
Submitted:
22 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
Cancer is one of the most significant global health problems and a leading cause of death worldwide. The origins of cancer are diverse and may stem from natural biological processes over time or directly result from anthropogenic activities. The complete elimination of all cancer-causing events within a living organism is highly unlikely. A more promising strategy would be to prevent tumors entirely by making the organism an unlivable environment where cancerous cells cannot survive. Surprisingly, this plausible alternative remains virtually unexplored. For the most part, it is not about the odds of cancer cells emerging in a hostile environment, rather than about their ability to adapt and persist within it. Plants have a broad-spectrum mechanism of defense against pathogens called non-host resistance (NHR), when an entire plant species is resistant to all isolates of a microbial species. While the NHR is effective against pathogens whereas cancer cells are perceived by an organism as “self” or “altered self”, they are still de facto “foreign intruders” since they generate neoantigens, novel proteins absent from normal tissues. Considering this, what are the theoretical possibilities of making a human organism an absolute non-host for cancer? As nearly all basic mechanisms and components of the NHR in plants have similarities to cancer responses in vertebrates, the task might be more feasible than it appears.
Keywords:
non-host resistance
; plant immunity
; cancer
; cancer immunity
; non-cancer environment
1. Introduction
Cancer is among the most important health problems and a leading cause of death worldwide. There is no universal cure against this complex disease as it is highly diverse and heterogenous. A typical process of tumorigenesis involves multiple steps, including disrupted cell cycle regulation, mutations in growth factors and signaling pathways, inactivation of tumor suppressor genes, activation of oncogenes, DNA repair deficiencies, mutations in DNA repair genes, evasion of apoptosis, resistance to immune surveillance, limitless replicative potential, and so forth (Hanahan and Weinberg, 2000: Hanahan, 2022).
Each of these and many other reasons for cancer onset are extremely complex, with thousands of possible ways to intervene without assuredly knowing if the intervention will be successful. Ideally, the root causes and processes would have to be eliminated as a whole because they are interconnected, often derive from and initiate one another.
Since cancer-related events occur naturally over time or arise consequentially from anthropogenic activities, the possibility of a simultaneous and total halt to their development appears highly unlikely. A more realistic alternative would be to prevent the product of their combined activities – tumor – from happening, by creating unlivable conditions for the mass, so that “it does not feel like growing” due to the hostile environment. This credible option is, surprisingly, still largely unexplored.
In nature, all species have their unique habitat, a specific ecological niche (Chase and Leibold, 2003) outside of which they cannot occur. Put simply, crocodiles will not survive in Arctic because they are ectothermic species. Cancer cells have their niche as well, a specialized microenvironments where they reside (Plaks et al. 2015). Although metastases are considered a spread of cancer cells away from their primary niche, similar favorable environments are selected and eventually colonized at the new locations (Oskarsson, 2014).
The assumption of no niche for cancer prompts more questions, the most important of them being how to create an all-embracing and hostile for the emergence and proliferation of cancer cells environment within an individual organism. So that if any of the cancer-causing events occur, they will have little or no chance to transform into a tumor. Back to the crocodile analogy, can the “Arctic” environment be created at the organismal level permitting normal cells to thrive and at the same time completely blocking development of the “crocodile” cells?
Plants have a broad-spectrum mechanism of defense against pathogens called non-host resistance (NHR), when an entire plant species is resistant to all isolates of a microbial species (Nurnberger and Lipka, 2005). For example, immunity of rice against all rust and powdery mildew fungi (Wu et al. 2023). Or general resistance of monocotyledonous plants against broadly successful in dicots plant pathogen Agrobacterium tumefaciens, a cause of the neoplastic disease crown gall (De Cleene and De Ley, 1976). While NHR is effective against pathogens and cancer cells are perceived by an organism as “self” or “altered self”, tumors are still de facto “foreign intruders” since they generate neoantigens, novel proteins absent from normal tissues. Considering this, would that be theoretically possible to make an animal organism an absolute non-host for cancer? As nearly all basic mechanisms and components of the NHR in plants have similarities to cancer responses in animals, this challenging task may be more achievable than it first appears.
2. Main Text
The evolution operates at all levels, from molecular to organismal, obeying the same law of natural selection. In nature, species that confront an unfavorable environment can adapt, enter dormancy, migrate, or face extinction. The ability of cancer cells to undergo the first three states and also to overcome harsh treatment conditions is well known (Park and Nam, 2020; Madukwe, 2023). But while cancer cells can survive hostile surroundings, will they, given a choice, likely emerge de novo in adverse or in the favorable environment? Surprisingly, the literature on this simple query is not as extensive as it would imply, if any, and mostly relates to the ability of cancer cells to adapt and resist different stress conditions (Chen and Xie, 2018).
Furthermore, would cancer cells emerge at all if the entire organismal environment is hostile to their appearance and proliferation? Doubling back on the crocodile example, could this very species hypothetically originate in the Arctic? No, it could not, because its body temperature and heat sources depend on its habitat. A similar answer is expected to apply to cancer cells: while they can post-factum resist the adverse environment, they are unlikely to arise in it. Reiterating the point, it is about the odds of cancer cells emerging in an extremely hostile environment rather than about their ability to adapt and persist within it.
There are only a few comprehensive, all-organismic conditions that can potentially satisfy a requirement of being favorable for normal but hostile for cancer cells. Among them are oxygen levels, aerobic in normal cells and hypoxic in solid tumor environments (Emami et al. 2021) and nutrient dependencies, particularly glucose availability: while normal cells require a balanced supply of nutrients, cancer competes for nutrients like glucose and glutamine, relying on glycolysis and much higher glycose uptake (Pavlova and Thompson, 2017). None of these, however, can prevent cancer cells from appearing: they can emerge and grow in well-oxygenated and survive in limited glycose environment (Chen et al. 2023; Nwosu et al. 2023). Other known candidates for a stable “Arctic” background on the organismal level are many but more specific, targeting individual cancer-associated processes rather than being broadly detrimental to all cancer cells.
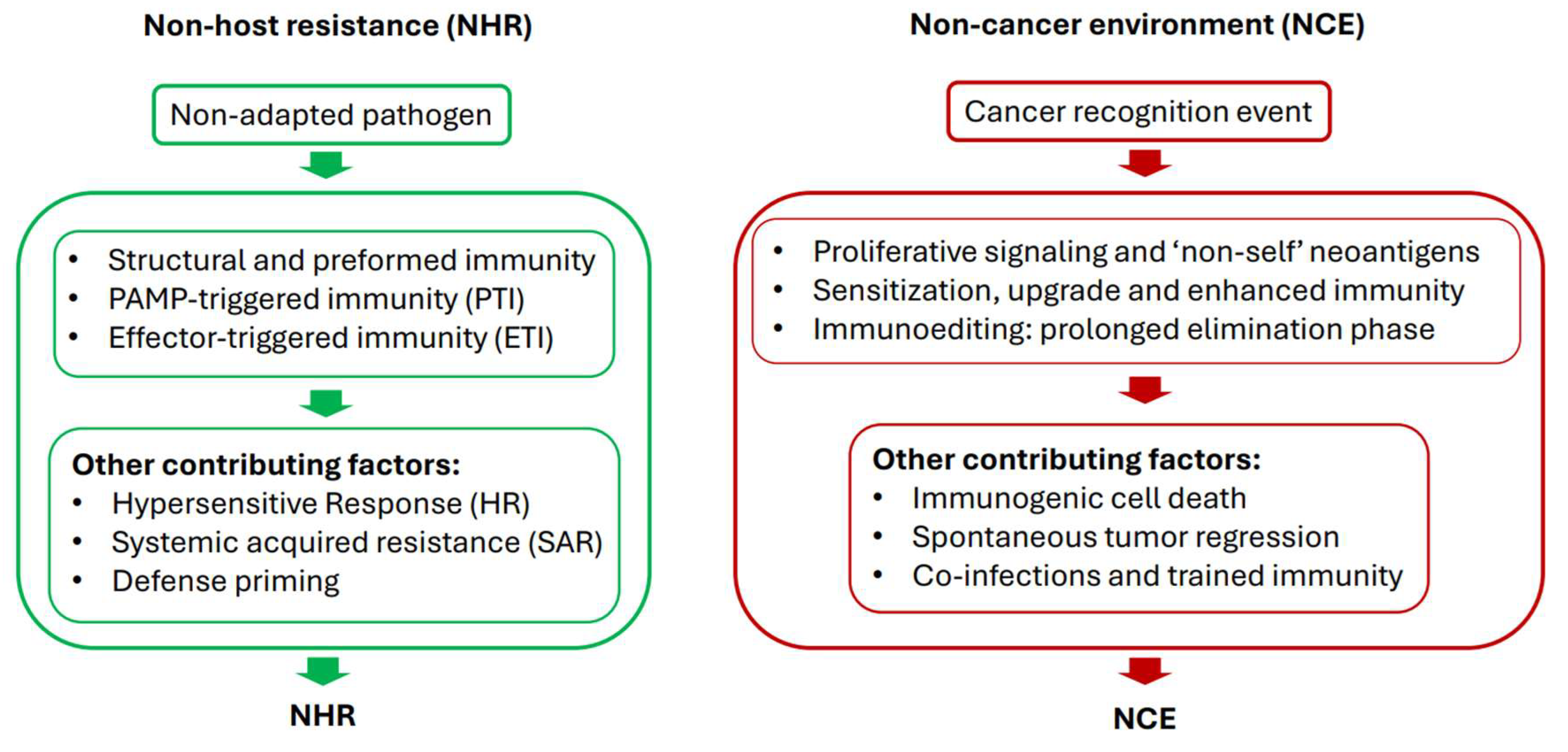
The NHR is a remarkably durable and long-lasting immunity and the sole reason why most plants in the natural environment are healthy. It is triggered by a combination of factors, including molecular motifs conserved within a microbial species (pathogen-associated molecular patterns or PAMPs), pathogens’ virulence factors (effectors), and subsequent downstream signaling (Nurnberger and Lipka, 2005). When enabled, NHR can also spark hypersensitive response (HR) – a rapid, localized, and programmed cell death at the point of infection (Balint-Kurti, 2019).
In addition, plants have constitutive immunity as a part of the NHR: preformed mechanical barriers (leaf topography, waxes, cell wall, volatiles, etc.); various antimicrobial compounds (AMPs) (terpenes, flavonoids, tannins, phytoalexins, phytoanticipins, saponins, etc); and defense proteins, participating in both constitutive and induced immunity (Panstruga and Moscou, 2020). Altogether, these processes ensure the inability of a particular pathogen to infect a given host species.
In vertebrates, cancer cells trigger the immune system by analogous mechanisms, such as damage-associated molecular patterns (DAMPs), (Hernandez et al. 2016); molecules that may act as “effectors” by suppressing immune cells; metabolic enzymes; and various enzymes eliciting host immune responses (Yu et al. 2018). They also induce cytotoxicity of the natural killer (NK) cells and cytotoxic T lymphocytes (CTL) prompting immunogenic cell death (ICD), (Troitskaya et al. 2022) – a concept related to the HR in plants. Like plants, animals produce a wide variety of AMPs and some of them were shown to have anticancer activity (Qu et al. 2024). Then why all these and other processes associated with multilayered cancer defense, are not capable of creating an environment similar to the non-host resistance in plants?
For that, cancer cells would have to be recognized by an organism as strictly external, “non-self” entities rather than “self” or “altered self” as they are normally perceived, which causes the immune system to stumble. Although cancer cells can still be called “foreign intruders” since they generate neoantigens (Efremova et al, 2017), their identity for the host is not straightforward: they are like a rental apartment a neighbor erected in your own backyard using your building materials.
Besides, unlike plant pathogens which need to penetrate preformed and mechanical barriers, these intruders are of domestic origin and thus often do not have to cope with the first line of organismal defenses but only with the pre-formed genetic defenses and innate immune system.
A sequence of the NHR in plants involves pre-invasive and post-invasive defense responses (Panstruga and Moscou, 2020). Pre-invasive responses against cancer in the same manner as they present in plants (physical barriers preventing pathogens from entering the plant) do not occur in animals. Instead, they are a part of the comprehensive (i.e. post-invasive) organismal defenses intended to suppress malignancy at the cellular (e.g. DNA repair, apoptosis, cell-cycle checkpoint, etc), and systemic levels (e.g. immune surveillance). Since this strategy against post-invasive cancer, although robust, ultimately fails, can all the sophisticated organismic responses to cancer assault be rechanneled for it prevention, thus generating a total non-cancer environment (NCE), an animal analog of the NHR?
Would dramatically enhanced sensitivity of all existent anti-cancer processes do the trick? In naked mole-rat (Heterocephalus glaber), cells stop dividing due to the hypersensitivity to contact inhibition (Seluanov et al, 2009), resulting in a longer life span and suppression of cancer (Caulin and Maley, 2011). The early contact inhibition is controlled by cyclin-dependent kinase inhibitor p16 in contrast to the regular contact inhibition that is triggered by accumulation of p27, another cyclin-dependent kinase inhibitor (Seluanov et al. 2009). Contact inhibition of proliferation is one of the mechanisms of cellular defenses against cancer (Ribatti, 2017).
Therapeutic upregulation of another inherent anti-cancer pathway, DNA repair, is also feasible by enhancing activity of DNA repair enzymes (Song et al. 2024). Modulation of apoptosis, an evolutionary conserved mechanism of cell death (Singh et al. 2017), toward high apoptotic sensitivity is possible as well. Highly sensitive pro-apoptotic signaling, or priming for apoptosis, is known to occur in developing brain and is responsible for the hypersensitivity of the brain tissue to damage or stress (Singh et al. 2017). Further, enhancing functional activity of tumor-suppressor protein p53, the “guardian of the genome” (Lane and Crowford, 1979; Lane, 1992; Linzer and Levine, 1979), through regulation of its inhibitors MDM2 (murine double minute 2 gene) and MDM4 have been reported (Toledo and Wahl, 2007). Therapeutically increasing the sensitivity of the innate immune system against cancer, for example, by means of enhancing cytotoxicity of the NK is a promising strategy in cancer immunotherapy (Stenger and Miller, 2024). So is increasing the sensitivity of the adaptive immune system against cancer by multiple approaches, including adoptive cell therapy (Ruan and Wang, 2025).
Thus, considering that many if not all non-immune and immune defenses against cancer can be reinforced and elevated to a higher level, an establishment of the NCE in animals equivalent or comparable to the NHR in plants deems possible. One way to accomplish that would be a combined rather than individual, intensification and continuous preservation of all these processes at the elevated level by virtue of some broad-based elicitor(s) capable of acting on main components of the anti-cancer defenses. Importantly, this would not be a therapeutic treatment of the existing malignancy, but instead an initiation of the more preemptive and persistent defensive state of an organism as compared to its ordinary safeguarded status.
This “upgraded” biological condition could be reminiscent of a highly adaptive, “fight or flight” response intended for an immediate survival under acute stress (Dhabhar, 2019). However, unlike the short-term “fight or flight” response, it would have to be a more comprehensive and lasting state constantly supported by all aspects of the innate and adaptive immunity. Perhaps cognitive, physiological, hormonal, metabolic, and immune mechanisms involved in the sudden stress response along with specific modulations of the anti-cancer processes mentioned above can offer an answer on what integrative catalyst or substance could possibly be utilized to achieve this ameliorated organismal condition of the absolute NCE.
An alternative or rather broadened way of carrying out this task could be to incorporate approaches used for artificial induction or engineering the NHR in plants (Figure 1). The NHR is believed to be a multi-gene trait, and different methodologies currently exist to elucidate its genetic basis and components, including chemical genetics, mutagenesis-aided forward genetics, reverse genetics, exploring near-host variations, and other strategies (Wu et al. 2023). Importantly, the NHR is transferable from resistant to susceptible plants by transformation methods, genome editing or crossbreeding (Gill et al. 2015). For example, the expression of non-host genes that are targets of pathogens effectors enhanced disease resistance in host plants: expression of AtPUB33, an E3 ubiquitin ligase and a non-host Arabidopsis ortholog of the Phytophthora infestans effector target in potato called StUBK, increased resistance to P. infestans in two different host species, potato and Nicotiana benthamiana (McLellan et al. 2022). In addition to genetic engineering, various chemical elicitors and biological priming can be implemented to trigger defense responses in both non-host and host plants (Karasov et al. 2017; Malik et al. 2020; Naz et al. 2024).
Since many genes and processes involved in the cellular and immune responses to oncogenesis are increasingly known, a strategy resembling that used for the induction of the NHR in plants can potentially be applied to generate a durable NCE state in animals. Whereas NHR exists as a multi-layered form of plant immunity that enacts as a whole almost immediately upon detecting a threat, its artificial induction is likely to be a sequential process in which a single step, for example transfer of the NHR genes from resistant to susceptible plants, could be sufficient to render immunity to a particular pathogen.
The NCE, on the other hand, being not there yet per se, is represented by a complex, interconnected but highly vulnerable protective system. It would have to be redesigned from the available organismal resources as a single and ready-to-act operational unit similar to the NHR and possessing a crucial ability to perceive malignancies as “foreign” to the immune system by recognizing cancer-related neoantigens and stress signals (Efremova et al 2017; Xie et al. 2023). This restructuring of the standard defense mechanisms to generate NHR-like comprehensive prevention system will likely require two major steps: i. instant signaling prompted by cancer-recognition events followed by ii. sensitization and upgrade of the existing anti-tumor network.
In line with the NHR, in which the major recognition event is PAMP-triggered immunity (PTI) activated by microbial molecular signatures, cancer-recognition signals initiating quick NCE response must be associated with different biomarkers indicating that transformation of normal cells into malignant is underway (Sanchez-Herrero et al. 2022; Passaro et al. 2024). It is essential that these cancer fingerprints are of the “non-self” origin and recognized by the immune system as antigens thus stimulating it for the attack (Fang et al. 2022). Yet, this recognition alone frequently fails because cancers routinely escape immune destruction (Messerschmidt et al. 2016). The escape process, called “immunoediting” (Dunn et al. 2002) strongly reminds the zigzag model illustrating plant response to infection (Jones and Dangl, 2006).
It consists of three phases, each presenting a distinct barrier subsequently overpowered by cancer cells evading the organism in the end: elimination, equilibrium and escape (Dunn et al 2002; Messerschmidt et al. 2016). While eliminating tumors along the way, the immunoediting also plays a dual role of creating cancer phenotypes that escape immunosurveillance (Dunn et al. 2002). Likewise, the “zigzag” arrangement of the plant immune system can completely stop pathogen’s colonization (PTI), result in effector-triggered susceptibility (ETS), or in effector-triggered immunity (ETI). The latter can drive pathogens to avoid elimination by gaining new effectors through natural selection (ETS), (Jones and Dangle, 2006). This phase can circle back to ETI again due to plant regrouping its alleles of resistance (R) genes – a direct result of the continuous co-evolution of host R-genes and pathogen effectors (Jones and Dangle, 2006).
In contrast, after the third phase of immunoediting (immune escape), the immune system is considered defeated and can no longer block tumors (Dunn et al. 2002; Messerchmidt et al. 2016). Extending the first phase of immunoediting (elimination of the newly formed tumor) by enhancing activity of immune cells, improved neoantigen presentation, vaccination, oncolytic viruses, etc. - would be a major breakthrough en route to the formation of NCE.
Animals and humans do not have R-genes on a par with those in plants; the closest parallels are pattern recognition receptors (PRRs) of the innate immune response, for example intracellular NOD-like receptors (NLRs) sensing cancer-derived DAMPs; and transmembrane T-cell Receptors (TCRs) of the adaptive immune system recognizing cancer-related peptides. The TCRs can recognize tumor-related neoantigens, which will lead to a clonal proliferation of the specific T cell clones and tumor killing (Sim and Sun, 2022), similarly to a formation of new alleles of resistance (R) genes effective against modified pathogen’s effectors. Complete tumor elimination, however, is limited due to the heterogeneity of neoantigens within a single tumor. Consequently, a reversal of immune escape, the same as in the zigzag pattern, does not normally happen and has to be induced therapeutically (Liu et al. 2023).
In rare cases, however, spontaneous tumor regression (SR) occurs naturally (Cui et al. 2003). Although SR mechanisms are currently elusive, they bear a resemblance to NHR and mirror the proposed state of NCE by robust activation of all anti-tumor resources.
One of the reported features associated with the SR is coinfection with pathogenic microorganisms promoting a defensive mode of the immune system by stimulation of antigen-presenting cells (APC) (Kaufman et al. 2015; Kucerova and Cervinkova, 2016). The involvement of pathogens in SR response is another hallmark linking it to the NHR and potentially to the establishment of NCE, as infection could be that very signal triggering cancer-recognition events by presentation of cancer biomarkers via APC. Interestingly, although the evidence is almost nonexistent, tumor reversion can also occur in plants infected by a pathogenic bacterium A. tumefaciens. It was demonstrated in the late 1950s that crown-gall tumor cells could be transformed into benign by series of graftings to healthy plants (Braun, 1959).
A second major step to rearrange standard defense mechanisms against cancer presumably requires sensitization and upgrade of the existing anti-tumor network. As evident from the research on contact inhibition, therapeutic upregulation, priming, and enhancement of sensitivity of cellular and immune responses (Toledo and Wahl, 2007; Seluanov et al, 2009; Singh et al. 2017; Liu et al. 2023; Song et al. 2024; Stenger and Miller, 2024; Ruan and Wang, 2025), as well as from the studies on SR (Kucerova and Cervinkova, 2016; Radha and Lopus, 2021), this task is feasible. Moreover, as all organismal resources and arrays of defense mechanisms against cancer are readily available and highly modifiable, their recurrent, preventive, and stress-induced upgrade would deem natural. Just like an upgrade of an existing version of a computer program by running scripts and commands to change its configuration.
As noted earlier, one example of the command to step up and maximize could come from cancer-associated, presumably non-carcinogenic, microbial infections by way of DAMPs and activated APCs. Antitumor efficacy can also be augmented by beneficial microbes: a recent study showed that a new strain of the bacterial genus Hominenteromicrobium, a probiotic member of the human gut microbiome, activated tumor-specific CD8+ T cells, enhancing antitumor responses in mice (Lin et al 2025). Hormonal activation, signals inducting apoptosis, ischemia, or epigenetic changes can also contribute to the upgrade of anti-tumor network (Greger et al. 1989). The more precise mechanisms would likely stem from deciphering the SR response (Ricci and Cerchiari, 2010).
After signals to launch NCE-like response are initiated and the status is implemented through optimization of anti-cancer defenses, further actions will be necessary to make the process sustainable and, more importantly, reproducible. So that any uncontrollably proliferating cells will be treated by an organism as non-adapted invaders and the ability to enhance the body’s anti-cancer defenses will be retained in the future. Plant immune system can be primed to generate faster, stronger, and long-lasting defense response upon pathogen’s attack (Pastor et al. 2013). Defense priming in plants can be triggered not only by pathogens, but also by beneficial microorganisms, alterations in primary metabolism, synthetic and chemical compounds (Conrath et al. 2006; Pastor et al. 2013). Formerly called sensitization (Conrath, 2011), defense priming involves amplification of extracellular stimuli by mitogen-activated protein kinases (MPKs); chromatin modifications; systemic acquired resistance (SAR); molecular modifications of primary metabolism; rapid accumulation of reactive oxygen species (ROS); and other signaling processes and pathways (Pastor et al. 2013; Conrath, 2011, Conrath et al, 2006). Defense priming, as a form of induced resistance, can also contribute to the NHR.
Against this plant background, a fairly new concept of trained immunity, an enhanced immunity in vertebrates that follows initial exposure to a pathogen (Netea et al. 2011; Li et al. 2025), may be potentially viewed as a process able to provide consistency to the NCE. The idea of trained immunity in vertebrates initially came from similarities with long known SAR phenomenon in plants, when plants are broadly and systemically protected for extended periods of time after initial local infection (Netea et al. 2011). One of the key characteristics of trained immunity is a broad and rapid response to PAMPs and DAMPs leading to reprogramming of different cell types of the innate immune system and heightened state of immune cell effectors (Li et al. 2025). Same as in plants, trained immunity in cancer can be artificially induced by therapeutic agents like β-glucan or Bacillus Calmette-Guerin (BCG) vaccination, or happen naturally after infections, for example, with influenza A virus or hepatitis B virus (Li et al. 2025).
Summarizing, potential steps or conditions imperative for unveiling the NCE state include initial proliferating signaling followed by instant sensitization, upgrade and preservation of the continuing elevated status of all available anti-cancer resources. In a general way, those are the same requirements needed for the establishment of NHR in plants (Figure 2). Since the NHR trait is inherited, it may well be that NCE state is intrinsic likewise and only needs to be “uncaged” from idleness by yet unidentified but readily available factor(s).
The NHR in plants has been known for many years and experimentally tested (Nürnberger and Lipka, 2025); each of the potential components and manifestations that may prompt and accompany the NCE state (sensitization, priming, vaccines, ICD, SR, elimination phase of immunoediting, etc.) has been studied and empirically verified as well. It is therefore possible, based on the available data, to develop exploratory in vitro and in vivo models simulating NCE conditions and their effect on the process of tumorigenesis.
3. Conclusions
While individual enigmas pertaining hypothetical establishment of the NCE state and even the order of the proposed events could be learned from the NHR and mechanisms of plant immunity in general, linking them together into the closely tied, vigilant, and sensitized protective network is a challenging task. Nevertheless, both the concept and the obvious possibility of the natural occurrence of the NCE appear strongly plausible, especially keeping in mind the resilience and efficiency of the comparable NHR process in plants. Moreover, within the current accumulated knowledge of the processes related to the NHR and to the body's natural and induced defenses against cancer, this hypothesis appears to be experimentally testable. The testing could be approached in multiple ways including genetically engineering animals with traits that are thought to confer cancer resistance or enhancing their immune response by a combination of immunotherapies and trained immunity (Figure 1).
Author Contributions
The author confirms sole responsibility for the conception, design, analysis, interpretation, drafting, and final approval of the manuscript. The author consents to the publication of the manuscript.
Funding
This study was supported by the United States Department of Agriculture, the Agricultural Research Service, CRIS numbers 8042-21500-003-000D.
Informed Consent Statement
No human participants were used in this study.
Data Availability Statement
No datasets were generated or analyzed during the current study.
Declaration on the Use of Artificial Intelligence
Generative AI has not been used for writing this manuscript. During the preparation of this work the author used Google Gemini and Google search tools to search and review available literature on the subject, to analyze data as part of the research process, and to improve language and readability. After using the tools, the author thoroughly reviewed and evaluated the content as needed and takes full responsibility for the content of the publication.
Conflicts of Interest
The author declares that he has no competing interests.
Abbreviations
| AMPs | antimicrobial compounds |
| APC | antigen-presenting cells |
| CTL | cytotoxic T lymphocytes |
| DAMPs | damage-associated molecular patterns |
| ETI | effector-triggered immunity |
| ETS | effector-triggered susceptibility |
| HR | hypersensitive response |
| ICD | immunogenic cell death |
| NCE | non-cancer environment |
| NHR | non-host resistance |
| NK | natural killer cells |
| PAMPs | pathogen-associated molecular patterns |
| PTI | PAMP-triggered immunity |
| R genes | resistance genes |
| SAR | systemic acquired resistance |
| SR | spontaneous tumor regression |
| TAA | tumor-associated antigens |
| TCR | T-cell Receptors |
References
- Balint-Kurti, P. The plant hypersensitive response: concepts, control and consequences. Molecular Plant Pathology 2019, 20, 1163–1178. [Google Scholar] [CrossRef]
- Braun, A.C. A Demonstration of the recovery of the crown-gall tumor cell with the use of complex tumors of single-cell origin. Proc Natl Acad Sci USA 1959, 45, 932-938. [Google Scholar] [CrossRef]
- Caulin, A.F.; Maley, C.C. Peto’s Paradox: Evolution’s Prescription for Cancer Prevention. Trends Ecol Evol. 2011, 26, 175–182. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Ecological Niches Linking Classical and Contemporary Approaches; University of Chicago Press, 2003; p. 4. [Google Scholar]
- Chen, M.; Xie, S. Therapeutic targeting of cellular stress responses in cancer. Thorac Cancer 2018, 9, 1575–1582. [Google Scholar] [CrossRef]
- Chen, Z.; Han, F.; Du, Y.; Shi, H.; Zhou, W. Hypoxic microenvironment in cancer: molecular mechanisms and therapeutic interventions. Signal Transduction and Targeted Therapy 2023, 8, 70. [Google Scholar] [CrossRef]
- Conrath U, Beckers GJM, Flors V, Garcia-Agustin P, Jakab G, Mauch F, Newman MA, Pieterse CM, Poinssot B, Pozo MJ, et al. (2006). Priming: getting ready for battle. Mol Plant Microbe Interact 19 1062–1071 Conrath, U., 2011. Molecular aspects of defence priming. Trends Plant Sci. 16,524–531.
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 8 16, 524–31. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z; Willingham, MC; Hicks, AM; Alexander-Miller, MA; Howard, TD; Hawkins, GA; Miller, MS; Weir, HM; Du, W; DeLong, CJ.; DeLong. Spontaneous regression of advanced cancer: Identification of a unique genetically determined, age-dependent trait in mice. PNAS 2003, 100, 6682–6687. [Google Scholar] [CrossRef] [PubMed]
- De Cleene, M.; De Ley. J. Bot. Rev. 1976, 10 42, 389–466. [CrossRef]
- Dhabhar, F. The Short-Term Stress Response – Mother Nature’s Mechanism for Enhancing Protection and Performance Under Conditions of Threat, Challenge, and Opportunity. Front Neuroendocrinol. 2018, 49, 175–192. [Google Scholar] [CrossRef]
- Dunn, GP; Bruce, AT; Ikeda, H; et al. Cancer immunoediting: From immunosurveillance to tumor escape. Nat Immunol. 2002, 12 3, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Finotello, F.; Rieder, D.; Trajanoski, Z. Neoantigens Generated by Individual Mutations and Their Role in Cancer Immunity and Immunotherapy. Front Immunol. 2017, 28;8, 1679. [Google Scholar] [CrossRef]
- Emami Nejad, A.; Najafgholian, S.; Rostami, A.; Sistani, A.; Shojaeifar, S.; Esparvarinha, M.; Nedaeinia, R.; Haghjooy Javanmard, S.; Taherian, M.; Ahmadlou, M.; Salehi, R.; Sadeghi, B.; Manian, M. The role of hypoxia in the tumor microenvironment and development of cancer stem cell: a novel approach to developing treatment. Cancer Cell International 2021, 14 21(1), 62. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Liu, Z; Gao, M.; Tu, K.; Xu, Q.; Zhang, Y. Targeting Nutrient Dependency in Cancer Treatment. Front Oncol. 2022, 15 12, 820173. [Google Scholar] [CrossRef]
- Fang, X.; Guo, Z.; Liang, J.; Wen, J.; Liu, Y.; Guan, X.; Li, H. Neoantigens and their potential applications in tumor immunotherapy (Review). Oncol Lett. 2022, 16 23(3), 88. [Google Scholar] [CrossRef]
- Gill, U.S.; Lee, S.; Mysore, K.S. Host versus nonhost resistance: distinct wars with similar arsenals. Phytopathology 2015, 17 105(5), 580–7. [Google Scholar] [CrossRef]
- Greger, V.; Passarge, E.; Höpping, W.; Messmer, E.; Horsthemke, B. Epigenetic changes may contribute to the formation and spontaneous regression of retinoblastoma. Hum Genet. 1989, 18 83, 155–8. [Google Scholar] [CrossRef]
- 19. Hanahan D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hernandez, C.; Huebener, P.; Schwabe, R.F. Damage-associated molecular patterns in cancer: a double-edged sword. Oncogene 2016, 21 35, 5931–5941. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 22 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Karasov, T.L.; Chae, E.; Herman, J.J.; Bergelson, J. Mechanisms to Mitigate the Trade-Off between Growth and Defense. Plant Cell. 2017, 29, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic viruses: a new class of immunotherapy drugs. Nat Rev Drug Discov. 2015, 14, 642–662. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, P.; Cervinkova, M. Spontaneous regression of tumour and the role of microbial infection – possibilities for cancer treatment. Anticancer Drugs 2016, 27, 269–277. [Google Scholar] [CrossRef]
- Lane, D.P.; Crawford, L.V. T antigen is bound to a host protein in SV40-transformed cells. Nature 1979, 26 278, 261–3. [Google Scholar] [CrossRef]
- Lane, D.P. Cancer. p53, guardian of the genome. Nature 1992, 358, 15–6. [Google Scholar] [CrossRef]
- Li, S.; Zou, Y.; McMasters, A.; Chen, F.; Yan, J. Trained immunity: A new player in cancer immunotherapy. eLife 2025, 28 14, e104920. [Google Scholar] [CrossRef]
- Lin, N.YT.; Fukuoka, S.; Koyama, S. Microbiota-driven antitumor immunity mediated by dendritic cell migration. Nature 2025, 644, 1058–1068. [Google Scholar] [CrossRef]
- Linzer, D.I.; Levine, A.J. Characterization of a 54K dalton cellular SV40 tumor antigen present in SV40-transformed cells and uninfected embryonal carcinoma cells. Cell. 1979, 30 17, 43–52. [Google Scholar] [CrossRef]
- Liu, S.; Sun, Q.; Ren, X. Novel strategies for cancer immunotherapy: counter-immunoediting therapy. J Hematol Oncol. 2023, 16, 38. [Google Scholar] [CrossRef]
- Madukwe, J.C. Overcoming drug resistance in cancer. Cell 2023, 32 186, 1515–1516. [Google Scholar] [CrossRef]
- Malik, N.AA.; Kumar, I.S.; Kalaivani Nadarajah, K. Elicitor and Receptor Molecules: Orchestrators of Plant Defense and Immunity. Int J Mol Sci. 2020, 21(3), 963. [Google Scholar] [CrossRef]
- McLellan, H.; Harvey, S.E.; Steinbrenner, J.; Armstrong, M.R.; He, Q.; Clewes, R.; Pritchard, L.; Wang, W.; Wang, S.; Nussbaumer, T.; et al. Exploiting breakdown in nonhost effector–target interactions to boost host disease resistance. Proc. Natl. Acad. Sci. USA 2022, 119(35), e2114064119. [Google Scholar] [CrossRef] [PubMed]
- Messerschmidt, J.L.; Prendergast, G.C.; Messerschmidt, G.L. How Cancers Escape Immune Destruction and Mechanisms of Action for the New Significantly Active Immune Therapies: Helping Nonimmunologists Decipher Recent Advances. The Oncologist 2016, 21, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Naz, M; Zhang, D; Liao, K; Chen, X; Ahmed, N; Wang, D; Zhou, J; Chen, Z. The Past, Present, and Future of Plant Activators Targeting the Salicylic Acid Signaling Pathway. Genes (Basel) 2024, 15(9), 1237. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Quintin, J.; van der Meer, J.W.M. Trained Immunity: A Memory for Innate Host Defense. Cell Host & Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef]
- Nürnberger, T.; Lipka, V. Non-host resistance in plants: new insights into an old phenomenon. Mol Plant Pathol. 2005, 6, 335–45. [Google Scholar] [CrossRef]
- Nwosu, ZC; Ward, MH; Sajjakulnukit, P; Poudel, P; Ragulan, C; Kasperek, S; Radyk, M; Sutton, D; Menjivar, RE; Andren, A; Apiz-Saab, JJ; Tolstyka, Z; Brown, K; Lee, HJ; Dzierozynski, LN; He, X; Ps, H; Ugras, J; Nyamundanda, G; Zhang, L; Halbrook, CJ; Carpenter, ES; Shi, J; Shriver, LP; Patti, GJ; Muir, A; Pasca di Magliano, M; Sadanandam, A; Lyssiotis, CA. Uridine-derived ribose fuels glucose-restricted pancreatic cancer. Nature 2023, 618, 151–158. [Google Scholar] [CrossRef]
- Oskarsson, T.; Batlle, E.; Massagué, J. Metastatic Stem Cells: Sources, Niches, and Vital Pathways. Cell Stem Cell. 2014, 14, 306–321. [Google Scholar] [CrossRef]
- Panstruga, R.; Moscou, M. J. What is the molecular basis of nonhost resistance? Mol Plant Microbe Interact. 2020, 33, 1253–1264. [Google Scholar] [CrossRef]
- Park, S-Y; Nam, J-S. The force awakens: metastatic dormant cancer cells. Experimental & Molecular Medicine 2020, 42 52, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Passaro, A.; Al Bakir, M.; Hamilton, E.G.; Diehn, M.; André, F.; Roy-Chowdhuri, S.; Mountzios, G.; Wistuba, I.I.; Swanton, C.; Peters, S. Cancer Biomarkers - Emerging Trends and Clinical Implications for personalized treatment. Cell. 2024, 187(7), 1617–1635. [Google Scholar] [CrossRef] [PubMed]
- Pastor, V.; Luna, E.; Mauch-Mani, B.; Ton, J.; Flors, V. Primed plants do not forget. Environmental and Experimental Botany 2013, 94, 46–56. [Google Scholar] [CrossRef]
- Pavlova, N.; Thompson, C. B. The emerging hallmarks of cancer metabolism. Cell Metab. 2016, 45 23, 27–47. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The Cancer Stem Cell Niche: How Essential is the Niche in Regulating Stemness of Tumor Cells? Cell Stem Cell. 2015, 16, 225–238. [Google Scholar] [CrossRef]
- Qu, B.; Yuan, J.; Liu, X.; Zhang, S.; Ma, X.; Lu, L. Anticancer activities of natural antimicrobial peptides from animals. Front Microbiol. 2024, 14, 1321386. [Google Scholar] [CrossRef]
- Radha, G.; Lopus, M. The spontaneous remission of cancer: current insights and therapeutic significance. Translational Oncology 2021, 48 14, 101166. [Google Scholar] [CrossRef]
- Ribatti, D. A revisited concept: Contact inhibition of growth. From cell biology to malignancy. Exp Cell Res. 2017, 359(1), 17–19. [Google Scholar] [CrossRef]
- Ricci, S.B.; Cerchiari, U. Spontaneous regression of malignant tumors: Importance of the immune system and other factors (Review). Oncol Lett. 2010, 1, 941–945. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, L. Adoptive cell therapy against tumor immune evasion: mechanisms, innovations, and future directions. Front Oncol. 2025, 15, 1530541. [Google Scholar] [CrossRef]
- Sánchez-Herrero, E.; Serna-Blasco, R.; Robado de Lope, L.; González-Rumayor, V.; Romero, A.; Provencio, M. Circulating Tumor DNA as a Cancer Biomarker: An Overview of Biological Features and Factors That may Impact on ctDNA Analysis. Front Oncol. 2022, 12, 943253. [Google Scholar] [CrossRef]
- Seluanov, A.; Hine, C.; Azpurua, J.; Feigenson, M.; Bozella, M.; Mao, Z.; Catania, K.C.; Gorbunova, V. Hypersensitivity to contact inhibition provides a clue to cancer resistance of naked mole-rat. Proceedings of the National Academy of Sciences of the United States of America 2009, 106, 19352–19357. [Google Scholar] [CrossRef] [PubMed]
- Sim, M. J.W.; Sun, P.D. T Cell recognition of tumor neoantigens and insights into T cell immunotherapy. Frontiers in Immunology 2022, 13, 833017. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol. 2019, 20(3), 175–193. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Park, C.; Cabanting, F.E.B.; Jun, Y.W. Therapeutic upregulation of DNA repair pathways: strategies and small molecule activators. RSC Med Chem. 2024, 56 15, 3970–3977. [Google Scholar] [CrossRef]
- Stenger, T.D.; Miller, J.S. Therapeutic approaches to enhance natural killer cell cytotoxicity. Front. Immunol. 2024, 15, 1356666. [Google Scholar] [CrossRef]
- Toledo, F.; Wahl, G.M. MDM2 and MDM4: p53 regulators as targets in anticancer therapy. Int J Biochem Cell Biol. 2007, 39, 1476–1482. [Google Scholar] [CrossRef]
- Troitskaya, S.; Novak, D.D.; Richter, V.A.; Koval, O.A. Immunogenic Cell Death in Cancer Therapy. Acta Naturae 2022, 14(1), 40–53. [Google Scholar] [CrossRef]
- Wu, Y.; Sexton, W.; Yang, B.; Xiao, S. Genetic approaches to dissect plant nonhost resistance mechanisms. Mol Plant Pathol. 2023, 24(3), 272–283. [Google Scholar] [CrossRef]
- Xie, N.; Shen, G.; Gao, W.; Huang, Z.; Huang, C. Neoantigens: promising targets for cancer therapy. Sig Transduct Target Ther 2023, 8, 9. [Google Scholar] [CrossRef]
- Yu, C-P.; Fu, S-F.; Chen, X.; Ye, J.; Ye, Y.; Kong, L-D.; Zhu, Z. The Clinicopathological and Prognostic Significance of IDO1 Expression in Human Solid Tumors: Evidence from a Systematic Review and Meta-Analysis. Cellular Physiology and Biochemistry 2018, 49, 134–143. [Google Scholar] [CrossRef]
Figure 1.
Initiation of the non-cancer environment (NCE) as an analog to the non-host resistance (NHR) in plants. TSGs, tumor suppressor genes.
Figure 1.
Initiation of the non-cancer environment (NCE) as an analog to the non-host resistance (NHR) in plants. TSGs, tumor suppressor genes.
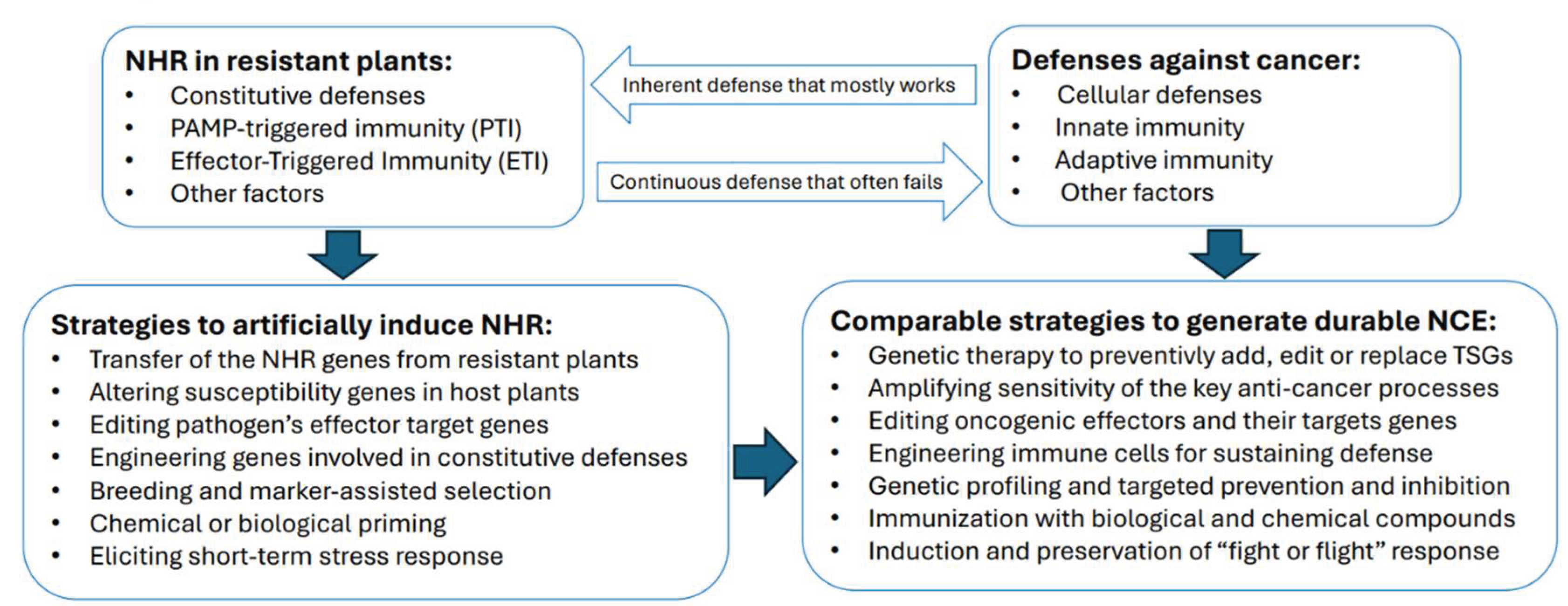
Figure 2.
Conditions essential for unveiling NHR and NCE states.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



