
Submitted:
06 February 2026
Posted:
06 February 2026
You are already at the latest version
Abstract
The whole framework of host innate and adaptive immunological pathways is proposed. In the innate immunity, γδ T cells can be categorized into several groups. Clonal anergy and tolerance pathway is related to Vγ2 chain γδ T cells. Host innate immunological pathway against viruses is related to Vγ8 chain γδ T cells. Host innate immunological pathway against intracellular micro-organisms is related to Vγ9 chain γδ T cells. Host innate immunological pathway against extracellular micro-organisms is related to Vγ4 chain γδ T cells. Host innate immunological pathway against helminths is related to Vγ3 chain γδ T cells. Host innate immunological pathway against insects is related to Vγ5 chain γδ T cells. In the adaptive immunity, there are five eradicable immune reactions and four tolerable immune reactions. In the tolerable immune reactions, TH3 is related to interleukin-35 producing CD4 T cells, and TH4 is related to interleukin-32 producing CD4 T cells.
Keywords:
TH3
; γδ T cells
; TH4
; interleukin-35
; interleukin-32
Introduction
Overview of host innate immunity and adaptive immunity
The immune system is a highly coordinated biological network that protects the host from pathogenic microorganisms while preserving tolerance to self-components. This system is classically divided into two interdependent arms: innate immunity and adaptive immunity. Together, these two branches provide both immediate defense and long-term, antigen-specific protection, forming an integrated and dynamic host defense system. Understanding the distinct characteristics and cooperative functions of innate and adaptive immunity is essential for elucidating immune homeostasis, host–pathogen interactions, and the mechanisms underlying immune-mediated diseases. My previous framework of host immunity is needed to be updated.[1]
Innate immunity constitutes the first line of defense against invading pathogens and is evolutionarily conserved across multicellular organisms. It relies on germline-encoded receptors, known as pattern recognition receptors, that detect conserved molecular structures shared by broad classes of microorganisms, referred to as pathogen-associated molecular patterns, as well as danger-associated molecular patterns released from damaged host cells. Innate immune responses are rapid, occurring within minutes to hours following pathogen exposure, and involve physical barriers such as the skin and mucosal surfaces, as well as cellular and humoral components including macrophages, neutrophils, dendritic cells, natural killer cells, and the complement system. Although innate immunity lacks antigen specificity and immunological memory, it plays a crucial role in containing early infection, shaping tissue inflammation, and instructing subsequent adaptive immune responses through cytokine production and antigen presentation[2].
Adaptive immunity, in contrast, is characterized by antigen specificity, clonal expansion, and immunological memory. This branch of the immune system is mediated primarily by T lymphocytes and B lymphocytes, which express highly diverse antigen receptors generated through somatic gene rearrangement. Adaptive immune responses are slower to develop during initial antigen exposure, typically requiring several days; however, they provide highly specific and potent effector functions, including cytotoxic T-cell–mediated killing of infected cells and antibody-mediated neutralization of extracellular pathogens. A defining feature of adaptive immunity is its ability to generate long-lived memory cells, which confer enhanced protection upon subsequent encounters with the same antigen, forming the basis of vaccination.
Despite their distinct properties, innate and adaptive immunity are not independent systems but function in close coordination. Innate immune cells, particularly dendritic cells, serve as critical intermediaries by capturing antigens and presenting them to naïve T cells, thereby initiating adaptive immune responses. Conversely, adaptive immune cells can modulate innate immunity through cytokine secretion and antibody-mediated opsonization, enhancing pathogen clearance. This bidirectional communication ensures an appropriate immune response tailored to the nature, location, and persistence of the antigenic challenge.
Dysregulation of either innate or adaptive immunity can result in pathological conditions, including chronic inflammation, autoimmunity, immunodeficiency, and cancer. Consequently, a comprehensive understanding of both immune arms and their regulatory mechanisms is fundamental for advancing immunological research and developing targeted therapeutic strategies. This review aims to synthesize current knowledge on innate and adaptive immune pathways, emphasizing their functional integration and relevance to immune tolerance and disease pathogenesis.
Mechanism of clonal anergy of host immune regulation
The immune system is fundamentally tasked with discriminating between self-antigens and foreign antigens in order to maintain host integrity while providing effective defense against pathogens. Under physiological conditions, immune cells that encounter self-antigens do not elicit immune responses, thereby preventing autoimmunity. A central principle underlying this specificity is the clonal selection mechanism, whereby each individual T or B lymphocyte expresses a unique antigen receptor and is therefore capable of recognizing only a single antigenic determinant. When a clonally distinct T or B cell recognizes a self-antigen, it enters a state of functional unresponsiveness known as clonal anergy, characterized by the absence of effector immune responses[2,3].
Clonal anergy is mediated through distinct cellular mechanisms involving γδ T cells and IgD-expressing B cells. γδ T cells arise developmentally earlier in the thymus than conventional αβ T cells. TRGV2 γδ T cells have TCR γδ chain in the first chromosome location, so the TRGV2 γδ T cells can be responsible for the clonal anergy. This temporal distinction has important implications for immune tolerance. When a developing clonal T cell encounters a self-antigen, particularly a protein antigen, it preferentially differentiates into a TRGV2 γδ T cell. As a consequence, the subsequent αβ T-cell repertoire is selectively shaped to avoid recognition of self-antigens, thereby preserving immune tolerance. Recognition of self-antigens by TRGV2 γδ T cells results in clonal anergy rather than immune activation, reinforcing central and peripheral tolerance mechanisms. Other γδ T cells are responsible for innate immunity which should react earlier than the adaptive αβ T-cells.
An analogous process operates within the B-cell compartment. Mature B lymphocytes co-express immunoglobulin D (IgD) and immunoglobulin M (IgM) on their cell surface. Antigen recognition through IgD serves a tolerogenic function: when IgD binds self-antigens, B cells undergo clonal anergy without initiating antibody-mediated immune responses. In contrast, recognition of foreign antigens through surface IgM triggers B-cell activation, leading to proliferation and differentiation. Activated IgM-bearing B cells subsequently undergo immunoglobulin class-switch recombination to generate IgG, IgE, or IgA antibodies, enabling effective humoral immunity against pathogens. This functional dichotomy between IgD- and IgM-mediated signaling highlights the critical role of IgD in maintaining B-cell tolerance.
Experimental evidence supports the tolerogenic roles of certain γδ T cells and IgD-expressing B cells. Multiple studies have demonstrated that IgD administration ameliorates autoimmune arthritis in animal models, suggesting a direct immunoregulatory effect. Similarly, certain γδ T cells have been shown to protect against graft-versus-host disease following organ transplantation, further underscoring their importance in immune tolerance.
γδ T cells are heterogeneous and can be subdivided based on their δ-chain usage and tissue distribution. γδ1 T cells are predominantly localized in the intestinal mucosa, where they contribute to clonal anergy toward dietary antigens and innate immunity against pathogens in the GI tracts. This mechanism underlies oral tolerance, explaining why common food proteins typically do not provoke immune responses. γδ2 T cells circulate mainly in peripheral blood and are primarily responsible for inducing tolerance to systemic self-antigens and for innate immunity against pathogens in the blood circulation. γδ3 T cells are enriched in the liver, an organ with intrinsic immune-tolerant properties and a central role in protein metabolism. These cells are essential for maintaining hepatic immune tolerance, particularly toward endogenous and metabolized antigens. γδ3 T cells are also responsible for the innate immunity against pathogens entering hepatic-spleen circulation.
The framework of innate immunological pathways
Based on the above host eradicable adaptive immune responses, there are five immunological pathways: TH1, TH2a, TH2b, TH22, and THαβ. γδ T cells play the vital roles in mediating innate immune reactions. There are mainly five types of γ chains of the γδ T cells, and there are mainly three types of δ chains of the γδ T cells. In our previous work, we pointed out that the three subtypes of δ chains of the γδ T cells are related to the organ-tissue locations of these cells. The δ1 γδ T cells are found in intestine or skin, the δ2 γδ T cells are found in blood, and the δ3 γδ T cells are found in liver. The functions of γ chain subtypes can be related to the host immunity fighting against different types of pathogens. Excluding the pseudogenes, there are six γ chain subtypes in human including Vγ2, Vγ3, Vγ4, Vγ5, Vγ8, and Vγ9. Thus, there are five or six innate immunological pathways which will be described below.
Clonal anergy and tolerance pathway is related to Vγ2 chain γδ T cells
The γδ T cells related to clonal anergy is the Vγ2 chain T cells. Vγ2 chain appears first in the γ chain sequence in the chromosome. Thus, it is reasonable that Vγ2 chain is associated with the function of clonal anergy. Because TCR γδ chain rearrange before the TCR αβ chain in thymus, it can assure that TCR against self-antigens can be produced first. Thus, these Vγ2 chain T cells can become clonal anergy which won’t let host immune reaction activated against self-antigens. Besides, TCR αβ chain against self-antigens are also eliminated during positive and negative selection of CD4 and CD8 T cells with double assurance. This kind of mechanism is like IgD and IgM double expressing B cells. When self-antigens bind to IgD, the IgM of B cells won’t be activated against self-antigens. It also won’t undergo following isotype class switch to IgG, IgE, or IgA. In brief, this is the clonal anergy and tolerance mechanism of the Vγ2 chain γδ T cells[2,4].
Host innate immunological pathway against viruses is related to Vγ8 chain γδ T cells
The eradicable host adaptive immune response against viruses and prions is THαβ immune reaction. In comparison with anti-viral THαβ immune reaction, the host innate immune response is mediated by Vγ8 chain γδ T cells. It is worth noting that immune tolerance with self-antigens and innate immune reactions are both mediated by limited diversity of the subtypes of immune cells including γδ T cells, invariant Natural Killer T cells (iNKT cells), or Mucosa-associated invariant T cells (MAIT cells). Other important immune cells related to Vγ8 chain γδ T cells include natural killer cells (NK cells), invariant NKT10 cells (iNKT10), MAIT10 cells, and innate lymphoid cells (ILC10 cells). Among these cells, many have invariant cells to correlated with the limited antigen diversity. These cells can produce interleukin-10 which is the central mediator of anti-viral immune response. Besides, B-1a cells are the immunoglobulin-M (IgM) producing B cells that provides the antibody for the innate immune response against viruses. These above cell types are related to host anti-virus immune response. Other infectious particles such as prions may also induce Vγ8 chain γδ T cells as well as the above cell types. For example, CMV infection can induce the activation of Vγ8 chain γδ T cells[5].
Host innate immunological pathway against intracellular micro-organisms is related to Vγ9 chain γδ T cells
Vγ9 chain γδ T cells are the subtypes of γδ T cells which mediate the innate immune reaction against intracellular micro-organisms including intracellular bacteria. Other intracellular micro-organisms including intracellular fungi and protozoa may also involve the activity of Vγ9 chain γδ T cells. In fact, γ9δ2 γδ T cells are the major populations of γδ T cells in our blood circulation. Previous studies reporting intracellular pathogens involving the innate γ9δ2 γδ T cells include the immune reactions against salmonella, shigella, and other intracellular bacteria. These immune cells can expand dramatically and exceed all other lymphocytes within a few days in tuberculosis, salmonellosis, ehrlichiosis, brucellosis, tularemia, listeriosis, toxoplasmosis, and malaria. Other immune cells involving this branch of immunological pathway include iNKT1 cells, ILC1 cells, MAIT1 cells, and B1-b IgM secreting B cells. Macrophages are the main effector cells in the innate immune reaction against intracellular micro-organisms. All γ9δ2 γδ T cells can recognize the same small microbial compound (E)-4-hydroxy-3-methyl-but-2-enyl phosphate (HMB-PP), a natural intermediate of the non-mevalonate pathway of isopentenyl pyrophosphate (IPP) biosynthesis. HMB-PP is an essential metabolite in many pathogenic intracellular bacteria or protozoa including Mycobacterium tuberculosis and malaria parasites. But, HMB-PP is absent in the host cells. Thus, HMB-PP can be the key antigen target of γ9δ2 γδ T cells against intracellular bacteria, fungi, or protozoa[6,7].
Host innate immunological pathway against extracellular micro-organisms is related to Vγ4 chain γδ T cells
As for extracellular bacteria infection, the host innate immune reaction is mainly Vγ4 chain γδ T cells. The TRGV4 γδ T cells can produce interleukin-17 which is related to TH17 or TH22 adaptive immune response. Other extracellular micro-organisms including extracellular fungi or extracellular protozoa can also stimulate Vγ4 chain γδ T cells in host innate immunity. The major effector cells of this branch of host innate immune reactions are neutrophils which can perform phagocytosis to digest the extracellular micro-organisms. Other immune cells related to TRGV4 γδ T cells are iNKT17 cells, MAIT17 cells, ILC3, and IgM producing marginal zone B-1 cells (MZ B-1 cells). IgM producing MZ B cells located in the marginal zone of spleen are the major B lymphocytes which encounter the portal circulation from intestines to produce natural IgM antibodies against extracellular bacteria, protozoa, and fungi from gastroenterol tracts. iNKT17 cells, MAIT17 cells, and ILC3 can all produce interleukin-17 for the anti-extracellular micro-organism immune reaction[8].
Host innate immunological pathway against helminths is related to Vγ3 chain γδ T cells
The host innate immunity against helminths is related to TRGV3 γδ T cells. TRGV3 γδ T cells are mainly located in mucosa of gastroenterol tracts or lungs. Thus, TRGV3 γδ T cells can react to helminths (endoparasites) which mainly live in GI tracts or respiratory tracts. TRGV3 γδ T cells are mucosa associated γδ T cells. They can react with Langerhans cells which are related to trigger TH2 immunity via antigen presentation. The main effector cells for this host innate immunity tract are mast cells and eosinophils. Other immune cells related to this innate immunological pathway include ILC2, MAIT2, iNKT2 cells, and IgM T2 cells[9,10].
Host innate immunological pathway against insects is related to Vγ5 chain γδ T cells
The innate immune reaction against insects is related to Vγ5 chain γδ T cells. These TRGV5 γδ T cells are skin residents. They are also called DETC T cells. They can react with Langerhans’ cells in the skin to trigger the innate immune response after insect bites. These insects are also called ectoparasites. The main effector cells of this host innate immunological pathway are basophils and mast cells. Other immune cells related to this innate immunological pathway include ILC2, MAIT2, iNKT2 cells, and IgM T2 cells[11,12]. The framework of host innate immune pathways is drawn in Figure 1.
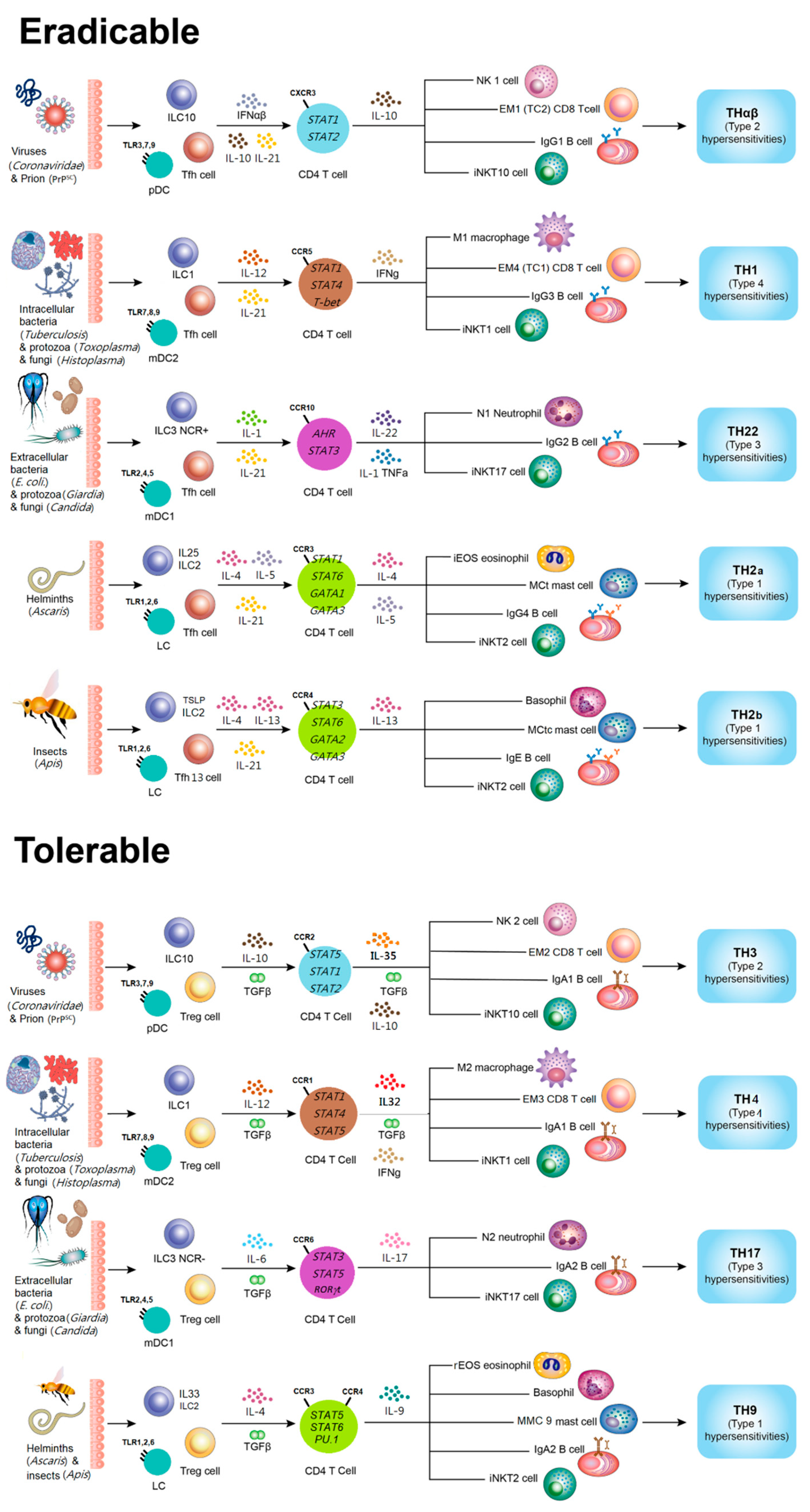
The Framework of Host adaptive Immunological Pathways
Host adaptive immune responses can be systematically classified into eradicable and tolerable pathways according to their functional objectives and regulatory mechanisms. Eradicable host immune reactions are primarily initiated by follicular helper T cells (Tfh). These CD4⁺ T cells are characterized by the expression of the chemokine receptor CXCR5 and the secretion of interleukin-21 (IL-21). Activation of Tfh cells is regulated by the transcription factors BCL6 and STAT5B, which collectively promote the maturation of B cells within germinal centers. Through this interaction, Tfh cells induce antibody production and immunoglobulin class switching to IgG, thereby establishing highly effective humoral immunity against a wide range of pathogens.
Among eradicable pathways, the TH1 immune response is specialized for the elimination of intracellular microorganisms, including viruses and certain bacteria. This pathway involves type 2 myeloid dendritic cells, type 1 innate lymphoid cells, M1-polarized macrophages, interferon-γ (IFN-γ)-secreting CD4⁺ T cells, cytotoxic CD8⁺ T cells, invariant natural killer T cells of the iNKT1 subset, and IgG3-producing B cells. Interleukin-12 (IL-12) is the principal cytokine that drives TH1 differentiation. Effector M1 macrophages activated in this pathway destroy infected host cells through lipid membrane peroxidation and other free radical-mediated processes. Consequently, TH1 immunity is closely associated with type 4 delayed-type hypersensitivity reactions.
The TH2 pathway is responsible for protective immunity against parasitic infections. It can be further divided into TH2a responses, which target endoparasites such as helminths, and TH2b responses, which act against ectoparasites. Each subtype engages distinct effector cell populations and cytokine profiles, including eosinophils, basophils, mast cells, and IgE- or IgG4-producing B cells. Dysregulation of TH2 immunity contributes to type 1 immediate hypersensitivity and classical allergic diseases. In addition, the TH22 pathway coordinates immunity against extracellular bacteria and fungi. Neutrophils, IL-22-secreting CD4⁺ T cells, iNKT17 cells, and IgG2-producing B cells participate in this response, which is mechanistically linked to type 3 immune complex-mediated hypersensitivity. The THαβ pathway is specialized for the recognition and removal of infectious particles. Natural killer cells, IL-10-producing CD4⁺ T cells, cytotoxic CD8⁺ T cells, and IgG1-producing B cells cooperate to eliminate pathogens, and this pathway is implicated in type 2 cytotoxic hypersensitivity reactions.
Tolerable host immune reactions are mediated by regulatory T cells (Treg), which initiate immune modulation rather than direct destruction. These cells express CD25 and secrete transforming growth factor-β (TGF-β), promoting antibody class switching to IgA through STAT5α and STAT5β activation. The TH4 pathway represents a regulated form of TH1 immunity. It responds to intracellular microorganisms using macrophages, IFN-γ/TGF-β-secreting CD4⁺ T cells, cytotoxic CD8⁺ T cells, iNKT1 cells, and IgA1-producing B cells. This pathway also contributes to type 4 hypersensitivity. The TH9 pathway confronts parasites under regulatory control, engaging eosinophils, basophils, mast cells, IL-9-secreting CD4⁺ T cells, iNKT2 cells, and IgA2-producing B cells, and is likewise related to type 1 allergic hypersensitivity.
Furthermore, TH17 and TH3 pathways constitute major components of tolerable immunity against extracellular microorganisms and infectious particles, respectively. TH17 reactions involve IL-17-producing CD4⁺ T cells, neutrophils, iNKT17 cells, and IgA2-producing B cells and are associated with type 3 hypersensitivity. The TH3 pathway is dominated by IL-10/TGF-β-producing regulatory CD4⁺ T cells, NK cells, CD8⁺ T cells, and IgA1-producing B cells and is implicated in type 2 cytotoxic hypersensitivity. A schematic representation of this integrated framework of host immunological pathways is illustrated in Figure 2.
Based on our previous immunological pathway framework, we need to know the key cytokines in TH3 and TH4 immune responses. TH3 is a well-known immunological reaction, it is the T cells producing interleukin-10 and TGF-β. However, Interleukin-10 is the central cytokine of THαβ immunity, and TGF-β is the central cytokine of Treg cells. Thus, there should be another cytokine representing TH3 immunological pathway. Recently, a new subset of regulatory T cells other than interleukin-10 producing Tr1 cells or TGF-β producing Treg cells was discovered. It is called iTr35 cells with massively producing interleukin-35[13,14]. iTr35 cells can suppress TH1, TH2, and TH17 related cytokines but it can up-regulate interleukin-10 which is the key cytokine in THαβ immunity[14]. The TH3 immune reaction is the tolerable pathway of the eradicable THαβ anti-viral immunity. So, interleukin-35 secreting iTr35 cells can be a good candidate of the key cytokine of TH3 immunity[15,16,17,18]. Indeed, TH3 condition with interleukin-10 and TGF-β can induce the production of interleukin-35. Interleukin-35 can also have positive feedback to increase the production of interleukin-10. Besides, iTr35 cells are identified as a regulatory CD4 T cell stable clone. These suggest that interleukin-35 or iTr35 cells are the key components of the TH3 immune response[19,20,21]. Previous literatures also pointed out the important role of interleukin-35 in the chronic virus infection including hepatitis B, hepatitis C, EBV, and influenza virus[22,23,24]. Thus, iTr35 cells can be the chronic tolerable anti-viral immune cells[25,26]. iTr35 cells are also related to tolerable immunity like systemic sclerosis[27]. iTr35 cells are actually TH3 cells.
TH4 immunological pathway is formerly called TH1-like immune reaction in our previous articles. Here, we rename the pathway TH4 immune pathway with identifying the key cytokines related to the TH4 immunological pathway. In the TH4 immunity, the tolerable immune response of the eradicable TH1 immunity, is against intracellular micro-organisms including intracellular bacteria, protozoa, and fungi[28]. Macrophages are the key effector cells in the TH1 and TH4 immunity fighting against intracellular micro-organisms. TH4 central cytokine should have the ability to shift M1 macrophages to M2 macrophages. In this study, we found that several cytokines are induced by interleukin-12 with TGFβ include interleukin-32, interleukin-19, and platelet factor 4. Interleukin-32 is mainly produced by CD4 T cells. Interleukin-32 has been reported in the host immunity against intracellular bacteria, protozoa, and fungi[28,29,30,31,32,33,34,35,36]. Interleukin-19 is mainly produced by monocytes or macrophages[37]. Interleukin-19 is related to tolerable immunity like fibrosis reaction[38]. Platelet factor 4, which is also named CXCL4, is mainly produced by platelets. All these cytokines can let macrophage switch from M1 macrophages to M2 macrophages[39,40,41,42]. Platelet factor 4 is more likely a chemokine rather than a cytokine and PF4 also has a regulatory function[43]. Because T helper cell major cytokines are mainly produced by CD4 T cells, interleukin-32 can be the most important cytokine standing for the TH4 immunological pathway[44]. Interleukin-32 is both a intracellular and an extracellular cytokine[45]. However, all these above cytokine-chemokines are important components of the TH4 immunity.
Once we know the key cytokines in TH3 and TH4 immunological pathways, the framework of immunity can be completed. There are study reports saying PF4 and interleukin-32 are related to TH1-Treg immune reaction. This so-called TH1-Treg immune reaction is actually TH4 immune response. Interleukin-32 is also reported to type 4 delayed type hypersensitivities which is TH1 dominant autoimmune disorders including type 1 diabetes[46]. In addition, interleukin-32 is often produced by solid cancer cells[47,48]. In our previous paper, we proposed that TH1-like immunity (TH4 immunity) is the pro-tumor immunological pathway. And, the levels of interleukin-32 are related to the prognosis of cancer patients[49]. Thus, it is reasonable that solid tumors can secrete interleukin-32[47,50]. Based on the findings of this study, TH3 may also be called TH35 and TH4 may also be called TH32. In our previous paper, we pointed out that CCR1 is the chemokine receptor of TH1-like immunity (TH4 immunity)[51]. Actually, PF4(CXCL4) is a chemokine ligand for CCR1[52]. TH3 or TH4 are better names according to the history of immunology. In terms of pathogen defense, TH3 is related to tolerable anti-infectious particle immunity including viruses and prions, and TH4 is related to tolerable anti-intracellular micro-organism immunity including intracellular bacteria, protozoa, and fungi. This study can provide more information and strategies to combat viruses or intracellular micro-organisms clinically.
Conclusion
The host innate immunological pathways can be categorized into six groups. Clonal anergy and tolerance pathway is related to Vγ2 chain γδ T cells. Host innate immunological pathway against viruses is related to Vγ8 chain γδ T cells. Host innate immunological pathway against intracellular micro-organisms is related to Vγ9 chain γδ T cells. Host innate immunological pathway against extracellular micro-organisms is related to Vγ4 chain γδ T cells. Host innate immunological pathway against helminths is related to Vγ3 chain γδ T cells. Host innate immunological pathway against insects is related to Vγ5 chain γδ T cells. In the adaptive immunity, there are five eradicable immune reactions and four tolerable immune reactions. In the tolerable immune reactions, TH3 is related to interleukin-35 producing CD4 T cells, and TH4 is related to interleukin-32 producing CD4 T cells. Once we know the mechanism of the host innate and adaptive immune response pathways, we can develop more strategies against infectious diseases as well as hypersensitivity reactions.
References
- Hu, W.C. A Framework of All Discovered Immunological Pathways and Their Roles for Four Specific Types of Pathogens and Hypersensitivities. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Chen, J.Y.; Shih, L.J.; Liao, M.T.; Tsai, K.W.; Lu, K.C.; Hu, W.C. Understanding the Immune System's Intricate Balance: Activation, Tolerance, and Self-Protection. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tsai, K.W.; Lu, K.C.; Shih, L.J.; Hu, W.C. Cancer as a Dysfunctional Immune Disorder: Pro-Tumor TH1-like Immune Response and Anti-Tumor THalphabeta Immune Response Based on the Complete Updated Framework of Host Immunological Pathways. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Lal, G. Differentiation and functional plasticity of gamma-delta (gammadelta) T cells under homeostatic and disease conditions. Mol Immunol 2021, 136, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Vermijlen, D.; Brouwer, M.; Donner, C.; Liesnard, C.; Tackoen, M.; Van Rysselberge, M.; Twite, N.; Goldman, M.; Marchant, A.; Willems, F. Human cytomegalovirus elicits fetal gammadelta T cell responses in utero. J Exp Med 2010, 207, 807–821. [Google Scholar] [CrossRef]
- Eberl, M.; Hintz, M.; Reichenberg, A.; Kollas, A.K.; Wiesner, J.; Jomaa, H. Microbial isoprenoid biosynthesis and human gammadelta T cell activation. FEBS Lett 2003, 544, 4–10. [Google Scholar] [CrossRef]
- Guo, F.; Song, Y.; Dong, S.; Wei, J.; Li, B.; Xu, T.; Wang, H. Characterization and anti-tuberculosis effects of gammadelta T cells expanded and activated by Mycobacterium tuberculosis heat-resistant antigen. Virulence 2025, 16, 2462092. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, Q.; Hao, Y.; Tao, Z.; Song, W.; Chen, S.; Qin, L.; Song, W.; Shan, Y. Infiltrated IL-17A-producing gamma delta T cells play a protective role in sepsis-induced liver injury and are regulated by CCR6 and gut commensal microbes. Front Cell Infect Microbiol 2023, 13, 1149506. [Google Scholar] [CrossRef]
- Parthasarathy, A.; Li, T.; Edelblum, K.L. Crosstalk between the microbiota and intestinal gammadelta T cell compartments in health and IBD. Gut Microbes 2026, 18, 2604908. [Google Scholar] [CrossRef]
- Li, G.Q.; Xia, J.; Zeng, W.; Luo, W.; Liu, L.; Zeng, X.; Cao, D. The intestinal gammadelta T cells: functions in the gut and in the distant organs. Front Immunol 2023, 14, 1206299. [Google Scholar] [CrossRef]
- Girardi, M.; Lewis, J.M.; Filler, R.B.; Hayday, A.C.; Tigelaar, R.E. Environmentally responsive and reversible regulation of epidermal barrier function by gammadelta T cells. J Invest Dermatol 2006, 126, 808–814. [Google Scholar] [CrossRef]
- Girardi, M.; Lewis, J.; Glusac, E.; Filler, R.B.; Geng, L.; Hayday, A.C.; Tigelaar, R.E. Resident skin-specific gammadelta T cells provide local, nonredundant regulation of cutaneous inflammation. J Exp Med 2002, 195, 855–867. [Google Scholar] [CrossRef]
- Ye, C.; Yano, H.; Workman, C.J.; Vignali, D.A.A. Interleukin-35: Structure, Function and Its Impact on Immune-Related Diseases. J Interferon Cytokine Res 2021, 41, 391–406. [Google Scholar] [CrossRef]
- Yang, C.; Dong, L.; Zhong, J. Immunomodulatory effects of iTr35 cell subpopulation and its research progress. Clin Exp Med 2024, 24, 41. [Google Scholar] [CrossRef]
- Al-Khikani, F.H.O. Role of interleukin-35 in viral infections: Promising immunological branch. Journal of Medical Society 2024, 38, 169–177. [Google Scholar] [CrossRef]
- Collison, L.W.; Chaturvedi, V.; Henderson, A.L.; Giacomin, P.R.; Guy, C.; Bankoti, J.; Finkelstein, D.; Forbes, K.; Workman, C.J.; Brown, S.A.; et al. IL-35-mediated induction of a potent regulatory T cell population. Nat Immunol 2010, 11, 1093–1101. [Google Scholar] [CrossRef]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Kochetkova, I.; Golden, S.; Holderness, K.; Callis, G.; Pascual, D.W. IL-35 stimulation of CD39+ regulatory T cells confers protection against collagen II-induced arthritis via the production of IL-10. J Immunol 2010, 184, 7144–7153. [Google Scholar] [CrossRef] [PubMed]
- Collison, L.W.; Delgoffe, G.M.; Guy, C.S.; Vignali, K.M.; Chaturvedi, V.; Fairweather, D.; Satoskar, A.R.; Garcia, K.C.; Hunter, C.A.; Drake, C.G.; et al. The composition and signaling of the IL-35 receptor are unconventional. Nat Immunol 2012, 13, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Niedbala, W.; Wei, X.Q.; Cai, B.; Hueber, A.J.; Leung, B.P.; McInnes, I.B.; Liew, F.Y. IL-35 is a novel cytokine with therapeutic effects against collagen-induced arthritis through the expansion of regulatory T cells and suppression of Th17 cells. Eur J Immunol 2007, 37, 3021–3029. [Google Scholar] [CrossRef]
- Olson, B.M.; Sullivan, J.A.; Burlingham, W.J. Interleukin 35: a key mediator of suppression and the propagation of infectious tolerance. Front Immunol 2013, 4, 315. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Wang, W. IL-35: A Novel Immunomodulator in Hepatitis B Virus-Related Liver Diseases. Front Cell Dev Biol 2021, 9, 614847. [Google Scholar] [CrossRef]
- Shao, X.; Ma, J.; Jia, S.; Yang, L.; Wang, W.; Jin, Z. Interleukin-35 Suppresses Antiviral Immune Response in Chronic Hepatitis B Virus Infection. Front Cell Infect Microbiol 2017, 7, 472. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, S.; Xu, G.; Feng, J.; Han, T.; Zhao, F.; She, Y.L.; Liu, S.; Ye, L.; Zhu, Y. Gene Expression and Antiviral Activity of Interleukin-35 in Response to Influenza A Virus Infection. J Biol Chem 2016, 291, 16863–16876. [Google Scholar] [CrossRef]
- Shamji, M.H.; Layhadi, J.; Perera-web, A.; Yan, R.; Durham, S.R. IL-35-Producing T Cells (iTR35) Inhibit Th2 Effector Function, Induce Infectious Tolerance and Are Elevated Following Grass Pollen Sublingual Immunotherapy. Journal of Allergy and Clinical Immunology 2014, 133. [Google Scholar] [CrossRef]
- Sullivan, J.A.; Tomita, Y.; Jankowska-Gan, E.; Lema, D.A.; Arvedson, M.P.; Nair, A.; Bracamonte-Baran, W.; Zhou, Y.; Meyer, K.K.; Zhong, W.; et al. Treg-Cell-Derived IL-35-Coated Extracellular Vesicles Promote Infectious Tolerance. Cell Rep 2020, 30, 1039–1051 e1035. [Google Scholar] [CrossRef]
- Yang, C.; Lu, C.; Pan, J.; Zhao, C.; Chen, Z.; Qin, F.; Wen, J.; Wei, W.; Lei, L. The role of iTr35 cells in the inflammatory response and fibrosis progression of systemic sclerosis. Rheumatology (Oxford) 2023, 62, 3439–3447. [Google Scholar] [CrossRef]
- Braga, Y.L.L.; Neto, J.R.C.; Costa, A.W.F.; Silva, M.V.T.; Silva, M.V.; Celes, M.R.N.; Oliveira, M.A.P.; Joosten, L.A.B.; Ribeiro-Dias, F.; Gomes, R.S.; et al. Interleukin-32gamma in the Control of Acute Experimental Chagas Disease. J Immunol Res 2022, 2022, 7070301. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.S.; Silva, M.V.T.; Dos Santos, J.C.; de Lima Silva, L.L.; Batista, A.C.; Machado, J.R.; Teixeira, M.M.; Dorta, M.L.; de Oliveira, M.A.P.; Dinarello, C.A.; et al. IL-32gamma promotes the healing of murine cutaneous lesions caused by Leishmania braziliensis infection in contrast to Leishmania amazonensis. Parasit Vectors 2017, 10, 336. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Dias, F.; Saar Gomes, R.; de Lima Silva, L.L.; Dos Santos, J.C.; Joosten, L.A. Interleukin 32: a novel player in the control of infectious diseases. J Leukoc Biol 2017, 101, 39–52. [Google Scholar] [CrossRef]
- Guimaraes de Matos, G.; Barroso de Figueiredo, A.M.; Diniz Goncalves, P.H.; Luiz de Lima Silva, L.; Bastista, A.C.; Borges, C.L.; Maria de Almeida Soares, C.; Joosten, L.A.B.; Ribeiro-Dias, F. Paracoccidioidesbrasiliensis induces IL-32 and is controlled by IL-15/IL-32/vitamin D pathway in vitro. Microb Pathog 2021, 154, 104864. [Google Scholar] [CrossRef] [PubMed]
- Koeken, V.; Verrall, A.J.; Ardiansyah, E.; Apriani, L.; Dos Santos, J.C.; Kumar, V.; Alisjahbana, B.; Hill, P.C.; Joosten, L.A.B.; van Crevel, R.; et al. IL-32 and its splice variants are associated with protection against Mycobacterium tuberculosis infection and skewing of Th1/Th17 cytokines. J Leukoc Biol 2020, 107, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Deng, W.; Xie, J. The Biology and Role of Interleukin-32 in Tuberculosis. J Immunol Res 2018, 2018, 1535194. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Azam, T.; Lewis, E.C.; Joosten, L.A.; Wang, M.; Langenberg, D.; Meng, X.; Chan, E.D.; Yoon, D.Y.; Ottenhoff, T.; et al. Mycobacterium tuberculosis induces interleukin-32 production through a caspase- 1/IL-18/interferon-gamma-dependent mechanism. PLoS Med 2006, 3, e277. [Google Scholar] [CrossRef]
- Ribeiro-Dias, F.; Oliveira, I.B.N. A Critical Overview of Interleukin 32 in Leishmaniases. Front Immunol 2022, 13, 849340. [Google Scholar] [CrossRef]
- Schenk, M.; Krutzik, S.R.; Sieling, P.A.; Lee, D.J.; Teles, R.M.; Ochoa, M.T.; Komisopoulou, E.; Sarno, E.N.; Rea, T.H.; Graeber, T.G.; et al. NOD2 triggers an interleukin-32-dependent human dendritic cell program in leprosy. Nat Med 2012, 18, 555–563. [Google Scholar] [CrossRef]
- Gabunia, K.; Autieri, M.V. Interleukin-19 can enhance angiogenesis by Macrophage Polarization. Macrophage (Houst) 2015, 2, e562. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, S.; Wang, K.; Zhang, M.; Li, M.; Zan, Y.; Huang, Q.; Wu, S.; Zhao, W.; Xu, W.; et al. Interleukin-19 Aggravates Pulmonary Fibrosis via Activating Fibroblast through TGF-beta/Smad Pathway. Mediators Inflamm 2022, 2022, 6755407. [Google Scholar] [CrossRef]
- Erbel, C.; Tyka, M.; Helmes, C.M.; Akhavanpoor, M.; Rupp, G.; Domschke, G.; Linden, F.; Wolf, A.; Doesch, A.; Lasitschka, F.; et al. CXCL4-induced plaque macrophages can be specifically identified by co-expression of MMP7+S100A8+ in vitro and in vivo. Innate Immun 2015, 21, 255–265. [Google Scholar] [CrossRef]
- Gleissner, C.A. Macrophage Phenotype Modulation by CXCL4 in Atherosclerosis. Front Physiol 2012, 3, 1. [Google Scholar] [CrossRef]
- Schaffner, A.; Rhyn, P.; Schoedon, G.; Schaer, D.J. Regulated expression of platelet factor 4 in human monocytes--role of PARs as a quantitatively important monocyte activation pathway. J Leukoc Biol 2005, 78, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, R.; Toyoda, E.; Maehara, M.; Wasai, S.; Omura, H.; Watanabe, M.; Sato, M. Effect of Platelet-Rich Plasma on M1/M2 Macrophage Polarization. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Fleischer, J.; Grage-Griebenow, E.; Kasper, B.; Heine, H.; Ernst, M.; Brandt, E.; Flad, H.D.; Petersen, F. Platelet factor 4 inhibits proliferation and cytokine release of activated human T cells. J Immunol 2002, 169, 770–777. [Google Scholar] [CrossRef]
- Sanna, F.C.; Benesova, I.; Pervan, P.; Krenz, A.; Wurzel, A.; Lohmayer, R.; Muhlbauer, J.; Wollner, A.; Kohl, N.; Menevse, A.N.; et al. IL-2 and TCR stimulation induce expression and secretion of IL-32beta by human T cells. Front Immunol 2024, 15, 1437224. [Google Scholar] [CrossRef]
- Heinhuis, B.; Netea, M.G.; van den Berg, W.B.; Dinarello, C.A.; Joosten, L.A. Interleukin-32: a predominantly intracellular proinflammatory mediator that controls cell activation and cell death. Cytokine 2012, 60, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Hanna, S.J. Mini review: Interleukin-32 as a key mediator of type 1 diabetes pathogenesis. Front Immunol 2025, 16, 1641698. [Google Scholar] [CrossRef] [PubMed]
- Paz, H.; Tsoi, J.; Kalbasi, A.; Grasso, C.S.; McBride, W.H.; Schaue, D.; Butterfield, L.H.; Maurer, D.M.; Ribas, A.; Graeber, T.G.; et al. Interleukin 32 expression in human melanoma. J Transl Med 2019, 17, 113. [Google Scholar] [CrossRef]
- Sun, Y.; Qian, Y.; Chen, C.; Wang, H.; Zhou, X.; Zhai, W.; Qiu, L.; Zhou, X.; Ning, H.; Zhao, Y.; et al. Extracellular vesicle IL-32 promotes the M2 macrophage polarization and metastasis of esophageal squamous cell carcinoma via FAK/STAT3 pathway. J Exp Clin Cancer Res 2022, 41, 145. [Google Scholar] [CrossRef]
- Yan, H.; He, D.; Huang, X.; Zhang, E.; Chen, Q.; Xu, R.; Liu, X.; Zi, F.; Cai, Z. Role of interleukin-32 in cancer biology. Oncol Lett 2018, 16, 41–47. [Google Scholar] [CrossRef]
- Han, L.; Chen, S.; Chen, Z.; Zhou, B.; Zheng, Y.; Shen, L. Interleukin 32 Promotes Foxp3(+) Treg Cell Development and CD8(+) T Cell Function in Human Esophageal Squamous Cell Carcinoma Microenvironment. Front Cell Dev Biol 2021, 9, 704853. [Google Scholar] [CrossRef]
- Chu, Y.T.; Liao, M.T.; Tsai, K.W.; Lu, K.C.; Hu, W.C. Interplay of Chemokines Receptors, Toll-like Receptors, and Host Immunological Pathways. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Kausar, F.; Day, A.; Osborne, M.; Hussain, K.; Mueller, A.; Lin, J.; Tsuchiya, T.; Kanegasaki, S.; Pease, J.E. CXCL4/Platelet Factor 4 is an agonist of CCR1 and drives human monocyte migration. Scientific Reports 2018, 8. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
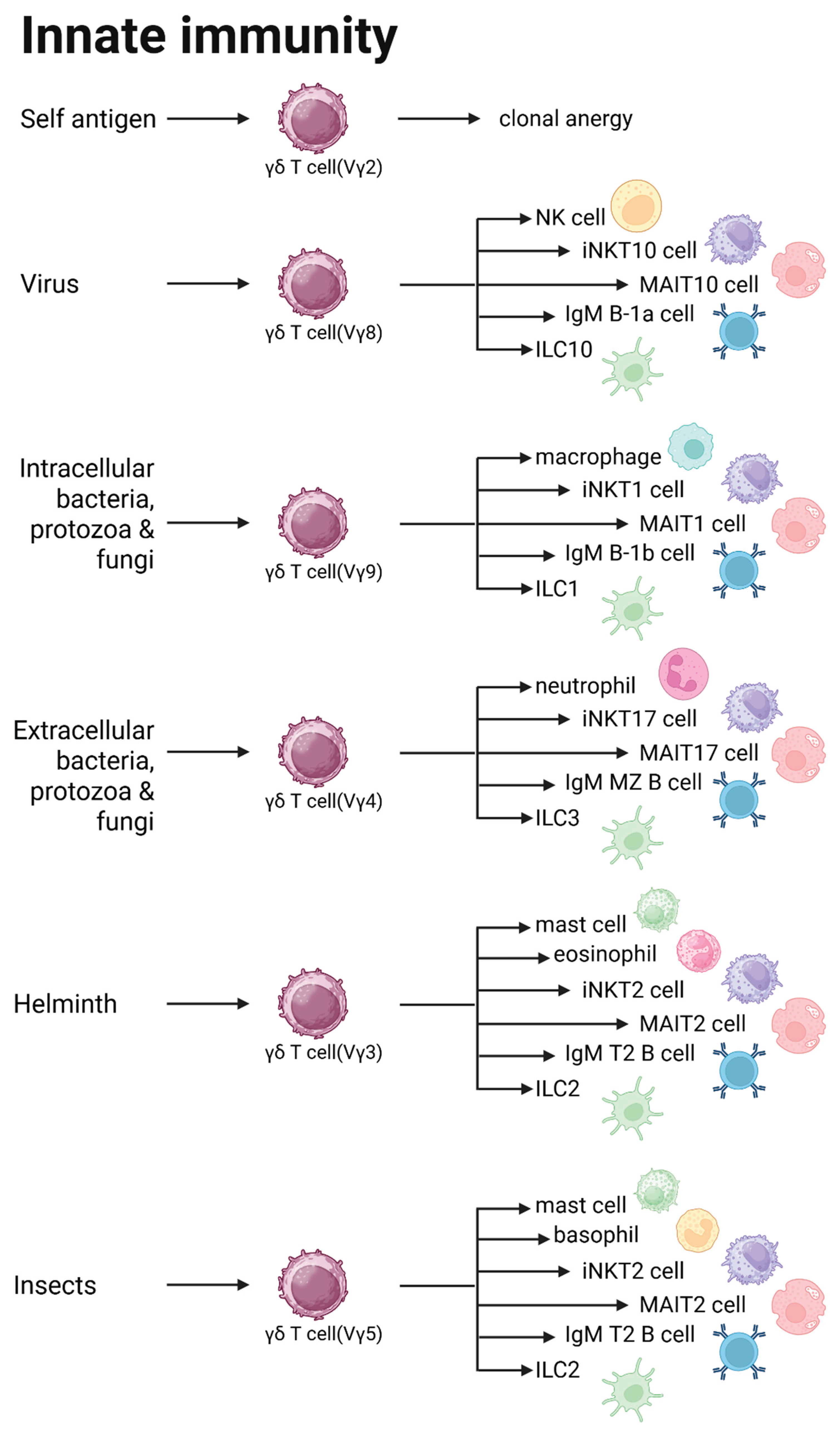
Figure 2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



