
Submitted:
04 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Knowledge on the biology of the coastal spotted dolphin Stenella attenuata graffmani from western South America, one of the least studied delphinid taxa, is summarized for the first time. Information was obtained largely opportunistically during field research targeting other species or during whale watching. Life history is discussed including distribution (Colombia, Ecuador, Peru, Chile), external morphology and pigmentation, cranial characteristics, reproduction and growth, feeding, behavioral ecology, parasites and pathology, but also genetic diversity, population structure and fisheries interactions. As a (sub)tropical dolphin its habitual austral range is the Piura Department, northern Peru, but under El Niño conditions it can reach Lima Province (Cerro Azul at 13°02’S). No evidence was found for Chile. The coastal subspecies is heavily spotted and substantially bigger, as reflected in body length and all cranial measurements. Condylobasal length, mandibular ramus length and height do not even overlap with the offshore S. a. attenuata morph. Phylogenetic analysis of the hypervariable part of mtDNA control region, revealed Ecuadorian coastal spotted dolphins to be distributed across seven different clades, with most belonging to the three largest clades, while only one haplotype was found in each of the remaining four. This pattern suggests a complex evolutionary history, possibly indicating substructures within the population or an ongoing evolutionary process. The presence of haplotypes in multiple clades may reflect high genetic diversity, likely influenced by dolphin migration between nearby geographic areas, facilitating haplotype exchange and contributing to the observed distribution.
Keywords:
Delphinidae
; life history
; Colombia
; Ecuador
; Peru
; SE Pacific
; bycatch
; distribution
Introduction
The oceanic dolphin genus Stenella Gray, 1866 (Delphinidae) has been well-studied in the offshore Eastern Tropical Pacific (e.g. Douglas et al., 1984; Dizon et al., 1994; Hohn et al., 1985; Gerrodette and Palacios, 1996; Perrin, 1975, 2001, 2018; Gerrodette and Forcada, 2005) but received minimal focussed attention in coastal Pacific South America (e.g. Dizon et al., 1994; Van Waerebeek et al., 1998; Hamilton et al., 2009). Specifically, the distribution and biology of the pantropical spotted dolphin Stenella attenuata (Gray, 1846) from coastal waters of western South America, i.e. Colombia, Ecuador, Peru and Chile, has not been studied in any detail, partially due to a scarcity of documented cases, until recently (Londoño, 2005; Valencia, 2006; García et al., 2006, 2008; Hamilton et al., 2009; Martínez et al., 2021; Cortez-Casamayor et al., 2024).
Swedish zoologist Einar Lönnberg (1934) described a new delphinid species Prodelphinus graffmani from a spotted dolphin captured by J. Holger Graffman in nearshore waters 20 miles north of Acapulco, Guerrero, Mexico (Figure 1). Taxonomist William Perrin (1969, 1975), followed by Rice (1998), synonymized Prodelphinus graffmani (Lönnberg, 1934) with Stenella attenuata (Gray, 1846). Subsequently, Perrin (2001) then designated two subspecies, a cosmopolitan pelagic Gray’s pantropical spotted dolphin Stenella attenuata attenuata (Gray, 1846) (hereafter: SAA) and an eastern Pacific coastal spotted dolphin Stenella attenuata graffmani (Lönnberg, 1934) (hereafter: SAG), or briefly, coastal spotted dolphin (Dizon et al., 1994; Carwardine, 2020). In Latin America they are referred to as delfín manchado costero or delfín moteado costero. Castro and Van Waerebeek (2019) first assigned heavily spotted stranded specimens in Ecuador as SAG. No other S. attenuata subspecies have been described world-wide.
The distribution of SAG in the NE Pacific has been characterized as coastal and inshore waters within about 25 km from land, between the Gulf of California and Colombia (Perrin, 1969, 1975, 2018; Rice, 1998; Flórez-González et al. 2004; Jefferson et al., 2015). A continuous distribution is supported along the coasts, with observations from Ensenada (31º30’N), NW Baja California, Mexico (Vidal et al., 1993), Guaymas, Gulf of California (Mexico), Guatemala (Ortiz-Wolford et al., 2021), El Salvador (Ibarra-Portillo et al., 2020), Nicaragua (de Weerdt, 2021), Costa Rica (Rodríguez-Fonseca and Cubero-Pardo, 2001), Panama (Aguilar et al., 1997; Justines, 1997; García et al., 2008) and Colombia (Perrin, 2018; Hamilton et al., 2009).
SAG individuals of the eastern Pacific are heavily spotted, significantly more than SAA offshore stocks, hence the dorsal cape may be practically obliterated by dense lightly-colored spotting in large individuals. Spotting may extend to dorsal fin and flippers. The smallish dorsal fin is strongly falcate and relatively narrow (Perrin, 1969, 1975, 2001; Carwardine, 2020). All mensural cranial characters of the coastal form are substantially greater than those of offshore stocks, concomitant with the larger body size. SAG has slightly lower tooth counts while tooth width is significantly greater (Perrin, 1969, 1975, 2001; Douglas et al., 1984; Perrin et al., 1987, 1994).
Based on haplotypic diversity, it was proposed that SAG distributed along the Eastern Pacific is divided into four different populations: northern Mexico, Central America, Costa Rica and Ecuador, being the southern populations that diverged first from the offshore form (Escorza-Treviño et al., 2005). These authors included seven spotted dolphin samples (unknown subsp.) from unspecified locations in Ecuador and pooled them with specimens from Panama. However, they admitted that some structure might exist within this area which could be unveiled as the sampling gaps are filled (Escorza-Treviño et al. 2005). Since the latter study included few samples from Ecuador, further efforts are needed to properly characterize the southern population molecularly. The northern Mexican stock did not significantly differentiate from offshore (SAA) animals and further sampling was recommended (Escorza-Treviño et al. 2005).
In the Southeast Pacific, Stenella attenuata has been briefly reported from captures and strandings in Ecuador and Peru (Félix and Samaniego, 1994; Van Waerebeek and Reyes, 1994; Chiluiza et al., 1998; Félix et al., 2011; Castro and Van Waerebeek, 2019). Spotted dolphins accounted for 9.42% of the stranding records on the coast of Ecuador between 1996 and 2009 and recorded throughout the year with no seasonal pattern (Félix et al., 2011). The species is observed with regularity over the shelf on the central coast of Ecuador between Isla de la Plata (01º15´S) - Puerto López (01°33’S) and Salinas (02º10’S) (FF and CC, unpublished information) although most probably its distribution is continuous along the entire coast of the country. Stenella attenuata is observed in waters of the Colombian Pacific throughout the year; however, its relative abundance is higher during the first quarter, when ocean productivity is greater (Rodríguez-Rubio et al., 2003). During research cruises conducted in both semesters between 2006 and 2009, S. attenuata exhibited the second highest relative abundance (0.21 groups/100 km), after the striped dolphin Stenella coeruleoalba, and was associated with waters characterized by higher chlorophyll-a concentrations and a stronger, shallower thermocline (Herrera, 2009). In Colombia, Stenella attenuata graffmani is resident in the waters surrounding Gorgona Island and is present year-round, typically in small groups of up to ten individuals, although it is occasionally observed forming herds of more than one hundred individuals (Capella et al., 2014). It can also be found at the Negritos shoals in Bahía Málaga, the Gulf of Tribugá and Cupica, and the Utría Bight (Herrera et al., 2011a).
Life history, genetics and population structure information are essential for developing conservation and management strategies. Here we compile spotted dolphin records from Peru, Ecuador and Colombia, hence update distribution range and status, document (scarce) natural history information, and review literature. We also include a genetic analysis (mtDNA) of samples collected in central Ecuador aiming at defining stock identity and population structure.
Material and Methods
Study Area
The study area that was examined for potential SAG records comprises the western South American (WSA) coast including continental shelf waters from Chocó, northern Colombia, Ecuador, Peru extending for approximately 8,000 km, to southern Chile (Figure 2). The climate across this vast region ranges from tropical in the north (Colombia and Ecuador), subtropical in northern and central Peru, temperate in southern Peru and most of Chile, and subantarctic in southern Chile. Surface circulation in the north is dominated by the North Equatorial Current and North and South Equatorial Counter currents, while from Ecuador to southern Chile it is influenced by the Peruvian or Humboldt Current (Wyrtki, 1966). In addition to seasonal shifts in the position of the Intertropical Convergence Zone, the region is regularly affected by El Niño–Southern Oscillation (ENSO) events (NOAA, 2025). During its warm phase (El Niño), sea surface temperature can rise by up to 4 °C above the mean, whereas during its cold phase (La Niña) it may fall by up to 2 °C below the mean (Fiedler, 2002). This cycle is also associated with changes in winds, rainfall, thermocline depth, circulation, and biological productivity, with significant effects on the distribution of marine species, including seabirds, small pelagic fish, and marine mammals (Barber & Chávez, 1983; Ramírez & Urquizo, 1985; Fiedler, 2002).
Source of Data
While significant information is available on natural history parameters, and especially morphological variation and reproductive biology, for the SAA offshore ETP populations (e.g. Hohn et al., 1985; Perrin and Hohn, 1994; Perrin et al., 1987; Perrin, 1975, 2001), equivalent knowledge for the SAG subspecies remains minimal (Douglas et al., 1984; Perrin, 2018). Original data utilized in this paper are derived mainly from spotted dolphin specimens bycaught in small-scale fisheries (Van Waerebeek et al., 1988; Félix and Samaniego, 1994; Van Waerebeek and Reyes, 1994) and stranding surveys implemented along much of the Peruvian and Ecuadorian coasts in 1985–2017 (e.g. Chiluiza et al., 1998; Félix et al., 2011; Castro and Van Waerebeek, 2019; Van Waerebeek et al., 2018; and others see below). Of skulls curated at Museo de Ballenas, Salinas, at the Parque Nacional Machalilla, Puerto López (Ecuador) and at Museo de Delfines, CEPEC, Pucusana, Peru, standard cranial measurements were taken with callipers (to nearest 0.5 mm) according to Perrin (1975), with a few additions, ensuring optimal comparability with published data from the ETP.
Sighting data off central Ecuador were collected during commercial whale watching trips and research surveys targeting humpback (Megaptera novaeangliae) and Bryde’s whales (Balaenoptera edeni/brydei) (Félix et al., 2025). Sightings in Colombia have been conducted aboard oceanographic vessels of the General Maritime Directorate (DIMAR) during ERFEN cruises, as well as from smaller research vessels, tourism boats, and fishing vessels (Herrera, 2009; Herrera and Flórez-González, 2018; Herrera et al., 2007, 2011b). Off northern Ecuador and off Colombia equivalent data were collected by the Southwest Fisheries Science Center (SWFSC) during the Eastern Tropical Pacific Cetacean and Ecosystem surveys conducted between 1986 and 2005 (Hamilton et al., 2009) and were downloaded from the Ocean Biodiversity Information System (OBIS).
Genetic Analysis
Twenty-nine skin samples, collected from 2006 to 2024 on the central coast of Ecuador, were used for genetic analysis (Table S1). Dolphins were thought to be SAG and included stranded (n= 13) and free-ranging (n= 16) individuals. Samples were collected by swabbing the dorsolateral region using a 3 m extendable tube with a 5 cm-wide strip of #35 sandpaper at the tip, a modified version of the technique described by Bayas-Rea et al. (2018). This technique enabled sampling of dolphins both above and below water. Small epidermal tissue samples, ranging from a few mm² to approximately 1 cm², were collected and preserved in 95% ethanol. All genetic analyses were conducted at the Molecular Laboratory of the Pontificia Universidad Católica del Ecuador (PUCE), Quito.
DNA was extracted, and the hypervariable region of mtDNA control region (CR) (~602 bp; n = 21 and ~727 bp; n = 8) was amplified using two primer sets: (1) H00034 (Rosel et al., 1994) and L15824 (Rosel et al., 1999) and (2) t-pro-whale and Dlp8G (Dalebout et al., 1998). Forward and reverse strands of each sequenced sample were manually edited and aligned to obtain a consensus sequence using MEGA 11 (Tamura et al., 2021). To validate sequence identity, all consensus sequences were evaluated with the BLAST algorithm in GenBank (Altschul et al., 1997). Finally, multiple sequence alignment for each marker was performed using CLUSTAL W, implemented in MEGA 11 (Tamura et al., 2021). Since sequences varied in length, all molecular analyses were based on a 405 bp truncated alignment. Genetic diversity was estimated by calculating the number of haplotypes using DNAsp v6 (Rozas et al., 2017). Haplotype diversity (h), nucleotide diversity (π), Tajima’s D test (Tajima, 1989), and Fu’s Fs test (Fu, 1997) for selective neutrality were estimated using Arlequin v3.5 (Excoffier & Lischer, 2010). Significance of both neutrality tests was assessed through randomization (10,000 iterations).
To investigate the phylogeographic relationships among Ecuadorian haplotypes and other populations of S. attenuata elsewhere, 101 mtDNA CR sequences from different geographic regions available in the GenBank database were included (Courbis et al., 2014; Escorza-Treviño et al., 2005; Leslie et al., 2016; Oremus et al., 2015), along with 27 sequences obtained from this study (Table S2). Genealogical relationships were inferred using a median-joining network implemented in Network v.10 software (Bandelt et al., 1999). To infer phylogenetic relationships, Bayesian inference with MrBayes v.3.2.7 was used (Ronquist et al., 2012). The analysis included 57 mtDNA CR sequences of S. attenuata (accession numbers in Table 1) (Escorza-Treviño et al., 2005; Leslie et al., 2016), along with two sequences of S. longirostris (NC_032301.1 and KX857382.1), one sequence of S. coeruleoalba (NC_012053.1), one sequence of S. frontalis (NC_060612.1), and one sequence of Lagenorhynchus albirostris (NC_005278.1) as an outgroup. The best-fitting model for nucleotide evolution (HKY+I+G) was inferred under the AICc criterion using jModelTest v.2.1.10 (Darriba et al., 2012). Posterior probabilities of the tree and parameters in the evolutionary model were approximated using Markov Chain Monte Carlo (MCMC). Two independent runs of four chains were carried out for 5,000,000 generations, with a 100,000 burn-in and sampling every 5,000 generations. The effective sample size (ESS > 200) values were evaluated using Tracer v.1.6 (Rambaut et al., 2018) to ensure adequate mixing and convergence of the posterior distribution and parameters. Additionally, the potential scale reduction factor (PSRF = 1) and the average standard deviation of split frequencies between chains (≤ 0.01) were examined. The phylogenetic tree was visualized and edited using Fig (Tree) v.1.4.2 (Rambaut, 2014).
Results and Discussion
Distribution Off Western South America
Colombia
Perrin (1975) indicated that the southernmost known specimen of S. attenuata (USNM 258641) from Colombia was collected at Isla Gorgona (03°0’N, 78°20’W), one of the earliest specimens. Valencia (2006) studied the behavior of spotted dolphins around Isla Gorgona, without discussing subspecific status. García et al. (2006) reported 11 sightings of SAA and indeterminate subsp. and one confirmed sighting of SAG (n= 3) in Colombian waters, including towards Malpelo Island (04°00’12”N, 81°36’27”W), ca. 500 km offshore. A follow-up abstract indicated the high risk of fisheries interactions for coastal spotted dolphins (García et al., 2008). The pantropical spotted dolphin was the third most abundant cetacean recorded and the second delphinid after the striped dolphin (Stenella coeruleoalba) in both coastal and oceanic waters of Colombia during surveys conducted between 2001 and 2010 from the mainland to Malpelo Island (Herrera et al., 2011 b).
Palacios et al. (2012) estimated abundance of S. attenuata in the Colombian EEZ, without distinguishing between subspecies. In contrast with common bottlenose dolphin Tursiops truncatus (Montagu) that occurred nearshore, Londoño (2005) did not observe spotted dolphins inside Bahía Málaga (03°58’N, 77°19’W), Valle del Cauca Department, but at a mean distance from the coastline of 9.1 km (range 1–30 km), indicating separate habitats. Herrera et al. (2011 b) also indicate that it is possible to find the species in the external part of the bay at Negritos Bank. Only one small group (n=5) was sighted at 1 km from shore, while mean group size was 22 (range 2-100). Spotted dolphins were present off Bahía Malaga in both dry and wet seasons and presumably year-round. A similar situation was found in the Parque Nacional Utría, Chocó Department (Suárez, 1994). However, in none of these studies was it attempted to distinguish between coastal and oceanic subspecies. SAG would have a continuous distribution along the entire coast of Colombia according to sightings registered by the SWFSC (Hamilton et al., 2009) (Figure 2).
Ecuador
Stenella attenuata has a regular occurrence in offshore waters off Ecuador (Table 1 and Table 2; Figure 2), but there is no indication of abundance in coastal waters. However, Gerrodette and Palacios (1996) estimated the abundance of spotted dolphins within the Ecuadorian EEZ at 1,864 animals (CI 95% 567-6427), remaining unclear what proportion corresponds to the coastal SAG. The species has been reported also off the Galápagos Islands (Perrin et al. 1987; Day, 1994; Palacios and Salazar, 2002; Denkinger et al., 2013) especially outside the Galápagos Marine Reserve (40 nm offshore) where it was regularly seen in mixed aggregations with spinner dolphins (Stenella longirostris) (Palacios and Salazar, 2002) and would correspond to the offshore subspecies. Among the four small odontocete species captured in artisanal drift gillnets in 1993, only 2% were identified as S. attenuata (Félix and Samaniego, 1994). In contrast, Castro and Rosero (2010) reported that spotted dolphins accounted for 22% of the cetacean bycatch from small-scale fishing fleets along the central coast of Ecuador. This discrepancy is likely related to the fishing depth at which different fleets operated, as the continental shelf is narrower in some areas. Most probably virtually all specimens bycaught in the Ecuadorian small-scale fisheries belong to SAG.
The neritic subspecies seems to primarily occur over the shelf along the ocean-exposed central provinces of Santa Elena and Manabí, although this could be explained by the lower research effort in northern Ecuador and the southwestern Gulf of Guayaquil, the most productive zone along the Ecuadorian coast (Chinacalle et al., 2021). No SAG catches were reported, for example, for the southern port of Puerto Bolívar (Van Waerebeek et al., 1997) but some remains were found in the northeastern part of the gulf, at Playas (Chiluiza et al., 1998).
Peru
The first specimen records of SAG in central and northern Peru date from, respectively, 1986 and 1989, (Table 3), decades before free-ranging animals were sighted in northern Peru (Pacheco et al., 2019; Martínez et al., 2021; Cortez-Casamayor, 2024). Among thousands of small cetaceans taken in Peru’s mostly small-scale coastal fisheries, examined in 1985-2010 (e.g. Read et al., 1988; Van Waerebeek et al., 1988, 1994; Reyes, 2009; Mangel et al., 2010; Tzika et al., 2010), only four individuals of S. attenuata were authenticated (Table 3). Stranding surveys in southern and south-central Peru (Tacna and Ica Departments) did not encounter any spotted dolphin records (Van Waerebeek and Reyes, 1994; Van Waerebeek et al., 2018; Reyes, 2009; Pizarro-Neyra, 2010; Pizarro-Neyra et al., 2021; Santillán, 2021). Surveys of beaches and fishing ports in Peru’s three northern coastal departments, i.e. Tumbes, Piura and Lambayeque (Van Waerebeek et al., 1988; Van Waerebeek and Reyes, 1994; unpublished CEPEC archives) yielded a single skull from Bahía Sechura in 1989 (Table 3).
Seasonal observer effort from humpback whale-watching trips in nearshore waters off El Ñuro (04°14’S, 81°10.5′W) and Los Órganos (04°29.5’S, 81°20.778’W), Piura Province, did not result in any spotted dolphin sightings in winter and early spring months (Silva-Buse, 2016; Pacheco et al., 2019). However, in 2018-2019, a single spotted dolphin was sighted among a short-beaked common dolphin Delphinus delphis pod off Los Órganos thrice, in September, October and January (Martínez et al., 2021). The peculiarity of these encounters suggests these are most likely re-sightings of a single individual. The habitual southern range of SAG is probably Piura, with the cold coastal Humboldt Current acting as barrier. The three capture records in central Peru, in February and April 1986, occurred during the rapid warming phase of an exceptionally lengthy 1986-1988 El Niño event (NOAA, 2025). An increase of 85% in sightings of spotted dolphins in northern Peru was associated with warm waters intrusion during a strong El Niño event in 2023 (Cortez-Casamayor et al., 2024). The documented austral range for SAG is off Cerro Azul (13°02’S) at the southern end of the Lima Department, based on a net-entangled mother-calf pair (Figure 3). The veteran fisher who hand-harpooned AJR-036 from a pod of three animals just off the beach of Punta Negra, Lima, (Table 3) emphasized that he had ‘never seen dolphins of this type before’.
Chile
Although Sielfeld (1980) reported no S. attenuata specimens in Chilean collections, he later listed S. attenuata for the country, without supporting evidence (Sielfeld, 1983). A cetacean manual (Cárdenas et al., 1986) stated that the spotted dolphin was ‘probably’ present in littoral waters of northern Chile. The primary range of the oceanic SAA was mapped to range south to the Peru-Chile border (18°21’S) (Jefferson et al., 2015; Perrin, 2018) while Rice (1998) indicated ‘about 35°S off Talca’ as southern range. However, all these remained without specific, authenticated records. Two authoritative national reviews of Chilean cetaceans (Aguayo, 1975; Aguayo et al., 1998a) did not include the species. Also, no spotted dolphins were encountered offshore, between Valparaiso (33°00’S,71°37’W) and Easter Island (27°09’S,109°W), during five sighting surveys in winter months (Aguayo et al., 1998b). Comprehensive field work in 1986-1988 and 2017-2021, including vessel-based surveys, stranding and bycatch monitoring in the Antofagasta region, were also negative for S. attenuata (Guerra-Correa et al., 1987; Findlay et al., 1998; García-Cegarra et al., 2021). The same results were found during dolphin surveys off central and southern Chile (e.g. Findlay et al., 1998; Gibbons, 2010; Schweber and Thomas, 2010; Gibbons and Waring, 2013; Siciliano and Higuera, 2015). We are confident that S. attenuata, and especially SAG is normally absent from Chile’s coastal waters. A far offshore summer presence of SAA off northern Chile, beyond the 200 nm EEZ, remains a possibility.
Life History
Pigmentation, External Morphology
Perrin (1969, 1975) described the strong developmental and individual variation in coloration found in SAG specimens from the ETP off Central America. He grouped individuals in five general categories of increasing spotting concurrent with larger body size: Newborn stage, Two-tone, Speckled, Mottled and Fused stage in the largest dolphins (see Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13 and Figure 14 in Perrin, 1969). The degree of body spotting in S. attenuata diminishes in animals the further west one goes in the ETP (Perrin, 1975; 2018). The coastal morphotype encountered in Peru and Ecuador (Figure 4) shows heavy spotting fully congruent with the Eastern Pacific SAG holotype (Figure 1). An adult female (Figure 3) and subadult male in Peru showed heavy white mottling on dorsum, flanks, tailstock and anogenital area. The lower half of the head, rostrum (minus white lips and tip), flippers and anterior ventral field was dark gray (Figure 6). Two calves that lacked any spotting (Two-tone stage), had grey upper flanks and dorsal field while the abdomen was whitish with a white anogenital blaze (Figure 3 and Figure 5). They closely resembled a calf Indo-Pacific bottlenose dolphin Tursiops aduncus, due to the more attenuate rostrum compared to T. truncatus.
A subadult male (AJR-036; Table 3) showed darker spots on a dark background of upper rostrum, flanks, abdomen, dorsal fin and flippers. A few lighter spots were present on the upper flanks. Dark bands extended from rostrum, around eyes and towards the blowhole. All three Peruvian animals had pale lips, and a smallish, falcate dorsal fin. Adult animals photographed at sea in Ecuador also show white lips and most have a pronounced narrow, falcate dorsal fin, although there exists important individual variation in dorsal fin shape (Figure 4). An adult female had a slight postanal keel (Figure 3). Males of adult size, from Ecuador, had a more pronounced postanal keel (Figure 7).
In central Peru, a male of 188 cm (standard body length, SL) had a body mass of 84 kg. The SL of specimens collected in Ecuador ranged between 92–228 cm, with females ranging 179-228 cm and males 92-200 cm (Table 3). In the NE Pacific, average lengths of adult male coastal and offshore subspecies were 223 cm (range 197–246) and 200 cm (range 166–240), respectively (Perrin, 2001). Average lengths of adult females were 207 cm (range, 179–227) and 182 cm (range, 163–215), coastal and offshore respectively (Perrin, 2001). The Peruvian adult female (211 cm; 99 kg) and the Ecuadorian females thus cluster with the ETP coastal stock. In the Gulf of California, males measured 146–227 cm (n=9) and females 182–221 cm (n=6) (Vidal et al., 1993). No adult males have been collected in Ecuador for comparison.
Cranial Characteristics
The earlier studies of geographic variation in cranial morphology of eastern Pacific S. attenuata integrated relatively large samples (e.g. Perrin, 1975; Perrin et al., 1994), however they did not include coastal specimens from South America. Here we present descriptive statistics of standard craniometrics for 9 SAG skulls (8 adults, 1 subadult) from Ecuador and Peru (Table 4), including mean, SD, minimum and maximum values, subsample sizes (because variable), both for absolute measurements (in mm) and the same expressed as percent of condylobasal length (% CBL). For comparison, equivalent data for SAA from the offshore ETP (Perrin, 1975) are juxtaposed, based on 65–70 adults (Table 4).
Jointly with graphic evidence (Figure 8), such information is immensely helpful in the (sub)species identification of dolphin skulls, especially those that are damaged or of unknown origin. To allow statistical analysis, however, our preliminary sample will need to be increased with additional skulls from South America. Also, access to raw craniometric data from Central and North America will be required. However, a simple comparison of means and ranges for adult skulls (Table 4) shows that, for all variables, the SAG skulls from South America are consistently larger than the offshore SAA collected by Perrin (1975). Three cranial variables, the CBL, mandibular ramus length and ramus height do not even have overlapping ranges, so important are the differences in size (Table 4). With a few measurements individual skulls can readily be assigned to their respective subspecies. Similarly, skulls from inshore specimens in the Northeast Pacific are also substantially larger than those from offshore areas of the ETP (Perrin, 1975; Douglas et al. 1984; Perrin and Hohn, 1994; Perrin, 2018). Perrin (1975) reported that tooth width is the character of the skull where the greatest divergence is found between (northern) inshore and offshore forms, with SAG having the widest teeth. Our sample (lacking teeth) does not allow evaluation. There are other significant differences in the feeding apparatus of the two forms, with offshore specimens having a more attenuate rostrum, smaller temporal fossa and smaller teeth (Perrin, 1975), which also explains the differences in size (non-overlapping) of the mandibles between the southern SAG and SAA (this paper). Our findings are consistent with the conclusions by Perrin (1975), that the significant variations suggest feeding differences between inshore and offshore morphs.
Reproduction and Growth
The reproductive biology of both male and female S. attenuata from the Eastern Tropical Pacific is supported by large sample sizes (Hohn et al., 1985; Myrick et al., 1986; Kellar et al., 2013). No information is published on reproductive parameters of the SAG subspecies in South America. In Peru, one small calf (KVW-344) without neonatal folds measured 106 cm for a weight of 19 kg, while a 84 cm near-term foetus from Ecuador, collected from a 218 cm female, is conserved in formalin at the Museo de Ballenas, Salinas. This suggests that length at birth for SAG may approach 90–95cm. Neonates of SAA in the ETP measure some 80-85 cm (Perrin and Hohn, 1994; Perrin, 2018) or 83.2–85.4 cm (Hohn and Hammond, 1985), smaller than those of the coastal stock.
A lactating female of 211 cm (KVW-343; Table 3) with abundant milk in its mammaries, presented only two corpora albicantia, one regressing, in the right ovary, and was thought to be still young. The left ovary was smooth, without corpora. Ovary dimensions were 40 x 13 x 7 mm (left) and 50 x 21 x 19 mm (right). Largest diameters of uterine horns measured 21 mm (left) and 36 mm (right). The female calf (KVW-344) with unerupted teeth had milk in its forestomach and oesophagus. It showed a healed umbilicus and lacked foetal rostral hairs and foetal folds. Age was estimated as a few months, hence the 20 April capture date was consistent with a summer parturition. Its smooth, fully immature, ovaries measured 17 x 9 x 6 mm (left) and 17 x 8 x 6 mm (right).
The macroscopic absence of sperm in both epididymides of a 188 cm male (AJR-036, Peru) confirmed its sexual immaturity. Its testes measured 97 x 17 x 12 mm (left) and 103 x 10 x 15 mm (right), with a mass of 34 g and 33 g, respectively.
Feeding
Nothing has been published on the diet of SAG in South America. The forestomach contents of an immature male (AJR-036) from Peru consisted of undigested fish flesh, small fish bones (possibly Peruvian anchovy Engraulis ringens) and small squid beaks. Nine otoliths were retrieved from 6 unidentified small fish remains. The main and pyloric stomachs, which were empty, weighed 390 g and 190 g respectively. Perrin (1975) indicated that morphological differences in the feeding apparatus, including tooth width (wider in SAG), length and width of temporal fossa (larger in SAG) suggest feeding differences between inshore and offshore populations of the ETP. Interestingly, in a case of parallel evolution, significantly wider teeth also differentiate inshore from offshore forms of common bottlenose dolphins in the eastern Pacific. The two forms show discrete diets (Walker, 1981; Van Waerebeek et al., 1990).
In Ecuador, SAG has been observed feeding on small pelagic fish, on occasions with large aggregations of marine bird flocks including blue-footed booby Sula nebouxii, red-footed booby Sula sula, and frigate birds Fregata sp., among others, as well as with other species of cetaceans, including Bryde's whales and common bottlenose dolphins (Félix et al., 2025). On two occasions, prey was identified as anchovies (Anchoa sp.) but it is not discarded that SAG targets other small pelagic fish such as mackerel (Scomber japonicus) or sardines (Opisthonema spp.), which are more abundant in Ecuadorian coastal waters than anchovies (Romero et al., 2020). Feeding behavior was observed on 11 occasions (18.6% of sightings) in Ecuadorian waters between 18 and 57 m in depth (mean= 46.75, n=4). Empty stomachs were found in a fresh adult female and an immature male, both stranded in 2006.
Helminth Parasites and Pathology
There is no published information available on parasites, epibionts and diseases of SAG. Here we present some initial findings.
Peruvian specimens
An adult female was infested with five Anisakis sp. nematodes in the forestomach and one in the pyloric stomach. We found no helminth parasites in the gastrointestinal tract of the subadult male AJR-036. Its kidneys were also unaffected. The female KVW-344 presented a moderate infestation of small-sized Nasitrema sp. trematodes in the cranial sinuses while its calf had none. The subadult male’s sinuses were infested with the same trematode. Its pterygoid bones showed some fenestration, but any potential link is unclear. Fully adult skull KVW-1999 had (anteriad) several maxillary alveoli filled-in with trabecular osseous tissue, presumably age-related.
None of the appendages of the three freshly dead individuals examined in Peru (Table 3) carried semi-stalked barnacles Xenobalanus globicipitis, although this phoront is very common in other Peruvian small odontocetes (e.g. Van Waerebeek et al., 1993; Reyes and Van Waerebeek, 1995). Also, no whale lice (Cyamidae) were found. All three animals had good body condition and no macroscopic pathologies were observed. However, due to the extensive spotting, certain dermatopathies e.g. poxviral tattoo skin disease, cutaneous herpes and papillomaviral warts (e.g. Van Bressem et al., 2007, 2009) could have escaped detection in the field.
Ecuadorian specimens
In Ecuador, only a handful of the hundred SAG recorded and photographed at sea carried X. globicipitis in their dorsal fins (less than 2%), although a high proportion of photographs were of insufficient quality for an exact evaluation. Anyhow, this datum indicates a low prevalence of the semi-stalked barnacle in SAG on the central coast of Ecuador, contrasting with the offshore common bottlenose dolphin which shows a prevalence of 42.3%, primarily on the dorsal fin (Félix and Castro, 2023), but only 4% in offshore T. truncatus of Peru (Van Waerebeek et al., 1990). The sampling period (because of possible seasonality), speed of travel and feeding ecology (cf. depth of diving) may all play an important role. Deep-diving Ziphiidae and Globicephala spp., for instance, do not carry Xenobalanus barnacles (KVW, unpublished data).
We recorded two cases of unusual barnacle infestation. The first case involved a calf with (reddish) stalked barnacles Conchoderma auritum in its mouth (Figure 9a), probably attached to exposed teeth due to a mandibular deviation or fracture. The second case was an immature dolphin that had an unusually high number (estimate n= 25) semi-stalked barnacles attached to its dorsal fin (Figure 9b). The animal appeared unhealthy and swam slowly, which may explain the unusual number and large size reached by the barnacles in this specimen. Additionally, an animal stranded in 2017 was found with five X. globicipitis barnacles along the trailing edge of the tail flukes.
Behavioral Ecology
Except, perhaps, for a limited study at Gorgona Island (Valencia, 2006) of an indeterminate form, but probably SAG, the behavioral ecology in South America remains unstudied. As off California, habitat of SAG in Pacific South America is thought to be largely limited to neritic waters and in northern part of South America (Colombia, Ecuador) may appear partially sympatric with the offshore ecotype of T. truncatus (Van Waerebeek et al., 2017; Félix and Castro, 2023). However, competitive exclusion may force upon a parapatric distribution.
In Colombia, Herrera et al. (2011b) reported spotted dolphins in deep waters around Malpelo Island and also over the shelf, suggesting that both subspecies could be recorded in their surveys from mainland to Malpelo Island. At Gorgona Island, individuals are frequently sighted in small herds with an average group size of eight dolphins. Groups with small calves are observed year-round, suggesting that reproduction occurs continuously in the region (Capella et al., 2014). On the central coast of Ecuador SAG groups have been recorded in depths ranging from 10 to 718 m (mean 75.8 m, SD = 124), with 60% of the sightings at depths between 30 and 60 m and only 5.6% in waters deeper than 100 m (Figure 10). Since most sightings in Ecuador were made onboard whale-watching boats, there is a sampling bias towards coastal areas. In the southern part of Ecuador SAG has been recorded in the outer estuary of the Gulf of Guayaquil but not in the inner estuary where inshore type T. truncatus is resident (e.g. Félix, 1997), suggesting that SAG avoids brackish waters.
Group size of SAG in central Ecuador varied widely from 3 to 500 (mean 121.22, SD=129.62, n= 52). (Figure 11). Most groups had 100 or fewer individuals. Large aggregations appeared to be integrated by several smaller units (50-100 individuals) that likely joined temporarily for feeding or social activities. High variability in group size has also been reported in other coastal areas in the Eastern Pacific (e.g. García and Dawson, 2003; Palacios & Salazar, 2002). Herrera et al. (2011b) reported significantly smaller group sizes around Malpelo Island (mean = 17.3, SD = 7.3, range = 8–30, n=6) compared to those recorded along the crossing route to the mainland (mean = 42.1, SD = 33.6, range = 3–100, n=14), suggesting spatial variation in social aggregation patterns consistent with niche separation.
SAG groups were also seen to be associated with other cetacean species in Ecuador on 14 occasions (Table 2): nine times with humpback whales (15.2 %), four with Bryde’s whales (6.78%), and on one occasion with both humpback whales and offshore bottlenose dolphins (1.69%). Interactions with humpback whales appeared to be social, while Bryde’s whales were typically linked to feeding activities. Interspecific associations between spotted dolphins and other species of cetaceans have been reported in Brazil (Rossi-Santos et al., 2009), Hawaii (Psarakos et al., 2003) and Galápagos (Palacios and Salazar, 2002). Further pelagic surveys could shed more light on SAG ecology, and in particular document interspecific relations.
Genetic Diversity and Population Structure
Two of the 29 samples from the coast of Ecuador did not amplify for mtDNA CR (Table S1). Haplotype diversity (h) was estimated at 0.8746 ± 0.0473, and nucleotide diversity (π) at 0.230550 ± 0.126309, showing that the population has high genetic diversity concordant with a large population. Tajima's D (-0.40581, p = 0.327) and Fu's Fs (-2.12380, p = 0.2033) provide no strong evidence of a recent evolutionary event in the population. Among the 27 sequences of the 405 bp mtDNA CR, 26 polymorphic sites were identified, revealing 14 haplotypes (Table 5). The nucleotide sequences were deposited in GenBank under accession numbers OQ867166–OQ867185 and PV131008–PV131014.
Of the 14 distinct haplotypes, six were unique (H1, H3, H4, H6, H11, and H13). Most of them (H3, H4, H6, H11, and H13) were identified in only one individual. The most common haplotypes were H1 and H7, found in 8 and 6 specimens, respectively. Eight haplotypes (H2, H5, H7, H8, H9, H10, H12, and H14) were previously reported in more than one location within the Eastern Tropical Pacific (ETP), Ecuador + Panama, California, and Costa Rica, indicating slight maternal genetic flow between these locations. However, as it was indicated earlier, it was not possible to distinguish between Ecuadorian and Panamanian sequences used for comparison. Haplotype H8 showed six mutational steps from H16, a haplotype from California, and H28 was identified as an offshore haplotype. H15 networked with distant localities, including Hawaii, ETP coastal regions, the Solomon Islands, and offshore stocks (Figure 12), suggesting it is one of the most ancestral haplotypes. Ecuadorian haplotypes are shared with at least one or two localities in the ETP and are primarily located at the periphery of the haplotype network, suggesting they are likely recent haplotypes. The haplotype network reveals the presence of unique haplotypes along the central coast of Ecuador and non-differentiated groups, indicating low genetic flow with other localities in the ETP and even lower connectivity with populations in the central and western Pacific (Figure 12).
According to the phylogenetic analysis, the Ecuadorian dolphins were distributed across seven different clades, with most belonging to the three largest clades, while only one haplotype was found in each of the remaining four (Figure 13). The Ecuadorian sequences primarily group with those previously identified by Escorza-Treviño et al. (2005) and labelled Ecuador-Panama (all labelled as ECU in the phylogenetic tree). This pattern suggests a complex evolutionary history, possibly indicating substructures within the population or an ongoing evolutionary process. The presence of haplotypes in multiple clades may reflect high genetic diversity, likely influenced by dolphin migration between nearby geographic areas, facilitating haplotype exchange and contributing to the observed distribution.
Fisheries Interactions and Other Threats
Bycatches of SAG are well-documented in coastal fisheries, mainly in artisanal gillnet fisheries in Ecuador and Peru (Van Waerebeek and Reyes, 1994; Castro and Van Waerebeek, 2019; Félix and Samaniego, 1994; this paper), in the bottom-trawl fishery for shrimp off Panama (Justines, 1997; García et al., 2008), and by harpooning for bait in artisanal fisheries in Chocó, Colombia (Avila et al., 2008; García et al., 2008). Of four Peruvian specimens, at least three had been killed in fisheries, one by harpooning. Of seven SAG specimens found stranded in Ecuador in 2009-2016, six (86%) bore evidence of net entanglement (Figure 14), underscoring the taxon’s high vulnerability to fisheries pressure (Castro and Van Waerebeek, 2019). In Ecuador, Castro and Rosero (2010) documented two additional cases of bycatch in Machalilla National Park, on the central coast, and Félix et al. (2011) reported that two of eleven stranding records found in Ecuador were probably a product of bycatch and three were suspected collisions with boats. Data on bycatches in Colombia are lacking. However, a study on the interaction between cetaceans and purse-seine fisheries targeting small pelagic fish found no evidence of interaction in 18 fishing sets conducted along the 1,300 km of the Colombian Pacific coast (Herrera and Flórez-González, 2018). No other threats have been identified, but dedicated field research is needed. For instance, potentially high contaminant loads in SAG, linked to neritic habitats, have not been studied.
Conclusions
Available evidence demonstrates that the spotted dolphins documented either free-ranging, taken in small-scale fisheries or stranded along the coasts of western South America, belong to S. a. graffmani, bar one dubious case from Puerto López, Ecuador.
We determine the known austral range of S. a. graffmani to the southern border of the Lima Department, at 13°02’S 76°29’W, which is 1,870 km and 1,360 km farther southeast than previously reported austral ranges, i.e. respectively the southern Colombia border (Perrin, 2018) and Playas, Ecuador (Chiluiza et al., 1998). We argued that the Peruvian southern range extension into the Lima Department was temporally related to a strong El Niño warm water intrusion into the coastal Humboldt Current Ecosystem which is concordant with recent records in northern Peru (Cortez-Casamayor et al., 2024). Abundant data from sighting surveys, strandings and bycatches support the notion that SAG is absent from southern Peru and Chile. The habitual southern range in non-El Niño years includes neritic waters off Ecuador’s Guayas and Manabí provinces (01°-02°S), perhaps including also Tumbes and Piura departments of northern Peru, in the months with higher SST. North of Ecuador, indications are of a continuous coastal distribution throughout the tropical waters of Colombia and Central America, into Baja California. Habitat of S. a. graffmani is thought to be largely limited to neritic waters and in Peru may be sympatric with the coastal ecotype of common bottlenose dolphin (Van Waerebeek et al., 1990, 2017), while off central Ecuador sympatry occurs with the offshore T. truncatus.
Phylogenetic analysis of mt-DNA control region (602 and 727 bp) of 29 Ecuadorian SAG samples confirms the presence of population structure, consistent with previous findings (Escorza-Treviño et al., 2005) and indicate limited gene flow with adjacent (northern) stocks. The SAG southern stock currently exhibits high genetic diversity and represents the most abundant neritic cetacean along the Ecuadorian coast and likely along the Colombian coast as well. Nevertheless, small-scale gillnet fisheries pose a persistent bycatch threat to the subspecies across its entire distribution range.
It is recommended that any spotted dolphin specimens becoming available in the study region be fully necropsied and sampled as to increase the currently unusually scarce life history record. All skeletal material, including postcranials, should also be collected.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
We express our sincere gratitude to the Pacific Whale Foundation for their continued support of cetacean research in the Eastern Tropical Pacific. We are especially thankful to Palo Santo Travel for providing logistical support and facilitating access to coastal areas for sample collection in Ecuador. We acknowledge the collaboration of local authorities of Machalilla National Park, fishermen, and community members who reported strandings and bycatch events along the coasts of Ecuador, as well as to Mr. Aurelio of Hotel Mandala, for access to skulls in their care. Julio C. Reyes, Santiago Zambrano and Andrew J. Read are thanked for their ‘historical’ contributions to field work in Peru in the 1980s, and later (JCR). Biopsy samples of free-ranging dolphins were collected following the guidelines specified under the Framework Contract for Access to Genetic Resources (permit number MAAE-DBI-CM-2021-0165) issued by the Ministerio del Ambiente, Agua y Transición Ecológica of Ecuador. Support by the Colombian Dirección General Marítima DIMAR, the Fundación Yubarta and WWF Colombia are also acknowledged. The Centro Peruano de Estudios Cetológicos (CEPEC) is an all-volunteer study group dedicated to conservation biology of cetaceans.
Conflicts of Interest
The authors declare that no conflict of interest exists.
References
- Aguayo, A. Progress report on small cetacean research in Chile. Journal Fisheries Research Board of Canada 1975, 32, 1123–1143. [Google Scholar] [CrossRef]
- Aguayo-Lobo, A.; Torres Navarro, D.; Acevedo Ramírez, J. Los mamíferos marinos de Chile: I. Cetacea. Ser. Cient. INACH 1998a, 48, 19–159. [Google Scholar]
- Aguayo, A.; Bernal, R.; Olavarria, C.; Vallejos, V.; Hucke, R. Observaciones de cetáceos realizadas entre Valparaiso e isla de Pascua, Chile, durante los inviernos de 1993, 1994 y 1995. Revista de Biología Marina y Oceanografía 1998b, 33(1), 101–123. [Google Scholar]
- Aguilar, A.; Forcada, J.; Gazo, M.; Badosa, E. Los Cetáceos del Parque Nacional de Coiba (Panamá). In Flora y Fauna del Parque Nacional de Coiba (Panamá); Castroviejo, S., Velayos, M., Eds.; Agencia Española de Cooperación Internacional (AECI): Madrid, 1997; pp. 75–102. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Avila, I.C.; Garcia, C.; Bastidas, J.C. A note on the use of dolphins as bait in the artisanal fisheries off Bahía Solano, Chocó, Colombia. J. Cetacean Res. Manag. 2008, 10, 179–182. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Barber, R. T.; Chavez, F. P. Biological consequences of El Niño. Science 1983, 222, 1203–1210. [Google Scholar] [CrossRef]
- Bayas-Rea, R.d.L.Á.; Félix, F.; Montufar, R. Genetic divergence and fine scale population structure of the common bottlenose dolphin (Tursiops truncatus, Montagu) found in the Gulf of Guayaquil, Ecuador. PeerJ 2018, 6, e4589. [Google Scholar] [CrossRef] [PubMed]
- Capella, A.J.; Flórez González, L.; Falk, P. E.; Herrera, J.C.; Tobón, I.C.; Hernández, E.; Recalde, A. Plan básico para el manejo de los mamíferos marinos en el PNN Gorgona, Pacífico colombiano; Parques Nacionales Naturales y WWF-Colombia: Cali, Colombia, 2014; p. 72 p. [Google Scholar]
- Cárdenas, J. C.; Stutzin, M. E.; Oporto, J. A.; Cabello, C.; Torres, D. Manual de identificación de los cetáceos chilenos. Proyecto WH-445 Cetáceos-Chile, WWF and CODEFF, Santiago; 1986. [Google Scholar]
- Carwardine, M. Handbook of Whales, dolphins and Porpoises; Bloomsbury Wildlife, London, 2020; p. 528pp. [Google Scholar]
- Castro, C.; Rosero, P. Small cetacean interaction with fishing gear in Machalilla National Park, Ecuador. Document SC/62/BC3, 2010; presented to the Scientific Committee of the International Whaling Commission. [Google Scholar]
- Castro, C.; Van Waerebeek, K. Strandings and mortality of cetaceans due to interactions with fishing nets in Ecuador, 2001-2017. Document SC/68A/HIM/17 presented to the IWC Scientific Committee Meeting, Nairobi, May 2019; 2019; p. 11 pp. [Google Scholar]
- Chiluiza, D.; Aguirre, W.; Félix, F.; Haase, B. Varamientos de mamíferos marinos en la costa continental ecuatoriana período 1987-1995. Acta Oceanográfica del Pacífico. INOCAR 1998, 9(1), 209–217. [Google Scholar]
- Chinacalle-Martínez, N.; García-Rada, E.; López-Macías, J.; Pinoargote, S.; Loor, G.; Zevallos-Rosado, J.; Cruz, P.; Pablo, D.; Andrade, B.; Robalino-Mejía, C.; et al. Oceanic primary production trend patterns along coast of Ecuador. Neotropical Biodivers. 2021, 7, 379–391. [Google Scholar] [CrossRef]
- Cortez-Casamayor, S.; Rodriguez, K.A.; Gubbins, S.; Cortés, D.; Gutierrez, C.; Silva, S.; Alcorta, B.; Pacheco, A.S. Occurrence of pantropical spotted dolphin (Stenella attenuata) in northern Peru: an El Niño effect? Mammalia 2024, 89, 182–188. [Google Scholar] [CrossRef]
- Courbis, S.; Baird, R.W.; Cipriano, F.; Duffield, D. Multiple Populations of Pantropical Spotted Dolphins in Hawaiian Waters. J. Hered. 2014, 105, 627–641. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Dalebout, M.L.; VAN Helden, A.; VAN Waerebeek, K.; Baker, C.S. Molecular genetic identification of southern hemisphere beaked whales (Cetacea: Ziphiidae). Mol. Ecol. 1998, 7, 687–694. [Google Scholar] [CrossRef]
- Day, D. List of cetaceans seen in Galápagos. Noticias de Galápagos 1994, 53, 5–6. [Google Scholar]
- De Weerdt, J.; Ramos, E.A.; Pouplard, E.; Kochzius, M.; Clapham, P. Cetacean strandings along the Pacific and Caribbean coasts of Nicaragua from 2014 to 2021. Mar. Biodivers. Rec. 2021, 14, 1–9. [Google Scholar] [CrossRef]
- Denkinger, J.; Oña, J.; Alarcón, D.; Merlen, G.; Salazar, S.; Palacios. From whaling to whalewatching: cetacean presence and species diversity in the Galapagos Marine Reserve. Science and Conservation in the Galapagos Islands: Frameworks & Perspectives, Social and Ecological Interactions in the Galapagos Islands 2013. [Google Scholar] [CrossRef]
- Dizon, A. E.; Perrin, W. F.; Akin, P. A. Stocks of dolphins (Stenella spp. and Delphinus delphis) in the eastern tropical Pacific: a phylogeographic classification. NOAA Technical Report NMFS 1994, 119, 1–20. [Google Scholar]
- Douglas, M.E.; Schnell, G.D.; Hough, D.J. Differentiation between Inshore and Offshore Spotted Dolphins in the Eastern Tropical Pacific Ocean. J. Mammal. 1984, 65, 375–387. [Google Scholar] [CrossRef]
- Escorza-Treviño, S.; Archer, F.I.; Rosales, M.; Lang, A.; Dizon, A.E. Genetic differentiation and intraspecific structure of Eastern Tropical Pacific spotted dolphins, Stenella attenuata, revealed by DNA analyses. Conserv. Genet. 2005, 6, 587–600. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Félix, F. Organization and social structure of the bottlenose dolphin Tursiops truncatus in the Gulf of Guayaquil, Ecuador. Aquatic Mammals 1997, 23(1), 1–16. [Google Scholar]
- Félix, F.; Samaniego, J. Incidental catches of small cetaceans in the artisanal fisheries of Ecuador. Report International Whaling Commission (Special Issue 15) 1994, 475–480. [Google Scholar]
- Félix, F.; Castro, C. Occurrence, abundance and some ecological aspects of the offshore bottlenose dolphin off Ecuador’s central coast. Lat. Am. J. Aquat. Mamm. 2023. [Google Scholar] [CrossRef]
- Félix, F.; Haase, B.; Denkinger, J.; Falconí, J. Varamientos de mamíferos marinos registrados en la costa continental de Ecuador entre 1996 y 2009. Acta Oceano Del. Pac 2011, 16, 61–73. [Google Scholar]
- Félix, F.; Castro, C.; Barragán-Tabares, L.; Orellana-Vásquez, H.; Platt, M. THE BRYDE’S WHALE ON THE CENTRAL COAST OF ECUADOR, DISTRIBUTION, BEHAVIOR AND ECOLOGICAL INSIGHTS. J. Cetacean Res. Manag. 2025, 26, 87–100. [Google Scholar] [CrossRef]
- Fiedler, P. Environmental change in the eastern tropical Pacific Ocean: review of ENSO and decadal variability. Mar. Ecol. Prog. Ser. 2002, 244, 265–283. [Google Scholar] [CrossRef]
- Findlay, K.; Pitman, R.; Tsurui, T.; Sakai, K.; Ensor, P.; Iwakami, H.; Ljungblad, D.; Shimada, H.; Thiele, D.; Van Waerebeek, K.; Hucke-Gaete, R.; Sanino-Vattier, G.P. 1997/1998 IWC-Southern Ocean whale and ecosystem research (IWC-SOWER) blue whale cruise, Chile. Document SC/50/Rep2 presented to the IWC Scientific Committee, Muscat, Oman, May 1998; 1998. [Google Scholar]
- Flórez-González, L.; Capella, J.; Falk-Fernández, P. Guia de campo de los mamíferos acuaticos de Colombia; Editorial Sepia Ltda: Santiago de Cali, Colombia, 2004; p. 124 pp. [Google Scholar]
- Fu, Y.-X. Statistical Tests of Neutrality of Mutations Against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Dawson, S.M. Distribution of pantropical spotted dolphins in Pacific coastal waters of Panama. Lat. Am. J. Aquat. Mamm. 2003, 2, 29–38. [Google Scholar] [CrossRef]
- García, C.; Soler, G.; Avila, I.C.; Bessudo, S. Observaciones preliminares de la presencia y distribución estacional de Mamíferos Marinos del Pacífico colombiano. p. 123. Resúmenes Primera Reunión Internacional sobre el Estudio de los Mamíferos Acuáticos SOMEMMA – SOLAMAC, Mérida, México, 2006. [Google Scholar]
- García, C.; Avila, I.C.; Palacios, D.; Gerrodette, T.; Suárez, M.; Soler, G.; Rasmussen, K.; May-Collado, L.; Parson, C.; Trujillo, F.; Bessudo, S. Presence, distribution and threats of Stenella attenuata and Tursiops truncatus in the Pacific waters of Panamá and Colombia. In Resumen 310 en Memorias XIII Reunión de Trabajo de Especialistas en Mamíferos Acuáticos de América del Sur y 7° Congreso SOLAMAC; Montevideo, Uruguay, 2008. [Google Scholar]
- Garcia-Cegarra, A.M.; Toro, F.; Gonzalez-Borasca, V. Citizen science as a tool to assess cetacean diversity in the Atacama Desert coast. Ocean Coast. Manag. 2021, 213. [Google Scholar] [CrossRef]
- Gerrodette, T.; Forcada, J. Non-recovery of two spotted and spinner dolphin populations in the eastern tropical Pacific Ocean. Mar. Ecol. Prog. Ser. 2005, 291, 1–21. [Google Scholar] [CrossRef]
- Zamorano-Abramson, J.; Gibbons, J.; Capella, J. DIVERSITY AND SUMMER DISTRIBUTION OF CETACEANS IN INLET WATERS OF NORTHERN AISÉN, CHILE. An. del Inst. de la Patagon. 2010, 38, 151–157. [Google Scholar] [CrossRef]
- Gibbons, J. A.; Waring, G. T. Observations of coastal and offshore dolphins in Chilean waters. 2013. [Google Scholar]
- Marine Ecology Progress Series 493, 283–295.
- Gray, J.E. The Zoology of the Voyage of H.M.S. Erebus and Terror; Richardson, J., Gray, J.E., Eds.; London, 1846. [Google Scholar]
- Gerrodette, T.; Palacios, D. Estimates of cetacean abundance in EEZ. Waters of the Eastern Tropical Pacific. Administrative Report LJ 96-10; NMFS - SWFSC: La Jolla, CA, 1996. [Google Scholar]
- Guerra-Correa, C.; Van Waerebeek, K.; Portflitt, G.; Luna, G. Presencia de cetáceos frente a la segúnda región de Chile. Estudios Oceanológicos 1987, 6, 87–96. [Google Scholar]
- Hamilton, T.A.; Redfern, J.; Barlow, J.; Balance, L.T.; Gerrodette, T.; Holt, R.S.; Forney, K.A.; Taylor, B.L. NOAA-TM-NMFS-SWFSC-440; Atlas of cetacean sightings for Southwest Fisheries Science Center cetacean and ecosystem surveys: 1986-2005. 2009.
- Herrera, J.C.; Ávila, I.; Falk, P.; Soler, G.; García, C.; Tobón, I.; Capella, J. Los Mamíferos Marinos en el Santuario de Fauna y Flora Malpelo y Aguas hacia el Continente, Pacífico Colombiano. DIMAR-CCCP y UAESPNN. Santuario de Fauna y Flora Malpelo: descubrimiento en marcha, (Ed.) DIMAR 2007, 113–130. [Google Scholar]
- Herrera, J. C.; Flórez-González, L.; Giraldo, A. Mamíferos Acuáticos. In Colombia Pacífico: Una visión sobre su biodiversidad marina; Cantera, J., Londoño, E., Eds.; Editorial Universidad del Valle: Cali, 2011a; pp. 393–402. [Google Scholar]
- Carmona, J.C.H.; Alzueta, J.J.C.; Soler, G.A.; Bessudo, S.; García, C.; González, L.F. OCURRENCIA Y TASAS DE ENCUENTRO DE MAMÍFEROS MARINOS EN LAS AGUAS DE LA ISLA MALPELO Y HACIA EL CONTINENTE. Bull. Mar. Coast. Res. 2016, 40. [Google Scholar] [CrossRef]
- Herrera, J.C.; Flórez-González, L. Registro de cetáceos durante los cruceros de evaluación pesquera de pequeños pelágicos sobre la plataforma continental del Pacífico de Colombia. In Cruceros de evaluación de pequeños pelágicos en el Pacífico colombiano; Zapata, L.A., Herrera, J.C., Eds.; Ministerio de Agricultura y Desarrollo Rural, Parques Nacionales Naturales, AUNAP, Universidad del Valle, HARMAR S.A., y WWF-Colombia: Cali, 2018; pp. 141–149. [Google Scholar]
- Hohn, A.A.; Chivers, S.J.; Barlow, J. REPRODUCTIVE MATURITY AND SEASONALITY OF MALE SPOTTED DOLPHINS, STENELLA ATTENUATA, IN THE EASTERN TROPICAL PACIFIC. Mar. Mammal Sci. 1985, 1, 273–293. [Google Scholar] [CrossRef]
- Hohn, A. A.; Hammond, P. S. Early postnatal growth of the spotted dolphin, Stenella attenuata, in the offshore eastern tropical Pacific. Fishery Bulletin 1985, 83(4), 553–566. [Google Scholar]
- Ibarra-Portillo, R.; Barraza, J.E.; Pineda, L. Registros de varamientos de cetáceos en El Salvador entre 1995-2019. Real. Y Reflex. 2020, 20, 210–231. [Google Scholar]
- Jefferson, T.A.; Webber, M.A.; Pitman, R.L. Marine Mammals of the World: A Comprehensive Guide to Their Identification, 2nd Edition ed; Elsevier: San Diego, CA, 2015. [Google Scholar]
- Justines, G. Métodos de monitoreo para estimar la mortalidad incidental de mamíferos marinos en el Pacífico de Panamá. Comisión Permanente del Pacífico Sur, Quito; (unpublished); 1997; p. 100pp. [Google Scholar]
- Kellar, N.M.; Trego, M.L.; Chivers, S.J.; Archer, F.I. Pregnancy patterns of pantropical spotted dolphins (Stenella attenuata) in the eastern tropical Pacific determined from hormonal analysis of blubber biopsies and correlations with the purse-seine tuna fishery. Mar. Biol. 2013, 160, 3113–3124. [Google Scholar] [CrossRef]
- Leslie, M. S.; Archer, F. I.; Morin, P. A. Mitogenome and nuclear DNA differentiation in spinner (Stenella longirostris) and pantropical spotted dolphins (S. attenuata) from the Eastern Tropical Pacific Ocean. bioRxiv 2016, 091215. [Google Scholar]
- Londoño, C. Ecología del delfín Tursiops truncatus y Stenella attenuata en el sector de Bahía Málaga, Pacífico colombiano; Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia: Medellín, Colombia, 2005. [Google Scholar]
- Lönnberg, E. Prodelphinus graffmani n.sp. a new dolphin from the Pacific coast of Mexico. Arkiv för Zoologi 1934, 26A(19), 1–11. [Google Scholar]
- Mangel, J.C.; Alfaro-Shigueto, J.; Van Waerebeek, K.; Cáceres, C.; Bearhop, S.; Witt, M.J.; Godley, B.J. Small cetacean captures in Peruvian artisanal fisheries: High despite protective legislation. Biol. Conserv. 2010, 143, 136–143. [Google Scholar] [CrossRef]
- Martínez, L.; Silva, S.; Alcorta, B.; Pacheco, A.S. First records of the pantropical spotted dolphin Stenella attenuata, within groups of common dolphins Delphinus delphis in northern Peru. Rev. De Biol. Mar. Y Oceanogr. 2021, 56, 78–82. [Google Scholar] [CrossRef]
- Myrick, A.C., Jr.; Hohn, A. A.; Barlow, J.; Sloan, P. A. Reproductive biology of female spotted dolphins, Stenella attenuata, from the Eastern Tropical Pacific. Fishery Bulletin 1986, 84(2), 247–259. [Google Scholar]
- NOAA. Climate Prediction Service. 2025. [Google Scholar]
- 10 December 2025. Available online: https://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php.
- Oremus, M.; Leqata, J.; Baker, C.S. Resumption of traditional drive hunting of dolphins in the Solomon Islands in 2013. R. Soc. Open Sci. 2015, 2, 140524. [Google Scholar] [CrossRef]
- Ortiz-Wolford, J.; Corona-Figueroa, M.F.; Dávila, V.; Cabrera, A.A. Cetacean stranding records along the Pacific coastline of Guatemala, 2007–2021: Implications for management, conservation and research. Mar. Policy 2021, 134. [Google Scholar] [CrossRef]
- Pacheco, A.S.; Silva, S.; Alcorta, B.; Gubbins, S.; Guidino, C.; Sanchez-Salazar, F.; Petit, A.; Llapapasca, M.A.; Balducci, N.; Larrañaga, E.; et al. Cetacean Diversity Revealed from Whale-Watching Observations in Northern Peru. Aquat. Mamm. 2019, 45, 116–122. [Google Scholar] [CrossRef]
- Palacios, D. M.; Salazar, S. Cetáceos. En: Reserva Marina de Galápagos. In Línea Base de la Biodiversidad; Danulat, E., Edgar, G.J., Eds.; Fundación Charles Darwin/Servicio Parque Nacional Galápagos: Santa Cruz, Galápagos, Ecuador, 2002; pp. pp 291–304. [Google Scholar]
- Palacios, D.M.; Herrera, J.C.; Gerrodette, T.; Garcia, C.; Soler, G.A.; Avila, I.C.; Bessudo, S.; Hernandez, E.; Trujillo, F.; Forez-Gonzalez, L.; et al. Cetacean distribution and relative abundance in Colombia’s Pacific EEZ from survey cruises and platforms of opportunity. J. Cetacean Res. Manag. 2012, 12, 45–60. [Google Scholar] [CrossRef]
- Perrin, W.F. Color pattern of the Eastern Pacific spotted porpoise Stenella graffmani Lönnberg (Cetacea, Delphinidae). Sci. Contrib. New York Zoöl. Soc. 1970, 54, 135–149. [Google Scholar] [CrossRef]
- Perrin, W.F. Variation of spotted and spinner dolphin (genus Stenella) in the eastern tropical Pacific and Hawaii. Bulletin Scripps Institute of Oceanography 1975, 21, 1–206. [Google Scholar]
- Perrin, W.F. Stenella attenuata. Mamm. Species 2001, 1–8. [Google Scholar] [CrossRef]
- Perrin, W.F.; Würsig, B.; Thewissen, J. Encyclopedia of Marine Mammals; Elsevier: Amsterdam, NX, Netherlands; ISBN, 2009. [Google Scholar]
- Perrin, W.F.; Mitchell, E.D.; Mead, J.G.; Caldwell, D.K.; Caldwell, M.C.; van Bree, P.J.H.; Dawbin, W.H. REVISION OF THE SPOTTED DOLPHINS, STENELLA SPP. Mar. Mammal Sci. 1987, 3, 99–170. [Google Scholar] [CrossRef]
- Perrin, W.F.; Hohn, A.A. Pantropical spotted dolphin. In Handbook of Marine Mammals; Ridgway, S.H., Harrison, R., Eds.; Academic Press: London, 1994; Volume 5, p. pp. 71-98 71-98. [Google Scholar]
- Perrin, W.F.; Schnell, G. D.; Hough, D. J.; Gilpatrick, J., Jr.; Kashiwada, J. V. Reexamination of geographic variation in canial morphology of the pantropical spotted dolphin, Stenella attenuata, in the eastern Pacific. Fishery Bulletin 1994, 92, 324–346. [Google Scholar]
- Pizarro-Neyra, J. Varamiento de cetáceos en Tacna, Perú (2002-2010). Revista peru Biología 2010, 17(2), 253–255. [Google Scholar] [CrossRef]
- Pizarro-Neyra, J.; Mamani-Fernández, J.; Vizcarra, J.K. Varamiento de Cetáceos en la costa sur del Perú (2016-2021). Cienc. Desarro. 2021, 20, 29–40. [Google Scholar] [CrossRef]
- Psarakos, S.; Herzing, D.L.; Marten, K. Mixed-species associations between Pantropical spotted dolphins (Stenella attenuata) and Hawaiian spinner dolphins (Stenella longirostris) off Oahu, Hawaii. Aquat. Mamm. 2003, 29, 390–395. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.4, a graphical viewer of phylogenetic trees; Institute of Evolutionary Biology: University of Edinburgh, 2018. [Google Scholar]
- Ramírez, P.; Urquizo, W. Los cetáceos mayores y el fenómeno El Niño 1982-1983. In Vol. Extraordinario. El Niño. Su impacto en la Fauna Marina. Boletín IMARPE; Arntz, W., Landa, A., Tarazona, J., Eds.; Callao, Peru, 1985; pp. 201–206. [Google Scholar]
- Read, A.J.; Van Waerebeek, K.; Reyes, J.C.; McKinnon, J.S.; Lehman, L.C. The exploitation of small cetaceans in Coastal Peru. Biol. Conserv. 1988, 46, 53–70. [Google Scholar] [CrossRef]
- Reyes, J. C. Ballenas, delfines y otros cetáceos del Perú; Squema-Ediciones: Lima, 2009; p. 150pp. [Google Scholar]
- Reyes, J.C.; Van Waerebeek, K. Aspects of the Biology of Burmeister's porpoise from Peru. In Report of the International Whaling Commission (Special Issue 16); 1995; pp. 349–364. [Google Scholar]
- Rice, D. W. Marine Mammals of the World. In Systematics and Distribution; The Society for Marine Mammalogy, 1998; Volume Special Publication no. 4. [Google Scholar]
- Rodríguez-Fonseca, J.; Cubero-Pardo, P. Cetacean strandings in Costa Rica (1966-1999). 2001, 49, 667–72. [Google Scholar]
- Rodríguez-Rubio, E.; Schneider, W.; Abarca del Rio, R. On the seasonal circulation within the Panama Bight derived from satellite observations of wind, altimetry and sea surface temperature. Geophysical Research Letters 2003, 30(7), 1410. [Google Scholar] [CrossRef]
- Romero, A.; Ponce, G.; Hurtado, M.; Prado, J.; Armijos, G.; Vilela, K.; González, N.; Aguilar, F.; Muñoz, A.; Jurado, V. Estimación hidroacústica de los principales peces pelágicos pequeños en el Ecuador y su distribución geoespacial, durante diciembre de 2019 (INP 2019-12-02V). Instituto Nacional de Pesca, Ecuador. 2020. Available online: https://www.institutopesca.gob.ec/wp-content/uploads/2018/01/Estimaci%C3%B3n-Hidroac%C3%BAstica-de-los-principales-peces-pel%C3%A1gicos-peque%C3%B1os-en-el-Ecuador-y-su-distribuci%C3%B3n-geoespacial-durante-diciembre-de-2019.pdf.
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rosel, P.E.; Dizon, A.E.; Heyning, J.E. Genetic analysis of sympatric morphotypes of common dolphins (genus Delphinus). Mar. Biol. 1994, 119, 159–167. [Google Scholar] [CrossRef]
- Rosel, P.E.; France, S.C.; Wang, J.Y.; Kocher, T.D. Genetic structure of harbour porpoisePhocoena phocoenapopulations in the northwest Atlantic based on mitochondrial and nuclear markers. Mol. Ecol. 1999, 8, S41–54. [Google Scholar] [CrossRef] [PubMed]
- Rossi-Santos, M.R.; Santos-Neto, E.; Baracho, C.G. Interspecific cetacean interactions during the breeding season of humpback whale (Megaptera novaeangliae) on the north coast of Bahia State, Brazil. J. Mar. Biol. Assoc. United Kingd. 2009, 89, 961–966. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Corrales, L.A.S. Cetáceos del sur del Perú. Cienc. Desarro. 2021, 20, 3–11. [Google Scholar] [CrossRef]
- Siciliano, S.; Higuera, D. Distribution and ecology of dolphins off the southern Chilean coast: implications for conservation. Marine Mammal Science 2015, 31(4), 1378–1391. [Google Scholar]
- Schweber, C.; Thomas, P. O. Cetacean distribution in Chilean coastal waters: a baseline for conservation efforts. Ocean & Coastal Management 2010, 53(12), 750–759. [Google Scholar]
- Sielfeld, W. Mamíferos marinos en colecciones y museos de Chile. Anales del Instituto de la Patagonía 1980, 11, 273–280. [Google Scholar]
- Sielfeld, W. Mamíferos marinos de Chile. Ediciones de la Universidad de Chile, Santiago; 1983. [Google Scholar]
- Silva-Buse, S. Gran diversidad. In (ed.) Fondo Editorial, Universidad Científica del Sur). Ballenas en el Norte del Perú; Industria Gráfica Cimagraf S.A.C.: Lima, Peru, 2016; pp. 138–171. [Google Scholar]
- Suárez, M. Aspectos ecológicos y del compotrtamiento de los delfines Tursiops truncatus (Montagu, 1821) y Stenella attenuata (Gray, 1846) en el Parque Nacional Utría, Chocó, Colombia. Tesis de grado en biología Universidad Nacional de Colombia, (unpublished). Santa Fe de Bogotá, 1994; p. 107 pp. [Google Scholar]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tzika, A.C.; D’aMico, E.; Alfaro-Shigueto, J.; Mangel, J.C.; Van Waerebeek, K.; Milinkovitch, M.C. Molecular identification of small cetacean samples from Peruvian fish markets. Conserv. Genet. 2010, 11, 2207–2218. [Google Scholar] [CrossRef]
- Valencia, B. Presencia y comportamiento del delfín moteado pantropical (Stenella attenuata) en el Parque Nacional Natural Gorgona, Pacífico Colombiano. Tesis de grado, (unpublished). Universidad del Valle, Cali, Colombia, 2006. [Google Scholar]
- Van Bressem, M.-F.; Reyes, J.C.; Félix, F.; Echegaray, M.; Siciliano, S.; Di Beneditto, A.P.; Flach, L.; Viddi, F.; Avila, I.C.; Herrera, J.C.; et al. A preliminary overview of skin and skeletal diseases and traumata in small cetaceans from South American waters. Lat. Am. J. Aquat. Mamm. 2007, 6, 7–42. [Google Scholar] [CrossRef]
- Van Bressem, M.; Van Waerebeek, K.; Aznar, F.; Raga, J.; Jepson, P.; Duignan, P.; Deaville, R.; Flach, L.; Viddi, F.; Baker, J.; et al. Epidemiological pattern of tattoo skin disease: a potential general health indicator for cetaceans. Dis. Aquat. Org. 2009, 85, 225–237. [Google Scholar] [CrossRef]
- Van Waerebeek, K.; Reyes, J.C.; Luscombe, B.A. Revisión de la distribución de pequeños cetáceos frente al Perú. In Recursos y Dinámica del Ecosistema de Afloramiento Peruano. Boletín del Instituto del Mar del Perú, Callao; Salzwedel, H., Landa, A., Eds.; 1988; Volumen extraordinario, pp. 345–351. [Google Scholar]
- Van Waerebeek, K.; Reyes, J.C.; Read, A.J.; McKinnon. Preliminary observations of bottlenose dolphins from the Pacific coast of South America. The Bottlenose Dolphin. [CrossRef]
- Van Waerebeek, K.; Reyes, J.C.; Alfaro, J.O. Helminth parasites and phoronts of dusky dolphins Lagenorhynchus obscurus (Gray 1828) from Peru. Aquatic Mammals 1993, 19(30), 159–169. [Google Scholar]
- Van Waerebeek, K.; Reyes, J.C. Post-ban small cetacean takes off Peru: a review. Report of the International Whaling Commission (Special Issue 15) 1994, 503–520. [Google Scholar]
- Van Waerebeek, K.; Van Bressem, M. F.; Félix, F.; Alfaro, J.; García-Godos, A.; Chávez, L.; Ontón, K.; Montes, D.; Bello, R. Mortality of dolphins and porpoises in coastal fisheries off Peru and southern Ecuador in 1994. Biological Conservation 1997, 81, 43–49. [Google Scholar] [CrossRef]
- Van Waerebeek, K.; Félix, F.; Haase, B.; Palacios, D. M.; Mora-Pinto, D. M.; Muñoz-Hincapié, M. Inshore records of the striped dolphin, Stenella coeruleoalba, from the Pacific coast of South America. Report of the International Whaling Commission 1998, 48, 525–532. Available online: https://www.researchgate.net/publication/230630858.
- Van Waerebeek, K.; Reyes, J.C.; Sanino, G.; Félix, F.; Van Bressem, M-F.; Santillán, L.; Montes, D.; García-Godos, I.; Echegaray, M.; Venegas-Abad, A. Common bottlenose dolphins Tursiops truncatus of Pacific South America, a synoptic review of population identification data. Document SC/67A/SM/10, IWC Scientific Committee Meeting, Bled, Slovenia, May 2017; 2017. [Google Scholar]
- Van Waerebeek, K.; Apaza, M.; Reyes, J.C.; Alfaro-Shigueto, J.; Santillán, L.; Barreda, E.; Altamirano-Sierra, A.; Astohuaman-Uribe, J.; Ortiz-Alvarez, C.; Mangel, J. Beach cast small cetaceans bear evidence of continued catches and utilisation in coastal Peru. Document SC/67B/HIM/01 presented to IWC Scientific Committee Meeting, Bled, Slovenia, April 2018; 2018. [Google Scholar]
- Vidal, O.; Findley, L. T.; Leatherwood, S. Annotated checklist of the marine mammals of the Gulf of California. Proceedings of the San Diego Society of Natural History 1993, 28, 1–16. [Google Scholar]
- Walker, W.A. Geographical variation in morphology and biology of bottlenose dolphins (Tursiops) in the eastern north Pacific. In Southwest Fisheries Center (U.S.) Administrative report LJ-81-03C; 1981; p. 54 pp. [Google Scholar]
- Available online: https://repository.library.noaa.gov/view/noaa/22873.
- Wyrtki, K. Oceanography of the eastern Equatorial Pacific Ocean. Oceanography and Marine Biology Annual Review 1966, 4, 33–68. [Google Scholar]
Figure 1.
Holotype specimen of the stocky and heavily spotted Stenella graffmani Lönnberg (1934) from Acapulco, Mexico. The left-side mandibular and labial damage, due to firearm shots, exposes underlying connective tissues (pale colouration). Reproduced from Lönnberg (1934) 1.
Figure 1.
Holotype specimen of the stocky and heavily spotted Stenella graffmani Lönnberg (1934) from Acapulco, Mexico. The left-side mandibular and labial damage, due to firearm shots, exposes underlying connective tissues (pale colouration). Reproduced from Lönnberg (1934) 1.

Figure 2.
The Southeast Pacific region and records of SAG in the period 1986-2024. Record from Colombia were obtained from the scientific literature.
Figure 2.
The Southeast Pacific region and records of SAG in the period 1986-2024. Record from Colombia were obtained from the scientific literature.
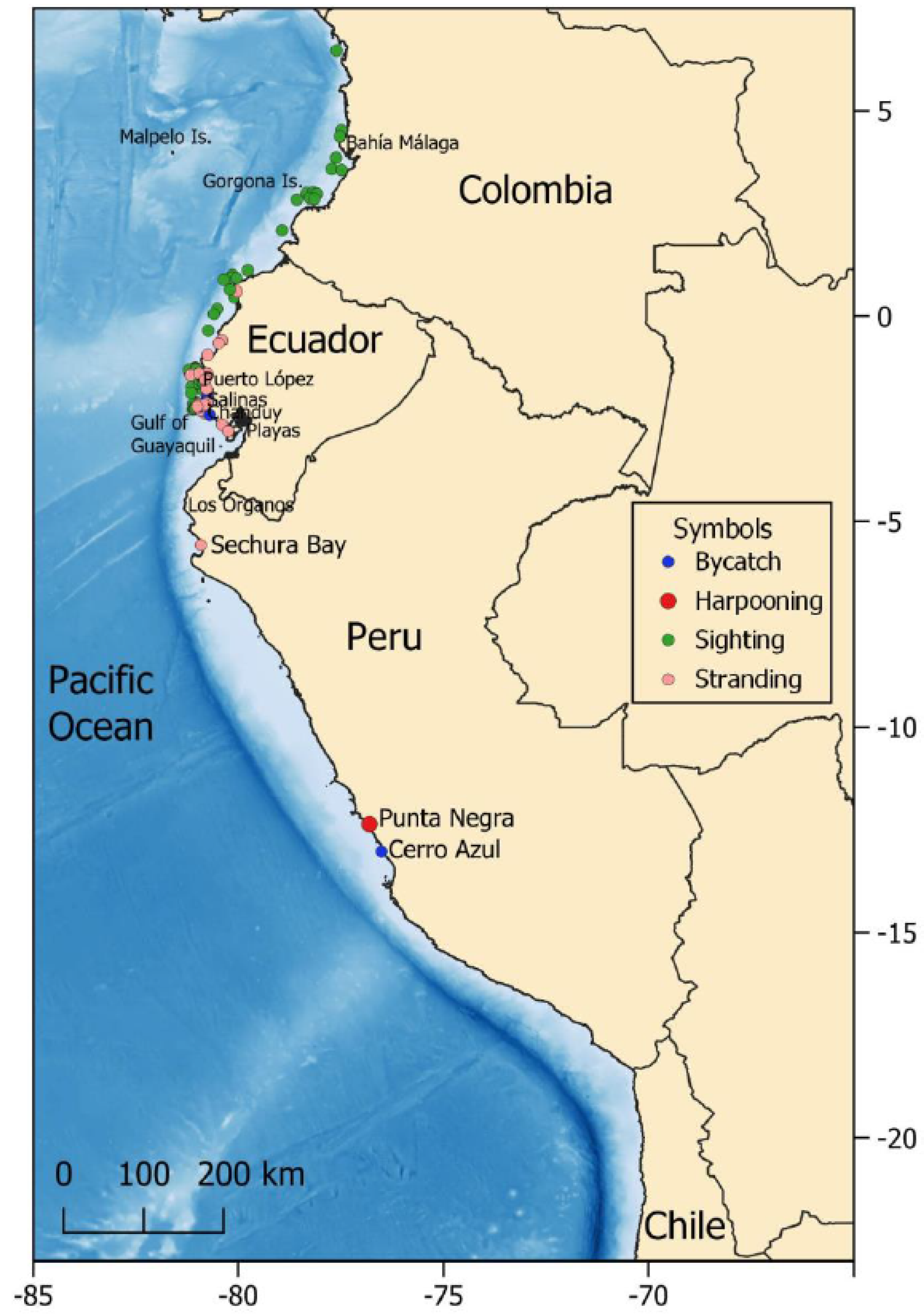
Figure 3.
Southernmost record of Stenella attenuata graffmani in the SE Pacific: adult female (KVW-343) with calf (KVW-344) landed at Cerro Azul artisanal port in Lima Department, Peru.
Figure 3.
Southernmost record of Stenella attenuata graffmani in the SE Pacific: adult female (KVW-343) with calf (KVW-344) landed at Cerro Azul artisanal port in Lima Department, Peru.

Figure 4.
Photographs of three adult S. a. graffmani taken on the central coast of Ecuador in which the heavily spotted pattern is evident. Note the important variation in dorsal fin shape, with the strongly falcate morph (bottom) commonly seen.
Figure 4.
Photographs of three adult S. a. graffmani taken on the central coast of Ecuador in which the heavily spotted pattern is evident. Note the important variation in dorsal fin shape, with the strongly falcate morph (bottom) commonly seen.

Figure 5.
A calf of SAG with no spots on the body leaping by its mother.

Figure 6.
Adult coastal spotted dolphin photographed off Ecuador showing a dark rostrum with white lips.
Figure 6.
Adult coastal spotted dolphin photographed off Ecuador showing a dark rostrum with white lips.

Figure 7.
An adult male SAG showing a pronounced postanal keel.

Figure 8.
SAG skull curated at CEPEC, Peru. A) dorsal view and mandibles, B) ventral view, C) lateral vie and left mandible.
Figure 8.
SAG skull curated at CEPEC, Peru. A) dorsal view and mandibles, B) ventral view, C) lateral vie and left mandible.
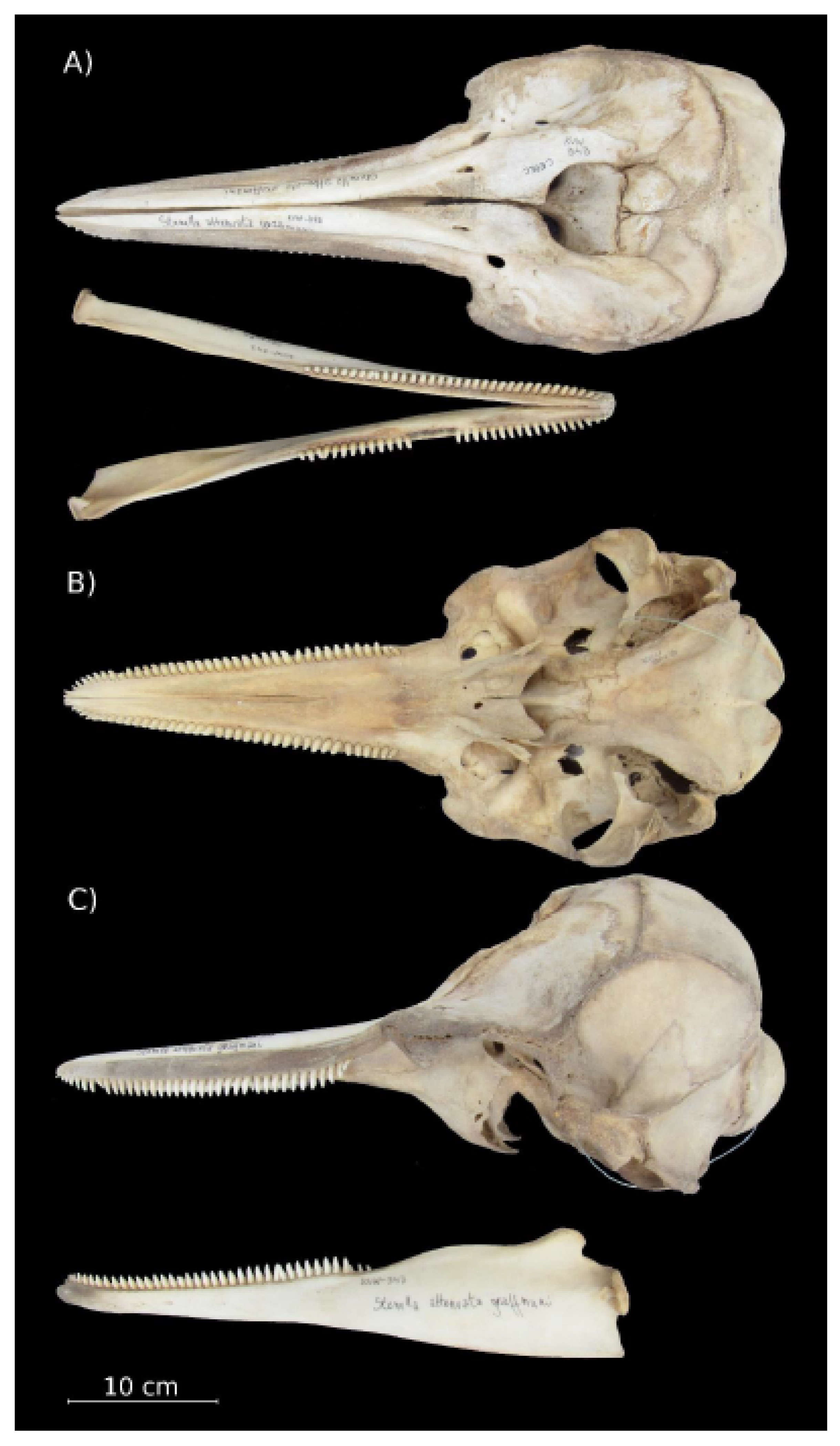
Figure 9.
Two extreme cases of SAG dolphins infested with the stalked barnacles Conchoderma auritum in the mouth of a calf (left) and the semi-stalked barnacle X. globicipitis: on the dorsal fin of an immature animal (right).
Figure 9.
Two extreme cases of SAG dolphins infested with the stalked barnacles Conchoderma auritum in the mouth of a calf (left) and the semi-stalked barnacle X. globicipitis: on the dorsal fin of an immature animal (right).

Figure 10.
Frequency distribution of records of SAG in Ecuador according to depth (n=56).
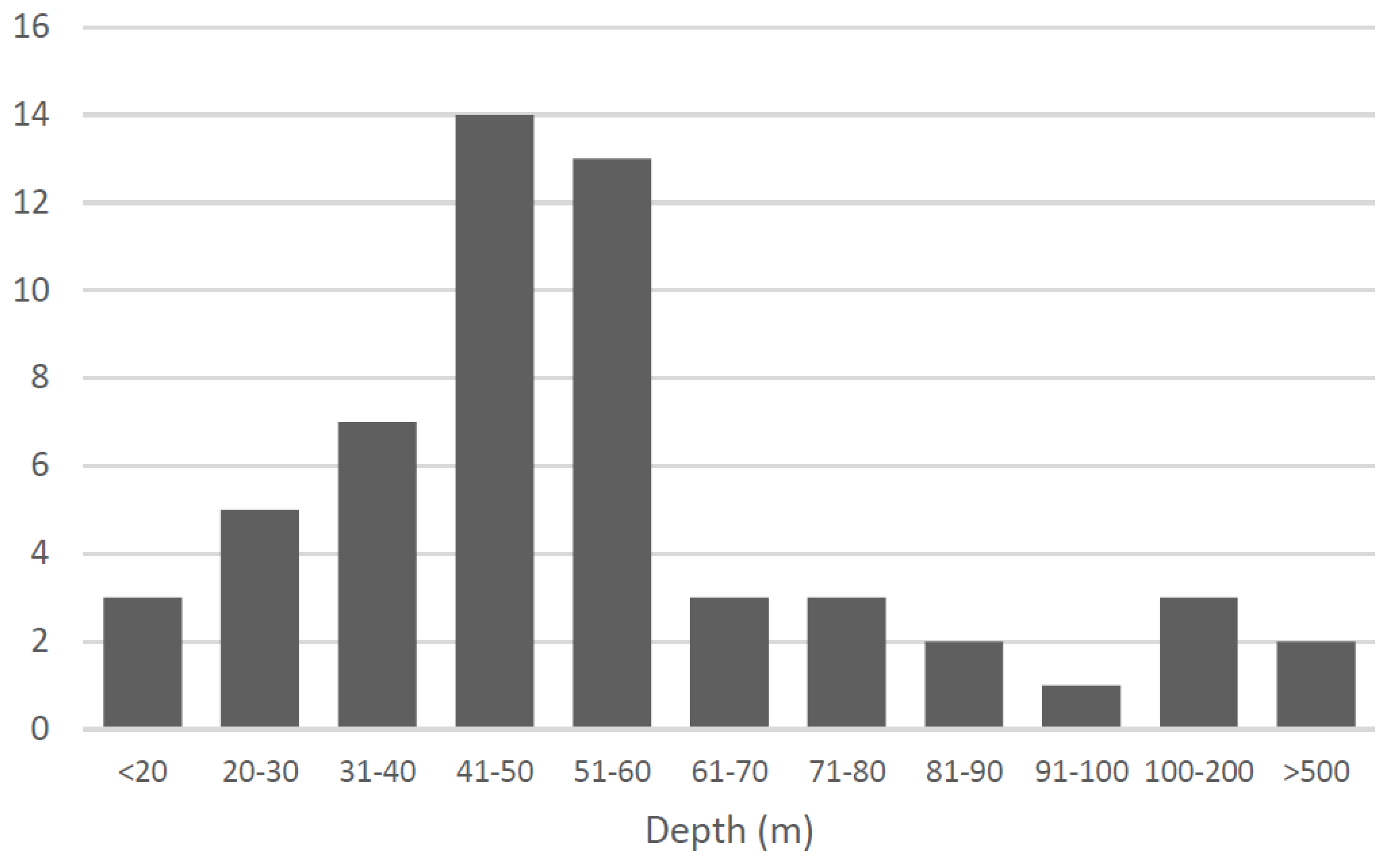
Figure 11.
Frequency distribution of SAG group size (n=52).
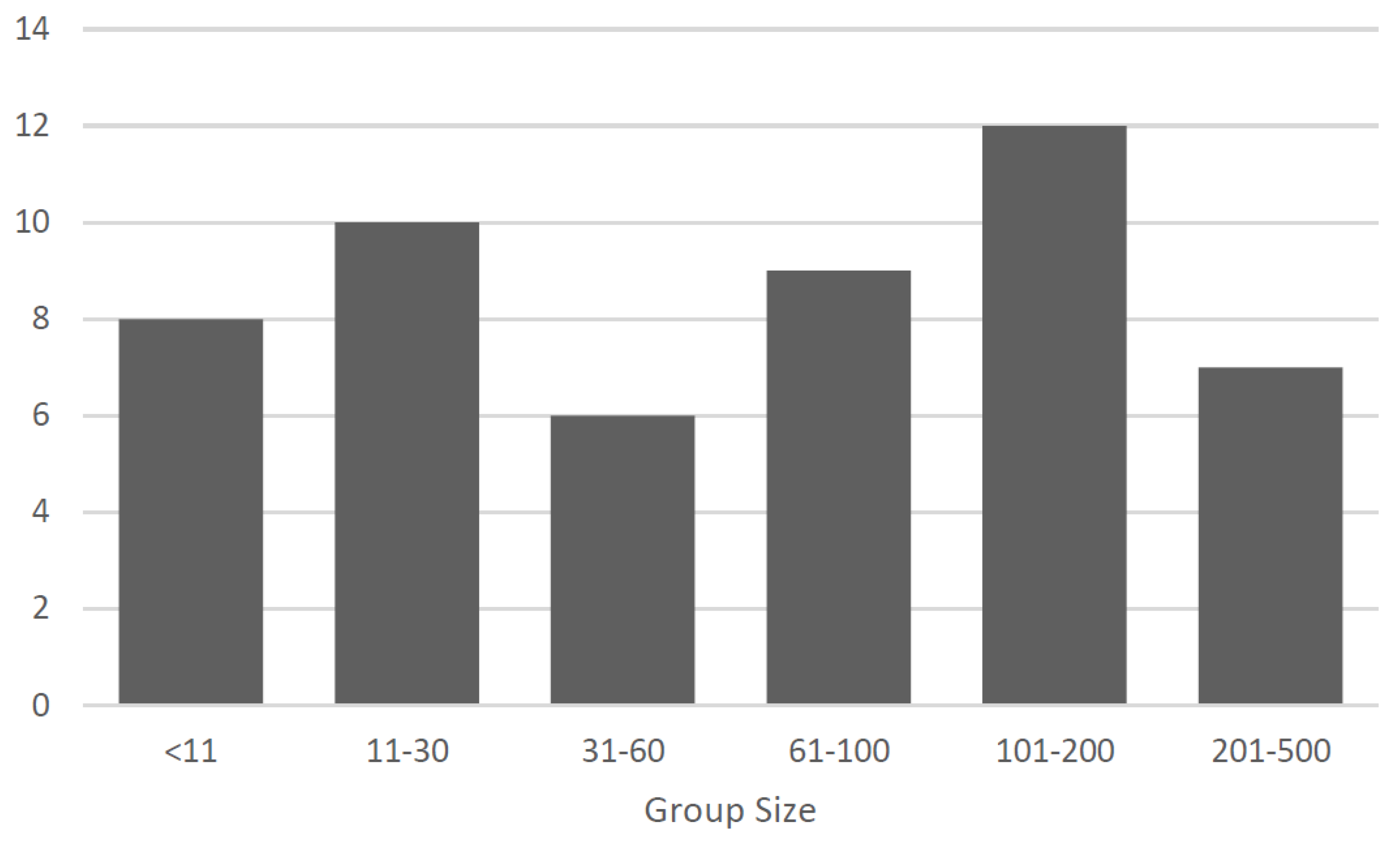
Figure 12.
Median-joining network of Stenella attenuata mtDNA CR. Circle size represents the number of individuals sharing the same haplotype. Circles are colored based on the geographic regions indicated in the legend. The white circle represents a missing or intermediate haplotype. Branch length is proportional to the number of mutational steps between haplotypes, with hatch marks indicating the total number of mutations. ETP: Eastern Tropical Pacific.
Figure 12.
Median-joining network of Stenella attenuata mtDNA CR. Circle size represents the number of individuals sharing the same haplotype. Circles are colored based on the geographic regions indicated in the legend. The white circle represents a missing or intermediate haplotype. Branch length is proportional to the number of mutational steps between haplotypes, with hatch marks indicating the total number of mutations. ETP: Eastern Tropical Pacific.
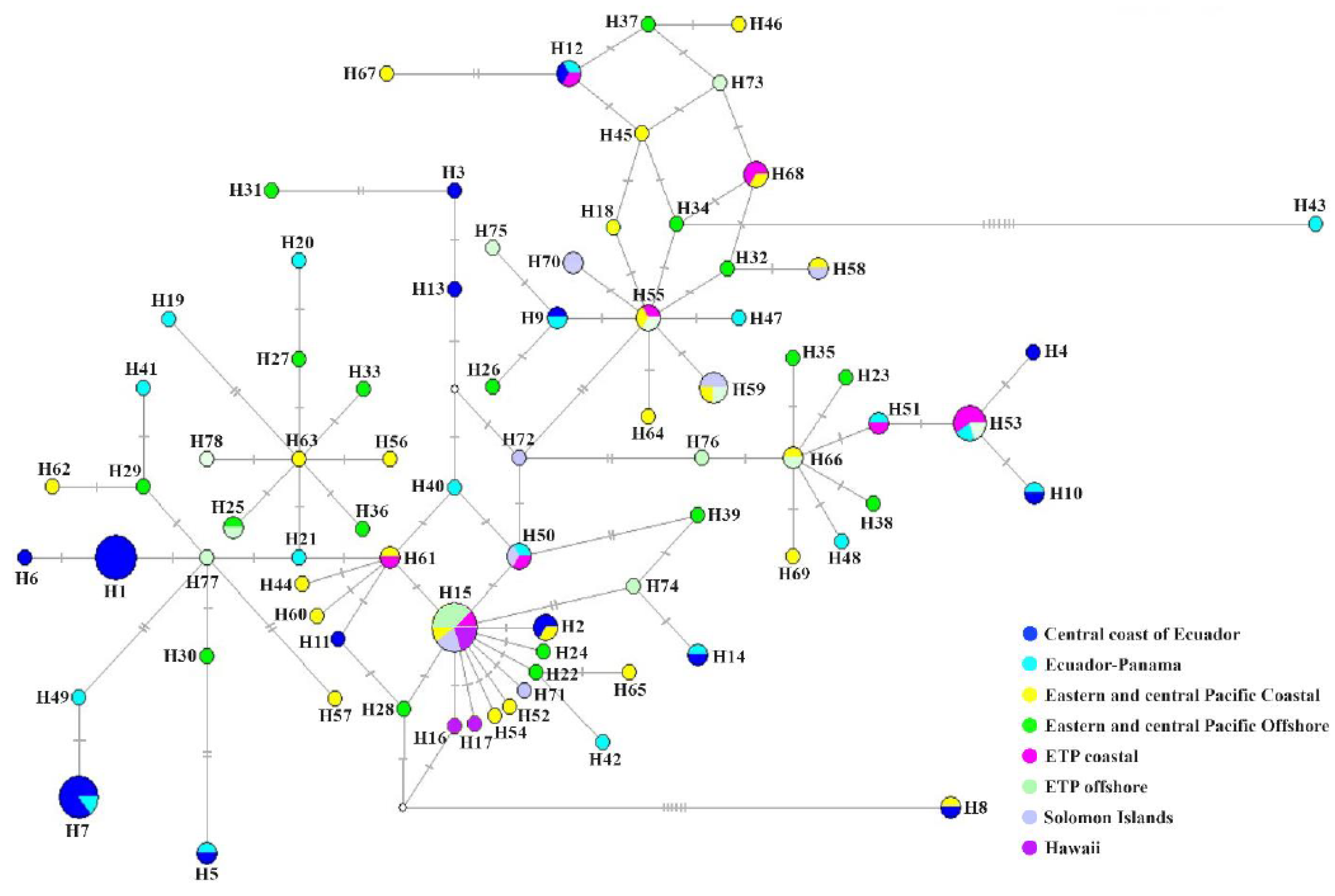
Figure 13.
Bayesian phylogenetic tree of spotted dolphins mtDNA CR. Bayesian phylogenetic tree inferred from a 405 bp segment of mtDNA CR. Posterior probability values are shown above the main branches. Sequences obtained from GenBank are labeled with accession numbers. The outgroup includes Lagenorhynchus albirostris (accession number NC_005278.1). Haplotypes representing sequences identified in the central coast of Ecuador (this study) are shown in violet. Ecuadorian sequences previously identified by Escorza-Treviño et al. (2005) are shown in black. ECU: Ecuador; ETP: Eastern Tropical Pacific.
Figure 13.
Bayesian phylogenetic tree of spotted dolphins mtDNA CR. Bayesian phylogenetic tree inferred from a 405 bp segment of mtDNA CR. Posterior probability values are shown above the main branches. Sequences obtained from GenBank are labeled with accession numbers. The outgroup includes Lagenorhynchus albirostris (accession number NC_005278.1). Haplotypes representing sequences identified in the central coast of Ecuador (this study) are shown in violet. Ecuadorian sequences previously identified by Escorza-Treviño et al. (2005) are shown in black. ECU: Ecuador; ETP: Eastern Tropical Pacific.
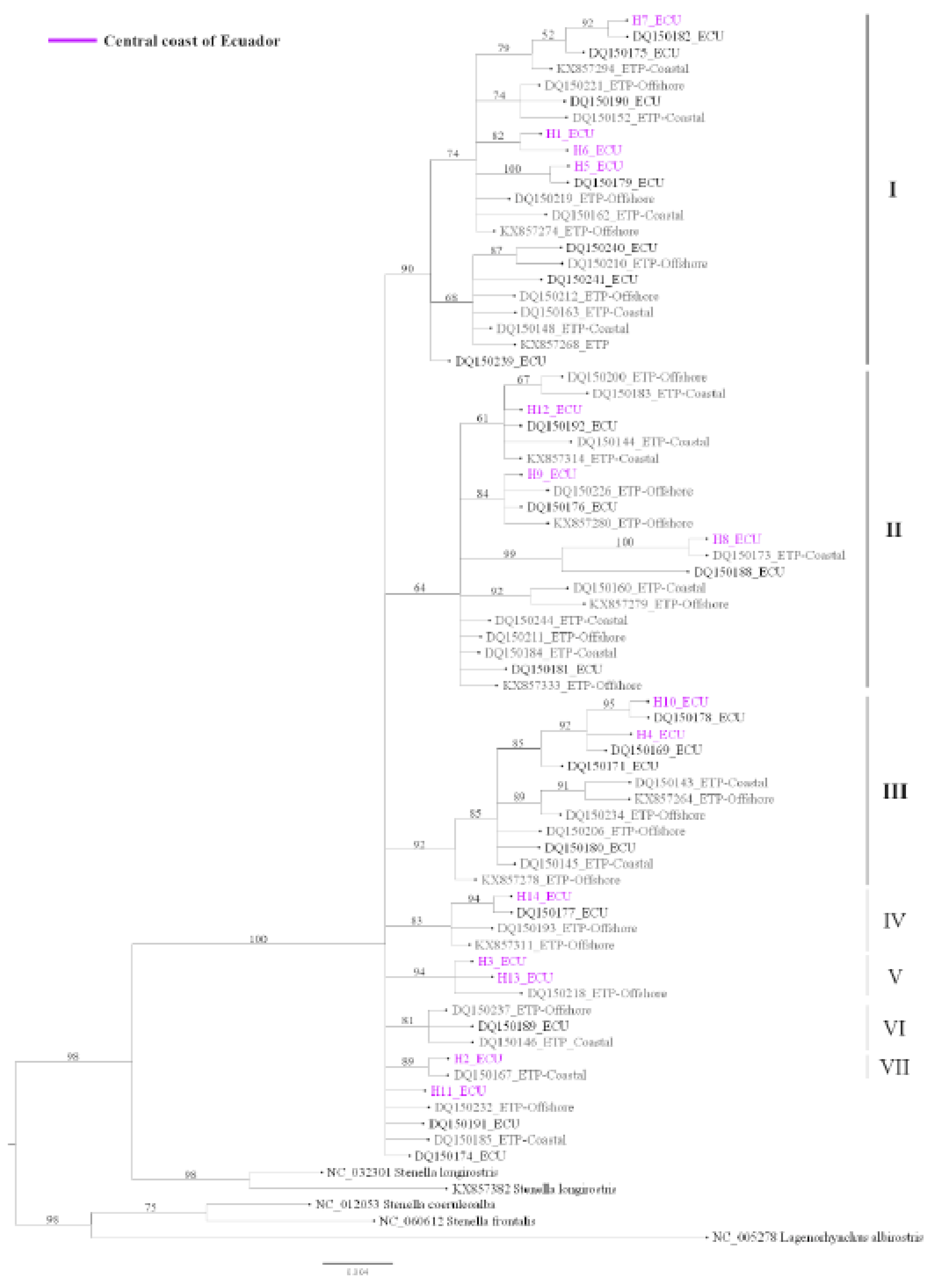
Figure 14.
Large-bodied, heavily spotted coastal form Stenella attenuata graffmani found stranded at Barbasquillo, Manta, Ecuador, on 07/12/2016. Tail flukes were necrotized from net entanglement. Photo MAE- PWF.
Figure 14.
Large-bodied, heavily spotted coastal form Stenella attenuata graffmani found stranded at Barbasquillo, Manta, Ecuador, on 07/12/2016. Tail flukes were necrotized from net entanglement. Photo MAE- PWF.


Table 1.
Stranding records, chronologically ordered, of S. attenuata graffmani on the coast of Ecuador, 1993–2021.
Table 1.
Stranding records, chronologically ordered, of S. attenuata graffmani on the coast of Ecuador, 1993–2021.
| Date | Site | Position | Sex | Length (cm) | Source |
|---|---|---|---|---|---|
| 5/10/1993 | San Vicente | 3º35'S. 80º24'W | 212 | Chiluiza et. al. (1998) | |
| 21/11/1993 | Anconcito | 2º22'S, 80º47'W | Chiluiza et. al. (1998) | ||
| 5/4/1994 | Montañita | 1º49'S 80º45'W | 204 | Chiluiza et. al. (1998) | |
| 30/4/1995 | Libertador Bolívar | 1º56'S, 80º44'W | Chiluiza et. al. (1998) | ||
| 12/7/1995 | Olón | 1º50'S, 80º34'W | 191 | Chiluiza et. al. (1998) | |
| 2/9/1995 | Playas | 2º37'S, 80º23'W | Chiluiza et. al. (1998) | ||
| 3/9/1995 | Monte Verde | 2º03'S, 80º44'W | Chiluiza et. al. (1998) | ||
| 25/5/2007 | Ayangue | 1º58'S, 80º45'W | M | » 200 | Félix et al. (2011) |
| 14/4/2006 | Mar Bravo | 2º15'S, 80º56'W | 190 | Félix et al. (2011) | |
| 23/12/2002 | San José | 1º46'S, 80º46'W | M | 193 | Félix et al. (2011) |
| 21/3/2004 | Mar Bravo | 2º13'S, 80º57'W | »120 | Félix et al. (2011) | |
| 14/9/2004 | Punta Carnero | 2º17'S, 80º54'W | Félix et al. (2011) | ||
| 19/9/2004 | Mar Bravo | 2º14'S, 80º56'W | F | 228 | Félix et al. (2011) |
| 11/11/2005 | Playa FAE | 2º11'S, 80º59'W | >200 | Félix et al. (2011) | |
| 16/2/2006 | Punta Carnero | 2º17'S, 80º54'W | M | 92 | Félix et al. (2011) |
| 18/11/2006 | Mar Bravo | 2º16'S, 80º55'W | M | 181 | Félix et al. (2011) |
| 2/12/2006 | Mar Bravo | 2º15'S, 80º55'W | F | 218 | Félix et al. (2011) |
| 13/5/2007 | Diablica | 2º18'S, 80º54'W | Félix et al. (2011) | ||
| 13/1/2008 | Mar Bravo | 2º14'S, 80º57'W | F | 186 | Félix et al. (2011) |
| 31/10/2009 | Punta Carnero | 2º17'S, 80º54'W | F | 179 | Félix et al. (2011) |
| 2/1/2010 | Mar Bravo | 2º15'S, 80º56W | 195 | This report | |
| 1/5/2015 | Diablica | 2º17'S, 80º54W | » 200 | This report | |
| 31/8/2017 | San Pablo | 2º06'S, 80º44W | F | 190 | This report |
| 27/6/2018 | Cauchiche | 2º48'S, 80º14'W | This report | ||
| 10/7/2021 | Mar Bravo | 2º14'S, 80º57'W | Chocho et al. (2023) |
Table 2.
Sightings of S. attenuata graffmani (n=59) on the central coast of Ecuador, 2000–2024.
| Date | Position | Group size | Remarks |
|---|---|---|---|
| 23/7/2000 | 1°18'S, 81°02'W | 6-10 | With humpback whales |
| 2/7/2002 | 2º09'S, 81º02'W | With humpback whales | |
| 1/8/2003 | 2º16'S, 81º06'W | 30-40 | Feeding |
| 23/9/2004 | 2º15'S, 81º04'W | ~100 | With humpback whales |
| 2/10/2004 | 2º06'S, 81º00'W | 100-200 | Several subgroups |
| 14/8/2005 | 2°08'S, 80°53'W | 60-80 | |
| 18/8/2005 | 2°11'S, 81°01'W | ~80 | |
| 21/8/2005 | 2°10'S, 80°58'W | ~60 | Feeding |
| 9/7/2007 | 2°08'S, 81°00´W | 3 | Feeding |
| 15/7/2008 | 2°10'S, 81°00´W | ~30 | |
| 11/8/2008 | 2°06'S, 81°03'W | With humpback whales | |
| 11/7/2010 | 2°09'S, 80°59’W | 50-100 | |
| 28/7/2010 | 80-100 | ||
| 29/7/2010 | 2°08'S, 81°01'W | ~50 | |
| 4/8/2010 | 2°11'S, 80°58'W | ~60 | |
| 5/8/2010 | ~40 | ||
| 12/8/2010 | 2°07.'S, 81°05'W | With humpback whales and common bottlenose dolphins | |
| 13/8/2010 | 2°12'S, 81°03'W | 300-400 | |
| 14/8/2010 | 60-80 | ||
| 23/8/2010 | 2°13'S, 81°03'W | 6-8 | With humpback whales |
| 19/9/2010 | 2°08'S, 81°00’W | 15-20 | |
| 2021-03-05 | 1°36'S, 81°00'W | 15-20 | |
| 2022-03-25 | 1°47'S, 80°51'W | ~100 | |
| 2022-12-20 | 1°26'S, 80°56'W | 4 | |
| 2023-01-22 | 1°23'S, 81°04'W | 18-20 | |
| 2023-02-25 | 1°27'S, 80°59'W | ~200 | Feeding |
| 7/3/2023 | 1°19'S, 80°58'W | ~200 | |
| 8/3/2023 | 1°24'S, 80°58'W | ~100 | Feeding |
| 26/2/2024 | 1°25'S, 81°07'W | ~500 | |
| 28/2/2024 | 1°25'S, 81°08'W | ~500 | |
| 14/3/2024 | 1°26'S, 80°59'W | ~200 | Feeding , with Bryde's whales |
| 14/3/2024 | 1°33'S, 81°11'W | ~500 | Feeding |
| 17/3/2024 | 1°19'S, 80°59'W | ~200 | Feeding |
| 3/4/2024 | 1°24'S, 81°11'W | ~50 | |
| 11/4/2024 | 1°21'S, 80°57'W | ~20 | |
| 11/4/2024 | 1°16'S, 81°04'W | ~50 | |
| 18/4/2024 | 1°25'S, 80°57'W | ~200 | |
| 18/4/2024 | 1°35'S, 80°53'W | 5 | With Bryde's whales |
| 1/5/2024 | 1°25'S, 80°56'W | ~300 | |
| 2/5/2024 | 1°25'S, 80°55'W | ~300 | Feeding |
| 4/6/2024 | 1°26'S, 80°54'W | 10 | With Bryde's whales |
| 27/7/2024 | 1°33'S, 80°56'W | ~300 | With humpback whales |
| 15/8/2024 | 1°19'S, 81°00'W | ~20 | With humpback whales |
| 17/8/2024 | 1°24'S, 80°57'W | ~20 | |
| 19/8/2024 | 1°21'S, 80°59'W | ~200 | |
| 24/8/2024 | 1°16'S, 81°02'W | ~200 | |
| 26/8/2024 | 1°25'S, 80°56'W | ~20 | With Bryde's whales |
| 3/6/2024 | 1°35'S, 80°52'W | ~200 | Feeding |
| 8/9/2024 | 1°25'S, 80°56'W | ~100 | |
| 11/9/2024 | 1°24'S, 80°54'W | ~200 | |
| 20/9/2024 | 1°26'S, 80°54'W | ~20 | |
| 20/9/2024 | 1°26'S, 80°55'W | ~20 | |
| 21/9/2024 | 1°23'S, 80°58'W | ~200 | |
| 21/9/2024 | 1°24'S, 80°57'W | ~200 | Feeding |
| 22/9/2024 | 1°25'S, 80°56'W | ~100 | Feeding |
| 1/10/2024 | 1°28'S, 80°52'W | ~200 | With humpback whales |
| 19/10/2024 | 1°17'S, 81°1'W | 10 | |
| 26/10/2024 | 1°24'S, 80°55'W | 3 | |
| 28/10/2024 | 1°26'S, 80°54'W | 5 |
Table 3.
Specimen records of S. attenuata graffmani from Peru, ordered chronologically. Abbreviations: Standard body length (SL), Not available (NA).
Table 3.
Specimen records of S. attenuata graffmani from Peru, ordered chronologically. Abbreviations: Standard body length (SL), Not available (NA).
| Specimen | Date | Location | Coord. | Sex, maturity | SL (cm) | Circumstances | Voucher, Museo de Delfines, CEPEC |
|---|---|---|---|---|---|---|---|
| KVW-343 | 20/04/1985 | Landed at Cerro Azul port, Lima Department | 13°02'S, 76°29'W | Adult female (lactating) | 211 | Gillnet bycatch | skull |
| KVW-344 | 20/04/1985 | Landed at Cerro Azul port, Lima Dept. | 13°02'S, 76°29'W | Female. Small calf. Milk in stomach. | 106 | Gillnet bycatch, landed with KVW-343 | skull |
| AJR-036 | 24/02/1986 | Taken at Punta Negra, landed at Pucusana port, Lima Dept. | 12°22'S, 76°48'W | Male. Sexually immature; cranially subadult | 188 | Harpooned; collected by A.J. Read | skull |
| KVW-1999 | 28/12/1989 | Playa Michallo, Bahía Sechura (sur), Piura Dept. | 05°35'S, 80°57'W | Sex unknown; cranially mature | NA | Beach pick-up (Santiago Zambrano) | skull |
Table 4.
Standard craniometrics of adult specimens of S. a. graffmani compared to equivalent data for a large sample (n= 75-80, depending on variable) of S. a. attenuata presented by Perrin (1975). Values in blue indicate non-overlapping between the two morphs. % CBL = percent of condylobasal length. .
Table 4.
Standard craniometrics of adult specimens of S. a. graffmani compared to equivalent data for a large sample (n= 75-80, depending on variable) of S. a. attenuata presented by Perrin (1975). Values in blue indicate non-overlapping between the two morphs. % CBL = percent of condylobasal length. .
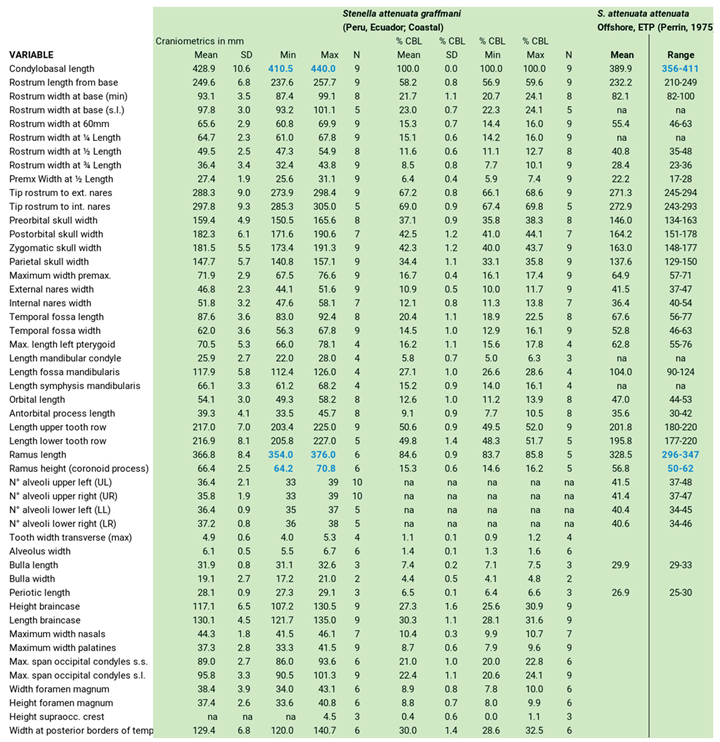 |
Table 5.
Nucleotide and haplotype frequencies of the control region of S. a. graffmani from central Ecuador. Polymorphic sites and haplotype frequencies of the 405 bp mtDNA CR are shown. Sequence identity was assessed relative to the reference haplotype (HR; GenBank accession number NC_012051.1). Numbers in the first three rows indicate the positions of polymorphic sites. Dots (.) denote identity with HR.
Table 5.
Nucleotide and haplotype frequencies of the control region of S. a. graffmani from central Ecuador. Polymorphic sites and haplotype frequencies of the 405 bp mtDNA CR are shown. Sequence identity was assessed relative to the reference haplotype (HR; GenBank accession number NC_012051.1). Numbers in the first three rows indicate the positions of polymorphic sites. Dots (.) denote identity with HR.
| Haplotype | Polymorphic sites | Haplotype frequency | |||||||||||||||||||||||||
| 1 | 2 | 5 | 7 | 8 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| 6 | 6 | 1 | 9 | 8 | 0 | 3 | 3 | 4 | 6 | 7 | 2 | 3 | 4 | 4 | 5 | 5 | 6 | 6 | 7 | 1 | 5 | 5 | 7 | 7 | 9 | ||
| 4 | 4 | 6 | 9 | 5 | 5 | 7 | 0 | 1 | 7 | 1 | 9 | 4 | 5 | 2 | 1 | 2 | 5 | 2 | 3 | 3 | |||||||
| Hap_R | C | T | T | T | G | T | G | A | T | T | A | T | A | T | A | C | T | T | T | A | C | C | T | A | C | A | |
| Hap_1 | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | C | . | . | G | 8 |
| Hap_2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | . | T | . | . | . | T | . | 2 |
| Hap_3 | . | . | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | C | . | . | . | . | . | . | . | 1 |
| Hap_4 | . | . | . | . | . | C | . | . | . | . | . | C | . | . | . | T | . | . | C | . | T | T | . | G | T | . | 1 |
| Hap_5 | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | T | . | . | C | . | . | T | . | . | T | G | 1 |
| Hap_6 | . | . | . | . | A | . | . | C | . | . | . | . | . | . | . | . | . | . | C | . | . | . | C | . | . | G | 1 |
| Hap_7 | . | . | . | . | A | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | T | . | . | . | G | 6 |
| Hap_8 | T | C | . | . | . | . | A | . | . | . | . | . | G | . | T | . | C | . | . | G | . | . | . | . | T | . | 1 |
| Hap_9 | . | C | . | . | . | . | A | . | . | . | G | . | . | . | . | . | . | C | C | . | . | T | . | . | T | . | 1 |
| Hap_10 | . | . | . | . | . | C | . | . | C | . | . | C | . | . | . | T | . | C | C | . | T | T | . | G | T | . | 1 |
| Hap_11 | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | . | C | . | . | . | . | . | . | . | 1 |
| Hap_12 | . | C | . | . | . | . | A | . | . | C | . | C | G | . | . | . | . | C | C | . | . | T | . | . | T | . | 1 |
| Hap_13 | . | . | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | C | C | . | . | T | . | . | . | . | 1 |
| Hap_14 | . | . | . | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | G | . | . | . | . | T | . | 1 |
| 1 | Published in Arkiv för Zoologi which has ceased to exist. It was a Swedish scientific journal on zoology published between 1903 and 1974. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



