and AA
Significant differences (p < 0.05) were recorded in the sourdough bread samples regarding the total phenolic content (TPC) and antioxidant activity (Table 4). Among the formulations, SDGF2 exhibited the highest TPC (87.3 mg GAE/100 g) and the highest ABTS (179.7 mg TE/100 g) and DPPH (3.8 µmol TE/g) values, indicating good antioxidant potential. SDGF1 gluten free sourdough bread showed intermediate values for all parameters, whereas SDGF3 gluten free sourdough bread presented the lowest TPC and antioxidant activity. Specifically, SDGF1 exhibited a total phenolic content of 75.9 mg GAE/100 g, with antioxidant activity values of 154.1 mg TE/100 g (ABTS) and 3.0 µmol TE/g (DPPH). In contrast, SDGF3 showed lower values, presenting a TPC of 57.1 mg GAE/100 g, ABTS activity of 121.5 mg TE/100 g, and DPPH activity of 2.5 µmol TE/g. The significant elevation of TPC and AA observed in the SDGF2 gluten free sourdough bread can be attributed to the complex metabolic flux of L. paracasei SP5 within the rice-buckwheat matrix. In cereal-based systems, a vast majority of phenolic compounds are non-extractable, because they are covalently bonded to cell wall polysaccharides via ester or ether bonds. The fermentation process, particularly when catalyzed by a high-density immobilized culture, facilitates the synthesis of microbial enzymes such as various esterases various glycosidases [32]. These enzymes act as biochemical catalysts that hydrolyze the ester-linked phenolic fragments and facilitate the liberation of them from the dietary fiber complex of the buckwheat and rice flours. This enzymatic liberation increases the concentration of free phenolic acids, such as ferulic, p-coumaric and caffeic acids, which directly correlates with the higher GAE values reported [33]. Furthermore, the higher antioxidant capacity measured in the SDGF2 gluten free sourdough bread reflects a synergistic relationship between microbial acidification and phytochemical stability. The intense acidification creates an optimal environment for the stabilization of buckwheat’s native flavonoids, such as rutin and quercetin, which are otherwise prone to oxidative degradation during the thermal processing of baking [34]. The immobilization of L. paracasei SP5 on the trahanas matrix provides a protective micro-environment that maintains higher metabolic activity and cell viability compared to free L. paracasei SP5 cells. This sustained activity ensures a continuous release of metabolites throughout the fermentation stages, leading to a more thorough transformation of the dough’s chemical profile [35]. Studies have shown that such targeted lactic acid fermentation not only increases the bioaccessibility of these compounds but also promotes the conversion of phenolic acids into more potent antioxidant derivatives, such as vinyl derivatives or dihydro-derivatives, through microbial decarboxylation and reductase activities [36]. The discrepancy between the control gluten free sourdough bread (SDGF3) and the inoculated samples further underscores the role of lactic acid bacteria as bio-refineries. While the control sample relies solely on the endogenous enzymes of the flours and yeast, the SDGF2 gluten free sourdough bread benefits from the specialized metabolic pathways of the L. paracasei SP5 strain, which are amplified by the carrier’s nutrient-rich dairy-wheat composition. The trahanas carrier likely contributes additional nitrogenous sources and fermentable carbohydrates that fuel the production of bioactive exopolysaccharides (EPS) [13]. These EPS molecules can act as secondary antioxidants by scavenging reactive oxygen species, thereby contributing to the overall antioxidant capacity [13]. Consequently, the use of immobilized L. paracasei SP5 represents a superior technological strategy for enhancing the functional and nutraceutical profile of gluten-free matrices, transforming them from simple starch-based products into bioactive-rich functional foods.
3.6. Shelf Life
Gluten-free breads are highly perishable and susceptible to spoilage, with visible fungal growth and rope development generally appearing aproximately within 3 to 5 days when produced without a bioprotective agent [37]. The increased microbial stability observed in the SDGF2 gluten free sourdough bread, which preserved for 7.0 days regarding mould spoilage and for 7.5 days regarding rope spoilage is closely associated with its efficient acidification profile (Table 2). This is evidenced by a pH value of 4.55 and a total titratable acidity (TTA) of 12.9 mL, representing the highest acidity levels among all tested gluten free sourdough breads. In addition, the elevated lactic acid content in SDGF2 gluten free sourdough bread plays a key role in preservation, as it works in conjunction with acetic acid to suppress spoilage microorganisms through intracellular acidification, particularly when the low pH favors the undissociated state of these organic acids [23]. These chemical conditions provided an effective environment explaining the extension of its shelf-life against the proliferation of fungi and Bacillus species [38]. SDGF1 gluten free sourdough bread successfully extended the shelf-life to 6.0 days for both mould and rope resistance. This is a notable improvement over the 4.0–4.5 days observed in the control SDGF3 gluten free sourdough bread, confirming that the presence of L. paracasei SP5 even in free form induces enough acidification to inhibit common spoilage organisms. However, the one-day difference between SDGF1 and SDGF2 gluten free sourdough breads is statistically significant and it demonstrates that the “free” environment does not protect the bacteria as effectively against the rising acidity or thermal stress of baking, potentially leading to a slightly lower accumulation of protective metabolites toward the end of the proofing stage.
In contrast, the lower metabolic output in the control SDGF3 gluten free sourdough bread, with lactic and acetic acid levels of only 1.13 g/kg and 0.49 g/kg respectively, resulted in earlier spoilage within 4.0–4.5 days. These findings suggest that the immobilization of L. paracasei SP5 on the trahanas carrier optimizes fermentation kinetics, facilitating a metabolite profile that improves both the technological and bioprotective properties of gluten-free sourdough bread [13,33].

Table 5.
Appearance of rope and mould spoilage in the produced gluten free sourdough breads.
| Bread sample | Mould spoilage | Rope spoilage |
|---|---|---|
| days | ||
| SDGF1 | 6.0 ± 0.5ᵇ | 6.0 ± 0.5ᵇ |
| SDGF2 | 7.0 ± 0.5ᵃ | 7.5 ± 0.5b |
| SDGF3 | 4.0 ± 0.5ᶜ | 4.5 ± 0.5c |
Different superscript letters in a column indicate statistically significant differences (ANOVA, Duncan’s multiple range test, p<0.05.
4. Challenges and Perspectives
The application of Lacticaseibacillus paracasei SP5, particularly in immobilized form, demonstrates considerable potential for advancing the production of high quality and functionally enhanced gluten free sourdough bread. The technological, microbiological and functional improvements observed in this study highlight the feasibility of this biotechnological approach and support its relevance for clean-label bakery innovation. Nevertheless, further optimization is required to facilitate large-scale industrial implementation.
A major challenge involves the industrial scale-up of immobilized starter systems, which necessitates maintaining homogeneous cell distribution, consistent metabolic activity, and structural integrity of the carrier throughout large-scale dough mixing, fermentation, and baking. Continued progress in carrier design, freeze-drying, and stabilization technologies is expected to enhance reproducibility, improve storage stability, and ensure reliable and consistent performance under industrial processing conditions.
In addition, the inherent structural limitations of gluten-free doughs, including reduced viscoelasticity and limited gas retention capacity, remain critical technological constraints. These limitations could be further addressed through optimized fermentation kinetics, hydration strategies, and synergistic formulation approaches combining fermentation with structuring agents. The enhanced antifungal activity, improved loaf volume and extended shelf life achieved with immobilized L. paracasei SP5 are fully consistent with current consumer demand for preservative free and clean label bakery products, highlighting the industrial relevance of this approach. Likewise, future research should be focused on refining immobilization methodologies, including advanced encapsulation or structured carrier systems, in order to enhance scalability, process control, and starter performance consistency. Overall, L. paracasei SP5 seems to be a multifunctional and commercially promising starter culture capable of supporting the development of next generation gluten free breads with improved technological performance, stability and functional value.
5. Conclusions
The present study highlights the potential of L. paracasei SP5, particularly when immobilized in traditional trahanas, as a natural and effective strategy for the ameriolation of gluten free sourdough bread quality. The use of Lacticaseibacillus paracasei SP5 as a sourdough starter markedly improved the overall quality, stability, and functional value of gluten-free sourdough bread. In particular, gluten free sourdough bread produced with free and immobilized L. paracasei SP5 on trahanas exhibited intensified acidification and the highest concentrations of lactic and acetic acids. These outcomes led into enhanced microbial stability, extending in the highest levels the shelf life of the sourdough bread against mould and rope spoilage compared. In parallel, significant technological improvements were recorded, including higher loaf volume and height, reduced baking loss, and improved moisture retention. The immobilized formulation also exhibited the highest functional potential, with markedly increased total phenolic content and antioxidant activity, demonstrating the capacity of targeted sourdough fermentation to enhance both technological and functional properties of gluten-free bread.
Future research should explore sensory characteristics and underlying biochemical mechanisms to further refine the functional and organoleptic properties of gluten-free breads and enable their commercial exploitation.
Author Contributions
Conceptualization, I.M. and S.P.; methodology, S.P.; I.M. and A.B.; software, S.P. and A.B.; validation, S.P. and I.M.; investigation, I.M. and S.P; resources, S.P. and I.M.; data curation, S.P. and I.M.; writing—original draft preparation, S.P.; writing—review and editing, S.P.; supervision, S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Haixi, L.; Ahmed, S.; Shahbaz, M.; Abid, J.; Jahangir, M.; Khan, S. Quality Improvement in Gluten-Free Bread: A Comprehensive Review of Modern Techniques and Ingredients. Food Reviews International 2025, 1–35. [CrossRef]
- Melini, V.; Melini, F. Gluten-Free Diet: Gaps and Needs for a Healthier Diet. Nutrients 2019, 11, 170.
- Sciarini, L.S.; Ribotta, P.D.; León, A.E.; Pérez, G.T. Influence of Gluten-Free Flours and Their Mixtures on Batter Properties and Bread Quality. Food Bioprocess Technol 2010, 3, 577–585. [CrossRef]
- Usman, M.; Patil, P.; Ray, A.; Karrar, E.; Xu, M. From Tradition to Innovation: Enhancing the Nutritional and Functional Quality of Chapati Through Modern Techniques. Food Reviews International 2025, 1–39. [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E.; Hernández-Ledesma, B. Pseudocereal Grains: Nutritional Value, Health Benefits and Current Applications for the Development of Gluten-Free Foods. Food and chemical toxicology 2020, 137, 111178. [CrossRef]
- Gobbetti, M.; Minervini, F.; Pontonio, E.; Di Cagno, R.; De Angelis, M. Drivers for the Establishment and Composition of the Sourdough Lactic Acid Bacteria Biota. International Journal of Food Microbiology 2016, 239, 3–18. [CrossRef]
- Oleinikova, Y.; Amangeldi, A.; Zhaksylyk, A.; Saubenova, M.; Sadanov, A. Sourdough Microbiota for Improving Bread Preservation and Safety: Main Directions and New Strategies. Foods 2025, 14, 2443. [CrossRef]
- Landis, E.A.; Oliverio, A.M.; McKenney, E.A.; Nichols, L.M.; Kfoury, N.; Biango-Daniels, M.; Shell, L.K.; Madden, A.A.; Shapiro, L.; Sakunala, S. The Diversity and Function of Sourdough Starter Microbiomes. elife 2021, 10, e61644. [CrossRef]
- Kiousi, D.E.; Efstathiou, C.; Tegopoulos, K.; Mantzourani, I.; Alexopoulos, A.; Plessas, S.; Kolovos, P.; Koffa, M.; Galanis, A. Genomic Insight into Lacticaseibacillus Paracasei SP5, Reveals Genes and Gene Clusters of Probiotic Interest and Biotechnological Potential. Frontiers in microbiology 2022, 13, 922689. [CrossRef]
- Lapponi, M.J.; Méndez, M.B.; Trelles, J.A.; Rivero, C.W. Cell Immobilization Strategies for Biotransformations. Current Opinion in Green and Sustainable Chemistry 2022, 33, 100565. [CrossRef]
- Kazakos, S.; Mantzourani, I.; Plessas, S. Quality Characteristics of Novel Sourdough Breads Made with Functional Lacticaseibacillus Paracasei SP5 and Prebiotic Food Matrices. Foods 2022, 11, 3226. [CrossRef]
- Dan, H.; Li, H.; Li, C.; Fang, Z.; Hu, B.; Chen, H.; Wang, C.; Chen, S.; Hui, T.; Wu, W.; et al. Application of Sourdough in Gluten-Free Bakery Products. Critical Reviews in Food Science and Nutrition 2025, 65, 3048–3068. [CrossRef]
- Kazakos, S.; Bampali, I.; Mantzourani, I.; Plessas, S. Cereal-Based Derivatives as Functional Fortifiers of Sourdough Bread: Nutritional, Antioxidant and Quality Insights. Applied Sciences 2026, 16, 646. [CrossRef]
- Plessas, S.; Pherson, L.; Bekatorou, A.; Nigam, P.; Koutinas, A.A. Bread Making Using Kefir Grains as Baker’s Yeast. Food chemistry 2005, 93, 585–589. [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2019, 8, 64. [CrossRef]
- Ryan, L.A.M.; Dal Bello, F.; Arendt, E.K. The Use of Sourdough Fermented by Antifungal LAB to Reduce the Amount of Calcium Propionate in Bread. International Journal of Food Microbiology 2008, 125, 274–278. [CrossRef]
- Banicod, R.J.S.; Tabassum, N.; Javaid, A.; Kim, Y.-M.; Khan, F. Lactic Acid Bacteria–Derived Secondary Metabolites: Emerging Natural Alternatives for Food Preservation. Probiotics & Antimicro. Prot. 2025. [CrossRef]
- Moore, M.M.; Juga, B.; Schober, T.J.; Arendt, E.K. Effect of Lactic Acid Bacteria on Properties of Gluten-Free Sourdoughs, Batters, and Quality and Ultrastructure of Gluten-Free Bread. Cereal Chem 2007, 84, 357–364. [CrossRef]
- Moroni, A.V.; Dal Bello, F.; Arendt, E.K. Sourdough in Gluten-Free Bread-Making: An Ancient Technology to Solve a Novel Issue? Food microbiology 2009, 26, 676–684. [CrossRef]
- Chochkov, R.; Savova-Stoyanova, D.; Papageorgiou, M.; Rocha, J.M.; Gotcheva, V.; Angelov, A. Effects of Teff-Based Sourdoughs on Dough Rheology and Gluten-Free Bread Quality. Foods 2022, 11, 1012. [CrossRef]
- Seyedahmadi, S.; Gharekhani, M.; Tariverdi, S.; Bakhshabadi, H. Enhancing the Quality of Rice-Based Gluten-Free Bread Using Sourdoughs Fermented with Lactobacillus Fermentum and Lactobacillus Plantarum. Scientific Reports 2025, 15, 26543. [CrossRef]
- Diowksz, A.; Sadowska, A. Impact of Sourdough and Transglutaminase on Gluten-Free Buckwheat Bread Quality. Food Bioscience 2021, 43, 101309. [CrossRef]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Gänzle, M. Exploiting Synergies of Sourdough and Antifungal Organic Acids to Delay Fungal Spoilage of Bread. International journal of food microbiology 2019, 302, 8–14.
- Castro-Alba, V. Fermentation of Quinoa, Canihua and Amaranth to Degrade Phytate and Improve Mineral Bioavailability. 2019.
- Nascimento, A.P.S.; Barros, A.N. Sustainable Innovations in Food Microbiology: Fermentation, Biocontrol, and Functional Foods. Foods 2025, 14, 2320. [CrossRef]
- Alper, A.; Altan, A. Recent Advances in the Use of Sourdough Fermentation to Improve the Quality of Gluten-Free Bakery Products. In Sourdough Microbiota and Starter Cultures for Industry; Ceresino, E.B., Juodeikiene, G., Miescher Schwenninger, S., Ferreira Da Rocha, J.M., Eds.; Springer International Publishing: Cham, 2024; pp. 461–492 ISBN 978-3-031-48603-6.
- Özdestan, Ö.; Üren, A. Biogenic Amine Content of Tarhana: A Traditional Fermented Food. International Journal of Food Properties 2013, 16, 416–428. [CrossRef]
- Agirbasli, Z.; Elvan, M.; Harsa, S. Immobilization Techniques Applicable to Sourdough. In Sourdough Microbiota and Starter Cultures for Industry; Ceresino, E.B., Juodeikiene, G., Miescher Schwenninger, S., Ferreira Da Rocha, J.M., Eds.; Springer International Publishing: Cham, 2024; pp. 257–279 ISBN 978-3-031-48603-6.
- Cappa, C.; Lucisano, M.; Raineri, A.; Fongaro, L.; Foschino, R.; Mariotti, M. Gluten-Free Bread: Influence of Sourdough and Compressed Yeast on Proofing and Baking Properties. Foods 2016, 5, 69. [CrossRef]
- Lynch, K.M.; Coffey, A.; Arendt, E.K. Exopolysaccharide Producing Lactic Acid Bacteria: Their Techno-Functional Role and Potential Application in Gluten-Free Bread Products. Food research international 2018, 110, 52–61.
- Agirbasli, Z.; Elvan, M.; Harsa, S. Immobilization Techniques Applicable to Sourdough. In Sourdough Microbiota and Starter Cultures for Industry; Ceresino, E.B., Juodeikiene, G., Miescher Schwenninger, S., Ferreira Da Rocha, J.M., Eds.; Springer International Publishing: Cham, 2024; pp. 257–279 ISBN 978-3-031-48603-6.
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected Lactic Acid Bacteria Synthesize Antioxidant Peptides during Sourdough Fermentation of Cereal Flours. Appl Environ Microbiol 2012, 78, 1087–1096. [CrossRef]
- Gänzle, M.G. Enzymatic and Bacterial Conversions during Sourdough Fermentation. Food microbiology 2014, 37, 2–10. [CrossRef]
- Wu, X.; Fu, G.; Li, R.; Li, Y.; Dong, B.; Liu, C. Effect of Thermal Processing for Rutin Preservation on the Properties of Phenolics & Starch in Tartary Buckwheat Achenes. International Journal of Biological Macromolecules 2020, 164, 1275–1283. [CrossRef]
- Gabriele, M.; Arouna, N.; Árvay, J.; Longo, V.; Pucci, L. Sourdough Fermentation Improves the Antioxidant, Antihypertensive, and Anti-Inflammatory Properties of Triticum Dicoccum. International Journal of Molecular Sciences 2023, 24, 6283. [CrossRef]
- Dapčević-Hadna\djev, T.; Stupar, A.; Stevanović, D.; Škrobot, D.; Maravić, N.; Tomić, J.; Hadna\djev, M. Ancient Wheat Varieties and Sourdough Fermentation as a Tool to Increase Bioaccessibility of Phenolics and Antioxidant Capacity of Bread. Foods 2022, 11, 3985. [CrossRef]
- Moroni, A.V.; Dal Bello, F.; Arendt, E.K. Sourdough in Gluten-Free Bread-Making: An Ancient Technology to Solve a Novel Issue? Food microbiology 2009, 26, 676–684. [CrossRef]
- Luz, C.; D’Opazo, V.; Mañes, J.; Meca, G. Antifungal Activity and Shelf Life Extension of Loaf Bread Produced with Sourdough Fermented by Lactobacillus Strains. J Food Process Preserv 2019, 43. [CrossRef]
Figure 1.
Crumb and crust moisture content (%) of the produced gluten free sourdough breads at 0 and 24 h storage.
Figure 1.
Crumb and crust moisture content (%) of the produced gluten free sourdough breads at 0 and 24 h storage.
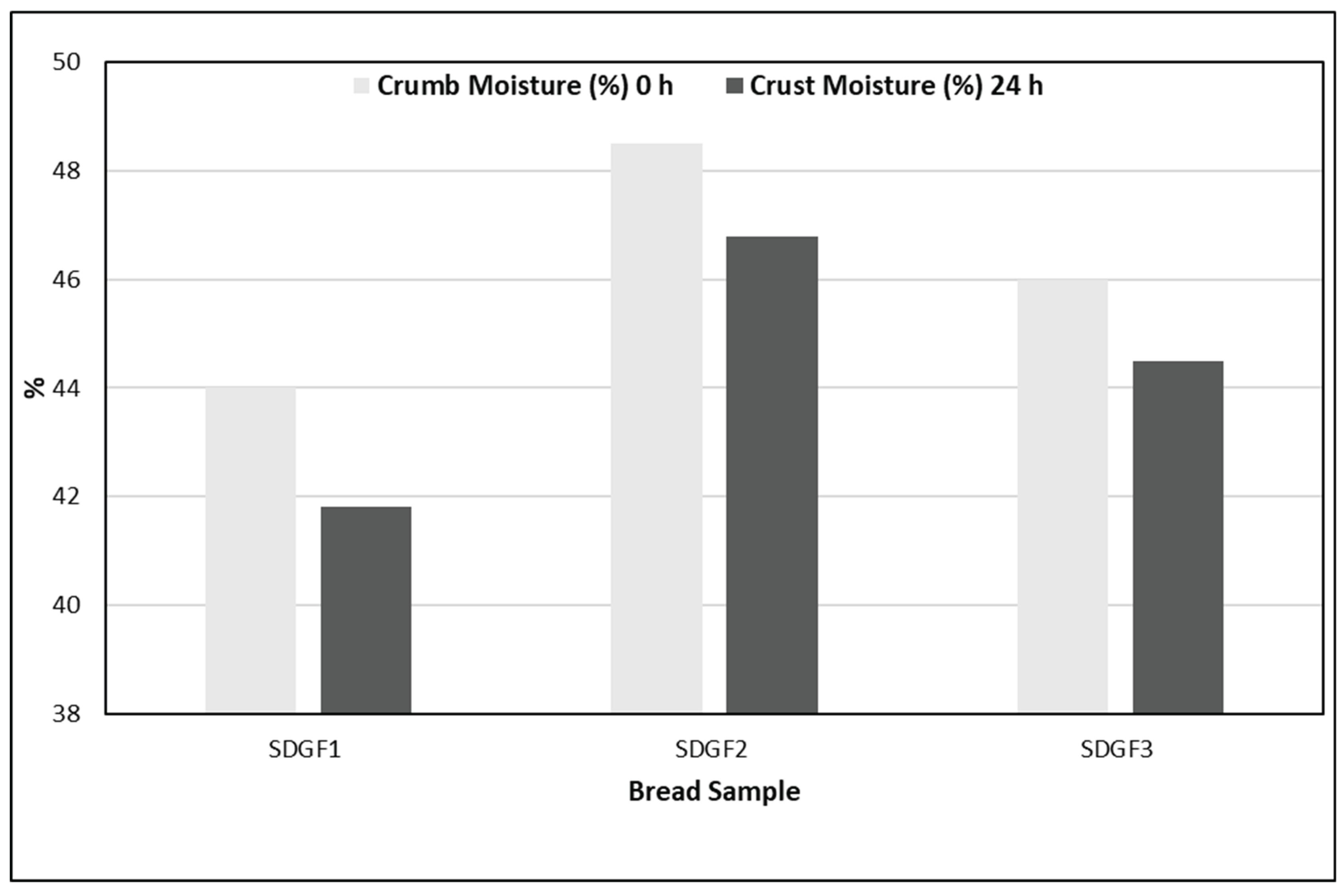
Table 1.
Antifungal Activity.
| Sample | Inhibition zone (mm) | ||
|---|---|---|---|
| A. niger | P. roqueforti | F. culmorum | |
| Control (no LAB) | 0 | 0 | 0 |
| Free cells | 8.2 ± 0.3b | 7.5 ± 0.4b | 7.0 ± 0.3b |
| Immobilized cells | 12.5 ± 0.4a | 11.8 ± 0.3a | 11.0 ± 0.4a |
Different superscript letters in a column indicate statistically significant differences (p<0.05).
Table 2.
pH total titratable acidity and organic acid levels of the produced gluten free sourdough breads.
Table 2.
pH total titratable acidity and organic acid levels of the produced gluten free sourdough breads.
| Bread Sample | pH | TTA | Lactic acid (g/kg) | Acetic acid |
|---|---|---|---|---|
| (mL) | (g/kg) | |||
| SDGF1 | 4.80 ± 0.02b | 11.4 ± 0.5b | 1.45± 0.07b | 0.62± 0.02b |
| SDGF2 | 4.55 ± 0.01c | 12.9 ± 0.3a | 2.20± 0.08a | 0.76± 0.05a |
| SDGF3 | 6.00 ± 0.02a | 6.4 ± 0.4c | 1.13± 0.15c | 0.49± 0.03c |
Different superscript letters in a column indicate statistically significant differences (p<0.05).
Table 3.
Levels of height, specific volume and baking loss of the produced gluten free sourdough breads.
Table 3.
Levels of height, specific volume and baking loss of the produced gluten free sourdough breads.
| Bread Sample | Height (cm) |
Specific Volume (mL/g) | Baking Loss (%) |
|---|---|---|---|
| SDGF1 | 5.35 ± 0.10ᵃ | 2.79 ± 0.06ᵃ | 17.90 ± 0.30a |
| SDGF2 | 5.50 ± 0.08ᵃ | 2.84 ± 0.04ᵃ | 17.42 ± 0.12a |
| SDGF3 | 4.88 ± 0.12b | 2.51 ± 0.09b | 18.55 ± 0.40a |
Different superscript letters in a column indicate statistically significant differences (p<0.05).
Table 4.
Total phenolic content (TPC) and antioxidant activity (AA) (on dry weight basis) of the produced gluten free sourdough breads.
Table 4.
Total phenolic content (TPC) and antioxidant activity (AA) (on dry weight basis) of the produced gluten free sourdough breads.
| Bread Sample | TPC (mg GAE/100 g) | ABTS (mg TE/100 g) | DPPH (µmol TE/g) |
|---|---|---|---|
| SDGF1 | 75.9 ± 3.4b | 154.1 ± 4.8b | 3.0 ± 0.2b |
| SDGF2 | 87.3 ± 2.2a | 179.7 ± 2.8a | 3.8 ± 0.1a |
| SDGF3 | 57.1 ± 1.5c | 121.5 ± 1.3c | 2.5 ± 0.1c |
Different superscript letters in a column indicate statistically significant differences (ANOVA, Duncan’s multiple range test, p<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



