
Submitted:
22 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
Ethiopian finches (Fringillidae) provide a continental-scale system for examining how environmental gradients shape functional trait evolution. This review synthesizes current knowledge on the evolution of beak traits in Ethiopian finches across the country’s highlands, integrating evidence from diet composition, habitat heterogeneity, thermoregulatory demands, and human-mediated environmental change. Beak diversification reflects the interaction of lineage-specific phylogenetic constraints, functional trade-offs, and plastic responses, producing adaptive divergence comparable in magnitude to classic island radiations despite ongoing gene flow. Anthropogenic pressures, including agricultural expansion, habitat fragmentation, and urbanization, interact with natural gradients to favor generalized morphologies while eroding specialized traits. Genetic analyses highlight key loci, such as BMP4 and ALX1, that may enable rapid adaptation to environmental and anthropogenic pressures. This synthesis emphasizes the need for empirical research to test trait-environment relationships and clarifies gaps in current knowledge, providing a framework for future ecological and evolutionary studies in Ethiopian finches, with implications for conservation and management of finch populations in rapidly changing landscapes.
Keywords:
Ethiopian finches
; beak morphology
; adaptive evolution. trait-environmental association
; phenotypic diversification
; ecological gradients
1. Introduction
Morphological traits in nature often serve multiple functions simultaneously (Arnolds, 1983), meaning that evolution must navigate inherent trade-offs rather than optimizing a single purpose. Bird beak morphology represents a prominent and well-studied example of evolutionary adaptation in vertebrates, linking phenotypic variation with specific ecological and environmental pressures. Since the realization of Darwin’s finches from the Galápagos Islands as a model system for adaptive radiation, studies of avian beaks have generated foundational insights into how morphological traits evolve in response to variation in diet, habitat, and climate (Yalin et al., 2024; Grant & Grant, 2008). The immense diversity of beak shapes, from robust, seed-cracking (granivorous) forms to slender, probing structures, reflects prolonged natural selection acting on multiple functional demands across heterogeneous environments. Beak morphology integrates various ecological functions, including preening and parasite removal (Clayton et al., 2004), nest construction (Collias & Collias, 1964), singing (Podos, 2001), and thermoregulation (Tattersall et al., 2016), highlighting its multifunctional nature and the complex selective pressure that shapes it (Meier et al., 2017; Friedman et al., 2017).
Recent molecular and functional research further demonstrates that beak evolution is shaped not only by ecological pressures but also by intrinsic factors that constrain or guide morphological change. These intrinsic factors include phylogenetic history, determining the inherited craniofacial framework available to selection; genetic constraints, such as limited variation in key developmental genes, restricting the range of possible beak shapes and sizes; and developmental pathways, influencing how variation in size and shape can be expressed across life stages (Bright et al., 2016; Felice & Goswami, 2018; Navalón et al., 2020). These factors collectively modulate the trajectory of morphological change, indicating that adaptation is contingent on both evolutionary legacy and the immediate environmental context (Peterson et al., 2019; Badyaev et al., 2008). In classic adaptive radiation systems, such as Darwin's finches, beak size and shape correlate closely with available food resources, and microevolutionary shifts have been documented over just a few generations in response to changes in resource abundance, illustrating the rapidity with which natural selection can alter phenotypic distributions (Grant & Grant, 2002; Herrel et al., 2005).
Ethiopia provides a uniquely powerful continental context for examining these processes. The country is characterized by pronounced altitudinal gradients from lowland savannas to highland plateaus exceeding 4,500 m, and encompasses a mosaic of habitat types including forests, shrublands, grasslands, and afro-alpine moorlands (Sebsebe et al., 2021; Friis et al., 2010). These environmental gradients impose distinct selective pressures on avian populations, influencing diet composition, foraging strategies, and morphological traits. Variation in rainfall and aridity can alter seed availability and quality, favoring particular beak shapes and sizes in different habitats. Habitat structure may further mediate ecological interactions, such as competition and foraging mode. Together, these factors shape adaptive variation in beak morphology, particularly depth, length, and robustness.
The Ethiopian Fringillidae, commonly known as true finches, comprise a diverse group of small to medium-sized passerine birds that occupy a wide range of habitats across the country (Ash & Miskell, 2009). Notable species include the Ethiopian Siskin (Serinus nigriceps), the Brown-rumped Seedeater (Crithagra tristriata), and the Yellow-rumped Serin (Crithagra xanthopygia), among others, which are typically associated with the highland and montane regions of the Ethiopian Highlands (Ash & Miskell, 2009; Jose, 2004). These species exhibit remarkable variation in body size, plumage, and beak morphology across different ecological niches and strategies. Finches inhabiting open grassland and dry lowlands are typically associated with granivorous diets, requiring stout, conical beaks capable of cracking hard seed coats. Whereas, species in wetter, forested areas may supplement their diet with insects or nectar, favoring longer and slimmer beaks that allow for the precise handling of these food items (Ash & Miskell, 2009; Jose, 2004). In addition to foraging, beak morphology in these finches influences thermoregulation, social interactions, and nest construction, highlighting the multifunctional nature of this trait (Meier et al., 2017; Peterson et al., 2019).
Despite their ecological significance, studies of Ethiopian finches remain limited and fragmented, often focusing on isolated populations or single species. Empirical data on quantitative trait-environment relationships, particularly across multiple species and environmental gradients in Ethiopia, remain scarce, representing a critical gap in understanding the adaptive evolution of these finches. This review, therefore, consolidates current knowledge on beak morphology in Ethiopian Fringillidae, emphasizing how environmental factors such as diet composition, habitat heterogeneity, and climatic variation relate to morphological diversity. By integrating regional ecological data with global principles of avian evolution, it highlights potential recurring patterns in trait–environment associations and provides context for prioritizing future research on the evolutionary ecology of Ethiopia’s finch fauna.
2. Phylogenetic and Biogeographic Context
Ethiopian finches (family Fringillidae) form a phylogenetically heterogeneous assemblage shaped by multiple independent colonization events into the Ethiopian highlands (Lerner et al., 2011; Zuccon et al., 2012). Unlike classic island radiations such as Darwin’s finches, which originated from a single ancestral lineage and diversified rapidly under strong ecological opportunity (Grant & Grant, 1986), Ethiopian finches derive from multiple evolutionary lineages spanning different subfamilies (Fringillinae and Carduelinae) and genera, with the genus Crithagra representing the most diverse highland lineage (Clement et al., 1993; Del & Collar, 2014). This deep evolutionary structure establishes lineage-specific craniofacial architectures that constrain the range and direction of beak morphological evolution across species, reflecting broader macroevolutionary limits on avian skull and beak diversification (Abzhanov et al., 2006; Bright et al., 2016).
These phylogenetic constraints are evident in the persistence of distinct beak architectures among lineages despite shared environments. For instance, taxa such as Spinus nigriceps retain relatively slender, pointed bills, whereas highland Serins such as Crithagra ankoberensis consistently exhibit deeper, more robust beaks (Ash & Miskell, 1998; Clement et al., 1993). Such differences reflect an inherited developmental and genetic framework that modulates how selection can act on beak size and shape, emphasizing that adaptive responses are contingent on evolutionary history rather than solely on contemporary ecological conditions (Badyaey et al., 2008; Bright et al., 2016).
Despite these constraints, Ethiopian finches potentially exhibit patterns of morphological convergence across phylogenetically distant taxa. Comparable beak forms recur in unrelated species occupying comparable ecological settings, indicating that shared selective regimes can produce repeated evolutionary outcomes even in the absence of close ancestry (Losos, 2011; Schluter, 2000). Such potential convergence is likely facilitated by conserved developmental pathways implicated broadly in avian craniofacial evolution, including those involving genes such as BMP4 and Alx1, which may enable parallel responses to selection, while lineage-specific regulation limits the precise phenotypic space accessible to each clade (Abzhanov et al., 2004; Abzhanov et al., 2006; Mallarino et al., 2011).
Biogeography further structures these evolutionary dynamics. The Ethiopian highland functions as a continental ‘‘sky island’’ system composed of isolated plateaus, mountain massifs, and deeply incised valleys (Friis et al., 2010; Sebsebe et al., 2021). This topographic complexity promotes spatial subdivision of populations and intermittent isolation without the absolute barriers characteristic of oceanic islands (Fjeldså & Bowie, 2008; Rahbek et al., 2019). As a result, populations may diverge morphologically while retaining limited gene flow, producing fine-scale differentiation rather than rapid, lineage-wide radiations (Nosil, 2012). Patterns of endemism among Ethiopian finches are best interpreted within this phylogeographic framework. Several highland-restricted taxa occupy narrow geographic ranges shaped by historical dispersal, isolation, and long-term persistence within montane refugia (Ash & Atkins, 2009; Fjeldså et al., 2012). These distributions reflect the combined influence of lineage history and landscape structure, rather than single-founder events or recent ecological specialization alone (Wiens & Donoghue, 2004). Recurrent isolation across highland blocks creates opportunities for both divergences within lineages and convergence among lineages exposed to similar environments (Schluter, 2000). Taken together, the phylogenetic diversity and complex biogeography of Ethiopian finches define a continental evolutionary system in which adaptive change occurs within strong historical and spatial constraints; this context is essential for interpreting trait-environment relationships, as beak morphological variation emerges from the interaction of inherited developmental limits, repeated isolation, and exposure to heterogeneous selective regimes across the Ethiopian landscape (Nosil, 2012; Felice & Goswami, 2018; Rahbek et al., 2019).
3. Trait-Environment Associations in Ethiopian Finches
3.1. Diet and Foraging Ecology
Diet and foraging ecology constitute primary axes along which natural selection shapes beak morphology in birds, reflecting functional demands with food acquisition, handling, and processing (Grant & Grant, 2008; Schluter, 2000; Benkman, 2003). However, the evolutionary consequences of dietary specialization are rarely determined by food type alone but are influenced by environmental context, interspecific interactions, and phylogenetic constraints that modulate both the direction and magnitude of morphological divergence (Losos, 2011; Bright et al., 2016). As a result, similar diets can be associated with different beak forms, while similar beak morphologies may arise under contrasting ecological conditions.
Within this broader evolutionary framework, Ethiopian finches provide a particularly informative system for examining how dietary selection interacts with steep environmental gradients and complex ecological settings to generate adaptive variation in beak morphology (Grant & Grant, 2008; Schluter, 2000). Although detailed functional-morphology studies in Ethiopian finches are limited, general finch biomechanics consistently show that granivorous species feeding on hard seeds evolve deep, robust, conical beaks optimized for high bite force and mechanical efficiency (Grant & Grant, 2002; Herrel et al., 2005; Parchman & Benkman, 2002). Several Ethiopian finches rely heavily on large, tough seeds, suggesting they exhibit similar functional adaptations, reflecting the same seed-cracking principle documented across lineages (Ash & Miskell, 2009; Jose, 2004). By contrast, finches that forage on small, soft, or spatially dispersed resources, such as smaller seeds, insects, or floral resources, typically exhibit slender, pointed, or elongated beaks. These beaks enhance precision, probing ability, and access to concealed food items such as arthropods within foliage or seeds embedded in complex plant structures (Homberger, 2001; Parchman & Benkman, 2002).
Across both dietary strategies, habitat structure and climatic pressures interact with feeding requirements to produce coordinated morphological divergence along multiple ecological axes rather than isolated responses to diet alone (Benkman, 2003; Losos, 2011). Beak forms specialized for force generation are favored in arid lowlands and highland grasslands, where seed hardness, seasonal scarcity, and reduced dietary diversity impose strong mechanical constraints on foraging (Grant & Grant, 2008; Herrel et al., 2005). Conversely, beaks optimized for precision and reach are advantageous in complex habitats, such as forests or shrublands, where food resources are dispersed or concealed. This interplay between diet, environment, and functional trade-offs generates structured variation across populations, linking local resource conditions to population-level differentiation (Badyaev et al., 2008; Schluter, 2000; Podos et al., 2004).
Comparative studies of avian adaptive radiations, such as Darwin’s finches in the Galápagos, show that beak morphology often reflects dietary niches and functional performance, with measurable differences among species adapted to contrasting food types (Grant & Grant, 2002, 2008). In addition, broad comparative analyses across modern birds indicate that similar beak shapes can be associated with disparate dietary regimes, suggesting that dietary selection may act on particular functional axes rather than producing uniform changes in overall beak form (Bright et al., 2016; Losos, 2011; Schluter, 2000). Building on the general functional framework of finch beak morphology, we can examine how dietary selection acts within Ethiopian finch populations, revealing fine-scale differentiation driven by local ecological conditions. Granivorous species with robust, seed-cracking beaks and insectivorous or nectarivorous species with slender, precision-oriented beaks (Grant & Grant, 2002; Herrel et al., 2005; Parchman & Benkman, 2002; Homberger, 2001; Podos, 2001) exhibit substantial intraspecific variation in relation to locally abundant food resources, including differences in seed size distributions, plant species composition, and foraging substrates (Grant & Grant, 2008; Badyaev et al., 2008). Whether these differences represent heritable divergence or phenotypic plasticity remains unresolved, but their persistence underscores the importance of ecological heterogeneity in maintaining morphological diversity.
Intraspecific competition further modulates selective pressures. Overlap in resource use increases the fitness costs of competition, intensifying selection on traits that improve foraging efficiency on underexploited resources (Schluter, 2000; Grant & Grant, 2006). Divergence in beak morphology, therefore, facilitates resource partitioning by allowing species to specialize on different seed sizes, food types, or foraging strata, reducing direct competitive interference (Benkman, 2003). These patterns illustrate that functional differentiation emerges from the interaction of multiple ecological pressures, producing predictable trait distributions across sympatric species while respecting lineage-specific constraints (Schluter, 2000; Losos, 2011). Importantly, evolutionary change is not random across all aspects of beak shape but is biased toward functional dimensions that enhance performance on alternative resources while remaining developmentally accessible within a given lineage (Alberch, 1982; Schluter, 2000; Wake & Larson, 1987). As a result, species occupying similar dietary niches may diverge along morphological dimensions, reflecting the combined influence of ecological opportunity and lineage-specific developmental constraints rather than identical adaptive endpoints (Losos, 2011).
3.2. Habitat and Environmental Gradients
Habitat structure and elevational gradients exert strong but variable selective pressures on beak morphology, shaping foraging performance through ecological trade-offs rather than deterministic relationships between environment and form (Bock & von Wahlert, 1965; Schluter, 2000; Losos, 2011). Variation in vegetation complexity, from multi-layered canopies and dense understory in montane forests to sparse, low-lying cover in grasslands, creates diverse structural environments that challenge foraging efficiency (MacArthur & MacArthur, 1961; Bock & Jones, 2004; Bell, 2000). Differences in resource distribution, such as clumped hard seeds versus dispersed insects or nectar, and physical constraints across elevation, including temperature, season length, and terrain steepness, further shape the availability and predictability of food resources (Endler, 1986; Benkman, 2003). These factors generate heterogeneous selective landscapes in which similar ecological conditions can produce divergent morphological outcomes due to historical contingency and constraint (Losos, 2011). As a result, finches frequently exhibit slender or moderately elongated beaks in structurally complex habitats. These morphologies are favored where foraging success depends on precision, reach, and maneuverability within heterogeneous substrates (Werner & Sherry, 1987; Norberg, 1990). Individuals that can efficiently extract concealed or dispersed food items, such as foliage-dwelling insects or seeds embedded in complex plant architecture, gain a selective advantage by reducing handling time and energy expenditure (Homberger, 2001; Werner & Sherry, 1987). This advantage comes at the expense of reduced mechanical force, highlighting a functional trade-off between precision and power (Figure 1) (Herrel et al., 2005; Benkman, 2003).
In contrast, open habitats, including grasslands, savannas, and agro-ecosystems, consistently favor shorter, deeper, and more robust beaks. These environments impose mechanical demands associated with ground foraging, hard seed processing, and temporally clumped resources. Robust beaks enhance bit force and efficiency but reduce versatility, suggesting that habitat openness selects for mechanical optimization over foraging flexibility (Benkman, 2003). Importantly, similar beak forms arise across distantly related finch taxa occupying comparable open habitats, indicating repeated adaptive responses rather than shared ancestry (Grant & Grant, 2008; Parchman & Benkman, 2002). These patterns demonstrate how habitat complexity interacts with dietary demands to shape beak morphology, such that mechanical force is prioritized in granivores of open habitats while precision is favored in insectivores of complex forests, producing population-level differentiation that reflects both ecological pressures and lineage-specific constraints (Badyaev, et al., 2008).
Elevation further modifies these habitat-driven pressures by constraining resource diversity, seasonality, and energetic budgets. High-elevation environments, characterized by short growing seasons and lower primary productivity, tend to favor generalized or mechanically efficient beak forms capable of exploiting a narrow and unpredictable resource base (Bock, 1980; Grant & Grant, 2008). At lower elevations, prolonged productivity and higher plant diversity permit finer-scale specialization, allowing beak morphology to track subtle differences in resource structure (Schluter, 2000). These patterns suggest that elevation gradients do not merely scale habitat effects but reshape the adaptive landscape by altering the balance between specialization and flexibility (Bock, 1980; Schluter, 2000). Selective pressures also operate at microhabitat scales. Variation in canopy cover, disturbance intensity, and vegetation layering within the same elevational zone can produce divergent foraging contexts, promoting intraspecific variation in beak morphology (Werner & Sherry, 1987; Badyaev et al., 2008). Whether such variation reflects genetic differentiation or phenotypic plasticity remains unresolved, but its persistence implies that fine-scale ecological heterogeneity can maintain morphological diversity even in the absence of geographic isolation (Homberger, 2003; Badyaev et al., 2008).
Taken together, these patterns indicate that beak morphology in Ethiopian finches is likely shaped by interaction among environmental gradients, function trade-offs, and phylogenetic constraints (Schluter, 2000; Losos, 2011). Similar ecological conditions repeatedly favor comparable beak designs, yet lineage-specific developmental pathways limit the range of achievable morphologies. This tension between convergence and constraint is a common feature of avian morphological evolution, shaping the adaptive trajectories of beak form across diverse taxa (Bright et al., 2016). Ethiopian finches situated across steep environmental gradients and heterogeneous habitats allow direct observation of how ecological variation drives adaptive divergence while preserving evolutionary contingency.
3.3. Climatic Factors and Thermoregulation
Climatic variables, particularly temperature, precipitation, and seasonality, shape beak morphology by modifying the selective landscape along multiple ecological axes (Danner & Greenberg, 2015). Beak morphology is inherently multifunctional, and modifications that enhance one function often impose trade-offs on others. High temperatures and arid conditions elevate thermal stress and increase evaporative water loss, favoring relatively larger, more vascularized beaks that enhance heat dissipation (Symonds & Tattersall, 2010; Greenberg et al., 2012). This increase in surface area typically produces longer, flatter, or thinner beaks, which trade mechanical leverage, thereby limiting the force that can be applied to crack hard seeds or manipulate other food resources (Herrel et al., 2005; Parchman & Benkman, 2002).
Conversely, cooler or high-elevation environments promote the evolution of smaller, more compact beaks that minimize heat loss while maintaining functional efficiency for feeding (Greenberg et al., 2012). These patterns illustrate that climatic selection acts within functional trade-offs, producing context-dependent outcomes rather than uniform morphological responses. Comparative studies across birds further indicate that selection for thermoregulation can occur partly independently of foraging demands, resulting in beak forms that balance heat dissipation with mechanical performance (Friedman et al., 2017; van de Ven et al., 2016; Danner et al., 2017). In Ethiopian finches, steep elevational gradients and pronounced seasonal variation are likely to influence the balance among thermoregulation, feeding efficiency, and water conservation. Rather than favoring a single optimal beak form, these gradients may generate shifting selective regimes in which similar climatic pressures are associated with different morphological solutions depending on dietary demands and habitat structure.
Seasonal fluctuations in precipitation further influence selective pressures by altering food availability. Prolonged dry periods intensify energetic and mechanical constraints, whereas wetter seasons relax these constraints and permit a broader range of viable functional strategies (Benkman, 2003; Nosil, 2012). Taken together, climatic gradients interact with dietary and habitat-based selection to shape the adaptive landscape of beak morphology across Ethiopian finch assemblages (Figure 1).
3.4. Human-Mediated Environmental Change
Human activities are rapidly modifying landscapes occupied by Ethiopian finches, creating novel selective pressures that overlay and in some cases override natural ecological and climatic gradients (Asefa et al., 2017). Agricultural expansion, deforestation, overgrazing, habitat fragmentation, and urbanization alter vegetation structure and resource availability across highland and mid-elevation regions, potentially reshaping trait-environment relationships historically structured by natural gradients (Newmark, 2002). Analogous studies in other continental birds suggest that such anthropogenic pressures amplify selective trade-offs between feeding mechanics and thermoregulation, favoring phenotypically plastic individuals capable of exploiting novel food sources (Friedman et al., 2017; Evans et al., 2011; Alberti et al., 2017; Hendry et al., 2008; Charmantier et al., 2008).
In other avian systems, such conditions have been shown to coincide with reduced dietary specialization and the maintenance of generalized foraging strategies, often supported by beak morphologies capable of producing adequate bite force across a narrow range of mechanically similar food types (Shochat et al. 2006; Benton et al., 2003; Devictor et al., 2008; De León et al., 2011).
The conversion of native grasslands, shrublands, and forest edges into agricultural mosaics dominated by crops such as teff, wheat, barley, and sorghum modifies seed size, hardness, diversity, and seasonal availability. Compared to heterogeneous native floras, cultivated and ruderal plant assemblages tend to produce more homogeneous and predictable food resources. In other avian systems, such conditions have been shown to coincide with reduced dietary specialization and the persistence of generalized foraging strategies, often involving beak morphologies capable of generating sufficient bite force across a limited range of mechanically similar food types (Shochat et al. 2006; Benton et al., 2003; Devictor et al., 2008; De León et al., 2011). Despite limited direct evidence from Ethiopian finches, these patterns suggest that agricultural landscapes may function as ecological filters, disproportionately disadvantaging specialists adapted to diverse native seed assemblages while permitting broader functional tolerance. Endemic highland species such as Ankober Serin (Crithagra ankoberensis) and Yellow-throated Seedeater (Serinus flavigula), which rely on native seed plants that decline under agricultural expansion, potentially increase their vulnerability to landscape transformation. By contrast, generalist species are more likely to exploit anthropogenic food sources, contributing to shifts in community composition and functional trait distributions (Deng et al., 2025).
Human-modified landscapes, those shaped by deforestation and overgrazing, also alter physical and microclimatic conditions by simplifying vegetation, reducing native seed-producing plants, and increasing exposure, solar radiation, and thermal variability (Meseret et al., 2023). These changes interact with the multifunctional role of the Ethiopian finch beak as both a feeding apparatus and a thermoregulatory organ, thereby reshaping functional trade-offs that govern beak morphology (Fig. 1) (Tattersall et al., 2016). Beak forms optimized for seed processing or thermoregulation under natural conditions may perform differently in fragmented or cultivated habitats, where food density, exposure, and energetic demands are altered (Grant & Grant, 2006; Herrel et al., 2005). As a result, finches in human-modified areas may exhibit a shift toward more generalized beak shapes capable of handling a narrower range of predictable food resources while maintaining thermal balance, potentially reducing the prevalence of highly specialized morphologies (Shochat et al., 2006; Benton et al., 2003). In a similar pattern, Ethiopian finches such as Crithagra ankoberensis and Serinus flavigula may experience selection for beak structures that balance feeding efficiency and thermoregulation in response to scarce or homogenized resources and altered microclimatic conditions. With scarce or homogenized resources and altered microclimates selecting for beak structures that balance feeding efficiency and thermoregulation. Habitat fragmentation further compounds these effects by reducing population connectivity and limiting gene flow, particularly for endemic and range-restricted species confined to small, isolated populations (Fahrig, 2003; Haddad et al., 2015). Reduced connectivity may constrain adaptive responses to rapidly changing selective pressures, while altered competitive dynamics in fragmented habitats can facilitate the expansion of generalist species that outcompete specialists, destabilizing previously adaptive trait-environment associations (Devictor et al., 2008; Bonte et al., 2012).
Urbanization introduces an additional and distinct selective regime by replacing natural substrates with artificial surfaces, increasing access to anthropogenic food sources, and generating persistent urban-island effects (Yeh, 2004; Aronson et al., 2014). In towns and cities across the Ethiopian Highlands, finches exploit human-provided foods that are often energy-dense, mechanically uniform, and spatially predictable. These conditions tend to favor beak morphologies that balance moderate bite force with efficient handling rather than extreme specialization (Shochat et al., 2006). Collectively, across agricultural transformation, habitat simplification, fragmentation, and urbanization, reconfigure Ethiopian landscapes into hybrid selective regimes in which resource predictability, mechanical demands, thermal conditions, and demographic constraints jointly shape patterns of beak morphological variation. Rather than reflecting simple extensions of natural gradients, these environments impose novel combinations of pressures that may decouple historically adaptive trait-environmental associations.
4. Conclusion
Finches exhibit extensive and finely structured variation in beak morphology, reflecting adaptive response to complex ecological, climatic, and anthropogenic gradients. Evidence from comparative studies of finches globally, combined with knowledge of local ecological and climatic patterns, suggests that variation in Ethiopian finch beak morphology likely arises from the interplay of dietary specialization, habitat heterogeneity, and thermal environment. Granivorous species possess deep, robust, conical beaks, while insectivorous or nectar-feeding species likely show slender, elongated beaks; elevational and thermal gradients may further influence the size and shape of these beaks. Human-altered environments, shaped by agriculture, habitat fragmentation, overgrazing, and urbanization, favor generalist or versatile forms, while phylogenetic history and phenotypic plasticity modulate adaptive trajectories. These patterns indicate that Ethiopian Fringillidae provide a compelling framework for understanding how environmental gradients and human activities jointly shape morphological diversity, functional trade-offs, and evolutionary trajectories of finch beak traits. Future research should focus on quantifying trait-environment relationships across species and elevations, distinguishing plastic from genetic contributions to beak variation, and assessing the vulnerability of specialist taxa to environmental change. Such studies will clarify the evolutionary ecology of continental adaptive systems and inform conservation strategies for highland bird populations.
Ethics Approval and Consent
Ethics approval was not required for this study as it did not involve human participants or vertebrate animals.
Data Availability
No new data were generated or analysed in support of this review article.
Competing Interests
The author declares no conflict of interest.
Author Contributions
A.T. conceived the study, performed the literature review and synthesis, and wrote the manuscript.
Funding
This review received no external funding. .
Acknowledgments
I sincerely thank Mr. Tewodros Kabtamu for his guidance and support during the preparation of this manuscript. Portions of this manuscript were revised for clarity and grammar using ChatGPT (OpenAI). The author reviewed and verified all content.
References
- Abzhanov, A.; Kuo, W.P.; Hartmann, C.; Grant, B.R.; Grant, P.R.; Tabin, C.J. The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches. Nature 2006, 442, 563–567. [Google Scholar] [CrossRef]
- Abzhanov, A.; Protas, M.; Grant, B.R.; Grant, P.R.; Tabin, C.J. Bmp4 and Morphological Variation of Beaks in Darwin's Finches. Science 2004, 305, 1462–1465. [Google Scholar] [CrossRef]
- Alberch, P. Developmental constraints in evolutionary processes. In Evolution and development; Bonner, J. T., Ed.; Springer: Berlin, Germany, 1982; pp. 313–332. [Google Scholar]
- Alberti, M.; Marzluff, J.M.; Shulenberger, E.; Bradley, G.; Ryan, C.; Zumbrunnen, C.; Hoelzel, A.R. Global urban signatures of phenotypic change in animal and plant populations. Proceedings of the National Academy of Sciences 2017, 114, 8957–8962. [Google Scholar] [CrossRef]
- Arnold, S.J. Morphology, performance, and fitness. Am. Zool 1983, 23, 347–361. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Clilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Asefa, A.; Davies, A.B.; McKechnie, A.E.; Kinahan, A.A.; van Rensburg, B.J. Effects of anthropogenic disturbance on bird diversity in Ethiopian montane forests. Condor 2017, 119, 416–430. [Google Scholar] [CrossRef]
- Ash, J.S.; Atkins, J.; Ash, C. Birds of Ethiopia and Eritrea: An atlas of distribution. 2009. Available online: https://api.semanticscholar.org/CorpusID:129869873.
- Badyaev, A.V.; Schwabl, H.; Young, R.L.; A Duckworth, R.; Navara, K.J.; Parlow, A. Adaptive sex differences in growth of pre-ovulation oocytes in a passerine bird. Proc. R. Soc. B: Biol. Sci. 2005, 272, 2165–2172. [Google Scholar] [CrossRef]
- Badyaev, A.V.; Young, R.L.; Hill, G.E.; Duckworth, R.A. Evolution of sex-biased maternal effects in birds. Proceedings of the Royal Society B 2008, 275, 915–923. [Google Scholar]
- Bell, H.L. Foraging behavior and microhabitat use in forest birds. Journal of Avian Biology 2000, 31, 355–363. [Google Scholar]
- Benkman, C.W. Divergent selection drives the adaptive radiation of crossbills. Evolution 2003, 57, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Bock, C.E.; Jones, Z.F. Avian habitat evaluation: should counting birds count? Frontiers in Ecology and the Environment 2004, 2, 403–410. [Google Scholar] [CrossRef]
- Bock, W.J. The Definition and Recognition of Biological Adaptation. Am. Zoöl. 1980, 20, 217–227. [Google Scholar] [CrossRef]
- Bock, W.J.; von Wahlert, G. Adaptation and the Form-Function Complex. Evolution 1965, 19, 269. [Google Scholar] [CrossRef]
- Bonte, D.; Van Dyck, H.; Bullock, J.M.; Coulon, A.; Delgado, M.; Gibbs, M.; Lehouck, V.; Matthysen, E.; Mustin, K.; Saastamoinen, M.; et al. Costs of dispersal. Biol. Rev. 2012, 87, 290–312. [Google Scholar] [CrossRef]
- Bright, J.A.; Marugán-Lobón, J.; Cobb, S.N.; Rayfield, E.J. The shapes of bird beaks are highly controlled by nondietary factors. Proc. Natl. Acad. Sci. 2016, 113, 5352–5357. [Google Scholar] [CrossRef]
- Charmantier, A.; McCleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptive Phenotypic Plasticity in Response to Climate Change in a Wild Bird Population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef]
- Cheng, Y.; Miller, M.J.; Lei, F. Molecular Innovations Shaping Beak Morphology in Birds. Annu. Rev. Anim. Biosci. 2024, 13, 99–119. [Google Scholar] [CrossRef]
- Clayton, D.H.; Moyer, B.R.; E Bush, S.; Jones, T.G.; Gardiner, D.W.; Rhodes, B.B.; Goller, F. Adaptive significance of avian beak morphology for ectoparasite control. Proc. R. Soc. B: Biol. Sci. 2005, 272, 811–817. [Google Scholar] [CrossRef]
- Clement, P.; Harris, A.; Davis, J. Finches and sparrows: An identification guide; Helm Field Guides, 1993. [Google Scholar]
- De León, L.F.; McNew, S.M.; Hendry, A.P. Evolutionary divergence in Darwin’s finches: Recent advances and future directions. Evolutionary Ecology 2011, 25, 305–328. [Google Scholar]
- De León, L.F.; Podos, J.; Gardezi, T.; Herrel, A.; Hendry, A.P. Darwin's finches and their diet niches: the sympatric coexistence of imperfect generalists. J. Evol. Biol. 2014, 27, 1093–1104. [Google Scholar] [CrossRef]
- del Hoyo, J.; Collar, N.J. HBW and BirdLife International illustrated checklist of the birds of the world. In Non-passerines; Lynx Edicions, 2014; Volume 1. [Google Scholar]
- Deng, J.; Zhu, Y.; Luo, Y.; Zhong, Y.; Tu, J.; Yu, J.; He, J. Urbanization drives biotic homogenization of the avian community in China. Integr. Zoöl. 2025, 20, 60–72. [Google Scholar] [CrossRef]
- Devictor, V.; Julliard, R.; Jiguet, F. Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 2008, 117, 507–514. [Google Scholar] [CrossRef]
- Endler, J.A. Natural selection in the wild; Princeton University Press, 1986. [Google Scholar]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Feijó, A.; Wen, Z.; Cheng, J.; Ge, D.; Xia, L.; Yang, Q. Divergent selection along elevational gradients promotes genetic and phenotypic disparities among small mammal populations. Ecol. Evol. 2019, 9, 7080–7095. [Google Scholar] [CrossRef]
- Felice, R.N.; Goswami, A. Developmental origins of mosaic evolution in the avian cranium. Proc. Natl. Acad. Sci. 2018, 115, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Fjeldså, J.; Bowie, R.C.K. New perspectives on the origin and diversification of Africa’s forest avifauna. Afr. J. Ecol. 2008, 46, 235–247. [Google Scholar] [CrossRef]
- Friedman, N.R.; Miller, E.T.; Ball, J.R.; Kasuga, H.; Remeš, V.; Economo, E.P. Evolution of a multifunctional trait: shared effects of foraging ecology and thermoregulation on beak morphology, with consequences for song evolution. Proc. R. Soc. B: Biol. Sci. 2019, 286, 20192474. [Google Scholar] [CrossRef]
- Friis, I.; van Breugel, P. Atlas of the potential vegetation of Ethiopia. In Biologiske Skrifter; The Royal Danish Academy of Sciences and Letters, 2010; Vol. 58. [Google Scholar]
- Grant, P.R.; Grant, B.R. Ecology and evolution of Darwin’s finches; Princeton University Press, 1986. [Google Scholar]
- Grant, P.R.; Grant, B.R. Unpredictable Evolution in a 30-Year Study of Darwin's Finches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Grant, P.R.; Grant, B.R. Evolution of Character Displacement in Darwin's Finches. Science 2006, 313, 224–226. [Google Scholar] [CrossRef]
- Grant, P.R.; Grant, B.R. The beak of the finch: A story of evolution in our time; Princeton University Press, 2008. [Google Scholar]
- Greenberg, R.; Cadena, V.; Danner, R.M.; Tattersall, G. Heat Loss May Explain Bill Size Differences between Birds Occupying Different Habitats. PLOS ONE 2012, 7, e40933. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Hendry, A.P.; Farrugia, T.J.; Kinnison, M.T. Human influences on rates of phenotypic change in wild animal populations. Mol. Ecol. 2008, 17, 20–29. [Google Scholar] [CrossRef]
- Herrel, A.; Podos, J.; Huber, S.K.; Hendry, A.P. Bite performance and morphology in a population of Darwin's finches: implications for the evolution of beak shape. Funct. Ecol. 2005, 19, 43–48. [Google Scholar] [CrossRef]
- Homberger, D.G. The case of the cockatoo bill, horse hoof, rhinoceros horn, whale baleen, and turkey beard: The integument as a model system to explore the concepts of homology and non-homology. In Vertebrate functional morphology: Horizon of research in the 21st century; Dutta, H. M., Munshi, J. S. Datta, Eds.; Oxford & IBH Publishing; Science Publishers, 2001; pp. 317–343. [Google Scholar]
- Homberger, D.G. The comparative biomechanics of a prey–predator relationship: The adaptive morphologies of the feeding apparatus of Australian black-cockatoos and their foods as a basis for the reconstruction of the evolutionary history of the Psittaciformes. In Vertebrate biomechanics and evolution; Bels, V. L., Gasc, J.-P., Casinos, A., Eds.; BIOS Scientific Publishers, 2003; pp. 203–228. [Google Scholar]
- Lamichhaney, S.; Berglund, J.; Almén, M.S.; Maqbool, K.; Grabherr, M.; Martinez-Barrio, A.; Promerová, M.; Rubin, C.-J.; Wang, C.; Zamani, N.; et al. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature 2015, 518, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Lamichhaney, S.; Han, F.; Webster, M.T.; Andersson, L.; Grant, B.R.; Grant, P.R. Rapid hybrid speciation in Darwin’s finches. Science 2018, 359, 224–228. [Google Scholar] [CrossRef]
- Lerner, H.R.; Meyer, M.; James, H.F.; Hofreiter, M.; Fleischer, R.C. Multilocus Resolution of Phylogeny and Timescale in the Extant Adaptive Radiation of Hawaiian Honeycreepers. Curr. Biol. 2011, 21, 1838–1844. [Google Scholar] [CrossRef]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef]
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef]
- MacArthur, R.H.; MacArthur, J.W. On Bird Species Diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Mallarino, R.; Campàs, O.; Fritz, J.A.; Burns, K.J.; Weeks, O.G.; Brenner, M.P.; Abzhanov, A. Closely related bird species demonstrate flexibility between beak morphology and underlying developmental programs. Proc. Natl. Acad. Sci. 2012, 109, 16222–16227. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.I.; Marques, D.A.; Mwaiko, S.; Wagner, C.E.; Excoffier, L.; Seehausen, O. Ancient hybridization fuels rapid cichlid fish adaptive radiations. Nat. Commun. 2017, 8, 14363. [Google Scholar] [CrossRef]
- Navalón, G.; Marugán-Lobón, J.; Bright, J.A.; Cooney, C.R.; Rayfield, E.J. The consequences of craniofacial integration for the adaptive radiations of Darwin’s finches and Hawaiian honeycreepers. Nat. Ecol. Evol. 2020, 4, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Newmark, W.D. Conserving Biodiversity in East African Forests; Springer Nature: Durham, NC, United States, 2002; ISBN 9783642076299. [Google Scholar]
- Norberg, U.M. Vertebrate flight: mechanics, physiology, morphology, ecology and evolution; Springer-Verlag: Berlin, Germany, 1990. [Google Scholar]
- Nosil, P. Ecological speciation; Oxford University Press, 2012. [Google Scholar]
- Parchman, T.L.; Benkman, C.W. Diversifying coevolution between crossbills and black spruce. Evolution 2002, 56, 1663–1678. [Google Scholar] [CrossRef]
- Podos, J. Correlated evolution of morphology and vocal signal structure in Darwin's finches. Nature 2001, 409, 185–188. [Google Scholar] [CrossRef]
- Podos, J.; Southall, J.A.; Rossi-Santos, M.R. Vocal mechanics in Darwin's finches: correlation of beak gape and song frequency. J. Exp. Biol. 2004, 207, 607–619. [Google Scholar] [CrossRef]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogues-Bravo, D.; Rasmussen, C.M.Ø.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Schluter, D. The ecology of adaptive radiation; Oxford University Press, 2000. [Google Scholar]
- Schluter, D. The ecology of adaptive radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Schneider, R.A. Cellular, Molecular, and Genetic Mechanisms of Avian Beak Development and Evolution. Annu. Rev. Genet. 2024, 58, 433–454. [Google Scholar] [CrossRef]
- Demissew, S.; Friis, I.; Weber, O. Diversity and endemism of the flora of Ethiopia and Eritrea: state of knowledge and future perspectives. Rendiconti Lince- Sci. Fis. e Nat. 2021, 32, 675–697. [Google Scholar] [CrossRef]
- Shochat, E.; Warren, P.; Faeth, S.; Mcintyre, N.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef]
- Tattersall, G.J.; Andrade, D.V.; Abe, A.S. Heat Exchange from the Toucan Bill Reveals a Controllable Vascular Thermal Radiator. Science 2009, 325, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, G.J.; Arnaout, B.; Symonds, M.R.E. The evolution of the avian bill as a thermoregulatory organ. Biol. Rev. 2017, 92, 1630–1656. [Google Scholar] [CrossRef]
- Tattersall, G.J.; Arnaout, B.; Symonds, M.R.E. The evolution of the avian bill as a thermoregulatory organ. Biol. Rev. 2016, 92, 1630–1656. [Google Scholar] [CrossRef]
- Van De Ven, T.M.F.N.; O Martin, R.; Vink, T.J.F.; E McKechnie, A.; Cunningham, S.J. Regulation of Heat Exchange across the Hornbill Beak: Functional Similarities with Toucans? PLOS ONE 2016, 11, e0154768. [Google Scholar] [CrossRef]
- Vans, K.L.; et al. The evolutionary ecology of dispersal and adaptation to urban environments. Functional Ecology 2011, 25, 405–416. [Google Scholar]
- Pol, J.L.V. A guide to endemic birds of Ethiopia and Eritrea. 2004. [Google Scholar] [CrossRef]
- Wake, D.B.; Larson, A. Multidimensional Analysis of an Evolving Lineage. Science 1987, 238, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.K.; Sherry, T.W. Behavioral feeding specialization in Pinaroloxias inornata, the “Darwin's Finch” of Cocos Island, Costa Rica. Proc. Natl. Acad. Sci. 1987, 84, 5506–5510. [Google Scholar] [CrossRef]
- Werner, T.K.; Sherry, T.W. Behavioral feeding specialization in some Tennessee warblers. Ecology 1987, 68, 189–196. [Google Scholar]
- Wiens, J.J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef]
- Yeh, P.J. Rapid evolution of a sexually selected trait following population establishment in a novel habitat. Evolution 2004, 58, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Zuccon, D.; Cibois, A.; Pasquet, E.; Ericson, P.G.P. Nuclear and mitochondrial phylogenetic analyses of the Fringillidae (Aves: Passeriformes). Molecular Phylogenetics and Evolution 2012, 62, 157–164. [Google Scholar]
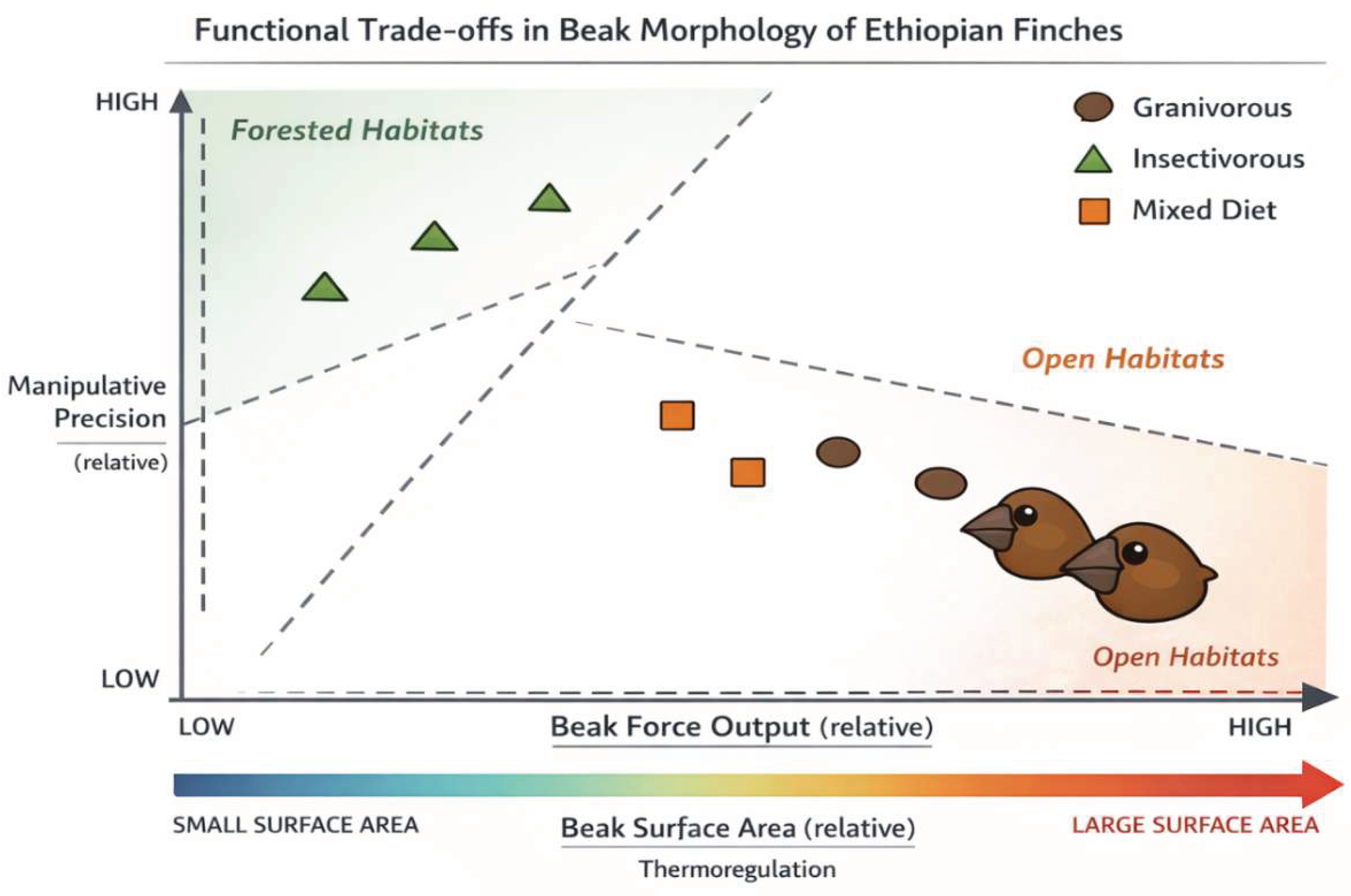
Figure 1.
Conceptual morphospace illustrating functional trade-offs in Ethiopian finch beak morphology between relative beak force output, manipulative precision, and thermoregulatory surface area. Symbols represent generalized dietary strategies rather than individual species, with shape indicating diet type and dashed regions denoting forested versus open habitats. Positions reflect relative functional tendencies synthesized from the literature rather than empirical measurements, highlighting how ecological context structures beak design trade-offs.
Figure 1.
Conceptual morphospace illustrating functional trade-offs in Ethiopian finch beak morphology between relative beak force output, manipulative precision, and thermoregulatory surface area. Symbols represent generalized dietary strategies rather than individual species, with shape indicating diet type and dashed regions denoting forested versus open habitats. Positions reflect relative functional tendencies synthesized from the literature rather than empirical measurements, highlighting how ecological context structures beak design trade-offs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



