Submitted:
24 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
Two seminal studies were published almost simultaneously in 1988 in the same scientific journal. Both spurred the field of brain energy metabolism research in new directions, culminating in long-lasting debates that appear to split its practitioners into two factions that seem unwilling to agree on what metabolic processes are responsible for providing adenosine triphosphate (ATP) to the active brain. The first study used rat hippocampal slices to demonstrate the ability of lactate to support neuronal function as the sole oxidative mitochondrial substrate. The second study demonstrated that upon brain stimulation, glucose consumption is not accompanied by oxygen consumption i.e., a non-oxidative glucose utilization or what has become known as “aerobic glycolysis.” Consequently, for almost four decades, researchers in this field have been divided between those who profess that non-oxidative glucose metabolism supports neural activity and those who profess that this activity is supported by oxidative lactate metabolism. Hypotheses for both concepts were offered, “The Efficiency Tradeoff Hypothesis” and the “Astrocyte Neuron Lactate Shuttle Hypothesis,” respectively. To bridge the gap between the two groups, a recent Editorial was published, authored by over twenty leading investigators. The editorial received two separate responses from investigators who support the non-oxidative glucose consumption as the main source of energy to support neuronal activity, signaling that the gap has not being bridged. The present review attempts to cover the principal disagreements that divide this utmost important field of research with the aim of uncovering points of agreement and possible hurdles that, if removed, could close this rift.
Keywords:
active brain energy metabolism
; glucose
; glycolysis
; lactate
; non-oxidative consumption
; oxidative consumption
1. At the Beginning
Most of our knowledge about the processes of brain energy metabolism is based on decades of research performed on a tissue of lesser complexity, the skeletal muscle. This research led to the drawing of the aerobic and anaerobic glycolytic pathways, and with the use of additional tissues, such as liver, the tricarboxylic acid (TCA) cycle and the mitochondrial oxidative phosphorylation (OXPHOS) were also discovered. Through normal science, these energy-producing metabolic pathways have become common knowledge. Although some research was performed using brain tissue in the mid-1920s to the early 1930s, it was guided mainly by results of studies performed using skeletal muscle. Therefore, upon discovering that lactate, in the presence of oxygen, disappears from brain tissue it was simply considered as a mechanism that rids the brain from this useless glycolytic end-product [1,2,3,4,5,6]. Although later the possibility that lactate could be used, not just removed from the tissue in the presence of oxygen was indicated that possibility was not pursued further [7,8]. For over eight decades, glycolysis has been taught in schools and universities almost unchanged from its original 1940 drawing, where its aerobic version ends with pyruvate and the anaerobic one ends with lactate. The division of glycolysis into two parallel pathways, one which occurs in the presence of oxygen (aerobic), while the other takes place in the absence of oxygen (anaerobic), was based on half a century of research that was performed by some of the best scientists of their time, who used mainly preparations of skeletal muscle [9]. By the time the elucidation of the series of enzymatic reactions, substrates and products of the glycolytic pathway was published in 1940, not much was known about the role of mitochondria in adenosine triphosphate (ATP) production. However, Krebs and Johnson [10] already published their studies on the TCA cycle, where they postulated pyruvate to be the entry substrate of that pathway. Embden, Meyerhof, and Parnas, who drew the glycolytic pathway, were aware of that postulation and the role of the TCA cycle in respiration. Along with the general acceptance that without oxygen, glucose hydrolysis ends with lactate, the useless end-product of the process, glycolysis was offered as a metabolic pathway that under oxidative conditions is composed of ten enzymatic reactions. Two molecules of ATP are invested in the breakdown of one molecule of hexose (glucose) into two triose molecules (pyruvate) and the production of four molecules of ATP. To account for lactate production in the absence of oxygen, the fathers of the glycolytic pathway added eleventh enzymatic reaction, which converts pyruvate to lactate by the enzyme lactate dehydrogenase (LDH). Consequently, glycolysis was proposed to have two different outcomes, an aerobic one, ending with pyruvate, and anaerobic one, ending with lactate. This division is still being taught and recited both in textbooks and biochemistry courses in classrooms and online today.
2. The Revolution of the 1980s
Since its discovery in 1780 by Carl Wilhelm Scheele, lactate carried the negative reputation of milk spoiler, a useless product of anaerobic glycolysis, and even a poisonous substance. Then, Brooks published his notable study, showing that lactate is shuttled from fast twitching muscle fibers, where it is produced, to slow twitching ones, where it is utilized as an energy substrate during sustained exercise in mammals [11]. Independently, Schurr et al. [12] demonstrated in vitro the ability of neurons to sustain their function utilizing lactate as their sole oxidative energy substrate. Less than a month later, Fox et al. [13] published their seminal study, claiming that brain activity is supported by a non-oxidative glucose consumption (glycolytic ATP production). Three studies published in a span of three years appeared to shake the tranquility of the field of energy metabolism, and especially the field of brain energy metabolism. Suddenly, lactate was viewed in a completely new light, and brain activity needs, believed to require great increases in ATP supply, could be supported by the inefficient glycolytic pathway alone. Naturally, before such revolutionary ideas could be accepted both had faced much skepticism. While skepticism is part and parcel of scientific research, the two key findings, that lactate is an oxidative energy substrate, and that the brain does not require more than a meager amount of ATP to support its activity, have divided the scientists researching brain energy metabolism for the past four decades. While our knowledge about the many roles of lactate in the brain expanded over this period [14,15,16,17,18,19], one particular construct from the original chart of the glycolytic pathway 85 years ago stubbornly remained unchanged, despite it being erroneous, i.e., the assigning of two different end-products, pyruvate or lactate, depending on the presence or absence of oxygen, respectfully. Embden, Meyerhof, and Parnas, had neither the knowledge nor the tools available to us today, forcing them to use some guesswork to fit their knowledge to the final proposal of the glycolytic pathway, its enzymes, substrates and products. Since at that time lactate was considered a useless end product of anaerobiosis, glucose hydrolysis that led to lactate production was assumed to be a process that took place in the absence of oxygen, a process of no return. However, in the presence of oxygen the lack of lactate production, the product of pyruvate reduction by lactate dehydrogenase (LDH), meant that aerobic glycolysis is one step shorter than the anaerobic pathway. Their decision to draw aerobic glycolysis as a pathway ending with pyruvate was based on the suggestion by Krebs and Johnson [10] that pyruvate is the substrate of the tricarboxylic acid (TCA) cycle [12]. More than eight decades later, the original division of glycolysis into aerobic and anaerobic pathways is still taught and cited as such [20,21,22,23] despite the overwhelming evidence that neither oxygen nor mitochondria could prevent glycolysis from proceeding through eleven enzymatic steps, ending with the production of lactate [12,24,25,26,27,28]. Brooks et al. [27] summarized this paradox as follows:
“Contemporary textbooks of biochemistry and physiology abound the 19th-century concepts that metabolism is either “anaerobic” (without O2) or “aerobic” (with O2). Glycolytic flux from glucose and glycogen is typically depicted in textbooks as progressing to pyruvate and then to the tricarboxylic acid (TCA) cycle. However, if oxygen is absent, textbooks assert that glycolysis progresses to lactate. This is a convenient motif, typically copied by one textbook author from another and then through serial editions of texts. Amazingly, some textbook authors who advanced the idea of lactate production due to oxygen limitation were biochemists who worked with cells in high-glucose-containing culture media under fully aerobic conditions of one atmosphere pressure in which the partial pressure of oxygen was at least 50% greater than in the arterial blood of individuals at sea-level altitudes. Because the maintenance of cells in such preparations required daily changing of the incubation media to maintain high [glucose], low [lactate], and physiological pH, the observations should have informed the investigators that lactate was produced under fully aerobic conditions. Contemporaneously, physiologists measured lactate [L] to pyruvate [P] concentration ratios (L/P) of 10 in muscles and blood in resting mammals, including humans, and further observed the L/P to rise more than 100 during submaximal exercise. However, did any of the textbook authors ever look to determine whether isolated mitochondria oxidize lactate? For many textbook authors, the answer is “no.” However, for a few others, the answer is “yes”. Moreover, some investigators also looked for the presence of a mitochondrial monocarboxylate (lactate) transporter (mMCT) and a lactate dehydrogenase (mLDH) enzyme. Unfortunately, while some attempts failed, fortunately, other attempts to observe mitochondrial lactate oxidation and the presence of mLDH and mMCT were successful. Importantly, lactate oxidation in human muscle mitochondrial preparations was observed. Regrettably, for a time, positive results were overlooked or castigated as being “controversial” and dismissed for failing to fit with established paradigms of metabolic regulation.”
Passarella [29] expresses a similar sentiment in his recent review:
“…the existence of mitochondrial L-lactate metabolism, mediated by specific L-lactate transporters and a mLDH, remains largely overlooked in contemporary biochemistry textbooks and in several recent reviews and research papers.”
A significant number of scientists in the field of energy metabolism persistently hold to the erroneous concept that oxygen could prevent the conversion of pyruvate to lactate, a concept on which a great deal of “common knowledge” relies. When wrong common knowledge becomes entrenched in the minds of the investigators who used it in their studies and their teachings, in the minds of their students and therefore in the mind of the public, rectifying it could be an overwhelming task. Consequently, “aerobic glycolysis” can end both with pyruvate, but also with lactate under aerobic conditions, where the latter signifies a non-oxidative glucose consumption [13]. This convoluted situation could be straightened up easily if all agreed that glycolysis under any condition, always begins with the phosphorylation of D-glucose by the enzyme hexokinase, and ends with the reduction of pyruvate to L-lactate by LDH. Either prefix, aerobic or anaerobic, are not necessary to describe the pathway that, thermodynamically [30,31], must and does always end with lactate. Oxygen, even at the highest possible concentration, cannot block the LDH reaction, as is clear from the glycolytic process of red blood cells for their ATP production or the activity of the Na+/K+-ATPase pump that is supported by its own ATP-producing glycolytic pathway with lactate as an end-product. Nevertheless, as illustrated below by some relatively recent studies, the concept of aerobic glycolysis as a pathway of ten enzymatic reactions ending with pyruvate is still deeply rooted. Table 1 lists several recent publications that exemplify the continued reliance of many in the research field of brain energy metabolism on the original 1940 paradigm of glycolysis, according to which under aerobic conditions the pathway proceeds through ten enzymatic reactions, hydrolyzing a molecule of glucose into two triose molecules, namely pyruvate.
_________________________________________________________________________________________________
Table 1. Six recent publications, four of which are highly cited, where the original depiction of aerobic glycolysis by Embden, Meyerhof and Parnas, is still promoted.
______________________________________________________________________________________________________
1. McNair et al. [32], in a study titled “Metabolic Characterization of Acutely Isolated Hippocampal and Cerebral Cortical Slices Using [U-13C]Glucose and [1,2-13C]Acetate as Substrates,” found a high production of lactate by both hippocampal and cortical slices that reached its maximum after 15 minutes of incubation with [U-13C]Glucose, a production that did not increase throughout the following 75 minutes. While they correctly pointed out that lactate production is due to glycolytic activity, they failed to explain why lactate production, as measured by its content in the incubation medium, did not continue to increase above the initial level measured at 15 minutes, which indicates lactate utilization by the hippocampal and the cerebral cortical slices. Clearly, the investigators followed the concept that pyruvate is the glycolytic end-product that enters the TCA cycle and never considered lactate as the possible mitochondrial substrate. (Cited 43 times since its publication).
2. Yellen [33] published the article “Fueling thought: Management of glycolysis and oxidative phosphorylation in neuronal metabolism,” where the author defined glycolysis as the partial metabolism of glucose to pyruvate or lactate. That it occurs in the cytosol (and possibly is subcompartmented within the cytosol, perhaps to the intracellular surface of the plasma membrane). He detailed that metabolism of glucose to pyruvate yields two net phosphorylations of ADP to ATP and two 2e– (two-electron) reductions of NAD+ to NADH, and that sustaining glycolysis requires reoxidation of NADH to NAD+ either through the coordinated reduction of pyruvate to lactate by lactate dehydrogenase (LDH) or through the action of one of the mitochondrial NADH shuttles. (Cited 411 times since its publication).
3. Jang et al. [34], in their study titled “Metabolomics and isotope tracing,” used the isotope [1,2-13C]glucose to trace glycolytic products. While tracing lactate, they relied on the concept that pyruvate is the glycolytic end-product that enters the TCA cycle, as illustrated in Figure 6D of the article. In a personal communication, Prof. Rabinowitz stated: “At the whole-body level, we see that a majority of the glycolytic process goes straight from glucose to pyruvate to Acetyl-CoA in the fed state (some also to lactate), whereas in the fasted state it mainly proceeds to lactate (which enters the bloodstream) and then back to pyruvate (presumably extra-mitochondrially) and to Acetyl-CoA.” However, the study’s results could very easily be explained by accepting the concept that glycolysis ends with lactate, the monocarboxylate that is transported by MCT into the mitochondria, where it is readily converted back to pyruvate via the mitochondrial LDH (mLDH), which then enters the TCA cycle via acetyl-CoA. (Cited 941 times since its publication).
4. Casson et al. [35], in a detailed review article, summarized the progress in research of “Retinal energy metabolism in health and glaucoma,” graphically illustrated glycolysis as a ten enzymatic reactions pathway that ends with pyruvate, while at the same time, accepting the idea that retinal lactate is produced glycolytically and is utilized as the mitochondrial oxidative substrate. (Cited 125 times since its publication).
5. Westi et al. [36], in a detailed study titled “Divergent Cellular Energetics, Glutamate Metabolism, and Mitochondrial Function Between Human and Mouse Cerebral Cortex,” presented glycolysis graphically as a pathway that begins with the phosphorylation of glucose and ends with the production of pyruvate, although they also used β-hydroxybutyrate (BHB) as a mitochondrial oxidative substrate, which is not different from the ability of their mitochondrial preparation to use lactate, which unfortunately, they did not consider as a potential mitochondrial substrate. (Cited 20 times since its publication)
6. Rae et al. [37], in an encompassing review titled “Brain energy metabolism: A roadmap for future research,” emphatically promote the concept of ten glycolytic enzymatic reactions with pyruvate as its final product and the mitochondrial substrate of the TCA cycle. Among the 26 co-authors of this review are many established investigators in the research field of brain energy metabolism. Their review minimizes the role of lactate as a significant energy substrate and question the validity of the astrocyte neuron lactate shuttle hypothesis. (Cited 80 tomes since its publication)
____________________________________________________________________________________________________________
The fact that many within this field continue to argue for two separate glycolytic outcomes, while accepting the fact that even under conditions where the pathway is supposed to end with pyruvate, it ends with lactate (aerobic glycolysis), requires a re-evaluation of the accepted dogma. ‘Habit of mind’ [38] has been discussed before as a possible explanation for the resistance to shift away from said dogma when scientific findings justify a paradigm shift [12,25,28,29,39]. Clearly, neither the thermodynamic argument nor a psychological explanation could overcome this resistance for such a shift. Therefore, it is up to a scientific experiment to show which glycolysis is the real pathway: a) the one composed of eleven sequential enzymatic reactions that begins with the phosphorylation of one equivalent of glucose and ends with the formation of two equivalents each of lactate, ATP and NAD+ or: b) the glycolytic pathway that ends after its tenth enzymatic step, the conversion of phosphoenolpyruvate (PEP) to pyruvate, in the presence of both oxygen and mitochondria. While there are ample experiments to support the former, there are no experiments that support the latter. The research field of brain energy metabolism will thrust forward only when its practitioners accept glycolysis as a pathway that begins with glucose and ends with lactate.
3. Evidence in Support of Lactate as the Only Triose end-Product of Glycolysis
Thermodynamically, glycolysis must flow through all its eleven enzymatic reactions, including the very last one, the LDH reaction (Figure 1). Park et al. [30] studied several metabolic pathways in three different organisms, including mammals, concluded that all metabolic pathways, their metabolite concentrations, fluxes and free energies, imply an efficient enzyme usage, which achieved by maintaining a negative ΔG throughout. While the investigators did not specifically measure or discuss the last enzymatic reaction of glycolysis, i.e., the conversion of pyruvate to lactate by the cytosolic LDH (cLDH), the significant free energy gained by this reaction going forward (ΔG = -6.0 Kcal) should agree with their general conclusion. Therefore, the glycolytic pathway should always end with lactate. Nevertheless, upon examining Figure 6D in the paper by Park et al. [30], two questions arise: 1. Why does glycolysis in this figure ends with pyruvate prior to its conversion to Acetyl-CoA and its entry into the TCA cycle, while lactate is shown as a side end-product that goes nowhere? 2. Could the investigators, using their tools and approaches, decipher whether the glycolytic process proceeds from glucose to pyruvate to Acetyl-CoA or it proceeds from glucose to lactate and then to pyruvate (intra-mitochondrially) and Acetyl-CoA?
In a personal communication with one of the authors (Prof. Rabinowitz), the answers to these two questions were: “At the whole-body level, we see that a majority of the glycolytic process goes straight from glucose to pyruvate to Acetyl-CoA in the fed state (some also to lactate), whereas in the fasted state it mainly proceeds to lactate (which enters the bloodstream) and then back to pyruvate (presumably extra-mitochondrially) and to Acetyl-CoA.” These answers do not align with the principle of their study according to which efficient enzyme usage of a metabolic pathway is achieved by maintaining a negative ΔG throughout. Claiming that glycolysis stops at pyruvate rather than proceeds to lactate contradicts this principle. In addition, the presumption, held by many, that the conversion of lactate back to pyruvate occurs extra-mitochondrially, is wrong, since that enzymatic reaction in the cytosol could not be catalyzed by cLDH against its negative ΔG. In contrast, intra-mitochondrially, lactate conversion to pyruvate by mitochondrial LDH (mLDH) does proceed fluently [27,40,41,42,43,44,45,46,47]. A study by Cai et al. [48] demonstrated that lactate also plays a role in suppressing glycolysis, while activating the mitochondrial electron transport chain independently of its metabolism.
Accepting glycolysis as a pathway that always ends up with lactate means that its prefixes, aerobic and anaerobic, are meaningless [26,28]. Accordingly, lactate is always the substrate for the mitochondrial TCA cycle and OXPHOS under oxidative conditions. Moreover, when ample supply of lactate is available to the brain, lactate is the preferred substrate for oxidative energy production over glucose [49,50,51,52,53,54,55,56]. In a recent study, using ultrasensitive sensors, Li et al. [57] showed that a lactate pool exists inside mitochondria at levels significantly higher than those in the cytosol. Moreover, only when lactate is the final glycolytic product that the pathway fulfills its cyclic purpose of renewing NAD+ supply [46]. Obviously, the conversion of lactate back to pyruvate, the substrate of the TCA cycle, occurs in the mitochondrion upon lactate transport via a monocarboxylate transporter (MCT) into the organelle and its oxidation by mLDH, a reaction that is not part of the glycolytic pathway.
Yet old habits are still at the center of the debate in the field of brain energy metabolism, where it has always been easy to accept the fact that without oxygen the glycolytic pathway ends with lactate [27]. And since without oxygen, mitochondrial activity is impossible, the only metabolic pathway capable of producing ATP is glycolysis. However, Otto Warburg described aerobic glucose consumption in cancerous cells, where lactate is the main product, the process by which these cells produce most of the energy required for their proliferation [58,59]. This phenomenon was discovered more than a decade before the glycolytic pathway sequence was elucidated, but the scientists of the day studying energy metabolism seemed to consider it unique to cancerous cells. Yet nowadays ‘aerobic glycolysis’ is a term used to describe glycolytic glucose consumption that ends with the production of lactate by normal, noncancerous cells, that are fully oxygenated. Despite the term’s popularity, it is a metabolic process that defies decades of accepted concept, according to which aerobic glycolysis is supposed to end with pyruvate, as every biochemistry textbook describes. While oxygen alone cannot stop glycolysis at the production of pyruvate (see glycolysis of red blood cells), when mitochondria are present the textbook states that pyruvate is the glycolytic end-product. Yet, aerobic glycolysis in today’s jargon means a process that ends with lactate despite the presence of both oxygen and mitochondria, as was first claimed by Fox et al. [13]. The claim was based on an indirect estimate of oxygen consumption that was calculated from measurement of changes in brain blood oxygen level and/or the rate of cerebral blood flow (CBF). Regardless, a significant number of investigators have accepted aerobic glycolysis as the metabolic process that provides the necessary additional ATP for the active brain, and to rationalize the brain’s choice of an inefficient ATP-producing process for that task, the efficiency tradeoff hypothesis was offered [60]. The arguments for and against the validity of the ‘aerobic glycolysis’ concept have been already reviewed [28]. Additionally, the results of two recent studies [61,62] show that myelin can store enough oxygen, which makes it available for the active brain. These studies support the findings of Hu and Wilson [49] that oxygen is consumed upon brain activation, possibly explaining the non-reliance of the active brain on an immediate availability of blood-borne oxygen [63]. In essence, these studies challenge not only the efficiency tradeoff hypothesis, but they also highlight the inadequacy of techniques such as BOLD fMRI and CBF measurements to gauge brain oxygen consumption accurately, since these techniques cannot sense and measure consumption of oxygen stored in brain tissue outside the vasculature.
4. The roles of MCT and mLDH in Mitochondrial Lactate Metabolism
The existence of mLDH in rat heart, kidney, liver, and lymphocytes mitochondrial preparations was first reported by Brandt et al. [64]. However, they reported that the presence of mLDH in brain mitochondrial preparation was inconclusive. By 1999, Brooks et al. [40] expanded our knowledge of mLDH in muscle, heart and liver tissues by showing that there are differences in the isoform composition of LDH, not only between cLDH and mLDH, but also between liver, heart and muscle mLDH. Van Hall [65] confirmed these findings and suggested that not only the isoform composition of LDH, but also its compartmentalization and its association with specific cellular structures, determines whether it reduces pyruvate or oxidizes lactate. An in vitro study, employing rat hippocampal slices and LDH inhibitors [41], strongly suggested that mLDH, not cLDH, participates in the oxidation of lactate to pyruvate that enters the TCA cycle after its conversion to acetyl CoA. Later Atlante et al. [66] and Hashimoto et al. [42] demonstrated the existence of brain mLDH. Passarella et al. [43] summarized the mitochondrial lactate dehydrogenase research affair. For lactate to be oxidized intra-mitochondrially, it must be transported into the organelle first. Pierre and Pellerin [67] described the distribution, function, and regulation of the specific monocarboxylate transporters (MCTs) of the central nervous system. Ueno et al. [68] described the distribution and function of the four brain MCTs, belonging to the SLC16 family of solute carriers, that catalyzes proton-coupled lactate transport. MCT1 is expressed almost ubiquitously in the brain, in endothelial cells, astrocytes, and oligodendrocytes. Astrocytes release lactate via MCT4 and possibly into oligodendrocytes via MCT1. MCT2 catalyzes the proton-coupled transport of monocarboxylates such as lactate, pyruvate, and ketone bodies, across the plasma membrane, showing the highest affinity for lactate followed by pyruvate. MCT2 is expressed mainly in neurons but also found in astrocytes. Pierre et al. [69] found that MCT1 is strongly expressed in astrocytes of mice, while MCT2 is expressed in neurons, findings that are consistent with the ANLS hypothesis, where lactate is transported out of astrocytes via MCT1 and is taken up by neurons via MCT2. MCT4 is a low-affinity, high-capacity transporter and is expressed mainly in astrocytes [70] and is known to facilitate lactate excretion in cells that have high rate of glycolysis [71]. MCT2 is known to be a transporter with a higher affinity for most monocarboxylates than MCT1 [72]. In short, the distribution of MCTs in the plasma membrane of neurons and astrocytes suggests a significant role of these transporters in the shuttling of energy metabolites between these two cell types. Hashimoto et al. [42] used confocal laser scanning microscopy and immunoblotting after immunoprecipitation from cell lysates to demonstrate that the location of MCT1, MCT2 and LDH is in neuronal mitochondria, and additionally that these MCTs and the LDH are associated with cytochrome oxidase (COX) in rat brain mitochondria.
The LDH enzyme exists in several isomers. These isomers differ from one another based on the type of the four subunits that compose them. Their subunit composition also determines the direction the reaction they catalyze goes, pyruvate to lactate or vice versa. There are several genes that code the synthesis of the LDH protein, two are found in most tissues and organs, including the brain, LDHA and LDHB [73]. The former’s product is LDHA, also known as LDH-M (M for muscle), and the latter is LDHB, also known as LDH-H (H for heart). Accordingly, five different isomers could be formed, each composed of 4 subunits of LDH; LDH-1, composed of four M subunits, LDH-2, composed of M subunits and one H subunit, LDH-3, composed of two M and two H subunits, LDH-4, composed of 3 H subunits and one M subunit, and LDH-5, composed of four H subunits. LDH-M has higher affinity to pyruvate and therefore catalyzes its conversion to lactate, with an oxidation of NADH to NAD+, while LDH-H has higher affinity to lactate and its conversion to pyruvate with a reduction of NAD+ to NADH. Studies showed the colocalization of MCTs and LDH isoforms in the inner mitochondrial membrane of rat derived L6 skeletal muscle cell line [74] and in mitochondria from untransformed primary breast cell line, HMEC 184 [75]. Nevertheless, it is now recognized that mitochondrial LDH (mLDH) is a constituent of the mitochondrial proteome [76]. Li et al. [77], using ultrasensitive sensors, showed that lactate is highly enriched in mitochondria at levels that significantly exceed those in the cytosol. Therefore, the overwhelming evidence that demonstrates the existence of a complete mitochondrial machinery that imports and metabolizes lactate cannot be ignored or discounted any longer.
5. Conclusions
The past four decades have brought about a paradigm shift in the field of brain energy metabolism research. Unfortunately, significant number of investigators within this field continue to cling to an 85-year-old paradigm, relying on it for the interpretation of their studies. Consequently, a long-standing debate between those who accept the new paradigm and those who reject it, continues to divide this utmost important endeavor of understanding how the one organ that consumes more energy than any other, controls its energy needs in health, during rest and activity, and how the loss of said control leads to numerous brain disorders. At the center of the debate stands the first metabolic pathway to be fully explicated, namely, glycolysis. All enzymes, cofactors, substrates and products were identified and arranged in the order they comprise the pathway. Although it was clear then that anaerobically this pathway ends with the conversion of pyruvate to lactate, it was assumed that the presence of oxygen should make this last step unnecessary. The idea of two different glycolytic outcomes had become deeply rooted in textbooks, school and college classes, and minds, both novice and expert.
Despite the continuous accumulation of evidence in support of the required paradigm shift described above, the research community of brain energy metabolism remained divided. A recent study in humans [78] concluded that “…when lactate is made available in circulation – whether through passive infusion or exercise – the healthy human brain preferentially metabolizes it over glucose to sustain energetic needs. This sparing effect of lactate on glucose appears to permit glucose to fulfil non-energetic requirements of the brain.” It is too early to determine if findings such as these would sway more investigators to accept the new paradigm.
The publication of a recent Editorial attempted to bridge this divide [79]. Considering the responses that followed shortly after its publication [80,81], it appears that additional efforts to build a bridge are needed.
This monograph is another attempt to correct this erroneous concept that hinders the progress of understanding the utmost important processes that energize the brain both under rest and during activity, in health and disease. It reviews the discoveries that made us question the old archetype and lists the scientific evidence that has brought about a paradigm shift according to which glycolysis is a metabolic pathway that begins with glucose and always ends with lactate, in the presence or absence of oxygen or mitochondria (Figure 2).
Until all investigators of brain energy metabolism agree that there is only one glycolytic pathway, independent of oxygen and mitochondria that always ends with the production of lactate, the progress in our understanding of the most crucial processes that energize the brain will be hindered.
Funding
This research received no external funding.
Acknowledgments
The author thanks Paige Hessel for her comments, suggestions, and corrections.
Conflicts of Interest
The author declares that they have no competing interests.
References
- Holmes, EG; Holmes, BE. Contributions to the study of brain metabolism. II. Carbohydrate metabolism. Biochem. J. 2025, 199(5), 836–39. [Google Scholar] [CrossRef]
- Holmes, EG; Holmes, BE. Contributions to the study of brain metabolism. III. Carbohydrate metabolism relationship of glycogen and lactic acid. Biochem. J. 1926, 20(6), 1196–203. [Google Scholar] [CrossRef]
- Holmes, EG; Holmes, B E. Contributions to the study of brain metabolism. IV. Carbohydrate metabolism of the brain tissue of depancreatised cats. Biochem. J. 1927, 21(2), 412–18. [Google Scholar] [CrossRef] [PubMed]
- Ashford, CA; Holmes, EG. Contributions to the study of brain metabolism. V. Role of phosphates in lactic acid production. Biochem. J. 1929, 23(4), 748–59. [Google Scholar] [CrossRef]
- Holmes, EG. Oxidations in central and peripheral nervous tissue. Biochem. J. 1930, 24(4), 914–25. [Google Scholar] [CrossRef]
- Holmes, EG; Ashford, CA. Lactic acid oxidation in brain with reference to the “Meyerfof cycle.”. Biochem. J. 1930:249, 4, 1119–27. [Google Scholar] [CrossRef]
- Holmes, EG. The metabolism of brain and nerve. Ann. Rev. Biochem. 1932:1, 487–506. [Google Scholar] [CrossRef]
- Holmes, EG. The relation between carbohydrate metabolism and the function of the grey matter of the central nervous system. Biochem. J. 1933, 27(2), 523–36. [Google Scholar] [PubMed]
- Schurr, A. Cerebral glycolysis: a century of persistent misunderstanding and misconception. Front. Neurosci. 2014, 8, 360. [Google Scholar] [CrossRef] [PubMed]
- Krebs, H. A.; Johnson, W. A. The role of citric acid in intermediary metabolism in animal tissue. Enzymologia 1937, 4, 148–56. [Google Scholar]
- Brooks, GA. “Lactate: glycolytic product and oxidative substrate during sustained exercise in mammals – the “lactate shuttle” in Comparative Physiology and Biochemistry: Current Topics and Trends, Vol. A. Respiration-Metabolism–Circulation, ed R. Gilles (Berlin: Springer-Verlag), 208–218.
- Schurr, A; West, CA; Rigor, BM. Lactate-supported synaptic function in the rat hippocampal slice preparation. Science 1988, 240(4857), 1326–8. [Google Scholar] [CrossRef]
- Fox, PT; Raichle, ME; Mintun, MA; Dence, C. Nonoxidative glucose consumption during focal physiologic neural activity. Science 1988, 241(4864), 462–4. [Google Scholar] [CrossRef] [PubMed]
- Barros, LF. Metabolic signaling by lactate in the brain. Trends in neurosciences 2013, 36(7), 396–404. [Google Scholar] [CrossRef]
- Proia, P; Di Liegro, CM; Schiera, G; Fricano, A; Di Liegro, I. Lactate as a Metabolite and a Regulator in the Central Nervous System. International Journal of Molecular Sciences 2016, 17(9), 1450. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P; Allaman, I. Lactate in the brain: from metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–49. [Google Scholar] [CrossRef] [PubMed]
- Cai, M; Wang, H; Song, H; Yang, R; Wang, L; Xue, X; Sun, W; Hu, J. Lactate Is Answerable for Brain Function and Treating Brain Diseases: Energy Substrates and Signal Molecule. Front. Nutr. 2022, 9, 800901. [Google Scholar] [CrossRef]
- Wu, A; Lee, D; Xiong, W.-C. Lactate Metabolism, Signaling, and Function in Brain Development, Synaptic Plasticity, Angiogenesis, and Neurodegenerative Diseases. Internatl. J. Molec. Sci,. 2023, 2023:24(17), 13398. [Google Scholar] [CrossRef]
- Zhu, X; Chen, W; Pinho, RA; Thirupathi, A. Lactate-induced metabolic signaling is the potential mechanism for reshaping the brain function - role of physical exercise. Front. Endocrinol. 2025, 16, 1598419. [Google Scholar] [CrossRef]
- 13.1: Glycolysis - Biology LibreTexts.
- Chaudhry, R; Varacallo, MA. Biochemistry, Glycolysis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island (FL), 8 Aug 2023. [Google Scholar] [PubMed]
- Kierans, SJ; Taylor, CT. Glycolysis: A multifaceted metabolic pathway and signaling hub. J. Biol. Chem. 2024, 300(11), 107906. [Google Scholar] [CrossRef]
- Fuentes-Lemus, E; Usgame, K; Fierro, A; López-Alarcón, C. Enzymes of glycolysis and the pentose phosphate pathway as targets of oxidants: Role of redox reactions on the carbohydrate catabolism. Redox Biochemistry and Chemistry 2025:11, 100049. [Google Scholar] [CrossRef]
- Rogatzki, MJ; Ferguson, BS; Goodwin, ML; Gladden, LB. Lactate is always the end product of glycolysis. Front. Neurosci. 2015, 9:22. [Google Scholar] [CrossRef]
- Schurr, A. Glycolysis Paradigm Shift Dictates a Reevaluation of Glucose and Oxygen Metabolic Rates of Activated Neural Tissue. Front. Neurosci. 2018, 12, 700. [Google Scholar] [CrossRef]
- Schurr, A; Passarella, S. Aerobic Glycolysis: A DeOxymoron of (Neuro)Biology. Metabolites 2022, 2022:12(1), 72. [Google Scholar] [CrossRef]
- Brooks, GA; Curl, CC; Leija, RG; Osmond, AD; Duong, JJ; Arevalo, JA. Tracing the lactate shuttle to the mitochondrial reticulum. Exp. Mol. Med. 2022, 54(9), 1332–47. [Google Scholar] [CrossRef]
- Schurr, A. How the ‘Aerobic/Anaerobic Glycolysis’ Meme Formed a ‘Habit of Mind’ Which Impedes Progress in the Field of Brain Energy Metabolism. Internatl. J. Molec. Sci. 2024, 25(3), 1433. [Google Scholar] [CrossRef]
- Passarella, S. Revisiting Concepts of Mitochondrial Transport and Energy Metabolism in Health and Cancer. Acad. Biol. 2025, 3(4). [Google Scholar] [CrossRef]
- Park, J; Rubin, S; Xu, YF; Amador-Noguez, D; Fan, J; Shlomi, T; Rabinowitz, J. Metabolite concentrations, fluxes and free energies imply efficient enzyme usage. Nat Chem Biol 2016, 12, 482–9. [Google Scholar] [CrossRef]
- Chen, J; Zaidi, M; Chaudhary, J; Erfani, Z; Al Nemri, S; Plautz, EJ; Wen, X; Seeley, EH; Bartnik-Olson, BL; Corbin, IR; Park, JM. Comparative Evaluation of Hyperpolarized [13C] pyruvate and [13C] lactate for Imaging Neuronal and Glioma Metabolism. ACS sensors 2025. [Google Scholar] [CrossRef] [PubMed]
- McNair, LF; Kornfelt, R; Walls, AB; Andersen, JV; Aldana, BI; Nissen, JD; Schousboe, A; Waagepetersen, HS. Metabolic Characterization of Acutely Isolated Hippocampal and Cerebral Cortical Slices Using [U-13C]Glucose and [1,2-13C]Acetate as Substrates. Neurochem. Res. 2017, 42(3), 810–26. [Google Scholar] [CrossRef] [PubMed]
- Yellen, G. Fueling thought: Management of glycolysis and oxidative phosphorylation in neuronal metabolism. J. Cell Biol. 2018, 217(7), 2235–46. [Google Scholar] [CrossRef] [PubMed]
- Jang, C; Chen, L; Rabinowitz, JD. Metabolomics and isotope tracing. Cell 2018, 3173(4), 822–37. [Google Scholar] [CrossRef] [PubMed]
- Casson, RJ; Chidlow, G; Crowston, JG; Williams, PA; Wood, JP. Retinal energy metabolism in health and glaucoma. Progress in retinal and eye research 2021, 81, 100881. [Google Scholar] [CrossRef]
- Westi, EW; Jakobsen, E; Voss, CM; Bak, LK; Pinborg, LH; Aldana, BI; Andersen, JV. Divergent cellular energetics, glutamate metabolism, and mitochondrial function between human and mouse cerebral cortex. Molec. Neurobio 2022, 59, 7495–512. [Google Scholar] [CrossRef]
- Rae, CD; Baur, JA; Borges, K; Dienel, G; Díaz-García, CM; Douglass, SR; Drew, K; Duarte, JM; Duran, J; Kann, O; Kristian, T. Brain energy metabolism: A roadmap for future research. J. Neurochem. 2024, 168, 910–54. [Google Scholar] [CrossRef] [PubMed]
- Margolis, H. Paradigms and Barriers: How Habits of Mind Govern Scientific Beliefs; The University of Chicago Press: Chicago and London, 1993. [Google Scholar]
- Schurr, A. The Feud over Lactate and Its Role in Brain Energy Metabolism: An Unnecessary Burden on Research and the Scientists Who Practice It. Internl. J. Molec. Sci. 2025, 26(9), 4429. [Google Scholar] [CrossRef] [PubMed]
- Brooks, GA; Dubouchaud, H; Brown, M; Sicurello, JP; Butz, CE. Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc. Nat. Acad. Sci. 1999, 96(3), 1129–34. [Google Scholar] [CrossRef]
- Schurr, A; Payne, R S. Lactate, not pyruvate, is neuronal aerobic glycolysis end product: an in vitro electrophysiological study. Neuroscience 2007, 147(3), 613–9. [Google Scholar] [CrossRef]
- Hashimoto, T; Hussien, R; Cho, HS; Kaufer, D; Brooks, GA. Evidence for the mitochondrial lactate oxidation complex in rat neurons: demonstration of an essential component of brain lactate shuttles. PloS 2008, 3(8), e2915. [Google Scholar] [CrossRef]
- Passarella, S; Paventi, G; Pizzuto, R. The mitochondrial L-lactate dehydrogenase affair. Front. Neurosci. 2014, 8, 407. [Google Scholar] [CrossRef]
- Passarella, S; Schurr, A. l-Lactate transport and metabolism in mitochondria of Hep G2 cells—The Cori cycle revisited. Front. Oncol. 2018, 8, 120. [Google Scholar] [CrossRef]
- Young, A; Oldford, C; Mailloux, RJ. Lactate dehydrogenase supports lactate oxidation in mitochondria isolated from different mouse tissues. Redox biol. 2020, 28, 101339. [Google Scholar] [CrossRef] [PubMed]
- Glancy, B; Kane, DA; Kavazis, AN; Goodwin, ML; Willis, WT; Gladden, LB. Mitochondrial lactate metabolism: history and implications for exercise and disease. J. Physiol. 2021, 599(3), 863–88. [Google Scholar] [CrossRef]
- Leija, RG; Arevalo, JA; Xing, D; Vázquez-Medina, JP; Brooks, GA. The mitochondrial lactate oxidation complex: endpoint for carbohydrate carbon disposal. Am. J. Physiol-Endocrinol. Metab. 2025, 2025:328(1), E126-36. [Google Scholar] [CrossRef]
- Cai, X; Ng, CP; Jones, O; Fung, TS; Ryu, KW; Li, D; Thompson, CB. Lactate activates the mitochondrial electron transport chain independently of its metabolism. Molec. cell 2023, 2023:83(21), 3904–20. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y; Wilson, GS. A temporary local energy pool coupled to neuronal activity: fluctuations of extracellular lactate levels in rat brain monitored with rapid-response enzyme-based sensor. J Neurochem. 1997, 69(4), 1484–90. [Google Scholar] [CrossRef]
- Schurr, A; Miller, JJ; Payne, RS; Rigor, BM. An increase in lactate output by brain tissue serves to meet the energy needs of glutamate-activated neurons. J. Neurosci. 1999, 19(1), 34–9. [Google Scholar] [CrossRef]
- Bouzier-Sore, AK; Voisin, P; Canioni, P; Magistretti, PJ; Pellerin, L. Lactate is a preferential oxidative energy substrate over glucose for neurons in culture. J. Cereb. Blood Flow Metab 2003, 23(11), 1298–306. [Google Scholar] [CrossRef]
- Smith, D; Pernet, A; Hallett, WA; Bingham, E; Marsden, PK; Amiel, SA. Lactate: a preferred fuel for human brain metabolism in vivo. J. Cereb. Blood Flow Metab. 2003, 23(6), 658–64. [Google Scholar] [CrossRef] [PubMed]
- Schurr, A. Lactate: the ultimate cerebral oxidative energy substrate? J. Cereb. Blood Flow Metab. 2006, 26(1), 142–52. [Google Scholar] [CrossRef]
- Van Hall, G; Stømstad, M; Rasmussen, P; Jans, Ø; Zaar, M; Gam, C; Quistorff, B; Secher, NH; Nielsen, HB. Blood lactate is an important energy source for the human brain. J. Cereb. Blood Flow Metab. 2009, 29(6), 1121–29. [Google Scholar] [CrossRef]
- Chen, YJ; Mahieu, NG; Huang, X; Singh, M; Crawford, PA; Johnson, SL; Gross, RW; Schaefer, J; Patti, GJ. Lactate metabolism is associated with mammalian mitochondria. Nature Chem. Biol. 2016, 12(11), 937–43. [Google Scholar] [CrossRef]
- Hui, S; Ghergurovich, JM; Morscher, RJ; Jang, C; Teng, X; Lu, W; Esparza, LA; Reya, T; Zhan, L; Yanxiang Guo, J; White, E. Glucose feeds the TCA cycle via circulating lactate. Nature 2017, 551(7678), 115–8. [Google Scholar] [CrossRef]
- Li, X; Zhang, Y; Xu, L; Wang, A; Zou, Y; Li, T; Huang, L; Chen, W; Liu, S; Jiang, K; Zhang, X. Ultrasensitive sensors reveal the spatiotemporal landscape of lactate metabolism in physiology and disease. Cell Metab. 2023, 35(1), 200–11. [Google Scholar] [CrossRef]
- Warburg, O. The metabolism of carcinoma cells. J. Cancer Res. 1925, 9(1), 148–63. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123(3191), 309–14. [Google Scholar] [CrossRef]
- Theriault, JE; Shaffer, C; Dienel, GA; Sander, CY; Hooker, JM; Dickerson, BC; Barrett, LF; Quigley, KS. A functional account of stimulation-based aerobic glycolysis and its role in interpreting BOLD signal intensity increases in neuroimaging experiments. Neurosci. Biobehav. Rev. 2023, 153;105373. [Google Scholar] [CrossRef]
- Morelli, AM; Scholkmann, F. The Significance of Lipids for the Absorption and Release of Oxygen in Biological Organisms. Adv. Exp. Med. Biol. 2023, 93–9. [Google Scholar] [CrossRef]
- Vervust, W; Safaei, S; Witschas, K; Leybaert, L; Ghysels, A. Myelin sheaths can act as compact temporary oxygen storage units as modeled by an electrical RC circuit model. Proc. Nat Acad. Sci. 2025, 122(20), e2422437122.e2422437122. [Google Scholar] [CrossRef] [PubMed]
- Schurr, A. Where Is the Oxygen? The Mirage of Non-Oxidative Glucose Consumption During Brain Activity. NeuroSci. 2025, 6(4), 126. [Google Scholar] [CrossRef] [PubMed]
- Brandt, RB; Laux, JE; Spainhour, SE; Kline, ES. Lactate dehydrogenase in rat mitochondria. Archives of Biochemistry and Biophysics 1987, 259(2), 412–22. [Google Scholar] [CrossRef]
- Van Hall, G. Lactate as a fuel for mitochondrial respiration. Acta Physiol. Scand. 2000, 168(4), 643–6. [Google Scholar] [CrossRef]
- Atlante, A; de Bari, L; Bobba, A; Marra, E; Passarella, S. Transport and metabolism of L-lactate occur in mitochondria from cerebellar granule cells and are modified in cells undergoing low potassium dependent apoptosis. Biochim. et Biophys. Acta 2007, 2007:1767(11), 1285–99. [Google Scholar] [CrossRef]
- Pierre K. Pellerin L. Monocarboxylate transporters in the central nervous system: distribution, regulation and function. J. Neurochem. 2005, 94(1), 1–14. [CrossRef]
- Ueno, M; Chiba, Y; Murakami, R; Miyai, Y; Matsumoto, K; Wakamatsu, K; Takebayashi, G; Uemura, N; Yanase, K. Distribution of Monocarboxylate Transporters in Brain and Choroid Plexus Epithelium. Pharmaceutics 2023, 15(8), 2062. [Google Scholar] [CrossRef]
- Pierre, K; Pellerin, L; Debernardi, R; Riederer, B; Magistretti, P. Cell-specific localization of monocarboxylate transporters, MCT1 and MCT2, in the adult mouse brain revealed by double immunohistochemical labeling and confocal microscopy. Neuroscience 2000, 1009(3), 617–27. [Google Scholar] [CrossRef]
- Lundquist, AJ; Llewellyn, GN; Kishi, SH; Jakowec, NA; Cannon, PM; Petzinger, G.M; Jakowec, MW. Knockdown of Astrocytic Monocarboxylate Transporter 4 in the Motor Cortex Leads to Loss of Dendritic Spines and a Deficit in Motor Learning. Mol. Neurobiol. 2022, 59(2), 1002–17. [Google Scholar] [CrossRef] [PubMed]
- Dimmer, K-S; Friedrich, B; Lang, F; Deitmer, JW; Bröer, S. The low-affinity monocarboxylate transporter MCT4 is adapted to the export of lactate in highly glycolytic cells. Biochem. J. 2000, 350(1), 219–227. [Google Scholar] [CrossRef]
- Halestrap, A.P. The SLC16 gene family—Structure, role and regulation in health and disease. Mol. Asp. Med. 2013, 34, 337–349. [Google Scholar] [CrossRef]
- Valvona, CJ; Fillmore, HL; Nunn, PB; Pilkington, GJ. The regulation and function of lactate dehydrogenase a: therapeutic potential in brain tumor. Brain Pathol. 2016, 26(1), 3–17. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T; Hussien, R; Brooks, GA. Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: evidence of a mitochondrial lactate oxidation complex. Am. J. Physiol. Endocrinol. Metabol. 2006, 290(6), E1237-44. [Google Scholar] [CrossRef] [PubMed]
- Hussien, R; Brooks, GA. Mitochondrial and plasma membrane lactate transporter and lactate dehydrogenase isoform expression in breast cancer cell lines. Physiol. Genom. 2011, 43(5), 255–64. [Google Scholar] [CrossRef]
- https://www.broadinstitute.org/mitocarta/mitocarta30-inventory-mammalian-mitochondrial-proteins-and-pathways.
- Li, X; Zhang, Y; Xu, L; Wang, A; Zou, Y; Li, T; Huang, L; Chen, W.; Liu, S; Jiang, K; Zhang, X. Ultrasensitive sensors reveal the spatiotemporal landscape of lactate metabolism in physiology and disease. Cell Metab. 2023, 35(1), 200–211. [Google Scholar] [CrossRef]
- Koep, JL; Duffy, JS; Carr, JM; Brewster, ML; Bird, JD; Monteleone, JA; Monaghan, TD; Islam, H; Steele, AR; Howe, CA; MacLeod, DB. Preferential lactate metabolism in the human brain during exogenous and endogenous hyperlactataemia. The Journal of Physiology 2025, 603(22), 6783–800. [Google Scholar] [CrossRef]
- Bolaños, JP; Alberini, CM; Almeida, A; Barros, LF; Bonvento, G; Bouzier-Sore, AK; Dringen, R; Hardingham, GE; Hirrlinger, J; Magistretti, PJ; Marsicano, G; et al. Embracing the modern biochemistry of brain metabolism. J. Neurochem. 2025, 169(70), e70166. [Google Scholar] [CrossRef] [PubMed]
- Dienel, GA; Rothman, DL; Mangia, S. Comment on the Editorial “Embracing the Modern Biochemistry of Brain Metabolism”. Journal of Neurochemistry 2025, 169(8), e70197. [Google Scholar] [CrossRef] [PubMed]
- Andersen, JV; Aldana, BI; Bak, LK; Behar, KL; Borges, K; Carruthers, A; Cumming, P; Derouiche, A; Díaz-García, CM; Drew, KL; Duarte, JM; et al. Embracing scientific debate in brain metabolism. Journal of Neurochemistry 2025, 169(9), e70230. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A schematic presentation of the glycolytic free energy change through the eleven enzymatic steps of the pathway. The first step of glycolysis, the phosphorylation of glucose to glucose-6-phosphate by hexokinase, produces the largest free energy change (ΔG = -8.0 Kcal), the last step, the conversion of pyruvate to lactate by lactate dehydrogenase (LDH) produces the second largest free energy change (ΔG = -6.0 Kcal). The conversion of pyruvate to lactate also produces NAD+, keeping up with the cyclical nature of the pathway, while supplying the redox cofactor for additional redox reactions. NAD+ = nicotinamide adenine dinucleotide; NADH = nicotinamide adenine dinucleotide, reduced form; cLDH = cytosolic lactate dehydrogenase. ATP = adenosine triphosphate; G6P = glucose 6-phosphate; F6P = fructose 6-phosphate; FDP = fructose 1,6-biphosphate; GAP = glyceraldehyde 3-phosphate; 3PGA = 3-phosphoglycerate; 2PGA = 2-phosphoglycerate; PEP - phosphoenolpyruvate; *non-equilibrium reaction; ΔG = free energy change in Kcal.
Figure 1.
A schematic presentation of the glycolytic free energy change through the eleven enzymatic steps of the pathway. The first step of glycolysis, the phosphorylation of glucose to glucose-6-phosphate by hexokinase, produces the largest free energy change (ΔG = -8.0 Kcal), the last step, the conversion of pyruvate to lactate by lactate dehydrogenase (LDH) produces the second largest free energy change (ΔG = -6.0 Kcal). The conversion of pyruvate to lactate also produces NAD+, keeping up with the cyclical nature of the pathway, while supplying the redox cofactor for additional redox reactions. NAD+ = nicotinamide adenine dinucleotide; NADH = nicotinamide adenine dinucleotide, reduced form; cLDH = cytosolic lactate dehydrogenase. ATP = adenosine triphosphate; G6P = glucose 6-phosphate; F6P = fructose 6-phosphate; FDP = fructose 1,6-biphosphate; GAP = glyceraldehyde 3-phosphate; 3PGA = 3-phosphoglycerate; 2PGA = 2-phosphoglycerate; PEP - phosphoenolpyruvate; *non-equilibrium reaction; ΔG = free energy change in Kcal.
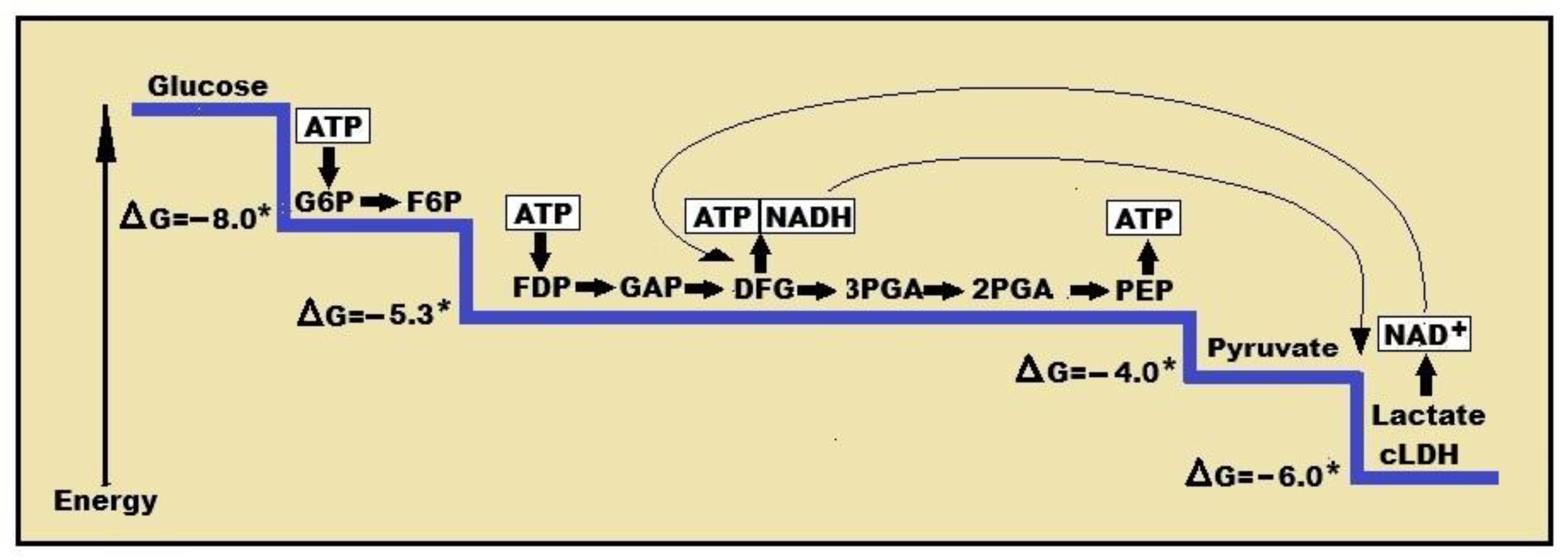
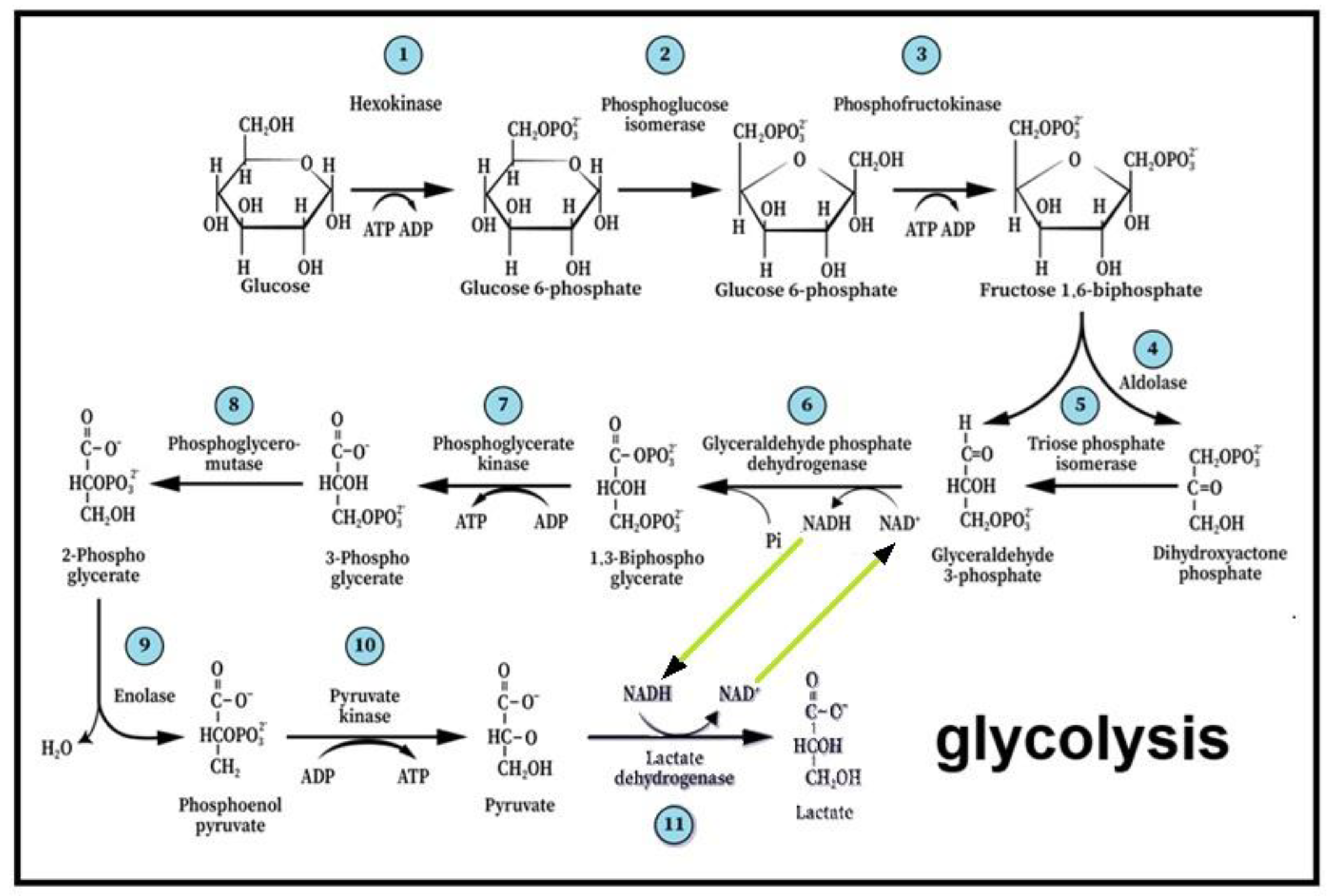
Figure 2.
Glycolysis, the first elucidated metabolic pathway, describes the breakdown of glucose through 11 enzymatic reactions, where the hexose molecule is cleaved into two triose molecules. In the process, two molecules of ATP are invested in phosphorylation along with the reduction of one NAD+ equivalent to NADH and the yield of four ATP molecules. The last step of the pathway is the production of its end-products, lactate and NAD+. This last step and the reproduction of NAD+ create the cyclical nature of the pathway, supplying the sixth step with a renewed supply of the reducing power. The enzyme responsible for the eleventh step is cytosolic lactate dehydrogenase (cLDH), which differs from the mitochondrial enzyme (mLDH) that is responsible for the conversion of lactate to pyruvate, the substrate of the tricarboxylic acid (TCA) cycle.
Figure 2.
Glycolysis, the first elucidated metabolic pathway, describes the breakdown of glucose through 11 enzymatic reactions, where the hexose molecule is cleaved into two triose molecules. In the process, two molecules of ATP are invested in phosphorylation along with the reduction of one NAD+ equivalent to NADH and the yield of four ATP molecules. The last step of the pathway is the production of its end-products, lactate and NAD+. This last step and the reproduction of NAD+ create the cyclical nature of the pathway, supplying the sixth step with a renewed supply of the reducing power. The enzyme responsible for the eleventh step is cytosolic lactate dehydrogenase (cLDH), which differs from the mitochondrial enzyme (mLDH) that is responsible for the conversion of lactate to pyruvate, the substrate of the tricarboxylic acid (TCA) cycle.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



