
Submitted:
05 February 2026
Posted:
06 February 2026
You are already at the latest version
Abstract
This critical review examines the evolution of mathematical modeling approaches for aerobic digestion processes in food industry waste management, highlighting their role in operational optimization and dynamic prediction. Starting from mass conservation principles, simple kinetic models such as first-order and Monod models are analyzed. These models assume homogeneity and perfect mixing but fail to capture the heterogeneity of effluents rich in variable carbohydrates, proteins, and lipids. Structural limitations, such as numerical rigidity, parametric non-identifiability, and idealized assumptions that underestimate spatial gradients and stochastic fluctuations, are discussed. In continuous systems, coupled substrate-biomass-oxygen dynamics, washout phenomena, and extensions to partial differential equations for real heterogeneity are explored. Structured models such as ASM incorporate multicomponent fractions but face parameterization crises exacerbated by data scarcity in industrial settings, where less than 25% of plants use formal modeling. Emerging paradigms include hybrid mechanistic-machine learning approaches for prediction under perturbations, multiscale modeling , and spatially explicit modeling. A table distributes approaches by food matrix, revealing the dominance of simple kinetics in composting and ASM in activated sludge. Finally, a progressive selection framework based on operational objectives is proposed, balancing complexity with predictive robustness and experimental validation, emphasizing that sophistication must be justified to overcome barriers such as sensor costs and stochastic variability, thus promoting sustainable industrial adoption.
Keywords:
mathematical modeling
; aerobic digestion
; food waste
; hybrid models
; parametric identifiability
1. Introduction
Composting organic waste is an important process in the environmental management of the food industry, and aerobic digestion, in its different operating configurations such as activated sludge, extended aeration reactors, and sequential batch, exhibits high efficiency in removing chemical oxygen demand, simultaneous nitrification, and the ability to be adapted to variable loads [1]. The establishment of these processes as the preferred technology for high-density organic waste has historically been closely linked to the development of mathematical modeling frameworks that allow for the formalization of the underlying biological mechanisms, the optimization of reactor design, and the prediction of the dynamic system's behavior in response to operational disturbances [2].
The first models were based on simplified pseudo-first- order kinetics, which assumed linear relationships between degradation rate and substrate concentration, without considering biomass dynamics or enzyme saturation processes [3]. The progressive incorporation of mechanistic models of microbial growth, particularly Monod kinetics and its extensions with Haldane-type inhibition, represented a conceptual advance by introducing a nonlinear saturating dependence between specific growth rate and substrate availability [4]. Subsequently, the development of complex structured models such as the Activated Sludge Models with multiple microbial populations, differentiated substrate fractions, and detailed nitrogen and phosphorus balances significantly expanded the descriptive capacity of these frameworks [5].
However, despite over twenty years of intensive research into advanced structured models, a significant gap still exists between theoretical sophistication and demonstrable industrial adoption [6]. Recent structural identifiability analysis based on Fisher information theory and global sensitivity analysis has revealed that models with 15-25 kinetic parameters often contain subsets of parameters that are structurally unidentifiable, even with noise-free data, and that multiple radically different parameter sets can reproduce the same calibration data but can diverge catastrophically in their predictions under operational disturbances [7]. This identifiability crisis is exacerbated when applied in industry, where data is scarce, manual monitoring is usually performed only once or twice a day on a few basic parameters, and this differs greatly from the high-resolution continuous sensors on which advanced structured models operate [8].
Field studies and industry surveys have documented that less than 25% of food processing plants use formal mathematical models to support operational decisions, and when they do, simple empirical approaches predominate over high-dimensional models [9]. The barriers identified include limited technical capacity for advanced calibration using global optimization algorithms, a lack of sufficiently high-quality data for robust parameterization, stochastic variability in organic load that invalidates static calibrations, and prohibitive costs for advanced sensor infrastructure , particularly for small and medium-sized enterprises (SMEs), which represent more than 60% of the sector [10].
In the food industry in particular, these difficulties are compounded by the inherent heterogeneity of the effluents produced. Unlike the relatively homogeneous municipal wastewater, food production effluents have a complex and fluctuating composition, consisting of mixtures of short- and long-chain carbohydrates, proteins with varying rates of hydrolysis, lipids that are difficult to biodegrade aerobically, and phenolic compounds that act as inhibitors [11]. They are also subjected to sudden variations over time, with magnitudes between 200 and 800% in chemical oxygen appetite, linked to discontinuous production cycles, cleaning operations that discharge concentrated alkaline and acidic solutions, and intermittent discharges during production line changes [12]. These properties directly challenge basic assumptions of classical models such as the spatial homogeneity assumed by perfect mixing theories, the stationarity of the influence, and the stability of the kinetic parameters [13].
Over the past decade, hybrid modeling paradigms have emerged that combine mechanistic model-based frameworks with machine learning techniques for difficult-to-model components, such as predicting stochastic organic shocks and optimizing aeration strategies under energy constraints [14]. Simultaneously, the development of digital twins for wastewater treatment plants promises to bridge the gap between research models and operational tools through real-time simulation and early fault detection [15]. However, these emerging approaches face critical challenges, including the need for massive amounts of data for robust training, which exceeds typical industry availability; the poor interpretability of black-box models, hindering operational acceptance; and the very high costs of implementing sensors [16].
Scrutiny of modern literature shows that the justification for more complex modeling must be based on incremental increases in the quantitative analysis of its predictive power versus cost in parameters, grounded in experimental data and well-defined operational objectives [17]. Over-parameterized models run without strict calibration protocols often lead to lower predictability beyond the calibration conditions compared to simple formulations with well-identifiable parameters validated by independent experiments [18]. This observation is particularly relevant in industrial settings, where information is limited, stochastic variability over time is high, and spatial gradients are not accounted for in large-scale reactors.
This critical review examines the development of mathematical models for the aerobic digestion of food waste from an industrial perspective, discussing the trade-offs between simplicity, realism, identifiability, and predictive capacity, as well as interpretability and flexibility. It considers the evolution from empirical kinetics to multicomponent models, spatial formulations, and hybrid models based on the integration of mechanism with machine learning. The aim is to provide quantitative tools for choosing the appropriate model complexity, emphasizing that predictive robustness in real-world systems depends more on the structural consistency between assumptions and physical system characteristics, the practical identifiability of parameters, and rigorous experimental validation than on the number of implemented equations.
2. Mathematical Foundations of Aerobic Digestion
Mathematical modeling of aerobic digestion processes is based on universal physical principles, including conservation principles, which allow us to describe the changes in open biological systems transformed through complex biochemical reactions. In environmental engineering and food science, the principle of mass conservation is the backbone of most dynamic models developed to predict the aerobic biodegradation of organic waste [19].
Several reviews have reported that the dominant mechanistic models for aerobic digestion are derived from mass balances formulated at the macroscopic scale, under simplifying assumptions that allow for mathematical solutions but limit their application to the system of interest [20]. This is a particularly critical problem when models are used for food industry effluents, which are highly heterogeneous in physicochemical terms, with a significant particulate fraction and temporal fluctuations in organic loading.
In contrast to homogeneous municipal wastewater, the substrate derived from food production has complex distributions of carbohydrates, proteins, and lipids, as well as solids with varying degrees of biodegradability. This complexity leads to nonlinear dynamics, biological process-transport phenomenon couplings, and spatial dependencies that cannot be adequately described by zero-dimensional formulations. Therefore, although the conservation principle offers a convenient mathematical foundation, its direct application often results in models that are not very predictive of plant conditions [21].
In this context, the literature has evolved toward a progressive diversification of modeling approaches, ranging from empirical kinetic formulations to structured multicomponent frameworks, spatially resolved models, and, more recently, hybrid data-driven approaches. To conceptually organize this progression and provide a comprehensive analytical framework, an epistemological hierarchy of mathematical models is introduced, based on their structural complexity, their capacity to represent the real system, and the requirements for identification and validation.
As Figure 1 illustrates, the lower layers of the hierarchy are based directly on conservation principles and possess a high degree of interpretability and mathematical simplicity; however, they encounter limitations in representing the structural and spatial heterogeneity of food effluents. In contrast, the higher levels are constructed from explicit biochemical processes, spatial gradients, and data-derived functional relationships, increasing the system's representation to a higher level of dimensional and computational complexity, as well as parametric fragility.
This conceptual framework allows us to understand classical mass balances not as isolated formulations, but as the lowest level of a methodological progression that naturally leads to heterogeneous, spatial, and hybrid models, driven by real-world constraints. Thus, the proposed hierarchy serves as an organizing principle that guides the mathematical analysis in the subsequent subsections, where the differential foundations, structural aspects, and numerical challenges associated with each level are formalized.
The Principle of Conservation of Mass in the Mathematical Formulation of Aerobic Processes
The principle of conservation of mass states that the total mass of a component in a system can only change due to inputs, outputs, or internal reactions. This principle forms the basis of the most relevant formal frameworks in biological modeling, including Activated Sludge Models (ASM) for aerobic wastewater treatment, as well as the Anaerobic Digestion Model (ADM) initially developed for anaerobic and composting processes [22]. For an arbitrary component , the mass balance is expressed by Equation (1):
where denotes the system volume; and are the inlet and outlet flow rates, respectively; and represent the inlet and outlet concentrations of component ; and is the net reaction rate. This general formulation enables the extension of the analysis to more complex dynamics in aerobic digestion processes, accounting for stochastic variations and heterogeneities inherent to food waste streams.
Equation (1) forms the basis for defining dynamic models and allows the biochemical processes of the system to be expressed in a concrete mathematical language. In practice, this equation becomes a system of ordinary differential equations under assumptions generally accepted in the literature, such as constant volume, perfect mixing, and homogeneity of the medium. These assumptions have been widely adopted in models of activated sludge, aerobic reactors, and composting systems, mainly due to their mathematical simplicity and ease of implementation [23].
However, the resulting mathematical equation has significant structural limitations. The generation-consumption terms that model microbial kinetics and the biodegradation process are typically defined using nonlinear functions dependent on substrate, oxygen, and biomass concentrations [24]. When rapid processes, such as the oxidation of soluble substrates, and slow processes, such as the hydrolysis of particulate fractions, occur simultaneously, the dynamical system can be numerically very stiff. Mathematically, this stiffness is reflected in the eigenvalue spectrum of the linearized system around the steady state. If we consider the Jacobian eigenvalues of the ODE system, the stiffness can be measured through the stiffness ratio defined in Equation (2):
Where large values of R, typically greater than , indicate the coexistence of very different timescales. Under such conditions, explicit methods of numerical integration become unstable or inefficient, requiring the use of implicit or semi-implicit schemes, which entails an increase in computational cost and greater complexity in the numerical analysis.
where high values of , typically greater than , indicate the coexistence of very disparate time scales. Under such conditions, explicit methods of numerical integration become unstable or inefficient, making it necessary to resort to implicit or semi-implicit schemes, with the consequent increase in computational cost and complexity of the numerical analysis [25].
These restrictions were exacerbated in food wastewater, where 20 to 40% of dissolved solids (DSP) were in the form of particulate solids, as has been reported for fruit, meat, and dairy wastewater [26]. Separation into dissolved and particulate fractions facilitates additional sequential processes such as disintegration and hydrolysis, which are rarely explicitly modeled in conventional mass balance models. Consequently, additional delays and nonlinearities arise, affecting the model's prediction, particularly in situations of high organic loads or low aeration.
While most aerobic models assume complete mixing and spatial homogeneity, oxygen, temperature, and humidity gradients are observed that strongly influence microbial activity in real-world environments [27]. Ignoring these gradients can lead to incorrect calculations of retention times, degradation rates, and gas emissions, even in environments much more practical than industrial reactors and compost heaps.
Table 1 provides a summary of the key mathematical assumptions used in mass balances for aerobic digestion, showing their frequency of use in the literature, the associated mathematical limitations, and the specific effect on food effluents. This table does not present novel results, but rather offers an analysis based on a comparison within the review article genre itself, allowing for a clear and structured visualization of the recurring shortcomings in current approaches.
The evidence presented in Table 1 indicates that, while these assumptions simplify the mathematical formulation and numerical solution, they greatly limit the applicability of the models when confronted with the biochemical and structural complexity of real feed effluents. In this sense, although mass balances strictly based on Equation (1) are formally correct, they are insufficient to describe the spatial and multi-scale dynamics of aerobic digestion, which is why these approaches must be extended to partial differential equations or coupled hybrid models.
3. Kinetic Models and Their Limitations in Industrial Applications
Kinetic models are the mathematical foundation of aerobic digestion models, defining the functional form of the generation and consumption terms in the mass balance equations. From a formal perspective, selecting kinetic law is equivalent to imposing a hypothesis about the dominant degradation mechanism, the role of biomass, and the limiting regime of the system as a closed system. In industrial applications, this selection involves a trade-off between analytical simplicity, computational stability, and biological fidelity, which is particularly critical in the treatment of adapted food effluents.
3.1. First-Order Kinetics in the Modeling of Aerobic Processes
First-order kinetics remains widely used in aerobic process modeling, particularly in feasibility studies and applications where computational simplicity is prioritized over biological realism. This persistence is not solely due to historical reasons, but also to practical needs, such as the lack of sufficient experimental data, to use models that can be easily calibrated. However, recent analyses have highlighted that this decision is often made without a critical analysis of the structural implications or the effect on the interpretability of the estimated parameters [32].
The mathematical formulation of these kinetics is expressed by Equation (3):
where represents the concentration of biodegradable substrate and is an apparent kinetic constant. Equation (3) indicates that the degradation rate is only a function of the available substrate, which implicitly assumes that the active biomass does not limit the overall kinetics of the process. Recent research has highlighted that this assumption is rarely met in real aerobic systems, even under apparently stable conditions, due to the physiological differentiation of microbial communities and the presence of slowly biodegradable fractions [33].
Direct integration of Equation (3) leads to the analytical solution given by Equation (4):
The solution to Equation (4) is globally stable and monotonically increasing. These characteristics have led to its use as a descriptive tool, since the parameter can be related to a characteristic time of the process . However, recent work has indicated that this interpretation is erroneous, given that it is not a tangible physiological parameter, but an aggregate quantity that represents several unresolved processes and is implicit in the model [34].
From a mechanistic perspective, first-order kinetics can be interpreted as a limiting case of more general formulations based on the explicit dynamics of biomass. Specifically, Monod-type kinetics, which is widely used in models of the ASM family and remains relevant in recent developments, expresses the consumption rate through Equation (5):
where , and retain their usual meaning. Under the assumption of approximately constant biomass, , and in the low substrate concentration regime, , Equation (5) can be approximated by a first-order expansion, giving rise to Equation (6):
Substituting Equation (6) into the mass balance leads to an equation formally equivalent to Equation (3), with . This result, critically revisited in recent work on structural identifiability, demonstrates that first-order kinetics represents an asymptotic approximation, valid only under very restrictive conditions that are rarely sustained in complex aerobic systems [35].
3.2. Degradation of Kinetics and Fundamental Dynamic Limitations
A direct consequence of Equation (3) is that the modulus of the substrate degradation rate is given by Equation (7):
Equation (7), which is directly derived from equation (3), indicates that the degradation rate is at its maximum at time 0 and decreases monotonically over time. From a dynamic point of view, this property imposes a very rigid structure on the model, which cannot overcome an initial growth acceleration associated with microbial adaptation, a characteristic frequently observed in real aerobic systems and in recent experimental studies [36].
The analysis of the curvature of the timeline with the solution of Equation (4) for the trajectory corroborates this restriction. By differentiating this solution twice, or equivalently by differentiating Equation (3) again, we arrive at the second derivative given in Equation (8):
The strict positivity of Equation (8) implies that the solution has no inflection points, which confirms the mathematical impossibility of the model generating abrupt changes between regimes dominated by biomass, hydrolysis, or oxygen constraints. Recent analyses have shown that this structural constraint produces systematic errors in predictions of stabilization times and in the evaluation of control strategies when applied to higher-order models outside their domain of validity [36].
A more general representation of the dynamic process is obtained using coupled systems of equations for the substrate and biomass, such as that described by Equation (9):
These formulations allow for the explicit capture of latency and growth-with-decay phases and form the basis of recent developments aimed at improving the parametric identifiability and predictability of aerobic models [36]. Reducing the system defined by Equation (9) to a single equation of the form of Equation (3) eliminates these essential dynamics and reinforces the empirical nature of the parameter .
In the food industry, the elimination of complex waste products with complex physical and chemical properties introduces a level of complexity. Recent studies have established that a significant portion of the substrate decomposes over very different timescales, allowing the use of multifraction models that extend the exponential solution of Equation (4). This approximation is expressed through Equation (10):
Equation (10) specifies that there are fast and slow degradation fractions, and this is observed in recurring types of food waste. The use of a single exponential term, as in Equation (4), amounts to imposing the conjecture , which has been recognized as one of the main sources of bias in estimating overall efficiencies and sorting times for industrial aerobic processes in recent studies [37].
4. Microbial Growth Models and Limitations in Parameter Calibration
Microbial growth constitutes the kinetic core of mathematical models applied to aerobic processes, as it establishes the functional link between substrate availability, biomass generation, and the overall rate of organic matter degradation [38]. In industrial applications, the formulation adopted to describe this growth not only conditions the model's predictive capacity but also its mathematical stability, the identifiability of its parameters, and its transferability across effluent scales and matrices [39]. In the case of food wastewater, characterized by high compositional heterogeneity and temporal variability, these limitations are significantly accentuated.
4.1. Formulation and Mathematical Properties of the Monod Model
The Monod model is the most widely used kinetic formulation to describe substrate-limited microbial growth in aerobic systems [40]. The functional relationship between the specific microbial growth rate and the substrate concentration is expressed by Equation (11):
In Equation (11), represents the maximum specific growth rate and the half-saturation constant, defined as the substrate concentration for which . This formulation presents well-defined mathematical properties, such as continuity, increasing monotonicity, and asymptotic saturation, which has favored its integration into complex dynamic models and standardized frameworks such as ASM models.
By coupling Equation (11) with the biomass and substrate mass balances, the nonlinear dynamic system described by Equations (12) and (13) is obtained:
where is the active biomass concentration, the endogenous decay coefficient and the biomass-substrate yield. The system defined by Equations (11)-(13) exhibits multiple time scales and nonlinear dynamics that can generate numerical rigidity when coupled with rapid processes, such as oxygen transfer or acid-base reactions, a frequent situation in long-term industrial simulations [41].
From a critical point of view, although the Monod model adequately captures kinetic saturation at low and moderate substrate concentrations, its formulation assumes the existence of a single limiting substrate, a hypothesis that is rarely met in complex food effluents [18].
4.2. Formal Comparison with Alternative Kinetic Models
To overcome some of the structural limitations of the Monod model, alternative kinetic formulations have been proposed that modify the functional dependence between the growth rate and the state variables. One of the earliest extensions is the Tessier model, which introduces an exponential dependence described by Equation (14):
In Equation (14), is a scaling parameter associated with the sensitivity of growth to changes in substrate concentration. This formulation exhibits a more gradual transition toward saturation, which can improve empirical fitting in systems where the microbial response does not follow a strict hyperbolic relationship. However, recent studies have indicated that this improved fit is often achieved at the cost of reduced physiological interpretability of the parameters [42].
Another relevant formulation is the Contois model, which explicitly incorporates the biomass concentration into the kinetic limitation, as expressed in Equation (15):
Equation (15) is particularly relevant in systems with high cell concentrations or mass transfer limitations, common conditions in industrial aerobic reactors and food effluents with high particulate organic loads. However, the explicit introduction of the variable in the denominator increases the nonlinearity of the system and exacerbates the problems of parametric identifiability when experimental data are limited [43].
In the presence of substrate inhibition, models such as Andrews or Haldane extend the classical Monod formulation by including an inhibitory term, as shown in Equation (16):
where is the inhibition constant. Equation (16) describes the decrease in growth rate at high substrate concentrations, a phenomenon reported in effluents with toxic compounds or high organic loads. However, recent analyses have shown that the inclusion of inhibitory terms often introduces highly correlated parameters, whose robust estimation requires extensive and highly informative datasets, which are uncommon in real-world industrial applications [44].
4.3. Structural Identifiability and Parametric Calibration Problems
From a mathematical perspective, one of the main challenges associated with microbial growth models defined by Equations (11), (14)-(16) is the structural identifiability of their parameters. In nonlinear models, parameters such as , , and can exhibit strong correlations, leading to non-uniqueness problems in parametric estimation.
Formally, this difficulty manifests as an ill-conditioned Fisher information matrix, meaning that small perturbations in the experimental data generate large variations in the estimated parameter values. Recent studies have shown that even when the overall model fit is satisfactory, the confidence intervals for the parameters can be excessively wide, compromising the model's predictive ability outside of calibration conditions [45].
4.4. Experimental Implications and Model Validation
The validation of microbial growth kinetic models should not be limited to the visual fitting of substrate consumption or biomass growth curves. It is necessary to systematically evaluate the stability of the estimated parameters, the model's sensitivity to disturbances, and its predictive capacity under operating conditions different from those used for calibration.
Recent comparative studies have shown that, in complex matrices such as dairy, meat, or brewery effluents, models like Contois or formulations with inhibition can provide statistically superior fits to the Monod model, but only when sufficient experimental data are available to support the added complexity [46]. In the absence of this information, introducing more complex models can create a false sense of accuracy without any real improvement in predictive ability.
4.5. Critical Considerations on the Selection of the Kinetic Model
From a critical perspective, the selection of a microbial growth model for industrial applications should not be based solely on its mathematical simplicity or the quality of the statistical fit. It should explicitly consider physiological consistency, parametric identifiability, and the operational context in which the model will be used.
In heterogeneous food wastewater, the uncritical adoption of the Monod model or its extensions can lead to empirical parameters that are difficult to interpret and predictions that are not robust to changes in effluent composition. In this regard, recent literature converges on the need to adopt progressive modeling approaches, in which kinetic complexity is increased only when there is sufficient experimental evidence to justify its introduction [47].
To summarize the structural and application differences between the main models discussed, Table 2 presents a systematic comparison of their mathematical characteristics and their relevance in food contexts.
5. Dynamic Models in Continuous Systems: Flow Integration and Multiscalarity
5.1. Coupled Substrate-Biomass-Oxygen Dynamics in Continuous Reactors
The mathematical modeling of continuous aerobic processes introduces additional complexity compared to closed or batch systems, due to the explicit presence of inlet and outlet flows that interact with the internal biological kinetics. In this context, mass balances are no longer governed solely by reaction terms, but incorporate hydraulic processes that modify the stability, parametric identifiability, and dynamic structure of the system. This approach is representative of continuous stirred-tank reactors widely used in the biological treatment of industrial effluents and wastewater, where the assumption of perfect mixing is combined with nonlinear kinetic hypotheses derived from microbial growth.
Under conditions of constant volume and complete mixing, the dynamic balance of the soluble substrate is expressed by Equation (17):
where represents the concentration of biodegradable substrate, is the biomass-substrate yield, the specific microbial growth rate, the substrate concentration in the influent, and the hydraulic retention time. Equation (17) highlights the direct competition between biological consumption processes and hydraulic dilution, an interaction that is crucial for the operational stability of the reactor, especially under variable load conditions, as documented in recent studies on complex food effluents [48].
The dynamics of active biomass are described by Equation (18):
where is the active biomass concentration, the endogenous decay coefficient, and the biomass present in the influent. Equation (18) reveals that net biomass growth depends not only on microbial kinetics but also on the hydraulic conditions of the system. Several recent studies have indicated that the explicit omission of the dilution term leads to a systematic overestimation of biological activity and errors in predicting steady states in continuous reactors [49].
In aerobic systems, oxygen availability constitutes an additional constraint that introduces a different timescale, associated with gas -liquid mass transfer. The evolution of dissolved oxygen can be described by Equation (19):
where is the dissolved oxygen concentration, the saturation concentration, the volumetric transfer coefficient, and the oxygen consumption rate associated with biodegradation. The inclusion of Equation (19), along with Equations (17) and (18), results in a nonlinear, multiscale dynamic system in which rapid physical transfer processes coexist with slower biological processes. This disparity in scales has been identified as one of the main sources of numerical rigidity in the simulation of continuous aerobic systems, conditioning the choice of appropriate time integration methods [50].
5.2. Hydraulic Stability, Washout and Dynamic Bifurcations
From a mathematical perspective, the system defined by Equations (17)-(19) exhibits dynamic properties substantially different from those observed in batch models. In particular, the presence of external flows alters the structure of steady-state equilibria and can induce instability phenomena associated with critical hydraulic conditions. A typical manifestation of this behavior is the washout phenomenon, in which biomass is carried out of the reactor due to insufficient retention time.
From Equation (18), this regime is reached when the condition expressed in Equation (20) is satisfied:
Equation (20) defines a critical threshold of hydraulic retention time below which equilibrium with positive biomass loses stability compared to trivial equilibrium . Recent studies have shown that this critical point is mathematically associated with a transcritical bifurcation, the correct identification of which is essential for the design and control of aerobic reactors subjected to variations in organic loading [51].
When kinetic formulations are expanded to incorporate multiple substrates, inhibition, or intracellular storage, the system can exhibit multiple steady-state equilibria and non-monotonic dynamic regimes. These behaviors have been reported in industrial aerobic processes with effluents of varying composition, reinforcing the need for stability and bifurcation analyses as an integral part of the modeling process [52].
5.3. Space-Dependent Extension and Heterogeneity Effects
The assumption of perfect mixing implicit in Equations (17)-(19) is an idealization that is not always met in real systems. In large-scale industrial reactors, the presence of spatial gradients of concentration, oxygen, and temperature can significantly affect the efficiency of the biological process. To capture these effects, dynamic models can be extended to space-dependent formulations based on partial differential equations.
The dynamics of the soluble substrate in a one-dimensional domain can be expressed by Equation (21):
where represents the fluid velocity field and the effective diffusion or dispersion coefficient. Equation (21) formally extends the balance defined in Equation (17) by incorporating advection and diffusion processes, allowing the representation of phenomena such as hydraulic dead zones, mass transfer limitations, and spatial heterogeneities, widely documented through CFD simulations coupled with biological models [53].
From a mathematical point of view, the inclusion of spatial terms increases the dimensionality of the system and tightens the computational requirements but provides a more faithful representation of real processes when spatial heterogeneity dominates reactor dynamics.
5.4. Multicomponent Formalization in ASM Models
The activated sludge models developed by the International Water Association represent the most comprehensive formalization of continuous-flow dynamics models. In these models, substrate, biomass, oxygen, and nutrient dynamics are integrated through coupled stoichiometric and kinetic balances, conceptually extending Equations (17)-(21) to a multicomponent and multiscale framework.
In compact notation, this general formulation is expressed by Equation (22):
where is the vector of state concentrations, the stoichiometric matrix, and the vector of process rates. Equation (22) generalizes the individual balances and allows the incorporation of additional processes such as nitrification, denitrification, polymer storage, and associated oxygen consumption.
However, recent studies have indicated that the high structural complexity of ASM models introduces significant challenges in terms of parametric identifiability and experimental data requirements, particularly when applied to complex industrial effluents. This limitation reinforces the need for critical and contextualized use of these models, prioritizing consistency between the adopted mathematical structure and the information available for their calibration and validation [54].
6. Advanced Structured Models: Mathematical Complexity and Parametric Identification Problems
The evolution from aggregate kinetic models to structured formulations represents an explicit attempt to increase the biological realism of aerobic processes by incorporating the functional heterogeneity of microbial biomass. In this context, the total biomass is broken down into fractions with differentiated metabolic roles, leading to expressions such as Equation (23):
This disaggregation allows us to distinguish between metabolically active biomass, the inert non-biodegradable fraction, and intracellular reserve compounds, which are conceptually consistent with microbiological observations and the structure of advanced models such as ASM3. However, from a mathematical and experimental perspective, the introduction of these latent variables substantially increases the dimensionality of the dynamic system and exacerbates problems of structural and practical identifiability.
Several recent studies have shown that, even with dense time series data for substrate and total biomass, the simultaneous estimation of the fractions defined in Equation (10) is not unique, resulting in multiple parameters sets that are equally compatible with the observed data [55]. This non-uniqueness manifests itself in almost linearly dependent sensitivity matrices and a poorly conditioned Fisher information matrix, which severely limits the interpretability of the estimated parameters.
From an applied perspective, this limitation is intensified in complex food wastewater, where the direct experimental quantification of active, inert, and reserve biomass requires advanced techniques such as dynamic respirometry, isotopic markers, or biochemical fractionation, which are not always feasible at an industrial scale. Consequently, increased theoretical rigor does not necessarily translate into a proportional improvement in the model's predictive capacity.
6.1. Mass Balances in Ideal Reactors and Their Industrial Limitations
Most advanced structured models are based on dynamic balances formulated under the assumption of a perfectly mixed reactor. For a continuous stirred-tank reactor, the overall mass balance for a generic component can be expressed by Equation (24):
where it represents the concentration of the component in the reactor, its concentration in the influent, the normalized volumetric flow rate, and the reaction term that groups the biochemical transformations.
Although Equation (11) is a cornerstone of dynamic modeling, its validity relies on the assumption of spatial homogeneity, which is rarely met in large-scale industrial aerobic systems. Recent studies based on in situ measurements and CFD simulations have demonstrated the systematic presence of spatial temperature gradients exceeding 20 °C and dissolved oxygen gradients ranging from anoxic conditions to near-saturation levels, even in reactors formally designed as CSTRs [56].
These spatial heterogeneities invalidate the strict application of Equation (24) and generate differentiated microenvironments in which aerobic, nitrifying, and denitrifying processes coexist simultaneously. In the treatment of meat by-products and protein-rich waste, this functional compartmentalization favors the emergence of specialized microbial niches, whose dynamics cannot be captured by zero-dimensional models, even those that include complex kinetics.
6.2. Energy Balance and Thermodynamic Simplifications
The extension of structured models to the thermal description of the process is usually done through global energy balances formulated as Equation (25):
where is the effective density of the medium, its heat capacity, the temperature, the heat generation terms and the losses to the environment.
While this formulation allows for the incorporation of the thermal effect of microbial activity, its common use involves significant thermodynamic simplifications. Biological heat generation is frequently modeled as proportional to the overall substrate degradation rate, assuming constant metabolic efficiency. However, recent work has shown that microbial energy efficiency varies substantially throughout the process due to the ecological succession of consortia, changes in the dominant metabolic pathway, and local oxygen limitations.
In composting systems for fish waste and meat by-products, these variations manifest as abrupt thermal peaks that can reach temperatures close to 70 °C, followed by prolonged plateaus. Such thermal profiles cannot be adequately reproduced using constant coefficients in Equation (12), highlighting the need for energy models coupled to microbial dynamics and substrate structure.
6.3. Parametric Sensitivity and Predictive Fragility in Structured Models
Increased structural complexity leads to greater sensitivity of the model to parametric uncertainty. Table 3 summarizes typical ranges of kinetic parameters used in advanced composting and aerobic digestion models, as well as their impact on the system's dynamic response, according to recent reviews of the specialized literature [57].
This high sensitivity, combined with identifiability problems and the difficulty of obtaining independent experimental data, limits the predictive robustness of structured models when applied outside of carefully controlled conditions.
7. The Crisis of Parameterization and the Role of Environmental Corrections
One of the main bottlenecks in the mathematical modeling of aerobic processes lies not in the formulation of kinetic equations or dynamic balances, but in the reliable parameterization of increasingly complex models. Despite significant progress in the biological and physicochemical structuring of models, recent literature reveals a persistent gap between theoretical complexity and parametric robustness. In a comprehensive systematic review, Zhu et al. [57]. They report that less than 26% of the studies analyzed incorporate formal sensitivity or identifiability analyses, despite employing high-dimensional nonlinear models.
This methodological deficit is particularly problematic because most advanced models exhibit structural or practical non-identifiability, which compromises the physical interpretation of parameters and severely limits predictive capacity outside of calibration conditions.
7.1. Parametric Non-Identifiability and Mathematical Limits of Calibration
From a mathematical perspective, the problem can be formalized by considering a dynamic model described by a system of ordinary differential equations with unknown state variables and parameters, where typically . In this context, parametric estimation using maximum likelihood or least squares methods is based on the Fisher information matrix, defined as in Equation (26):
Where represents the parameter vector and the likelihood function associated with the experimental data.
In kinetic and structured models of aerobic processes, the matrix defined in Equation (13) is frequently singular or severely ill-conditioned, with condition numbers greater than . This implies that small perturbations in the experimental data can induce large variations in the estimated parameters, making it impossible to obtain unique and stable solutions. Recent studies have shown that this non-identifiability persists even when high-time-resolution data are available, due to intrinsic functional correlations between kinetic parameters such as , , and [58].
Empirical evidence supports this diagnosis. In wastewater treatment processes in the juice and beverage industry, multiple parameter sets have been observed to generate statistically indistinguishable fits with high coefficients of determination, typically during the calibration phase. However, when these same models are subjected to validation scenarios with changes in organic load or flow rate, the predicted dynamic trajectories diverge significantly, revealing a structural fragility of the model in the face of operational disturbances [59]. This phenomenon highlights that a good statistical fit does not constitute a guarantee of mechanistic validity or predictive capacity.
7.2. Environmental Corrections: Between Empirical Pragmatism and Mechanistic Incoherence
Given the limitations of direct parameterization, a widespread practice is to introduce environmental correction functions designed to capture the effect of external variables such as temperature, humidity, or oxygen availability. However, Valenzuela et al. [60] They indicate that only 46% of the reviewed models explicitly implement this type of correction, and that in most cases these are independent empirical formulations.
The most common approach involves modifying the specific growth rate using multiplicative factors, as expressed in Equation (27):
where , and represent dimensionless correction functions for temperature, humidity and oxygen, respectively.
Although this formulation is computationally convenient and improves the local fit of the model, it has profound conceptual limitations. First, it implicitly assumes that environmental effects are separable and independent, which contradicts experimental evidence showing nonlinear interactions between temperature, water availability, and oxygen transfer. Second, it restricts the environmental influence solely to the growth rate, ignoring that other fundamental kinetic parameters also depend on environmental conditions.
Both the half-saturation constant and the maximum growth rate should be considered multivariate functions of the environmental state, as conceptually expressed in Equations (28) and (29):
These dependencies reflect changes in microbial affinity for the substrate, metabolic efficiency, and the structure of the biological consortium, phenomena widely documented in recent studies on aerobic processes and composting [61].
From a mathematical perspective, introducing empirical environmental corrections can mask identifiability problems by artificially increasing the number of degrees of freedom in the model, without necessarily improving its mechanistic consistency. As a result, models are obtained that adequately reproduce historical data, but whose extrapolation to new operating conditions proves unreliable.
8. Advanced Mathematical Modeling of Aerobic Processes in Complex Systems
The most recent scientific literature agrees that classical approaches to aerobic digestion modeling, based on zero-dimensional mass balances and aggregate kinetics such as those described in Equations (3), (11), and (17), have structural limitations when applied to complex and highly variable industrial systems, particularly food industry effluents. While these models have proven useful under controlled conditions, their predictive capacity degrades significantly in the face of rapid load variations, compositional heterogeneity, and spatial gradients, as has been systematically documented in recent reviews [62].
In response to these limitations, since 2020 a paradigm shift has been observed towards approaches that seek to integrate greater structural complexity without sacrificing parametric identifiability or operational applicability. This shift does not imply abandoning the conservation principles formulated in Equation (1), but rather extending them through hybrid, multiscale, and spatially explicit frameworks capable of absorbing additional experimental information and representing emergent dynamics not captured by traditional models.
8.1. Hybrid Mechanistic-Data-Based Approaches
One of the most significant developments in recent aerobic process modeling is the integration of mechanistic models with machine learning techniques and advanced statistical modeling. In these approaches, kinetic equations and dynamic balances derived from first principles, such as the systems defined by Equations (17)-(19), provide the physical structure of the model, while empirical components are used to represent unobservable or poorly characterized processes, such as abrupt changes in substrate biodegradability or physiological variations in biomass.
From a formal point of view, these hybrid models can be expressed by a coupled dynamical system in the following way:
where it represents the mechanistic core based on mass balances and biological kinetics, consistent with Equations (17)-(22), and corresponds to an empirical term learned from experimental data using neural networks, Gaussian processes or other nonlinear regression methods.
Recent studies have shown that this type of formulation consistently improves prediction under dynamic conditions, especially in non-stationary load scenarios, without losing interpretability when empirical learning is restricted to structured residuals of the mechanistic model [63]. However, from a critical perspective, these approaches require rigorous regularization and cross-validation strategies, since the indiscriminate incorporation of empirical terms can reintroduce overfitting problems like those discussed in Section 7 for over-parameterized models.
8.2. Spatially Explicit Modeling and Overcoming the Perfect Mixture Hypothesis
Another fundamental aspect of emerging paradigms is the explicit incorporation of spatial dimension. As discussed previously, the assumption of perfect mixing implicit in Equations (17)-(19) and in the general formulation of Equation (22) is an idealization that is rarely met in real, large-scale aerobic systems. Recent experimental measurements and simulations have confirmed the persistent presence of spatial gradients of oxygen, temperature, and substrate, especially in reactors with high particulate organic loading and in food waste composting piles [64].
The natural extension of classical balances leads to formulations based on partial differential equations, which generalize the mass balance defined in Equation (24) towards a spatiotemporal description of the type:
where is the effective diffusion or dispersion coefficient, the velocity field associated with aeration or percolation, and the spatial position-dependent biological reaction term.
Equation (31) allows for the explicit representation of phenomena such as diffusive limitations in solid matrices, local anoxic zones, and decouplings between intrinsic kinetics and transport, which cannot be captured by zero-dimensional models. However, its practical implementation introduces significant computational challenges and spatial identifiability problems, which has motivated the development of order reduction and coupling techniques with simplified CFD models.
8.3. Multiscale Approaches and Reinterpretation of Kinetic Parameters
A third emerging paradigm involves reinterpreting classical kinetic parameters, such as and , not as effective constants, but as emergent quantities resulting from the spatiotemporal average of underlying microbiological processes. This perspective is consistent with the evidence presented in Section 6 and Section 7, which showed that the parametric variability observed in the literature cannot be explained solely by experimental errors.
From a formal point of view, this idea can be expressed by an effective definition of the specific growth rate:
where represents the spatial domain of the system. Equation (32) directly connects spatial formulations such as Equation (31) with the aggregate models discussed in Section 3 to 5, providing a conceptual framework for understanding why parameters estimated under different operating conditions are not directly transferable.
Recent studies combining omics data, dynamic respirometry, and multiscale modeling have shown that this reinterpretation improves consistency between levels of description and reduces empirical dependence on ad hoc correction factors such as those defined in Equation (27) [65].
8.4. Critical Evaluation of New Paradigms
Despite their potential, emerging paradigms do not constitute a universal solution. Hybrid and multiscale models increase data requirements, computational complexity, and the need for rigorous validation. In the absence of sufficient experimental data, these approaches can reproduce the same problems of non-identifiability and predictive fragility discussed in Section 7, albeit under a more sophisticated technological guise.
Consequently, recent literature converges on the idea that progress in aerobic process modeling should not be measured by the number of equations or parameters incorporated, but rather by the coherence between the adopted mathematical structure, the available experimental data, and the model's operational objective. In this sense, new paradigms should be understood as complementary extensions of classical models, not as automatic replacements, reinforcing the need for a critical and progressive approach in the design of models for complex food wastewater effluents [66].
9. Choice of Modeling Approach in Aerobic Digestion
The choice of modeling approach for aerobic digestion processes represents much more than a simple technical decision; it is the central element that determines the model's actual capacity to guide, optimize, and clarify operations in complex, dynamic, and highly heterogeneous systems such as those that characterize waste treatment in the food industry. While there is a tendency to assume that greater modeling sophistication automatically leads to more robust results, evidence shows that this paradigm rarely holds true in practice. In fact, multiple reviews have indicated that the indiscriminate increase in the number of modeled parameters and processes often leads to problems of identifiability and low transferability, and even to reduced operational utility of the model when experimental validation is insufficient [67]. It is imperative to recognize that the goal of mathematical modeling is not the exhaustive recreation of every detail of the process, but rather the generation of reliable information that allows for predicting, controlling, and improving results in real-world industrial operating scenarios [68].
A critical analysis of the literature reveals that, in many cases, the models used are not only selected without justification but rarely undergo formal stages of sensitivity and parametric uncertainty assessment, even though robust methodologies for this have been reported for years [68]. In more than two-thirds of the cases reviewed, the correlation between the complexity of the model and the accuracy obtained is not even discussed, perpetuating poorly substantiated practices that hinder inter-study comparisons and the consolidation of best practices in the field. This is a particularly serious problem with food waste, whose compositional profiles vary widely with origin and seasonality, creating additional challenges for models that assume homogeneity, isotropy, or only consider soluble substrate as raw material [69].
To address these limitations, the specialized literature emphasizes that the starting point for any modeling process must be a thorough characterization of the operational objective and the nature of the residue. Only in this way can an informed decision be made as to whether a simple kinetic model, which ignores the disintegration and hydrolysis of particulate fractions, will suffice, as may occur in approximate simulations for pilot plants under constant conditions, or whether, on the contrary, models integrating sub-stages such as solubilization, cell destruction, and component recalcitrance will be essential , in line with the postulates of the ASM family and contemporary trends in multi-stage modeling [69]. In this regard, it is important to note that the use of Monod equations or more advanced kinetics must be supported not only by theoretical knowledge but also by experimental evidence, since the joint identifiability of parameters related to biomass, substrate, and product can rarely be guaranteed without independent microbial dynamics data [70].
Another key criticism is that, despite the proliferation of models incorporating space-time balances, coupled transport, and even thermal gradient or pH phenomena, most of these developments lack the rigorous validation that can be provided by experimentation at real-world plant scale. They often rely on laboratory data which, while useful for fine-tuning, are not representative of the inherent variability of the industrial environment [71]. Therefore, it is strongly recommended that all modeling processes be accompanied by a comprehensive validation protocol that combines global sensitivity testing, uncertainty analysis, and comparative experimentation across scales. Similarly, the applicability range of each model must be clearly acknowledged, and unvalidated extrapolations beyond the domains for which it was calibrated must be avoided.
Faced with these challenges, hybrid strategies that integrate mechanistic models with machine learning algorithms are particularly promising. This allows for real-time parameter adjustments, the identification of nonlinear patterns that are difficult to model analytically, and an improved model capacity to adapt to unforeseen disturbances, such as abrupt variations in waste flows [72]. However, it should not be overlooked that these approaches, while attractive from a predictive standpoint, present significant challenges in terms of interpretation, computational requirements, and the risk of overfitting, especially when large volumes of representative, labeled data are unavailable. Therefore, the recommended pragmatic solution is to favor models of incremental complexity, incorporating greater detail only when rigorous quantitative analysis demonstrates a real improvement in predictive capacity and usefulness for decision-making [73].
In summary, the true scientific contribution to aerobic digestion modeling transcends the mere construction of complex models and lies in the development of transparent, evidence-based decision-making processes, systematically subjected to uncertainty analysis, and open to iterative refinement as new data or knowledge become available. For readers and professionals facing the task of selecting or building models in this field, it is recommended to explicitly define operational objectives, thoroughly characterize the waste, initially employ simple models subjected to sensitivity assessment and experimental validation, and only scale to higher levels of complexity based on the results obtained, resorting to hybrid and data- driven approaches where traditional systems prove clearly insufficient or unfeasible. This perspective, supported by reference literature and recent experience, fosters not only greater scientific robustness but also the generation of plausible solutions to the specific challenges of waste management in the contemporary food industry.
Table 4 summarizes the distribution of mathematical modeling approaches according to the type of food matrix, revealing clear patterns in the selection of modeling strategies conditioned by the physical characteristics of the waste. First-order kinetic models dominate in solid waste composting systems, where mathematical simplicity and the limited availability of continuous experimental data favor empirical formulations. In contrast, liquid effluents treated with activated sludge show a clear preference for models based on Monod kinetics and structured extensions (ASM), reflecting the greater availability of online sensors and the need for dynamic control in these systems.
Hybrid approaches that integrate machine Learning with mechanistic frameworks show increasing penetration in matrices of high compositional variability, particularly mixed municipal solid waste and large-scale composting systems, where traditional formulations exhibit documented predictive limitations. Partial differential equation (PDE)-based models incorporating spatially resolved heat and mass balances are concentrated in industrial composting applications, consistent with the experimental evidence of thermal and oxygen gradients discussed.
10. Model Selection Framework: From Theory to Industrial Application
The reviewed literature reveals a recurring disconnect between mathematical sophistication and operational utility. Although the advanced approaches described in previous sections represent significant theoretical achievements, their practical application in food wastewater treatment plants requires a systematic selection strategy based on operational criteria, data availability, and design or control objectives.
Recent critical studies have shown that the uncritical adoption of complex models without experimental justification frequently leads to greater predictive uncertainty than simpler, properly calibrated formulations [78]. This modeling paradox reflects structural problems of parametric non-identifiability, discussed extensively in Section 7 and Section 8, which limit the transferability of models beyond their specific calibration conditions [79].
This section synthesizes a pragmatic decision framework that connects specific operational objectives with appropriate modeling approaches, explicitly considering the unavoidable trade-off between biological realism, data requirements, computational cost, and predictive robustness. Table 5 summarizes this integrative synthesis, linking specific industry objectives with mathematical model recommendations based on the evidence analyzed throughout this review.
Decision Matrix: Operational Objectives and Recommended Models
The selection of the modeling approach should be guided primarily by the operational objective for which the model will be used. As shown in Table 5, different industrial purposes demand different levels of mathematical complexity and types of experimental validation. This matrix was constructed through cross-sectional analysis of the mathematical formulations presented above, integrating the identified structural limitations and methodological recommendations from recent specialized literature.
For the preliminary design of reactors, the main objective is to estimate required volumes and hydraulic retention times under constant load conditions, simple first-order or Monod-based kinetic models are sufficient, requiring only COD degradation data from batch experiments. However, these models do not capture substrate inhibition phenomena or variable load effects.
When the objective is to optimize the hydraulic retention time (HRT) in continuous systems, it is necessary to explicitly incorporate hydraulic dynamics through coupled substrate-biomass balances in CSTR reactors (Equations 17-18). These models allow the identification of critical washout conditions (Equation 20) and the evaluation of operational stability under load variations, but they assume perfect mixing, a limit discussed previously and experimentally demonstrated by Zhang et al (2023).
For advanced dynamic control applications requiring real-time prediction under variable load conditions, such as those typical in dairy or meat effluent treatment plants with intermittent discharges, the models in the ASM (Activated) family are ideal. Sludge Models provide the necessary multi-component structure (Equation 22). However, their successful implementation depends critically on the availability of detailed COD fractionation (soluble/particulate, biodegradable/inert) and continuous dissolved oxygen, nitrogen, and phosphorus data. The high parametric dimensionality of these models introduces severe structural identifiability problems when data are limited [80].
Hybrid mechanistic-ML approaches (Equation 30) emerge as a robust alternative for prediction under highly variable loads or in matrices with fluctuating composition, characteristic of waste from the beverage or fruit processing industries. These models combine fundamental mass balances with learned empirical components, improving predictive capacity without completely losing physical interpretability. However, they require dense time series for training and present a risk of overfitting when appropriate regularization techniques are not applied [81].
Finally, composting processes with spatial resolution, where significant thermal and oxygen gradients have been experimentally documented [82]Formulations based on partial differential equations (Equation 31) that incorporate convective-diffusive transport are required. These models, although mathematically rigorous, present high computational cost and problems of spatial parameter identifiability, limiting their application to research studies or the design of novel systems rather than to routine operational control [83].
Table 5 systematically integrates this hierarchy of complexity with practical requirements, providing an operational guide for engineers and scientists facing modeling decisions in real industrial contexts.
11. Conclusions
This comprehensive review underscores that mathematical modeling of aerobic digestion processes in food waste management has progressed from simplified empirical kinetics, such as first-order and Monod kinetics, to hybrid and multiscale paradigms that integrate biochemical mechanisms with machine learning and partial differential equations. These paradigms address structural limitations such as parametric non-identifiability and numerical rigidity inherent in idealized assumptions of homogeneity and perfect mixing. However, a critical gap persists between theoretical complexity and industrial adoption. Factors such as scarce data, stochastic variability in heterogeneous effluents with fluctuating particulate fractions and loads, along with sensor costs , limit applicability, with less than 25% of plants employing formal models. Emerging approaches, such as hybrid mechanistic-ML models represented in Equation (30), offer promise for robust prediction under perturbations, capturing nonlinearities and spatial gradients, but require rigorous validation, regularization against overfitting, and sensitivity analysis to preserve physical interpretability. The proposed framework prioritizes progressive selection based on operational objectives, from preliminary design with simple kinetics to dynamic control with ASM or PDE, balancing biological realism with practicality, and cautions against overparameterization that erodes predictability. Future research should focus on integrating omics data and digital twins to bridge gaps, encouraging adoption in SMEs through accessible tools and standardized calibration protocols. Ultimately, success lies in models not as theoretical ends in themselves, but as operational tools that enhance environmental sustainability and efficiency in the food industry, aligning with global goals for circular waste management and emissions reduction.
References
- Manea, E.E.; Bumbac, C.; Dinu, L.R.; Bumbac, M.; Nicolescu, C.M. Composting as a Sustainable Solution for Organic Solid Waste Management: Current Practices and Potential Improvements. Sustainability 2024, Vol. 16, Page 6329 2024, 16, 6329. [CrossRef]
- Molina-Peñate, E.; Sánchez, A. An Overview of the Technological Evolution of Organic Waste Management over the Last Decade. Processes 2025, Vol. 13, Page 940 2025, 13, 940. [CrossRef]
- Bavaro, T.; Robescu, M.S.; Ma, Y.; Eser, B.E. Bridging Disciplines in Enzyme Kinetics: Understanding Steady-State, Transient-State and Performance Parameters. Catalysts 2025, Vol. 15, Page 1139 2025, 15, 1139. [CrossRef]
- Amirian, M.M.; Irwin, A.J.; Finkel, Z. V. Extending the Monod Model of Microbal Growth with Memory. Front. Mar. Sci. 2022, 9, 963734.
- Altowayti, W.A.H.; Shahir, S.; Eisa, T.A.E.; Nasser, M.; Babar, M.I.; Alshalif, A.F.; AL-Towayti, F.A.H. Smart Modelling of a Sustainable Biological Wastewater Treatment Technologies: A Critical Review. Sustainability 2022, Vol. 14, Page 15353 2022, 14, 15353. [CrossRef]
- Drissi Elbouzidi, A.; Frédéric, R.; Pellerin, R.; Lamouri, S.; Ait El Cadi, A. Leveraging Digital Twins for Enhanced Sustainable Warehouse Management. Cleaner Logistics and Supply Chain 2025, 17, 100287. [CrossRef]
- Kala, Z. Global Sensitivity Analysis of Structural Reliability Using Cliff Delta. Mathematics 2024, Vol. 12, Page 2129 2024, 12, 2129. [CrossRef]
- Pavlovic, M.; Ilic, S.; Antonic, N.; Culibrk, D. Monitoring the Impact of Large Transport Infrastructure on Land Use and Environment Using Deep Learning and Satellite Imagery. Remote Sensing 2022, Vol. 14, Page 2494 2022, 14, 2494. [CrossRef]
- Barasoain-Echepare, Í.; Zárraga-Rodríguez, M.; Podhorski, A.; Villar-Rosety, F.M.; Besga-Oyanarte, L.; Jaray-Valdehierro, S.; Fernández-Arévalo, T.; Sancho, L.; Ayesa, E.; Gutiérrez-Gutiérrez, J.; et al. Mathematical Model for Optimal Agri-Food Industry Residual Streams Flow Management: A Valorization Decision Support Tool. Mathematics 2024, Vol. 12, Page 2753 2024, 12, 2753. [CrossRef]
- Jalal, A.N.; Oniga, S.; Ujvari, B. Advanced Big Data Solutions for Detector Calibrations for High-Energy Physics. Electronics 2025, Vol. 14, Page 2088 2025, 14, 2088. [CrossRef]
- Diangelakis, A.; Lyberatos, G.; Matsakas, L.; Nikoli´c, I.N.; Miji´c, K.M.; Mitrovi´c, I.M. Characteristics of Food Industry Wastewaters and Their Potential Application in Biotechnological Production. Processes 2025, Vol. 13, Page 2401 2025, 13, 2401. [CrossRef]
- Gutiérrez González, J.A.; Fernández Mohedano, A.; Raposo Bejines, F. Evaluating Alternative Oxidants for Artificial Chemical Oxygen Demand Removal Performance from Wastewater Treatment Plants. Resources 2025, 14, 46.
- Liu, X.; Coutu, A.; Mottelet, S.; Pauss, A.; Ribeiro, T. Overview of Numerical Simulation of Solid-State Anaerobic Digestion Considering Hydrodynamic Behaviors, Phenomena of Transfer, Biochemical Kinetics and Statistical Approaches. Energies 2023, Vol. 16, Page 1108 2023, 16, 1108. [CrossRef]
- Rojas-Flores, S.J.; Liza, R.; Nazario-Naveda, R.; Díaz, F.; Delfin-Narciso, D.; Cardenas, M.G.; Alviz-Meza, A. Machine Learning and Hybrid Approaches in the Energy Valorization of Contaminated Sludge: Global Trends and Perspectives. Processes 2026, Vol. 14, Page 363 2026, 14, 363. [CrossRef]
- Saratkar, S.Y.; Langote, M.; Kumar, P.; Gote, P.; Weerarathna, I.N.; Mishra, G. V. Digital Twin for Personalized Medicine Development. Front. Digit. Health 2025, 7, 1583466.
- Safari, A.; Daneshvar, M.; Anvari-Moghaddam, A. Energy Intelligence: A Systematic Review of Artificial Intelligence for Energy Management. Applied Sciences 2024, Vol. 14, Page 11112 2024, 14, 11112. [CrossRef]
- Belik, I.; Bhattacharya, P.; Knudsen, E.S. A Case for Simulated Data and Simulation-Based Models in Organizational Network Research. Res. Policy 2024, 53, 105058. [CrossRef]
- Casagli, F.; Turolla, A.; Batstone, D.J.; Capson-Tojo, G.; Ficara, E.; García, J.; Gonzalez-Flo, E.; Laurent, J.; Lorenz, T.; Pierrelée, M.; et al. Modelling Challenges to Unlock the Power of Phototrophic Systems for Wastewater Valorization. Biotechnol. Adv. 2025, 85, 108709. [CrossRef]
- Abilmazhinov, Y.; Shakerkhan, K.; Meshechkin, V.; Shayakhmetov, Y.; Nurgaliyev, N.; Suychinov, A. Mathematical Modeling for Evaluating the Sustainability of Biogas Generation through Anaerobic Digestion of Livestock Waste. Sustainability (Switzerland) 2023, 15, 5707.
- Gandhi, B.P.; Lag-Brotons, A.J.; Ezemonye, L.I.; Semple, K.T.; Martin, A.D. Development of Mass-Conserving Atomistic Mathematical Model for Batch Anaerobic Digestion: Framework and Limitations. Fermentation 2024, 10, 299.
- Jiang, L.; Chen, M.; Huang, Y.; Peng, J.; Zhao, J.; Chan, F.; Yu, X. Effects of Different Treatment Processes in Four Municipal Wastewater Treatment Plants on the Transport and Fate of Microplastics. Science of The Total Environment 2022, 831, 154946. [CrossRef]
- Ghorbani Bam, P.; Rezaei, N.; Roubanis, A.; Austin, D.; Austin, E.; Tarroja, B.; Takacs, I.; Villez, K.; Rosso, D. Digital Twin Applications in the Water Sector: A Review. Water (Switzerland) 2025, 17, 2957.
- Mate-Kole, E.M.; Dewji, S.A. Mathematical Complexities in Radionuclide Metabolic Modelling: A Review of Ordinary Differential Equation Kinetics Solvers in Biokinetic Modelling. Journal of Radiological Protection 2024, 44, 021001. [CrossRef]
- Lazar, D.; Lu, Z.-H.; Yumnam, P.; Debnath, P. A Review on Mathematical Modeling of Different Biological Methods of Hydrogen Production. Hydrogen 2023, Vol. 4, Pages 881-916 2023, 4, 881–916. [CrossRef]
- Gooneie, A.; Schuschnigg, S.; Holzer, C. A Review of Multiscale Computational Methods in Polymeric Materials. Polymers 2017, Vol. 9, Page 16 2017, 9, 16. [CrossRef]
- Suárez-Carreño, F.; Rosales-Romero, L. Convergency and Stability of Explicit and Implicit Schemes in the Simulation of the Heat Equation. Applied Sciences 2021, Vol. 11, Page 4468 2021, 11, 4468. [CrossRef]
- Liu, P.; Wen, S.; Zhu, S.; Hu, X.; Wang, Y. Microbial Degradation of Soil Organic Pollutants: Mechanisms, Challenges, and Advances in Forest Ecosystem Management. Processes 2025, 13, 916.
- Walling, E.; Vaneeckhaute, C. Greenhouse Gas Emissions from Inorganic and Organic Fertilizer Production and Use: A Review of Emission Factors and Their Variability. J. Environ. Manage. 2020, 276, 111211. [CrossRef]
- Metcalf & Eddy, A. Wastewater Engineering Treatment and Resource Recovery; McGraw-Hill Education., 2014; ISBN 0073401188.
- Liang, C.; Das, K.C.; McClendon, R.W. The Influence of Temperature and Moisture Contents Regimes on the Aerobic Microbial Activity of a Biosolids Composting Blend. Bioresour. Technol. 2003, 86, 131–137. [CrossRef]
- Dominguillo-Ramírez, D.; Aburto, J.; Hugo Leon-Santiesteban, H.; Martinez-Hernandez, E. Neural Network Model for Predicting the Biomethane Yield in an Anaerobic Digester Using Biomass Composition Profiles. Fuel 2023, 344, 128053. [CrossRef]
- Peng, M.Q.; Chen, T.H.; Jin, T.; Su, Y.C.; Luo, S.T.; Xu, H. A Novel First-Order Kinetic Model for Simultaneous Anaerobic–Aerobic Degradation of Municipal Solid Waste in Landfills. Processes 2024, Vol. 12, Page 2225 2024, 12, 2225. [CrossRef]
- Rossetti, I.; Conte, F.; Ramis, G. Kinetic Modelling of Biodegradability Data of Commercial Polymers Obtained under Aerobic Composting Conditions. Eng 2021, Vol. 2, Pages 54-68 2021, 2, 54–68. [CrossRef]
- Lyubimov, V. V. A Method of Qualitative Analysis for Determining Monotonic Stability Regions of Particular Solutions of Differential Equations of Dynamic Systems. Mathematics 2023, Vol. 11, Page 3142 2023, 11, 3142. [CrossRef]
- Salazar, Y.; Valle, P.A.; Rodríguez, E.; Soto-Cruz, N.O.; Páez-Lerma, J.B.; Reyes-Sánchez, F.J. Mechanistic Modelling of Biomass Growth, Glucose Consumption and Ethanol Production by Kluyveromyces Marxianus in Batch Fermentation. Entropy 2023, 25, 497.
- Pigłowska, M.; Kurc, B.; Rymaniak, Ł.; Lijewski, P.; Fuć, P. Kinetics and Thermodynamics of Thermal Degradation of Different Starches and Estimation the OH Group and H2O Content on the Surface by TG/DTG-DTA. Polymers 2020, Vol. 12, Page 357 2020, 12, 357. [CrossRef]
- Solon-Biet, S.M.; Clark, X.; Bell-Anderson, K.; Rusu, P.M.; Perks, R.; Freire, T.; Pulpitel, T.; Senior, A.M.; Hoy, A.J.; Aung, O.; et al. Toward Reconciling the Roles of FGF21 in Protein Appetite, Sweet Preference, and Energy Expenditure. Cell Rep. 2023, 42, 113536. [CrossRef]
- Nyholm, N. A Mathematical Model for Microbial Growth under Limitation by Conservative Substrates. Biotechnol. Bioeng. 1976, 18, 1043–1056.
- Jin, Q. Building Microbial Kinetic Models for Environmental Application: A Theoretical Perspective. Applied Geochemistry 2023, 158, 105782. [CrossRef]
- Liu, S. How Cells Grow. Bioprocess Engineering 2017, 629–697. [CrossRef]
- Wilhelmsen, Ø.; Aasen, A.; Skaugen, G.; Aursand, P.; Austegard, A.; Aursand, E.; Gjennestad, M.A.; Lund, H.; Linga, G.; Hammer, M. Thermodynamic Modeling with Equations of State: Present Challenges with Established Methods. Ind. Eng. Chem. Res. 2017, 56, 3503–3515. [CrossRef]
- Jin, Q.; Wu, Q.; Shapiro, B.M.; McKernan, S.E. Limited Mechanistic Link Between the Monod Equation and Methanogen Growth: A Perspective from Metabolic Modeling. Microbiol. Spectr. 2022, 10. [CrossRef]
- Ferdowsi, M.; Avalos Ramirez, A.; Jones, J.P.; Heitz, M. Elimination of Mass Transfer and Kinetic Limited Organic Pollutants in Biofilters: A Review. Int. Biodeterior. Biodegradation 2017, 119, 336–348. [CrossRef]
- Ahmed, M.A.; Amin, S.; Mohamed, A.A. Current and Emerging Trends of Inorganic, Organic and Eco-Friendly Corrosion Inhibitors. RSC Adv. 2024, 14, 31877–31920. [CrossRef]
- Sellán, S.; Villaverde, E.; Structural, A.F.; Montefusco, F.; Pedersen, M.G.; Procopio, A.; Díaz-Seoane, S.; Sellán, E.; Villaverde, A.F. Structural Identifiability and Observability of Microbial Community Models. Bioengineering 2023, Vol. 10, Page 483 2023, 10, 483. [CrossRef]
- Wade, M.J. Not Just Numbers: Mathematical Modelling and Its Contribution to Anaerobic Digestion Processes. Processes 2020, Vol. 8, Page 888 2020, 8, 888. [CrossRef]
- Lai, J.C.; Then, Y.L.; Hwang, S.S.; Tam, Y.C.; Chua, C.C.N. Modelling Temperature Profiles in Food Waste Composting: Monod Kinetics Under Varied Aeration Conditions. Process Integration and Optimization for Sustainability 2025 9:3 2025, 9, 839–853. [CrossRef]
- Sivaramakrishnan, R.; Kumar, S.; Hosseinzadeh, H.; Balan, V. Harnessing Microalgae from Wastewater for Biocrude via Hydrothermal Liquefaction: A Sustainable Pathway to Biofuels and Biochemicals. Renewable and Sustainable Energy Reviews 2026, 231, 116759. [CrossRef]
- Vandekerckhove, T.G.L.; Bodé, S.; De Mulder, C.; Vlaeminck, S.E.; Boon, N. 13C Incorporation as a Tool to Estimate Biomass Yields in Thermophilic and Mesophilic Nitrifying Communities. Front. Microbiol. 2019, 10, 396249.
- Ingólfsson, H.I.; Bhatia, H.; Aydin, F.; Oppelstrup, T.; López, C.A.; Stanton, L.G.; Carpenter, T.S.; Wong, S.; Di Natale, F.; Zhang, X.; et al. Machine Learning-Driven Multiscale Modeling: Bridging the Scales with a Next-Generation Simulation Infrastructure. J. Chem. Theory Comput. 2023, 19, 2658–2675. [CrossRef]
- Fekih-Salem, R.; Daoud, Y.; Abdellatif, N.; Sari, T. A Mathematical Model of Anaerobic Digestion with Syntrophic Relationship, Substrate Inhibition, and Distinct Removal Rates. 2021, 20, 1621–1654. [CrossRef]
- Bapat, P.M.; Bhartiya, S.; Venkatesh, K. V.; Wangikar, P.P. Structured Kinetic Model to Represent the Utilization of Multiple Substrates in Complex Media during Rifamycin B Fermentation. Biotechnol. Bioeng. 2006, 93, 779–790. [CrossRef]
- Soares, W.S.; dos Santos Magalhães, E.; Govender, N. Enhancing Particle Breakage and Energy Utilization in Ball Mills: An Integrated DEM and SPH Approach. Mining 2025, Vol. 5, Page 18 2025, 5, 18. [CrossRef]
- Tiar, S.M.; Bessedik, M.; Abdelbaki, C.; ElSayed, N.B.; Badraoui, A.; Slimani, A.; Kumar, N. Steady-State and Dynamic Simulation for Wastewater Treatment Plant Management: Case Study of Maghnia City, North-West Algeria. Water 2024, Vol. 16, Page 269 2024, 16, 269. [CrossRef]
- Wang, M.; Chen, X.; Tang, Y.Q.; Nie, Y.; Wu, X.L. Substrate Availability and Toxicity Shape the Structure of Microbial Communities Engaged in Metabolic Division of Labor. mLife 2022, 1, 131. [CrossRef]
- Kang, J.E.; Lee, S.H.; Hong, J.K.; Kim, J.J. Computational Fluid Dynamics Simulation of High-Resolution Spatial Distribution of Sensible Heat Fluxes in Building-Congested Area. Atmosphere 2024, Vol. 15, Page 681 2024, 15, 681. [CrossRef]
- Zhu, G.; Zhang, J.-Y.; Ferreira, V.; Martinez, F.; Jaimes, B.; Carlos Vesga Ferreira, J.; Florez Martinez, A.; Erickson Barbosa Jaimes, J. Linear Optimization Model with Nonlinear Constraints to Maximize Biogas Production from Organic Waste: A Practical Approach. Applied Sciences 2025, Vol. 15, Page 10453 2025, 15, 10453. [CrossRef]
- de la Fuente, I.M. Quantitative Analysis of Cellular Metabolic Dissipative, Self-Organized Structures. International Journal of Molecular Sciences 2010, Vol. 11, Pages 3540-3599 2010, 11, 3540–3599. [CrossRef]
- Kang, M.-S.; Choi, Y.-J.; Hong, K.-H.; Kang, M.-S.; Choi, Y.-J.; Hong, K.-H. Evaluation of the Complex Impact of Major Factors and Derivation of Important Priorities in the Wastewater Treatment Process for Nutrient Removal Using Multiple Regression Analysis. Processes 2025, Vol. 13, Page 833 2025, 13, 833. [CrossRef]
- Valenzuela, K.; Prieto, A.L.; Suárez, F.; de la Fuente, A. Modeling Long-Term Dynamics of Carbon Dioxide and Oxygen in High-Altitude Wetlands. J. Environ. Manage. 2025, 393, 126853. [CrossRef]
- Davies, B.; Szyniszewski, S.; Dias, M.A.; de Waal, L.; Kisil, A.; P Smyshlyaev, V.; Cooper, S.; Kamotski, I. V.; Touboul, M.; Craster, R. V.; et al. Roadmap on Metamaterial Theory, Modelling and Design. J. Phys. D Appl. Phys. 2025, 58, 203002. [CrossRef]
- Roeva, O.; Chorukova, E.; Kabaivanova, L. Advanced Mathematical Modeling of Hydrogen and Methane Production in a Two-Stage Anaerobic Co-Digestion System. Mathematics 2025, Vol. 13, Page 1601 2025, 13, 1601. [CrossRef]
- Muldbak, M.; Gargalo, C.; Krühne, U.; Udugama, I.; Gernaey, K. V. Digital Twin of a Pilot-Scale Bio-Production Setup. Computer Aided Chemical Engineering 2022, 49, 1417–1422. [CrossRef]
- Vereecken, H.; Schnepf, A.; Hopmans, J.W.; Javaux, M.; Or, D.; Roose, T.; Vanderborght, J.; Young, M.H.; Amelung, W.; Aitkenhead, M.; et al. Modeling Soil Processes: Review, Key Challenges, and New Perspectives. Vadose Zone Journal 2016, 15, 1–57. [CrossRef]
- Nordanger, H.; Morozov, A.; Stenhammar, J.; Nordanger, H.; Morozov, A.; Stenhammar, J. Interplay between Brownian and Hydrodynamic Tracer Diffusion in Suspensions of Swimming Microorganisms. J. Fluid Mech. 2023, 974, A25. [CrossRef]
- Zhao, T.; Wang, S.; Ouyang, C.; Chen, M.; Liu, C.; Zhang, J.; Yu, L.; Wang, F.; Xie, Y.; Li, J.; et al. Artificial Intelligence for Geoscience: Progress, Challenges, and Perspectives. The Innovation 2024, 5, 100691. [CrossRef]
- Wang, J.; Qiao, Z. A Comprehensive Review of Landfill Leachate Treatment Technologies. Front. Environ. Sci. 2024, 12, 1439128. [CrossRef]
- Ginocchi, M.; Ponci, F.; Monti, A. Sensitivity Analysis and Power Systems: Can We Bridge the Gap? A Review and a Guide to Getting Started. Energies 2021, Vol. 14, Page 8274 2021, 14, 8274. [CrossRef]
- Gartner, T.E.; Jayaraman, A. Modeling and Simulations of Polymers: A Roadmap. Macromolecules 2019, 52, 755–786. [CrossRef]
- Santander, M.; Chica, V.; Correa, H.A.M.; Rodríguez, J.; Villagran, E.; Vaillant, F.; Escobar, S. Unravelling Cocoa Drying Technology: A Comprehensive Review of the Influence on Flavor Formation and Quality. Foods 2025, Vol. 14, Page 721 2025, 14, 721. [CrossRef]
- Yuan, X.; Niu, G.; Yin, J.; Xie, Y. Data Assimilation in Hydrological Models: Methods, Challenges and Emerging Trends. Hydrology 2025, Vol. 12, Page 323 2025, 12, 323. [CrossRef]
- Nawaz, W.; Li, Z. Complexity to Resilience: Machine Learning Models for Enhancing Supply Chains and Resilience in the Middle Eastern Trade Corridor Nations. Systems 2025, Vol. 13, Page 209 2025, 13, 209. [CrossRef]
- Javed, H.; El-Sappagh, S.; Abuhmed, T. Robustness in Deep Learning Models for Medical Diagnostics: Security and Adversarial Challenges towards Robust AI Applications. Artificial Intelligence Review 2024 58:1 2024, 58, 12-. [CrossRef]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The Links between Gut Microbiota and Obesity and Obesity Related Diseases. Biomedicine & Pharmacotherapy 2022, 147, 112678. [CrossRef]
- Henze, M.; Gujer, W.; Mino, T.; van Loosedrecht, M. Activated Sludge Models ASM1, ASM2, ASM2d and ASM3. Water Intelligence Online 2006, 5, 9781780402369–9781780402369. [CrossRef]
- Pastor-Poquet, V.; Papirio, S.; Steyer, J.P.; Trably, E.; Escudié, R.; Esposito, G. Modelling Non-Ideal Bio-Physical-Chemical Effects on High-Solids Anaerobic Digestion of the Organic Fraction of Municipal Solid Waste. J. Environ. Manage. 2019, 238, 408–419. [CrossRef]
- Ponsá, S.; Puyuelo, B.; Gea, T.; Sánchez, A. Modelling the Aerobic Degradation of Organic Wastes Based on Slowly and Rapidly Degradable Fractions. Waste Management 2011, 31, 1472–1479. [CrossRef]
- Zhang, J.; Wu, Z.; Huang, Y.; Zhan, X.; Zhang, Y.; Cai, C. Industrial-Scale Composting of Swine Manure with a Novel Additive-Yellow Phosphorus Slag: Variation in Maturity Indicators, Compost Quality and Phosphorus Speciation. Bioresour. Technol. 2023, 384. [CrossRef]
- Stoudt, S.; de Valpine, P.; Fithian, W. Nonparametric Identifiability in Species Distribution and Abundance Models: Why It Matters and How to Diagnose a Lack of It Using Simulation. Journal of Statistical Theory and Practice 2023 17:3 2023, 17, 39-. [CrossRef]
- Monday, C.; Zaghloul, M.S.; Krishnamurthy, D.; Achari, G. A Review of AI-Driven Control Strategies in the Activated Sludge Process with Emphasis on Aeration Control. Water 2024, Vol. 16, Page 305 2024, 16, 305. [CrossRef]
- Lwele, E.; Shenfield, A.; da Silva, C.E. AI-Based Surrogate Models for the Food and Drink Manufacturing Industry: A Comprehensive Review. Processes 2025, Vol. 13, Page 2929 2025, 13, 2929. [CrossRef]
- Lhaj, M.O.; Moussadek, R.; Sanad, H.; Manhou, K.; Lhaj, M.O.; Alaoui, M.M.; Zouahri, A.; Mouhir, L. Ecological and Microbial Processes in Green Waste Co-Composting for Pathogen Control and Evaluation of Compost Quality Index (CQI) Toward Agricultural Biosafety. Environments 2026, Vol. 13, Page 43 2026, 13, 43. [CrossRef]
- Jahin, A.; Zidan, A.H.; Bao, Y.; Liang, S.; Liu, T.; Zhang, W. Evaluating Mathematical Reasoning Across Large Language Models: A Fine-Grained Approach. 2025.
- Naghavi, M.; Ong, K.L.; Aali, A.; Ababneh, H.S.; Abate, Y.H.; Abbafati, C.; Abbasgholizadeh, R.; Abbasian, M.; Abbasi-Kangevari, M.; Abbastabar, H.; et al. Global Burden of 288 Causes of Death and Life Expectancy Decomposition in 204 Countries and Territories and 811 Subnational Locations, 1990–2021: A Systematic Analysis for the Global Burden of Disease Study 2021. The Lancet 2024, 403, 2100–2132. [CrossRef]
- Mbamba, C.K.; Lindblom, E.; Flores-Alsina, X.; Tait, S.; Anderson, S.; Saagi, R.; Batstone, D.J.; Gernaey, K. V; Jeppsson, U. Plant-Wide Model-Based Analysis of Iron Dosage Strategies for Chemical Phosphorus Removal in Wastewater Treatment Systems. Water Res. 2019, 155, 12–25.
- Awasthi, M.K.; Ganeshan, P.; Gohil, N.; Kumar, V.; Singh, V.; Rajendran, K.; Harirchi, S.; Solanki, M.K.; Sindhu, R.; Binod, P.; et al. Advanced Approaches for Resource Recovery from Wastewater and Activated Sludge: A Review. Bioresour. Technol. 2023, 384, 129250. [CrossRef]
Figure 1.
Conceptual hierarchy of mathematical modeling approaches in aerobic digestion.
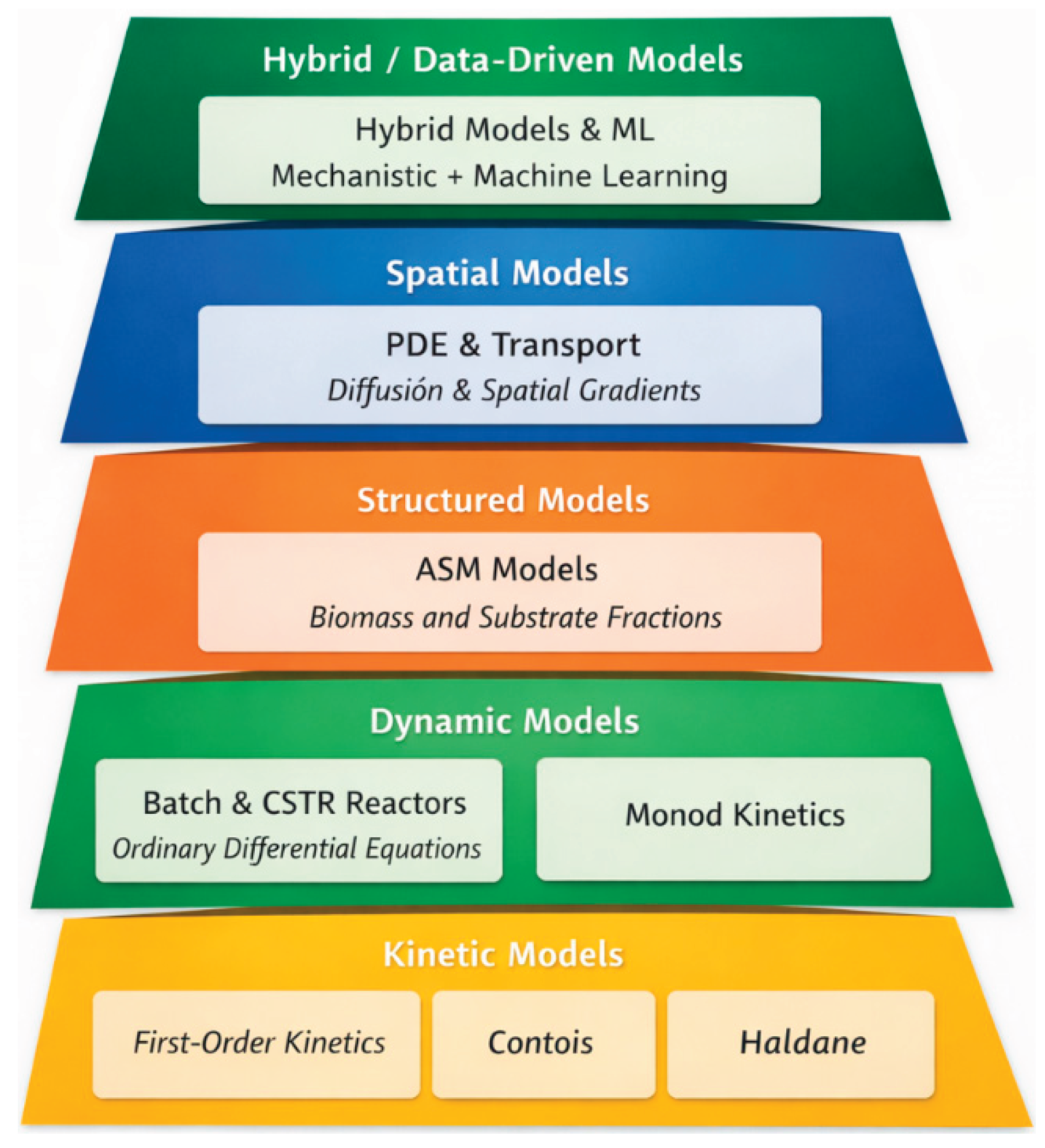

Table 1.
Common assumptions in mass balances and their impact on aerobic digestion models of food effluents.
Table 1.
Common assumptions in mass balances and their impact on aerobic digestion models of food effluents.
| Modeling assumption | Level of adoption in the literature | Direct mathematical implication | Consequence in food effluents | References |
|---|---|---|---|---|
| Perfect blend (0D model, no spatial resolution) | High (>60%) | Reduction of the system to ODEs without spatial gradients | Underestimation of heterogeneity in effluents with high solids content and oxygen-limited zones | (Walling & Vaneeckhaute, 2020) |
| Constant volume | Very high (>80%) | Elimination of volumetric variation terms | Errors in predicting processes involving evaporation, compaction, or gas generation | [29] |
| Homogeneous medium | High (~70%) | Absence of internal diffusion and convection | Inability to reproduce thermal and oxygen gradients in reactors and compost piles | [30] |
| Uniform continuous flow | Moderate (~45%) | Constant hydraulic forcing | Overestimation of efficiency in systems with intermittent discharges typical of the food industry | [31] |
Note: The 0D model (zero-dimensional model) is associated with statements without spatial resolution where all variables depend only on time and the system is considered to be completely mixed.
Table 2.
Comparison of kinetic models of microbial growth in aerobic systems.
| Model | Main dependency | Mathematical advantage | Structural limitation | Applicability in food effluents |
|---|---|---|---|---|
| Monod | Substrate (S) | Simplicity, stability | It does not consider inhibition or biomass | Suitable for simple effluents |
| Tessier | Substrate (S) | Smooth transition | Less physiological interpretation | Systems with gradual response |
| Contois | Substrate and biomass (S, X) | Captures density effects | Greater nonlinearity | Effluents with high biomass |
| Andrews/Haldane | Inhibitory substrate | Describe toxicity | Parameters that are difficult to identify | High organic loads |
Table 3.
Critical kinetic parameters and their sensitivity in structured aerobic models.
| Parameter | Units | Typical range | Reported sensitivity | Impact on food waste |
|---|---|---|---|---|
| day−1 | 2-12 | High | Variations of up to 400% between carbohydrates and lipids | |
| mg·L−1 | 5-50 | Medium-high | Strong dependence on substrate type | |
| mgX·mgS−1 | 0.3-0.8 | Average | Reductions of 30-50% in the presence of inhibitors | |
| b | day−1 | 0.05-0.3 | Low-medium | Non-linear increase with temperature |
| K h | day - 1 | 0.05-0.5 | High in solids | Biphasic kinetics in lignocellulosic materials |
Table 4.
Distribution of mathematical modeling approaches according to food matrix type.
| Matrix type | First-order models | Kinetic models | Heat/mass balances (PDE) | Hybrid approaches (ANN/ML) | Main references |
|---|---|---|---|---|---|
| Composting of food waste | High frequency (>12 studies) | Moderate | Low | Emergent | [30] |
| Composting of organic solid waste | High frequency (>10 studies) | Moderate | Moderate | Low | [74] |
| Composting of plant waste/vineyards | Moderate (5-8 studies) | High | Low | Very low | (Walling & Vaneeckhaute, 2020) |
| Aerobic activated sludge | Low | High frequency (>15 studies) | Moderate | Emergent | [75] |
| Mixed municipal solid waste | Moderate | Moderate | High | High (>5 studies) | (Pastor- Poquet et al., 2019) |
| Dairy/meat effluents | Low | High frequency | Low | Moderate | (Ponsá et al., 2011) |
| Large-scale composting systems | Moderate | Moderate | High frequency (>8 studies) | Moderate | [78] |
Table 5.
Framework for selecting mathematical models for aerobic digestion according to operational objectives and data availability.
Table 5.
Framework for selecting mathematical models for aerobic digestion according to operational objectives and data availability.
| Operational objective | Recommended mathematical model | Equations | Minimum required data | Main advantages | Critical limitations | References |
|---|---|---|---|---|---|---|
| Preliminary reactor design | First-order kinetics | Equation (3): |
Batch COD time series (≥3 replicates) | Mathematical simplicity; analytical solution; rapid calibration | No dynamic biomass capture; substrate inhibition failure; parameter lacks physiological interpretation | (Walling & Vaneeckhaute , 2020) |
| Load capacity estimate | Monod model ( batch ) | Equations (11-13) |
COD, total biomass, yield Y ( controlled batch experiments ) | Captures kinetic saturation; allows estimation of y ; based on structured models | Assumes a homogeneous microbial population; does not consider particulate fractions or inhibition | [29] |
| TRH optimization in CSTR | Continuous coupled balances | Equations (17-18) with hydraulic term | COD influent/effluent, biomass, flow rate (continuous data ≥7 days) | washout conditions ( Eq . 20); captures hydraulic-kinetic interaction; dynamic stability | It assumes a perfect blend; it ignores spatial gradients; it is sensitive to variations in |
( Naghavi et al., 2024) |
| Advanced control with variable load | ASM2d or ASM3 (multi-component) | Equation (22) |
COD fractionation (soluble/particulate), N, P, dissolved O2, pH (online sensors) | Standardized IWA framework; nitrification/denitrification capture; internationally validated | High dimensionality (>20 parameters); identifiability problems; requires costly experimental fractionation | [75] |
| Prediction under fluctuating composition | Mechanistic-ML hybrid | Equation (30): |
Dense time series (>500 points); multiple operating variables; cross-validation | Captures complex nonlinearities; adapts to disturbances; improves prediction in uncalibrated regimes | Risk of overadjustment; requires regularization; partial loss of physical interpretability | [85] |
| Composting with spatial gradients | PDE with transport | Equation (31) |
Spatial profiles of T, O2, humidity (≥5 positions); thermal properties of the medium | It represents real spatial heterogeneity; it captures diffusive limitation; it predicts anoxic zones | High computational cost; limited spatial identifiability; difficult experimental validation | [78] |
| Parametric sensitivity analysis | Extensions with environmental corrections | Equations (27–29) |
Factorial experiments T, humidity, O2 (orthogonal design) | Quantifies environmental effects; improves extrapolation; identifies dominant parameters | Assumes separability of effects (rarely valid); introduces additional parametric correlations | [86] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



