
Submitted:
28 January 2026
Posted:
05 February 2026
You are already at the latest version
Abstract
Introduction: Solid-state fermentation with culinary fungi (mushrooms commonly used in cooking) could be a scalable means of producing nutrient-dense protein foods, while improving digestibility and bioavailability and valorizing agricultural commodities and byproducts. Advancing the science of these processes has major implications for both nutrition security and resilient, sustainable food systems. This review synthesizes how substrate selection, fungal strain choice, and controllable growth conditions can transform underutilized or under-consumed plant materials into minimally processed, consumer-acceptable foods through enhanced sensory characteristics and nutritional profiles. Methods: We conducted a narrative review to synthesize peer-reviewed and applied literature to map substrate–strain–process combinations that could enhance nutritional composition, sensory performance, food safety, scalability, and sustainable food-system outcomes. Results: Across cereals, legumes, and oilseed meals, solid-state fermentation consistently increases protein concentration and quality, improves nutrient bioavailability, decreases anti-nutritional factors, and generates flavor-active metabolites while enhancing texture. Outcomes can be further optimized through manipulation of growth conditions such as moisture, temperature, particle size, aeration, light, residence time, and post-process thermal finishing. Discussion: Integrating culinary fungi with commodity crops enables circular use of side streams, shortens protein production cycles relative to animal sources, and can support rural economies while aligning with scientific dietary guidance. Factorial process studies linking growth stage inputs to sensory acceptance, standardized safety and regulatory frameworks and techno-economic analyses that quantify cost-per-nutrient and edible output per hectare would help extend this knowledge for maximal impact. Conclusions: Solid-state fermentation should be considered as an important strategy for improving diet quality and food security.
Keywords:
solid-state fermentation
; mushrooms
; mycelium
; food security
; nutrient-density
; protein bioavailability
; agricultural valorization
; sensory and flavor development
1. Introduction
Solid-state fermentation (SSF) is a bioprocess in which microorganisms, typically fungi, colonize solid substrates. SSF has emerged as a practical approach to transform edible base materials into nutrient-dense foods. Compared to other fermentation techniques, such as submerged liquid-state fermentation (LSF), SSF has received less academic and industry attention [1,2]. SSF and LSF are different but complementary platforms. LSF produces a more homogeneous, tightly controlled, concentrated mycelial biomass grown in liquid media, whereas SSF is a low-water transformation of a solid substrate into a composite, minimally processed food that includes the mycelium itself. Recent research highlights the capacity of SSF to improve protein quality by concentrating essential amino acids and enhancing digestibility of otherwise recalcitrant biomass [3], while generating savory flavor precursors under growth conditions that require relatively low water and energy inputs [1,2,4−7]. Importantly, these benefits extend beyond fungal protein concentration alone.
Many alternative protein production systems depend on extraction and intensive processing. By contrast, fungal mycelia grown on edible substrates enable integrated, minimally processed composites that preserve the nutritional and sensory properties of whole food (e.g., whole grains, legumes, and seeds). In addition, the use of commodity crops and their side streams as SSF substrates provides an avenue for additional valorization, resulting in significant nutritional, sensory, and economic opportunities for SSF production [5,8]. SSF could be an important lever to support alignment of dietary patterns with public health guidance, increase the consumption of whole and minimally-processed foods, reduce on-farm and food waste, add value to commodity crops, and increase access to protein-rich, plant foods.[5] Traditional exemplars of SSF fermentation such as tempeh (e.g. Rhizopus SSF of soybeans) and oncom (e.g. Neurospora or Rhizopus SSF of peanut or soy oil press cakes) have converted agricultural products and side streams into high-protein foods with greater digestibility for hundreds of years.[3] However, these foods are primarily eaten in East Asia and adoption elsewhere is low, in part due to hedonic rejection. In contrast, modern food manufacturers have recently introduced novel mycelium-forward products (e.g., Quorn™, Meati™, Prime Roots™) that have gained traction in Western markets. Recent articles and reviews largely frame SSF as a tool to produce meat analogues, or further examine methods of enhancing traditional tempeh or oncom[3,9,10] reflecting that mycelium product category visibility has been driven largely by submerged-fermentation mycelial biomass products, while SSF is gaining traction as a complementary platform for solid-matrix biotransformation.[11] Consequently, relatively few analyses consider how SSF can serve as a standalone method for producing sensory-optimized, minimally processed, nutrient-dense foods on inherently nutritious edible substrates. Even fewer examine how coordinated choices across substrate, strain, and growth-condition vectors could enable broader global adoption than traditional SSF foods. Such synthesis of information on combinations can be applied to expand product development and also open up additional market opportunities for crops and edible sidestreams [5,12].
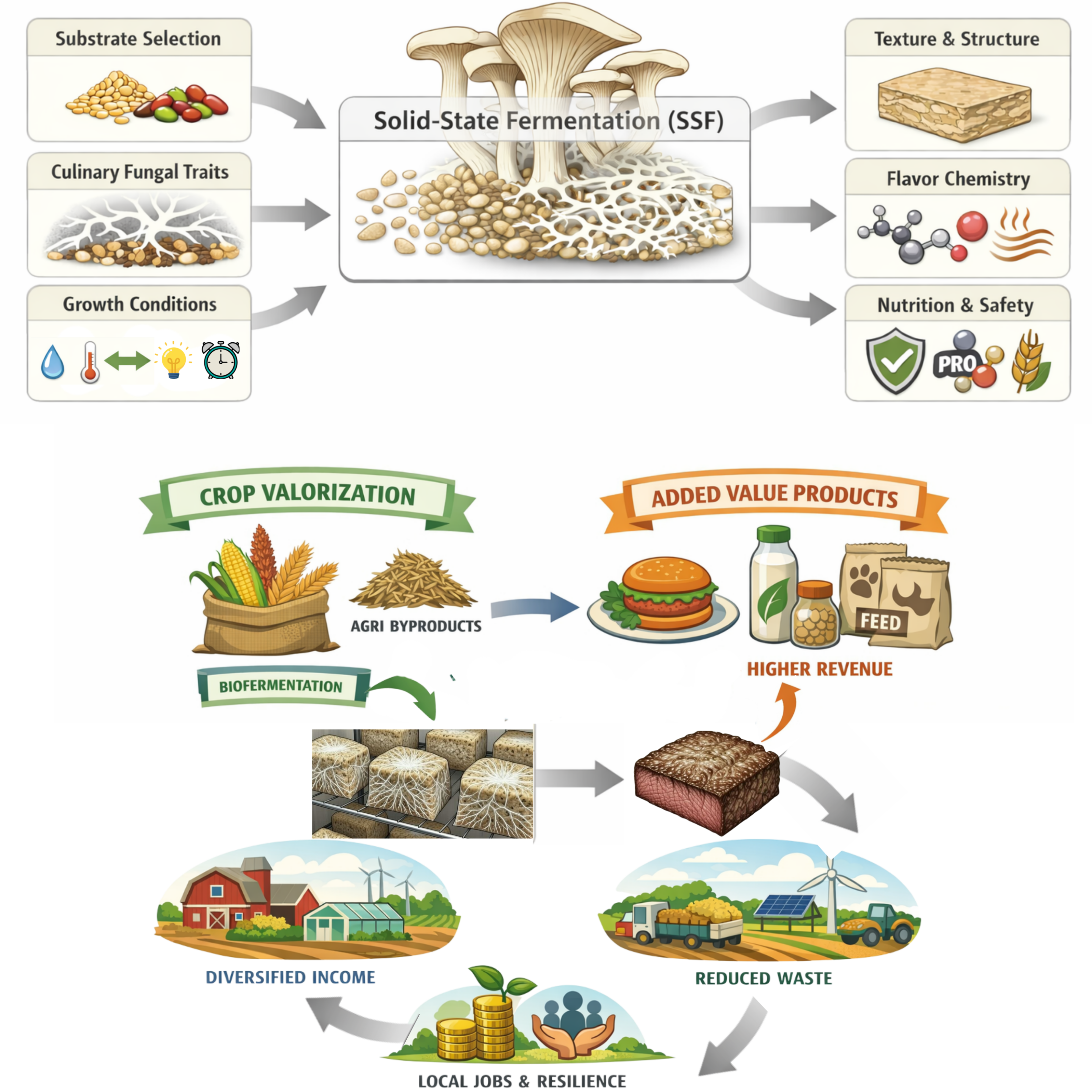
Despite nutritional and sustainability advantages, SSF remains less commercially utilized relative to LSF. This is due to various tunable factors such as heat/mass-transfer constraints, limited in-line monitoring, and variable substrate features that are standardized across processes used in LSF.[1,13] This review synthesizes practical solutions to mitigate the challenges of SSF and outlines a targeted research agenda to accelerate reliable, food-grade deployment, while summarizing the known nutritional and sensory results of SSF connected to different inputs and growth conditions. The information that follows can help outline key advancement steps for stakeholders—from scientists, product developers, regulators, to agricultural producers and others—working to create more nutrient dense, minimally processed foods. The following sections outline optimization pathways, with attention to edible substrate combinations and the use of culinary mushroom strains (mushrooms and edible fungi valued for their taste, texture, and use in cooking) to enhance final food matrices. This review complements recent SSF literature focused on “meat analogues,”[5] incorporating broader considerations of substrate composition, food-system impacts, commercial progress, and safety and regulatory frameworks. We synthesize quantitative nutrition effects—including protein concentration, accrual, and reductions in anti-nutrients—and relate these biochemical changes to nutrient availability and sensory attributes. Additional technoeconomic hotspots, such as moisture, particle size, aeration, and temperature influences on growth time and yield are identified as key parameters for improving unit cost and consumer willingness to try and buy. Finally, we situate SSF within global nutrition-security and circular-bioeconomy agendas, highlighting its capacity to upgrade crop value, diversify farm income, and provide minimally processed, nutrient-dense foods to broad populations. (Figure 1).
Advantages for Using Commonly Consumed Culinary Fungi for SSF
Most commercially available LSF and SSF use fungal strains (e.g. Fusarium. venenatum, and Rhizopus oligosporus) with no direct culinary applications beyond their incorporation into mycelium-based foods. In other words, these fungi are not commonly grown to produce mushroom fruiting bodies for human foods. However, mushroom fruiting bodies are widely cultivated for culinary use, lauded for their firm, meaty texture and savory flavors, making them highly coveted ingredients and even substitutes for meat [14]. For edible substrate SSF, the choice of fungal species can be altered in order to intentionally alter the taste and texture of the final output, and capitalizing on the sensory advantages of popular culinary mushrooms is an emerging area of advancement. The most relevant fungal traits for SSF include: (i) colonization kinetics and growth robustness on food-grade substrates; (ii) extracellular enzymes (e.g., amylases, cellulases/hemicellulases, pectinases, proteases, phytases) able to metabolize a range of plant materials; (iii) mycelial packing density and hyphal morphology (i.e., rhizomorph tendency, branching) and (iv) resultant production of sensory relevant metabolites (e.g., nucleotides, free glutamate/peptides, lipids, and volatiles) [15−20]. Culinary fungi substantially vary, and species selection should complement the growth substrate for optimal texture, flavor and nutritional outcomes. Below we highlight popular strains and their known benefits and drawbacks for SSF.
Pleurotus spp. (oyster mushrooms: Pleurotus. eryngii, Pleurotus. ostreatus, Pleurotus pulmonarius)
Among culinary mushrooms, Pleurotus ssp. consistently demonstrate strong performance on edible substrates, as shown by: the ability to rapidly colonize on hydrated grains, legumes and fibrous co-substrates; secrete robust carbohydrases and proteases which metabolize otherwise indigestible fibers, agricultural food, and lignocellulosic wastes while liberating amino acids for enhanced protein digestibility [21]; form dense, cohesive mycelial networks that can bind particles into sliceable slabs for meat analogues or be milled into flour to increase moisture retention of resultant foods [17,22,23].
SSF studies with Pleurotus ssp. on canola and cottonseed meals, or cereal–legume bases, report increased protein, reduction in anti-nutrient factors (e.g., phytate, sinapine, gossypol, glucosinolates, purines), and gains in antioxidant and polyphenol content [24−33]. This is because as Pleurotus ssp. mycelium develops, secreted enzymes break down cellulose and other structural components providing nutrients for fungal growth which improves end stage digestibility of the fermented substrate for humans [34] Together, rapid colonization, broad extracellular enzyme portfolios, and dense, cohesive mycelial networks, make Pleurotus ssp. a pragmatic genus for commercial, food-grade SSF, and one capable of converting agricultural side streams and whole crop substrates into nutrient-dense, sensory-forward, value-added foods.
Among individual species, Pleurotus eryngii (i.e. king oyster mushroom, or King Trumpet Mushroom) may be the strongest species within the genus, owing to robust rhizomorphic growth, dense mycelial mats, and tolerance for modest lipid inclusion [15,21,35] Studies of P. eryngii have demonstrated its ability to valorize low-acceptance, high yield, and resilient crops by increasing consumer acceptance of the resulting foods [24,36,37]. Solid-state fermentation of canola meal with P. eryngii for 12 days resulted in an 11-18% increase in protein content and improved in vitro digestibility, enhancing the overall nutritional profile.12 Among edible basidiomycetes, P. eryngii is especially attractive when the whole mycelium-base integrated platform is the desired consumable, owing to the production of a flavorful and dense mycelium that resembles steak and other whole meat cuts.5 Lastly, incorporating P. eryngii with Rhizopus or Aspergillus as a co-culture is a logical next step to widen substrate utilization and tailor for specific flavor/texture outcomes, as notably both Aspergillus and Rhizopus include oleaginous species that can enhance flavor profiles through lipid accumulation and enzymatic generation of flavor-active compounds.[38,39]
Lentinula edodes (shiitake)
Lentinula fruiting bodies are known for producing elevated levels of guanylic acid (i.e. Guanosine 5'-monophosphate, or 5′-GMP), free glutamate, and sulfur-derived volatiles (e.g., lanthionine) that drive savory flavor. Additionally, Lentinula mushroom is known for its cap with a firm, meat-like texture, and distinctive aroma profile. However, Lentinula requires lengthy colonizing time to achieve fruiting.[40,41] Lentinula mycelium can produce cohesive networks but is less rhizomorphic than P. eryngii, and the resulting mats are less dense, making it more suited to minces or used as flavorful additive where Lentinula may be valuable as a flavor driver contributing umami-active nucleotides rather than standalone foods or whole cut meat analogues. However, there is great opportunity to explore hybrid co-cultures (such as with rapidly colonizing Rhizopus) to add a favorable aroma while accelerating colonization and forming denser mycelial mats.[15]
Agaricus bisporus (button/portobello)
Agaricus is a ubiquitous culinary mushroom, valued for its plentiful growth and mild flavor. However, its mycelium typically forms a delicate network that lacks the packing density required to generate cohesive, whole-cut structures. Although Agaricus has a balanced essential amino acid profile, it generally has lower concentrations of flavor-active compounds compared to other culinary fungi making it less appealing for some meat analogues.[40,41] However, use of the fruiting body of Agaricus can be a reliable co-culture partner with structure and flavor supplied by Pleurotus or another companion fungi.
Hericium erinaceus (lion’s mane)
Hericium has gained increased popularity in culinary applications, largely due to its fibrous, fruiting bodies which have been explored in the production of seafood alternatives, especially shellfish such as crab and shrimp.[42] Its mycelium, which is typically cottony and slower to colonize, often fails to form dense substrate mats, but still contributes distinctive flavor and texture attributes from pinning through fruiting. Fruiting bodies have multiple applications from shredded or “pulled” meat applications, owing to lighter and fibrous textures, which make Hericium fruit valuable not only for seafood applications but also as a broader flavor adjunct.[15,40,41] In addition to culinary use, Hericium has attracted attention for its bioactive compounds and potential applications in functional foods and nutraceuticals, which could be leveraged to increase interest for SSF foods.[43−45].
Other Culinary Fungi—Co-Culture Candidates
Additional culinary basidiomycetes offer potential for SSF applications, though their growth characteristics and technoeconomic profiles differ from the more aggressive colonizers described above.[46] Flammulina velutipes (enoki) and Hypsizygus marmoreus (beech) are widely consumed mushrooms with mild, clean flavors and light coloration which can be advantageous for inclusion in neutral-tasting, light-colored matrices. Additionally, both species produce fine, sparsely rhizomorphic hyphae and slower substrate colonization than others Pleurotus ssp.[15,40,41] However, Flammulina can contribute subtle umami notes, polysaccharide enrichment, and textural softening,[47] while the mild aromatic compounds of Hypsizygus could be used to modulate flavor without overpowering, each offering attractive traits that could be well incorporated through innovative co-culturing.
Auricularia sp. (wood ear) are a common culinary mushroom with a distinct texture. This fungi produces gelatinous, polysaccharide-rich matrices with high levels of soluble fiber and β-glucans, offering unique opportunities for moisture retention, chewiness, and functional fiber enrichment in SSF composites.[48−50] Studies of Auricularia spp. describe moderate mycelial growth rates that are highly dependent on substrate composition and culture conditions, with colony expansion and mycelial density optimized within a narrow carbon–nitrogen range.[51,52] Additionally, Auricularia ssp. provides moderate mat cohesion[52] which makes Auricularia is most beneficial for SSF in co-culture systems where its mucilaginous texture can be complemented by a structurally robust companion strain.[51] However, its mild, earthy flavor and pigmentation can also be leveraged for specific sensory outcomes.
Graph 1. and β-1,6-glucans identified as major structural polysaccharides in its mycelium.[53] Grifola polysaccharides form branched glucan networks, yielding mycelium that is cohesive but not densely rhizomorphic. As a result, Grifola typically exhibits only moderate structural integrity and slower substrate colonization than more aggressive genera such as Pleurotus. However, its robust metabolic activity produces many functional bioactive metabolites which help give maitake its well-known favorable flavor profile, making it particularly attractive for both premium flavor development and nutritional enhancement.[54] However, because its mycelial architecture lacks the dense binding needed for strong matrix formation, Grifola may perform best as a co-culture, where its sensory and nutritional contributions can be leveraged without relying on it for rapid substrate conquest.
Nutritional Profile of Mycelium-Rich Foods and SSF Benefits
Edible mushroom mycelium is a nutritionally rich food source often compared to animal proteins,[5,11] with both mycelium and fruiting bodies high in protein (20–30% of dry weight, and some as high as 40%, depending on growth conditions) [34] with a balanced essential amino acid profile[56]. For example, the mycoprotein produced by Fusarium venenatum (e.g. Quorn™ foods) has a protein digestibility-corrected amino acid score (PDCAAS) of ~0.99644 , which can be comparable to chicken and beef, but is much higher than soybeans.[57−60] Many fungi are also reportedly “complete” proteins (having a balance of essential amino acids) but also have high levels of functional amino acids associated with flavor and animal-sourced foods (e.g. glutamate, GABA).[34] Alongside protein enrichment from the fungal biomass accretion itself, reproducible nutritional benefits of SSF include (i) liberation of bound polyphenols and consequent gains in antioxidant capacity [61−63]; (ii) reductions in anti-nutritional factors, especially phytic acid in cereals or legumes and glucosinolates and sinapine in Brassica meals which increases nutrient availability [64]; (iii) formation of flavor-active molecules (free amino acids, short peptides, and 5′-nucleotides) that increase umami perception; (iv) increases in dietary fiber (especially polysaccharides like β-glucans found in fungal cell walls, which have been associated with cholesterol-lowering and immune-modulating effects [11,34,65,66]; and (v) increases in essential nutrients such as B-vitamins, ergosterol (provitamin D2) [67,68], zinc, potassium, copper, and selenium. In contrast to other sources of these nutrients (plant and animal), the lipid content, and consequently the relative energy density, of mycelium is low, resulting in health-promoting, nutrient dense foods [34,57].
Two complementary mechanisms drive nutritional gains in SSF (Table 1). First, the fungus contributes its own mycelial biomass, producing high quality protein, fiber (typically 20–35% dry basis in mushroom mycelium), β-glucans and chitin[69−71], B-vitamins (notably riboflavin, folate, and when co-cultured or fortified vitamin B12[5]), and ergosterol (provitamin D2) which can be further modulated by UV exposure to stimulate greater endogenous production.[68] As mycelium preferentially metabolize carbohydrates to support growth, the relative protein content of the substrate increases, and concurrent mycelial activity releases peptides and amino acids that contribute to improved protein digestibility.[5,12,34,72,73]
The second mechanism is driven by secreted enzymes that biotransform the substrate. Many plant foods (e.g., legumes, wheat bran, spinach) contain “anti-nutrients” such as phytic acid, and tannins (Table 2) which bind essential minerals (i.e., iron, calcium) thus reducing their bioavailability.[74] However, the enzymatic activity of fungi can degrade these molecules, leading to improved nutrient absorption.[5,75−78] For example, fermentation of rapeseed (canola) meal by Pleurotus mycelium degrades sinapine and phytates, while concentrating protein[31,79], and was similar for Pleurotus fermentation of mixed grain or legumes.[5,25,80] Carbohydrases, proteases, and phytases improve protein digestibility by hydrolyzing macromolecules to release bound phenols, flavonoids, and peptides, and by degrading anti-nutritional factors including phytic acid (an iron- and zinc-chelating inhibitor of protein digestibility), sinapine and glucosinolates (bitter/pungent compounds), and gossypol (a toxic cottonseed phenolic).[28,29,70,71,81−83] Additionally, many industrial mycoprotein processes include an RNA reduction heat step to neutralize purines.[84,85] Overall, this leads to increased nutrient availability from common plant food bases and the mycelium itself.[86] Reactions can be tuned through substrate pretreatments (hydration, gelatinization, altered particle size), moisture (60–65%), temperature (24–30°C for Pleurotus), and aeration, which modulate colonization kinetics by influencing heat removal, gas exchange, water activity, permeability, hyphal ingress, and enzyme secretion.[39,87−93]
Substrate composition can also impact product nutrition (Table 1 & 2). In SSF of brassica meals (e.g., canola, rapeseed), Pleurotus ostreatus increased protein content (11–18% dry weight), but reduced sinapine (99.8%), glucosinolates (~98.8%), and phytate content (~56–76)[24]. In further studies using cottonseed press cake blended with Lathyrus (i.e., sweet peas) pericarp (80%/20%), P. ostreatus reported increased crude protein content (35%), and 1,3/1,6-β-glucans were five times greater, but total gossypol was decreased (89%). These impacts addressed a key safety and sensory constraint of cottonseed substrates, where the presence of gossypol prevents safe consumption for humans and palatability; mycelial fermentation is a promising alternative to genetic modification of cottonseed, the only other viable approved path for human consumption.[25,94] Comparable outcomes have been observed for P. ostreatus on soybeans, resulting in elevated protein without isolation and increased antioxidant levels.[38]
In cereals (e.g. oats, barley) fermented with Rhizopus oligosporus like tempeh, phytate content dropped by 74% and 89%, respectively. In a human crossover study, iron (non-heme iron) absorption from fermented whole-grain barley reached 5.5% versus 3.0% for boiled barley, demonstrating a bioavailability benefit consistent with phytate reduction.[37,95] Co-fermenting grass pea with flaxseed oil-cake strategically re-profiles lipids: α-linolenic acid (ALA, ω-3) rose 3.6–10× (dose-dependent) and the ω6:ω3 ratio improved from 11:1 to 0.5–2.5:1, with modest increases in sulfur amino acids—an approach that leverages lipid-rich inputs while the fungus primarily utilizes carbohydrate.[96]
SSF can also reduce allergenicity of certain foods (Table 1). Fermentation breaks down large proteins into smaller peptides and amino acids, degrading many of the IgE-binding epitopes of foods like soy, lupins, wheat, and peanuts.[97,98] For example, fermenting peanut flour with Rhizopus for 1–2 days was shown to significantly degrade peanut allergen proteins.[99,100]
SSF thus offers a pragmatic route to valorize crops and edible side streams while enhancing nutritional value.[64] Below, we add additional insight on sensory enhancement and circular-economy value creation, to complete the picture of how SSF with edible fungi addresses complementary priorities for nutrition security and sustainable food systems.
Sensory and Consumer Experience Advantages for SSF
Mushroom aroma is often attributed to C8 compounds such as 1-octen-3-ol and 1-octen-3-one, yet the volatile profile of mushrooms is far more complex, arising from a broader ensemble of lipid-derived alcohols, aldehydes, ketones, and sulfur chemistry.[101] This complexity can be leveraged in conjunction with SSF to build a novel sensory experience. As fungi colonize a substrate, their metabolism will reflect the consumption and modification of substrate derived nutrients creating, diminishing, or amplifying key sensory attributes. In addition, secreted fungal enzymes can further modify substrate chemistry, generating new volatile and non-volatile compounds that contribute to aroma, taste, and texture. These processes can be influenced by moisture levels and particle sizes which support oxygen diffusion and consistent colonization, as well as controlled post-fermentation through thermal processing to amplify Maillard/Strecker pathways.[102−104]
Flavor development under SSF is strongly influenced by proteolysis, ribonucleotide metabolism, and substrate-specific biochemical pathways. These processes can increase the abundance of taste-active compounds such as L-glutamate, guanosine 5′-monophosphate, and inosine 5′-monophosphate, the synergistic drivers of umami associated with culinary mushrooms and traditional SSF applications like tempeh.[15,40,41,105,106] Furthermore, studies specifically using Pleurotus spp. have also demonstrated enhanced umami intensity through enzymatic activity and nutrient turnover.[107] Additionally, SSF can also influence the development of thermally driven aromatics by building the precursor pool of reducing sugars, free amino acids, and peptides that participate in Maillard and Strecker chemistry during subsequent cooking. As a result, thermal finishing (e.g., baking, roasting, pan-searing) of fermented substrates often yields elevated levels of Strecker aldehydes (e.g., methional, phenylacetaldehyde), alkylpyrazines (e.g., 2-methyl-, 2,5-dimethylpyrazine), and heterocycles such as 2-acetylpyrrole, contributing roasted and nutty aromas.[103,104,108,109] Studies evaluating SSF-treated vs. mushroom-enriched baked foods reported increased intensities of nutty, roasted, and mushroom-like aroma, along with umami and sweet taste modifications. Furthermore, consumer evaluations showed that acceptability of fermented products was modulated by moisture content and fiber level.[103,108,110]
SSF can also remove anti-nutritive and/or odor-active precursors (Table 1 &2). Across grains, legumes, and oilseed press-cakes, SSF consistently reduces bitterness and astringency through the degradation of phytate, tannins, and sinapine.[74,111−113] In Brassica meals, observed reductions in sinapine and glucosinolates by Pleurotus effectively lower the pungent mustard-like notes and bitterness.[23,79] Fermentations using Rhizopus with or without lactic acid bacteria amendments can be used to suppress beany and grassy aromas while imparting mild acidity. Complementing the reduction in off-notes, SSF fungi can also increase umami flavor and roasted aromas through accrual of amino acids and reactant sugars, both Maillard chemistry precursors.
However, SSF does carry risks of negative sensory expression which must be controlled during growth. Under nutrient imbalance or suboptimal moisture conditions, fungi may produce elevated levels of volatile sulfur compounds, geosmin-like earthy notes, or lipid-derived aldehydes associated with rancidity.[102,114−117] Overly aggressive enzymatic activity may also weaken substrate structure, resulting in soft, crumbly, or overly fibrous textures that can detract from product quality.[2,108] SSF can be strategically tuned to enhance desirable flavors, suppress off-notes, and selectively reshape challenging substrates.
In addition to modulating aroma and taste through metabolic and biochemical pathways, SSF also alters the physical structure of the substrate. These structural effects play a central role in defining the texture of the final product. With continued growth, enzymatic hydrolysis of structural carbohydrates and the expansion of hyphal networks jointly modify substrate mechanics which impact sensory enjoyment and culinary functionality. Two primary mechanisms dominate texture formation during SSF. The first mechanism is the partial enzymatic hydrolysis of abundant structural polysaccharides within the substrate (e.g., hemicellulose and pectin). As these polysaccharides are broken down, the matrix becomes more hydrated and pliable, leading to progressive softening of the substrate. Increased hydration can improve particle binding and cohesion, resulting in a better “bite” sensation, and in more meat-like applications can contribute to greater tenderness and juiciness.[5,118,119] The second mechanism is mycelial structuring, in which the growing network of intertwined hyphae contributes directly to texture. As fungal growth progresses, given adequate permeability and interstitial space, the substrate is gradually overtaken and transitions from a loose or granular mixture (e.g. grains and legumes) into a biologically consolidated material. Hyphae extend, branch, and fuse, increasing their contact with substrate particles and establishing a cross-linked, fibrous scaffold throughout the substrate. The expanding network fills voids, wraps around substrate particles, and interlocks with neighboring hyphae, gradually forming the mycelial architecture that underpins matrix cohesion. Additionally, hyphal walls contribute to rigidity and elasticity, while the interconnected cords distribute mechanical stress and enhance adhesion. Collectively, these processes alter how the material fractures, compresses, and retains moisture, which creates a product with firmer, integrated, and more enjoyable textures. [15,17,23] When mature and dense, this hyphal network can be modulated to create a “whole-cut” mycelium steak or meat-like texture (e.g., Pleurotus slabs), where the grown matrix itself provides an anisotropic, meat-like bite. This is further complementation during fermentation by the liberation of γ-glutamyl peptides and glutathione which signal the sensation kokumi (“mouthfulness”), enhancing the perception of richness alongside umami.[120−123] Thus, SSF can help make plant-rich staples more palatable while simultaneously increasing nutritive value via concentration and increased bioavailability.
For consumer-experience, framing seems to greatly influence consumer acceptance of fermentation biomass foods. Notably, contextual framing, especially anchored in the familiar, or that employes heightened emotional and sometimes sensory primes seems to be most influential.[124] U.S. consumers appear more receptive to novel-protein foods when they are framed around benefits (e.g., “clean,” “sustainable,” “high-quality protein”) than when labeled with process-focused terms such as “fungal” or “fermented,” which can trigger perceived “off” flavors, neophobia, or processing aversion.[6,125−127] By contrast, when these foods are positioned as flavorful, or protein-enhanced versions of familiar foods (e.g. high-protein noodles, breads, burger patties), adoption improves, including increased acceptance of tempeh in markets with less historical demand.[128] Allergen- and label-sensitive consumers often value short ingredient lists which is another advantage for SSF foods, as seen with successful market leaders like Quorn.[55] In Western samples, tempeh often faces adoption barriers tied to cultural unfamiliarity and a fermented flavor profile with less enculturation, despite the strong nutritional profile. [110,124,129] A synthesis of Western consumer studies finds higher acceptance for alternatives that mimic conventional meat in texture and appearance, while tofu and tempeh tend to be perceived as niche or transitional products.[108,110,129−132] Strategic benefit-forward framing—such as culinary versatility, environmental impact, and protein quality and digestibility—combined with sensory optimization (umami/kokumi chemistry, roasted volatiles) and familiar formats (patties, cutlets, sliced “whole-cut” mycelium) improves stated liking and purchase intent.[6,8,133]. [103,104] Collectively, SSF foods, flours, and powders can be used to create minimally processed, clean-label platforms to boost protein, umami, and bioactives across familiar foods, aiding adoption. [15,40,86,118,119,134,135,136]
As a Strategy to Improve Diet Quality and Nutrition Security
As discussed above, across cereals, legumes, and oilseed meals, filamentous fungi reduce anti-nutritional factors (e.g., phytate, sinapine, glucosinolates) and increase protein quality, digestibility, and micronutrient accessibility.[3,9,24,37,113,137,138] Traditional examples such as tempeh and oncom illustrate the mechanism: fungal proteases and peptidases liberate amino acids, small peptides, and partially hydrolyze structural polysaccharides, which reduces structural rigidity and increases matrix deformability, improving digestibility.[10,63,104,109,121,137,139−141] As a result, many SSF foods resemble meat when cooked, similar to widely consumed culinary mushrooms, while offering additional nutritional and sensory benefits derived from their fibrous texture and strong umami attributes.[3,104,107,140] SSF therefore can provide an abundant source of minimally processed, protein-dense, fiber-rich, low-saturated-fat foods that meet public-health guidance without sacrificing eating pleasure.
For nutrition-security, SSF is attractive because it upgrades abundant, affordable substrates with a range of nutrient densities into foods that people choose for taste as well as health and more niche priorities like reducing food waste. Traditional foods such as tempeh (Rhizopus-fermented soybeans) and oncom (Neurospora- or Rhizopus-fermented press cakes) have demonstrated for centuries that SSF can reliably turn crops and side streams into foods with long shelf lives, extending their utility through global trade and minimizing food waste. Compared to conversion of imperfect, surplus or side stream materials into other products, SSF can streamline operations [1,2,97,142] and support immediate food applications with production cycles ranging from 36–48 hours for Rhizopus to 10–21 days for Pleurotus. This means that production can be matched to demand, sited close to crop processors, and leveraged for rapid acceleration in production of quality protein sources. In contrast, common extant animal sources—fed the same underlying crops—require 2-24 months to reach market weight. [97,139] Plant protein sources themselves take 90-120 days to harvest, but SSF can be grown on extant or rejected crops and side streams, which are typically low in protein quality pre-fermentation. Instead, SSF upgrades these existing plant materials into higher-quality protein foods on a days to weeks cadence. This is notable because many plant proteins, even high quality sources such as soy (PDCAAS=0.92), remain limited in essential amino acid balance and digestibility, whereas SSF can raise PDCAAS values >0.96 through amino acid liberation, anti-nutrient reduction, and improved digestibility. The practical implication is that SSF enables rapid, demand responsive production of nutrient-dense protein foods without waiting for new crop or animal cycles, strengthening resilience during routine operations and supply disruptions.[97,139]
SSF also adds edible output per hectare and lowers delivered cost-per-nutrient. Unlike submerged systems, SSF uses the edible substrate as both growth matrix and food, avoiding expensive clarification and separation steps and large steam-in-place bioreactors. SSF tray and rack or tunnel systems using low-tech humidifiers and gentle aeration are sufficient (such as those for on-farm mushroom cultivation), and thus enable co-location with producers and processors for efficient side-stream valorization. [1,2,142−144] Consider rapeseed (aka canola seed), where approximately 40% of the harvest produces oil and 60% remains as seed meal, or roughly 1.8 t/ha meal for every 3 t/ha grown. Typical concentration of protein in canola meal is about 38%, or 0.684 t protein/ha. The advent of Pleurotus increases the protein fraction by 11–18% (Table 1) while protein digestion inhibitors are reduced by 55–99%.[24,79] If assuming an average protein increase to 43.7% (15% relative) and total mass loss to respiration of 0–10%, this would result in a net increase in edible protein to between 0.708–0.787 t/ha. In other words, SSF would produce up to an additional 24–103kg protein/ha. On the human scale, that incremental gain alone would supply up to 5.6 adult-years of protein per hectare, per year (based on international recommendation levels [145]). This change would also increase iron and zinc bioavailability from phytate removal and improve sensory attributes that may drive higher real-world adoption compared to the standard plant crops grown on the same land.[9,24,37,79]
Collectively, by converting low-cost crops and agricultural side streams into nutrient-dense, desirable foods with minimal processing, short production cycles, and strong sensory performance, SSF can be a pragmatic strategy to improve diet quality and food security. This approach also aligns with public health priorities for higher protein, higher fiber, and lower saturated fat diets while enhancing circular-bioeconomy goals (co-location, waste minimization; see below) all without reliance on ultra-processing or long, fragile supply chains. [108,114,143,144]
Substrate Selection Strategies for Fungal SSF
Suitable substrates (Table 3) for SSF should provide carbohydrates, protein, nitrogen, and minerals, while minimizing factors that inhibit fungal growth (e.g., extreme pH, toxins).[3,34] Common substrate categories include cereals, pseudocereals, legumes, oilseed cakes, omega fatty-acid-dense seeds, aquatic biomass [146−148], and bran-rich material from agricultural side streams.[96] Effective substrate design often blends materials to achieve an optimal carbon to nitrogen ratio and moisture level, while targeting desired nutritional outcomes. For example, a mixture of 70% cereal grain and 30% legume press cake can supply the needed carbon and nitrogen while yielding a balanced protein-fiber end product. Modifying substrate composition can significantly influence fungal performance and the resulting nutritional and sensory attributes [34].
Growth Conditions for Optimizing SSF
An interacting set of variables impact the microenvironment around hyphae, influencing the physiochemistry of the mycelium, and the time to maturity. Numerous factors can be manipulated to manage the outcomes, including: substrate moisture, substrate nutrients, water activity (aw), temperature (and heat removal), particle size, bed depth, and aeration, agitation, inoculum load and distribution, and ambient relative humidity. Fungal growth typically requires high aw (appx. 0.95–0.99), but not as free water; excess and/or localized moisture collapses inter-particle pores, choking oxygen transfer and limiting enzymatic activity and biomass formation.[2,149] Heterogeneity in growth bed construction can further create moisture and temperature gradients that alter colonization rates and metabolite production. Gentle forced aeration (e.g., systematic agitation or vibration) helps distribute oxygen and moisture, and when paired with strategic bed geometry, can support robust growth and reproducible yield characteristics.[1,2,102]
Optimal conditions for growth often include temperatures between 24–30°C, with relative humidity (RH) between 85–90% to minimize evaporative losses, in shallow beds that facilitate easy heat removal. When temperatures exceed 35°C, common species (e.g. Pleurotus) experience slowed mycelial growth and thermal hotspots that can result in quality loss, reduced yields, or stunting, suggesting that forced-air systems should provide sufficient O2 while simultaneously delivering gentle cooling in trays or packed beds. Growth conditions impact time-to-maturity, with tempeh-style Rhizopus systems typically reach a cohesive, sliceable cake in as little as 24–48 hours at 30–35°C and 85–90% RH, whereas Pleurotus tray cultures require several days to weeks, influenced by bed depth, inoculum density, and heat management.[46,150]
Substrate engineering also strongly influences mycelia growth. Particle size and distribution can create interparticle air gaps that promote gas diffusion, providing ample surface for hyphal attachment and penetration. In contrast, overly fine particles increase bed resistance and restrict oxygen transfer, and excessively coarse fractions hinder colonization uniformity and may prevent fungal access to nutrient dense starch stores beyond hard cell wall barriers. Mild pre-treatments to the substrate (hydration, gelatinization for starch-rich cereals, or mild enzymatic conditioning) can accelerate colonization. Buffering, or initial acidification of the substrate (e.g., to pH 4.5, as in tempeh manufacture) provides an effective microbial hurdle, suppressing spoilage bacteria during early stages of fermentation while remaining permissive to fungal germination and early hyphal growth. Once the fungus established, pH tends to drift toward neutral pH (6.5-7) as organic acids are metabolized.[151]
Circadian and photic cues are increasingly recognized as major regulators of fungal metabolism and development, yet most food-grade SSF protocols employ steady, constant setpoints (entrainment of darkness, temperature and RH). Neurospora is the canonical fungal strain for the circadian clock model where endogenous transcription–translation feedback loops have been seen to entrain robustly to both light/dark and cool/warm temperature cycles with significant resultant effects on primary and secondary metabolism, conidiation, and stress responses.[152−154] These principles generalize across filamentous fungi with light acting through conserved photoreceptors to reprogram metabolism and development; thus, timed light/temperature cues can modulate metabolic flux—i.e., the rate of carbon passage—through glycolysis and the pentose-phosphate pathway, alter redox balance, and shift volatile and pigment profiles.[155,156] Although basidiomycetes used in foods are less clock-mapped than Neurospora, Pleurotus spp. clearly perceive blue light via White Collar-1/2 homologs, and blue-light exposure modulates primordia formation and central carbon metabolism, and are responsive during vegetative growth stages in SSF.[157,158] SSF process design could thus improve yields and sensory outcomes by moving beyond static setpoints and toward programmable circadian-optimized regimes to allow synchronized hyphal growth and enzyme secretion, reduce heat-of-metabolism bottlenecks, and potentially increase desirable flavors and pigments—fertile terrain for future hypothesis testing.
Food System Benefits: Sustainability and Food Security
SSF offers value-added endpoints for commodity crops and agricultural side streams alike. Side streams that are otherwise low-cost, aesthetically imperfect main crops, or edible by products are underutilized materials and a significant valorization opportunity for enhancing circular food systems, bolstering rural economies, and for improving nutrition security at both local and international scale. Valorization allows the same agricultural inputs (e.g. land, water, fertilizer) used to grow main crops to feed more people, improving the resource efficiency of the food system.[3,11,99,159−161] Many crops produce a large fraction of inedible or underutilized residues (e.g. nut hulls, cereal brans, seed cakes, fruit pomace) that are typically relegated to edible low-value streams animal feed, or inedible low-value streams like packing and fillers, when not fully lost as farm waste.[25,72,162] Utilization for SSF directly addresses the United Nations Sustainable Development Goals of Zero Hunger and Responsible Consumption/Production, by increasing the food supply without clearing additional land for cultivation and minimizing waste [3,11]. For one example, in global peanut production, hulls are >20% of the yield and are mostly used as feed or processed into packaging materials.[162] Through SSF, peanut hulls can feed flavorful nutrient dense mycelium, thus boosting the total human-edible output per hectare, increasing farm diversification options, and contributing to food security in peanut-growing regions.
SSF could further support food security through nimble and strategic deployment owing to low resource requirements and utilization of simple, modular infrastructure.[163] SSF cultures can be distributed in compact and dehydrated formats allowing a variety of available substrates to be inoculated with simple rehydration[164], enabling fast establishment of food production systems in resource-constrained or decentralized settings [165], including following natural and political disasters which is increasingly identified as an urgent priority for global food security.[166] SSF-relevant fungi can be transported as lightweight, lyophilized, shelf-stable inoculum or dried solid-state cultures; some, like Trichoderma, that retain viability for over three months.[167] This low-cost, and low-resource operation allows hyperlocal activation on regionally available foodstuffs, or waste and by-product streams, with a capacity to produce nutritionally enhanced protein foods quicker than legacy agricultural systems. Temporal decoupling of production and use, and efficient valorization of local resources together position SSF as a promising path to enhance food security during crises or in other constrained environments, including long-duration space missions where time, space, and resource efficiency are critical.[168,169]
Fungal fermentation has a relatively low environmental footprint compared to cultivation of many traditional protein sources. Cultivating mushrooms or mycelium emits fewer greenhouse gases and uses less water and land than raising livestock. For instance, Quorn’s Carbon Trust–certified footprint analysis results report a farm-to-factory-gate carbon footprint for Quorn mycoprotein (Fusarium venenatum) of 0.7 kg CO2 per kg mycoprotein, reporting a footprint that is 55× lower for carbon, 13.5× lower for water, and 5.5× lower for land use than beef.[170] Additionally, mushrooms and mycelium can be grown indoors, minimizing competition with field agriculture for land, extending production throughout the year and multiple regions, while buffering against seasonal disruptions. Fungi also exhibit high substrate-to-biomass conversion efficiency because they expend no energy on locomotion or thermoregulation, unlike livestock. In optimized systems of edible SSF such as tempeh, fungal biomass is predicted to comprise approximately 5.9% of the final product on a dry-weight basis, whereas harvested mycelial-mat systems, such as those using Pleurotus report yields of over 12% accrual above starting substrate mass.[2,171,172] These biomass gains are not all substrate conversion, as prolonged SSF with some fungi can incur dry-matter losses of an estimated 30.9%. Substrate utilization and biomass accrual will be tunable by growth conditions and strain mechanics, with commercial endpoints aiming for products with closer to 50% fungi and 50% plant-based remaining substrates.[173]
SSF for Farm Economies
SSF offers downstream and on-farm revenue diversification for farmers and local producers. The increasing global consumption of animal protein has resulted in larger proportions of arable land used to grow crops for animal feed, rather than human food. Over half of habitable land is currently used for agriculture, with over 80% estimated to be used for livestock between raising, grazing, and feedstock.[174,175] Over recent decades, approximately 36% of global crop calories grown are currently used for animal feed globally, with over a third of croplands devoted to feed crops. In the U.S., most corn in a high-concentration corn growing region, goes to livestock rather than human foods.[174,176,177] By creating additional human demand for crops and agricultural byproducts, SSF can increase revenue from the same raw materials for rural economies. Rather than selling these materials solely as animal feed, farmers could supply them to fermentation manufacturers or implement on-farm fermentation to reduce waste and lower transportation costs between process steps. Studies also show that upcycled side streams command higher prices when repurposed for human food than when sold as low-value feed or fuel.[25,72,162] Further, production of on-farm fungi dovetails with many national U.S. and international initiatives for rural development. For example, oyster mushroom cultivation is already promoted by the Food and Agriculture Organization of the U.N. and the United States Department of Agriculture for farm diversification as a low-investment, high-nutrition crop that farmers or cooperatives can grow on wastes.[178−180] SSF extends this by expanding the range of substrates for growth and the resulting end products, which can enrich local diets to combat malnutrition and command premium prices from staple commodity crops, giving farmers access to higher-value markets.
Commercial Developments and Emerging Mycelium-Based Foods
Mycoprotein products have been on the market for several decades, but a new wave of interest has increased the number of startup enterprises seeking to leverage fermentation technology to create novel foods. From legacy to latest, fungal foods now span industrial mycoprotein, traditional SSF staples (e.g. tempeh), whole-cut mycelium meats, and protein fortified flours and flavor dry ingredients. Quorn™ pioneered large-scale mycoprotein (submerged Fusarium venenatum) and proved mainstream appeal of its high protein and higher fiber chicken alternatives.[170] Southeast Asian classics like tempeh (soybeans + Rhizopus) and oncom (peanut and/or soy + Neurospora) demonstrate SSF’s ability to boost digestibility, vitamins, and savory flavor. Newer platforms on the market have shown thick Pleurotus mycelium “slabs” like the MyForest/Atlast sliceable bacon; Rhiza produced by Better Meat Co. as a fast-growing filamentous fungus on low-value plant potato waste; Neurospora strains producing cutlet-style products like Meati, and the Koji deli meat formats of Prime Roots that produce fibrous mycelia in submerged tanks or trays [170]. Aside from meat analogues, mycelium is increasingly used as a functional processing step—debittering and improving plant proteins (e.g., shiitake-treated pea and rice), fortifying flour, and to generate enzyme- or antioxidant-rich ingredients for breads, gravies and other foods—highlighting the versatile, clean-label potential.[63,181]
As consumers grow more conscious of health and sustainability, fungi-derived functional foods are poised to capture a significant share of the protein market. For example, Quorn notes that their increased fiber ‘may help regulate blood cholesterol levels’, and Meati emphasizes that their mycelium is packed with ‘protein, vitamin B12, and minerals, while being low in fat’. Alternatively, applications other than meat alternatives, such as high-protein SSF flour can be used to fortify breads, tortillas, and snacks, or dried and micronized into natural umami-rich seasoning powders.[15,40] Applications in wheat breads with Pleurotus ostreatus amendments showed increased protein and antioxidants while retaining good loaf and crumb texture [119]; similar approaches to functional enhancements in bread with P. eryngii increased nutrient density and observed acceptable liking scores from tasters,[134] as with Agaricus and P. sajor-caju fortified breads [135,136] cookies, and snacks. [118]
Compared with liquid-state fermentation (LSF), solid-state fermentation (SSF) is underutilized because its core engineering and control problems are harder to solve at scale. Heterogeneous moist beds impede heat and mass transfer, creating local hot spots, moisture/aw and O2/CO2 gradients that reduce reproducibility and complicate scale-up—issues largely tamed in stirred, well-mixed LSF bioreactors.[1,2,4,102] SSF also lacks mature on-line instrumentation: moisture, metabolic rate, and endpoint prediction are often inferred indirectly (respirometry, NIR) rather than measured through robust probes common to LSF.[1,97] Feedstock variability further raises transaction costs—agri-byproducts require food-grade presorting, particle-size/voidage control, and thermal pre-treatments for safe, uniform colonization—whereas LSF typically uses standardized liquid media.[142−144] Microbiological risk management differs as well: SSF depends on bed hygiene, defined GRAS (Generally Regarded as Safe, or comparable determinations in regions outside of the U.S.) starters (e.g., Rhizopus, Aspergillus oryzae), and early hurdles (acidification/heat) to suppress competitors and mycotoxin risks, whereas LSF leverages closed, CIP/SIP vessels and antibiotics-free asepsis.[139,182−184] Finally, capital ecosystems, regulatory familiarity, and workforce experience are deeper for LSF (ubiquitous tanks, sensors, etc.), so investment and know-how naturally pool there despite SSF’s potential low-capital expense (CAPEX), low-utilities advantages.[2,142] SSF and LSF are thus best viewed as complementary platforms: LSF excels at producing homogeneous biomass and more purified ingredients under tightly controlled, high-water conditions, whereas SSF enables low-water transformation of solid food matrices that can increase intake of both the substrate and fungal biomass as a minimally processed composite food, with distinct constraints and advantages.
Recent advances demonstrate that SSF can serve as a foundational platform for hybrid protein production systems. SSF-derived mycelium is a promising scaffold for anchorage-dependent cell cultures in the production of another emerging food technology—cell-cultivated meat (sometimes referred to as in-vitro, or lab-grown meat).[185] For example, Pleurotus ostreatus mycelium has been shown to support bovine satellite cell adhesion and proliferation, providing a structurally edible matrix with tunable porosity and mechanical integrity.[186] further validated the use of Ganoderma lucidum mycelium in such hybrid constructs, showing enhanced cell viability and nutrient diffusion compared to more common collagen-based scaffolds. These platforms not only reduce reliance on animal-sourced matrices but also enable sensory and nutritional optimization.
Mycelium scaffolds produced by SSF can be nutritionally crafted with substrate design and process control. For example, co-fermenting legumes and cereals with flaxseed press-cake or other ω-3–rich inputs shifts the lipid profile to be higher in α-linolenic acid with favorable ω-6:ω-3 ratios, while retaining fungal protein and dietary fiber.[96] Beyond composition, the intertwined anisotropic structures can function as a fungal connective-tissue armature that can be further shaped by moisture, particle size, voidage, and growth time for desired composition. This approach can offer sensory and structural enhancements when paired with pea-protein systems for meat analogues.[24]
Meat-like coloration can also be enhanced with pigments from fungal fermentation. For example, Neurospora generates carotenoids (e.g., neurosporaxanthin) and Monascus red pigments provide heat-stable red/pink hues suitable for raw and cooked analogues, offering cleaner-label routes to natural and edible colorants with nutritional value.[187−189] This approach potentially has direct applications within cell-cultivated meat and fish technologies, and represents a convergence of bioprocessing and tissue engineering. The ability to combine natural and familiar mushroom-derived processes into plant-based and other novel protein foods, can help meet consumer needs, public health nutritional and gastronomic goals.[190]
Food Safety and Regulatory Considerations
SSF with edible fungi is safe when producers pair pure, food-grade starter cultures with aseptic, well-controlled processes to suppress contaminants, prevent toxins, manage allergens, and comply with applicable approvals. Fast-colonizing strains give the inoculum a competitive edge able to outcompete adventitious agents and later spoilage organisms; together, with substrate pasteurization or sterilization and pre-acidification, the pathogenic risk is reduced. Careful development of Good Manufacturing Procedures (GMP) and Hazard Analysis and Critical Control Point (HACCP) frameworks can also further specify sterilization, clean inoculation, and end-point moisture as critical controls among others. Using defined monocultures improves reproducibility and safety.[5,139,191] Strain choice and other process conditions can also mitigate risk—edible species such as Pleurotus, Agaricus, Lentinula, and tempeh’s Rhizopus oligosporus are used in food fermentation without problematic mycotoxin production. Domesticated Aspergillus oryzae (koji mold) used for centuries in traditional foods and in modern commercial applications (e.g. Prime Roots deli meats) has a long history of safe food use. Comparative genomic studies showed that the domestication of koji derived from atoxigenic lineages.[192,193] Aside from the in-field use of atoxigenic Aspergillus flavus strains as biocontrol pre-harvest, post-harvest biodegradation/biotransformation approaches include fungal- and enzyme-mediated transformation of aflatoxins under controlled conditions, including reports using Trichoderma reesei.[182,192,194−197] Laccases/peroxidases produces from many strains have also been demonstrated in controlled LSF fermentation to oxidize aflatoxins.[198] Companies (e.g., MyForest/Atlast) have argued that mycelium is “substantially similar” to fruiting bodies of the same mushrooms, supporting low-novelty safety claims. Producers have also gone to great lengths to demonstrate methodological and end point safety, including to verify absence of mycotoxins in products such as Fusarium mycoprotein (Quorn) as part of GRAS submissions.[173,199−202] At industrial scale, safety programs track pH, moisture, and temperature with post-fermentation kill-steps (pasteurization/drying) and protective packaging that can help to prevent recontamination, and traceably document sourcing clean, food-grade inputs (especially for byproducts that could carry residues) remains essential. Regulatory pathways vary by region: in the U.S., many edible fungi qualify via GRAS or food-additive petitions (e.g., Fusarium venenatum mycoprotein; GRN 945), and oyster-mushroom mycelium has been the subject of a recent GRAS notice by Mushlabs (GRN 1152, submitted but FDA ceased review in 2024[203]); other firms (e.g., Better Meat Co.’s Rhiza mycoprotein) have received “no questions” letters.[204] In the EU, several mycoproteins entered via Novel Food authorization. Ongoing calls for harmonized standards tailored to mycelium protein emphasize process control, toxin/allergen risk management, and accurate labeling, reflecting regulators’ receptivity when dossiers are robust.[202,204,205]
2. Discussion and the Future of SSF
Solid-state fermentation using culinary mushroom fungal strains is a practical bridge between traditional fermentation and next-generation sustainable foods. By working directly on solid substrates, SSF—unlike LSF—can transform intact agricultural crops into minimally processed, flavor-forward, high-protein foods, enhancing bioavailability and palatability without reliance on isolates or ultra-processing. To achieve the goal of a more sustainable food system, SSF can help to reduce food waste and increase food supply efficiency by valorizing commodity crops and side streams, producing high-quality protein with a low environmental footprint, creating economic opportunities for agriculturalists and food manufacturers alike.[170]
Demand for protein foods is expected to outpace supply by as soon as 2050, creating an important and timely need for innovations that not only deliver more protein foods but that can effectively meet the acceptance needed to serve that demand.[206,207] Current gaps in understanding include the generalizable, direct, mediated, and synergistic quantitative links between (i) substrate precursors and enzyme portfolios, (ii) process variables and variable schema (moisture, particle size, temperature, aeration, mono- vs. co-culture), (iii) methods for environmental and stress provocation of chemical drivers of taste and aroma, (iv) perceived sensory outcomes and resultant substitution and adoption, and (v) and consequential techno-economic outcomes when optimizing for any of the above. Resolving these unknowns calls for factorial SSF studies that integrate targeted functional analytics, comparisons of pre/post cooking methods, tied to predictive process–product-adoption maps. Producing a validated design framework that maps these interdependencies would de-risk scale-up, guide co-culture selection, and accelerate minimally processed SSF foods toward nutrition security and commercialization. With optimized processes, SSF can be scaled to produce delicious, health-promoting foods that are cost-effective to manufacture and affordable for consumers, while increasing yield and shortening time-to-harvest—benefits that strengthen both farm economies and nutrition security. Continued optimization can also support the development of improved edible fungal strains that grow more rapidly, generate specific health-promoting compounds, and more efficiently utilize diverse inputs, enhancing nutritional profiles and expanding waste-valorization opportunities.
3. Conclusions
SSF is a promising solution to our pressing food challenges. It provides a route to improve human nutrition by merging the strengths of plant foods and fungal biotechnology, yielding products that can help diversify protein sources and mitigate malnutrition—with methods and ingredient lists consumers will recognize and that public health officials can endorse. SSF allows for sustainable food systems enabling greater consumption of minimally processed plant foods, along with enhanced circular approaches. While challenges in optimization, scaling and consumer education remain, the progress to date signals that fungal fermentation can indeed move from the realm of niche research and traditional practice into the mainstream of global food supply. With continued interdisciplinary effort, mushroom mycelium foods could emerge as a staple contributor to high-protein, functional foods that support human and planetary health, while enhancing rural economies in the years ahead.
Author Contributions
SG: Conceptualization; Scope framing; Methodology; Literature Review; Formal analysis; Writing the original draft; review & editing; Project administration; Supervision; Funding acquisition. SCF: Literature review; writing, review & editing; Illustration. AMW: writing, review & editing. DLK: Critical review, significant edits and additions. All authors approved the final manuscript and agree to be accountable for its contents.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Funding for open access was provided by Tufts University Hirsh Health Sciences Library’s Open Access Fund. No funders initiated this research, participated in the research or the decision to publish. This work corresponds to a preprint; submission to a verified preprint server is required by the Tufts University Hirsh Health Sciences Library Open Access Fund. This work is partially supported by the Agriculture and Food Research Initiative Sustainable Agricultural Systems program, project award no. 2021-699012-35978, from the U.S. Department of Agriculture's National Institute of Food and Agriculture.
Acknowledgments
Generative AI (ChatGPT, OpenAI; accessed Jan 2026) was used by the authors strictly for language clarity editing, table note reformatting, citation formatting for upload to Endnote, and to create illustrations for the graphical abstract. All authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
Author SG serves as the Executive Director of the Association for Meat, Poultry & Seafood Innovation (AMPS), a trade association focused on cell-cultivated meat with no financial or operational interests related to food fermentation or mycelium-based foods. SG previously provided short-term contracted, paid, scientific and product-design consulting through her company Haven Foods Consulting to an SSF company (Celium) which concluded in June 2025. SG holds no current role, equity, or financial interest in Celium or any SSF/mycelium company. The remaining authors declare no competing interests.
References
- Hölker U, Höfer M, Lenz J. Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Appl Microbiol Biotechnol. 2004;64(2):175-186. [CrossRef]
- Thomas L, Larroche C, Pandey A. Current developments in solid-state fermentation. Biochemical Engineering Journal. 2013;81:146-161. [CrossRef]
- Wijaya CH, Prabawati PI, et al. Oncom: A Nutritive Functional Fermented Food Made from Food Process Solid Residue. Applied Sciences. 2024;14(22):10702. [CrossRef]
- Hölker U, Lenz J. Solid-state fermentation--are there any biotechnological advantages? Curr Opin Microbiol. 2005;8(3):301-306. [CrossRef]
- Milcarz A, Harasym J. Solid State Fermentation—A Promising Approach to Produce Meat Analogues. Foods. 2025;14(10). [CrossRef]
- Elhalis H, See XY, Osen R, Chin XH, Chow Y. Significance of Fermentation in Plant-Based Meat Analogs: A Critical Review of Nutrition, and Safety-Related Aspects. Foods. 2023;12(17):3222. [CrossRef]
- Pandey A. Solid-state fermentation. Biochemical Engineering Journal. 2003;13(2):81-84. [CrossRef]
- Elhalis H, See XY, Osen R, Chin XH, Chow Y. The potentials and challenges of using fermentation to improve the sensory quality of plant-based meat analogs. (1664-302X (Print)). [CrossRef]
- Eklund-Jonsson C, Sandberg A-S, Hulthén L. Tempe Fermentation of Whole Grain Barley Increased Human Iron Absorption and In Vitro Iron Availability. The Open Nutrition Journal. 2008;2:42-47. [CrossRef]
- Ahnan-Winarno AD, Cordeiro L, Winarno FG, Gibbons J, Xiao H. Tempeh: A semicentennial review on its health benefits, fermentation, safety, and sustainability. Comprehensive Reviews in Food Science and Food Safety. 2021;20(2):1717-1767. [CrossRef]
- Holt RR, Munafo JP, Jr., Salmen J, et al. Mycelium: A Nutrient-Dense Food To Help Address World Hunger, Promote Health, and Support a Regenerative Food System. J Agric Food Chem. 2024;72(5):2697-2707. [CrossRef]
- Garrido-Galand S, Asensio-Grau A, Calvo-Lerma J, Heredia A, Andrés A. The potential of fermentation on nutritional and technological improvement of cereal and legume flours: A review. Food Res Int. 2021;145:110398. [CrossRef]
- GFI. Deep dive: Fermentation upstream bioprocess design. Good Food Institute2025.
- Chaffee O, Ardoin R. Consumer perceptions of plant-based and mushroom-based jerky: A focus on texture, main ingredient and protein information, and willingness to pay. Current Research in Food Science. 2025;10:101058. [CrossRef]
- Carrasco-González JA, Serna-Saldívar SO, Gutiérrez-Uribe JA. Nutritional composition and nutraceutical properties of the Pleurotus fruiting bodies: Potential use as food ingredient. Journal of Food Composition and Analysis. 2017;58:69-81. [CrossRef]
- Grabarczyk M, Mączka W, Wińska K, Uklańska-Pusz C. Mushrooms of the Pleurotus genus—properties and application. Biotechnology and Food Science. 2019;83(1):13-30.
- Yin Z, Sun-Waterhouse D, Wang J, Ma C, Waterhouse GIN, Kang W. Polysaccharides from edible fungi Pleurotus spp.: advances and perspectives. Journal of Future Foods. 2021;1(2):128-140. [CrossRef]
- Sabu A, Sarita S, Pandey A, Bogar B, Szakacs G, Soccol CR. Solid-state fermentation for production of phytase by Rhizopus oligosporus. Appl Biochem Biotechnol. 2002;102-103(1-6):251-260. [CrossRef]
- Manikandan A, Muthusamy S, Wang ES, et al. Breeding and biotechnology approaches to enhance the nutritional quality of rapeseed byproducts for sustainable alternative protein sources- a critical review. Frontiers in Plant Science. 2024;Volume 15 - 2024.
- Jatuwong K, Suwannarach N, Kumla J, Penkhrue W, Kakumyan P, Lumyong S. Bioprocess for Production, Characteristics, and Biotechnological Applications of Fungal Phytases. Frontiers in Microbiology. 2020;Volume 11 - 2020.
- Ritota M, Manzi P. Edible mushrooms: Functional foods or functional ingredients? A focus on Pleurotus spp. AIMS Agriculture and Food. 2023;8(2):391-439. [CrossRef]
- Rodrigues BJ, dos Santos Freitas MM, da Silva Martins LH, de Carvalho RN. Polysaccharides of mushroom Pleurotus spp.: New extraction techniques, biological activities and development of new technologies. Carbohydrate Polymers. 2020;229:115550. [CrossRef]
- Torres-Martínez BM, Vargas-Sánchez RD, Torrescano-Urrutia GR, et al. Pleurotus genus as a potential ingredient for meat products. Foods. 2022;11. [CrossRef]
- Heidari F, Øverland M, Hansen JØ, et al. Solid-state fermentation of Pleurotus ostreatus to improve the nutritional profile of mechanically-fractionated canola meal. Biochemical Engineering Journal. 2022;187:108591. [CrossRef]
- Eliopoulos C, Langousi I, Kougia E, et al. Solid-State Fermentation Initiated by Pleurotus ostreatus of a Cottonseed Cake and Lathyrus clymenum Pericarp Mixture: Impact on Nutritional Profile and Gossypol Content. Applied Sciences. 2024;14(12). [CrossRef]
- Lee I-H, Hung Y-H, Chou C-C. Solid-state fermentation with fungi to enhance the antioxidative activity, total phenolic and anthocyanin contents of black bean. International Journal of Food Microbiology. 2008;121(2):150-156. [CrossRef]
- Lee AML, Chin CFS, Seelan JSS, Chye FY, Lee HH, Rakib MRM. Metabolites profiling of protein enriched oyster mushroom (Pleurotus ostreatus (Jacq.) P. Kumm.) grown on oil palm empty fruit bunch substrate. LWT. 2023;181:114731. [CrossRef]
- Adebo OA, Gabriela Medina-Meza I. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules. 2020;25(4). [CrossRef]
- Pascual MM, Herbert LT, Campos M, Jurski V, Painefilú JC, Luquet CM. Nutritional improvement of wheat grains and soybeans by solid-state fermentation with Pleurotus ostreatus mycelium. Innovative Food Science & Emerging Technologies. 2025;102:104021. [CrossRef]
- Kaneko K, Aoyagi Y, Fukuuchi T, Inazawa K, Yamaoka N. Total purine and purine base content of common foodstuffs for facilitating nutritional therapy for gout and hyperuricemia. Biol Pharm Bull. 2014;37(5):709-721. [CrossRef]
- Żuchowski J, Pecio Ł, Jaszek M, Stochmal A. Solid-state fermentation of rapeseed meal with the white-rot fungi trametes versicolor and Pleurotus ostreatus. Appl Biochem Biotechnol. 2013;171(8):2075-2081. [CrossRef]
- Niu Y, Jiang M, Guo M, et al. Characterization of the Factors that Influence Sinapine Concentration in Rapeseed Meal during Fermentation. PLOS ONE. 2015;10(1):e0116470. [CrossRef]
- Zhang WJ, Xu ZR, Zhao SH, Jiang JF, Wang YB, Yan XH. Optimization of process parameters for reduction of gossypol levels in cottonseed meal by Candida tropicalis ZD-3 during solid substrate fermentation. Toxicon. 2006;48(2):221-226. [CrossRef]
- Diamantopoulou P, Fourtaka K, Melanouri EM, et al. Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms. Fermentation. 2023;9(7). [CrossRef]
- Ritota M, Manzi P. Pleurotus spp. cultivation on different agri-food by-products: Example of biotechnological application. Sustainability. 2019;11(18). [CrossRef]
- Boin E, Cláudia A, João N, Guerra MM. Consumer Acceptability and Descriptive Characterization of Fresh and Dried King Oyster (Pleurotus eryngii) and Hedgehog (Hydnum repandum) Mushrooms. Journal of Food Research. 2016;5:55-55. [CrossRef]
- Eklund-Jonsson C, Sandberg A-S, Alminger M. Reduction of phytate content while preserving minerals during whole grain cereal tempe fermentation. Journal of Cereal Science. 2006;44(2):154-160. [CrossRef]
- He M, Peng Q, Xu X, Shi B, Qiao Y. Antioxidant capacities and non-volatile metabolites changes after solid-state fermentation of soybean using oyster mushroom (Pleurotus ostreatus) mycelium. Frontiers in Nutrition. 2024;Volume 11 - 2024.
- Mumpuni A, Ekowati N, Purnomowati P, Purwati E. Growth and Protein Content Establishment of Pleurotus ostreatus on Liquid and Solid Medium. Biosaintifika: Journal of Biology & Biology Education. 2017;9:572. [CrossRef]
- Reis FS, Barros L, Martins A, Ferreira ICFR. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter-species comparative study. Food and Chemical Toxicology. 2012;50(2):191-197. [CrossRef]
- Wang X-M, Zhang J, Wu L-H, et al. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chemistry. 2014;151:279-285. [CrossRef]
- Meyer F, Hutmacher A, Lu B, Steiger N, Nyström L, Narciso JO. Vegan shrimp alternative made with pink oyster and lion's mane mushrooms: Nutritional profiles, presence of conjugated phenolic acids, and prototyping. Curr Res Food Sci. 2023;7:100572. [CrossRef]
- Srivastava A, Attri BL, Bijla S, Arjun AD, Sharma P. Utilization of Lion’s Mane mushroom (Hericium erinaceus) for vegan crab analogue: effects of pre-treatments on nutritional, sensory, and physicochemical properties. Journal of Food Science and Technology. 2025. [CrossRef]
- Gonkhom D, Luangharn T, Raghoonundon B, Hyde K, Stadler M, Thongklang N. Hericium: A review of the cultivation, health-enhancing applications, economic importance, industrial, and pharmaceutical applications. Fungal Biotec. 2021;1:115-127. [CrossRef]
- Ge F, Chen Y, Wang B, Zhou W, Du B, Hou L. Bioactive Polysaccharides from Hericium erinaceus: Extraction, Structure, Bioactivities, and Applications. Molecules. 2025;30(8):1850. [CrossRef]
- Górska N, Włodarczyk K, Różańska A. Tempeh and Fermentation—Innovative Substrates, Co-Cultures, and Nutritional Potential: A Review. Applied Sciences. 2025;15(16):8888. [CrossRef]
- Song W, Li M, Yue X, Zheng Y. Effect of extraction and purification on the structure and activity of Flammulina velutipes polysaccharides: a review. International Journal of Food Science and Technology. 2024;59(12):9571-9591. [CrossRef]
- Bandara AR. A review of the polysaccharide, protein and selected nutrient content of Auricularia, and their potential pharmacological value. Mycosphere. 2019;10(1):579-607. [CrossRef]
- Sone Y, Kakuta M, Misaki A. Isolation and Characterization of Polysaccharides of “Kikurage,” Fruit Body of Auricularia auricula-judae. Agricultural and Biological Chemistry. 1978;42(2):417-425. [CrossRef]
- Ma Z, Wang J, Zhang L. Structure and chain conformation of β-glucan isolated from Auricularia auricula-judae. Biopolymers. 2008;89(7):614-622. [CrossRef]
- Liu M, Jiang W, Huang K, et al. Sustainable Cultivation and Functional Bioactive Compounds of Auricularia Mushrooms: Advances, Challenges, and Future Prospects. Biology. 2025;14(11):1555. [CrossRef]
- Regis M, Geösel A. Cultivation of Auricularia species: a review of the history, health benefits, principles, practices, environmental conditions, research methods, and recent trends. Sydowia. 2023;76.
- Geng J, Wang G, Guo J, et al. Preparation and structural analysis of fucomannogalactan and β-1,6-glucan from Grifola frondosa mycelium. Frontiers in Chemistry. 2023;Volume 11 - 2023.
- Yang Y-M, Fu X, Cui F-J, Sun L, Zan X-Y, Sun W-J. Biochemical and structural characterization of a glucan synthase GFGLS2 from edible fungus Grifola frondosa to synthesize β-1, 3-glucan. Biotechnology for Biofuels and Bioproducts. 2023;16(1):163. [CrossRef]
- Cerletti C, Esposito S, Iacoviello L. Edible Mushrooms and Beta-Glucans: Impact on Human Health. Nutrients. 2021;13(7). [CrossRef]
- Miles PG, Chang S-T. Mushrooms: cultivation, nutritional value, medicinal effect, and environmental impact. CRC press; 2004.
- Lee DJ, Kang AN, Lee J, et al. Molecular characterization of Fusarium venenatum-based microbial protein in animal models of obesity using multi-omics analysis. Communications Biology. 2024;7(1):133. [CrossRef]
- Boye J, Wijesinha-Bettoni R, Burlingame B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br J Nutr. 2012;108 Suppl 2:S183-211. [CrossRef]
- Wolf WJ. Soybean proteins. Their functional, chemical, and physical properties. Journal of Agricultural and Food Chemistry. 1970;18(6):969-976. [CrossRef]
- Friedman M, Brandon DL. Nutritional and Health Benefits of Soy Proteins. Journal of Agricultural and Food Chemistry. 2001;49(3):1069-1086. [CrossRef]
- Chen G, Liu Y, Zeng J, Tian X, Bei Q, Wu Z. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. Journal of Cereal Science. 2020;93:102940. [CrossRef]
- Queiroz Santos VA, Nascimento CG, Schmidt CAP, Mantovani D, Dekker RFH, da Cunha MAA. Solid-state fermentation of soybean okara: Isoflavones biotransformation, antioxidant activity and enhancement of nutritional quality. LWT. 2018;92:509-515. [CrossRef]
- Verduzco-Oliva R, Gutierrez-Uribe JA. Beyond Enzyme Production: Solid State Fermentation (SSF) as an Alternative Approach to Produce Antioxidant Polysaccharides. Sustainability. 2020;12(2):495. [CrossRef]
- Mukherjee R, Chakraborty R, Dutta A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal - A Review. Asian-Australas J Anim Sci. 2016;29(11):1523-1529. [CrossRef]
- Bekirian C, Valsecchi I, Bachellier-Bassi S, et al. β-1,6-Glucan plays a central role in the structure and remodeling of the bilaminate fungal cell wall. Elife. 2024;13. [CrossRef]
- Colosimo R, Mulet-Cabero A-I, Warren FJ, Edwards CH, Finnigan TJA, Wilde PJ. Mycoprotein ingredient structure reduces lipolysis and binds bile salts during simulated gastrointestinal digestion. Food & Function. 2020;11(12):10896-10906. [CrossRef]
- Jiang Q, Zhang M, Mujumdar AS. UV induced conversion during drying of ergosterol to vitamin D in various mushrooms: Effect of different drying conditions. Trends Food Sci Technol. 2020;105:200-210. [CrossRef]
- Cardwell G, Bornman JF, James AP, Black LJ. A Review of Mushrooms as a Potential Source of Dietary Vitamin D. Nutrients. 2018;10(10). [CrossRef]
- Bashir KMI, Choi J-S. Clinical and Physiological Perspectives of β-Glucans: The Past, Present, and Future. International Journal of Molecular Sciences. 2017;18(9). [CrossRef]
- Kumari R, Koul B, Kumar V, Kumar A, Somal MK, Sachan RSK. Protease and chitinase activity of Trichoderma isolates and their synergy with biochar in enhancing chickpea defense related enzymes. Frontiers in Microbiology. 2025;Volume 16 - 2025.
- El-Shora HM, El-Sharkawy RM, Khateb AM, Darwish DB. Production and immobilization of β-glucanase from Aspergillus niger with its applications in bioethanol production and biocontrol of phytopathogenic fungi. Sci Rep. 2021;11(1):21000. [CrossRef]
- Scholtmeijer K, van den Broek LAM, Fischer ARH, van Peer A. Potential Protein Production from Lignocellulosic Materials Using Edible Mushroom Forming Fungi. J Agric Food Chem. 2023;71(11):4450-4457. [CrossRef]
- Gardeli C, Mela N, Dedousi M, et al. The Influence of Substrate and Strain on Protein Quality of Pleurotus ostreatus. Applied Sciences. 2024;14(10). [CrossRef]
- Ng T-B. Protease Inhibitors, Lectins, Antifungal Protein and Saponins in Soybean. In: Ng T-B, ed. Soybean - Biochemistry, Chemistry and Physiology. London: IntechOpen; 2011.
- Sharma NK, Beniwal V, Kumar N, Kumar S, Pathera AK, Ray A. Production of tannase under solid-state fermentation and its application in detannification of guava juice. Prep Biochem Biotechnol. 2014;44(3):281-290. [CrossRef]
- van de Lagemaat J, Pyle DL. Solid-state fermentation: a continuous process for fungal tannase production. Biotechnol Bioeng. 2004;87(7):924-929. [CrossRef]
- Asghar A, Afzaal M, Saeed F, et al. Valorization and food applications of okara (soybean residue): A concurrent review. Food Sci Nutr. 2023;11(7):3631-3640. [CrossRef]
- Terefe ZK, Omwamba MN, Nduko JM. Effect of solid state fermentation on proximate composition, antinutritional factors and in vitro protein digestibility of maize flour. Food Sci Nutr. 2021;9(11):6343-6352. [CrossRef]
- Heidari F. Fungal treatment to enhance the nutritional value of canola meal for animal feed [Thesis]. Minneapolis, MN, University of Minnesota; 2023.
- Rozan P, Villaum C, Bau HM, Schwertz A, Nicolas JP, Méjean L. Detoxication of rapeseed meal by Rhizopus Oligosporus sp-T3: A first step towards rapeseed protein concentrate. International Journal of Food Science & Technology. 1996;31(1):85-90.
- Mageshwaran V, Satankar V, Paul S. Solid-state fermentation for gossypol detoxification and nutritive enrichment of cottonseed cake: A scale-up of batch fermentation process. BioResources. 2024;19(1):1107-1118. [CrossRef]
- Bhanja Dey T, Kuhad RC. Enhanced production and extraction of phenolic compounds from wheat by solid-state fermentation with Rhizopus oryzae RCK2012. Biotechnology Reports. 2014;4:120-127. [CrossRef]
- Wu JY, Wee S, Ler SG, Henry CJ, Gunaratne J. Unraveling the impact of tempeh fermentation on protein nutrients: An in vitro proteomics and peptidomics approach. Food Chemistry. 2025;474:143154. [CrossRef]
- Wiebe MG. Myco-protein from Fusarium venenatum: a well-established product for human consumption. Appl Microbiol Biotechnol. 2002;58(4):421-427. [CrossRef]
- Lonchamp J, Akintoye M, Clegg PS, Euston SR. Sonicated extracts from the Quorn fermentation co-product as oil-lowering emulsifiers and foaming agents. European Food Research and Technology. 2020;246(4):767-780. [CrossRef]
- Yegin S. Solid-state fermentation as a strategy for improvement of bioactive properties of the plant-based food resources. Bioresour Bioprocess. 2025;12(1):140. [CrossRef]
- Petraglia T, Latronico T, Liuzzi GM, Fanigliulo A, Crescenzi A, Rossano R. Hydrolytic Enzymes in the Secretome of the Mushrooms P. eryngii and P. ostreatus: A Comparison Between the Two Species. Molecules. 2025;30(12). [CrossRef]
- El-Gendi H, Saleh AK, Badierah R, Redwan EM, El-Maradny YA, El-Fakharany EM. A Comprehensive Insight into Fungal Enzymes: Structure, Classification, and Their Role in Mankind's Challenges. J Fungi (Basel). 2021;8(1). [CrossRef]
- Alvarez-Cervantes J, Sanchez C, Diaz R, Diaz-Godinez G. Characterization of production of laccases, cellulases and xylanases of Pleurotus ostreatus grown on solid-state fermentation using an inert support. Revista Mexicana de Ingeniería Química. 2016;15(2):323-331. [CrossRef]
- Xu LN, Guo S, Zhang S. Effects of solid-state fermentation with three higher fungi on the total phenol contents and antioxidant properties of diverse cereal grains. FEMS Microbiol Lett. 2018;365(16). [CrossRef]
- Li M, Wang H, Ng TB. Isolation of a phytase with distinctive characteristics from an edible mushroom, Pleurotus eryngii. Protein Pept Lett. 2013;20(4):459-466.
- Hoa HT, Wang CL. The Effects of Temperature and Nutritional Conditions on Mycelium Growth of Two Oyster Mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology. 2015;43(1):14-23. [CrossRef]
- Aditya, Neeraj, Jarial RS, Jarial K, Bhatia JN. Comprehensive review on oyster mushroom species (Agaricomycetes): Morphology, nutrition, cultivation and future aspects. Heliyon. 2024;10(5):e26539. [CrossRef]
- Rathore KS, Pandeya D, Campbell LM, et al. Ultra-Low Gossypol Cottonseed: Selective Gene Silencing Opens Up a Vast Resource of Plant-Based Protein to Improve Human Nutrition. Critical Reviews in Plant Sciences. 2020;39(1):1-29. [CrossRef]
- Zhang Y, Wei R, Azi F, et al. Solid-state fermentation with Rhizopus oligosporus RT-3 enhanced the nutritional properties of soybeans. Frontiers in Nutrition. 2022;Volume 9 - 2022.
- Stodolak B, StarzySka-Janiszewska A, Mickowska B. Effect of Flaxseed Oil-Cake Addition on the Nutritional Value of Grass Pea Tempeh. Food Science and Technology Research. 2013;19(6):1107-1114. [CrossRef]
- Chen L, Song P, Jia F, Wang JS. Reducing the Allergenicity from Food by Microbial Fermentation. Advanced Materials Research. 2012;524-527:2302-2305. [CrossRef]
- Tahmasian A, Drew R, Broadbent JA, Juhász A, Nye-Wood M, Colgrave ML. Conventional solid-state fermentation impacts the white lupin proteome reducing the abundance of allergenic peptides. Food Chemistry. 2023;426:136622. [CrossRef]
- Mattison CP, Dupre RA, Clermont K, Gibbons JG, Yu JH. Proteomic characterization of peanut flour fermented by Rhizopus oryzae. Heliyon. 2024;10(15):e34793. [CrossRef]
- Verhoeckx KCM, Vissers YM, Baumert JL, et al. Food processing and allergenicity. Food Chem Toxicol. 2015;80:223-240. [CrossRef]
- Schmidberger L, Schieberle P. Changes in the Key Aroma Compounds of Raw Shiitake Mushrooms (Lentinula edodes) Induced by Pan-Frying as well as by Rehydration of Dry Mushrooms. Journal of Agricultural and Food Chemistry. 2020. [CrossRef]
- Rahardjo YS, Tramper J, Rinzema A. Modeling conversion and transport phenomena in solid-state fermentation: a review and perspectives. Biotechnol Adv. 2006;24(2):161-179. [CrossRef]
- Handoyo T, Morita N. Hypoallergenic buckwheat flour preparation by Rhizopus oligosporus and its application to soba noodle. Food Research International. 2006;39(5):598-605. [CrossRef]
- Feng XM, Passoth V, Eklund-Jonsson C, Alminger ML, Schnürer J. Rhizopus oligosporus and yeast co-cultivation during barley tempeh fermentation—Nutritional impact and real-time PCR quantification of fungal growth dynamics. Food Microbiology. 2007;24:393-402. [CrossRef]
- Jeleń H, Majcher M, Ginja A, Kuligowski M. Determination of compounds responsible for tempeh aroma. Food chemistry. 2013;141(1):459-465. [CrossRef]
- Amin MNG, Kusnadi J, Hsu J-L, Doerksen RJ, Huang T-C. Identification of a novel umami peptide in tempeh (Indonesian fermented soybean) and its binding mechanism to the umami receptor T1R. Food Chemistry. 2020;333:127411. [CrossRef]
- Zhang Y, Venkitasamy C, Pan Z, Wang W. Recent developments on umami ingredients of edible mushrooms – A review. Trends in Food Science & Technology. 2013;33(2):78-92. [CrossRef]
- Godschalk-Broers L, Sala G, Scholten E. Meat Analogues: Relating Structure to Texture and Sensory Perception. Foods. 2022;11(15). [CrossRef]
- Tao A, Zhang H, Duan J, et al. Mechanism and application of fermentation to remove beany flavor from plant-based meat analogs: A mini review. Frontiers in Microbiology. 2022;13:1070773. [CrossRef]
- Appiani M, Cattaneo C, Laureati M. Sensory properties and consumer acceptance of plant-based meat, dairy, fish and eggs analogs: a systematic review. Frontiers in Sustainable Food Systems. 2023;Volume 7 - 2023.
- Aguilar CN, Rodríguez R, Gutiérrez-Sánchez G, et al. Microbial tannases: advances and perspectives. Appl Microbiol Biotechnol. 2007;76(1):47-59. [CrossRef]
- Saad MM, Saad AM, Hassan HM, Ibrahim EI, Hassabo AA, Ali BA. Bioremoval of tannins and heavy metals using immobilized tannase and biomass of Aspergillus glaucus. Microb Cell Fact. 2024;23(1):209. [CrossRef]
- Cai S, Gao F, Zhang X, et al. Evaluation of γ- aminobutyric acid, phytate and antioxidant activity of tempeh-like fermented oats (Avena sativa L.) prepared with different filamentous fungi. J Food Sci Technol. 2014;51(10):2544-2551. [CrossRef]
- Dekkers BL, Boom RM, van der Goot AJ. Structuring Processes for Meat Analogues. Trends in Food Science & Technology. 2018;81:25-36. [CrossRef]
- Wang Y, Tuccillo F, Lampi AM, et al. Flavor challenges in extruded plant-based meat alternatives: A review. Compr Rev Food Sci Food Saf. 2022;21(3):2898-2929. [CrossRef]
- Landaud S, Helinck S, Bonnarme P. Formation of volatile sulfur compounds by microorganisms in fermented foods: mechanisms and control. Food Microbiology. 2008.
- Mattheis JP, Roberts RG. Identification of geosmin as a volatile metabolite of Penicillium expansum. Appl Environ Microbiol. 1992;58(9):3170-3172. [CrossRef]
- Espinosa-Páez E, Hernández-Luna CE, Longoria-García S, et al. Pleurotus ostreatus: A potential concurrent biotransformation agent/ingredient on development of functional foods (cookies). LWT. 2021;148:111727. [CrossRef]
- Losoya-Sifuentes C, Simões LS, Cruz M, et al. Development and characterization of Pleurotus ostreatus Mushroom—Wheat bread. Starch - Stärke. 2022;74:2100126. [CrossRef]
- Aoyama Y, Kawahara S, Miyake Y. Characterization of kokumi gamma-glutamyl peptides and volatile compounds in misos. Food Research International. 2023;170:112986. [CrossRef]
- Yamaguchi S, Ninomiya K. Umami and Food Palatability. The Journal of Nutrition. 2000;130(4S):921S-926S.
- Ohsu T, Amino Y, Nagasaki H, et al. Involvement of the Calcium-Sensing Receptor in Human Taste Perception of Kokumi Substances. Journal of Agricultural and Food Chemistry. 2010.
- Li Q, Zhang L, Lametsch R. Current progress in kokumi-active peptides, evaluation and preparation methods: a review. Critical Reviews in Food Science and Nutrition. 2022;62(5):1230-1241. [CrossRef]
- Bryant C, Szejda K, Parekh N, Deshpande V, Tse B. A Survey of Consumer Perceptions of Plant-Based and Clean Meat in the USA, India, and China. Frontiers in Sustainable Food Systems. 2019;Volume 3 - 2019.
- Bryant C, Barnett J. Consumer acceptance of cultured meat: A systematic review. Meat Sci. 2018;143:8-17. [CrossRef]
- Siegrist M, Hartmann C. Consumer acceptance of novel food technologies. Nat Food. 2020;1(6):343-350. [CrossRef]
- Siegrist M, Hartmann C. Impact of sustainability perception on consumption of organic meat and meat substitutes. Appetite. 2019;132:196-202. [CrossRef]
- Aaslyng MD, Højer R. Introducing Tempeh as a New Plant-Based Protein Food Item on the Danish Market. Foods. 2021;10(11). [CrossRef]
- Szenderák J, Fróna D, Rákos M. Consumer Acceptance of Plant-Based Meat Substitutes: A Narrative Review. Foods. 2022;11(9). [CrossRef]
- Jang J, Lee D-W. Advancements in plant based meat analogs enhancing sensory and nutritional attributes. npj Science of Food. 2024;8(1):50. [CrossRef]
- da Silva BD, da Costa Marques JM, Conte-Junior CA. The meat mimicry dilemma in the sensory acceptance of plant-based meat analogue: A systematic review. Food and Humanity. 2025;5:100734. [CrossRef]
- Ng GCF, Choy MJY, Tan VWK, et al. Comparative analysis of sensory, textural, microstructural, amino acids and protein digestibility properties of animal and alternative meat products in the Asian market. International Journal of Food Science & Technology. 2024;59:5837-5858.
- Kim A, Öström Å, Mihnea M, Niimi J. Consumers’ Attachment to Meat: Association between Sensory Properties and Preferences for Plant-Based Meat Alternatives. Food Quality and Preference. 2024;116:105134. [CrossRef]
- Cirlincione F, Venturella G, Gargano ML, et al. Functional bread supplemented with Pleurotus eryngii powder: A potential new food for human health. International Journal of Gastronomy and Food Science. 2022;27:100449. [CrossRef]
- Sławińska A, Sołowiej BG, Radzki W, Fornal E. Wheat bread supplemented with Agaricus bisporus powder: Effect on bioactive substances content and technological quality. Foods. 2022;11(23):3786. [CrossRef]
- Ishak W, Azman W, Abu Bakar N. Does oyster mushroom (Pleurotus sajor-caju) powder addition improve nutrient composition, sensory acceptability, and glycaemic index (GI) of flatbread (Tortilla)? Kuwait Journal of Science. 2021;48(2):1-13. [CrossRef]
- Sitanggang AB, Sinaga WSL, Wie F, Fernando F, Krusong W. Enhanced antioxidant activity of okara through solid-state fermentation of GRAS Fungi. Food Science and Technology (Campinas). 2019;40(4). [CrossRef]
- Yasar S, Tosun R, Sonmez Z. Fungal fermentation inducing improved nutritional qualities associated with altered secondary protein structure of soybean meal determined by FTIR spectroscopy. Measurement. 2020;161:107895. [CrossRef]
- Nout MJR, de Dreu MA, Zuurbier AM, Bonants-van Laarhoven TMG. Ecology of controlled soyabean acidification for tempe manufacture. Food Microbiology. 1987;4(2):165-172. [CrossRef]
- Zhang W, Deng Z, Liu T, Liang J, Liu J. Fermentation with edible mushroom mycelia improves flavor characteristics and techno-functionalities of soybean protein. Food Bioscience. 2024;59:104123. [CrossRef]
- Gbenle J, Mert M, Phasha NN, et al. Fungal-mediated solid-state fermentation ameliorates antinutritional factors but does not improve in vitro digestibility of marama (Tylosema esculentum) beans. Future Foods. 2025;11:100664. [CrossRef]
- Pandey A, Soccol CR, Larroche C. Current Developments in Solid-State Fermentation. Berlin, Heidelberg: Springer; 2008.
- Artola A, Font X, Moral-Vico J, Sánchez A. The role of solid-state fermentation to transform existing waste treatment plants based on composting and anaerobic digestion into modern organic waste-based biorefineries, in the framework of circular bioeconomy. Frontiers in Chemical Engineering. 2024;6:1463785. [CrossRef]
- Sánchez C. Reactive oxygen species and antioxidant properties from mushrooms. Synthetic and Systems Biotechnology. 2017;2(1):13-22. [CrossRef]
- Lonnie M, Hooker E, Brunstrom JM, et al. Protein for Life: Review of Optimal Protein Intake, Sustainable Dietary Sources and the Effect on Appetite in Ageing Adults. Nutrients. 2018;10(3). [CrossRef]
- Yuwanita R, Isnansetyo A, Budhiyanti SA, Istiqomah I. Nutritional improvement of Spirulina (Arthrospira platensis) by-product through simultaneous fermentation with Bacillus subtilis (T2A) & (T3P1), and Lactococcus formosensis (JAL 11). Discover Food. 2025;6(1):27.
- Ayllón-Parra N, Castellari M, Gou P, Ribas-Agustí A. Effects of solid-state fermentation with Pleurotus ostreatus on the nutritional and techno-functional properties of alternative protein ingredients. Food Chemistry. 2025;490:145090. [CrossRef]
- Bonilla Loaiza AM, Rodríguez-Jasso RM, Belmares R, et al. Fungal Proteins from Sargassum spp. Using Solid-State Fermentation as a Green Bioprocess Strategy. Molecules. 2022;27(12). [CrossRef]
- Gervais P, Molin P. The role of water in solid-state fermentation. Biochemical Engineering Journal. 2003;13(2):85-101. [CrossRef]
- Mitchell DA, Ruiz HA, Krieger N. A Critical Evaluation of Recent Studies on Packed-Bed Bioreactors for Solid-State Fermentation. Processes. 2023;11(3):872. [CrossRef]
- Soccol CR, Costa ESFd, Letti LAJ, Karp SG, Woiciechowski AL, Vandenberghe LPdS. Recent developments and innovations in solid state fermentation. Biotechnology Research and Innovation. 2017;1(1):52-71. [CrossRef]
- Roenneberg T, Dragovic Z, Merrow M. Demasking biological oscillators: Properties and principles of entrainment exemplified by the Neurospora circadian clock. Proceedings of the National Academy of Sciences. 2005;102(21):7742-7747. [CrossRef]
- Merrow M, Roenneberg T. The circadian cycle: is the whole greater than the sum of its parts? Trends in Genetics. 2001;17(1):4-7. [CrossRef]
- Baker CL, Loros JJ, Dunlap JC. The circadian clock of Neurospora crassa. FEMS Microbiol Rev. 2012;36(1):95-110. [CrossRef]
- Corrochano LM. Light in the Fungal World: From Photoreception to Gene Transcription and Beyond. Annual Review of Genetics. 2019;53(Volume 53, 2019):149-170. [CrossRef]
- Bayram Ö S, Bayram Ö. An Anatomy of Fungal Eye: Fungal Photoreceptors and Signalling Mechanisms. J Fungi (Basel). 2023;9(5). [CrossRef]
- Qi Y, Sun X, Ma L, Wen Q, Qiu L, Shen J. Identification of two Pleurotus ostreatus blue light receptor genes (PoWC-1 and PoWC-2) and in vivo confirmation of complex PoWC-12 formation through yeast two hybrid system. Fungal Biol. 2020;124(1):8-14. [CrossRef]
- Ye D, Du F, Hu Q, Zou Y, Bai X. Transcriptome Analysis Reveals Candidate Genes Involved in Light-Induced Primordium Differentiation in Pleurotus eryngii. International Journal of Molecular Sciences. 2021;23(1):435. [CrossRef]
- Andayani SN, Lioe HN, Wijaya CH, Ogawa M. Umami fractions obtained from water-soluble extracts of red oncom and black oncom—Indonesian fermented soybean and peanut products. Journal of Food Science. 2020;85(3):657-665. [CrossRef]
- Dissasa G. Cultivation of different oyster mushroom (Pleurotus species) on coffee waste and determination of their relative biological efficiency and pectinase enzyme production, Ethiopia. International Journal of Microbiology. 2022. [CrossRef]
- Hultberg M, Asp H, Bergstrand KJ, Golovko O. Production of oyster mushroom (Pleurotus ostreatus) on sawdust supplemented with anaerobic digestate. Waste Management. 2023;155:1-7. [CrossRef]
- Gerber S, Roberts SB. Peanut hulls, an underutilized nutritious culinary ingredient: valorizing food waste for global food, health, and farm economies—a narrative review. Frontiers in Nutrition. 2024;Volume 11 - 2024.
- Chilakamarry CR, Mimi Sakinah AM, Zularisam AW, et al. Advances in solid-state fermentation for bioconversion of agricultural wastes to value-added products: Opportunities and challenges. Bioresource Technology. 2022;343:126065. [CrossRef]
- Erismann Y, Brück WM, Andlauer W. Solid-State Fermentation of Agro-Industrial By-Products. Nutraceuticals. 2025;5(2):11. [CrossRef]
- Navarro ME, Brizuela NS, Flores NE, et al. Preservation of Malolactic Starters of Lactiplantibacillus plantarum Strains Obtained by Solid-State Fermentation on Apple Pomace. Beverages. 2024;10(3):52. [CrossRef]
- FAO. The Impact of Disasters on Agriculture and Food Security 2025. Rome: Food Agriculture Organization of the United, Nations;2025.
- Grzegorczyk M, Kancelista A, Laba W, Piegza M, Witkowska D. The effect of lyophilization and storage time on the survival rate and hydrolytic activity of Trichoderma strains. Folia Microbiol (Praha). 2018;63(4):433-441. [CrossRef]
- Coblentz M, Evans JD, Kothe CI, et al. Food fermentation in space: Opportunities and challenges. iScience. 2025;28(4):112189. [CrossRef]
- Mycsology Foods I, National A, Space A. STTR Phase I: A Discovery Platform to Optimize the Production of Nutrient-Rich Food via Solid-State Fermentation of Feedstocks Relevant to Spaceflight. Washington, DC: NASA Shared Services Center; 2025-09-10 2025.
- Shahid M, Shah P, Mach K, et al. The environmental impact of mycoprotein-based meat alternatives compared to plant-based meat alternatives: A systematic review. Future Foods. 2024;10:100410. [CrossRef]
- Purwandari FA, Fogliano V, Capuano E. Tempeh fermentation improves the nutritional and functional characteristics of Jack beans (Canavalia ensiformis (L.) DC). Food & Function. 2024;15(7):3680-3691. [CrossRef]
- Olagunju LK, Isikhuemhen OS, Dele PA, et al. Effects of the Incubation Period of Pleurotus ostreatus on the Chemical Composition and Nutrient Availability of Solid-State-Fermented Corn Stover. Animals (Basel). 2023;13(16). [CrossRef]
- Watson E. Atlast Food Co: Mycelium scaffold can unlock the ‘Holy Grail’ of plant-based meat: whole cuts. FoodNavigator-USA. 2019.
- FAO. Sustainability Pathways: Livestock and Landscapes. Food Agriculture Organization of the United, Nations;2012.
- Ritchie H, Roser M. Half of the world’s habitable land is used for agriculture. Our World in Data. 2019.
- Bond JK. Corn and Other Feed Grains - Feed Grains Sector at a Glance. US Department of Agriculture, Economic Research Service. 2025.
- Cassidy ES, West PC, Gerber JS, Foley JA. Redefining agricultural yields: from tonnes to people nourished per hectare. Environmental Research Letters. 2013;8(3):034015. [CrossRef]
- Marbaniang EK. Cultivation of Oyster Mushroom: Training Manual. Meghalaya, India: Krishi Vigyan Kendra, West Khasi Hills (ICAR-ATARI Zone VII);2024.
- FAO. Indoor oyster mushroom (Pleurotus spp.) cultivation for livelihood diversification and resilience: Uganda field practice note. Rome: Food Agriculture Organization of the United, Nations;2023.
- FAO. Make money by growing mushrooms. Rome: Food Agriculture Organization of the United, Nations;2004.
- Clark AJ, Soni BK, Sharkey B, et al. Shiitake mycelium fermentation improves digestibility, nutritional value, flavor and functionality of plant proteins. LWT. 2022;156:113065. [CrossRef]
- Machida M, Yamada O, Gomi K. Genomics of Aspergillus oryzae: learning from the history of Koji mold and exploration of its future. DNA Res. 2008;15(4):173-183. [CrossRef]
- Steinkraus K. Industrialization of Indigenous Fermented Foods, Revised and Expanded (2nd ed.). . CRC Press; 2004.
- Center BC. Fermentos: Ciencia y técnica de la fermentación. Planeta Gastro2026.
- Maseko KH, Regnier T, Bartels P, Meiring B. Mushroom mycelia as sustainable alternative proteins for the production of hybrid cell-cultured meat: A review. J Food Sci. 2025;90(2):e70060. [CrossRef]
- Ogawa M, Kermani AS, Huynh MJ, Baar K, Leach JK, Block DE. Edible mycelium as proliferation and differentiation support for anchorage-dependent animal cells in cultivated meat production. npj Science of Food. 2024;8(1):23. [CrossRef]
- Gmoser R, Ferreira JA, Lennartsson PR, Taherzadeh MJ. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol Biotechnol. 2017;4:4. [CrossRef]
- Wu H, Sakai K, Zhang J, McClements DJ. Plant-based meat analogs: color challenges and coloring agents. Food, Nutrition and Health. 2024;1(1):4. [CrossRef]
- Chaudhary V, Katyal P, Poonia AK, Kaur J, Puniya AK, Panwar H. Natural pigment from Monascus: The production and therapeutic significance. Journal of Applied Microbiology. 2022;133(1):18-38. [CrossRef]
- Future Market I. Mycoprotein Market Outlook (2024 to 2034): Demand for Sustainable Protein Sources Drives Growth. Future Market Insights;2024.
- McIntyre L. Section 3.9 Tempeh. In: McIntyre L, Fermented Foods working g, eds. Safety of fermented foods: Assessing risks in fermented food processing practices and advice on how to mitigate them. Environmental Health Services, BC Centre for Disease Control; 2024:5-13.
- Gibbons John G, Salichos L, Slot Jason C, et al. The Evolutionary Imprint of Domestication on Genome Variation and Function of the Filamentous Fungus Aspergillus oryzae. Current Biology. 2012;22(15):1403-1409. [CrossRef]
- Jeong E, Kwon YJ, Seo J-A. Distinction Between Aspergillus oryzae and Aflatoxigenic Aspergillus flavus by Rapid PCR Method Based on the Comparative Sequence Analysis of the Aflatoxin Biosynthesis Gene Cluster. Journal of Fungi. 2026;12(1):10. [CrossRef]
- Yue X, Ren X, Fu J, et al. Characterization and mechanism of aflatoxin degradation by a novel strain of Trichoderma reesei CGMCC3.5218. Front Microbiol. 2022;13:1003039. [CrossRef]
- EFSA Panel on Food Contact Materials E, Processing A, Silano V, et al. Safety evaluation of the food enzyme α-amylase from Aspergillus oryzae (strain DP-Bzb41). EFSA Journal. 2019;17(11):e05899.
- EPA. Framework for Risk Characterization of Environmental Problems. U.S. Environmental Protection Agency; 1997/09/01 1997.
- FDA. FDA Food Ingredient and Packaging Inventories 2025.
- Jia M, Yu X, Xu K, et al. The High-Efficiency Degradation of Multiple Mycotoxins by Lac-W Laccase in the Presence of Mediators. Toxins (Basel). 2024;16(11). [CrossRef]
- Niyigaba T, Küçükgöz K, Kołożyn-Krajewska D, Królikowski T, Trząskowska M. Advances in Fermentation Technology: A Focus on Health and Safety. Applied Sciences. 2025;15(6). [CrossRef]
- Dwipa RP, Maneeboon T, Chuaysrinule C, Mahakarnchanakul W. Interactive Inhibition of Aflatoxigenic Aspergillus flavus and Ochratoxigenic Aspergillus carbonarius by Aspergillus oryzae under Fluctuating Temperatures. Microbiology Research. 2022;13(1):114-127. [CrossRef]
- Watson E. There's more than one way to make meat from mycelium, says MyForest CEO: 'What we're doing is far cheaper than building fermenters'. FoodNavigator-USA. 2022.
- FDA. GRAS Notice No. GRN 000945: Agency Response Letter—Protein from mycelial biomass of Fusarium venenatum. College Park, MD: Center for Food Safety and Applied Nutrition, Office of Food Additive Safety; 2022/04/22 2022.
- FDA. GRN. 1152: Phoenix oyster mushroom (Pleurotus pulmonarius) mycelia biomass. In: FDA, ed2024.
- FDA. Recently Published GRAS Notices and FDA Letters. FDA. 2025.
- FDA. GRAS Notice No. GRN 001152: Agency Response Letter—Pleurotus mycelium biomass. College Park, MD: Center for Food Safety and Applied Nutrition, Office of Food Additive Safety.
- Henchion M, Hayes M, Mullen AM, Fenelon M, Tiwari B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods. 2017;6(7). [CrossRef]
- Qin J, Kurt E, LBassi T, Sa L, Xie D. Biotechnological production of omega-3 fatty acids: current status and future perspectives. Frontiers in Microbiology. 2023;Volume 14 - 2023.
Figure 1.
Conceptual pathway for whole-food solid-state fermentation with culinary fungi and food-system impacts.
Figure 1.
Conceptual pathway for whole-food solid-state fermentation with culinary fungi and food-system impacts.
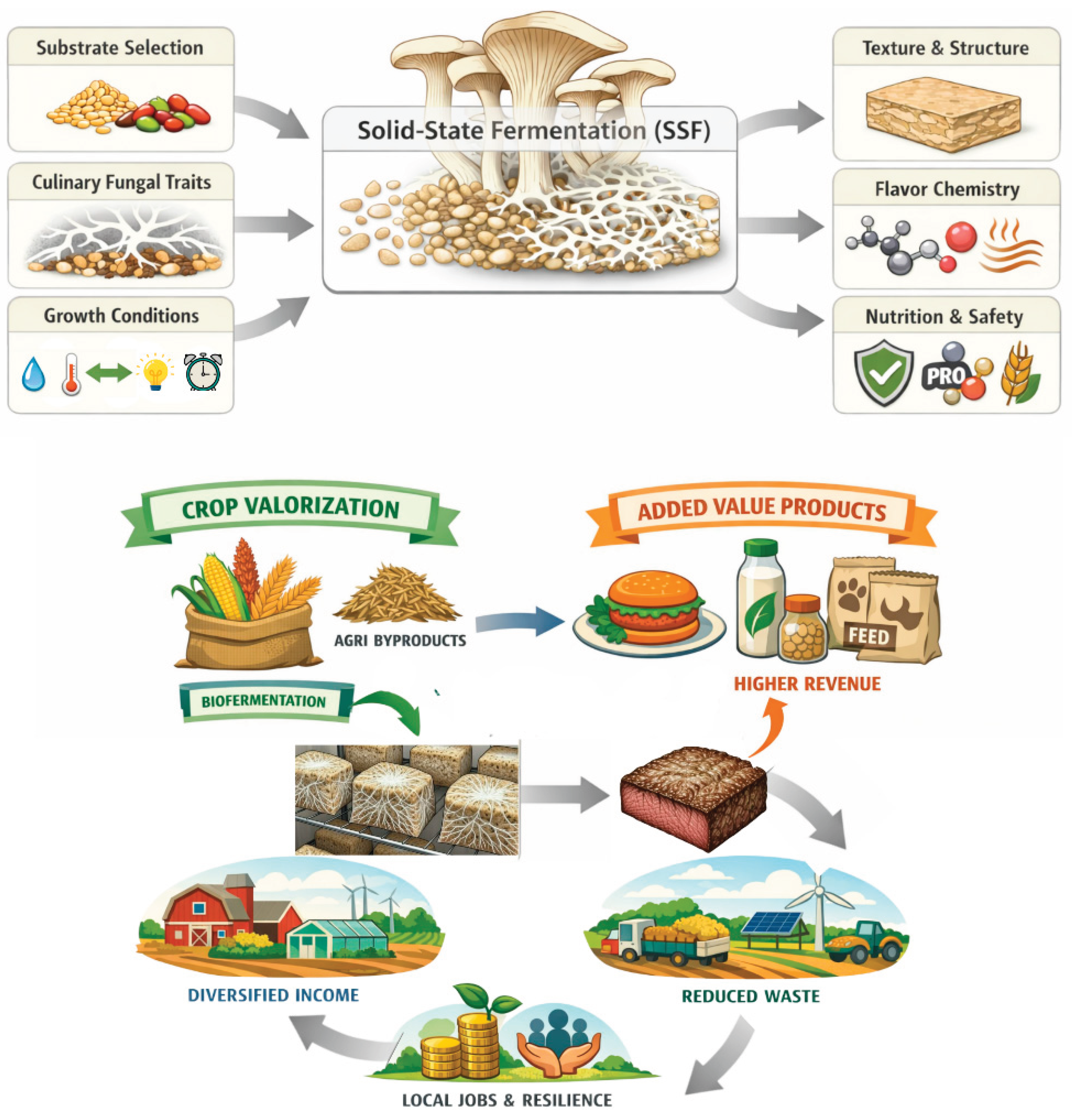

Table 1.
Summary of outcomes from edible-fungi solid-state fermentation (SSF) applied to diverse food substrates. The table compiles representative substrate × microorganism systems and the typical quantitative effects reported for key endpoints. Values reflect magnitudes reported in the cited studies and will vary with strain, inoculum and distribution, substrate composition/particle size, moisture/water activity, temperature, aeration/bed design, and process time; rows should therefore be interpreted as evidence exemplars rather than directly comparable benchmarks. Abbreviations: amino acids (AAs); essential amino acids (EAA,); β-glucans, 1,3/1,6-β-D-glucans; 2,2-diphenyl-1-picrylhydrazyl radical-scavenging (DPPH) assay; guanosine/inosine 5′-monophosphate 9 (5′-GMP/5′-IMP,); polyunsaturated fatty acids (PUFA); α-linolenic acid (ALA); omega-6:omega-3 ratio (ω-6:ω-3); phytic acid (myo-inositol hexakisphosphate; IP6).
Table 1.
Summary of outcomes from edible-fungi solid-state fermentation (SSF) applied to diverse food substrates. The table compiles representative substrate × microorganism systems and the typical quantitative effects reported for key endpoints. Values reflect magnitudes reported in the cited studies and will vary with strain, inoculum and distribution, substrate composition/particle size, moisture/water activity, temperature, aeration/bed design, and process time; rows should therefore be interpreted as evidence exemplars rather than directly comparable benchmarks. Abbreviations: amino acids (AAs); essential amino acids (EAA,); β-glucans, 1,3/1,6-β-D-glucans; 2,2-diphenyl-1-picrylhydrazyl radical-scavenging (DPPH) assay; guanosine/inosine 5′-monophosphate 9 (5′-GMP/5′-IMP,); polyunsaturated fatty acids (PUFA); α-linolenic acid (ALA); omega-6:omega-3 ratio (ω-6:ω-3); phytic acid (myo-inositol hexakisphosphate; IP6).
| Outcome target | Representative SSF system(s) | Quantitative effect examples | Notes/implications | Key sources (year) |
| Protein content and bioavailability Increases | Canola meal × Pleurotus ostreatus (≈12 d); Soybeans × P. ostreatus (long run); Peanut press cake (oncom) × Neurospora sitophila/Rhizopus oligosporus; Okara × R. oligosporus/A. oryzae; Tempeh soy × Rhizopus | Content: +11–18% (canola); +27–28% (soy, 31 d); 52.6–55.35% (peanut press cake, dry basis); Essential AAs of > 12.06 g/100 g; Digestibility increased quality; >50% of soy protein became free amino acids (tempeh) | Mix of concentration effect + de novo fungal protein; enables protein-dense composite foods. Fungal amino acid spectrum complements legumes/cereals Protease/peptidase action. | Heidari 2022; He 2024; Nuramlia 2024; Wijaya 2024 ; Sitanggang 2020; van Veen & Sohaefer 1950; Steinkraus 2004; Wu 2025; Yegin 2025; Kumar 2025 |
| β-glucan Increases | Cottonseed cake + Lathyrus (80:20) × P. ostreatus (11 d); Fusarium venenatum mycoprotein and binding of bile salts | 5× increase in Beta-glucan content; Mycoprotein fiber 24% (DW), 2/3 β-glucans and 1/3 chitin | Adds soluble fiber with immune-metabolic potential55; texture contribution. In vitro digestion of mycoprotein reduced lipolysis and bound bile salts, a mechanism associated with reduced blood cholesterol in humans | Eliopoulos 2024; Ritota & Manzi 2023; Colosimo 2020 |
| Reduced Anti-Nutrients (phytates, glucosinolates, sinapine, gossypol) | Canola meal × P. ostreatus; Oats/Barley × Rhizopus (tempeh-style); Cottonseed + Lathyrus × P. ostreatus | Phytates: −55–76% (canola); oats −74%, barley -89%; Glucosinolates : -98.8%; Sinapine:99.8%; Gossypol: total reduction 60-80%, with some reporting up to −89%, and free gossypol −12.45% | Increases Fe/Zn accessibility; supports bioavailability gains; Major removal of pungent/bitterness drivers; flavor improvement; increased protein digestibility | Heidari 2022; Cai 2014; Handoyo & Morita 2006; Ahnan-Winarno 2021; Eklund-Jonsson 2006; Eklund-Jonsson 2008; Niu 2015; Gbenle 2025; Eliopoulos 2024 |
| Aflatoxin Reduction (contaminated inputs) | Peanut press cake (black oncom) × Neurospora/Rhizopus | −50–70% | Requires tight process control and clean sourcing | Wijaya 2024 |
| Increased antioxidants and phenolics | Soy × P. ostreatus (long SSF); Okara × R. oligosporus/A. oryzae; Oats × Rhizopus; Oats × R. oryzae; Okara × Rh. oligosporus/Asp. oryzae; Soybeans × P. ostreatus | Soy phenolics 4.47×, DPPH 3.92×; Okara +260–550% antioxidant; Oats total phenolics increase | Liberation of bound phenolics; flavor co-benefits of reduced bitterness and astringency; longer shelf-stability | He et al 2024; Sitanggang 2020; Cai 2014; Ahnan-Winarno 2021; Verduzco-Oliva 2020; Chen 2020; Quieroz Santos 2018 |
| Omega fatty acids (ω-3) profile | Grass pea + flax press cake × Rhizopus (tempeh) | ALA increases by >10×; ratio of ω6:ω3 from 11:1 to 0.5–2.5:1 | Co-substrate strategy; retains PUFA while carbs fuel growth | Stodolak 2013; Ahnan-Winarno 2021 |
| Vitamin & Mineral content and absorption increases | Okara (red oncom) × Neurospora intermedia; Barley meal × Rhizopus (tempeh-style) vs. boiled barley | Ca 215 mg/100 g; P 66 mg/100 g; Fe 12.5 mg/100 g; non-heme Fe absorption 5.5% vs. 3.0% (+83% relative); Vitamin B1 raised to 150 µg/100 g | Demonstrates micronutrient density in fermented matrices | Adebo 2020; Asghar 2023; Wijaya 2024; Eklund-Jonsson et al. 2008; van Veen & Sohaefer 1950; Anhan-Winarno 2021; Yegin 2025 |
| Flavor balance improvements | Barley/oats × Rhizopus (with/without yeasts); SSF soy/cereal × Pleurotus/Rhizopus; Brassica meals × Pleurotus; cereal/legume tempeh × Rhizopus | Reduced grassy, beany flavor ratings; increased glutamate, 5′-GMP/IM savory flavor volatiles; increased pyrazines, Strecker aldehydes, 2-acetylpyrrole for cooking step flavor expression; Sensory bitterness falls as sinapine/IP6 decline | Sensory lift via proteolysis + ribonucleotides + Maillard on cooking | Reis 2012; Carrasco-Gonzalez 2017; Feng 2007; Jelen 2013; Handoyo and Morita 2006; Amin 2020; Tao 2022; Zhang 2024; He 2024; Heidari 2022; Eklund-Jonsson 2006 |
| Texture, moisture and cohesion improvements | Cereal/legume SSF flours; Pleurotus (esp. P. eryngii) | higher water holding capacity (WHC); better juice release; elastic, sliceable networks (hyphal entanglement); Dense, anisotropic mycelial mats; Moisture 55–65%; 2–4 mm particle size; O2 diffusion | Enables anisotropic, meat-like bite with minimal texturizing | Godschalk-Broers 2022; Dekkers 2018; Carrasco-González 2017; Ritota & Manzi 2023; He 2024 |
| Allergenicity | Buckwheat soba (tempeh-processing) × Rhizopus; (lupin proteomics in SSF, various); Lupin × Rhizopus; peanut press cake oncom × Neurospora/Rhizopus | Loss of allergen bands; reductions in IgE not always reported; Lupin: β-conglutin peptides reduce; Peanut: IgE binding slightly lower. | Promising but awaiting more research to be clinically validated | Handoyo & Morita 2006; Ahnan-Winarno et al (2021); Tahmasian 2023 |
Table 2.
Anti-nutrients and potentially harmful compounds in SSF feedstocks and fungal biomass, and mitigation pathways enabled by solid-state fermentation (SSF). NOTE: Effects of SSF are substrate-, strain-, and process-dependent. In some matrices, reductions in anti-nutritional factors or increased phytase activity do not necessarily translate into improved in vitro digestibility (Gbenle et al., 2025), and quinoa saponins may increase or decrease depending on processing conditions (Gautheron et al., 2024). Validate key endpoints analytically for the target substrate and product format.
Table 2.
Anti-nutrients and potentially harmful compounds in SSF feedstocks and fungal biomass, and mitigation pathways enabled by solid-state fermentation (SSF). NOTE: Effects of SSF are substrate-, strain-, and process-dependent. In some matrices, reductions in anti-nutritional factors or increased phytase activity do not necessarily translate into improved in vitro digestibility (Gbenle et al., 2025), and quinoa saponins may increase or decrease depending on processing conditions (Gautheron et al., 2024). Validate key endpoints analytically for the target substrate and product format.
| Compound | Foodstuff Source Examples | Effects on Consumption | Example Impacts of Combining Through SSF | References |
| Phytate | Brassica meals, cereals, okara, brans, spent grains, pomace, pseudocereals | Chelates Fe, Zn, Ca, Mg, lowering bioavailability | Fungal phytases (e.g., Rhizopus, Aspergillus, Pleurotus) produce phytases that hydrolyze phytate releasing bound minerals | Pandey 2008; Heidari 2022; Eklund-Jonsson 2006; Eklund-Jonsson 2008; Cai 2014; Sabu 2002; Manikandan 2024; Jatuwong 2020; Zhang 2022 |
| Glucosinolates | Brassica meals (canola, mustard), cruciferous vegetables | Goitrogenic effects, thyroid dysfunction, bitter taste, reduced animal performance | Fermentation with Rhizopus oligosporus, Lactobacillus spp., or Bacillus spp. to hydrolyze glucosinolates into less harmful compounds | Adebo 2022; Heidari 2022; Traka 2016; Zuchowski 2013 |
| Sinapine | Brassicas (Rapeseed, mustard seed, and others) | Bitter, astringent taste; reduced protein digestion; may cause allergic response | Solid-state fermentation with Trametes sp., Rhizopus oligosporus, or Bacillus subtilis degrades sinapine via laccase and other enzymes | Heidari 2022; Ahnan-Winarno 2021; Adebo 2024; Manikandan 2024; Niu 2015 |
| Bound phenolics | Legumes, cereals (wheat), rapeseed, canola, quinoa, fruits, vegetables | Reduced protein/mineral bioavailability, astringency, reduced digestibility | Fermentation with Rhizopus oligosporus, Aspergillus niger, Pleurotus ostreatus, or Lactobacillus plantarum to release bound phenolics | Cai 2014; Sitanggang 2020; Anhan-Winarno 2021; Yasar 2020; Pascual 2025; Verduzco-Oliva 2020; Bhanja Dey 2014; Yegin 2025 |
| Tannins | Legumes, cereals, nuts, tea, fruits, vegetables | Reduced protein digestibility, astringency, toxicity at high intake | Fermentation with tannase-producing fungi (e.g Penicillium glabrum, Aspergillus glaucus, A. niger, Rhizopus sppand others) to hydrolyze tannins | Aguilar 2002; Saad 2024; Górska 2025; Traka, 2016; Górska 2025; Sharma 2014; van de Lagemaat 2004 |
| Protease inhibitors/lectins | Legumes (soybeans, kidney beans, chickpeas), cereals | Inhibit digestive enzymes, reduce protein utilization, cause pancreatic hypertrophy | Fermentation with Rhizopus oligosporus, Aspergillus oryzae, Bacillus subtilis, or protease-secreting fungi to degrade inhibitors/lectins | Friedman 2001; Wolf 1970; Ng 2011; Górska 2025; Adebo 2022; Yasar 2020; Mukherjee 2016; Gbenle 2025 |
| Allergenic proteins (β-conglutin) | Lupin seeds (Lupinus angustifolius, L. mutabilis), fungal/yeast biomass | IgE binding in sensitized individuals and allergenic reactions, immune hypersensitivity, reduce nutrient intake through increased excretion | Proteolysis during SSF reduces specific peptides; Fermentation with Rhizopus oligosporus, Propionibacterium spp., or protease-producing fungi to hydrolyze allergenic proteins (should be validated with immunoassays) | Tahmasian 2023; Verhoeckx 2015; Mattison 2024; Wijaya 2024 |
| Gossypol | Cottonseed meal or cake | Toxicity (liver, reproductive, cardiac), binds lysine | Solid-state fermentation with Candida tropicalis, Saccharomyces cerevisiae, Aspergillus niger, Pleurotus spp., or Paecilomyces variotii | Mageshwaran 2024; Adebo 2022; Eliopoulos 2024; Zhang 2006 |
| Saponins | Legumes (soybeans, chickpeas), quinoa, green microalgae | Bitter taste, hemolytic activity, reduced nutrient absorption | Fermentation with Rhizopus oligosporus, Aspergillus spp., Saccharomyces cerevisiae, or lactic acid bacteria to degrade saponins. SSF can alter saponin profile (sometimes reducing, sometimes increasing saponins); pre-washing/dehulling and post-processing are often required for reliable debittering. | Górska et al., 2025; Traka, 2016; Gautheron 2024 |
| Purines (nucleic acids) | Yeast-rich biomass, fungi, legumes | Uric acid load (hyperuricemia) for gout | Fermentation with Aspergillus oryzae, Blastobotrys adeninivorans, Candida utilis, or low-purine yeast strains; enzymatic degradation. Downstream RNA reduction is described by commercial operators using heat-shock to activate endogenous RNases as a standard mitigation step for fungal biomass intended for high intake. | Kaneko 2014; Adebo 2022; |
| Chitin/Beta-glucans | Fungal biomass, mushrooms, yeasts, microalgae | Reduces apparent protein digestibility in vitro; Associated with indigestion; tough chewing texture; potential allergenicity | Fermentation with chitinase/β-glucanase-producing fungi (Mucor rouxii, Aspergillus terreus, Trichoderma spp.). Milling, alkaline/thermal pretreatments, or targeted enzymatic hydrolysis can further break down to improve digestion. | Yang 2023; Bekirian 2024; Zhang 2013; Kumar 2025; El-Shora 2021 |
| Mycotoxins | Contaminated grains, nuts, by products | Toxicity (carcinogenicity, nephrotoxicity, immunosuppression, reproductive toxicity); carcinogens, hepatotoxicity, immunosuppression | Fermentation with mycotoxin-degrading fungi (Rhizopus spp., a-toxigenic strains of Aspergillus spp.); laccase/peroxidase enzymatic detoxification (lactonases, peroxidases, laccases); Aflatoxin-degrading fungi (Trichoderma reesei, Aspergillus niger, Rhizopus spp.) | Adebo 2022; Wijaya 2024; Yue 2022; Jia 2024 |
Table 3.
Food-grade substrate classes used for solid-state fermentation (SSF) with culinary fungi. Above include observed changes in SSF foods from baseline composition (e.g., protein concentration, phytate/glucosinolate reduction, antioxidant/umami increases), and key formulation levers with representative applied examples. Substrate class boundaries overlap in practice; most edible SSF products are blended matrices designed to optimize C:N ratio, moisture/water activity, and aeration.
Table 3.
Food-grade substrate classes used for solid-state fermentation (SSF) with culinary fungi. Above include observed changes in SSF foods from baseline composition (e.g., protein concentration, phytate/glucosinolate reduction, antioxidant/umami increases), and key formulation levers with representative applied examples. Substrate class boundaries overlap in practice; most edible SSF products are blended matrices designed to optimize C:N ratio, moisture/water activity, and aeration.
| Substrate | Typical examples | Baseline nutrition | SSF Outcomes | Formulation notes | Applied examples |
| Whole cereal grains & pseudocereals | Wheat, rice, maize, barley, oats, millet; buckwheat, quinoa | High starch, moderate protein; bound phenolics & phytate common | Increase protein concentration and/or digestibility; Increase free phenolics/antioxidants; Decrease phytate; added umami/volatiles and other sensory gains via fermentation metabolites | Steam/gelatinize grains or hydrate to support colonization; adjust moisture (commonly ~60–65% wb); control particle size and aeration to modulate moisture; inoculate warm; mill post-SSF for high-protein flour | Whole-grain cereal tempeh fermentation reduced phytate (oats/barley; Rhizopus) (Eklund-Jonsson 2006); barley tempeh increased Fe absorption vs. boiled barley (Eklund-Jonsson 2008); Diverse cereal grains show increased phenolics/antioxidant under (Xu 2018); more grain-fermentation phenolics effects (Adebo 2020). phenolics effects (Adebo 2020). |
| Legumes | Soybeans, chickpeas, lupin, black bean; legume flours | High protein (often 30–50% db), PUFA, protease inhibitors, lectins, phytates; beany notes (bitter phenolics) | Increase protein quality & digestibility (proteolysis);increased free amino acids; decrease in many anti-nutrients; EAA profile improved; increased phenolics and antioxidants including phenol pigments | Balance moisture and oxygen; consider dehulling/cooking to reduce flatulence factors and improve texture; co-blend with cereals when additional carbon is needed | Soybeans × Rhizopus oligosporus SSF (Zhang 2022). Black bean SSF with fungi increased antioxidative activity and phenolics (Lee 2008). White lupin SSF reduced allergenic peptides (proteomics) (Tahmasian 2023). |
| Oilseed, meals and press cakes | Canola/rapeseed meal, cottonseed cake, sunflower, sesame, coconut; okara and other protein-rich byproducts | High protein (~30–50% db), variable lipids; constraints include sinapine/glucosinolates (Brassica) or gossypol (cottonseed); bitterness/astringency | Increased protein concentration and digestibility; reduced brassica anti-nutrients; detoxification of gossypol | Co-ferment with a cereal fraction when C:N is limiting; manage residual oil to avoid hydrophobic zones; verify food-grade sourcing (esp. cottonseed) | Canola meal × Pleurotus ostreatus SSF degraded sinapine and glucosinolates (Heidari M. 2022,). Canola-meal upgrading (Heidari F. 2022). Cottonseed cake + Lathyrus pericarp x P. ostreatus increased β-glucans and decreased gossypol (Eliopoulos 2024). Wheat grains and soybeans × P. ostreatus nutritional improvements (Pascual 2025). |
| Fiber- & bran-rich byproducts | Wheat/rice bran, corn fiber; fruit/veg pomace (apple, grape, pumpkin); spent brewers’ grains | High insoluble fiber, micronutrients; low protein; bound phenols | Increase extractable phenols and antioxidants; partial fiber depolymerization softening fiber matrix; increased moisture holding capacity | Use as a fraction (e.g., 10–40%) blended into cereal/legume/oilseed bases to maintain growth; fine grinding improves uniform colonization; ensure low contaminant load and safe water activity | Wheat SSF × Rhizopus oryzae increased production of phenols (Bhanja Dey 2014). SSF as platform to produce antioxidant polysaccharides (Verduzco-Oliva & Gutierrez-Uribe 2020); cereal-grain antioxidants via SSF across fungi (Xu 2018). |
| Omega-rich seeds & lipid sources | Flaxseed, chia seed; defatted seed meals; oil press cakes | ALA/PUFA, lignans | Can improve ω-3 content and ω6:ω3 ratio in the composite when lipid fraction is blended into a carbohydrate-supporting matrix; retains PUFA while fungus consumes carbs | Pre-mix finely; excess free oil can inhibit if pooled, limit oil to avoid hydrophobic zones; combine with cereal/legume | Grass pea tempeh + flaxseed oil-cake improved nutritional value (including lipid profile) (Stodolak et al. 2013). |
| Microalgae and Aquatic biomass | Chlorella vulgaris (typically blended with cereals); Arthrospira/Spirulina (often with stabilization) | High protein and pigments; bound phenols; sensory off-notes or fishy odors; benefits from a carbohydrate carrier | Increased water retention and gellification/emulsification; broader micronutrient enhancements | Use as minor fraction (e.g., 5–30%) in cereal/legume composite; pre-treatments help support integration | P. ostreatus SSF on oat and Chlorella + oat improved protein solubility (Ayllón-Parra et al., 2025). Arthrospira SSF (Yuwanita 2025). SSF of Sargassum macroalgae with Aspergillus oryzae (Bonilla Loaiza 2022) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



