
Submitted:
09 February 2026
Posted:
12 February 2026
You are already at the latest version
Abstract
Biofilm is a complex microbial community that provides protection to bacterial cells against various stress conditions, including harsh environments, antimicrobial treatments, and host immune responses. This protective mechanism enhances Campylobacter survival during food processing and storage and promotes transmission to humans. Despite its significance, the molecular mechanisms underlying Campylobacter biofilm formation and its impact on pathogen persistence remain poorly understood. In this study, we characterized the biofilm-forming ability of 18 C. jejuni and C. coli strains isolated from retail meat and performed whole-genome sequencing and comparative genomic analysis to identify strain-specific genes contributing to biofilm formation and maintenance. Phenotypic analysis revealed that C. jejuni strains YH001 and YH027 exhibited the strongest biofilm-forming capacity, producing the highest biomass among all isolates. Phylogenetic analysis indicated a close genetic relationship between these two strains, while pangenome analysis identified 19 unique genes/proteins specific to these strains. Functional annotation indicated their critical roles in adhesion, extracellular matrix production, and stress response. These findings demonstrate strain-specific biofilm formation in Campylobacter and highlight genetic determinants that may serve as targets for novel therapeutic approaches and intervention strategies to disrupt biofilms, improve food safety, and reduce persistent infections.
Keywords:
Introduction
Materials and Methods
Sample Preparation
Biofilm Formation
Biofilm Quantification
Genome Sequencing, Assembly, and Annotation
Pangenome & Phylogenetic Analysis
Results and Discussion
1. Determination of Biofilm-Forming Ability of C. jejuni and C. coli Food Isolates
2. Comparison Between Genetic Relatedness and Biofilm Formation of Campylobacter Isolates
3. Genetic Traits Associated with Biofilm Formation and Stability
Identification of Strain-Specific Genes Associated with Biofilm Formation
Conclusion
Supplementary Materials
Author Contributions
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. About Campylobacter Infection. Available online: https://www.cdc.gov/campylobacter/about/index.html (accessed on 28 January 2026).
- Tikhomirova, A.; McNabb, E.R.; Petterlin, L.; Bellamy, G.L.; Lin, K.H.; Santoso, C.A.; Daye, E.S.; Alhaddad, F.M.; Lee, K.P.; Roujeinikova, A. Campylobacter jejuni virulence factors: Update on emerging issues and trends. J. Biomed. Sci. 2024, 31, 45. [Google Scholar] [CrossRef]
- World Health Organization (WHO) Fact Sheet on Campylobacter. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter.
- Veronese, P.; Dodi, I. Campylobacter jejuni/coli Infection: Is It Still a Concern? Microorganisms 2024, 12, 2669. [Google Scholar] [CrossRef] [PubMed]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of polysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef]
- Lasa, I.; Penades, J.R. Bap: A family of surface proteins involved in biofilm formation. Res. Microbiol. 2006, 157, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.N.C.; Yildiz, F.H. Biofilm Matrix Proteins. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Panlilio, H.; Rice, C.V. The role of extracellular DNA in the formation, architecture, stability, and treatment of bacterial biofilms. Biotechnol. Bioeng. 2021, 118, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial Biofilms and Their Implications in Pathogenesis and Food Safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef]
- Buswell, C.M.; Herlihy, Y.M.; Lawrence, L.M.; McGuiggan, J.T.; Marsh, P.D.; Keevil, C.W.; Leach, S.A. Extended survival and persistence of Campylobacter spp. in water and aquatic biofilms and their detection by immunofluorescent-antibody and -rRNA staining. Appl. Environ. Microbiol. 1998, 64, 733–741. [Google Scholar] [CrossRef]
- Teh, A.H.; Lee, S.M.; Dykes, G.A. Does Campylobacter jejuni form biofilms in food-related environments? Appl. Environ. Microbiol. 2014, 80, 5154–5160. [Google Scholar] [CrossRef]
- Reeser, R.J.; Medler, R.T.; Billington, S.J.; Jost, B.H.; Joens, L.A. Characterization of Campylobacter jejuni biofilms under defined growth conditions. Appl. Environ. Microbiol. 2007, 73, 1908–1913. [Google Scholar] [CrossRef]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of mono- and mixed-culture Campylobacter jejuni biofilms. Appl. Environ. Microbiol. 2012, 78, 1033–1038. [Google Scholar] [CrossRef]
- Melo, R.T.; Mendonca, E.P.; Monteiro, G.P.; Siqueira, M.C.; Pereira, C.B.; Peres, P.; Fernandez, H.; Rossi, D.A. Intrinsic and Extrinsic Aspects on Campylobacter jejuni Biofilms. Front. Microbiol. 2017, 8, 1332. [Google Scholar] [CrossRef]
- Silha, D.; Sirotkova, S.; Svarcova, K.; Hofmeisterova, L.; Korycanova, K.; Silhova, L. Biofilm Formation Ability of Arcobacter-like and Campylobacter Strains under Different Conditions and on Food Processing Materials. Microorganisms 2021, 9, 2017. [Google Scholar] [CrossRef]
- Puning, C.; Su, Y.; Lu, X.; Golz, G. Molecular Mechanisms of Campylobacter Biofilm Formation and Quorum Sensing. Curr. Top. Microbiol. Immunol. 2021, 431, 293–319. [Google Scholar]
- Korkus, J.; Salata, P.; Thompson, S.A.; Paluch, E.; Bania, J.; Walecka-Zacharska, E. The role of cydB gene in the biofilm formation by Campylobacter jejuni. Sci. Rep. 2024, 14, 26574. [Google Scholar] [CrossRef]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef]
- He, Y.; Capobianco, J.; Armstrong, C.M.; Chen, C.Y.; Counihan, K.; Lee, J.; Reed, S.; Tilman, S. Detection and Isolation of Campylobacter spp. from Raw Meat. J. Vis. Exp. 2024. [Google Scholar] [CrossRef] [PubMed]
- He, Y.P.; Yao, X.M.; Gunther, N.W.; Xie, Y.P.; Tu, S.I.; Shi, X.M. Simultaneous Detection and Differentiation of Campylobacter jejuni, C. coli, and C. lari in Chickens Using a Multiplex Real-Time PCR Assay. Food Anal. Method. 2010, 3, 321–329. [Google Scholar] [CrossRef]
- He, Y.; Yan, X.; Reed, S.; Xie, Y.; Chen, C.Y.; Irwin, P. Complete Genome Sequence of Campylobacter jejuni YH001 from Beef Liver, Which Contains a Novel Plasmid. Genome Announc. 2015, 3, e01492-14. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Kanrar, S.; Reed, S.; Lee, J.; Capobianco, J. Whole Genome Sequences, De Novo Assembly, and Annotation of Antibiotic Resistant Campylobacter jejuni Strains S27, S33, and S36 Newly Isolated from Chicken Meat. Microorganisms 2024, 12, 159. [Google Scholar] [CrossRef]
- He, Y.; Reed, S.; Yan, X.; Zhang, D.; Strobaugh, T.; Capobianco, J.; Gehring, A. Complete Genome Sequences of Multidrug-Resistant Campylobacter coli Strains YH501, YH503, and YH504, from Retail Chicken. Microbiol. Resour. Announc. 2022, 11, e0023722. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.; He, Y.; Reed, S.; Strobaugh, T., Jr.; Irwin, P. Whole genome sequencing and analysis of Campylobacter coli YH502 from retail chicken reveals a plasmid-borne type VI secretion system. Genom. Data 2017, 11, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Silva, N.D.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206-14. [Google Scholar] [CrossRef]
- Arkin, A.P.; Cottingham, R.W.; Henry, C.S.; Harris, N.L.; Stevens, R.L.; Maslov, S.; Dehal, P.; Ware, D.; Perez, F.; Canon, S.; et al. KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef]
- Hall, B.G.; Nisbet, J. Building Phylogenetic Trees From Genome Sequences With kSNP4. Mol. Biol. Evol. 2023, 40, msad235. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Moore, R.M.; Harrison, A.O.; McAllister, S.M.; Polson, S.W.; Wommack, K.E. Iroki: Automatic customization and visualization of phylogenetic trees. PeerJ 2020, 8, e8584. [Google Scholar] [CrossRef]
- Ginestet, C. ggplot2: Elegant Graphics for Data Analysis. J. Roy. Stat. Soc. A 2011, 174, 245. [Google Scholar] [CrossRef]
- Sulaeman, S.; Le Bihan, G.; Rossero, A.; Federighi, M.; De, E.; Tresse, O. Comparison between the biofilm initiation of Campylobacter jejuni and Campylobacter coli strains to an inert surface using BioFilm Ring Test. J. Appl. Microbiol. 2010, 108, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef]
- Flemming, H.C.; van Hullebusch, E.D.; Little, B.J.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. Microbial extracellular polymeric substances in the environment, technology and medicine. Nat. Rev. Microbiol. 2025, 23, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Selles Vidal, L.; Kelly, C.L.; Mordaka, P.M.; Heap, J.T. Review of NAD(P)H-dependent oxidoreductases: Properties, engineering and application. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 327–347. [Google Scholar] [CrossRef]
- Pal, S.; Jain, D.; Biswal, S.; Rastogi, S.K.; Kumar, G.; Ghosh, A.S. The physiological role of Acinetobacter baumannii DacC is exerted through influencing cell shape, biofilm formation, the fitness of survival, and manifesting DD-carboxypeptidase and beta-lactamase dual-enzyme activities. FEMS Microbiol. Lett. 2024, 371, fnae079. [Google Scholar] [CrossRef]
- Peters, K.; Kannan, S.; Rao, V.A.; Biboy, J.; Vollmer, D.; Erickson, S.W.; Lewis, R.J.; Young, K.D.; Vollmer, W. The Redundancy of Peptidoglycan Carboxypeptidases Ensures Robust Cell Shape Maintenance in Escherichia coli. mBio 2016, 7, e00819-16. [Google Scholar] [CrossRef]
- Iwata, T.; Watanabe, A.; Kusumoto, M.; Akiba, M. Peptidoglycan Acetylation of Campylobacter jejuni Is Essential for Maintaining Cell Wall Integrity and Colonization in Chicken Intestines. Appl. Environ. Microbiol. 2016, 82, 6284–6290. [Google Scholar] [CrossRef]
- Das, T.; Sehar, S.; Manefield, M. The roles of extracellular DNA in the structural integrity of extracellular polymeric substance and bacterial biofilm development. Environ. Microbiol. Rep. 2013, 5, 778–786. [Google Scholar] [CrossRef]
- Greening, C.; Biswas, A.; Carere, C.R.; Jackson, C.J.; Taylor, M.C.; Stott, M.B.; Cook, G.M.; Morales, S.E. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 2016, 10, 761–777. [Google Scholar] [CrossRef]
- Kapfhammer, D.; Karatan, E.; Pflughoeft, K.J.; Watnick, P.I. Role for glycine betaine transport in Vibrio cholerae osmoadaptation and biofilm formation within microbial communities. Appl. Environ. Microbiol. 2005, 71, 3840–3847. [Google Scholar] [CrossRef]
- Typas, A.; Banzhaf, M.; Gross, C.A.; Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2011, 10, 123–136. [Google Scholar] [CrossRef]
- Doucette, C.D.; Schwab, D.J.; Wingreen, N.S.; Rabinowitz, J.D. alpha-Ketoglutarate coordinates carbon and nitrogen utilization via enzyme I inhibition. Nat. Chem. Biol. 2011, 7, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mah, T.F. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. J. Bacteriol. 2008, 190, 4447–4452. [Google Scholar] [CrossRef]
- Ardin, A.C.; Fujita, K.; Nagayama, K.; Takashima, Y.; Nomura, R.; Nakano, K.; Ooshima, T.; Matsumoto-Nakano, M. Identification and functional analysis of an ammonium transporter in Streptococcus mutans. PLoS ONE 2014, 9, e107569. [Google Scholar] [CrossRef] [PubMed]
- Neznansky, A.; Blus-Kadosh, I.; Yerushalmi, G.; Banin, E.; Opatowsky, Y. The Pseudomonas aeruginosa phosphate transport protein PstS plays a phosphate-independent role in biofilm formation. FASEB J. 2014, 28, 5223–5233. [Google Scholar] [CrossRef] [PubMed]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed]
- Epstein, W. The roles and regulation of potassium in bacteria. Prog. Nucleic Acid. Res. Mol. Biol. 2003, 75, 293–320. [Google Scholar]
- Gray, H.B.; Winkler, J.R. Electron flow through metalloproteins. Biochim. Biophys. Acta 2010, 1797, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
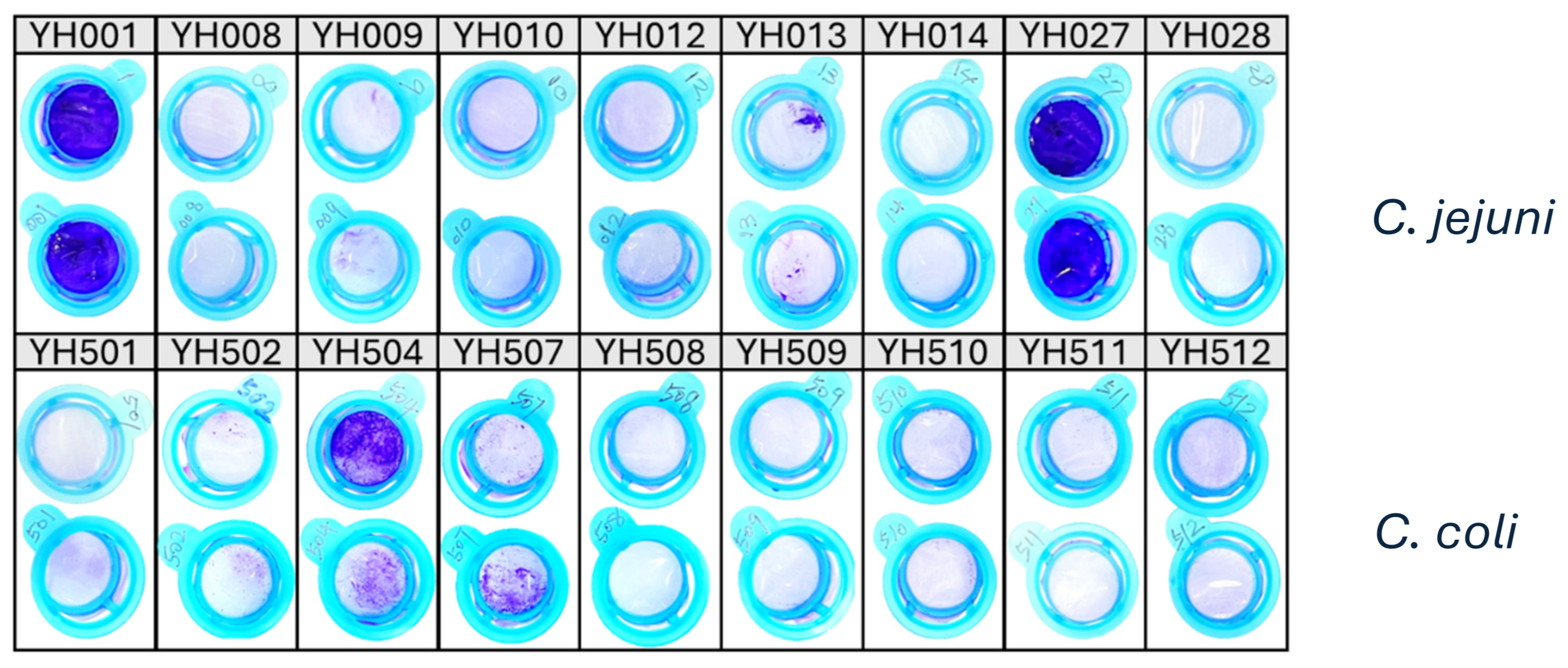
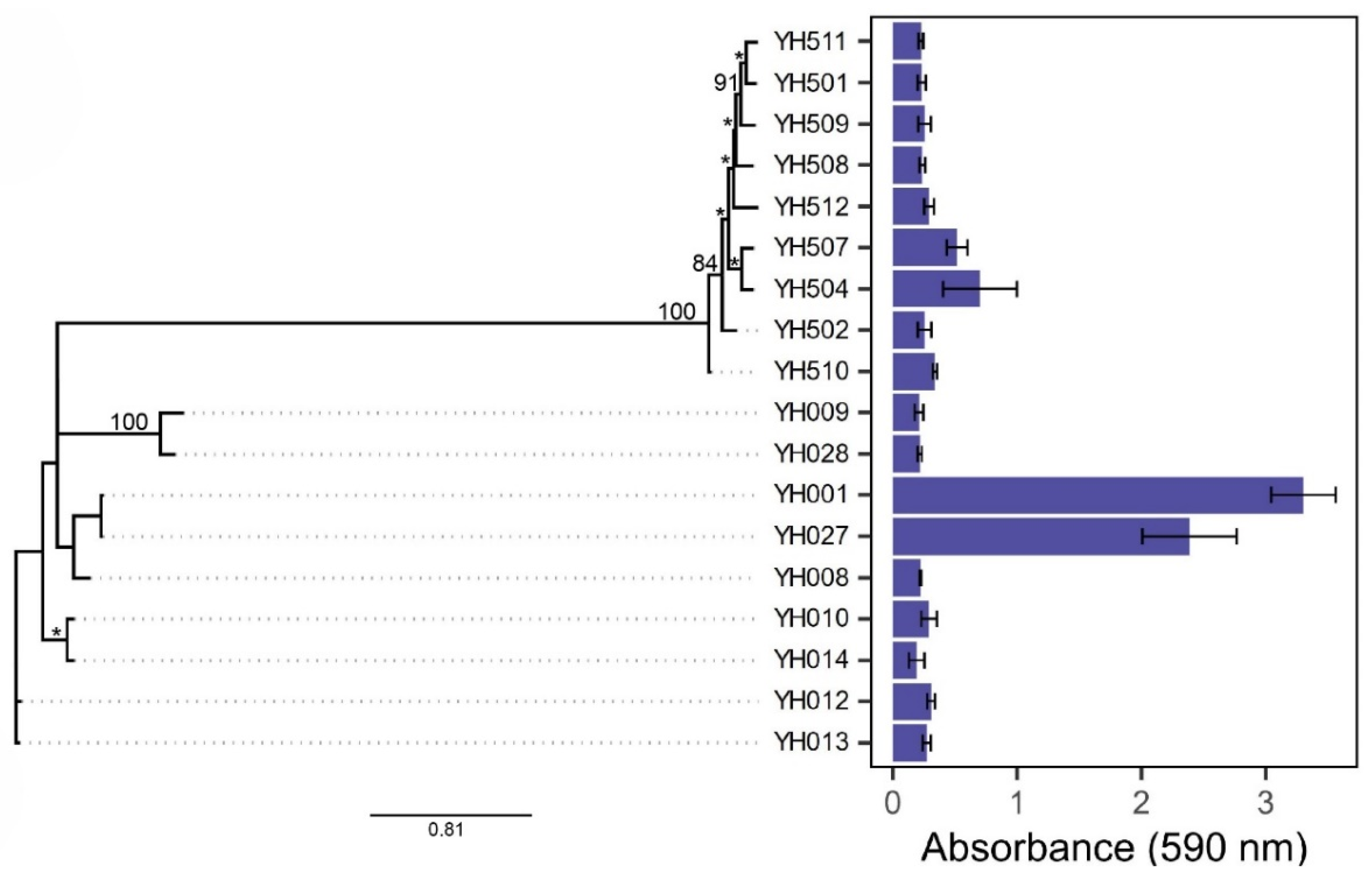
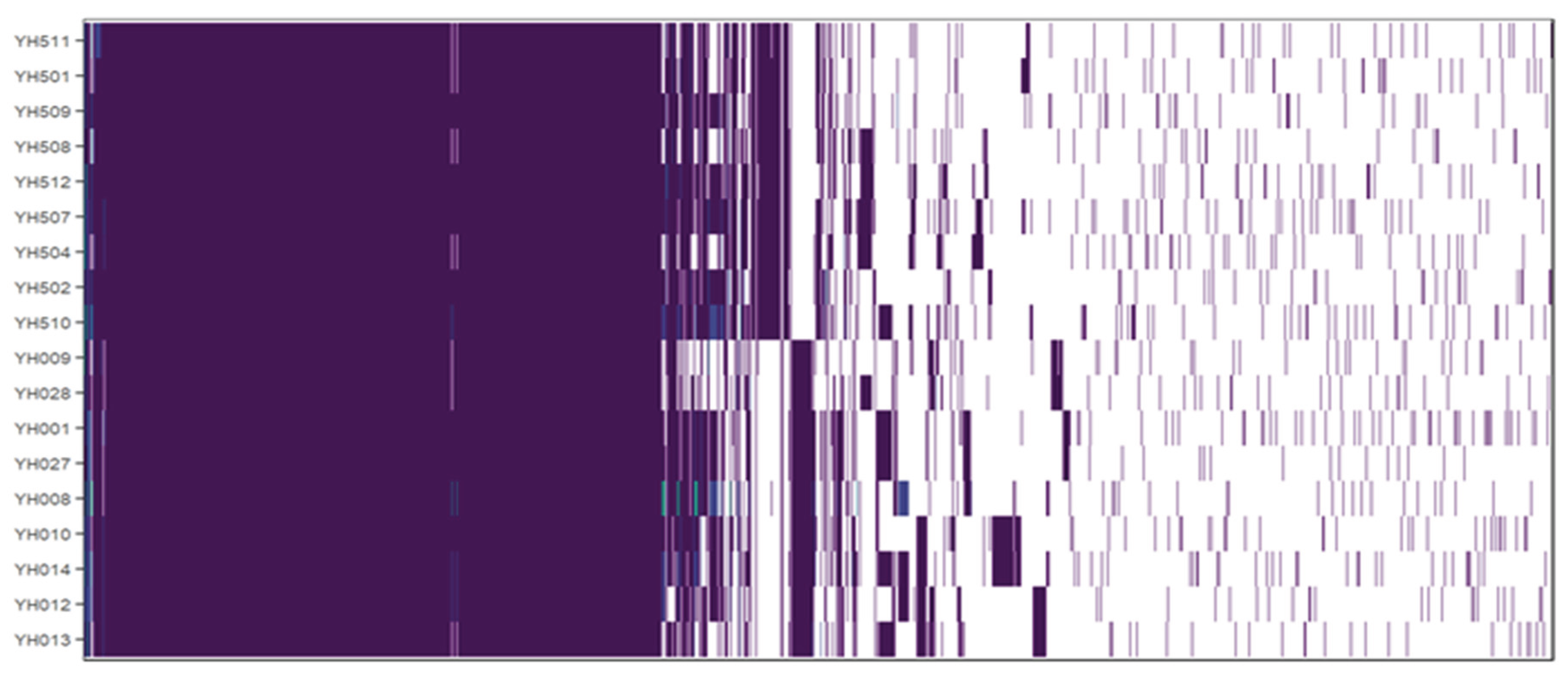

| Strain and Species | Source | Genome Size (bp) | %GC | Accession No. | Reference |
|---|---|---|---|---|---|
| C. jejuni YH001 | Veal livers | 1,712,361 | 30.5 | CP010058 | [22] |
| C. jejuni YH008 | Drumsticks | 1,792,424 | 30.5 | CP172380 | This work |
| C. jejuni (S27Cj) YH009 | Chicken thighs | 1,663,226 | 30.5 | CP131444 | [23] |
| C. jejuni (S33Cj) YH010 | Chicken thighs | 1,748,761 | 30.5 | CP131442 | [23] |
| C. jejuni YH012 | Chicken livers | 1,698,963 | 30.5 | CP172815 | This work |
| C. jejuni YH013 | Chicken livers | 1,691,848 | 30.5 | CP172379 | This work |
| C. jejuni YH014 | Chicken livers | 1,802,039 | 30.5 | CP172376 | This work |
| C. jejuni YH027 | Calf livers | 1,710,959 | 30.5 | CP172352 | This work |
| C. jejuni YH028 | Beef livers | 1,667,698 | 30.5 | CP172351 | This work |
| C. coli YH501 | Drumsticks | 1,668,523 | 31.5 | CP015528 | [24] |
| C. coli YH502 | Drumsticks | 1,718,974 | 31.0 | CP018900 | [25] |
| C. coli YH504 | Drumsticks | 1,722,143 | 31.0 | CP091644 | [24] |
| C. coli YH507 | Chicken livers | 1,756,096 | 31.0 | CP172392 | This work |
| C. coli YH508 | Chicken thighs | 1,703,740 | 31.5 | CP172391 | This work |
| C. coli YH509 | Chicken livers | 1,697,113 | 31.5 | CP172390 | This work |
| C. coli YH510 | Chicken livers | 1,812,356 | 31.0 | CP172387 | This work |
| C. coli YH511 | Chicken livers | 1,674,288 | 31.5 | CP172385 | This work |
| C. coli YH512 | Chicken livers | 1,754,135 | 31.5 | CP172384 | This work |
| Annotated Proteins | YH001 | YH027 | The Rest Strains |
|---|---|---|---|
| Putative Dihydrolipoamide dehydrogenase (EC 1.8.1.4); Mercuric ion reductase (EC 1.16.1.1); PF00070 family, FAD-dependent NAD(P)-disulphide oxidoreductase | 1 | 1 | 0 |
| D-alanyl-D-alanine carboxypeptidase (EC 3.4.16.4) | 1 | 1 | 0 |
| FIG00471123: hypothetical protein | 1 | 1 | 0 |
| DNA-binding protein Roi | 1 | 1 | 0 |
| FIG00470265: hypothetical protein | 1 | 1 | 0 |
| FIG00471635: hypothetical protein | 1 | 1 | 0 |
| FIG00470314: hypothetical protein | 1 | 1 | 0 |
| hydrogenase, (NiFe)/(NiFeSe) small subunit family | 1 | 1 | 0 |
| L-Proline/Glycine betaine transporter ProP | 1 | 1 | 0 |
| Uncharacterized membrane protein, YraQ family | 1 | 1 | 0 |
| Alpha-ketoglutarate permease | 1 | 1 | 0 |
| Putative efflux protein | 1 | 1 | 0 |
| Ammonium transporter | 1 | 1 | 0 |
| C4-dicarboxylate transporter | 1 | 1 | 0 |
| Sodium-dependent phosphate transporter | 1 | 1 | 0 |
| Multi antimicrobial extrusion protein (Na(+)/drug antiporter), MATE family of MDR efflux pumps | 1 | 1 | 0 |
| Hypothetical protein Cj0566 | 1 | 1 | 0 |
| Potassium-transporting ATPase A chain (EC 3.6.3.12) (TC 3.A.3.7.1) | 1 | 1 | 0 |
| Cytochrome c family protein | 1 | 1 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



