Submitted:
28 January 2026
Posted:
30 January 2026
You are already at the latest version
Abstract
Chordomas are rare, highly morbid tumors arising from notochordal progenitor cells along the spinal axis, associated with severe neurological complications and high recurrence rates. Their resistance to conventional therapies and limited options beyond surgical resection and high-dose radiation underscore the urgent need for novel therapeutic targets. Publicly available preliminary RNA sequencing data from the Chordoma Foundation identified chitinase-3-like 1 (CHI3L1), a secreted glycoprotein implicated in immune checkpoint regulation and epithelial-mesenchymal transition (EMT), as a promising candidate for chordoma immunotherapy. Yet, the comprehensive function of CHI3L1 in chordoma immune response remains unclear. To evaluate its presence in chordoma, we employed RNA-based analyses alongside enzyme-linked immunosorbent assays (ELISA) on commercially available chordoma cell lines (JHC7, U-CH12, U-CH1, U-CH1-N) and human chordoma tumor specimens. Our results demonstrate elevated CHI3L1 expression in chordoma cells relative to notochordal precursors, with comparative analyses revealing higher CHI3L1 expression in the primary tumor relative to recurrent samples. These findings suggest the potential role of CHI3L1 in chordoma tumorigenesis, emphasizing its relevance as a biomarker and therapeutic target for primary tumors. Future studies are necessary to elucidate the mechanistic role of CHI3L1 in chordoma immune evasion and to explore targeted interventions that may improve patient outcomes in this aggressive cancer.
Keywords:
chordoma
; chitinase-3-like 1
; tumorigenesis
; cancer
1. Introduction
1.1. Chordoma
Characterized by high local recurrence and distant metastasis after resection, chordomas are rare and slow-growing tumors of the spinal axis notable for their limited treatment modalities (1–3). Most often diagnosed in patients between 40 and 60 years of age, these highly morbid tumors are believed to arise from notochordal progenitor cells along the spinal axis and, thus, customarily manifest at the skull base, mobile spine, and sacrum (4,5). Clinical presentation varies based on the primary anatomic location of the tumor, though pain and neurological complications specific to the region are commonly indicated (6–10). Historically, overall survival rates decline to 62% at five years, then further decrease to 33% at ten, with recurrent and metastatic disease significantly diminishing patient outcomes (11,12).
Modern treatment paradigms primarily rely on maximal en bloc surgical resection followed by high-dose photon and proton radiation (13–22). Given that half of all resected chordomas reportedly recur after five years, and nearly 90% after ten, a substantial challenge then becomes evident: after exhausting maximal safe doses of radiation and surgical interventions, no proven clinical treatments remain (23). Despite advances in radiation and surgery, chordomas remain resistant to chemotherapy, highlighting the urgent need for targeted therapeutic agents and the growing interest in exploring molecular treatments that target specific signaling pathways (24).
Molecular markers have emerged as a pivotal frontier in chordoma treatment, offering significant potential to transform surgical decision-making and enhance patient outcomes (25–28). These markers are increasingly vital for accurate diagnosis and subtyping, guiding personalized treatment selection and prognostication. By providing critical insights into tumor behavior and assessing the likely course of the disease, they may help tailor the extent of surgery and inform multidisciplinary management.
1.2. Chitinase-3-Like 1
Immune checkpoints, one of the numerous avenues through which the immune system regulates cancer progression, may be exploited by tumor cells in order to evade the host immune response (29,30). Chitinase-3-like 1 (CHI3L1, also known as YKL-40), a member of the 18 glycosyl hydrolase (18-GH) family, is a secreted glycoprotein endogenously expressed in various cells, including immune cells (31). CHI3L1 has been correlated with poor prognosis and decreased survival rates of cancer patients, and implicated in mechanisms leading to the immunosuppressive tumor environment (32–35). In lung cancer, for instance, CHI3L1 modulates PD-L1 expression in immune cells, leading to PD-1-mediated T-cell inactivation and fostering an immunosuppressive tumor microenvironment (36). Furthermore, its role in enhancing tumor epithelial-mesenchymal transition (EMT) makes it an ideal target for cancer immunotherapy (37).
Preliminary studies in our laboratory have noted differential expression between human notochordal (where the cell of origin, notochordal cells, reside) and chordoma tissue and cell lines. Initial RNA sequencing analysis of publicly available transcriptome data from the Chordoma Foundation, derived from tissue deposited by the Xavier Lab at the Broad Institute, provides evidence for the expression of CHI3L1 (Figure 1). There is a substantial expression of CHI3L1 in chordoma tissue and a more discrete expression in notochordal tissue. However, its detailed involvement with the immune response has yet to be profiled in chordoma.
In light of this data, our objective is to assess the presence of CHI3L1 in human chordoma tissue and cell lines, with long-term aims encompassing the elaboration of its involvement in chordoma tumorigenesis. Given its potential contribution to the tumor’s aggressive behavior and high rates of recurrence in this manner, we believe that CHI3L1 may serve as a promising therapeutic target for immunotherapy. This study will be the first extensive study that intends to evaluate the capabilities of CHI3L1 as a biomarker of disease progression and potential target for novel therapeutic interventions in human chordoma.
2. Materials and Methods
To confirm the expression of CHI3L1 in chordoma tissue, we investigated its presence in tumor samples excised from patients and in commercially available chordoma cell lines (Figure 2).
2.1. Human Samples
Chordoma tissue samples were obtained from patients at a single institution and stored in a de-identified manner in accordance with institutional protocols. Institutional Review Board approval was established for both fresh frozen tissue collection (Protocol 862559) and acquisition of formalin-fixed, paraffin-embedded (FFPE) samples (Protocol 816619). Fresh tissue was apportioned and either stored by the surgical pathology department as FFPE cassettes or transported directly from the operating room to the laboratory in liquid nitrogen. FFPE scrolls were cut at 10um each. All samples were stored at -80°C until used for nucleic acid work.
2.2. Cell Culture
All commercial chordoma cell lines were purchased from the American Type Culture Collection (Manassas, VA, USA) and seeded in T-75 flasks (Fisher Scientific Cat. 07-202-001). JHC7 primary sacral chordoma cells were cultured in DMEM/F12 (ATCC Cat. 30-2006) media (40); primary U-CH12 sacral chordoma cells in IMDM/RPMI (ATCC Cat. 30-2005/30-2001), L-glutamine (ATCC Cat. 30-2214), and ITS-G (Fisher Scientific Cat. 41-400-045) (41); chordoma-derived U-CH1-N notochordal cells in Minimum Essential Medium Eagle (Sigma Aldrich StableCell Cat. M6074-500ML) media (42); and U-CH1, recurrent sacrococcygeal chordoma cells, in IMDM/RPMI and L-glutamine, with collagen coating (Corning Cat. 354236) as per supplier instructions (43–47). Media was supplemented with 10% fetal bovine serum (Cat. 30-2020) and 1% penicillin-streptomycin solution (Cat. 30-2300) from ATCC.
For all cell lines, once confluent, supernatant was collected from centrifuged media that had been utilized to culture the cells for an average of 1 week. Supernatant was stored at 4°C until analyzed via ELISA. Cells were flash-frozen to form 5 x 105 cell pellets, after which total RNA was extracted.
2.3. RNA Extraction
Total RNA from FFPE scrolls was isolated as instructed using the RNeasy FFPE Kit (Cat. 73504) from Qiagen. Total RNA from cell lines was isolated using TRIZOL Reagent (Cat. 15596026) from ThermoFisher Scientific. To validate RNA integrity and purity and select the highest-quality samples for downstream analyses, we conducted picochip and nanodrop assays to evaluate DV200 values, as well as the 260/280 and 260/230 ratios, respectively.
2.4. NanoString
Total RNA from 7 patient FFPE chordoma specimens (4 primary, 3 recurrent) was isolated according to the above procedure. RNA was sent for quantification of 770 genes using the NanoString nCounter PanCancer Pathways Panel. Differential gene expression was performed in NanoString’s proprietary user interface, nSolver, by selecting each of our samples for Advanced Analysis and using the default parameters. Genes were identified as differentially expressed if the gene exhibited |fold-change| > 0 and padj < 0.01. Results were visualized in RStudio using EnhancedVolcano and pheatmap. All scripts for data analysis and image generation can be found at this link <https://github.com/j-rdt/chi3l1-chordoma>.
2.5. Bulk RNA Sequencing
Total RNA from 5 patient chordoma specimens (2 primary from Chordoma Foundation, 1 locally-retrieved primary, 2 recurrent from Chordoma Foundation) was isolated according to the above procedure. RNA was sent to GeneWiz for bulk sequencing with rRNA depletion, ERCC spike-in, and read depth of 20M reads/sample. Data were stripped of identifiable personal health information. Bioinformatic analyses were performed in RStudio, using the DESeq2 package for differential gene expression analysis, EnhancedVolcano for visualizing the differential expression for each gene, and pheatmap for visualizing gene expression levels in each sample; default parameters were used for all packages (48–50). False discovery rate p-value adjustment was performed using the Benjamini-Hochberg procedure. Genes were identified as differentially expressed if the gene exhibited |log2(fold-change)| > 2 and padj < 0.01.
2.6. RT-PCR & qPCR
Reverse transcription (RT-PCR) & quantitative polymerase chain reaction (qPCR) were performed on RNA from primary sacral chordoma cell lines JHC7 (n = 8) and U-CH12 (n = 9), recurrent chordoma cell line U-CH1 (n = 12), and chordoma-derived notochordal-like cell line U-CH1-N (n = 7). RT-PCR was carried out using the Invitrogen™ SuperScript™ III First-Strand Synthesis System (ThermoFisher Scientific Cat. 18080051), and qPCR by the SYBR Green method (Applied Biosystems™ Cat. A25742). Primers were as follows: CHI3L1, forward 5’-GATGTGACGCTCTACGGCAT-3’ and reverse 5’-TGATAAAGTCCGGCGACTC-3’; GAPDH, forward 5’-GAGAAGGCTGGGGCTCATTTG-3’ and reverse 5’-GGTGCTAAGCAGTTGGTGGT-3’ (51). Samples without detectable amplification were assigned a maximum threshold cycle (Ct) value of 40 for normalization purposes. Relative expression (RE) values of CHI3L1 mRNA were calculated relative to housekeeping gene GAPDH in each sample using the 2-ΔCt method.
2.7. ELISA
CHI3L1 enzyme-linked immunosorbent assay (ELISA) measurements were obtained using the Human Chitinase 3-like 1/YKL-40 Quantikine ELISA Kit (R&D Systems Cat. DC3L10). Supernatant was collected from JHC7 (n = 6), U-CH12 (n = 9), U-CH1 (n = 9), and U-CH1-N (n = 6) cell lines according to the protocol described above. The ELISA plate included duplicates of controls, samples, and standards. Each well contained 50µL of its respective solution and was incubated for 2 hours at room temperature with the assay diluent RD1-34 from the kit. Post-incubation, wells were washed 4 times with the kit-supplied wash buffer and incubated overnight with the Human CHI3L1 Conjugate at 4ºC. The following day, the wells were washed 4 times with a prepared wash buffer and incubated with a substrate solution for 30 minutes at room temperature. After stop solution was added, the plate was read on the Glomax Discover Microplate Reader (Cat. GM3000) from Promega.
3. Results
3.1. Transcriptomics
The transition between primary and recurrent chordomas is hypothesized to involve the dysregulation of immune system surveillance. To assess this hypothesis, we evaluated the expression of key genes in patient-derived tumor tissue using a NanoString cancer transcriptomics panel. CHI3L1 was the most downregulated gene (Figure 3A). The higher gene expression of CHI3L1 in primary cells is an effect of one primary sample in particular (Figure 3B). Principal component analysis shows that this sample clusters with other primary cells (Figure 3C).
To validate the differential expression of CHI3L1 in chordoma, we performed whole transcriptome sequencing of human tissue provided by the Chordoma Foundation and locally retrieved at a single institution (3 primary, 2 recurrent). CHI3L1 was again the most downregulated gene in recurrent cells (Figure 4A). This result was driven by one sample in particular with extreme CHI3L1 expression (Figure 4B). Principal component analysis shows that this sample generally clusters with the other primary cells (Figure 4C). These results indicated CHI3L1 is a marker of interest in chordoma cells.
3.2. qPCR and ELISA
To assess the relative expression of CHI3L1 across chordoma cell lines, total RNA extracted from primary chordoma cell lines JHC7 and U-CH12, recurrent chordoma cell line U-CH1, and chordoma-derived notochordal-like U-CH1-N cells was used for RT-qPCR. Relative quantification showed significantly higher CHI3L1 mRNA levels in both primary chordoma cell lines compared to the recurrent U-CH1 line. Additionally, JHC7 cells exhibited significantly higher expression relative to notochordal-like U-CH1-N cells (Figure 5).
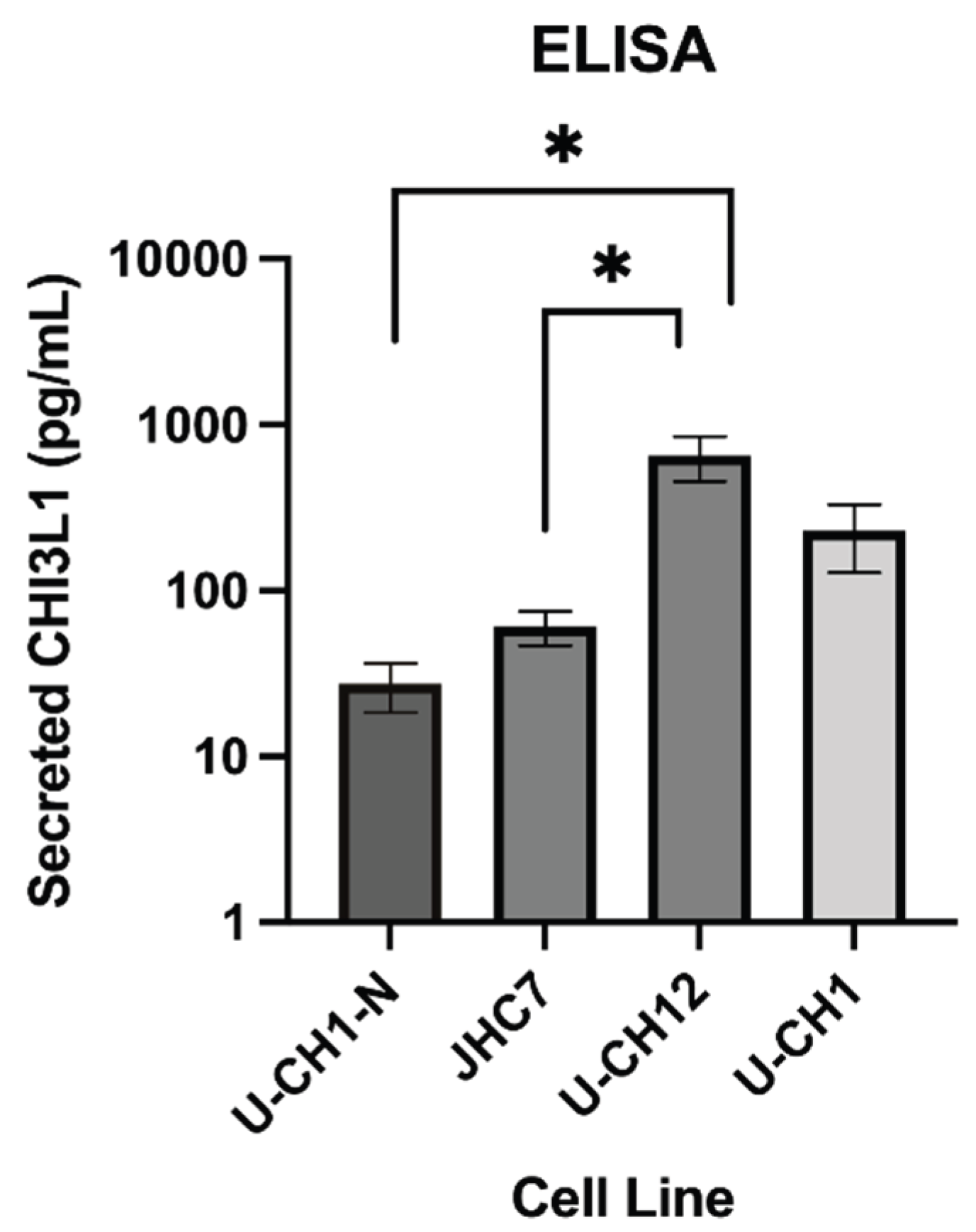
To evaluate secreted CHI3L1 levels in various chordoma cell lines and controls, an ELISA was performed on media supernatants after 1 week in culture. The results suggest higher levels of secreted CHI3L1 in the primary cell lines JHC7 and U-CH12, as well as the recurrent cell line U-CH1, compared to the low levels observed in the control notochordal UCH1-N cells (Figure 6).
4. Discussion
While en bloc resection yields the greatest long-term survival for chordoma patients, it is frequently linked to heightened morbidity. En bloc sacral amputation for sacral chordoma may result in the loss of bowel, bladder, and sexual function, along with impaired ambulation and chronic pain (15,52). In cases where sacral tumors invade the rectum, achieving negative margins to optimize patient survival may necessitate colon resection and colostomy, further diminishing quality of life. Cytoreductive chemotherapy or immunotherapy regimens aimed at reducing tumor volume may enable less invasive resections and, in turn, improve patient outcomes.
Similarly, patients with clival chordoma may also benefit from preoperative cytoreduction to facilitate surgical intervention. Although endoscopic surgical resection for clival chordoma is employed at some highly specialized centers, the anatomical complexity of the clivus often limits its success. As a result, radiation therapy remains the mainstay of treatment at many institutions (53–55). However, this procedure carries risks of cranial nerve complications, which can lead to vision loss, permanent swallowing dysfunction, and airway compromise. Given the substantial morbidity of current treatments and high recurrence rates, novel therapeutic strategies are urgently needed for patients with this disease.
Research into chordoma has historically been limited due to the tumor's rarity, with few dedicated studies and a scarcity of clinical trials. Its low incidence, coupled with the complexity of the disease, has hindered the development of targeted therapies, leaving patients with limited treatment options and poor prognoses. Current research efforts are largely focused on comprehending the molecular biology of chordoma, particularly the role of proteins such as brachyury, which is overexpressed in nearly all chordoma tumors (3). However, much remains to be understood about the underlying mechanisms driving tumor growth, recurrence, and progression.
Notably, previous studies have conflated primary and recurrent chordoma samples into a single analytical cohort, potentially obscuring meaningful molecular distinctions between these stages of disease progression. Preliminary data from our research center have begun to address this gap by exploring these stage-specific differences, isolating Chitinase-3-like 1 (CHI3L1), a glycoside hydrolase involved in tissue injury, remodeling, and repair, as a promising candidate for chordoma recurrence (31).
Dysregulated CHI3L1 expression has been documented in several solid cancers, with both circulatory and tissue levels strongly associated with chemoresistance and poor prognosis (36,56–58). As a result, CHI3L1 is increasingly regarded as a biomarker of tumor progression and therapeutic resistance. Moreover, CHI3L1 has been reported as a biomarker in bone and soft tissue sarcomas, highlighting its diagnostic and prognostic value (31,57,59). Importantly, recent investigations have identified that CHI3L1 significantly enhances the epithelial-mesenchymal transition (EMT) and promotes tumor development and metastasis, potentially through beta-catenin and MAPK signaling activation (31,60–62). Emerging evidence also implicates CHI3L1 in immune evasion through inhibition of T-cell co-stimulation via ICOS/ICOS ligand and CD28-B7 signaling, promotion of immunosuppressive myeloid cell accumulation, and suppression of tumor suppressive pathways involving PTEN and p53 (63,64).
Our study aims to address a critical gap in the chordoma research landscape by investigating the role of CHI3L1, a protein known to contribute to tumor growth and immune system evasion, in chordoma oncogenesis. By confirming the presence of CHI3L1, we seek to provide a foundation for further elucidation of the molecular mechanisms that drive this rare and aggressive cancer.
Preliminary qPCR and ELISA assays reveal elevated CHI3L1 expression in chordoma cells relative to notochordal precursor cells, underscoring its potential role in disease initiation and progression. Moreover, differential gene expression analyses between primary and recurrent chordoma cell lines and fresh human tissue samples indicate that some primary cells express elevated amounts of CHI3L1, whereas recurrent chordoma cells exhibit relatively reduced expression. This trend supports the hypothesis that CHI3L1 may play a role in the progression from notochordal progenitor cells to primary tumor cells, with its expression diminishing as chordoma cells replicate or differentiate. Taken together, these findings provide initial evidence of the importance of CHI3L1 as a molecular marker in early tumorigenesis, with possible therapeutic relevance most pronounced in the primary tumor setting.
In the long term, we aim to elucidate the mechanistic role of CHI3L1 in chordoma progression and immune evasion, with the goal of developing targeted therapeutic strategies that can impede or halt tumor growth. Robust in vitro and in vivo systems are crucial for the thorough appraisal of therapies and the appreciation of tumor-immune interactions. As such, additional studies require analysis of primary, recurrent, and metastatic cell lines, and the assembly of a broader patient sample to investigate the relevant mechanistic role of CHI3L1 across the chordoma disease progression. A larger pool of cell lines and tissue samples is already being prepared for NanoString and RNA-seq experiments at our institution to replicate these findings. Consistent with emerging evidence implicating CHI3L1 in immune evasion, ongoing studies at our center are investigating CHI3L1-mediated immune crosstalk in chordoma, with a particular focus on patient-derived CD8+ T cells and myeloid populations, using co-culture models with autologous chordoma cells and the JHC7 cell line. Proposed future research also aims to leverage in vivo animal models to elucidate the mechanisms underlying chordoma oncogenesis and evaluate therapeutic strategies. Humanized mouse models are particularly promising for exploring chordoma development and treatment within the context of the tumor microenvironment and immuno-oncology (65).
Given the rarity, aggressive nature, and limited treatment options available for chordoma, this research represents a pivotal step toward identifying new therapeutic strategies by advancing our understanding of its molecular drivers. By implicating CHI3L1 as a key element in chordoma pathogenesis and immune evasion, this study lays the groundwork for targeted interventions and frameworks that may reduce recurrence and improve long-term outcomes. As chordoma remains primarily a surgical disease, novel therapeutic approaches are especially promising and well-suited to this patient population. Understanding how CHI3L1 shapes the immunosuppressive tumor microenvironment may also inform new therapeutic avenues, particularly for recurrent or treatment-resistant cases. The molecular mechanisms central to this study may have broader relevance for other rare cancers that share similar immunological or signaling profiles, thereby extending the potential trajectory of our findings to improve survival and quality of life beyond chordoma.
Author Contributions
Conceptualization, M.M.M. and P.L.Z.S.; methodology, B.C. and C.G.; formal analysis, B.C., C.G., J.A., J.W., J.S.; investigation, B.C., C.G., J.A., J.D., K.G.; writing—original draft preparation, B.C., C.G., J.A., J.D., K.G.; writing—review and editing, B.C., C.G., A.F., C.S., M.J.M., W.L., M.M.M, P.L.Z.S.; supervision, J.W., J.S., M.P., S.K., J.A.E., C.L., Z.L.G., M.M.M, P.L.Z.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a Rhode Island Life Sciences Hub Small Grant, National Institutes of Health IDeA-CTR # U54GM115677, National Institutes of Health T35 Training Grant # HL094308, National Institute of General Medical Sciences of the National Institutes of Health IDeA # P20GM109035, and departmental funding from Brown University Health Neurosurgery.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (Protocol 862559, approved August 22, 2016; Protocol 816619, approved November 13, 2015).
Informed Consent Statement
All samples used in the study were either obtained for use in research with written informed consent from prospective registry participants, or accessed with local IRB approval to use historical tissue archives with waiver of informed consent for retrospective research.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author, as the complete data cannot be made available due to privacy restrictions. A formal institutional data-sharing request is required.
Acknowledgments
The authors would like to thank Christoph Schorl of the Genomics Core Facility, a Brown University BioMed Core Facility, for his assistance and guidance in the RNA analysis studies. The graphical abstract and Figure 2 were both created in BioRender: Campilan, B. (2026) https://biorender.com/k17pl2t and https://biorender.com/omo5qlm, respectively.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| CHI3L1 EMT FFPE qPCR RT-PCR ELISA MDPI |
Chitinase-3-like 1 Epithelial-mesenchymal transition Formalin-fixed, paraffin-embedded Quantitative polymerase chain reaction Reverse transcriptase polymerase chain reaction Enzyme-linked immunosorbent assay Multidisciplinary Digital Publishing Institute |
References
- Stacchiotti, S; Sommer, J. Building a global consensus approach to chordoma: a position paper from the medical and patient community. Lancet Oncol. 2015, 16(2), e71–83. [Google Scholar] [CrossRef]
- Tenny, S; Varacallo, M. Chordoma. In StatPearls [Internet]; StatPearls Publishing: Treasure Island (FL), 2024; Available online: http://www.ncbi.nlm.nih.gov/books/NBK430846/.
- Walcott, BP; Nahed, BV; Mohyeldin, A; Coumans, JV; Kahle, KT; Ferreira, MJ. Chordoma: current concepts, management, and future directions. Lancet Oncol. 2012, 13(2), e69–76. [Google Scholar] [CrossRef]
- Bakker, SH; Jacobs, WCH; Pondaag, W; Gelderblom, H; Nout, RA; Dijkstra, PDS; et al. Chordoma: a systematic review of the epidemiology and clinical prognostic factors predicting progression-free and overall survival. Eur Spine J 2018, 27(12), 3043–58. [Google Scholar] [CrossRef]
- Ulici, V; Hart, J. Chordoma. Arch Pathol Lab Med. 2022, 146(3), 386–95. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, JL; Espachs, P; Roca, G; Samper, D; Cubells, C; Vidal, F. Difficult management of pain following sacrococcygeal chordoma: 13 months of subarachnoid infusion. Pain 1994, 59(2), 317–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, S; Ulloa, R; Soffer, J; Alcazar-Felix, RJ; Snyderman, CH; Gardner, PA; et al. Chordoma: A Comprehensive Systematic Review of Clinical Trials. Cancers 2023, 15(24), 5800. [Google Scholar] [CrossRef]
- Court, C; Briand, S; Mir, O; Le Péchoux, C; Lazure, T; Missenard, G; et al. Management of chordoma of the sacrum and mobile spine. Orthop Traumatol Surg Res. 2022, 108(1), 103169. [Google Scholar] [CrossRef] [PubMed]
- Gardner, PA; Froelich, S; Gokaslan, ZL; MacDonald, SM; Peris Celda, M; Raza, SM; et al. Introduction. Chordoma: updates and advances. Neurosurg Focus 2024, 56(5), E1. [Google Scholar] [CrossRef]
- Xia, Y; Papali, P; Al-Mistarehi, AH; Hansen, LJ; Azad, TD; Ahmed, AK; et al. Outcomes After Definitive Surgery for Spinal and Sacral Chordoma in 101 Patients Over 20 Years. Neurosurgery [Internet]. 5 Aug 2024. Available online: https://journals.lww.com/10.1227/neu.0000000000003130.
- McPherson, CM; Suki, D; McCutcheon, IE; Gokaslan, ZL; Rhines, LD; Mendel, E. Metastatic disease from spinal chordoma: a 10-year experience. J Neurosurg Spine 2006, 5(4), 277–80. [Google Scholar] [CrossRef]
- Mukherjee, D; Chaichana, KL; Gokaslan, ZL; Aaronson, O; Cheng, JS; McGirt, MJ. Survival of patients with malignant primary osseous spinal neoplasms: results from the Surveillance, Epidemiology, and End Results (SEER) database from 1973 to 2003: Clinical article. J Neurosurg Spine 2011, 14(2), 143–50. [Google Scholar] [CrossRef]
- Boriani, S; Bandiera, S; Biagini, R; Bacchini, P; Boriani, L; Cappuccio, M; et al. Chordoma of the Mobile Spine: Fifty Years of Experience: Spine. 2006, 31(4), 493–503. [Google Scholar] [CrossRef]
- Catton, C; O’Sullivan, B; Bell, R; Laperriere, N; Cummings, B; Fornasier, V; et al. Chordoma: long-term follow-up after radical photon irradiation. Radiother Oncol. 1996, 41(1), 67–72. [Google Scholar] [CrossRef]
- Fourney, DR; Rhines, LD; Hentschel, SJ; Skibber, JM; Wolinsky, JP; Weber, KL; et al. En bloc resection of primary sacral tumors: classification of surgical approaches and outcome. J Neurosurg Spine 2005, 3(2), 111–22. [Google Scholar] [CrossRef] [PubMed]
- Frezza, AM; Botta, L; Trama, A; Dei Tos, AP; Stacchiotti, S. Chordoma: update on disease, epidemiology, biology and medical therapies. Curr Opin Oncol. 2019, 31(2), 114–20. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B. Operative Management of Sacral Chordoma. J Bone Jt Surg Am. 2005, 87(10), 2211. [Google Scholar]
- Gokaslan, ZL; Zadnik, PL; Sciubba, DM; Germscheid, N; Goodwin, CR; Wolinsky, JP; et al. Mobile spine chordoma: results of 166 patients from the AOSpine Knowledge Forum Tumor database. J Neurosurg Spine 2016, 24(4), 644–51. [Google Scholar] [CrossRef]
- Lockney, DT; Shub, T; Hopkins, B; Lockney, NA; Moussazadeh, N; Lis, E; et al. Spinal stereotactic body radiotherapy following intralesional curettage with separation surgery for initial or salvage chordoma treatment. Neurosurg Focus 2017, 42(1), E4. [Google Scholar] [CrossRef]
- Moojen, WA; Vleggeert-Lankamp, CLA; Krol, ADG; Dijkstra, SPD. Long-term Results: Adjuvant Radiotherapy in En Bloc Resection of Sacrococcygeal Chordoma is Advisable. Spine 2011, 36(10), E656–61. [Google Scholar] [CrossRef]
- Schroeder, C; De Lomba, WC; Leary, OP; De La Garza Ramos, R; Gillette, JS; Miner, TJ; et al. Multidisciplinary surgical considerations for en bloc resection of sacral chordoma: review of recent advances and a contemporary single-center series. Neurosurg Focus 2024, 56(5), E7. [Google Scholar] [CrossRef]
- Yamamoto, N; Tsuchiya, H. Treatment of chordoma—where is it going? J Spine Surg. 2019, 5(3), 387–9. [Google Scholar] [CrossRef]
- Wu, Z; Zhang, J; Zhang, L; Jia, G; Tang, J; Wang, L; et al. Prognostic factors for long-term outcome of patients with surgical resection of skull base chordomas—106 cases review in one institution. Neurosurg Rev. 2010, 33(4), 451–6. [Google Scholar] [CrossRef]
- Martinez Moreno, M; Wang, E; Schroeder, C; Sullivan, P; Gokaslan, Z. Shedding light on emerging therapeutic targets for chordoma. Expert Opin Ther Targets 2023, 27(8), 705–13. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C; Campilan, B; Leary, OP; Arditi, J; Michles, MJ; De La Garza Ramos, R; et al. Therapeutic Opportunities for Biomarkers in Metastatic Spine Tumors. Cancers 2024, 16(18), 3152. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y; Zhang, H. Immune microenvironment and immunotherapy for chordoma. Front Oncol. 2024, 14, 1374249. [Google Scholar] [CrossRef]
- Dridi, M; Krebs-Drouot, L; Meyronet, D; Dumollard, JM; Vassal, F; Jouanneau, E; et al. The Immune Microenvironment of Chordomas: An Immunohistochemical Analysis. Cancers 2021, 13(13), 3335. [Google Scholar] [CrossRef]
- Godinez, C; Campilan, B; Schroeder, C; Arditi, J; Michles, MJ; Herrera, BC; et al. Combinatorial therapies for epigenetic, immunotherapeutic, and genetic targeting of chordoma. J Neurooncol [Internet]. 31 Dec 2024. Available online: https://link.springer.com/10.1007/s11060-024-04920-y.
- Patel, SS; Schwab, JH. Immunotherapy as a Potential Treatment for Chordoma: a Review. Curr Oncol Rep. 2016, 18(9), 55. [Google Scholar] [CrossRef] [PubMed]
- Bishop, AJ; Amini, B; Lin, H; Raza, SM; Patel, S; Grosshans, DR; et al. Immune Checkpoint Inhibitors Have Clinical Activity in Patients With Recurrent Chordoma. J Immunother. 2022, 45(8), 374–8. [Google Scholar] [CrossRef]
- Zhao, T; Su, Z; Li, Y; Zhang, X; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct Target Ther 2020, 5(1), 201. [Google Scholar] [CrossRef]
- Yang, X; Fang, D; Li, M; Chen, J; Cheng, Y; Luo, J. Knockdown of Chitinase 3-Like-1 Inhibits Cell Proliferation, Promotes Apoptosis, and Enhances Effect of Anti-Programmed Death Ligand 1 (PD-L1) in Diffuse Large B Cell Lymphoma Cells. Med Sci Monit [Internet]. 27 Jan 2021, p. 27. Available online: https://www.medscimonit.com/abstract/index/idArt/929431.
- Libreros, S; Garcia-Areas, R; Iragavarapu-Charyulu, V. CHI3L1 plays a role in cancer through enhanced production of pro-inflammatory/pro-tumorigenic and angiogenic factors. Immunol Res 2013, 57(1–3), 99–105. [Google Scholar] [CrossRef]
- Libreros, S; Garcia-Areas, R; Shibata, Y; Carrio, R; Torroella-Kouri, M; Iragavarapu-Charyulu, V. Induction of proinflammatory mediators by CHI3L1 is reduced by chitin treatment: Decreased tumor metastasis in a breast cancer model. Int J Cancer 2012, 131(2), 377–86. [Google Scholar] [CrossRef]
- Kim, DH; Park, HJ; Lim, S; Koo, JH; Lee, HG; Choi, JO; et al. Regulation of chitinase-3-like-1 in T cell elicits Th1 and cytotoxic responses to inhibit lung metastasis. Nat Commun. 2018, 9(1), 503. [Google Scholar] [CrossRef]
- Ma, B; Akosman, B; Kamle, S; Lee, CM; He, CH; Koo, JS; et al. CHI3L1 regulates PD-L1 and anti-CHI3L1-PD-1 antibody elicits synergistic antitumor responses. J Clin Invest. 2021, 131(21), e137750. [Google Scholar] [CrossRef]
- Guetta-Terrier, C; Karambizi, D; Akosman, B; Zepecki, JP; Chen, JS; Kamle, S; et al. Chi3l1 Is a Modulator of Glioma Stem Cell States and a Therapeutic Target in Glioblastoma. Cancer Res. 2023, 83(12), 1984–99. [Google Scholar] [CrossRef]
- Dobin, A; Davis, CA; Schlesinger, F; Drenkow, J; Zaleski, C; Jha, S; et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29(1), 15–21. [Google Scholar] [CrossRef] [PubMed]
- Robinson, MD; McCarthy, DJ; Smyth, GK. edgeR : a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26(1), 139–40. [Google Scholar] [CrossRef]
- Hsu, W; Mohyeldin, A; Shah, SR; Ap Rhys, CM; Johnson, LF; Sedora-Roman, NI; et al. Generation of chordoma cell line JHC7 and the identification of Brachyury as a novel molecular target: Laboratory investigation. J Neurosurg. 2011, 115(4), 760–9. [Google Scholar] [CrossRef] [PubMed]
- Von Witzleben, A; Goerttler, LT; Marienfeld, R; Barth, H; Lechel, A; Mellert, K; et al. Preclinical Characterization of Novel Chordoma Cell Systems and Their Targeting by Pharmocological Inhibitors of the CDK4/6 Cell-Cycle Pathway. Cancer Res. 2015, 75(18), 3823–31. [Google Scholar] [CrossRef]
- Fujita, N; Suzuki, S; Watanabe, K; Ishii, K; Watanabe, R; Shimoda, M; et al. Chordoma-derived cell line U-CH1-N recapitulates the biological properties of notochordal nucleus pulposus cells. J Orthop Res. 2016, 34(8), 1341–50. [Google Scholar] [CrossRef]
- Karikari, IO; Gilchrist, CL; Jing, L; Alcorta, DA; Chen, J; Richardson, WJ; et al. Molecular characterization of chordoma xenografts generated from a novel primary chordoma cell source and two chordoma cell lines: Laboratory investigation. J Neurosurg Spine 2014, 21(3), 386–93. [Google Scholar] [CrossRef]
- Scheil, S; Brüderlein, S; Liehr, T; Starke, H; Herms, J; Schulte, M; et al. Genome-wide analysis of sixteen chordomas by comparative genomic hybridization and cytogenetics of the first human chordoma cell line, U-CH1. Genes Chromosomes Cancer 2001, 32(3), 203–11. [Google Scholar] [CrossRef] [PubMed]
- Brüderlein, S; Sommer, JB; Meltzer, PS; Li, S; Osada, T; Ng, D; et al. Molecular Characterization of Putative Chordoma Cell Lines. Sarcoma 2010, 2010, 1–14. [Google Scholar] [CrossRef]
- Presneau, N; Shalaby, A; Ye, H; Pillay, N; Halai, D; Idowu, B; et al. Role of the transcription factor T (brachyury) in the pathogenesis of sporadic chordoma: a genetic and functional-based study. J Pathol. 2011, 223(3), 327–35. [Google Scholar] [CrossRef]
- Liu, X; Nielsen, GP; Rosenberg, AE; Waterman, PR; Yang, W; Choy, E; et al. Establishment and characterization of a novel chordoma cell line: CH22. J Orthop Res. 2012, 30(10), 1666–73. [Google Scholar] [CrossRef]
- Love, MI; Huber, W; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15(12), 550. [Google Scholar] [CrossRef] [PubMed]
- Kolde, R. pheatmap: Pretty Heatmaps [Internet]. 2025. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html.
- Blighe, K. kevinblighe/EnhancedVolcano [Internet]. 2026. Available online: https://github.com/kevinblighe/EnhancedVolcano.
- Liu, K; Jin, M; Ye, S; Yan, S. CHI3L1 promotes proliferation and improves sensitivity to cetuximab in colon cancer cells by down-regulating p53. J Clin Lab Anal. 2020, 34(1), e23026. [Google Scholar] [CrossRef] [PubMed]
- Devin, C; Chong, PY; Holt, GE; Feurer, I; Gonzalez, A; Merchant, N; et al. Level-adjusted perioperative risk of sacral amputations. J Surg Oncol. 2006, 94(3), 203–11. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Miranda, JC; Gardner, PA; Snyderman, CH; Devaney, KO; Mendenhall, WM; Suárez, C; et al. Clival chordomas: A pathological, surgical, and radiotherapeutic review: Clival chordomas. In Head Neck; Eisele, DW, Ed.; June 2014; Volume 36, 6, pp. 892–906. [Google Scholar]
- Snyderman, CH; Gardner, PA. Current opinion in otolaryngology and head and neck surgery: clival chordoma and its management. Curr Opin Otolaryngol Head Neck Surg 2020, 28(2), 118–21. [Google Scholar] [CrossRef]
- Zhai, Y; Bai, J; Wang, S; Du, J; Wang, J; Li, C; et al. Differences in Dural Penetration of Clival Chordomas Are Associated with Different Prognosis and Expression of Platelet-Derived Growth Factor Receptor-β. World Neurosurg. 2017, 98, 288–95. [Google Scholar] [CrossRef]
- Chiang, YC; Lin, HW; Chang, CF; Chang, MC; Fu, CF; Chen, TC; et al. Overexpression of CHI3L1 is associated with chemoresistance and poor outcome of epithelial ovarian carcinoma. Oncotarget 2015, 6(37), 39740–55. [Google Scholar] [CrossRef]
- Johansen, JS; Schultz, NA; Jensen, BV. Plasma YKL-40: A Potential New Cancer Biomarker? Future Oncol. 2009, 5(7), 1065–82. [Google Scholar] [CrossRef]
- Choi, IK; Kim, YH; Kim, JS; Seo, JH. High serum YKL-40 is a poor prognostic marker in patients with advanced non-small cell lung cancer. Acta Oncol. 2010, 49(6), 861–4. [Google Scholar] [CrossRef] [PubMed]
- Thorn, AP; Harving, ML; Lausten, GS; Johansen, JS; Sørensen, MS; Petersen, MM. Plasma YKL-40 as a biomarker in patients with nonmetastatic bone and soft tissue sarcomas: a prospective exploratory clinical study. Int J Surg Oncol [Internet] 2020 June [cited 2025 July 2, 5(3). Available online: https://journals.lww.com/10.1097/IJ9.0000000000000087. [CrossRef]
- Geng, B; Pan, J; Zhao, T; Ji, J; Zhang, C; Che, Y; et al. Chitinase 3-like 1-CD44 interaction promotes metastasis and epithelial-to-mesenchymal transition through β-catenin/Erk/Akt signaling in gastric cancer. J Exp Clin Cancer Res. 2018, 37(1), 208. [Google Scholar] [CrossRef] [PubMed]
- Ma, B; Herzog, EL; Lee, CG; Peng, X; Lee, CM; Chen, X; et al. Role of Chitinase 3–like-1 and Semaphorin 7a in Pulmonary Melanoma Metastasis. Cancer Res. 2015, 75(3), 487–96. [Google Scholar] [CrossRef]
- Song, Z; Chen, E; Qian, J; Xu, J; Cao, G; Zhou, W; et al. Serum chitinase activity prognosticates metastasis of colorectal cancer. BMC Cancer 2019, 19(1), 629. [Google Scholar] [CrossRef]
- Ma, B; Kamle, S; Akosman, B; Khan, H; Lee, CM; Lee, CG; et al. CHI3L1 enhances melanoma lung metastasis via regulation of T cell co-stimulators and CTLA-4/B7 axis. Front Immunol. 2022, 13, 1056397. [Google Scholar] [CrossRef]
- Ma, B; Kamle, S; Sadanaga, T; Lee, CM; Lee, JH; Yee, DC; et al. Chitinase 3-like-1 Inhibits Innate Antitumor and Tissue Remodeling Immune Responses by Regulating CD47-SIRPα- and CD24-Siglec10-Mediated Phagocytosis. J Immunol Baltim Md 1950 2024, 213(9), 1279–91. [Google Scholar] [CrossRef]
- Campilan, B; Schroeder, C; Sagaityte, E; Arditi, J; Leary, OP; Gokaslan, ZL; et al. Animal model considerations for chordoma research: reproducing the tumor microenvironment in vivo with humanized mice. Front Oncol. 2024, 14, 1330254. [Google Scholar] [CrossRef]
Figure 1.
CHI3L1 gene expression in publicly available transcriptome data from human tissue and cell lines. Transcriptome data using the chordoma cell lines U-CH1 recurrent tumor (red, n = 3) and JHC7 primary tumor (blue, n = 2) or primary tissue samples from chordoma (yellow, n = 5) or notochord (green, n = 3) were assessed for expression of the CHI3L1 gene. All datasets were aligned to the GRCH38 human reference genome assembly using STAR version 2.6.1b, and libraries were normalized using the Bioconductor package edgeR (38,39). Distribution of the observed expression in each group is displayed in the above boxplot with the observed expression level (counts per million reads) for each sample overlaid. For human tissue samples, primary versus recurrent tumor status was unknown.
Figure 1.
CHI3L1 gene expression in publicly available transcriptome data from human tissue and cell lines. Transcriptome data using the chordoma cell lines U-CH1 recurrent tumor (red, n = 3) and JHC7 primary tumor (blue, n = 2) or primary tissue samples from chordoma (yellow, n = 5) or notochord (green, n = 3) were assessed for expression of the CHI3L1 gene. All datasets were aligned to the GRCH38 human reference genome assembly using STAR version 2.6.1b, and libraries were normalized using the Bioconductor package edgeR (38,39). Distribution of the observed expression in each group is displayed in the above boxplot with the observed expression level (counts per million reads) for each sample overlaid. For human tissue samples, primary versus recurrent tumor status was unknown.
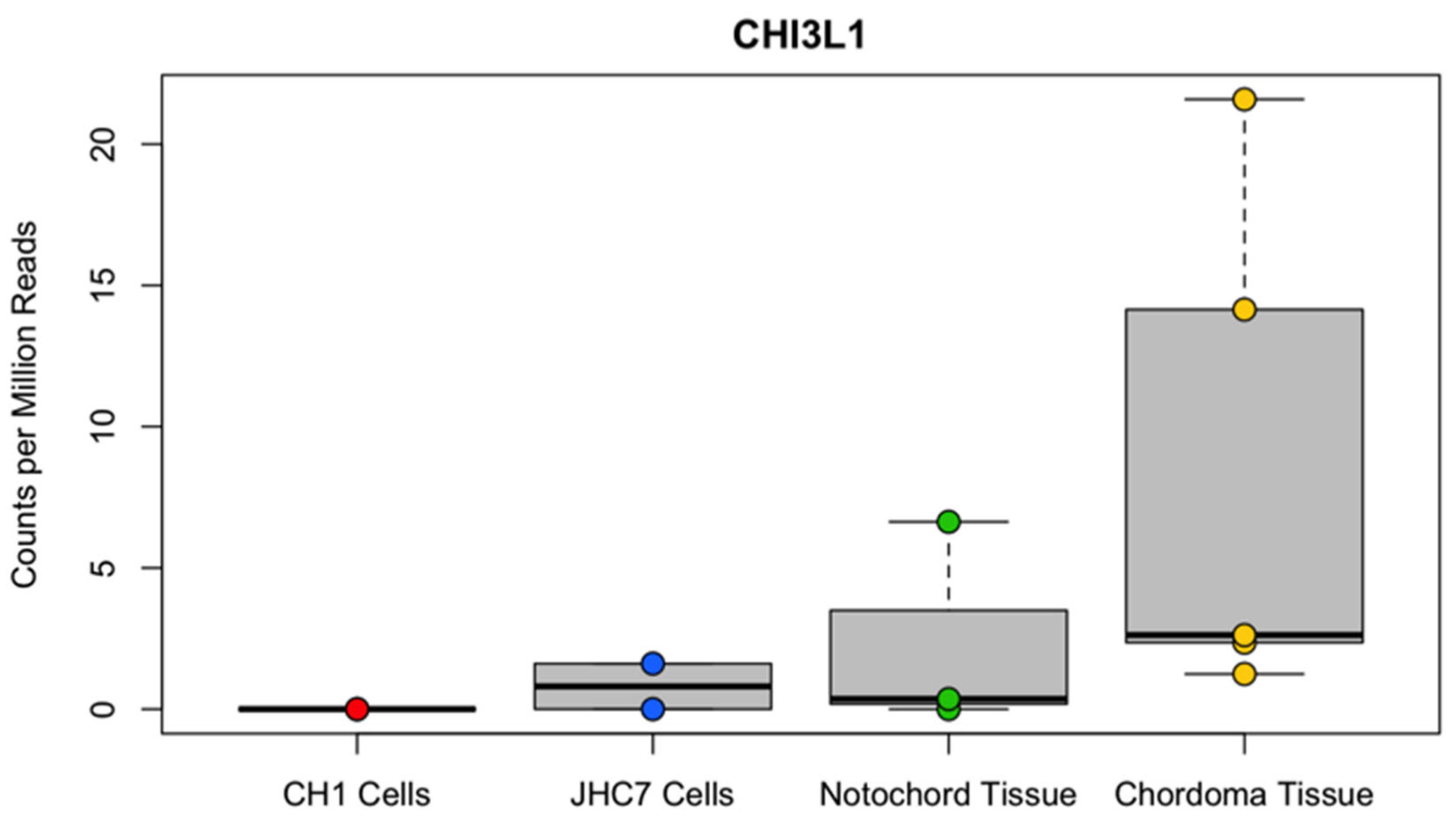
Figure 2.
Schematic of experimental methods. Bulk RNA sequencing was performed on total RNA obtained from the Chordoma Foundation and RNA extracted from intraoperatively collected patient specimens at a single institution. Total RNA derived from other formalin-fixed, paraffin-embedded (FFPE) patient samples from the same institution was profiled using NanoString. Commercially available chordoma cell lines were cultured, with extracted RNA analyzed by quantitative PCR (qPCR) and collected supernatant assessed by enzyme-linked immunosorbent assay (ELISA). Created in BioRender. Campilan, B. (2026) https://biorender.com/omo5qlm.
Figure 2.
Schematic of experimental methods. Bulk RNA sequencing was performed on total RNA obtained from the Chordoma Foundation and RNA extracted from intraoperatively collected patient specimens at a single institution. Total RNA derived from other formalin-fixed, paraffin-embedded (FFPE) patient samples from the same institution was profiled using NanoString. Commercially available chordoma cell lines were cultured, with extracted RNA analyzed by quantitative PCR (qPCR) and collected supernatant assessed by enzyme-linked immunosorbent assay (ELISA). Created in BioRender. Campilan, B. (2026) https://biorender.com/omo5qlm.
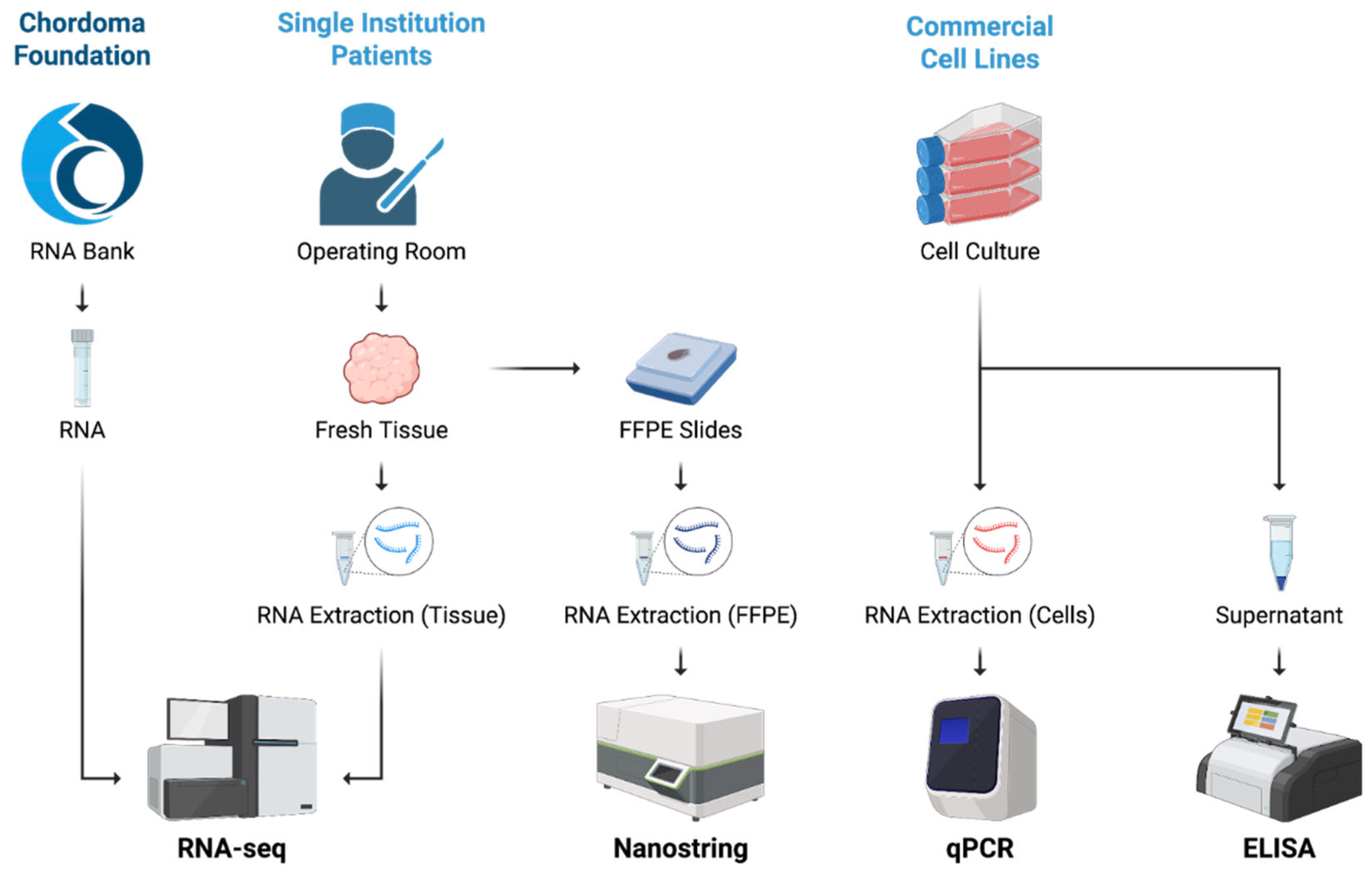
Figure 3.
Differential gene expression from the NanoString PanCancer Pathways Panel. A. Volcano plot of differential expression of genes in recurrent chordomas compared to primary, from patient-derived, fixed-formalin, paraffin-embedded tumor tissue. Red: |FC|>2 and p<0.05; Green: |FC|>2 and p>0.05; Blue: |FC|<2 and p<0.05; Black: |FC|<2 and p>0.05. B. Heatmap indicating the expression level of 56 differentially expressed genes in each sample. Blue indicates below-average gene expression, while red indicates above-average gene expression across all samples. CHI3L1 was enriched in the 4th primary chordoma sample. C. Principal component analysis of each sample’s transcriptome. Samples are colored by tumor type (primary vs recurrent).
Figure 3.
Differential gene expression from the NanoString PanCancer Pathways Panel. A. Volcano plot of differential expression of genes in recurrent chordomas compared to primary, from patient-derived, fixed-formalin, paraffin-embedded tumor tissue. Red: |FC|>2 and p<0.05; Green: |FC|>2 and p>0.05; Blue: |FC|<2 and p<0.05; Black: |FC|<2 and p>0.05. B. Heatmap indicating the expression level of 56 differentially expressed genes in each sample. Blue indicates below-average gene expression, while red indicates above-average gene expression across all samples. CHI3L1 was enriched in the 4th primary chordoma sample. C. Principal component analysis of each sample’s transcriptome. Samples are colored by tumor type (primary vs recurrent).

Figure 4.
Differential gene expression from whole transcriptome sequencing of patient-derived chordoma cell lines. A. Volcano plot of differential expression of genes in recurrent compared to primary chordomas from patient-derived, fixed-formalin, paraffin-embedded tumor tissue. Red: |FC|>2 and p<0.05; Green: |FC|>2 and p>0.05; Black: |FC|<2 and p>0.05. B. Heatmap indicating the expression level of 60 differentially expressed genes. Blue indicates below-average expression and red indicates above-average expression across all samples. C. Principal component analysis of each sample’s transcriptome. Samples are organized by tumor type (primary vs recurrent).
Figure 4.
Differential gene expression from whole transcriptome sequencing of patient-derived chordoma cell lines. A. Volcano plot of differential expression of genes in recurrent compared to primary chordomas from patient-derived, fixed-formalin, paraffin-embedded tumor tissue. Red: |FC|>2 and p<0.05; Green: |FC|>2 and p>0.05; Black: |FC|<2 and p>0.05. B. Heatmap indicating the expression level of 60 differentially expressed genes. Blue indicates below-average expression and red indicates above-average expression across all samples. C. Principal component analysis of each sample’s transcriptome. Samples are organized by tumor type (primary vs recurrent).
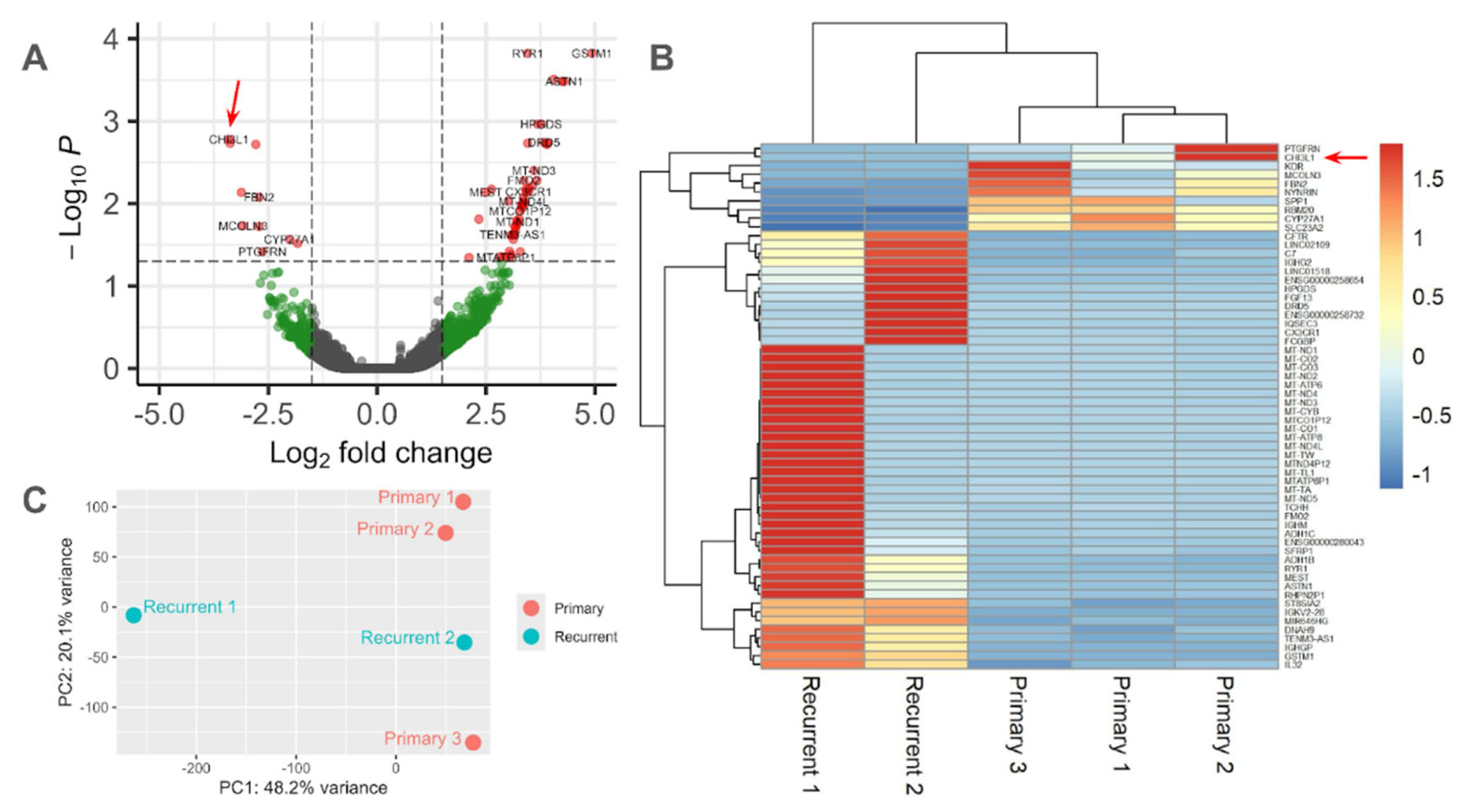
Figure 5.
Relative expression of CHI3L1 mRNA across different cell types measured by qPCR. Primary chordoma cell lines (JHC7 and U-CH12) exhibited significantly more CHI3L1 mRNA expression than the recurrent chordoma (U-CH1) cell line (JHC7 vs. U-CH1 unpaired t-test, p = 0.0112; U-CH12 vs. U-CH1 unpaired t-test, p = 0.0480). JHC7 cells exhibited significantly increased expression relative to notochordal-like U-CH1-N cells (unpaired t-test, p = 0.0390). Error bars represent the standard error of the mean.
Figure 5.
Relative expression of CHI3L1 mRNA across different cell types measured by qPCR. Primary chordoma cell lines (JHC7 and U-CH12) exhibited significantly more CHI3L1 mRNA expression than the recurrent chordoma (U-CH1) cell line (JHC7 vs. U-CH1 unpaired t-test, p = 0.0112; U-CH12 vs. U-CH1 unpaired t-test, p = 0.0480). JHC7 cells exhibited significantly increased expression relative to notochordal-like U-CH1-N cells (unpaired t-test, p = 0.0390). Error bars represent the standard error of the mean.
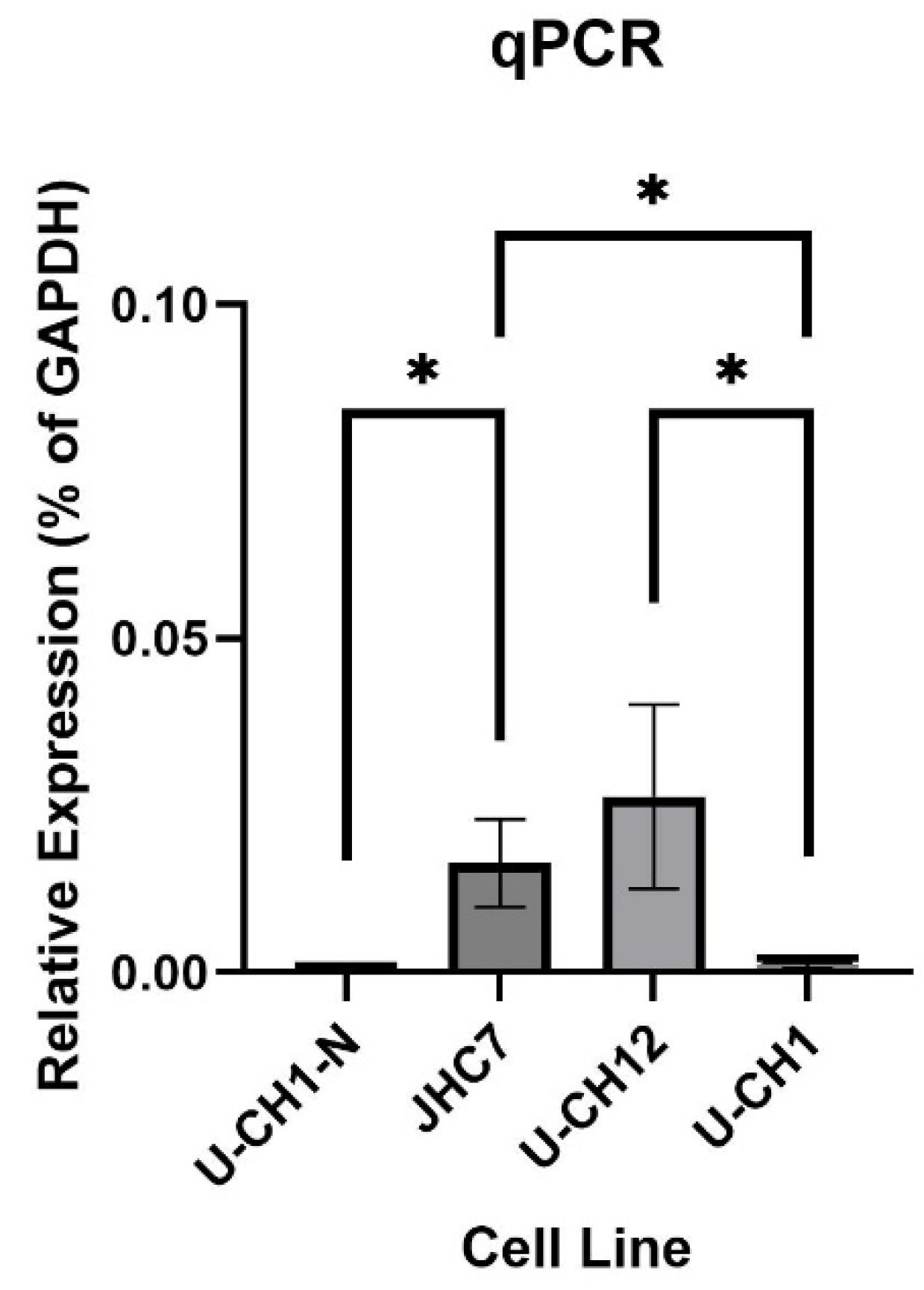
Figure 6.
Secreted CHI3L1 levels across different cell types measured by ELISA. After 1 week, secreted CHI3L1 was measured in the culture supernatant of notochordal-like cells (U-CH1-N), primary chordoma cells (JHC7 and U-CH12), and recurrent chordoma cells (U-CH1). Primary chordoma cells (U-CH12) demonstrated a significant difference in expression between JHC7 cells (U-CH12 vs. JHC7 unpaired t-test, p = 0.0175) and U-CH1-N (U-CH12 vs. U-CH1-N unpaired t-test, p = 0.0135). Error bars represent the standard error of the mean.
Figure 6.
Secreted CHI3L1 levels across different cell types measured by ELISA. After 1 week, secreted CHI3L1 was measured in the culture supernatant of notochordal-like cells (U-CH1-N), primary chordoma cells (JHC7 and U-CH12), and recurrent chordoma cells (U-CH1). Primary chordoma cells (U-CH12) demonstrated a significant difference in expression between JHC7 cells (U-CH12 vs. JHC7 unpaired t-test, p = 0.0175) and U-CH1-N (U-CH12 vs. U-CH1-N unpaired t-test, p = 0.0135). Error bars represent the standard error of the mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



