
Submitted:
29 January 2026
Posted:
31 January 2026
You are already at the latest version
Abstract
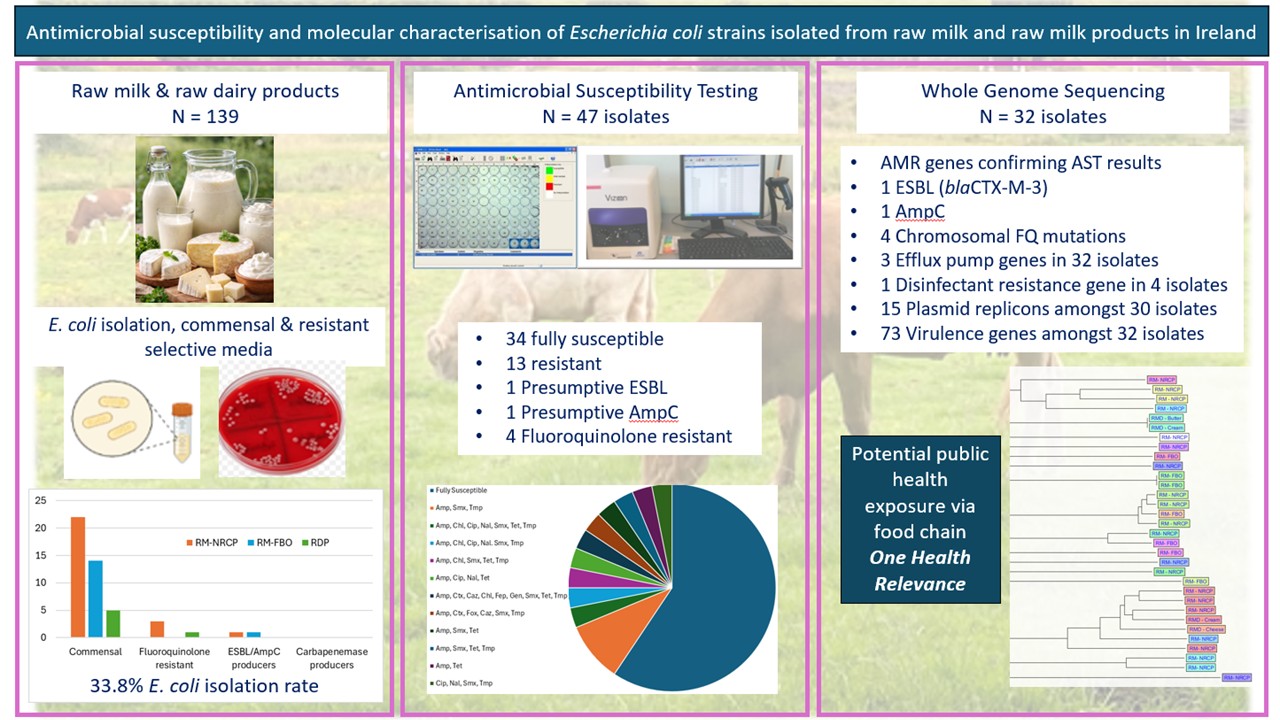
Background/Objectives: Escherichia coli is a ubiquitous commensal organism in humans, animals, and the environment, but certain strains harbour virulence and antimicrobial resistance (AMR) determinants that can cause significant disease. Food-producing animals, including dairy cattle, may act as reservoirs for AMR E. coli, and raw milk and raw milk products can serve as potential exposure pathways to humans. However, data on the prevalence and genomic characteristics of AMR E. coli in raw milk in Ireland are limited. This study aimed to investigate the prevalence of commensal and clinically relevant AMR E. coli in raw milk and raw milk products in Ireland and to characterise their antimicrobial susceptibility and genetic characteristics. Methods: A total of 139 raw milk and raw milk product samples were collected and analysed for commensal E. coli and fluoroquinolone resistant, extended-spectrum β-lactamase (ESBL)/AmpC β-lactamase and carbapenemase producing E. coli. Antimicrobial resistance patterns were determined in line with EU surveillance guidelines based on the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines which use Minimum Inhibitory Concentration (MIC) breakpoints. Whole-genome sequencing (WGS) was conducted on selected isolates to identify AMR genes (ARG), virulence factors, plasmid replicons, efflux pump, disinfectant resistance genes, pathotypes, Multi Locus Sequence Types (MLST) and phylogenetic diversity. Results: Forty-seven E. coli isolates were recovered (33.8% isolation rate). Thirteen isolates exhibited resistance to between two and nine antimicrobials, with ten classified as multidrug resistant (MDR). The highest resistance frequencies were to ampicillin, sulfamethoxazole, trimethoprim and tetracycline. Four fluoroquinolone resistant isolates, one ESBL producer (blaCTX-M-3) and one AmpC promoter mutation were identified; no carbapenemase producers were detected. WGS revealed diverse sequence types, multiple virulence determinants, widespread pervasive efflux pump genes and limited presence of the disinfectant resistance gene qacEΔ1. Conclusions: Raw milk and raw milk products in Ireland can harbour antimicrobial-resistant E. coli, including MDR and potentially pathogenic strains, highlighting the need for ongoing surveillance within the dairy supply chain.
Keywords:
Escherichia coli
; antimicrobial resistance
; raw milk
; dairy products
; multidrug resistance
; ESBL
; AmpC
; fluoroquinolone
; whole genome sequencing
1. Introduction
Most E. coli strains are ubiquitous commensal organisms, existing harmlessly in humans, animals and their environment and seldomly causing disease [1]. However, some strains can be pathogenic due to the presence of specific virulence factors encoded by genes located on the chromosome or mobile genetic elements[1,2]. These pathogens cause a wide range of conditions, from common illnesses like cystitis, urinary tract infections (UTIs), and gastroenteritis to life-threatening extraintestinal infections, including pneumonia, neonatal meningitis, haemorrhagic colitis or haemolytic-uremic syndrome.[3]. Pathogenic E. coli are classified into pathotypes based on their virulence factors and their association with different diseases in humans[4,5,6]. Genes encoding virulence factors confer the potential for mechanisms to cause disease such as adhesion, invasion and toxin production amongst others[2,7,8]. In addition, efflux pump also play an important role in AMR, virulence and disinfectant resistance through their ability to transport and expel a wide range of unwanted material from the bacterial cell[9,10,11,12].
Illnesses caused by pathogenic E. coli require treatment using antibiotics, including last resort antibiotics on those specific cases where an infection is caused by multidrug resistant (MDR) organisms or where all other treatment options have failed[13,14]. Resistance to antimicrobials classified by the World Health Organisation (WHO) as critically important (CIA) for human medicine, is of major concern since infections caused by these organisms are associated with the most adverse health consequences[15]. Extended spectrum β-lactamases (ESBLs) such as SHV, TEM, CTX-M and AmpC β-lactamases, are enzymes that confer resistance to the CIA cephalosporins and have been commonly identified in E. coli from food and food producing animals in Ireland, Europe and worldwide[16,17,18,19]. The spread of MDR or ESBL-producing E. coli due to the misuse and/or overuse of antibiotics in animal and human health as well as the environment is a serious concern[20].
Food-producing animals act as potential reservoirs of AMR bacteria and antimicrobial resistant genes (ARGs), which can be transmitted to other animals and humans through direct contact, the food chain or the environment[21,22,23,24,25,26]. This poses a significant public health threat requiring a one health approach linking human health, animal health and the environment with a strong focus on the emergence and spread of AMR and ARGs[21,23,24]. AMR can be intrinsic, often due to the bacterial cell wall structure, permeability, and the presence of natural efflux pumps that prevent antimicrobial agents from reaching their targets within the cell, but it can also be acquired, due to natural selection, through mechanisms such as horizontal gene transfer, driven by plasmids or due to genomic point mutations[27,28]. Bacteria usually acquire ARGs due to selective pressure exerted through the use and misuse of antimicrobials, resulting in them to be ineffective[20,29]. The use of heavy metals (e.g. copper and zinc) and biocides (e.g. acetic acid and hydrogen peroxide) in food production settings is also known to lead to co-resistance (resistance to several drugs which is linked genetically) through co-selection and co-transfer mechanisms[30,31]. In recent years, especially since the COVID-19 pandemic, non-corrosive biocides such as Quaternary ammonium compounds (QACs) are the most frequently used disinfectant in the food industry with a significant increase in their use[32,33,34,35]. Benzalkonium Chloride (BAC) and Dialkyldimethylammonium Chloride (DDAC) are commonly used QACs in the dairy industry where they are used to disinfect the udder, milking equipment, milk storage tanks and dairy processing machines[36]. However, it has been demonstrated that their overuse can lead to mutation, toxicity, spread of ARGs and therefore lead to increased AMR in bacteria[32,37].
Dairy cows account for a large proportion of Ireland’s cattle herd, with over 1.4 million recorded in December 2024 (Central Statistics Office, 2025)[38]. Mastitis, which is the inflammation of the mammary gland, is the most prevalent dairy cow disease worldwide and the main cause of antimicrobial use in the dairy industry[39]. E. coli has been reported to be one of the main pathogens responsible for mastitis[39,40,41]. While CIA should not be used as a first line of treatment, if milk culture results indicate that there is no alternative treatment or in exceptional circumstances, fluoroquinolones, macrolides and 3rd and 4th generation cephalosporins, may be used[42]. In addition to that, and prior to the introduction of the European Veterinary Medicines Regulation (Regulation (EU) 2019/6)[43] in 2022, treatment of the entire herd with intramammary, long-acting antimicrobials at the end of the lactation and during the dry period was carried out in the majority of Irish herds as a preventative measure[44,45].
According to the National Dairy Council, milk for sale in Ireland is generally pasteurised, however, it is legal to sell raw milk and there is a small sector of the population that consumes raw milk and raw milk products such as butter, cheese and cream[46]. Consumers could potentially acquire AMR and ARGs by consuming raw milk or raw milk dairy products[47,48]. Although there is limited information on the prevalence of AMR E. coli and ARG’s in raw milk and raw milk products in Ireland and Europe, AMR E. coli have been detected in milk from animals with mastitis and from healthy cows[49,50,51,52]. Therefore, the aim of this study was to investigate the prevalence of commensal E. coli and fluoroquinolone resistant, ESBL/AmpC β-lactamase and carbapenemase producing E. coli in raw milk and raw milk products in Ireland to help understand the risks of raw milk as a vehicle for AMR E. coli. AMR was determined using phenotypic methods and a subset of the strains were characterised using Whole Genome sequencing (WGS) to obtain a deeper understanding of the ARGs together with other related or significant genetic determinants, i.e. virulence factors, plasmid replicons, disinfectant resistance, efflux pumps genes, MLST and phylogenetic diversity and pathotype.
2. Materials and Methods
Sample Selection
A total of 139 samples received in our laboratories between March and September 2022 as part of different routine DAFM testing programs were selected for this study: A) 87 raw milk samples from individual dairy farms collected as part of the national residue control plan of Ireland (RM-NRCP). This milk is intended for pasteurisation or further processing. B) 38 raw milk samples from individual food business operators (RM-FBOs) approved for selling raw milk for human consumption. This milk is intended to be consumed raw. C) 14 raw dairy products (RDP) intended for human consumption: 5 butter, 7 cheeses and 2 cream samples.
Isolation of ESBL/AmpC β-Lactamase and Carbapenemase Producing E. coli
Samples were diluted 1:10 in buffered peptone water (BPW; Syntec Scientific, Dublin Ireland) and incubated for 18-22 h at 37oC. ESBL, AmpC and carbapenemase producing E. coli were isolated using the method previously described and recommended by the European Union Reference Laboratory for Antimicrobial Resistance (EURL-AR)[53]. In brief, the enriched BPW was spread plated onto appropriate selective agars i.e., MacConkey plates supplemented with 1 mg/L cefotaxime (E&O laboratories Ltd., Bonnybridge, UK) for ESBL and AmpC producing E. coli and ChromID CARBA and ChromID OXA-48 plates (BioMérieux, Basingstoke, UK) for the selective isolation of carbapenemase producing E. coli. All selective plates were incubated at 37°C for 18-22 h. Presumptive colonies on the chromogenic agars and those that grew up close to the ciprofloxacin disc were purified on the selective media from which they were obtained and on cystine–lactose–electrolyte-deficient (CLED) agar (W12515, Fannin Ltd, Dublin Ireland) as previously described[54].
Isolation of Fluoroquinolone Resistant E. coli
Isolation of fluoroquinolone resistant E. coli was carried out according to the method previously described[54]. In brief, a sterile cotton swab was dipped into enriched BPW and spread over the entire surface of a MacConkey agar plate (E&O laboratories Ltd, Bonnybridge, Scotland, UK.) which was streaked with the same swab after rotating the plate approximately 60 degrees. Immediately after the plate was inoculated, a 5 µg ciprofloxacin paper disk (CT0425B, Fannin Ltd, Dublin Ireland) was placed aseptically onto the middle of the agar plate. Plates were incubated as described above and presumptive colonies were those that grew up close to the ciprofloxacin disc. Presumptive colonies were purified as previously described[54].
Isolation of Commensal E. coli
Commensal E. coli was isolated from raw milk and raw milk products following the standard ISO 16649-2:2001[55]. Milk, cream and butter samples were diluted in maximum recovery diluent (MRD: SKU: KM0040, E&O Laboratories, Scotland, UK) while cheese samples were diluted in 2% tri-sodium citrate solution (Scientific Laboratory Supplies, Rathcoole, Dublin) as previously described[56]. A 1 ml volume of the required dilutions (Milk: 10o and 10-1, cream and butter: 10-1 and cheese: 10-1 and 10-2) was pipetted into the centre of a Petri dish followed by the addition of 15 ml Tryptone Bile X-Glucuronide (TBX) Medium (CM0945B, Fisher Scientific Ltd, Dublin, Ireland) which was tempered to 45 ± 2oC followed by gentle mixing. Once solidified, plates were incubated for 4 h at 37oC followed by 18 to 24 h at 44 ± 1oC.
Positive and Negative Controls
For each batch of samples tested, positive and negative controls were used i.e., E. coli NCTC 13353 (ESBL producer), E. coli NCTC 13476 (carbapenemase resistant) and E. cloacae 03-577qnr-AI (fluoroquinolone resistant). E. coli ATCC 25922 was used as the commensal positive control and as a negative control for the ESBL/AmpC β-lactamase producers and carbapenemase producing E. coli. These control isolates were obtained from the EURL-AR.
Bacterial Species Confirmation
Matrix Assisted Laser Desorption Ionization-Time of Flight (MALDI-TOF) Mass Spectrometry (Bruker Daltronics GmbH, Bremen, Germany) was used to confirm all E. coli isolates as previously described[57]. Confirmed E. coli isolates were maintained at -80oC pending further analyses.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility testing (AST) was carried out by determining the Minimum inhibitory concentration (MIC) using the Sensititre™ EU surveillance Salmonella/E. coli EUVSEC3 AST Plates (Thermo Scientific, MA, USA). A control strain of E. coli ATCC 25922 was included alongside each batch of strains that were tested. In addition, the negative control well was streaked onto blood agar for confirmation of no contamination. The interpretive breakpoints used in this study were those recommended by the EURL-AR and according to the Commission Implementing (EU) Decision 2020/1729[58,59,60]. Presumptive ESBL, AmpC or carbapenemase producing E. coli isolates were tested further with the second panel of antimicrobials, the Sensititre™ EU Surveillance ESBL EUVSEC2 AST plates, which included cefoxitin, cefepime, and clavulanate in combination with cefotaxime and ceftazidime, for the detection of ESBL and AmpC production. EUVSEC2 plates also included imipenem, meropenem and ertapenem to phenotypically verify presumptive carbapenemase producing E. coli (EUCAST 2013)[59]. Depending on the results obtained from the susceptibility testing performed with the second panel, the isolates were subsequently classified as presumptive ESBL, AmpC, ESBL/AmpC or carbapenemase producers according to EUCAST 2013 criteria[59,60]. Classification of presumptive chromosomal or plasmid mediated fluoroquinolone resistance was carried out as previously described[54].
Whole Genome Sequencing Analysis
Selection of Isolates
In total, 33 isolates were selected for WGS, and this included all phenotypically AMR resistant isolates (13) and 20 isolates that were fully susceptible to all antimicrobials.
Culture Preparation and DNA Extraction
E. coli isolates were recovered from frozen stocks on Columbia Agar and Horse blood (E & O Laboratories LTD) and incubated for 24 h at 37°C. A 1 μl loopful of pure culture was taken from the plate and re-suspended in 100 μl nuclease free water. DNA was extracted using the Roche MagNA pure 96 system and quality checked as previously described[17].
Library Preparation, Amplification, Denaturing and Sequencing of Libraries
Sample libraries for all isolates were prepared using the Illumina DNA prep kit (Illumina, Inc., Eindhoven, Netherlands) as per manufacturer’s instructions and using 30 μl of stock DNA to give an input range of 100–500 ng. The library was amplified by 5 PCR cycles with normalisation and denaturing performed as previously described[17]. The molarity of each sample was normalised to 2 nM and the same quantity of each normalised library was pooled to produce a pooled amplified library which was then loaded with 2% PhiX (control library). The isolates included in this study were distributed over four sequencing runs on the NextSeq 2000 (Illumina) using a NextSeq™ 1000/2000 P1 Reagent Cartridge (300 cycles) and NextSeq™ 1000/2000 P1 Flow Cell.
Data Analysis
Statistical Analysis
Using Epitools[61], a Chi-square test from cross tabulation of the raw data was employed to determine whether there was an association between RM-NRCP or RM-FBO recovery of commensal E. coli. Contingency tables were analysed, and results were considered statistically significant when p < 0.05. Due to the low sample numbers of butters, cheeses and creams that were received, statistical analysis was not performed on RDP.
Analysis of WGS Data
The following run metrics were used to check that each WGS run met the basic quality metrics requirements for raw sequence data: >70% bases higher than Q30 at 2 × 300 bp and cluster density of 800 -1200 k/mm2. In addition, the percentage of reads that aligned to the PhiX was also checked to ensure that the starting concentration of the libraries were not over or under-estimated.
MBioSEQ Ridom Typer software (formerly known as SeqSphere), version 10.04, 2024-07 (Ridom GmbH, Münster, Germany) was used to process and analyse WGS data, i.e., quality trimming, de novo assembly, quality assessment and analysis of WGS data. Raw reads (FASTQ) were automatically trimmed prior to assembly using the default settings for trimming i.e., trimmed at both ends until the average base quality was >30 in a window of bases. Paired-end raw read data were assembled using the SPAdes assembler’s algorithm (V3.15.4) set at the automatic mode to determine the coverage cut-off and minimum length of contigs. De novo assemblies with >30x coverage, 5.0 Mb ± 10% bp in size, an N50 ≥ 15,000 and core percentage of ≥95% were included in the downstream WGS analysis. Species confirmation, MLST using the Warwick scheme and core genome MLST (cgMLST) from assembled E. coli genomes was conducted using SeqSphere+. Additionally, the integrated NCBI AMRFinderPlus (AMR Gene Finder plus) software[62,63,64] and virulence factor database[65] (VFDB) were used to identify AMR and virulence, respectively. Furthermore, genes related to disinfectants and general efflux were also identified within AMRFinderPlus. In silico phenotyping was performed using the freely available online ClermonTyper, based on an In silico quadruplex PCR targeting the genes arpA, chuA, yjaA and TspE4.C2 and is accessible at http://clermontyping.iame-research.center/. Each FASTA file was uploaded, as input and the species/phylogroup was determined in two distinct ways: by in silico PCR and Mash. Mash is the species phylogroup of the nearest genome[66]. The presence or absence of the different amplicons constituted a profile that allowed for phylogroup assignment.
CGE tools (Center for Genomic Epidemiology) available at http://cge.cbs.dtu.dk/services, were used for further analysis of assembled data, i.e., in silico detection of plasmid replicons using PlasmidFinder 2.1[67,68] while PathogenFinder 1.1[69] was used to predict the potential of each E. coli isolate to be a human pathogen by uploading the FASTA file and choosing the proteobacteria as the phylum and gamma as the class. The number and type (pathogen specific or non-pathogen specific) of query strain orthologues in the protein family database are used to assign a human pathogen probability score (range 0.0 – 1.0), where a probability score > 0.5 is considered as the threshold for consideration as a human pathogen.
3. Results
Bacterial Culture
Forty-seven E. coli isolates were recovered from the 139 milk and dairy product samples tested (33.8% isolation rate) and this is summarised in Table 1. The percentage of commensal E. coli positive samples amongst the sample types was 29.9%, 39.5% and 42.9% from RM-NRCP, RM-FBO and RDP respectively. Although a higher proportion of commensal E. coli were recovered from RM-FBO samples compared to RM-NRCP, Chi-Squared analysis indicated that the difference was not significant (P > 0.05). In addition, four isolates were recovered from the MacConkey agar plates with the ciprofloxacin paper disk (3 RM-NRCP and one RDP from cream), and two isolates (RM-FBO and RM-NRCP) were isolated from MacConkey plates with cefotaxime. No presumptive carbapenemase producing E. coli were isolated from any of the samples tested.

Table 1.
E. coli isolated from raw milk and raw milk dairy products.
 |
Antimicrobial Susceptibility Testing
AST resulted in 34 isolates found fully susceptible to all antimicrobials tested and 13 isolates resistant to between two and nine antimicrobials (Table 2). The majority of resistant isolates were multidrug resistant (MDR) with ten of the 13 resistant isolates displaying resistance to at least one antimicrobial in three or more antimicrobial classes (Table 2). Ampicillin resistance was the highest (25.3%), followed by sulfamethoxazole (23.4%), trimethoprim (21.3%) and tetracycline (14.9%). Resistance levels were moderate for ciprofloxacin and nalidixic acid (8.5%) and chloramphenicol (8.5%) and only one isolate was resistant to gentamicin (2.1%). None of the isolates were resistant to colistin (polymyxin) or meropenem (carbapenem). Amongst the two presumptive ESBL/AmpC β-lactamase producers, the E. coli isolated from RM-NRCP was found to be a presumptive AmpC while the isolate from RM-FBO was found to be a presumptive ESBL producer when the EU Surveillance ESBL EUVSEC2 AST plate was used. The four presumptive fluoroquinolone isolates were indicative of phenotypic AMR chromosomal fluroquinolone resistance i.e., resistant to both ciprofloxacin and nalidixic acid[27].
Whole Genome Sequencing
Following WGS, one of the 33 sequenced genomes failed the quality metrics from the raw sequencing data. This was an isolate which was fully susceptible and was subsequently excluded from the WGS analysis. Over each run on the Miseq, the index reads were evenly distributed.
The four presumptive chromosomal fluoroquinolones were confirmed genotypically and three of the four displayed double mutations in gyrA and two in parC/parE (Table 3). The most frequent mutations were observed in codon 83 of gyrA and codon 80 of parC gene. The presumptive fluoroquinolone E. coli recovered from RDP, harboured a single mutation in codon 83 of gryA (Table 3). The presumptive AmpC β-lactamase producer encoded the upregulated AmpC promotor at position 42 (C-42T mutation), and harboured the blaTEM-1 gene, while the presumptive ESBL producer harboured the gene blaCTX-M-3 and was also phenotypically resistant to Gentamicin which was confirmed genotypically through identification of the ant(2'')-Ia gene (Table 3). Other aminoglycoside resistant genes responsible for encoding resistance to kanamycin and streptomycin were identified amongst the isolates, although none of them were phenotypically resistant to either kanamycin or streptomycin.
Ampicillin resistance was observed in 12 isolates which included the ESBL and AmpC isolates. BlaTEM-1 was identified in eight of the ampicillin-resistant commensal E. coli isolates, while blaTEM-30, blaTEM-39 in combination with blaOXA-1 and blaOXA-1, were identified in the other three ampicillin-resistant commensal E. coli isolates. Finally, four isolates resistant to chloramphenicol harboured the floR gene, seven isolates resistant to tetracyclines harboured the tet(A) or tet(B) genes, while 10 ten isolates resistant to both trimethoprim and sulfamethoxazole harboured the dfrA and sul1/sul2 genes (Table 3).
Efflux pump genes were present in all 32 E. coli genomes i.e., both fully susceptible and r isolates. Three efflux pump genes were identified i.e., the acrF, mdtM and emrE genes, either alone or in combination, in 29, 23 and 17 genomes, respectively (Table 3).
The resistance gene qacEΔ1, which confers resistance to disinfectants was identified in four of the sequenced E. coli isolates (Table 3). Three of these isolates were from RM-NRCP and two of these were fluoroquinolone resistant. The ESBL isolate from RM-FBO also harboured the qacEΔ1 gene.
Fifteen plasmid replicons were identified in 30 E. coli isolates i.e., IncFIB(AP001918), IncFII, IncFIA, IncFIC(FII), IncFIB(pB171), IncI1-I(Alpha), IncFII(pCoo), IncFII(29), IncX1, IncFII(pSE11), IncFII(pHN7A8), IncY, IncI2(Delta), ColpVC and IncX4, present respectively in 21, nine, seven, seven, four, four, four, two, and one of each of the rest of isolates. Twelve of the 13 AMR isolates harboured at least two plasmid replicons and the total number of plasmid replicons identified amongst all 12 isolates was six with IncFIB(AP001918) and IncFII, identified in 85% and 69% of resistant E. coli. The only resistant isolate without detectable plasmid replicons was the fluoroquinolone-resistant E. coli isolate recovered from RMD. Similarly, 18 of the 19 isolates that were fully susceptible harboured a total of 14 plasmid replicons, including major plasmid replicons such as IncFIB(AP001918), IncFIA, and IncFII. IncFIB(AP001918) was present in 53% of the fully susceptible isolates.
All 32 sequenced E. coli isolates were predicted to be human pathogens with an average probability score of 0.91 (range 0.84 to 0.96) as shown in Table 3. The strains with the highest average probability scores, i.e., 0.96, were the ESBL isolated from RM-FBO and the fluoroquinolone resistant isolate from RMD. The least probability score (0.84) was identified in an isolate from RM-NRCP which was fully susceptible. The genome sequences of the E. coli matched pathogenic families with a range of 53 to 755 per isolate i.e., 53 pathogenic families matched a fluoroquinolone resistant isolate from RM-NRCP, and 755 pathogenic families matched a fully susceptible isolate from RM-FBO (Table 3). In silico phylogrouping permitted the classification of E. coli isolates into four of the seven main phylogroups i.e., A, B1, C and E, in ten, seventeen, four and one isolate, respectively. Two of the fluoroquinolone resistant isolates were assigned to phylogroup A, whereas the other two were assigned into phylogroups B1 and C. The ESBL and AmpC producing isolates were assigned to phylogroups B1 and C, respectively (Table 3). There was 100% concordance between the in-silico PCR assay and the Mash genome-clustering tool.
In silico MLST typing yielded 22 different STs amongst the 32 E. coli, with ST10 the most common followed by ST58 identified in five and three isolates respectively. One isolate of unknown ST was also identified. The distribution of virulence genes amongst the 22 ST’s of E. coli isolates is shown in Figure 1. Major virulence determinants were identified in the sequenced E. coli isolates, including shiga-toxin producing (stx2A and stx2B), the cytolethal distending toxin operon (cdtABC), α-hemolysin (hlyA, hlyB, hlyD), the gene encoding arylsulfatase A enzyme (aslA) Cytotoxic Necrotizing Factor 1 (cnf1), EAST1 (astA/east1), high-affinity iron-acquisition systems (entB–E, entS, fepA–D/G, fes), iron uptake including the salmochelin siderophore system (iroBCDEN), the 8-gene cluster (chu genes) involved in heme transport and processing (chuA, chuS-Y), the type III secretion system (T3SS) effector genes (espL1, espL4, espP, espR1, espX1, espX4, espX5, espY1, espY2, espY3, espY4). Multiple fep genes encoding ferric enterobactin transport functions (fepA, fepB, fepC, fepD, fepG) were identified in all but one of the E. coli isolates, as was the ompA gene which affects adhesion and plays a key role in the resistance of bacteria to antibiotics. Genes involved in aerobactin biosynthesis (iucA–D, iutA), the outer membrane hemin receptor for heme uptake (chuA–Y) and key adherence factors such as fdeC, type 1 fimbriae (fimA–H), P fimbriae (papA–K, papG), sfaX and associated regulators were also identified. In general, all isolates within each ST displayed the same virulence profile. Amongst all 32 isolates, one isolate of ST10 (identified as fluoroquinolone resistant), harboured two stx2 gene toxins (stx2A and stx2B). The aggregative heat-stable enterotoxin 1 (EAST1) was identified in six isolates of ST10, ST58, ST88, ST410, ST8842 and ST17585 (Figure 1).
The neighbor joining phylogenetic tree (NJT) showed a wide range of diversity with 30 different MLST’s observed amongst the 32 E. coli isolates (
Figure 2
). Two clusters were identified, one of ST101 which included two isolates that were one allele different through cgMLST, and both were isolated from RM-FBO and both fully susceptible. The second cluster included two isolates of ST2539 isolated from RMD, both were indistinguishable to each other, and one was isolated from butter and the other from cream (
Figure 2
).
4. Discussion
Although raw milk and raw milk products are appealing to some consumers as a nutritious alternative to heat treated products, the lack of a thermal treatment poses a risk. Raw milk is not included in the AMR EU surveillance programme under Commission Implementing Decision (EU) 2020/1729[70] and to the authors’ knowledge, no previous work has examined AMR, virulence factors, plasmid replicons or disinfectant-resistance genes in E. coli from Irish raw milk. Only limited food safety data exist from the past two decades for raw milk in Ireland; notably, our group reported shiga toxin-producing E. coli (STEC) in 14% of raw milk filters and 1% of raw milk in 2017-2019[71]. In addition, the Food Safety Authority of Ireland (FSAI) reported STEC in 6% of raw milk filters in 2012–2013[72], while a study conducted between 2001 and 2003 detected E. coli O157 in 12.5% of milk filters sampled[73].
The consumption of raw milk and raw-milk cheeses in Europe represents a recognised public-health concern due to frequent contamination with E. coli, including STEC and AMR strains[74,75,76,77]. European studies consistently report E. coli in raw milk at variable but often significant levels (approximately 2–22%), while raw milk cheeses, particularly artisanal products, may show even higher levels[74,75,77,78]. Importantly, a substantial proportion of isolates display AMR, with MDR phenotypes reported in 27-40% of strains and commonly resistant to β-lactams, tetracyclines, aminoglycosides, and fluoroquinolones[76,77,79,80]. The repeated detection of virulent and resistant E. coli in unpasteurised dairy products, together with documented STEC outbreaks linked to raw milk cheeses in several European countries, underscores the dual risk of foodborne infection and dissemination of AMR via the raw milk food chain[81,82].
Our study identified E. coli that were MDR, and contained virulence, pathogenic, and disinfectant-resistance genes in raw milk and raw milk products. This reinforces the need for maintaining high farm hygiene measures, as E. coli levels in milk are known to reflect cleaning efficacy of milking equipment, and bulk milk storage temperatures[63,83,84]. Approximately one fourth of the E. coli isolates in our study (27.7%) exhibited AMR and 21% of these were classified as MDR. To the best of the authors knowledge, there is no previous research on AMR in raw milk and their products in Ireland. These results could therefore serve as a baseline for future research in Ireland. Encouragingly no carbapenemase producing E. coli were detected. The lower proportion of resistant isolates in raw milk destined for direct consumption, compared with milk intended for pasteurisation, may reflect greater compliance with the relevant hygiene requirements laid down in Annexes I and II of Regulation (EC) 852/2004[85] as well as those set out in Annex III (Section IX) of Regulation (EC) No 853/2004[86].
The sequencing results of this study identified a variety of resistance determinants in a subset of the isolates, while the efflux pump genes acrF, mdtM, and emrE were detected either alone or in combination in all 32 isolates. AcrF, for which encoding genes were found in all but three of the sequenced isolates, is a member of the Resistance Nodulation and cell Division (RND) family and represents one of the seven known RND transporters in E. coli. Through the tripartite efflux AcrEF-TolC system[87], it contributes to the extrusion of a broad array of hydrophobic and amphiphilic compounds, including a wide range of antimicrobials and biocides, although non-functional variants have also been described[88]. Genes encoding EmrE, which extrudes hydrophobic compounds[89,90] and belongs to the Small Multidrug Resistant (SMR) family, and MdtM, a Major Facilitator Superfamily (MFS) antiporter that actively expels neutral and cationic compounds including QACs[91,92,93], were also found in more than half of the isolates. This further highlights the potential of raw milk isolates to expel structurally diverse toxic agents, including polyaromatic and cationic compounds commonly encountered in clinical and environmental contexts[94]. The fact that all sequenced isolates harboured efflux pump genes, regardless of their antimicrobial susceptibility profiles, seems to indicate that efflux pump genes presence alone could not explain variations in antibiotic susceptibility. The coexistence of qacEΔ1 with efflux systems identified in four isolates is expected to likely enhance survival under disinfectant pressure and raises concerns regarding co-selection between biocides and antimicrobials[30,95,96,97].
Previously reported research showed that efflux pump mechanisms are widespread in E. coli but do not necessarily confer QAC tolerance unless induced under specific conditions[98,99]. Moreover, recent experimental work demonstrated that E. coli may develop increased tolerance to benzalkonium chloride through adaptive mechanisms independent of integron encoded qac genes[100]. Our findings highlight the need for follow-up phenotypic assays, for example QAC specific MIC testing, time-kill studies, and biofilm tolerance assessments investigations on the E. coli isolates carrying qacEΔ1, to determine whether this gene contributes to measurable reductions in QAC susceptibility. This is because although qacEΔ1 is frequently embedded within class 1 integrons, which are mobile genetic elements (MGE), its association with phenotypic QAC tolerance has often been reported to be weak and inconsistent [101,102,103]. Genetic detection alone cannot reliably predict disinfectant resistance, underscoring the importance of phenotypic validation[104,105]. Genes conferring resistance to other biocides, such as chlorhexidine, hydrogen peroxide, ethanol, isopropanol, peracetic acid, povidone-iodine and sodium hypochlorite[106] were not identified in this present study.
The widespread detection of plasmid replicons in both resistant and fully susceptible E. coli indicates that plasmid carriage is common in dairy-associated commensal populations and is not solely driven by AMR[107]. The high prevalence of plasmid replicons in both resistant and susceptible isolates, particularly IncF plasmids such as IncFIB(AP001918) and IncFII, seems to indicate that these lineages may serve as stable hosts for MGEs capable of acquiring resistance genes under selective pressure, even in the absence of phenotypic resistance[107,108,109]. Notably, IncFIB(AP001918), which is frequently associated with ESBL-producing E. coli in human infections, was also detected in a substantial proportion of fully susceptible isolates, suggesting that clinically relevant plasmids can persist in the raw milk microbiota in the absence of detectable resistance phenotypes[68,110]. The predominance of IncF plasmid replicons, particularly IncFIB(AP001918) and IncFII, among AMR isolates supports their established role as major vectors for resistance gene dissemination in E. coli[109]. Collectively, these findings underscore the importance of including plasmid surveillance in microbiological assessments of raw milk and raw milk products, as such foods may contribute to the silent maintenance and spread of resistance associated plasmids along the food chain[108]. IncL/M plasmid replicons, common in European outbreaks of ESBL [111] and NDM-1 carbapenemase producers [112,113], were not detected. However, the reference based approach using PlasmidFinder which depends on a precompiled database struggles to capture large plasmids greater than 50 kb[114] and short-read sequencing in this present study limited full plasmid reconstruction and characterisation of plasmid structures[115,116]. Chromosomal fluoroquinolone resistance was confirmed through mutations in gyrA and parC, which reduce susceptibility and can increase efflux pump expression[117,118,119]. Among the four fluoroquinolone-resistant isolates in this study, three encoded all three efflux pumps identified, while one encoded AcrF alone.
The co-occurrence of chromosomal AMR determinants, efflux pumps, plasmid replicons, virulence factors, and biocide-resistance genes in raw milk isolates reflects strong and overlapping selective pressures within dairy production environments[120,121]. Antimicrobial use in dairy cattle, particularly for the treatment of mastitis, can enrich intrinsic resistance and efflux-mediated tolerance, contributing to the selection of resistant bacterial populations[122]. Manure and slurry can further disseminate resistant bacteria and antimicrobial residues sustaining environmental selection and contaminating soil and water[123,124,125]. Routine exposure to biocides, heavy metals, and other environmental contaminants can promote co-selection through efflux activation and membrane adaptation[30]. Environmental pathways, including contaminated water, biofilms on milking surfaces, and movement of animals or personnel, facilitate ongoing circulation and persistence of resistant strains[126]. Collectively, these pressures allow chromosomal resistance traits to persist and spread independently of plasmid mediated exchange, highlighting the need for an integrated approach towards antimicrobial use, rigid hygiene measures at both farm and milk processing levels, and environmental management practices given that soil antibiotic resistomes are widespread and connected to human and environmental AMR reservoirs[127,128,129].
This present study revealed ST10 to be the most prevalent lineage, with all ST10 isolates belonging to phylogroup A, a human-associated lineage that has been linked to the dissemination of AMR via plasmids and other mobile genetic elements[107]. ST58 was the second most common and belonged to phylogroup B1, an emerging zoonotic lineage increasingly associated with MDR, ESBL production, and extraintestinal infections[68,109,110,130]. Among the isolates, one ST10 was MDR and fluoroquinolone resistant, while all three ST58 isolates were resistant to at least three antimicrobial classes. Apart from one fluoroquinolone isolate (ST10) that was recovered from cream, all remaining ST10 and ST58 isolates were obtained from raw milk, indicating that these lineages may serve as vehicles for resistance determinants along the dairy supply chain[68,109,130]. Additional resistant lineages included an AmpC β-lactamase-producing ST88 isolate of phylogroup C, an ESBL-producing ST1126 of phylogroup B1, and three more fluoroquinolone resistant isolates within phylogroups C (two isolates), and B1, demonstrating that AMR in raw milk-associated E. coli occurs across multiple phylogenetic backgrounds[68,109,110,130]. While no direct evidence of transmission to humans was assessed in this study, the presence of potentially high-risk lineages highlights the importance of continued surveillance of the dairy food chain within a One Health framework, encompassing human, animal, and environmental reservoirs to better understand and mitigate dissemination of strains of public health relevance.
Amongst the virulence genes identified, some were associated with functions including stress, survival, regulation, iron uptake, type-three secretion systems, invasion, adherence, colonisation and toxin production. The identification of major virulence determinants in the sequenced E. coli isolates highlights virulence profiles with public health relevance. Toxins such as Shiga-toxin are strongly linked to haemorrhagic colitis and haemolytic uremic syndrome, representing one of the most critical food and waterborne threats[131,132]. Likewise, cdtABC, hlyA, and cnf1 can contribute to epithelial damage, systemic inflammation, and severe extraintestinal infections, including urosepsis and neonatal meningitis[133,134,135] while it has also recently been reported that CNF1 promotes colorectal carcinogenesis[136]. Iron-acquisition systems including enterobactin, salmochelin, aerobactin, and heme-uptake pathways are key fitness determinants in host environments and are strongly associated with high-risk extraintestinal pathogenic E. coli lineages[137]. Adherence factors type 1 and P fimbriae enhance colonisation of the urinary and gastrointestinal tracts and facilitate progression to invasive disease[138,139]. In this study 30 isolates carried the fimA to fimH type 1 fimbrial genes, which encode adhesin proteins that are key contributors to biofilm formation and bacterial adherence[140]. The coexistence of these elements may enhance the ability of strains to persist in the environment, adapt to different hosts and contribute to the spread of AMR, highlighting the interconnected risks across human, animal and environmental health. This co-existence could not only limit therapeutic options but also increase the likelihood of horizontal gene transfer, environmental persistence, and foodborne transmission, underscoring the need for a comprehensive, up to date surveillance across the Irish dairy sector[141].
In conclusion, to the best of our knowledge, this is the first study to characterize AMR, STs, virulence, plasmid replicons, and disinfectant-resistance determinants in E. coli from raw milk and raw milk products in the Republic of Ireland. Although the overall prevalence of AMR was moderate, the detection of MDR isolates, ESBL/AmpC producers, high-risk sequence types (ST10, ST58), and Shiga-toxin genes indicate that raw milk can act as a vehicle for transmission of clinically significant E. coli through the food chain. The identification of isolates harbouring genes encoding for QAC-resistance highlights the potential selective effects of biocide use, which is not routinely monitored. Strengthening surveillance efforts, optimising hygiene practices and ensuring prudent use of antibiotics and biocides are therefore recommended to mitigate the emergence and spread of resistant and pathogenic E. coli in the dairy processing environment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Accession numbers along with quality metrics of assemblies are detailed in supplementary Table 1. The de novo assemblies consisted of an average of 299 contigs with an average N50 of 148,889 bp. The average coverage and genome size of the assembled reads were 148X and 5.00 bp, respectively.
Author Contributions
Conceptualization, M.G. and D.P.; methodology, M.T., D.K., S.H., G.M. and S.M.; software, D.P.; formal analysis, D.P. and M.G., investigation, D.P., M.T., D.K. G.M. and M.G., resources, G.M.; data curation, D.P., M.T., G.M. and M.G. writing—original draft preparation, D.P.; writing—review and editing, D.P, and M.G., visualization, D.P. and M.G., funding acquisition, M.G., supervision, G.M, D.P. and A.N.J. All authors have read and approved the final version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Sequencing data produced in this study were deposited to the NCBI Sequence Read Archive (SRA) repository and are available through the BioProject accession number PRJNA1415327.
Acknowledgments
We thank Brian Byrne and Kenneth Dygico for their assistance with the use of the Bruker MBioSEQ Ridom Typer software for WGS analysis. We are also grateful to Rebecca Cupial for her role in coordinating the WGS runs. Additional thanks to Elvira Ramovic or her help with uploading WGS data to NCBI.:
Conflicts of interest
The authors declare no conflicts of interest.
References
- Russo, T. A.; Johnson, J. R. Medical and economic impact of extraintestinal infections due to Escherichia coli: focus on an increasingly important endemic problem. Microbes Infect 2003, 5, 449–56. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W. M.; Rossen, J. W. A. Virulence Factors of Enteric Pathogenic Escherichia coli: A Review. Int J Mol Sci 2021, 22, 9922. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.; Tainter, C. R. Escherichia coli Infection. Available on line europepmc.org/books/n/statpearls/article-21320/?extid=31082016&src=med. 2025. (Accessed on: 15 January 2026).
- Naidoo, N.; Zishiri, O. T. Presence, Pathogenicity, Antibiotic Resistance, and Virulence Factors of Escherichia coli: A Review. Bacteria 2025, 4, 16. [Google Scholar] [CrossRef]
- Shpigel, N. Y.; Elazar, S.; Rosenshine, I. Mammary pathogenic Escherichia coli. Current Opinion in Microbiology 2008, 11, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Alhadlaq, M. A.; Aljurayyad, O. I.; Almansour, A.; Al-Akeel, S. I.; Alzahrani, K. O.; Alsalman, S. A.; Yahya, R.; Al-Hindi, R. R.; Hakami, M. A.; Alshahrani, S. D.; Alhumeed, N. A.; Al Moneea, A. M.; Al-Seghayer, M. S.; AlHarbi, A. L.; Al-Reshoodi, F. M.; Alajel, S. Overview of pathogenic Escherichia coli, with a focus on Shiga toxin-producing serotypes, global outbreaks (1982–2024) and food safety criteria. Gut Pathogens 2024, 16, 57. [Google Scholar] [CrossRef]
- Chatterjee, S. Chapter 9 - Genetic origins of microbial virulence. In Host Adaptation, Virulence, and Evolution; Parray, J. A., Singh, N., Li, W.-J., Eds.; Academic Press, 2025; pp. pp 153–177. ISBN 9780443315541. [Google Scholar]
- Leitão, J. H. Microbial Virulence Factors. Int J Mol Sci 2020, 21(15). [Google Scholar] [CrossRef] [PubMed]
- Gaurav, A.; Bakht, P.; Saini, M.; Pandey, S.; Pathania, R. Role of bacterial efflux pumps in antibiotic resistance, virulence, and strategies to discover novel efflux pump inhibitors. Microbiology (Reading) 2023, 169(5). [Google Scholar] [CrossRef]
- Kumawat, M.; Nabi, B.; Daswani, M.; Viquar, I.; Pal, N.; Sharma, P.; Tiwari, S.; Sarma, D. K.; Shubham, S.; Kumar, M. Role of bacterial efflux pump proteins in antibiotic resistance across microbial species. Microbial Pathogenesis 2023, 181, 106182. [Google Scholar] [CrossRef]
- Laborda, P.; Molin, S.; Johansen, H. K.; Martínez, J. L.; Hernando-Amado, S. Role of bacterial multidrug efflux pumps during infection. World Journal of Microbiology and Biotechnology 2024, 40, 226. [Google Scholar] [CrossRef]
- Shen, S.; He, Z.; Zhao, S.; Zhu, Z.; Wang, X.; Tian, Y.; Han, Y.; Hu, M.; Lu, C.; Li, A. Environmental high-risk efflux pumps mediate concurrent enhancement of resistance and virulence in reclaimed water from urban wastewater treatment plants. Journal of Hazardous Materials 2025, 493, 138236. [Google Scholar] [CrossRef]
- Marco-Fuertes, A.; Jordá, J.; Marin, C.; Lorenzo-Rebenaque, L.; Montoro-Dasi, L.; Vega, S. Multidrug-Resistant Escherichia coli Strains to Last Resort Human Antibiotics Isolated from Healthy Companion Animals in Valencia Region. Antibiotics (Basel) 2023, 12(11). [Google Scholar] [CrossRef] [PubMed]
- Zanichelli, V.; Sharland, M.; Cappello, B.; Moja, L.; Getahun, H.; Pessoa-Silva, C.; Sati, H.; Weezenbeek, C.; Balkhy, H.; Simão, M.; Gandra, S.; Huttner, B. The WHO AWaRe (Access, Watch, Reserve) antibiotic book and prevention of antimicrobial resistance. Bulletin of the World Health Organization 2023, 101, 290–296. [Google Scholar] [CrossRef]
- World Health Organisation. Critically Important Antimicrobials for Human Medicine: 6th Revision2018,Ranking of medically important antimicrobials for risk management of antimicrobial resistance due to non-human use.. ISBN 978-92-4-151552-8, 1-45.
- Husna, A.; Rahman, M. M.; Badruzzaman, A. T. M.; Sikder, M. H.; Islam, M. R.; Rahman, M. T.; Alam, J.; Ashour, H. M. Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines 2023, 11(11). [Google Scholar] [CrossRef] [PubMed]
- Prendergast, D. M.; Slowey, R.; Burgess, C. M.; Murphy, D.; Johnston, D.; Morris, D.; O' Doherty, Á.; Moriarty, J.; Gutierrez, M. Characterization of cephalosporin and fluoroquinolone resistant Enterobacterales from Irish farm waste by whole genome sequencing. Front Microbiol 2023, 14, 1118264. [Google Scholar] [CrossRef]
- Byrne, N.; O’Neill, L.; Dίaz, J. A. C.; Manzanilla, E. G.; Vale, A. P.; Leonard, F. C. Antimicrobial resistance in Escherichia coli isolated from on-farm and conventional hatching broiler farms in Ireland. Irish Veterinary Journal 2022, 75(1), 7. [Google Scholar] [CrossRef]
- Ewers, C.; de Jong, A.; Prenger-Berninghoff, E.; El Garch, F.; Leidner, U.; Tiwari, S. K.; Semmler, T. Genomic Diversity and Virulence Potential of ESBL- and AmpC-β-Lactamase-Producing Escherichia coli Strains From Healthy Food Animals Across Europe. Front Microbiol 2021, 12, 626774. [Google Scholar] [CrossRef]
- Ahmed, S. K.; Hussein, S.; Qurbani, K.; Ibrahim, R. H.; Fareeq, A.; Mahmood, K. A.; Mohamed, M. G. Antimicrobial resistance: Impacts, challenges, and future prospects. Journal of Medicine, Surgery, and Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Kaur, K.; Singh, S.; Kaur, R. Impact of antibiotic usage in food-producing animals on food safety and possible antibiotic alternatives. The Microbe 2024, 4, 100097. [Google Scholar] [CrossRef]
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosafety and Health 2021, 3(1), 32–38. [Google Scholar] [CrossRef]
- Kasimanickam, V.; Kasimanickam, M.; Kasimanickam, R. Antibiotics Use in Food Animal Production: Escalation of Antimicrobial Resistance: Where Are We Now in Combating AMR? Med Sci (Basel) 2021, 9(1). [Google Scholar] [CrossRef]
- Almansour, A. M.; Alhadlaq, M. A.; Alzahrani, K. O.; Mukhtar, L. E.; Alharbi, A. L.; Alajel, S. M. The Silent Threat: Antimicrobial-Resistant Pathogens in Food-Producing Animals and Their Impact on Public Health. Microorganisms 2023, 11(9), 2127. [Google Scholar] [CrossRef]
- Mediouni, M.; Diallo, A. B.; Makarenkov, V. Quantifying antimicrobial resistance in food-producing animals in North America. Frontiers in Microbiology 2025, 16, 2025. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, V.; Dapkevicius, M. L. E.; Caniça, M.; Tejedor-Junco, M. T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria Among Food-Producing Animals: Health Implications of Extended Spectrum β-lactamase (ESBL) Production. Animals (Basel) 2020, 10(12). [Google Scholar] [CrossRef]
- Munita, J. M.; Arias, C. A. Mechanisms of Antibiotic Resistance. Microbiol Spectr 2016, 4(2). [Google Scholar] [CrossRef] [PubMed]
- Devi, N. S.; Mythili, R.; Cherian, T.; Dineshkumar, R.; Sivaraman, G. K.; Jayakumar, R.; Prathaban, M.; Duraimurugan, M.; Chandrasekar, V.; Peijnenburg, W. J. G. M. Overview of antimicrobial resistance and mechanisms: The relative status of the past and current. The Microbe 2024, 3, 100083. [Google Scholar] [CrossRef]
- Nammi, J.; Pasala, R.; Andhe, N.; Vasam, R.; Poruri, A. D.; Sherikar, R. R. Antibiotic Misuse: An In-Depth Examination of Its Global Consequences and Public Health Challenges. Cureus 2025, 17(6), e85941. [Google Scholar] [CrossRef] [PubMed]
- Murray, L. M.; Hayes, A.; Snape, J.; Kasprzyk-Hordern, B.; Gaze, W. H.; Murray, A. K. Co-selection for antibiotic resistance by environmental contaminants. npj Antimicrobials and Resistance 2024, 2(1), 9. [Google Scholar] [CrossRef] [PubMed]
- Gillieatt, B. F.; Coleman, N. V. Unravelling the mechanisms of antibiotic and heavy metal resistance co-selection in environmental bacteria. FEMS Microbiol Rev 2024, 48(4). [Google Scholar]
- Lucchetti, D.; Delfino, D.; Di Giustino, P.; Droghei, B.; Mancuso, M.; Mauti, T.; Triolone, D.; Vaccari, S.; Neri, B.; Russo, K. Development of a QuEChERS-based method for the determination of quaternary ammonium compounds in different food matrices by LC-MS/MS. Journal of Food Composition and Analysis 2024, 136, 106760. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Avila-Novoa, M. G.; Iñiguez-Moreno, E.; Guerrero-Medina, P. J.; Gutiérrez-Lomelí, M. Antimicrobial activity of disinfectants commonly used in the food industry in Mexico. Journal of Global Antimicrobial Resistance 2017, 10, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Hachler, H.; Nuesch-Inderbinen, M.; Stephan, R. Characteristics of extended-spectrum beta-lactamase- and carbapenemase-producing Enterobacteriaceae Isolates from rivers and lakes in Switzerland. Appl Environ Microbiol 2013, 79(9), 3021–6. [Google Scholar] [CrossRef]
- Lu, Z.; Mahony, A. K.; Arnold, W. A.; Marshall, C. W.; McNamara, P. J. Quaternary ammonia compounds in disinfectant products: evaluating the potential for promoting antibiotic resistance and disrupting wastewater treatment plant performance. Environmental Science Advances 2024, 3(2), 208–226. [Google Scholar] [CrossRef]
- EURL-SRM, Analysis of Quaternary Ammonium Compounds (QACs) in Fruits and Vegetables using QuEChERS and LC-MS/MS. 2023. Available online: https://eurl-pesticides.eu/userfiles/file/EurlSRM/EurlSRM_meth_QAC_ShortMethod.pdf (accessed on 13 January 2026).
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic resistance genes in bacteria: Occurrence, spread, and control. Journal of Basic Microbiology 2021, 61(12), 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Central Statistics Office. Livestock Survey December 2024. 2025. Available online: https://www.cso.ie/en/releasesandpublications/ep/p-lsd/livestocksurveydecember2024/ (accessed on 09 January 2026).
- Morales-Ubaldo, A. L.; Rivero-Perez, N.; Valladares-Carranza, B.; Velázquez-Ordoñez, V.; Delgadillo-Ruiz, L.; Zaragoza-Bastida, A. Bovine mastitis, a worldwide impact disease: Prevalence, antimicrobial resistance, and viable alternative approaches. Veterinary and Animal Science 2023, 21, 100306. [Google Scholar] [CrossRef]
- Goulart, D. B.; Mellata, M. Escherichia coli Mastitis in Dairy Cattle: Etiology, Diagnosis, and Treatment Challenges. Front Microbiol 2022, 13, 928346. [Google Scholar]
- Zaatout, N. An overview on mastitis-associated Escherichia coli: Pathogenicity, host immunity and the use of alternative therapies. Microbiological Research 2022, 256, 126960. [Google Scholar] [CrossRef]
- Nobrega, D. B.; Naqvi, S. A.; Dufour, S.; Deardon, R.; Kastelic, J. P.; De Buck, J.; Barkema, H. W. Critically important antimicrobials are generally not needed to treat nonsevere clinical mastitis in lactating dairy cows: Results from a network meta-analysis. Journal of Dairy Science 2020, 103(11), 10585–10603. [Google Scholar] [CrossRef]
- EFSA. Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on veterinary medicinal products and repealing Directive 2001/82/EC. Official Journal of the European Union 20022 2019, L4/43, 43–167. [Google Scholar]
- More, S. J.; McCoy, F.; McAloon, C. I. The new Veterinary Medicines Regulation: rising to the challenge. Ir Vet J 2022, 75(1), 2. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; Manzanilla, E. G.; More, S. J.; O’Neill, L.; Bradford, L.; Carty, C. I.; Collins, Á. B.; McAloon, C. G. Current antimicrobial use in farm animals in the Republic of Ireland. Irish Veterinary Journal 2020, 73(1), 11. [Google Scholar] [CrossRef]
- National Dairy Council. Is all milk pasteurised? 2022. Available online: https://ndc.ie/accordion/is-all-milk-pasteurised/ (accessed on 13 January 2026).
- Paramasivam, R.; Gopal, D. R.; Dhandapani, R.; Subbarayalu, R.; Elangovan, M. P.; Prabhu, B.; Veerappan, V.; Nandheeswaran, A.; Paramasivam, S.; Muthupandian, S. Is AMR in Dairy Products a Threat to Human Health? An Updated Review on the Origin, Prevention, Treatment, and Economic Impacts of Subclinical Mastitis. Infection and Drug Resistance 2023, 16, 155–178. [Google Scholar] [CrossRef]
- Tóth, A. G.; Csabai, I.; Krikó, E.; Tőzsér, D.; Maróti, G.; Patai Á, V.; Makrai, L.; Szita, G.; Solymosi, N. Antimicrobial resistance genes in raw milk for human consumption. Sci Rep 2020, 10(1), 7464. [Google Scholar] [CrossRef]
- Majumder, S.; Jung, D.; Ronholm, J.; George, S. Prevalence and mechanisms of antibiotic resistance in Escherichia coli isolated from mastitic dairy cattle in Canada. BMC Microbiology 2021, 21(1), 222. [Google Scholar]
- Nüesch-Inderbinen, M.; Käppeli, N.; Morach, M.; Eicher, C.; Corti, S.; Stephan, R. Molecular types, virulence profiles and antimicrobial resistance of Escherichia coli causing bovine mastitis. Vet Rec Open 2019, 6(1), e000369. [Google Scholar] [CrossRef] [PubMed]
- Jalil, A.; Gul, S.; Bhatti, M. F.; Siddiqui, M. F.; Adnan, F. High Occurrence of Multidrug-Resistant Escherichia coli Strains in Bovine Fecal Samples from Healthy Cows Serves as Rich Reservoir for AMR Transmission. Antibiotics (Basel) 2022, 12(1). [Google Scholar] [CrossRef]
- Garzon, A.; Portillo, R.; Habing, G.; Silva-del-Rio, N.; Karle, B. M.; Pereira, R. V. Antimicrobial resistance of Escherichia coli from dairy farms participating in an antimicrobial stewardship educational program for farm employees. Journal of Dairy Science 2024, 107(3), 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R. S.; Cavaco, L. M.; Guerra, B.; Bortolaia, V.; Agersø, Y.; Svendsen, C. A.; Nielsen, H. N.; Kjeldgaard, J. S.; Pedersen, S. K.; Fertner, M.; Hasman, H. Evaluation and validation of laboratory procedures for the surveillance of ESBL-, AmpC-, and carbapenemase-producing Escherichia coli from fresh meat and caecal samples. Front Microbiol 2023, 14, 1229542. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, D. M.; O'Doherty, Á.; Burgess, C. M.; Howe, N.; McMahon, F.; Murphy, D.; Leonard, F.; Morris, D.; Harrington, C.; Carty, A.; Moriarty, J.; Gutierrez, M. Critically important antimicrobial resistant Enterobacteriaceae in Irish farm effluent and their removal in integrated constructed wetlands. Science of The Total Environment 2022, 806, 151269. [Google Scholar] [CrossRef] [PubMed]
- Microbiology of the Food Chain: Enumeration of ß-Glucuronidase-Positive Escherichia coli at 44˚C. 2001. Available online: https://cdn.standards.iteh.ai/samples/29824/cbb8efcaa00647c3807681d77c0fb144/ISO-16649-2-2001.pdf (accessed on 13 January 2025).
- O'Brien, M.; Hunt, K.; McSweeney, S.; Jordan, K. Occurrence of foodborne pathogens in Irish farmhouse cheese. Food Microbiology 2009, 26(8), 910–914. [Google Scholar] [CrossRef]
- Ramovic, E.; Madigan, G.; McDonnell, S.; Griffin, D.; Bracken, E.; NiGhallchoir, E.; Quinless, E.; Galligan, A.; Egan, J.; Prendergast, D. M. A pilot study using environmental screening to determine the prevalence of Mycobacterium avium subspecies paratuberculosis (MAP) and antimicrobial resistance (AMR) in Irish cattle herds. Ir Vet J 2020, 73, 3. [Google Scholar] [CrossRef]
- Commission Implementing Decision (EU) 2020/1729 of 17 November 2020 on the monitoring and reporting of antimicrobial resistance in zoonotic and commensal bacteria and repealing Implementing Decision 2013/652/EU. OJEU 2020, 387, 8–21.
- EUCAST guidelines for detection of resistance mechanisms and specific resistances of clinical and/epidemiological importance. 2013. Available online: https://rosco-diagnostica.com/wp-content/uploads/EUCAST_guidelines_detection_of_resistance_mechanisms_121222.pdf (accessed on 13 January 2026).
- EFSA (European Food Safety Authority), A. G., Beloeil P-A, GarciaFierro R, Guerra B, Rizzi V and Stoicescu A-V, , Manual for reporting 2023 antimicrobialresistance data under Directive 2003/99/EC and Commission Implementing Decision (EU)2020/1729. 2024, EFSA supporting publication 2024:EN-8585. 41 pp. doi:10.2903/sp.efsa.2024.EN-8585ISSN: 2397-8325. (Accessed on 13 January 2026).
- Sergeant, E. Epitools Epidemiological Calculators. 2018. Available online: https://epitools.ausvet.com.au/chisq (accessed on 13 January 2026).
- Feldgarden, M.; Brover, V.; Haft, D. H.; Prasad, A. B.; Slotta, D. J.; Tolstoy, I.; Tyson, G. H.; Zhao, S.; Hsu, C. H.; McDermott, P. F.; Tadesse, D. A.; Morales, C.; Simmons, M.; Tillman, G.; Wasilenko, J.; Folster, J. P.; Klimke, W. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob Agents Chemother 2019, 63(11). [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J. G.; Haendiges, J.; Haft, D. H.; Hoffmann, M.; Pettengill, J. B.; Prasad, A. B.; Tillman, G. E.; Tyson, G. H.; Klimke, W. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci Rep 2021, 11(1), 12728. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Fedorov, B.; Haft, D. H.; Prasad, A. B.; Klimke, W. Curation of the AMRFinderPlus databases: applications, functionality and impact. Microb Genom 2022, 8(6). [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Research 2016, 44(D1), D694–D697. [Google Scholar] [CrossRef] [PubMed]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: an easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb Genom 2018, 4(7). [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T. L. BLAST+: architecture and applications. BMC Bioinformatics 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrobial Agents and Chemotherapy 2014, 58(7), 3895–3903. [Google Scholar] [CrossRef]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. Correction: PathogenFinder - Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLOS ONE 2013, 8(12). [Google Scholar] [CrossRef]
- Commission Implementing Decision (EU) 2020/1729 of 17 November 2020 on the monitoring and reporting of antimicrobial resistance in zoonotic and commensal bacteria and repealing Implementing Decision 2013/652/EU. OJEU 2020, 387, 8–21.
- Nyambe, S.; Brehony, C.; Hselin, R.; Byrne, B.; Morris, *!!! REPLACE !!!*; Dearbhaile, *!!! REPLACE !!!*; Gutierrez, M. Occurrence and molecular characterisation of verocytotoxigenic Escherichia coli in raw milk and milk filters in Ireland. In Proceedings of the 11th International Symposium on Shiga Toxin (Verocytotoxin producing Escherichia coli infections, Banff centre for arts and creativity Banff, Alberta, Canada, 7th – 10th May, 2023. [Google Scholar]
- FSAI. Raw milk and raw milk filter microbiological surveillance programme. 2015. Available online: https://www.fsai.ie/publications/raw-milk-and-raw-milk-filter-microbiological-survey (accessed on 13 January 2026).
- Murphy, M.; Buckley, J. F.; Whyte, P.; O'Mahony, M.; Anderson, W.; Wall, P. G.; Fanning, S. Surveillance of dairy production holdings supplying raw milk to the farmhouse cheese sector for Escherichia coli O157, O26 and O111. Zoonoses Public Health 2007, 54(9-10), 358–65. [Google Scholar] [CrossRef]
- Stephan, R.; Schumacher, S.; Corti, S.; Krause, G.; Danuser, J.; Beutin, L. Prevalence and Characteristics of Shiga Toxin-Producing Escherichia coli in Swiss Raw Milk Cheeses Collected at Producer Level. Journal of Dairy Science 2008, 91(7), 2561–2565. [Google Scholar] [CrossRef]
- Hazards, E. P. o. B., Scientific Opinion on the public health risks related to the consumption of raw drinking milk. EFSA Journal 2015, 13(1), 3940. [CrossRef]
- Drugea, R. I.; Siteavu, M. I.; Pitoiu, E.; Delcaru, C.; Sârbu, E. M.; Postolache, C.; Bărăităreanu, S. Prevalence and Antibiotic Resistance of Escherichia coli Isolated from Raw Cow’s Milk. Microorganisms 2025, 209. [Google Scholar] [CrossRef]
- Imre, K.; Ban-Cucerzan, A.; Herman, V.; Sallam, K. I.; Cristina, R. T.; Abd-Elghany, S. M.; Morar, D.; Popa, S. A.; Imre, M.; Morar, A. Occurrence, Pathogenic Potential and Antimicrobial Resistance of Escherichia coli Isolated from Raw Milk Cheese Commercialized in Banat Region, Romania. Antibiotics (Basel) 2022, 11(6). [Google Scholar] [CrossRef]
- Morandi, S.; Silvetti, T.; Bonazza, F.; Brasca, M. Occurrence and diversity of Shiga toxin-producing Escherichia coli (STEC) in Italian Alpine raw milk cheeses and their development in the earlier stages of different cheese-making processes. LWT 2024, 213, 117029. [Google Scholar] [CrossRef]
- Condor, S.; Duma, M.; Crăciun, S.; Mihaiu, M.; Cîmpean, R.; Crisan-Reget, O. L.; Dan, S. D.; Condor, L.; Ionica, C.-N.; Tabaran, A. Quantification of Total Staphylococci and Escherichia coli in Milk and Dairy Products from Small Ruminants and Characterization of the Antimicrobial Resistance Profiles of Isolated Pathogenic Strains. Microorganisms 2025, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Heinsbroek, E.; Blakey, E.; Simpson, A.; Verlander, N. Q.; Greig, D. R.; Jorgensen, F.; Nelson, A.; Douglas, A.; Balasegaram, S.; Jenkins, C.; Elson, R. An outbreak of Shiga toxin-producing Escherichia coli serotype O103:H2 associated with unpasteurized soft cheese, England and Wales, 2022. Epidemiology and Infection 2024, 152, e172. [Google Scholar] [CrossRef]
- Shoaib, M.; He, Z.; Geng, X.; Tang, M.; Hao, R.; Wang, S.; Shang, R.; Wang, X.; Zhang, H.; Pu, W. The emergence of multi-drug resistant and virulence gene carrying Escherichia coli strains in the dairy environment: a rising threat to the environment, animal, and public health. Front Microbiol 2023, 14, 1197579. [Google Scholar] [CrossRef] [PubMed]
- Quinn, O. I.; Jenkins, C.; Greig, D. R.; Neale, S.; Jorgensen, F.; Yanshi; Inns, T.; Allison, L.; Browning, L.; Douglas, A.; Balasegram, S. An outbreak of Shiga Toxin-producing Escherichia coli Serotype O145:H28 Associated with Domestic Travel and Consumption of Unpasteurized Cheese, UK, 2023. Journal of Food Protection 2025, 88(4), 100470. [Google Scholar] [CrossRef]
- Laslo, É.; György, É. Evaluation of the microbiological quality of some dairy products. Acta Universitatis Sapientiae, Alimentaria 2018, 11(1), 27–44. [Google Scholar] [CrossRef]
- Younis, W.; Hassan, S.; Mohamed, H. M. A. Molecular characterization of Escherichia coli isolated from milk samples with regard to virulence factors and antibiotic resistance. Vet World 2021, 14(9), 2410–2418. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 852/2004 of the european parliament and of the council of 29 April 2004 on the hygiene of foodstuffs. Official Journal of the European Communities 2004, L139, 1–54.
- Commission Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Official Journal of the European Communities 2004, L 139/55, 55–205.
- Huang, L.; Wu, C.; Gao, H.; Xu, C.; Dai, M.; Huang, L.; Hao, H.; Wang, X.; Cheng, G. Bacterial Multidrug Efflux Pumps at the Frontline of Antimicrobial Resistance: An Overview. Antibiotics (Basel) 2022, 11(4). [Google Scholar] [CrossRef]
- Pugh, H. L.; Connor, C.; Siasat, P.; McNally, A.; Blair, J. M. A. E. coli ST11 (O157:H7) does not encode a functional AcrF efflux pump. Microbiology 2023, 169(4). [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, V.; Stone, T. A.; Deber, C. M.; Karplus, M. Structure of the EmrE multidrug transporter and its use for inhibitor peptide design. Proceedings of the National Academy of Sciences 2018, 115(34), E7932–E7941. [Google Scholar] [CrossRef]
- Schuldiner, S.; Granot, D.; Mordoch, S. S.; Ninio, S.; Rotem, D.; Soskin, M.; Tate, C. G.; Yerushalmi, H. Small is Mighty: EmrE, a Multidrug Transporter as an Experimental Paradigm. Physiology 2001, 16(3), 130–134. [Google Scholar] [CrossRef] [PubMed]
- Law, C. J.; Maloney, P. C.; Wang, D. N. Ins and outs of major facilitator superfamily antiporters. Annu Rev Microbiol 2008, 62, 289–305. [Google Scholar] [CrossRef]
- Law, C. J.; Alegre, K. O. Clamping down on drugs: the Escherichia coli multidrug efflux protein MdtM. Res Microbiol 2018, 169(7-8), 461–467. [Google Scholar] [CrossRef] [PubMed]
- Nagarathinam, K.; Nakada-Nakura, Y.; Parthier, C.; Terada, T.; Juge, N.; Jaenecke, F.; Liu, K.; Hotta, Y.; Miyaji, T.; Omote, H.; Iwata, S.; Nomura, N.; Stubbs, M. T.; Tanabe, M. Outward open conformation of a Major Facilitator Superfamily multidrug/H+ antiporter provides insights into switching mechanism. Nature Communications 2018, 9(1), 4005. [Google Scholar] [CrossRef] [PubMed]
- Banigan, J. R.; Gayen, A.; Cho, M. K.; Traaseth, N. J. A structured loop modulates coupling between the substrate-binding and dimerization domains in the multidrug resistance transporter EmrE. J Biol Chem 2015, 290(2), 805–14. [Google Scholar] [CrossRef]
- Walczak Ł, J.; Kwiatkowska, M.; Twarowski, B.; Kubacka, M.; Paluch, J.; Herbet, M. Disinfectant-induced bacterial resistance and antibiotic cross-resistance-mechanisms and clinical relevance. Clin Exp Med 2025, 26(1), 26. [Google Scholar]
- Sousa, M.; Machado, I.; Simões, L. C.; Simões, M. Biocides as drivers of antibiotic resistance: A critical review of environmental implications and public health risks. Environmental Science and Ecotechnology 2025, 25, 100557. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, J.; Zhao, K.; Yang, S.; Li, C.; Penttinen, P.; Ao, X.; Liu, A.; Hu, K.; Li, J.; Yang, Y.; Liu, S.; Bai, L.; Zou, L. Class 1 integron carrying qacEΔ1 gene confers resistance to disinfectant and antibiotics in Salmonella. International Journal of Food Microbiology 2023, 404, 110319. [Google Scholar] [CrossRef]
- Teelucksingh, T.; Thompson Laura, K.; Cox, G. The Evolutionary Conservation of Escherichia coli Drug Efflux Pumps Supports Physiological Functions. Journal of Bacteriology 2020, 202(22). [Google Scholar] [CrossRef]
- Pugh, H. L.; Connor, C.; Siasat, P.; McNally, A.; Blair, J. M. A. E. coli ST11 (O157:H7) does not encode a functional AcrF efflux pump. Microbiology (Reading) 2023, 169(4). [Google Scholar] [CrossRef]
- Nordholt, N.; Kanaris, O.; Schmidt, S. B. I.; Schreiber, F. Persistence against benzalkonium chloride promotes rapid evolution of tolerance during periodic disinfection. Nat Commun 2021, 12(1), 6792. [Google Scholar] [CrossRef]
- Paulsen, I. T.; Littlejohn, T. G.; Rådström, P.; Sundström, L.; Sköld, O.; Swedberg, G.; Skurray, R. A. The 3' conserved segment of integrons contains a gene associated with multidrug resistance to antiseptics and disinfectants. Antimicrobial Agents and Chemotherapy 1993, 37(4), 761–768. [Google Scholar] [CrossRef]
- Jaglic, Z.; Cervinkova, D. Genetic basis of resistance to quaternary ammonium compounds - the qac genes and their role: a review. Veterinární medicína 2012, 57(6), 275–281. [Google Scholar] [CrossRef]
- Romão, C.; Miranda, C. A.; Silva, J.; Mandetta Clementino, M.; de Filippis, I.; Asensi, M. Presence of qacEΔ1 Gene and Susceptibility to a Hospital Biocide in Clinical Isolates of Pseudomonas aeruginosa Resistant to Antibiotics. Current Microbiology 2011, 63(1), 16–21. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Helal, Z. QacE and QacEΔ1 Genes and Their Correlation to Antibiotics and Biocides Resistance Pseudomonas aeruginosa. Am. J. Biomed Sci 2015, 7, 52–62. [Google Scholar]
- Ransom, E. M.; Potter, R. F.; Dantas, G.; Burnham, C.-A. D. Genomic Prediction of Antimicrobial Resistance: Ready or Not, Here It Comes! Clinical Chemistry 2020, 66(10), 1278–1289. [Google Scholar] [CrossRef] [PubMed]
- Merchel Piovesan Pereira, B.; Wang, X.; Tagkopoulos, I. Biocide-Induced Emergence of Antibiotic Resistance in Escherichia coli. Front Microbiol 2021, 12, 640923. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int J Med Microbiol 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- San Millan, A. Evolution of Plasmid-Mediated Antibiotic Resistance in the Clinical Context. Trends Microbiol 2018, 26, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Mathers, A. J.; Peirano, G.; Pitout, J. D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin Microbiol Rev 2015, 28, 565–91. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M. S. M.; Fischer, J.; Wagenaar, J. A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D. J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. Journal of Antimicrobial Chemotherapy 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Adamczuk, M.; Zaleski, P.; Dziewit, L.; Wolinowska, R.; Nieckarz, M.; Wawrzyniak, P.; Kieryl, P.; Plucienniczak, A.; Bartosik, D. Diversity and Global Distribution of IncL/M Plasmids Enabling Horizontal Dissemination of β-Lactam Resistance Genes among the Enterobacteriaceae. Biomed Res Int 2015, 2015, 414681. [Google Scholar] [CrossRef]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic Features of blaNDM-1-Positive Enterobacteriaceae. Antimicrobial Agents and Chemotherapy 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: the phantom menace. Journal of Antimicrobial Chemotherapy 2012, 67, 1597–1606. [Google Scholar] [CrossRef]
- Lotfi, M.; Jalal, D.; Sayed, A. A. plsMD: A plasmid reconstruction tool from short-read assemblies. bioRxiv 2025, 03.17.643493. [Google Scholar] [CrossRef]
- Arredondo-Alonso, S.; Willems, R. J.; van Schaik, W.; Schürch, A. C. On the (im)possibility of reconstructing plasmids from whole-genome short-read sequencing data. Microb Genom 2017, 3, e000128. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-Alonso, S.; Top, J.; McNally, A.; Puranen, S.; Pesonen, M.; Pensar, J.; Marttinen, P.; Braat, J. C.; Rogers, M. R. C.; van Schaik, W.; Kaski, S.; Willems, R. J. L.; Corander, J.; Schürch, A. C. Plasmids Shaped the Recent Emergence of the Major Nosocomial Pathogen Enterococcus faecium. mBio 2020, 11(1). [Google Scholar] [CrossRef] [PubMed]
- Redgrave, L. S.; Sutton, S. B.; Webber, M. A.; Piddock, L. J. V. Fluoroquinolone resistance: mechanisms, impact on bacteria, and role in evolutionary success. Trends in Microbiology 2014, 22, 438–445. [Google Scholar] [CrossRef]
- Azzariti, S.; Bond, R.; Loeffler, A.; Zendri, F.; Timofte, D.; Chang, Y. M.; Pelligand, L. Investigation of In Vitro Susceptibility and Resistance Mechanisms in Skin Pathogens: Perspectives for Fluoroquinolone Therapy in Canine Pyoderma. Antibiotics (Basel) 2022, 11(9). [Google Scholar] [CrossRef]
- Tewawong, N.; Kowaboot, S.; Lektrakul, W.; Supcharoengoon, U.; Watanagul, N.; Pitaksajjakul, P. Mechanisms of fluoroquinolone resistance among Escherichia coli isolates from urinary tract infections in Thailand. PLOS ONE 2025, 20(5), e0325175. [Google Scholar] [CrossRef]
- Enshaie, E.; Nigam, S.; Patel, S.; Rai, V. Livestock Antibiotics Use and Antimicrobial Resistance. Antibiotics 2025, 14, 621. [Google Scholar] [CrossRef]
- Zhao, Y.; Niu, Y.; Zhao, M.; Huang, W.; Qin, Y. Prevalence of antibiotic resistance genes its association with microbiota in raw milk of northwest Xinjiang. Front Microbiol 2025, 16, 1595051. [Google Scholar] [CrossRef] [PubMed]
- de Jong, E.; McCubbin, K. D.; Speksnijder, D.; Dufour, S.; Middleton, J. R.; Ruegg, P. L.; Lam, T.; Kelton, D. F.; McDougall, S.; Godden, S. M.; Lago, A.; Rajala-Schultz, P. J.; Orsel, K.; De Vliegher, S.; Krömker, V.; Nobrega, D. B.; Kastelic, J. P.; Barkema, H. W. Invited review: Selective treatment of clinical mastitis in dairy cattle. J Dairy Sci 2023, 106, 3761–3778. [Google Scholar] [CrossRef] [PubMed]
- Huygens, J.; Daeseleire, E.; Mahillon, J.; Van Elst, D.; Decrop, J.; Meirlaen, J.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Presence of Antibiotic Residues and Antibiotic Resistant Bacteria in Cattle Manure Intended for Fertilization of Agricultural Fields: A One Health Perspective. Antibiotics (Basel) 2021, 10(4). [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Tan, J.; Khan, M. F.; Chugh, G.; Schmidt, O.; Ma, L.; Bu, D. Emerging Microbial and Enzymatic Approaches for Sustainable Antibiotic Biodegradation in Livestock Manure to Mitigate Water Pollution Risks. Water 2025, 16, 2960. [Google Scholar] [CrossRef]
- Qiu, T.; Huo, L.; Guo, Y.; Gao, M.; Wang, G.; Hu, D.; Li, C.; Wang, Z.; Liu, G.; Wang, X. Metagenomic assembly reveals hosts and mobility of common antibiotic resistome in animal manure and commercial compost. Environmental Microbiome 2022, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Trinchera, M.; De Gaetano, S.; Sole, E.; Midiri, A.; Silvestro, S.; Mancuso, G.; Catalano, T.; Biondo, C. Antimicrobials in Livestock Farming and Resistance: Public Health Implications. Antibiotics 2025, 14, 606. [Google Scholar] [CrossRef]
- Wellington, E. M. H.; Boxall, A. B. A.; Cross, P.; Feil, E. J.; Gaze, W. H.; Hawkey, P. M.; Johnson-Rollings, A. S.; Jones, D. L.; Lee, N. M.; Otten, W.; Thomas, C. M.; Williams, A. P. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. The Lancet Infectious Diseases 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, L.; Huang, Y.; Xu, X.; Liu, Z.; Li, S.; Zhu, L.; Hu, B.; Zhang, T. Global soil antibiotic resistance genes are associated with increasing risk and connectivity to human resistome. Nature Communications 2025, 16, 7141. [Google Scholar] [CrossRef]
- Li, X.; Bickel, S.; Wicaksono, W. A.; Lin, X.; Berg, G.; Zhu, Y. Unraveling antibiotic resistance dynamics at the soil–plant interface under climate change for One Health. One Health Advances 2025, 3, 16. [Google Scholar] [CrossRef]
- Partridge, S. R.; Kwong, S. M.; Firth, N.; Jensen, S. O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin Microbiol Rev 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga Toxin-Associated Hemolytic Uremic Syndrome: A Narrative Review. Toxins (Basel) 2020, 12, 67. [Google Scholar]
- Merrick, R.; Song, J.; Fina, L.; Sawyer, C.; Jenkins, C.; King, G.; Turner, D.; Thomas, D.; Williams, C. Long-term health outcomes of Shiga toxin-producing Escherichia coli O157 (STEC O157) infection and STEC-associated haemolytic uraemic syndrome (STEC-HUS), Wales, 1990–2020. Pediatric Nephrology 2025, 40, 2295–2310. [Google Scholar] [CrossRef]
- Garcia, T. A.; Ventura, C. L.; Smith, M. A.; Merrell, D. S.; O'Brien, A. D. Cytotoxic necrotizing factor 1 and hemolysin from uropathogenic Escherichia coli elicit different host responses in the murine bladder. Infect Immun 2013, 81, 99–109. [Google Scholar] [CrossRef]
- Zaki, M.; Bastawy, S.; Montasser, K. Molecular Study of E. coli Virulence Genes in Nosocomial Sepsis. Biosciences, Biotechnology Research Asia 2019, 16, 269–277. [Google Scholar] [CrossRef]
- Nagamatsu, K.; Hannan, T. J.; Guest, R. L.; Kostakioti, M.; Hadjifrangiskou, M.; Binkley, J.; Dodson, K.; Raivio, T. L.; Hultgren, S. J. Dysregulation of Escherichia coli α-hemolysin expression alters the course of acute and persistent urinary tract infection. Proc Natl Acad Sci U S A 2015, 112, E871-80. [Google Scholar] [CrossRef]
- Tozzi, M.; Fiore, A.; Travaglione, S.; Marcon, F.; Rainaldi, G.; Germinario, E. A. P.; Laterza, I.; Donati, S.; Macchia, D.; Spada, M.; Leoni, O.; Quattrini, M. C.; Pietraforte, D.; Tomasoni, S.; Torrigiani, F.; Verin, R.; Matarrese, P.; Gambardella, L.; Spadaro, F.; Carollo, M.; Pietrantoni, A.; Carlini, F.; Panebianco, C.; Pazienza, V.; Colella, F.; Lucchetti, D.; Sgambato, A.; Sistigu, A.; Moschella, F.; Guidotti, M.; Vincentini, O.; Maroccia, Z.; Biffoni, M.; De Angelis, R.; Bracci, L.; Fabbri, A. E. Coli cytotoxic necrotizing factor-1 promotes colorectal carcinogenesis by causing oxidative stress, DNA damage and intestinal permeability alteration. J Exp Clin Cancer Res 2025, 44, 29. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, X.; Xu, H.; Xu, Y.; Jielu, L.; Zhang, D.; Gao, S.; Liu, X. Roles of iron acquisition systems in virulence of extraintestinal pathogenic Escherichia coli: Salmochelin and aerobactin contribute more to virulence than heme in a chicken infection model. BMC microbiology 2012, 12, 143. [Google Scholar] [CrossRef]
- Bergsten, G.; Wullt, B.; Svanborg, C. Escherichia coli, fimbriae, bacterial persistence and host response induction in the human urinary tract. International Journal of Medical Microbiology 2005, 295, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Wullt, B.; Bergsten, G.; Samuelsson, M.; Svanborg, C. The role of P fimbriae for Escherichia coli establishment and mucosal inflammation in the human urinary tract. International Journal of Antimicrobial Agents 2002, 19, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Foroogh, N.; Rezvan, M.; Ahmad, K.; Mahmood, S. Structural and functional characterization of the FimH adhesin of uropathogenic Escherichia coli and its novel applications. Microbial Pathogenesis 2021, 161, 105288. [Google Scholar] [CrossRef] [PubMed]
- Aljohni, M. S.; Harun-Ur-Rashid, M.; Selim, S. Emerging threats: Antimicrobial resistance in extended-spectrum beta-lactamase and carbapenem-resistant Escherichia coli. Microbial Pathogenesis 2025, 200, 107275. [Google Scholar] [CrossRef]
Figure 1.
Distribution of virulence genes and E. coli sequence types amongst the 32 sequenced isolates.
Figure 1.
Distribution of virulence genes and E. coli sequence types amongst the 32 sequenced isolates.
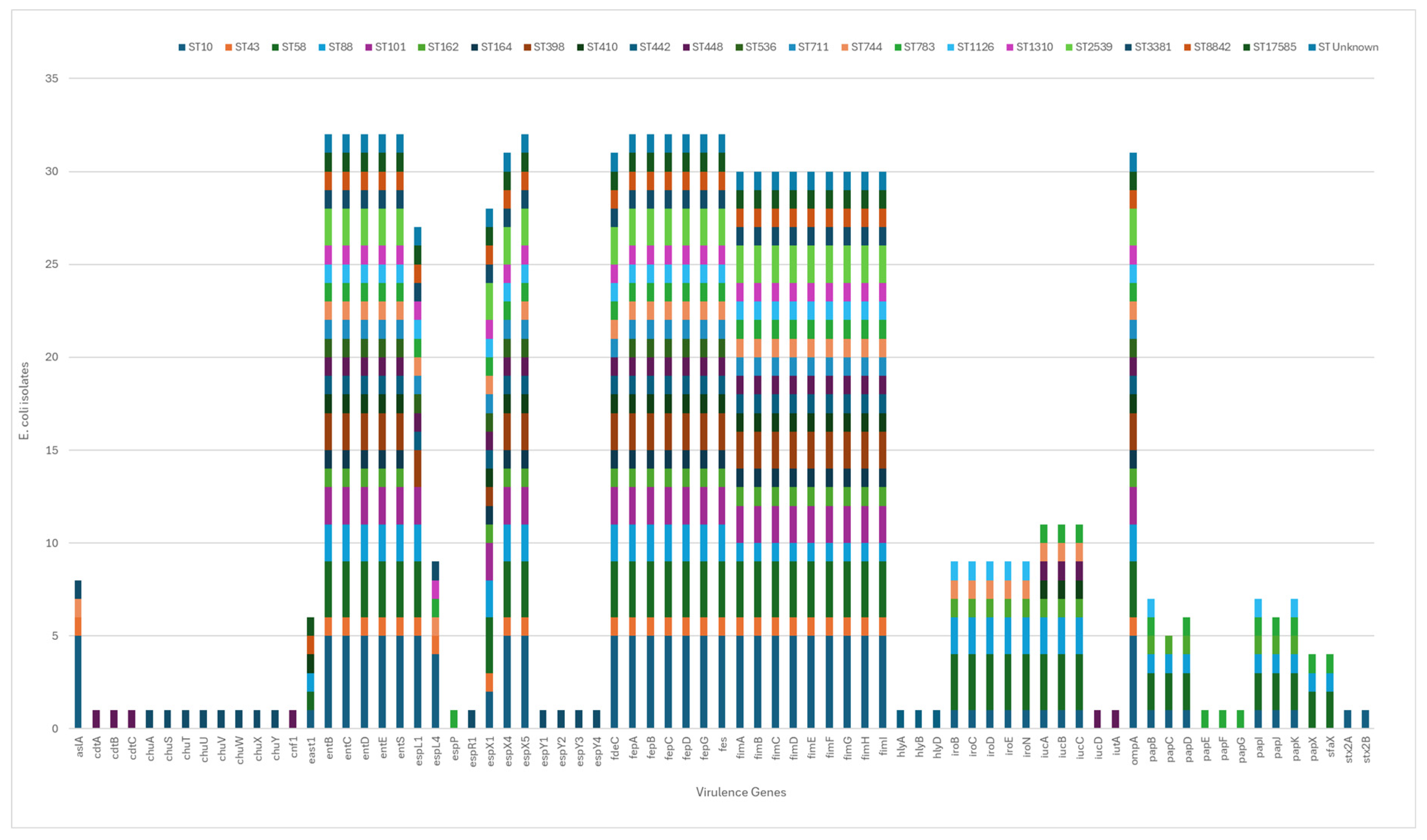
Figure 2.
Core genome MLST based Neighbor-joining phylogenetic tree of 32 E. coli isolates with distance based on 2513 core loci. Each coloured rectangle refers to the MLST (Warwick scheme) for a single strain with the source of each isolate written within each rectangle. The numbers within each coloured circle on the left-hand side refer to the ST for each isolate. The two red circles refer to the two isolates within each cgMLST as indistinguishable.
Figure 2.
Core genome MLST based Neighbor-joining phylogenetic tree of 32 E. coli isolates with distance based on 2513 core loci. Each coloured rectangle refers to the MLST (Warwick scheme) for a single strain with the source of each isolate written within each rectangle. The numbers within each coloured circle on the left-hand side refer to the ST for each isolate. The two red circles refer to the two isolates within each cgMLST as indistinguishable.
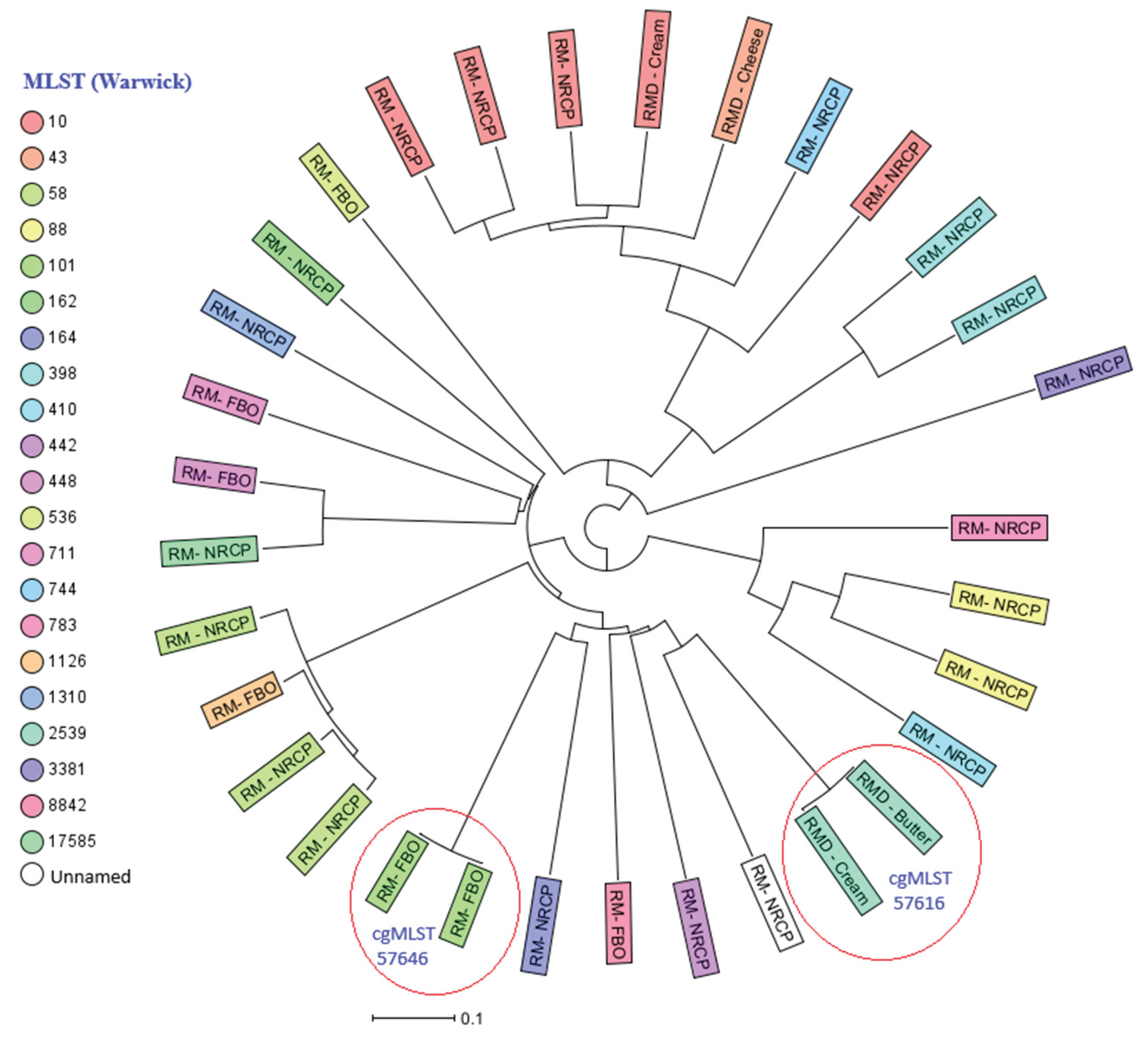
Table 2.
AMR profiles of commensal E. coli and presumptive ESBL/AmpC producing and fluroquinolone resistant E. coli isolates.
Table 2.
AMR profiles of commensal E. coli and presumptive ESBL/AmpC producing and fluroquinolone resistant E. coli isolates.
| Source | Phenotypic AMR profile | Commensal E. coli | ESBL/AmpC producers | Fluoroquinolone resistant |
| RM- NRCP | Amp, Chl, Cip, Nal, Smx, Tet, Tmp | 1 | ||
| Amp, Chl, Cip, Nal. Smx, Tmp | 1 | |||
| Amp, Ctx, Fox, Caz, Smx, Tmp | 1 | |||
| Amp, Chl, Smx, Tet, Tmp | 1 | |||
| Amp Cip Nal Tet | 1 | |||
| Amp Smx Tet Tmp | 1 | |||
| Amp Smx Tmp | 3 | |||
| Fully Susceptible | 17 | |||
| RM- FBO | Amp Ctx Caz Chl Fep Gen Smx Tet Tmp | 1 | ||
| Amp, Smx, Tet | 1 | |||
| Fully susceptible | 13 | |||
| RDP | Cip, Nal, Smx, Tmp | 1 | ||
| Amp, Tet | 1 | |||
| Fully Susceptible | 4 | |||
| Total no. isolates | 47 | 41 | 2 | 4 |
Table 3.
Phenotypic and genomic antimicrobial resistance of E. coli isolates, along with genes encoding efflux pumps, quaternary ammonium compounds resistance and plasmid replicons. .
Table 3.
Phenotypic and genomic antimicrobial resistance of E. coli isolates, along with genes encoding efflux pumps, quaternary ammonium compounds resistance and plasmid replicons. .
| Isolate ID | Source | Presumptive E. coli | AMR results | Aminoglycosides | Amphe-nicols | β-lactams | Quinolones | Sulfonamides | Tetracyclines | Diaminopy-rimidines | NCBI AMRFinderPlus | PlasmidFinder 2.1 | PathogenFinder 1.1 | Clermon Typing | ||
| GEN / KAN / STR | SF | TET | TMP | Efflux pump | Quaternary Ammo-nium | Plasmid identified | Patho-genicity | Patho-genic families | Phylo-group | |||||||
| 01 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | emrE, mdtM | - | IncFIB(pB171), IncFII(pHN7A8) | 0.94 | 535 | A |
| 02 | RM- NRCP | FQ | Amp, Cip, Nal, Tet | - | - | blaTEM-1 | gyrA_D87N, gyrA_S83L, parC_S80I | - | tet(B) | - | acrF | - | IncFIB(AP001918), IncFII | 0.86 | 54 | B1 |
| 03 | RM- NRCP | Commensal | Amp, Smx, Tmp | aph(3')-Ia, aph(3'')-Ib, aph(6)-Id | - | blaTEM-1 | - | sul2 | - | dfrA5 | acrF | - | IncFIB(AP001918), IncFII | 0.86 | 57 | B1 |
| 04 | RM- NRCP | AmpC | Amp, Ctx, Fox, Caz, Smx, Tmp | aph(3'')-Ib, aph(6)-Id | - | blaTEM-1, ampC_C-42T | - | sul2 | - | dfrA5 | acrF | - | IncFIB(AP001918), IncFII | 0.86 | 54 | C |
| 05 | RM- NRCP | Commensal | Amp, Smx, Tet, Tmp | aph(3'')-Ib, aph(6)-Id | - | blaTEM-1 | - | sul2 | tet(A) | dfrA5 | acrF, emrE, emrE, mdtM | - | IncFII, IncI1-I(Alpha) | 0.86 | 54 | B1 |
| 06 | RM- NRCP | Commensal | Amp, Smx, Tmp | aph(3'')-Ib, aph(6)-Id | - | blaTEM-1 | - | sul2 | - | dfrA5 | acrF, mdtM | - | IncFIB(AP001918), IncFII | 0.86 | 55 | B1 |
| 07 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIA | 0.94 | 693 | B1 |
| 08 | RMD (Butter) | Commensal | Fully susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIB(pB171), IncFII(pCoo) | 0.90 | 152 | B1 |
| 09 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, mdtM | - | IncX1 | 0.84 | 133 | A |
| 10 | RM- NRCP | Commensal | Fully susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIA, IncFIB(AP001918), IncFIC(FII) | 0.93 | 673 | A |
| 11 | RM- NRCP | Commensal | Fully susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIB(AP001918), IncFIC(FII) | 0.93 | 630 | A |
| 12 | RM- NRCP | FQ | Amp, Chl, Cip, Nal. Smx, Tmp | aph(3')-Ia, aadA1 | floR | blaTEM-39, blaOXA-1 | gyrA_D87N, gyrA_S83L, parC_S80I, parE_S458A | sul1, sul2 | tet(A) | dfrA36 | emrE, mdtM | qacED1 | ColpVC, IncFIA, IncFIB(AP001918), IncFII | 0.86 | 53 | C |
| 13 | RM- NRCP | Commensal | Amp, Smx, Tmp | aph(3'')-Ib, aph(6)-Id | - | blaTEM-1 | - | sul2 | - | dfrA5 | acrF, emrE, mdtM | - | IncFIB(AP001918), IncFII | 0.86 | 55 | A |
| 14 | RM- FBO | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, mdtM | - | IncY | 0.94 | 530 | A |
| 15 | RMD (Cream) | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIB(pB171), IncFII(pCoo) | 0.95 | 693 | B1 |
| 16 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, mdtM | - | IncFIB(AP001918), IncFII(pCoo) | 0.93 | 513 | E |
| 17 | RM- NRCP | Commensal | Amp, Tet | - | - | blaTEM-30 | - | - | tet(A) | - | acrF, emrE, mdtM | - | IncFIB(AP001918), IncFII | 0.86 | 54 | C |
| 18 | RM- NRCP | Commensal | Amp, Chl, Smx, Tet, Tmp | aph(3')-Ia, aph(3'')-Ib, aph(6)-Id | floR | blaOXA-1 | - | sul1, sul2 | tet(B) | dfrA36 | acrF, emrE, mdtM | qacED1 | IncFIB(AP001918) | 0.86 | 56 | C |
| 19 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIB(pB171), IncFII(pCoo) | 0.94 | 692 | B1 |
| 20 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIA, IncFIB(AP001918), IncFII(29), IncI2(Delta) | 0.94 | 720 | B1 |
| 21 | RM- FBO | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE, mdtM | - | IncFIB(AP001918) | 0.94 | 671 | B1 |
| 22 | RM- FBO | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, mdtM | - | IncFIB(AP001918), IncFIC(FII), IncI1-I(Alpha) | 0.93 | 754 | B1 |
| 23 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | emrE, emrE, mdtM | - | IncFIA, IncFIB(AP001918), IncFII(29), IncX4 | 0.94 | 701 | B1 |
| 24 | RMD (Cream) | FQ | Cip, Nal, Smx, Tmp | - | - | - | gyrA_S83L | sul2 | - | dfrA36 | acrF | - | None | 0.96 | 305 | A |
| 25 | RM- FBO | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, mdtM | - | IncFIB(AP001918), IncFII(pSE11) | 0.94 | 732 | B1 |
| 26 | RMD (Cheese) | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF | - | IncFIA, IncFIB(AP001918), IncFIC(FII) | 0.94 | 642 | A |
| 27 | RM- FBO | Commensal | Fully susceptible | - | - | - | - | - | - | - | acrF | - | IncFIB(AP001918), IncFIC(FII), IncI1-I(Alpha) | 0.93 | 755 | B1 |
| 28 | RM- FBO | ESBL | Amp, Ctx, Caz, Chl, Fep, Gen, Smx, Tet, Tmp | ant(2'')-Ia, ant(2'')-Ia, aph(3')-Ia, aadA1, aph(3'')-Ib, aph(6)-Id | floR | blaCTX-M-3 | - | sul1, sul2 | tet(A) | dfrA1, dfrA36 | acrF, mdtM | qacED1 | IncFIB(AP001918), IncFII | 0.96 | 289 | B1 |
| 29 | RM- FBO | Commensal | Amp, Smx, Tet | aph(3'')-Ib, aph(6)-Id | - | blaTEM-1 | - | - | tet(A) | - | acrF, emrE, emrE, mdtM | None | IncFIB(AP001918), IncFIC(FII), IncI1-I(Alpha) | 0.94 | 736 | B1 |
| 30 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF, emrE | None | IncFIA, IncFII | 0.95 | 720 | B1 |
| 31 | RM- NRCP | Commensal | Fully Susceptible | - | - | - | - | - | - | - | acrF | None | None | 0.95 | 526 | A |
| 32 | RM- NRCP | FQ | Amp, Chl, Cip, Nal, Smx, Tet, Tmp | aadA5, aph(3'')-Ib, aph(6)-Id | catA1, floR | blaTEM-1 | gyrA_D87N, gyrA_S83L, parC_A56T, parC_S80I | sul1 | tet(B) | dfrA17 | acrF | qacED1 | IncFIB(AP001918), IncFIC(FII) | 0.86 | 54 | A |
FQ: fluoroquinolone resistant; RM-FBO: raw milk from food business operator; RM -NRCP: raw milk from the national residue control plan; RMD: raw milk from dairy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



