Submitted:
03 February 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Panaeolus is a genus of small, dark-spored agarics within the family Galeropsidaceae Singer. Based on the majority of specimens collected from China, this study investigated the genus Panaeolus and identified 17 species. These include four new species: Panaeolus bambusicola, P. latifolius, P. praecox, and P. ovinus; and one new record for China: P. fraxinophilus. The new species and the newly recorded species for China are morphologically described and illustrated. A multi-locus phylogenetic analysis (ITS, nrLSU, tef1-α, rpb2) was conducted using maximum likelihood and Bayesian inference. Combined morphological and phylogenetic evidence supports the reduction of the genus Crucispora to a subgenus within Panaeolus, accommodating P. rhombispermus.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Sampling and Morphological Analyses
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Molecular Phylogeny
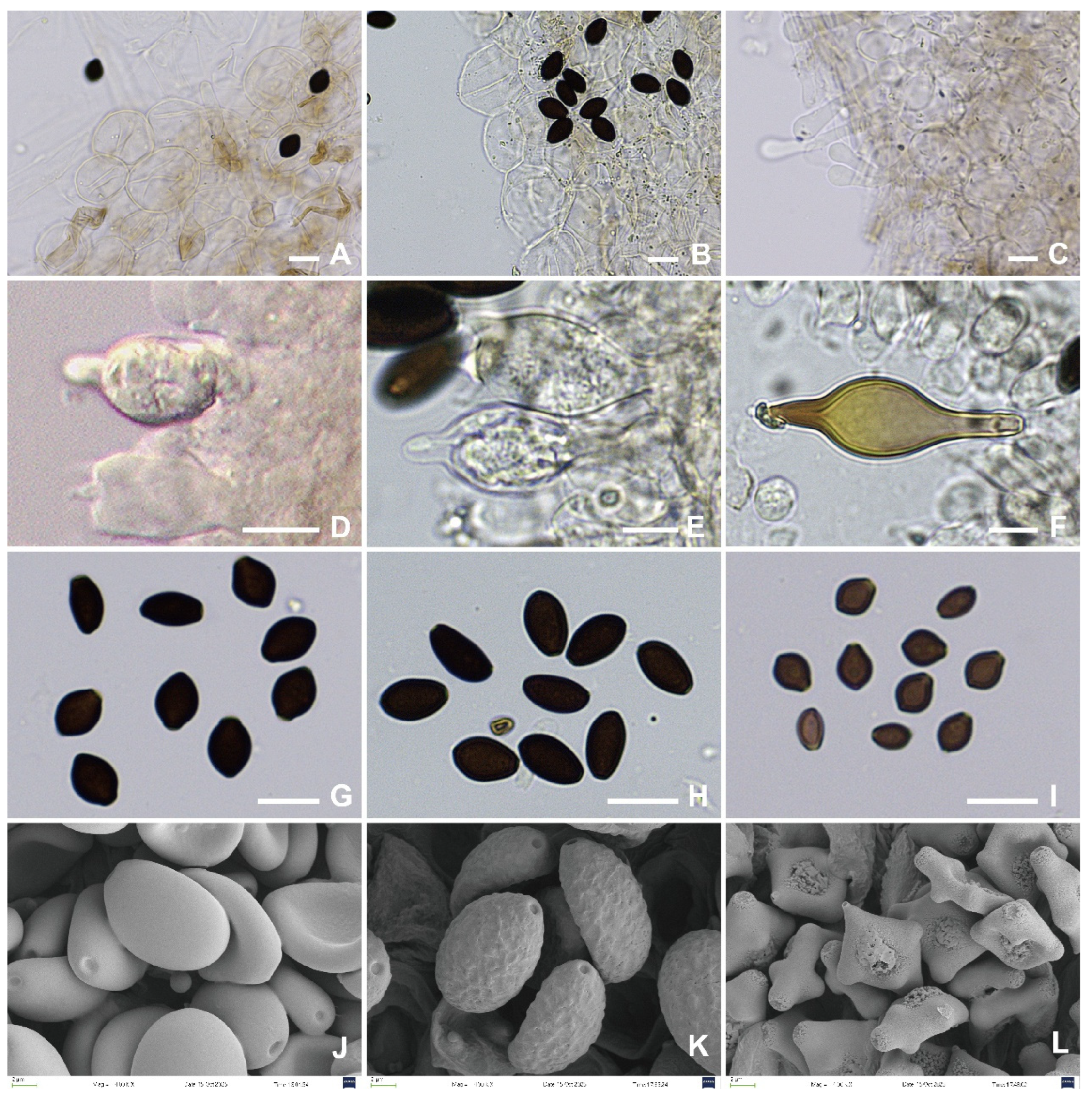
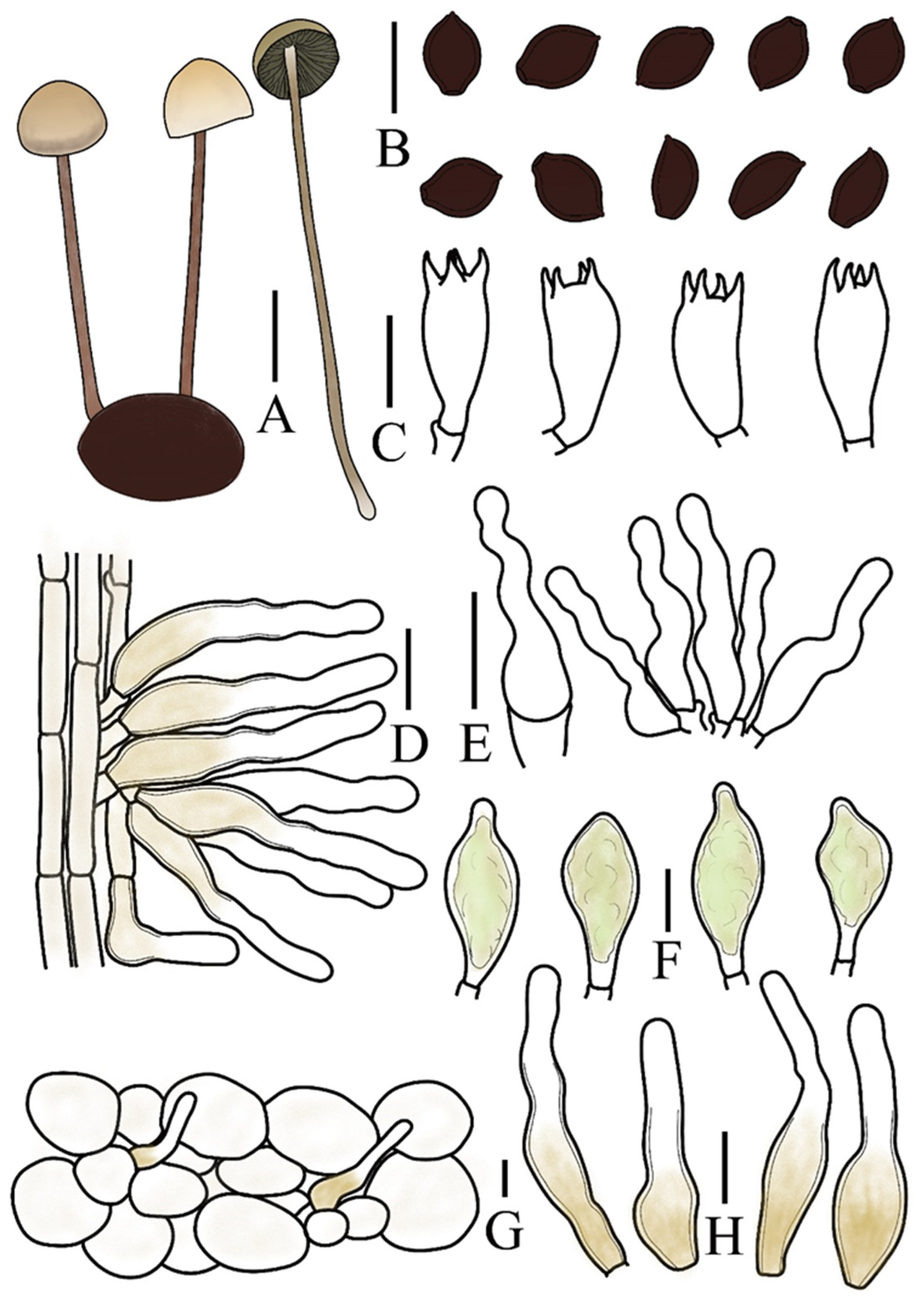
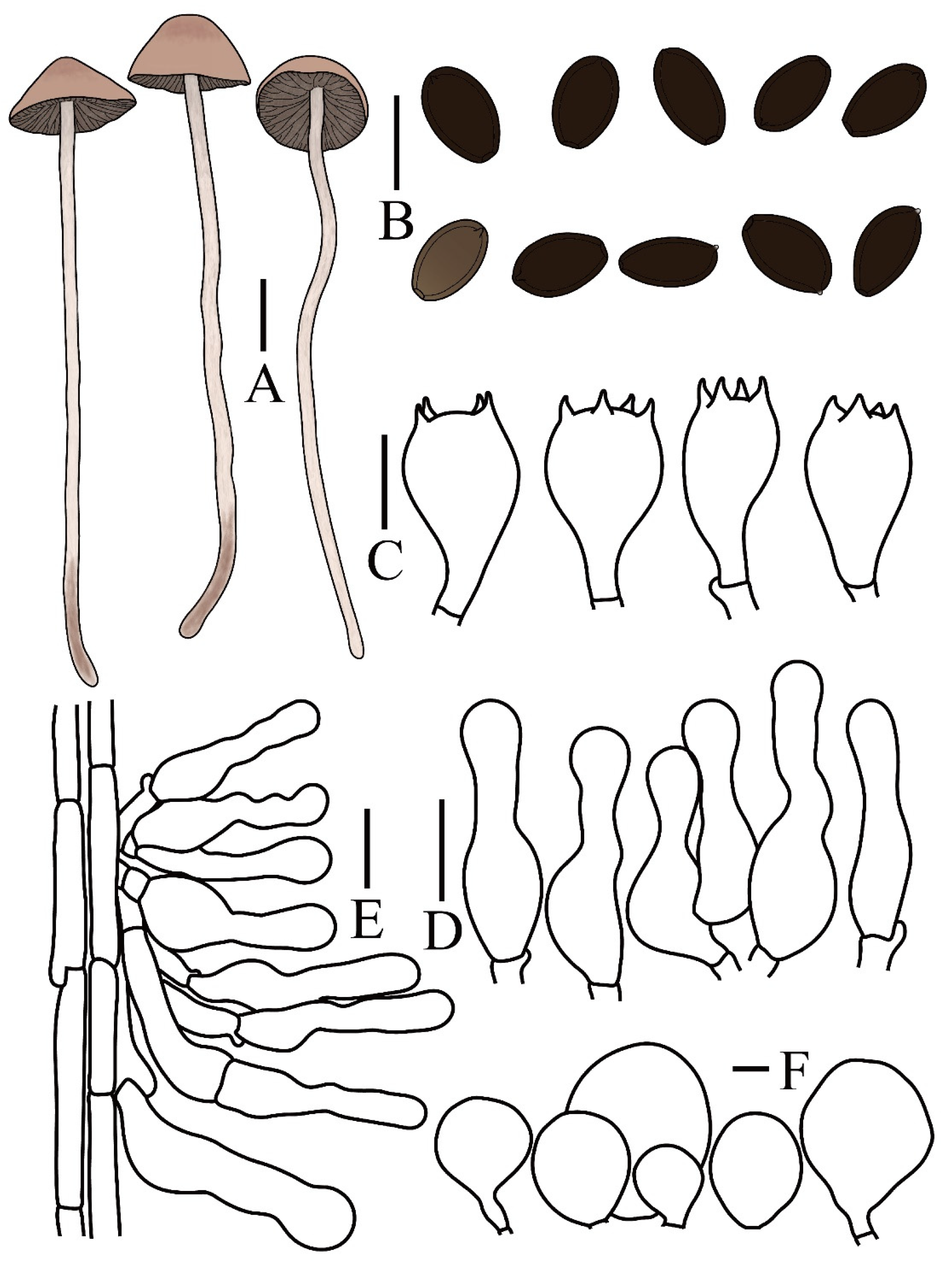
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fries, E. Epicrisis Systematis Mycologici: seu synopsis hymenomycetum; sumtibus auctoris: 1836; Volume 1.
- Quélet, L. Les champignons du Jura et des Vosges; Impr. et lithographie de H. Barbier, 1872. [Google Scholar]
- Pollock, S.H. A Novel Experience with Panaeolus A Case Study from Hawaii. Journal of Psychedelic Drugs 1974, 6, 85–89. [Google Scholar] [CrossRef]
- Watling, R. A Panaeolus poisoning in Scotland. Mycopathologia 1977, 61, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Stijve, T. Psilocin, psilocybin, serotonin and urea in Panaeolus cyanescens from various origin. Persoonia-Molecular Phylogeny and Evolution of Fungi 1992, 15, 117–121. [Google Scholar]
- Oláh, G.-M. Le genre Panaeolus: Essai taxinomique et physiologique; Laboratoire de cryptogamie du Muséum national d’histoire naturelle, 1969. [Google Scholar]
- Shaffer, R.L. The Agaricales in modern taxonomy. 1976.
- Tóth, A.; Hausknecht, A.; Krisai-Greilhuber, I.; Papp, T.; Vágvölgyi, C.; Nagy, L.G. Iteratively refined guide trees help improving alignment and phylogenetic inference in the mushroom family Bolbitiaceae. PLoS One 2013, 8, e56143. [Google Scholar] [CrossRef] [PubMed]
- Malysheva, E.; Moreno, G.; Villarreal, M.; Malysheva, V.; Svetasheva, T. The secotioid genus Galeropsis (Agaricomycetes, Basidiomycota): a real taxonomic unit or ecological phenomenon? Mycological Progress 2019, 18, 805–831. [Google Scholar] [CrossRef]
- Kalichman, J.; Kirk, P.M.; Matheny, P.B. A compendium of generic names of agarics and Agaricales. Taxon 2020, 69, 425–447. [Google Scholar] [CrossRef]
- He, M.-Q.; Yang, W.-Q.; Phurbu, D.; Liu, F.; Li, J.-X.; Cao, B.; Zhao, R.-L. Systematic study of Panaeolus (Agaricales, Galeropsidaceae) sensu lato and psilocybin-producing traits of species from China. IMA Fungus 2026, 17, e167329. [Google Scholar] [CrossRef]
- Horak, E. Contributions to the knowledge of the Agaricales sl (Fungi) of New Zealand. New Zealand Journal of Botany 1971, 9, 463–493. [Google Scholar] [CrossRef]
- Hongo, T. Notulae Mycologicae (12): Auctore. Bulletin of the Faculty of Education, Shiga University. Natural Sciences 1973, 37–43. [Google Scholar]
- Ostuni, S.; Voto, P.; Birkebak, J.; Meyer, M.G.; Geurin, Z.; Slot, J.C. Panaeolus oligotrophus: A new species from central Florida, with notes on Panaeolus pumilus and Crucispora rhombisperma. Mycologia 2025, 117, 1057–1066. [Google Scholar] [CrossRef]
- Li, Y.; Li, T.H.; Yang, Z.L.; Dai, Y.C.; Bau, T. Atlas of Chinese macrofungal resources; Central China Farmers’ Publishing House: Zhengzhou, 2015. [Google Scholar]
- Bau, T.; Sarentoya. Strophariaceae of China (V) Panaeolus s.l. Journal of Fungal Research 2010, 8, 131–168. [Google Scholar] [CrossRef]
- He, X.-S. A new record of Panaeolina in China. Mycosystema 2004, 150. [Google Scholar] [CrossRef]
- Ma, T. Taxonomy of Psilocybe sl and Panaeolus in Yunnan, Southwest China, with notes on related genus Protostropharia. Beijing: Chinese Academy of Forestry 2014.
- Hu, Y.; Mortimer, P.E.; Karunarathna, S.C.; Raspe, O.; Promputtha, I.; Yan, K.; Xu, J.; Hyde, K.D. A new species of Panaeolus (Agaricales, Basidiomycota) from Yunnan, Southwest China. Phytotaxa 2020, 434, 22–34-22–34. [Google Scholar] [CrossRef]
- Cao, B.; Phurbu, D.; Ralaiveloarisoa, A.; Liimatainen, K.; Niskanen, T.; Ramírez-Cruz, V.; Bradshaw, A.J.; Dentinger, B.T.; Ramírez-Guillén, F.; Cortés-Pérez, A. Fungal diversity notes 1919–2016: taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 2025, 1–227. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen handbook of colour. 1967.
- Weeks, R.A.; Singer, R.; Hearn, W.L. A new psilocybian species of Copelandia. Journal of Natural Products 1979, 42, 469–474. [Google Scholar] [CrossRef]
- Tj, W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols: a guide to methods and applications 1990, 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Molecular ecology 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Moncalvo, J.-M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Systematic biology 2000, 49, 278–305. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Molecular phylogenetics and evolution 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular biology and evolution 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zhao, D.; Ye, T.; Gao, F.; Jakovlić, I.; La, Q.; Tong, Y.; Liu, X.; Song, R.; Liu, F.; Lian, Z.m. PhyloSuite v2: The development of an all-in-one, efficient and visualization-oriented suite for molecular dating analysis and other advanced features. iMeta 2025, 4, e70095. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nature methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Wong, T.K.; Ly-Trong, N.; Ren, H.; Baños, H.; Roger, A.J.; Susko, E.; Bielow, C.; De Maio, N.; Goldman, N.; Hahn, M.W. IQ-TREE 3: Phylogenomic Inference Software using Complex Evolutionary Models. 2025.
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic biology 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Malysheva, E.F.; Malysheva, V.F.; Svetasheva, T.Y. Molecular phylogeny and taxonomic revision of the genus Bolbitius (Bolbitiaceae, Agaricales) in Russia. Mycological Progress 2015, 14, 64. [Google Scholar] [CrossRef]
- Song, H.-B.; Bau, T. Conocybe section Pilosellae in China: Reconciliation of taxonomy and phylogeny reveals seven new species and a new record. Journal of Fungi 2023, 9, 924. [Google Scholar] [CrossRef]
- Voto, P.; Angelini, C. Studies in the Panaeolus papilionaceus complex (Agaricales, Galeropsidaceae): two new species discovered in the Dominican Republic and Australia. 2024.
- Strauss, D.; Ghosh, S.; Murray, Z.; Gryzenhout, M. Global species diversity and distribution of the psychedelic fungal genus Panaeolus. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Consiglio G, M.M.C. Contributo alla conoscenza del genere Panaeolus sensu lato. Rivista di Micologia 2023, 66, 209–286. [Google Scholar]
- Voto, P.; Angelina, C. First record of Copelandia mexicana in Dominican Republic and notes on Panaeolus. Mycological Observations 2021, 1, 44–58. [Google Scholar]
- Asif, M.; Firdous, Q.; Izhar, A.; Niazi, A.R.; Sarwar, S.; Khalid, A.N. Molecular and morphological studies reveal a new species of Panaeolus (Agaricales, Basidiomycota) from Punjab, Pakistan. European journal of taxonomy 2023, 888, 77–96. [Google Scholar] [CrossRef]
- Chou, W.-N.; Yang, B.-H.; Hsieh, H.-M.; Ju, Y.-M. Crucispora rhombisperma newly recorded in Taiwan. Fungal Science 2021, 36, 33–36. [Google Scholar]
- Moser, M. Panaeolus alcidis, a new species from Scandinavia and Canada. Mycologia 1984, 76, 551–554. [Google Scholar] [CrossRef]
- Pegler, D.; Henrici, A. Panaeolus atrobalteatus sp. nov., a member of Panaeolus stirps Subbalteatus (Agaricales, Strophariaceae). 1998.

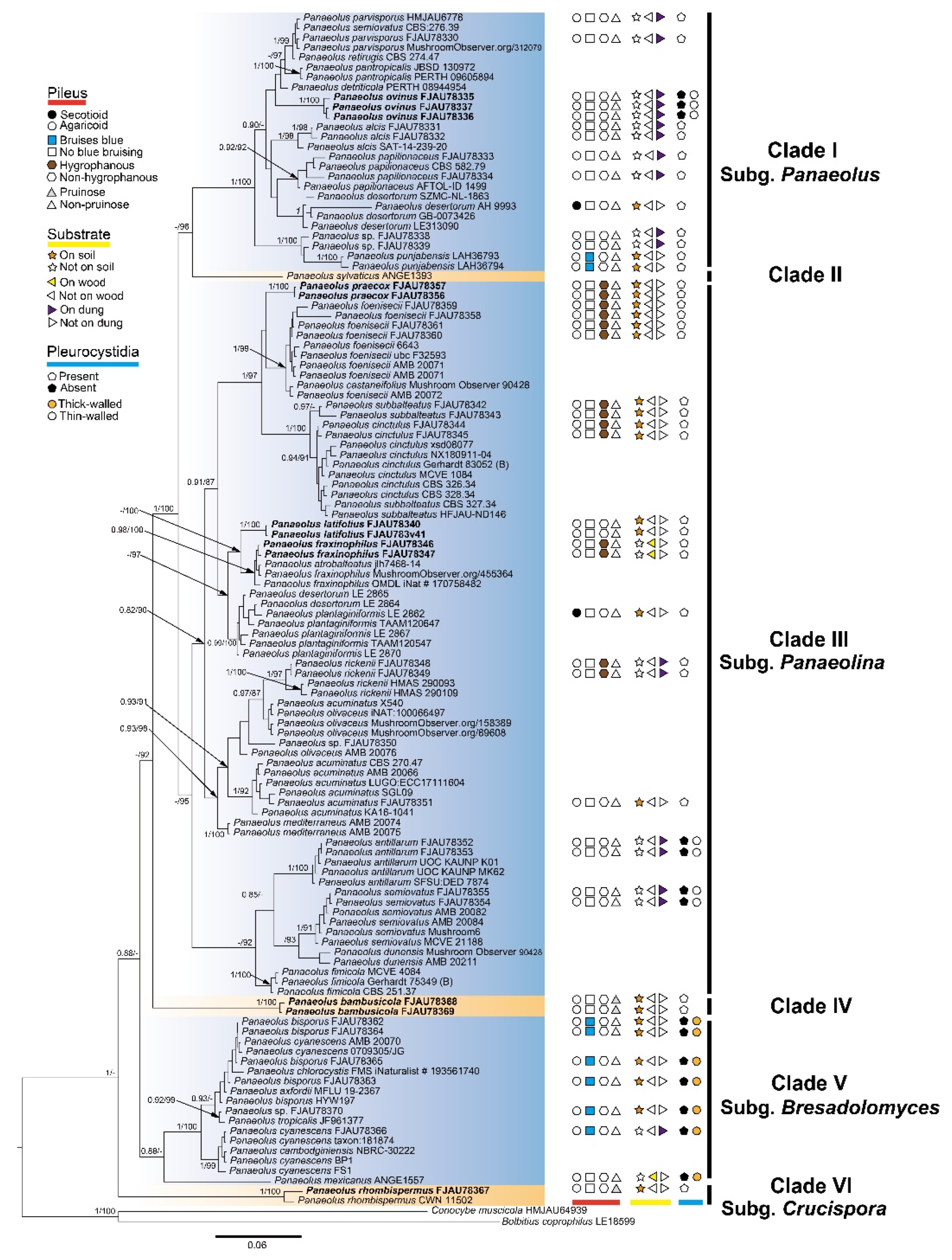
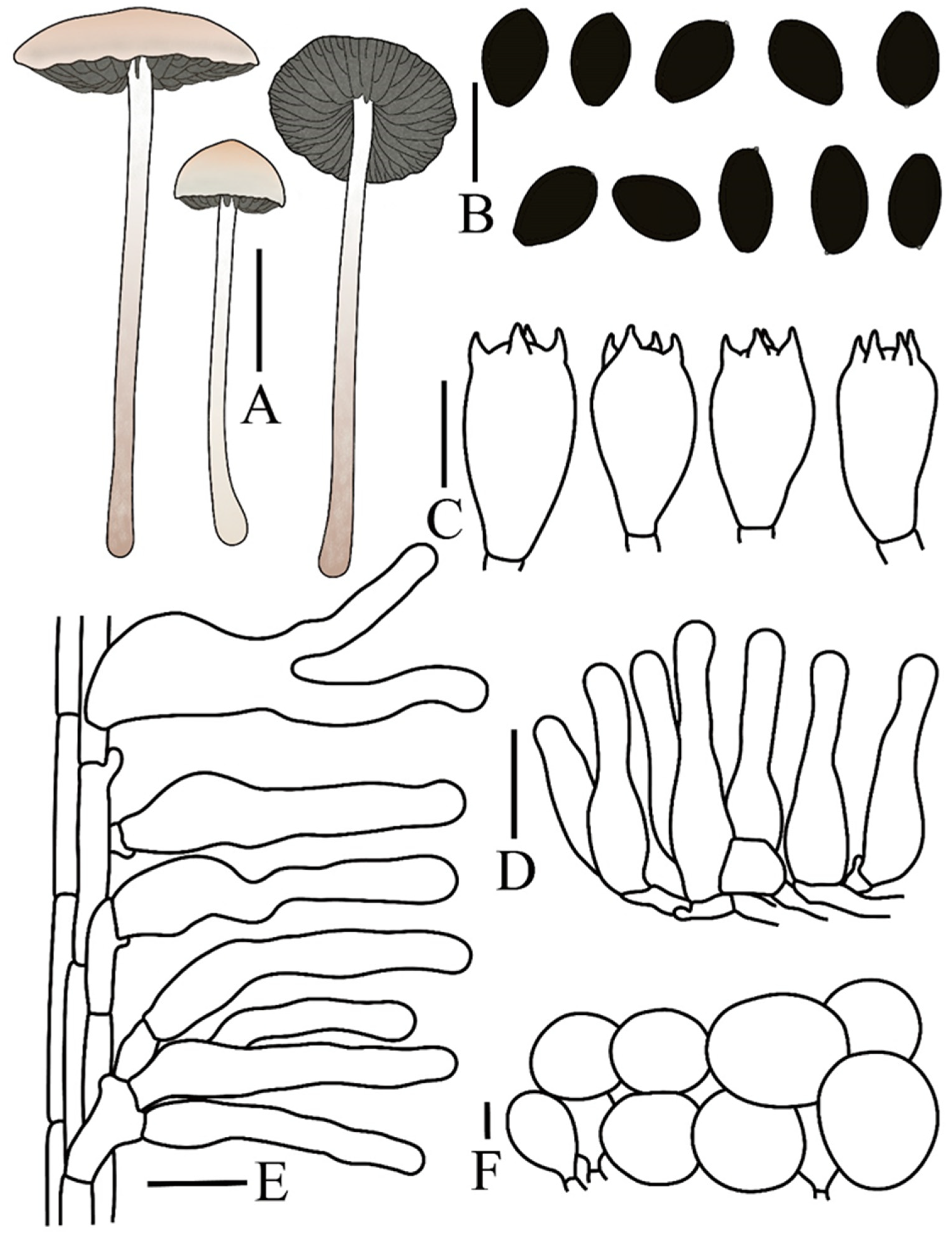
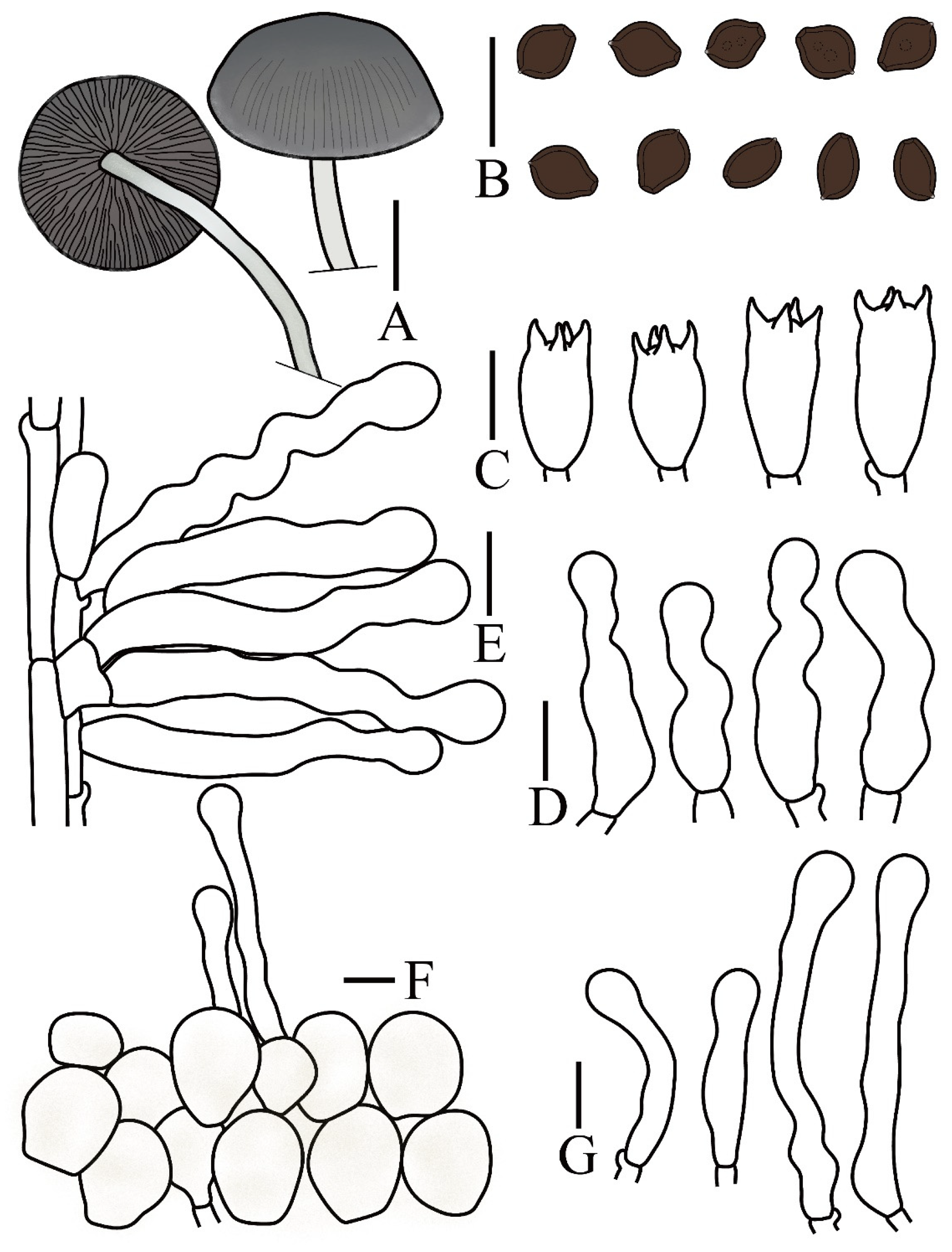
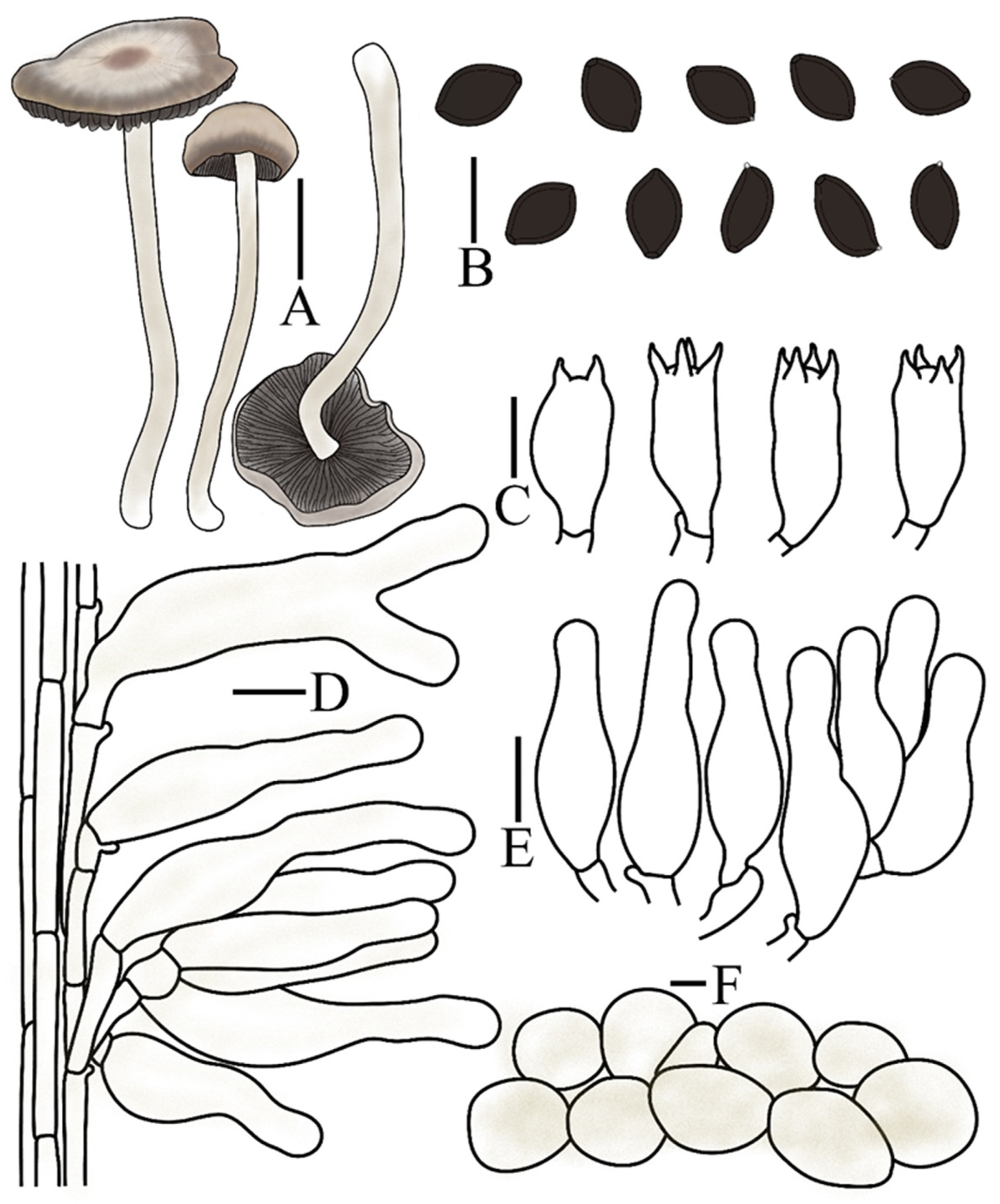

| Taxon | Voucher/Strain | Origin | ITS | nrLSU | tef1 | rpb2 | References |
|---|---|---|---|---|---|---|---|
| Bolbitius coprophilus | LE18599 | Russia | KR425526 | KR425556 | – | – | [33] |
| Conocybe muscicola | HMJAU64939 | China | OQ758113 | OQ758223 | OQ758309 | – | [34] |
| P. acuminatus | FJAU78351 | China | PX868543 | PX868580 | – | – | This study |
| P. acuminatus | CBS 270.47 | not indicated | MH856251 | MH867783 | – | – | [9] |
| P. acuminatus | KA16-1041 | Kyrgyzstan | MK351680 | – | – | – | [35] |
| P. acuminatus | X540 | Czech Republic | MW352021 | – | – | – | [35] |
| P. acuminatus | LUGO:ECC17111604 | Spain | MW376698 | – | – | – | [36] |
| P. acuminatus | SGL09 | not indicated | OR035540 | – | – | – | [35] |
| P. acuminatus | AMB 20066 | Italy | PP447481 | PP447517 | – | – | [37] |
| P. alcis | FJAU78331 | China | PX868551 | PX868579 | PX857983 | PX852305 | This study |
| P. alcis | FJAU78332 | China | PX868540 | – | – | – | This study |
| P. alcis | SAT-14-239-20 | USA(Alaska) | MW597122 | – | – | – | [35] |
| P. antillarum | FJAU78352 | China | PX868546 | PX868578 | PX857978 | – | This study |
| P. antillarum | FJAU78353 | China | PX868537 | PX868562 | PX893654 | PX852306 | This study |
| P. antillarum | UOC KAUNP MK62 | Sri Lanka | KP764810 | – | – | – | [35] |
| P. antillarum | UOC KAUNP K01 | Sri Lanka | KR867660 | – | – | – | [35] |
| P. antillarum | SFSU:DED 7874 | Thailand | MF497585 | – | – | – | [35] |
| P. atrobalteatus | jlh7468-14 | USA | PP808684 | – | – | – | – |
| P. axfordii | MFLU 19-2367 | China | NR169700 | – | – | – | [19] |
| P. bambusicola | FJAU78368 | China | PX868530 | PX868563 | PX893655 | PX852307 | This study |
| P. bambusicola | FJAU78369 | China | PX868522 | – | – | – | This study |
| P. bisporus | FJAU78362 | China | PX868547 | PX868577 | – | – | This study |
| P. bisporus | FJAU78363 | China | PX868550 | – | – | – | This study |
| P. bisporus | FJAU78365 | China | PX868532 | PX868561 | – | – | This study |
| P. bisporus | HYW197 | China | OR035518 | – | – | – | [35] |
| P. cambodginiensis | NBRC-30222 | Japan? | AB158633 | – | – | – | [35] |
| P. castaneifolius | Mushroom Observer 90428 | USA | KX010428 | – | – | – | [35] |
| P. chlorocystis | FMS iNaturalist # 193561740 | USA | PQ678531 | – | – | – | – |
| P. cinctulus | FJAU78344 | China | PX868557 | PX868570 | PX857981 | PX852303 | This study |
| P. cinctulus | FJAU78345 | China | PX868538 | – | PX893653 | PX852304 | This study |
| P. cinctulus | xsd08077 | not indicated | FJ478119 | – | – | – | [37] |
| P. cinctulus | CBS 326.34 | not indicated | MH855550 | MH867055 | – | – | [37] |
| Taxon | Voucher/Strain | Origin | ITS | nrLSU | tef1 | rpb2 | References |
| P. cinctulus | CBS 328.34 | not indicated | MH855552 | MH867057 | – | – | [37] |
| P. cinctulus | NX180911-04 | MN960188 | – | – | – | [37] | |
| P. cinctulus | MCVE 1084 | Italy | PP447482 | PP447518 | – | – | [37] |
| P. cinctulus | Gerhardt 83052 (B) | Germany | PP447483 | PP447521 | – | – | [37] |
| P. cyanescens | FJAU78366 | Timor-Leste | PX868520 | – | – | – | This study |
| P. cyanescens | taxon:181874 | not indicated | HM035084 | HM035084 | – | – | [37] |
| P. cyanescens | BP1 | India | MK855515 | – | – | – | [37] |
| P. cyanescens | FS1 | India | MK855516 | – | – | – | [37] |
| P. cyanescens | AMB 20070 | Italy | PP447484 | PP447522 | – | – | [37] |
| P. cyanescens | 0709305/JG | France | PP447485 | PP447523 | – | – | [37] |
| P. desertorum | SZMC-NL-1863 | Hungary | JX968154 | JX968271 | JX968387 | – | [8] |
| P. desertorum | LE 2865 | Uzbekistan | MH055383 | – | – | [35] | |
| P. desertorum | LE 2864 | Uzbekistan | MH055384 | – | – | – | [35] |
| P. desertorum | AH 9993 | Spain | MK397543 | MK397561 | – | – | [35] |
| P. desertorum | LE313090 | Russia | MK397566 | MK397591 | – | – | [37] |
| P. desertorum | GB-0073426 | Hungary | PP447486 | PP447524 | – | – | [37] |
| P. detriticola | PERTH 08944954 | Australia | NR199086 | MT571659 | – | – | – |
| P. dunensis | AMB 20210 | Italy | PP447489 | PP447527 | – | – | [37] |
| P. dunensis | AMB 20211 | Italy | PP447490 | – | – | – | [37] |
| P. fimicola | MCVE 4084 | Italy | JF908518 | – | – | – | [37] |
| P. fimicola | CBS 251.37 | not indicated | MH855904 | MH867411 | – | – | [37] |
| P. fimicola | Gerhardt 75349 (B) | Germany | PP447492 | PP447529 | – | – | [37] |
| P. foenisecii | FJAU78358 | China | PX868536 | – | – | – | This study |
| P. foenisecii | FJAU78359 | China | PX868542 | – | – | – | This study |
| P. foenisecii | FJAU78360 | China | PX868539 | PX868560 | PX893657 | PX852302 | This study |
| P. foenisecii | FJAU78361 | China | PX868554 | – | – | PX852298 | This study |
| P. foenisecii | 6643 | Italy | JF908520 | – | – | – | [37] |
| P. foenisecii | ubc F32593 | Canada | MG969989 | – | – | – | [37] |
| P. foenisecii | CBS 142.40 | not indicated | MH856067 | MH867557 | – | – | [37] |
| P. foenisecii | AMB 20071 | Italy | PP447493 | PP447530 | – | – | [37] |
| P. foenisecii | AMB 20072 | Italy | PP447494 | PP447531 | – | – | [37] |
| P. fraxinophilus | FJAU78346 | China | PX868544 | PX868575 | PX857980 | PX852308 | This study |
| P. fraxinophilus | FJAU78347 | China | PX868529 | PX868574 | PX857982 | PX852310 | This study |
| P. fraxinophilus | MushroomObserver.org/455364 | USA | OL629088 | – | – | – | [35] |
| Taxon | Voucher/Strain | Origin | ITS | nrLSU | tef1 | rpb2 | References |
| P. fraxinophilus | OMDL iNat # 170758482 | USA | OR987324 | – | – | – | [35] |
| P. latifolius | FJAU78340 | China | PX868523 | – | – | – | This study |
| P. latifolius | FJAU78341 | China | PX868533 | PX868565 | – | – | This study |
| P. mediterraneus | AMB 20074 | Italy | PP447496 | – | – | [37] | |
| P. mediterraneus | AMB 20075 | Italy | PP447497 | PP447533 | – | – | [37] |
| P. mexicanus | ANGE1557 | Dominican Republic | MZ856314 | – | OK546186 | [38] | |
| P. olivaceus | MushroomObserver.org/158389 | USA | MF629829 | – | – | – | [35] |
| P. olivaceus | MushroomObserver.org/89608 | USA | MH285992 | – | – | – | [35] |
| P. olivaceus | iNAT:100066497 | USA | ON314881 | – | – | – | [37] |
| P. olivaceus | AMB 20076 | Italy | PP447498 | PP447534 | – | – | [37] |
| P. ovinus | FJAU78335 | China | PX868559 | – | – | – | This study |
| P. ovinus | FJAU78336 | China | PX868534 | PX868566 | PX857973 | PX852299 | This study |
| P. ovinus | FJAU78337 | China | PX868535 | PX868567 | PX857974 | PX852297 | This study |
| P. pantropicalis | JBSD 130972 | Dominican Republic | PP590037 | – | – | – | [35] |
| P. pantropicalis | PERTH 09605894 | Australia | PP590039 | – | – | – | [35] |
| P. papilionaceus | FJAU78333 | China | PX868549 | PX868573 | PX857972 | PX852300 | This study |
| P. papilionaceus | FJAU78334 | China | PX868525 | – | – | – | This study |
| P. papilionaceus | AFTOL-ID 1499 | not indicated | DQ182503 | DQ470817 | – | – | [37] |
| P. papilionaceus | CBS 582.79 | not indicated | HM035081 | HM035081 | – | – | [37] |
| P. parvisporus | FJAU78330 | China | PX868556 | – | – | – | This study |
| P. parvisporus | MushroomObserver.org/312079 | USA | MH101639 | – | – | – | [35] |
| P. plantaginiformis | LE 2867 | Uzbekistan | MK397575 | MK397597 | – | – | [9] |
| P. plantaginiformis | LE 2870 | Uzbekistan | MK397576 | MK397598 | – | – | [9] |
| P. plantaginiformis | LE 2862 | Russia | MK397577 | MK397599 | – | – | [9] |
| P. plantaginiformis | TAAM120547 | Uzbekistan | PP447502 | PP447537 | – | – | [37] |
| P. plantaginiformis | TAAM120647 | Uzbekistan | PP447503 | – | – | – | [37] |
| P. praecox | FJAU78356 | China | PX868528 | PX868568 | PX857975 | PX852309 | This study |
| P. praecox | FJAU78357 | China | PX868558 | PX868569 | PX857976 | – | This study |
| P. punjabensis | LAH36793 | Pakistan | MZ265143 | ON116490 | – | – | [39] |
| P. punjabensis | LAH36794 | Pakistan | MZ823627 | ON116492 | – | – | [39] |
| Taxon | Voucher/Strain | Origin | ITS | nrLSU | tef1 | rpb2 | References |
| P. retirugis | CBS 274.47 | France | MH856255 | MH867787 | – | – | [37] |
| P. rhombispermus | FJAU78367 | China | PX868531 | PX868564 | PX926009 | PX926010 | This study |
| P. rhombispermus | CWN 11502 | China | MZ782082 | MZ781504 | – | – | [40] |
| P. rickenii | FJAU78348 | China | PX868548 | PX868572 | PX857979 | PX852296 | This study |
| P. rickenii | FJAU78349 | China | PX868541 | PX868571 | – | – | This study |
| P. rickenii | HMAS 290093 | China | MK966648 | – | – | – | – |
| P. rickenii | HMAS 290109 | China | MK966649 | – | – | – | – |
| P. semiovatus | FJAU78354 | China | PX868526 | – | – | – | This study |
| P. semiovatus | FJAU78355 | China | PX868524 | – | – | – | This study |
| P. semiovatus | MCVE 21188 | Italy | JF908515 | – | – | – | [37] |
| P. semiovatus | CBS 276.39 | not indicated | MH856012 | – | – | – | [35] |
| P. semiovatus | Mushroom6 | China | MT451924 | – | – | – | [35] |
| P. semiovatus | AMB 20084 | Italy | PP447509 | PP447542 | – | – | [37] |
| P. semiovatus | AMB 20082 | Italy | PP447511 | PP447541 | – | – | [37] |
| Panaeolus sp. | FJAU78338 | China | PX868555 | – | PX893656 | – | This study |
| Panaeolus sp. | FJAU78339 | China | PX868545 | – | – | – | This study |
| Panaeolus sp. | FJAU78350 | China | PX868521 | – | – | – | This study |
| Panaeolus sp. | FJAU78364 | China | PX868553 | – | – | – | This study |
| P. subbalteatus | FJAU78342 | China | PX868552 | PX868576 | PX857977 | PX852301 | This study |
| P. subbalteatus | FJAU78343 | China | PX868527 | – | – | – | This study |
| P. subbalteatus | CBS 327.34 | USA | MH855551 | MH867056 | – | – | [37] |
| P. subbalteatus | HFJAU-ND146 | China | MN622762 | – | – | – | [35] |
| P. sylvaticus | ANGE1393 | Dominican Republic | OQ311002 | – | – | – | [35] |
| P. tropicalis | not indicated | China | JF961377 | – | – | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



