
Submitted:
22 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
Background: Skin repair and wound healing are tightly regulated biological processes that require coordinated control of inflammation, redox balance, angiogenesis, and tissue remodeling. Natural extracts are increasingly investigated not as nonspecific bioactive mixtures, but as sources of chemically diverse phytochemicals capable of modulating defined molecular signaling pathways involved in skin repair. Methods: Representative botanical sources, including Aloe Vera, Centella asiatica, Curcuma longa, Calendula officinalis, and Panax ginseng, have been extensively studied in pre-clinical wound models, providing insight into how distinct phytochemical classes intersect with shared regulatory mechanisms. Results: Flavonoids, terpenoids, phenolic acids, alkaloids, and polysaccharides have been shown to influence inflammatory signaling, redox-sensitive pathways, growth factor-mediated responses, and cellular migration, thereby supporting phase-appropriate progression of wound healing. At the molecular level, modulation of pathways such as NF-κB, TGF-β, VEGF, and Nrf2 emerges as a recurring mechanistic theme, while excessive or poorly timed pathway activation is increasingly recognized as a source of impaired tissue quality or fibrotic risk. Advances in dermopharmaceutical formulation strategies, including hydrogels, nanoemulsions, and lipid-based carriers, have improved local delivery and stability of phytochemicals, but also introduced additional translational and regulatory constraints. Conclusions: This review provides a mechanism-driven and pharmaceutical-oriented synthesis of current evidence, highlighting how molecular specificity, temporal regulation, and formulation design collectively determine the therapeutic relevance of plant-derived extracts in skin wound healing.
Keywords:
wound healing
; phytochemicals
; molecular mechanisms
; inflammatory signaling
; redox homeostasis
; angiogenesis
; dermopharmaceutical formulations
1. Introduction
Effective skin repair depends on tightly regulated interactions between inflammatory resolution, redox control, angiogenesis, and tissue remodeling, which must occur within a restricted temporal window to ensure functional regeneration.[1] Therapeutic interventions that disturb this balance, either by insufficient modulation or by excessive pathway activation, risk delaying closure, impairing tissue quality, or promoting fibrosis.[2] From a pharmaceutical perspective, the challenge in wound healing therefore lies not in broadly stimulating repair processes, but in selectively and transiently modulating key molecular pathways in a manner that is compatible with the dynamic biology of the healing wound.[3]
Against this mechanistic background, natural extracts are increasingly examined as sources of chemically diverse constituents capable of interacting with defined signaling networks, rather than as nonspecific bioactive mixtures.[4] Mechanistic studies support the notion that phytochemicals modulate inflammatory signaling, growth factor, mediated responses, redox sensitive pathways, and cell migration in a context dependent manner.[4] However, the same properties that underlie this multifunctionality, pleiotropic targets, compositional complexity, and sensitivity to dose and timing, also complicate interpretation and translation.[5] In addition to pathway level modulation of inflammation and redox signaling, several plant derived extracts exhibit antimicrobial activity against wound-associated pathogens and have been reported to influence extracellular matrix production, including collagen synthesis, in preclinical models. Without careful mechanistic resolution and pharmaceutical framing, reported biological effects risk remaining correlative or irreproducible, particularly when extrapolated from simplified experimental models.[5] A further layer of complexity arises from formulation constraints.[6] Many phytochemicals exhibit limited solubility, stability, or skin penetration, necessitating delivery strategies that may substantially alter local exposure and pathway engagement.[6] While advanced dermopharmaceutical systems can address some of these barriers at the delivery level, they also introduce new variables related to pharmacodynamics, safety, and regulatory feasibility.[6] Consequently, meaningful evaluation of natural extracts in wound healing requires integrated consideration of molecular mechanisms, formulation design, and functional repair outcomes, rather than isolated assessment of biological activity or technological sophistication.[6]
Against this background, this review advances a mechanism driven and pharmaceutical-oriented perspective on plant derived extracts in skin repair and wound healing. Rather than cataloguing botanical species or reported bioactivities, the review focuses on how defined classes of phytochemicals modulate key signaling pathways, how these effects intersect with the phase dependent biology of wound healing, and where current evidence remains limited or overinterpreted. The manuscript is structured to progress from molecular mechanisms to formulation considerations and representative medicinal plants, followed by a critical appraisal of limitations and translational challenges, thereby providing a coherent framework for evaluating natural extracts as mechanistically defined and clinically contextualized therapeutic candidates.
2. Materials and Methods
Literature relevant to the molecular and pharmaceutical aspects of skin repair and wound healing was identified through targeted searches of PubMed, Web of Science, and Scopus. The primary focus was placed on studies published within the last 5–7 years to reflect current mechanistic understanding, while earlier publications were considered when they provided seminal insights into key signaling pathways or experimental models. Selection was restricted to peer reviewed primary research articles addressing cutaneous wound healing, with emphasis on in vivo studies or in vitro investigations supported by clear mechanistic rationale and functional relevance.
Inclusion criteria required that studies examined defined molecular pathways involved in inflammation, redox regulation, growth factor signaling, angiogenesis, or cell migration, and that they reported outcomes linked to tissue repair rather than wound closure alone. Purely descriptive studies, ethnopharmacological surveys, reviews, and opinion pieces were excluded. The retained literature was critically evaluated based on mechanistic depth, appropriateness of the experimental model, temporal and cell type resolution, and the presence of functional healing endpoints. When formulation strategies were discussed, evaluation considered whether delivery systems were mechanistically justified, aligned with the targeted phase of wound healing, and supported by translationally relevant outcomes rather than by physicochemical performance alone.
Figure 1.
Overview of the literature identification, selection, and critical evaluation workflow applied in this review.
Figure 1.
Overview of the literature identification, selection, and critical evaluation workflow applied in this review.
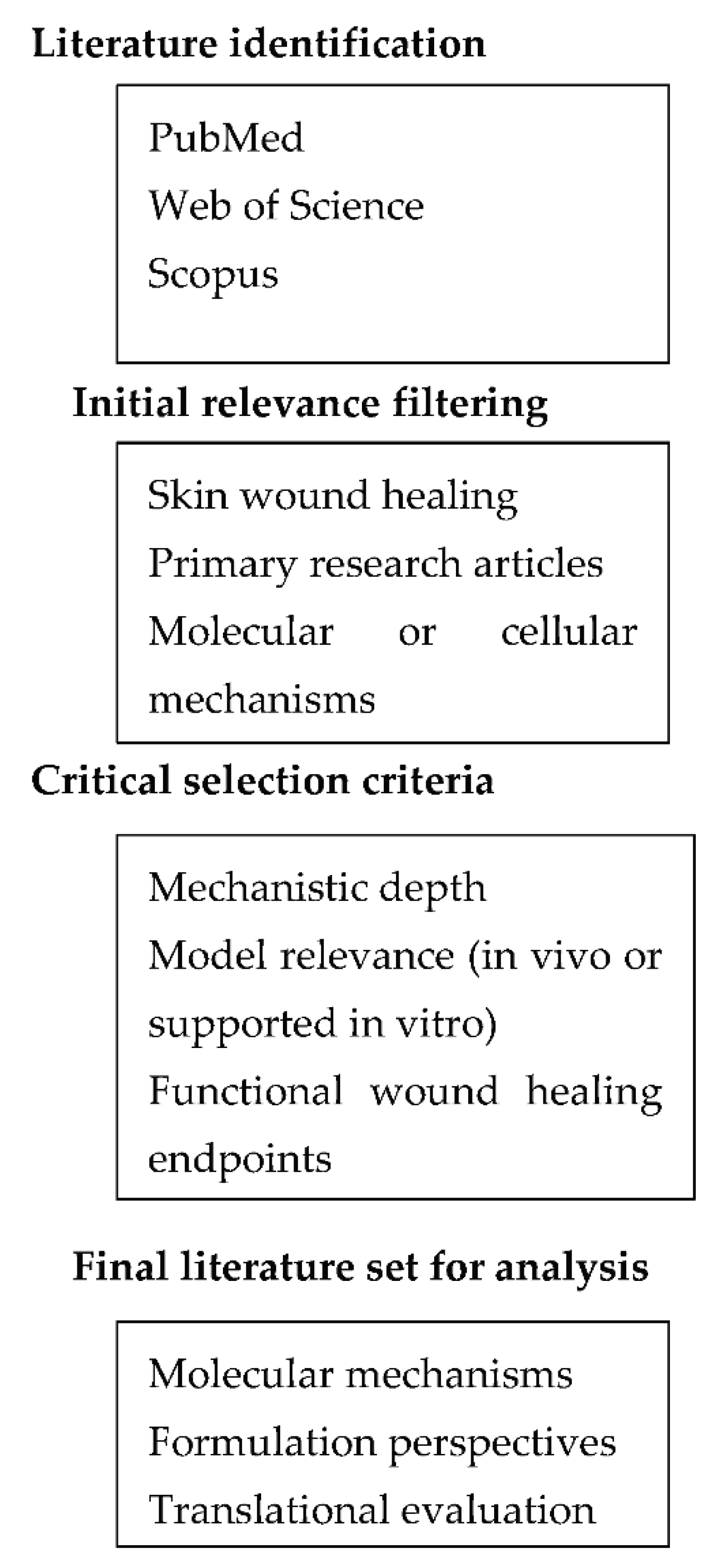
The diagram outlines the stepwise process used to identify primary research articles, apply mechanistic and translational relevance criteria, and integrate selected evidence into pathway, focused and pharmaceutical analyses.
3. Molecular Mechanisms of Skin Repair Modulated by Natural Extracts
This section synthesizes mechanistic evidence from primary preclinical studies addressing how plant derived phytochemicals modulate key molecular pathways across distinct phases of skin wound healing. Rather than presenting outcomes in isolation, the evidence is organized according to signaling axes and cellular processes that govern inflammatory resolution, redox homeostasis, angiogenesis, cellular migration, and tissue remodeling.
3.1. Modulation of Inflammatory Signaling Pathways
Inflammation is required for wound repair; however, the magnitude and timing of inflammatory signaling determine whether early host defense transitions efficiently toward tissue regeneration. Plant derived extracts influence this balance primarily by modulating, rather than abolishing, pro inflammatory signaling. Among the pathways implicated, the NF-κB axis occupies a central position due to its role in regulating early cytokine release, leukocyte recruitment, and oxidative stress responses. Multiple in vivo wound models show that several phytochemical classes attenuate excessive NF-κB activation in wound tissue, with concomitant reductions in TNF-α, IL-1β, and IL-6 expression, while preserving early inflammatory cues required for debris clearance and host defense; however, direct temporal analyses of early inflammatory kinetics remain limited.[7,8] This modulation is context- and dose dependent, and complete suppression of NF-κB signaling is neither observed nor desirable in physiologically relevant models of skin repair.[9] Beyond NF-κB, mitogen activated protein kinase (MAPK) pathways, including p38, JNK, and ERK, represent additional nodes through which natural compounds influence inflammatory signaling. Stress activated kinases such as p38 and JNK are frequently downregulated by flavonoids and phenolic acids, correlating with reduced inflammatory mediator production, whereas ERK signaling, more closely associated with cell survival and proliferation, tends to be preserved or indirectly enhanced. [9,10]
This differential regulation indicates that plant derived bioactives preferentially shift inflammatory signaling toward a pro healing phenotype rather than inducing broad pathway inhibition. However, in many studies’ MAPK modulation is inferred primarily from phosphorylation status, with limited integration of functional wound healing endpoints, most available data derive from keratinocyte and macrophage models. [10]
The cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) axis represents another frequently reported target of natural extracts. Terpenoids and triterpenes, in particular, have been shown to reduce COX-2 and iNOS expression in inflamed skin, thereby limiting prolonged prostaglandin and nitric oxide production associated with chronic inflammation. [8,9] Although these effects are frequently presented as beneficial, excessive emphasis on enzyme downregulation risks oversimplifying the roles of COX-2 and iNOS, which also contribute to angiogenesis and tissue remodeling at later stages of the healing cascade. Consequently, reviewer scrutiny often centers on whether COX-2 and iNOS modulation is linked to improved tissue architecture rather than solely to reduced inflammatory markers. [8]
A critical distinction in evaluating inflammatory modulation by natural extracts lies between anti-inflammatory and pro regenerative effects. Several studies indicate that specific phytochemicals facilitate the transition from a pro-inflammatory to a resolution phase by promoting macrophage polarization toward a resolution associated M2-like phenotype and increasing IL-10 production. [7,11] These changes contribute to the establishment of a microenvironment conducive to fibroblast activation and re epithelialization. Nonetheless, much of the supporting evidence remains correlative, particularly when derived from in vitro macrophage assays lacking in vivo validation. Overall, available evidence indicates that plant derived extracts can fine tune inflammatory signaling during wound healing, while highlighting the need for mechanistic studies that integrate temporal regulation, cell type specificity, and functional repair outcomes. [11] In selected models, attenuation of inflammatory signaling is accompanied by reduced microbial burden at the wound site, suggesting that immunomodulatory effects may indirectly contribute to antimicrobial defense without direct bactericidal activity.
3.2. Regulation of Growth Factors, Medicated Responses
Growth factor signaling represents a regulatory interface between inflammatory resolution and tissue regeneration during wound healing, with transforming growth factor-β (TGF-β) and vascular endothelial growth factor (VEGF) pathways exerting context dependent effects on repair outcomes. Preclinical wound models indicate that plant derived extracts influence these axes primarily by modulating downstream signaling dynamics rather than inducing sustained growth factor overexpression. In fibroblasts, several triterpenoid and saponin rich extracts enhance TGF-β/Smad2/3 signaling during the proliferative phase as assessed at early to intermediate post injury time points, where enhanced myofibroblast differentiation and extracellular matrix deposition are observed.[9,12] However, this effect is highly sensitive to temporal regulation, since prolonged or excessive activation of TGF-β signaling, particularly in the absence of Smad7, mediated negative feedback, is associated with fibrotic remodeling rather than functional tissue restoration.[2,12]
Beyond fibroblast activation, VEGF, dependent angiogenic signaling is a key determinant of oxygen and nutrient delivery to regenerating tissue. Phytochemicals such as polyphenols and curcuminoids have been reported to increase VEGF expression or signaling activity in endothelial cells with fibroblast and macrophage, derived VEGF contributing indirectly, often through redox sensitive mechanisms involving HIF-1α stabilization.[13] In vivo wound models indicate that moderate enhancement of VEGF signaling correlates with histologically detectable increases in capillary density and accelerated wound closure.[14] Nevertheless, angiogenic responses attributed to natural extracts are often inferred from surrogate measures, including VEGF expression or in vitro tube formation, which provide limited information on vessel maturation, perfusion, or long-term stability.[15] As a result, reviewer scrutiny often centers on whether angiogenesis is functionally integrated into tissue repair rather than merely amplified at the molecular level.[15]
Crosstalk between growth factor pathways further complicates interpretation of phytochemical effects. TGF-β signaling can indirectly influence angiogenesis through paracrine interactions between fibroblasts and endothelial cells, while VEGF driven neovascularization may reciprocally regulate fibroblast activity and collagen organization.[16] Certain extracts appear to coordinate these processes through a transient, pro-regenerative signaling milieu that supports keratinocyte migration and re-epithelialization without sustaining profibrotic cues. However, mechanistic evidence for such coordinated regulation remains limited, as many studies rely on single time point analyses and lack cell specific resolution within the wound microenvironment in vivo. [16]
From a translational perspective, modulation of growth factor signaling by natural extracts must be interpreted with caution. Acute wound models often overestimate pro regenerative benefits compared to chronic or impaired healing conditions, where dysregulated growth factor responses are already present.[5] Moreover, variability in extract composition and dosing complicates reproducibility and risk assessment, particularly with respect to fibrotic outcomes.[2] Overall, available evidence indicates that plant-derived compounds can fine tune growth factor mediated responses in skin repair, while highlighting the need for temporally resolved, cell specific studies linking signaling modulation to durable and functionally relevant healing outcomes.[3]
3.3. Oxidative Strss Control and Redox Homeostasis
Redox balance is a key regulator of phase specific wound progression, as reactive oxygen species (ROS) function both as damaging agents and as signaling mediators that coordinate inflammation, proliferation, and remodeling. In this context, the Nrf2 Keap1 pathway represents a central regulatory axis through which cells adapt to oxidative stress during skin repair. Beyond direct radical scavenging, several plant derived compounds modulate Nrf2 activation and nuclear translocation, thereby inducing a transcriptional program that enhances endogenous antioxidant defenses, an effect primarily supported by expression-based and short term functional assays.[17,18] Downstream targets such as heme oxygenase-1 (HO-1) and NAD(P)H quinone dehydrogenase 1 (NQO1) contribute to cytoprotection, redox buffering, and resolution of oxidative stress, particularly in keratinocytes and macrophages exposed to the inflammatory wound milieu.[19,20]
Phytochemical classes including flavonoids, phenolic acids, and certain terpenoids have been shown to influence Nrf2 signaling, often through redox sensitive modification of Keap1 cysteine residues or indirect modulation of upstream kinases.[18,20] In keratinocytes, Nrf2 activation has been associated with improved resistance to oxidative injury and enhanced migratory capacity, predominantly in in vitro or acute stress models.[4,20] In fibroblasts, redox homeostasis regulated via Nrf2-dependent pathways appears to facilitate survival and matrix producing activity during the proliferative phase.[17] However, these effects are highly context-dependent, since excessive or sustained Nrf2 activation during later remodeling stages can alter redox signaling required for growth factor responsiveness and matrix remodeling.[17,21]
A critical distinction that reviewers expect to see addressed is that between direct antioxidant activity and regulation of redox signaling. While many studies report reduced ROS levels following treatment with natural extracts, such observations alone do not establish therapeutic relevance. Low to moderate ROS concentrations are necessary for processes such as angiogenic signaling and growth factor activation, and indiscriminate ROS suppression may be counterproductive.[21] Consequently, mechanistic relevance is strongest in studies demonstrating that Nrf2-mediated transcriptional responses are linked to functional outcomes, such as improved wound closure rates, reduced oxidative damage to newly formed tissue, or enhanced tensile strength, rather than solely with biochemical markers of antioxidant capacity.[4,21]
Despite growing interest in redox targeted mechanisms, several limitations temper translational interpretation. Much of the available evidence is derived from in vitro models or short term in vivo studies that rely on expression-based readouts of Nrf2 or its target genes without assessing long term tissue integrity or scar quality.[4,18] Moreover, cell-type specificity within the wound microenvironment is often insufficiently resolved, complicating attribution of observed effects to keratinocytes, fibroblasts, or immune cells.[5] Overall, available evidence indicates that plant derived compounds can support wound healing by fine tuning redox homeostasis through Nrf2 dependent mechanisms, while highlighting the need for temporally resolved and cell specific studies linking redox signaling modulation to durable and functionally meaningful repair outcomes.[4]
3.4. Effects on Cellular Proliferation and Migration
Effective wound closure requires tightly coordinated proliferation and migration of multiple skin-resident cell populations, notably keratinocytes, fibroblasts, and endothelial cells. Plant-derived compounds influence these processes through modulation of intracellular signaling pathways that govern cytoskeletal dynamics, cell cycle progression, and cell matrix interactions, rather than by uniformly stimulating cell growth. In keratinocytes, several phytochemicals activate pathways such as ERK1/2 and PI3K/Akt, supporting cell survival and directional migration rather than increased proliferative index at the wound edge, thereby facilitating early re epithelialization.[22,23] Notably, these effects appear to be most pronounced under stress condition, supporting a role in restoring impaired migratory capacity rather than driving supraphysiological proliferation.[22]
Fibroblast responses to natural extracts are intrinsically context dependent reflecting the dual requirement for cell expansion and controlled matrix deposition. Phytochemical-mediated modulation of signaling networks, including TGF-β, dependent and focal adhesion, related pathways, has been linked to enhanced fibroblast migration, predominantly in preclinical wound models.[23,24] However, increased fibroblast proliferation alone does not equate to improved healing, as excessive or poorly regulated fibroblast activity may predispose to fibrosis, particularly in TGF-β, enriched microenvironments.[12] In selected models, attenuation of inflammatory signaling is accompanied by reduced microbial burden at the wound site, suggesting that immunomodulatory effects may indirectly contribute to antimicrobial defense without direct bactericidal activity. Consequently, mechanistic relevance is strongest in studies linking fibroblast migration and proliferation to improved granulation tissue organization and subsequent remodeling, rather than to cell number increases in isolation.[24]
Endothelial cell proliferation and migration underpin angiogenic sprouting and vascular network formation, processes that are indirectly influenced by phytochemicals through growth factor signaling and redox modulation. In vitro assays such as scratch wound closure, transwell migration, and tube formation are commonly used to demonstrate these effects, yet they provide limited insight into vessel maturation or functional perfusion.[25] In vivo evidence for enhanced endothelial migration and proliferation is more compelling when accompanied by histological or functional assessments of neovascular integration within regenerating tissue.[25]
Across cell types, a recurring limitation in the literature is the reliance on short term or single cell assays to infer regenerative benefit.[26] Proliferation markers or migration rates measured in isolation may overestimate therapeutic relevance if not contextualized within the multicellular and temporally regulated wound environment. Moreover, dose dependence and extract composition are rarely standardized, complicating translational interpretation. Overall, available evidence indicates that plant derived compounds can support wound healing by facilitating coordinated cell proliferation and migration, provided these effects are tightly regulated and integrated into the broader sequence of tissue repair events rather than treated as standalone regenerative drivers.[26] To facilitate integration of the mechanistic evidence discussed above, the principal signaling pathways, phytochemical classes, cellular targets, and functional outcomes implicated in plant mediated wound repair are summarized in Table 1.
4. Evidence from Key Medicinal Plants
The wound healing activity of medicinal plants can be understood through their convergence on a restricted set of molecular and cellular processes regulating inflammatory resolution, redox control, angiogenesis, and tissue remodeling. Among the most extensively studied botanical sources, Aloe Vera, Centella asiatica, Curcuma longa, Calendula officinalis, and Panax ginseng provide representative examples of how distinct phytochemical profiles intersect with shared regulatory pathways, albeit with varying degrees of mechanistic clarity and translational maturity. Extracts from Aloe Vera are predominantly associated with modulation of early inflammatory responses and redox homeostasis, largely attributed to polysaccharides and phenolic constituents.[27] Preclinical wound models report attenuation of sustained NF-κB activation during early inflammatory stages, together with improved keratinocyte migration under oxidative stress conditions, consistent with Nrf2-dependent cytoprotective signaling discussed in Section 3.3.[27] However, despite widespread use in topical formulations, mechanistic attribution remains complicated by extract heterogeneity and limited resolution of cell type specific effects. Accordingly, the translational relevance of Aloe Vera currently resides primarily in supportive wound care applications rather than in pathway specific therapeutic modulation. Beyond inflammatory and redox modulation, Aloe Vera extracts have been reported to limit microbial colonization in wound environments, primarily as a secondary consequence of improved barrier restoration and immune regulation rather than direct antimicrobial activity. In contrast, Centella asiatica represents one of the most mechanistically substantiated botanical sources in wound healing, largely due to its triterpenoid constituents. These compounds consistently engage TGF-β/Smad signaling in fibroblasts, promoting granulation tissue formation and collagen deposition, as outlined in Section 3.2.[28,29] Nevertheless, dose dependence and long-term scar quality remain insufficiently addressed, particularly when extracts are incorporated into advanced delivery systems that can amplify pathway activation beyond physiologically optimal ranges.[29] Curcuma longa derived preparations, rich in curcuminoids, intersect multiple regulatory axes, including inflammatory signaling, redox control, and angiogenic responses. Consistent with the mechanistic framework discussed in Section 3.1 and Section 3.3, curcuminoids modulate inflammatory and redox sensitive pathways and indirectly influence VEGF associated angiogenic signaling under hypoxic conditions.[30] While these multimodal effects are attractive from a mechanistic standpoint, translational interpretation is constrained by poor intrinsic bioavailability and heavy reliance on formulation driven enhancement, as discussed in Section 5.[30] Consequently, reported therapeutic benefits are often inseparable from the employed delivery strategy, complicating attribution to the botanical extract alone. In selected preclinical models, curcuminoid-based interventions have also been associated with improved collagen fiber organization and tensile properties of the regenerated tissue, effects that appear to depend on tightly regulated growth factor signaling rather than generalized enhancement of matrix deposition. Extracts of Calendula officinalis are predominantly discussed in the literature for their anti-inflammatory and antimicrobial effects with reported effects on fibroblast migration and early angiogenic signaling. However, the available evidence is dominated by descriptive outcomes, such as accelerated wound closure, without sufficient mechanistic dissection linking these effects to defined signaling pathways.[31] As a result, Calendula occupies an intermediate evidentiary position: supportive of wound repair processes, yet mechanistically under characterized, despite consistent phenotypic outcomes, relative to more intensively studied species.[31] Reported antimicrobial effects of Calendula officinalis extracts are predominantly descriptive and insufficiently linked to defined molecular targets, limiting their mechanistic interpretation and translational relevance despite consistent phenotypic outcomes. Finally, Panax ginseng offers a distinct mechanistic profile through ginsenoside mediated modulation of angiogenesis, cell proliferation, and redox sensitive signaling. Available preclinical data indicate coordinated effects on endothelial and fibroblast responses, including VEGF, related pathways and stress-adaptive signaling cascades.[32] While these findings align with a pro regenerative framework, translational advancement remains limited by variability in ginsenoside composition and insufficient comparative studies against simpler botanical or synthetic interventions.[14,32] These botanical examples indicate that therapeutic relevance in wound healing is determined not by plant identity alone, but by alignment of phytochemical composition, mechanistic engagement, and formulation strategy with controlled and phase-appropriate modulation of repair processes. Across species, a recurring limitation lies in the extrapolation of preclinical findings without adequate consideration of dose, timing, and extract standardization. [33] Meaningful progress in this area will depend less on expanding the list of candidate plants and more on integrating mechanistic rigor with translationally relevant evaluation frameworks. [33]
5. Dermopharmaceutical and Formulation Perspectives
Although preclinical studies provide mechanistic insight into the wound healing activity of plant derived extracts, translation into dermopharmaceutical products remains limited by formulation specific constraints. A substantial proportion of wound relevant phytochemicals is characterized by poor aqueous solubility, chemical instability, and limited skin penetration, collectively limiting effective local exposure at the wound site.[34] In addition, the complex and variable composition of botanical extracts complicates dose standardization, reproducibility and therapeutic predictability.[33] From a reviewer perspective, addressing these physicochemical and biopharmaceutical limitations is essential to justify claims of pharmaceutical relevance.[33]
Most advanced delivery systems investigated to date are designed to address physicochemical and biopharmaceutical limitations rather than to directly augment the intrinsic biological activity of the extracts. Hydrogel based formulations are among the most frequently studied systems in preclinical wound models, primarily due to their ability to maintain hydration, enable sustained release, and retain hydrophilic or macromolecular constituents at the wound interface[35] When mechanistically justified, hydrogels may also modulate wound microenvironmental factors such as hydration, pH, and oxygen diffusion, indirectly supporting cellular migration and matrix remodeling.[35] Accordingly, the contribution of hydrogels is more accurately interpreted as supportive, since many reported benefits arise from carrier-associated effects rather than from altered extract pharmacodynamics.[35]
Lipid-based carriers, including nanoemulsions, liposomes, and solid lipid nanoparticles, are predominantly investigated to improve the solubility, stability, and cutaneous delivery of lipophilic phytochemicals such as terpenoids and curcuminoids.[34,36] In preclinical models, these systems can increase local drug deposition and prolong residence time within superficial skin layers, which may be most relevant during the early inflammatory phase of wound healing.[36] A recurring concern is whether improvements in penetration metrics translate into measurable gains in wound healing quality particularly when functional endpoints such as tensile strength, scar quality, or long term tissue integrity are not assessed alongside pharmacokinetic improvements.[36]
Formulation studies frequently obscure the distinction between technological complexity and demonstrable therapeutic advancement. Claims that nanocarriers or complex delivery platforms inherently enhance wound healing are often insufficiently supported when compared to simpler formulations or appropriate controls or clinically relevant comparators.[37] Moreover, increased bioavailability may carry unintended consequences, including off target effects or excessive activation of pathways such as TGF-β signaling, potentially increasing fibrotic risk.[12] Mechanistic alignment between the delivery system, the bioactive constituents, and the targeted phase of wound healing therefore represents a critical but frequently underdeveloped aspect of formulation studies.[37]
Translation into regulated dermopharmaceutical products introduces additional challenges that extend beyond formulation performance. The scalability of advanced delivery systems, long term safety of nanomaterials, and regulatory classification of botanical based formulations remain significant hurdles.[36,38] Regulatory agencies require well defined composition, stability, and safety profiles, particularly under pharmaceutical rather than cosmetic regulatory frameworks.[38] Accordingly, the translational value of dermopharmaceutical formulations will depend on mechanism guided design and rigorous evaluation of therapeutic benefit and regulatory feasibility, rather than on formulation novelty per se.[33,38] Importantly, formulation-driven enhancement of biological responses should be interpreted within the context of phase-specific wound biology, as delivery strategies optimized for early inflammatory modulation may not be appropriate during proliferative or remodeling stages. Key formulation challenges, mechanistic rationales, and translational limitations associated with plant derived wound healing products are summarized in Table 2.
6. Limitations, Knowledge Gaps, and Translational Challenges
Despite substantial progress in delineating how plant derived extracts modulate wound healing pathways, several systemic limitations continue to constrain interpretation and translational advancement. A recurring limitation across mechanistic studies is the reliance on simplified experimental models that fail to recapitulate the temporal, multicellular, and phase dependent organization of skin repair.[39] Pathway modulation based readouts therefore limit insight into how these signals are coordinated across successive phases of healing.[39,40] Moreover, mechanistic conclusions are frequently extrapolated from in vitro or acute in vivo models without adequate validation in chronic or impaired wound settings where dysregulated inflammation, oxidative stress, and growth factor signaling, fundamentally alter therapeutic responsiveness.[5,39]
From both botanical and formulation perspectives, heterogeneity represents a central and unresolved challenge. Variability in extract composition, plant source, processing methods, and dosing complicates cross study comparison and undermines reproducibility, even when nominally identical species are investigated.[33] While advanced delivery systems can mitigate some bioavailability and stability issues, they also introduce additional layers of complexity, including altered pharmacodynamics, potential off target effects, and regulatory uncertainty.[36] Importantly, improvements in penetration or local concentration do not inherently translate into superior healing outcomes particularly when pathway activation exceeds the narrow therapeutic window required for regenerative rather than fibrotic responses.[36] Collectively, these issues highlight a persistent gap between mechanistic plausibility and functionally meaningful repair.[39]
Translational barriers in wound healing are further amplified by the multifactorial nature of clinical wounds and the absence of standardized evaluation frameworks. Endpoints such as accelerated closure are frequently prioritized over long term measures of tissue quality, tensile strength, or scar architecture, limiting clinical relevance.[39] In addition, regulatory requirements for botanical based dermopharmaceuticals demand well defined composition, safety, and manufacturing consistency, criteria that are challenging to meet for multicomponent natural extracts incorporated into complex formulations.[38] Addressing these gaps will require a shift toward temporally resolved, cell specific, and outcome oriented study designs, coupled with greater emphasis on standardization and comparative benchmarking.[33,40] Such an approach is necessary to move the field beyond isolated proof of concept observations toward reproducible and clinically translatable wound-healing strategies.[39]
7. Future Perspectives and Research Directions
Progress in the development of plant-derived therapeutics for skin repair depends less on expanding candidate lists than on the application of mechanism-guided experimental design. Priority should be given to moving beyond crude extracts toward active fractions, defined phytochemical combinations, or single lead compounds with clearly delineated molecular targets.[41] Such an approach allows more rigorous evaluation of dose–response relationships, temporal activity windows, and pathway-specific effects, particularly for signaling axes with narrow therapeutic ranges such as TGF-β–mediated matrix deposition or redox-sensitive Nrf2 signaling.[2,12] Mechanistic studies must be designed to resolve cell-type–specific responses within the wound microenvironment recognizing that keratinocytes, fibroblasts, endothelial cells, and immune cells may respond divergently to the same bioactive stimulus.[1,3,5]
Temporally resolved and functionally anchored study designs are required to capture the dynamic progression of wound healing.[40] Single time-point measurements or expression-based readouts rarely provide sufficient evidence for therapeutic relevance; pathway modulation should instead be linked to integrated functional outcomes, including tissue architecture, tensile strength, vascular maturity, and scar quality, assessed across distinct healing phases.[40] Greater reliance on chronic and impaired wound models is necessary to assess whether proposed mechanisms remain operative under clinically relevant pathological conditions.[5,39]
From a translational standpoint, formulation strategies should be guided by biological phase and target pathway rather than by technological complexity. Delivery systems should be aligned with the intended target pathway, cellular compartment, and phase of wound healing, rather than selected on the basis of novelty or technological complexity.[37,36] Parallel efforts in standardization, comparative benchmarking against existing therapies, and regulatory-aligned study design will be necessary to bridge preclinical findings with clinical feasibility.[33,38] Future progress will also depend on integrating comparative benchmarking against existing standard of care treatments to contextualize mechanistic benefits within clinically meaningful performance thresholds. Collectively, these considerations support a more disciplined integration of natural product chemistry, wound biology, and pharmaceutical science, positioning plant derived compounds as mechanistically defined and clinically contextualized therapeutic candidates rather than broadly acting remedies. [41]
8. Conclusions
The therapeutic relevance of plant derived extracts in skin repair depends less on the breadth of reported bioactivities than on the precision with which specific molecular pathways are modulated within defined temporal windows of the healing process. Anti-inflammatory, redox regulating, angiogenic, and pro migratory effects contribute meaningfully to repair only when coordinated with the phase dependent biology of the wound and constrained within narrow functional thresholds. Uncontrolled pathway activation, whether inflammatory suppression, growth factor signaling, or redox modulation, carries a clear risk of impaired tissue quality or fibrotic remodeling, reinforcing the importance of mechanistic discrimination over nonspecific pathway stimulation. Pharmaceutical relevance emerges at the intersection of molecular mechanism, formulation design, and translational feasibility. Advances in delivery systems can mitigate intrinsic limitations of phytochemicals, but do not replace the requirement for mechanistic clarity and functional validation. Botanical complexity, extract variability, and regulatory constraints limit interpretability when descriptive outcomes are not supported by standardized, outcome-oriented evaluation. Within this framework, natural extracts are best viewed not as inherently regenerative agents, but as sources of chemically defined modulators whose value depends on controlled and context appropriate integration into evidence based wound healing strategies.
Author Contributions
Conceptualization, N.T. and E.R.; methodology, N.T.; software, V.S.G..; validation, N.T., and V.K.; formal analysis, N.T.; investigation, V.K and N.T..; resources, V.K.; data curation, N.T.; writing—original draft preparation, N.T.; writing—review and editing, N.T and V.K..; visualization, V.S.G..; supervision, E.R.; project administration, N.T and V.K..; funding acquisition, N.T and E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, Z.; Bian, X.; Luo, L.; Björklund, Å.K.; Li, L.; Zhang, L.; Chen, Y.; Guo, L.; Gao, J.; Cao, C.; et al. Spatiotemporal Single-Cell Roadmap of Human Skin Wound Healing. Cell Stem Cell 2025, 32, 479–498.e8. [CrossRef]
- Yao, L.; Rathnakar, B.H.; Kwon, H.R.; Sakashita, H.; Kim, J.H.; Rackley, A.; Tomasek, J.J.; Berry, W.L.; Olson, L.E. Temporal Control of PDGFRα Regulates the Fibroblast-to-Myofibroblast Transition in Wound Healing. Cell Rep. 2022, 40, 111192. [CrossRef]
- Almet, A.A.; Liu, Y.; Nie, Q.; Plikus, M.V. Integrated Single-Cell Analysis Reveals Spatially and Temporally Dynamic Fibroblast States during Wound Healing. J. Invest. Dermatol. 2025, 145, 645–659.e25. [CrossRef]
- Barakat, M.; Han, C.; Chen, L.; David, B.P.; Shi, J.; Xu, A.; Skowron, K.J.; Johnson, T.; Woods, R.A.; Ankireddy, A.; et al. Non-Electrophilic NRF2 Activators Promote Wound Healing in Human Keratinocytes and Diabetic Mice and Demonstrate Selective Downstream Gene Targeting. Sci. Rep. 2024, 14, 25258. [CrossRef]
- Theocharidis, G.; Thomas, B.E.; Sarkar, D.; Mumme, H.L.; Pilcher, W.J.R.; Dwivedi, B.; Sandoval-Schaefer, T.; Sîrbulescu, R.F.; Kafanas, A.; Mezghani, I.; et al. Single-Cell Transcriptomic Landscape of Diabetic Foot Ulcers. Nat. Commun. 2022, 13, 181. [CrossRef]
- Bahloul, B.; Ben Bnina, E.; Hamdi, A.; Castillo Henríquez, L.; Baccar, D.; Kalboussi, N.; Abbassi, A.; Mignet, N.; Flamini, G.; Vega-Baudrit, J.R. Investigating the Wound-Healing Potential of a Nanoemulsion–Gel Formulation of Pituranthos tortuosus Essential Oil. Gels 2024, 10, 155. [CrossRef]
- Li, S.; Zhang, Y.; Wang, Z.; et al. Puerarin Improves Diabetic Wound Healing via Regulation of Macrophage M2 Polarization Phenotype. Burns Trauma 2022, 10, tkac046. [CrossRef]
- Chen, L.-Y.; et al. Therapeutic Potential of Luteolin on Impaired Wound Healing in Streptozotocin-Induced Rats. Biomedicines 2021, 9, 761. [CrossRef]
- Zhang, M.; Li, Y.; Chen, X.; et al. Centella asiatica Enhances Diabetic Wound Healing by Decreasing Macrophage-Driven Inflammation via the AKT/MAPK/NF-κB Pathway. Front. Pharmacol. 2025, 16, 1382451. [CrossRef]
- Sun, R.; Liu, Y.; Wang, J.; et al. Integrated Network Pharmacology and Experimental Validation to Explore the Mechanisms Underlying Naringenin Treatment of Chronic Wounds. Sci. Rep. 2023, 13, 26043. [CrossRef]
- Li, S.; Zhang, Y.; Wang, L.; Zhao, X.; Hu, Y.; et al. SZC-6 Promotes Diabetic Wound Healing in Mice by Modulating the M1/M2 Macrophage Ratio and Inhibiting the MyD88/NF-κB Pathway. Pharmaceuticals 2025, 18, 1143. [CrossRef]
- Ma, J.; Liu, Y.; Zhang, X.; et al. Asiaticoside Promotes Cutaneous Wound Healing by Regulating TGF-β/Smad Signaling in Dermal Fibroblasts. Int. J. Mol. Sci. 2021, 22, 1307. [CrossRef]
- Dai, C.; Li, Q.; Maynard, H.D.; et al. Curcumin Promotes Angiogenesis and Wound Healing through HIF-1α/VEGF Signaling. Biomed. Pharmacother. 2021, 135, 111178. [CrossRef]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of Ginsenosides in Angiogenesis and Wound Healing via VEGF Signaling. J. Ginseng Res. 2022, 46, 1–12. [CrossRef]
- Cheng, W.; Chen, Y.; Liu, J.; et al. VEGF Expression Does Not Necessarily Predict Functional Angiogenesis during Cutaneous Wound Healing. Wound Repair Regen. 2020, 28, 326–337. [CrossRef]
- Liu, S.; Zhang, H.; Duan, E. Epithelial–Mesenchymal and Fibroblast–Endothelial Crosstalk in Wound Healing. Signal Transduct. Target. Ther. 2021, 6, 243. [CrossRef]
- Wu, J.; Deng, L.; Yin, L.; Mao, Z.; Gao, X. Curcumin Promotes Skin Wound Healing by Activating Nrf2 Signaling Pathways. Turk. J. Med. Sci. 2023, 53, 1127–1135. [CrossRef]
- Xu, C.; Hu, L.; Zeng, J.; et al. Gynura divaricata (L.) DC. Promotes Diabetic Wound Healing by Activating Nrf2 Signaling. J. Ethnopharmacol. 2024, 323, 117638. [CrossRef]
- Huang, Y.; et al. Sulforaphane Promotes Diabetic Wound Healing by Regulating Macrophage Efferocytosis and Polarization via Nrf2 Signaling. Int. Immunopharmacol. 2025, 150, 114243. [CrossRef]
- Ning, Y.; Yuan, Z.; Wang, Q.; He, J.; Zhu, W.; Ren, D.; Wo, D. Epigallocatechin-3-Gallate Promotes Wound Healing Response in Diabetic Mice by Activating Keratinocytes and Promoting Re-Epithelialization via the K16/NRF2/KEAP1 Signaling Axis. Phytother. Res. 2024, 38, 1013–1027. [CrossRef]
- Fan, J.; Liu, H.; Wang, J.; et al. Procyanidin B2 Improves Endothelial Progenitor Cell Function and Promotes Diabetic Wound Healing via Nrf2 Activation. J. Cell. Mol. Med. 2021, 25, 652–665. [CrossRef]
- Wang, X.; et al. Icariin Enhances Cutaneous Wound Healing by Promoting Keratinocyte Proliferation and Migration through AKT and ERK Signaling. Pharmacol. Res. 2023, 192, 106674. [CrossRef]
- Yang, L.; et al. Artocarpin Accelerates Wound Healing by Enhancing Fibroblast and Keratinocyte Proliferation and Migration through ERK and p38 Signaling. Sci. Rep. 2023, 13, 14058. [CrossRef]
- Zhang, L.; et al. Rhein Enhances Skin Wound Healing by Shifting PI3K/Akt-Mediated Fibroblast Proliferation and Migration. Eur. J. Pharmacol. 2024, 937, 175245. [CrossRef]
- Zare, M.; et al. Crocus sativus Petal Extract Promotes Fibroblast and Endothelial Cell Migration and Angiogenesis during Diabetic Wound Healing. Sci. Rep. 2024, 14, 19106. [CrossRef]
- Ong, Y.S.; et al. Flavonoid C-Glycosides from Oil Palm (Elaeis guineensis) Leaves Stimulate Fibroblast Proliferation and Migration. Antioxidants 2025, 14, 326. [CrossRef]
- Liu, Y.; et al. Aloe vera Polysaccharides Promote Cutaneous Wound Healing by Regulating NF-κB-Mediated Inflammation and Nrf2-Dependent Antioxidant Responses. Int. J. Biol. Macromol. 2021, 182, 1235–1246. [CrossRef]
- Bylka, W.; et al. Asiaticoside and Madecassoside Regulate TGF-β/Smad Signaling and Collagen Deposition during Cutaneous Wound Healing. Phytomedicine 2020, 77, 153268. [CrossRef]
- Sun, B.; et al. Temporal Regulation of TGF-β Signaling by Centella asiatica Extract Prevents Excessive Fibrosis during Wound Repair. J. Ethnopharmacol. 2022, 289, 115002. [CrossRef]
- Li, X.; et al. Curcumin Accelerates Cutaneous Wound Healing by Modulating NF-κB, Nrf2 and VEGF Signaling Pathways. Biomed. Pharmacother. 2021, 138, 111427. [CrossRef]
- Preethi, K.C.; et al. Wound Healing Activity of Calendula officinalis Flower Extracts: In Vivo Evidence and Mechanistic Limitations. J. Ethnopharmacol. 2020, 260, 112981. [CrossRef]
- Kim, J.H.; et al. Ginsenoside Rg1 Promotes Angiogenesis and Fibroblast Proliferation during Cutaneous Wound Healing via VEGF-Related Signaling. Front. Pharmacol. 2021, 12, 702737. [CrossRef]
- Heinrich, M.; Appendino, G.; Efferth, T.; Fürst, R.; Izzo, A.A.; Kayser, O.; Pezzuto, J.M.; Viljoen, A. Best Practice in Research—Overcoming Common Challenges in Phytopharmacological Research. J. Ethnopharmacol. 2020, 246, 112230. [CrossRef]
- Puglia, C.; Offerta, A.; Tirendi, G.G.; Tarico, M.S.; Curreri, S.; Bonina, F. Design of Solid Lipid Nanoparticles for the Delivery of Natural Compounds: Physicochemical and Biopharmaceutical Limitations. Pharmaceutics 2021, 13, 170. [CrossRef]
- Boateng, J.; Catanzano, O. Advanced Therapeutic Dressings for Effective Wound Healing—A Review. J. Pharm. Sci. 2021, 110, 266–279. [CrossRef]
- Santos, A.C.; Veiga, F.; Ribeiro, A.J. Lipid-Based Nanocarriers as Dermal Delivery Systems for Natural Compounds: From Improved Penetration to Biological Relevance. Nanomaterials 2022, 12, 240. [CrossRef]
- Jafari, S.M.; McClements, D.J. Nanotechnology Approaches for Increasing the Bioavailability of Natural Compounds: A Critical Review of Functional Relevance. Trends Food Sci. Technol. 2023, 131, 164–178. [CrossRef]
- European Medicines Agency (EMA). Guideline on Quality of Herbal Medicinal Products; EMA: Amsterdam, The Netherlands, 2018.
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound Repair and Regeneration: Mechanisms, Signaling, and Translation. Sci. Transl. Med. 2014, 6, 265sr6.
- Hardman, M.J.; Ashcroft, G.S. Progress in Wound Healing: Moving Beyond Single Time-Point Analyses. J. Invest. Dermatol. 2022, 142, 247–254.
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216.

Table 1.
Molecular Mechanisms Targeted by Plant Derived Phytochemicals during Distinct Phases of Skin Wound Healing.
Table 1.
Molecular Mechanisms Targeted by Plant Derived Phytochemicals during Distinct Phases of Skin Wound Healing.
| Wound Healing Phase | Molecular Pathway /Axis | Primary Cellular Targets | |
| Representative Phytochemical Classes | |||
| Early inflammatory phase | NF-κB signaling | Flavonoids, phenolic acids, terpenoids | Keratinocytes, macrophages |
| Early inflammatory phase | MAPK pathways (p38, JNK, ERK) | Flavonoids, phenolic acids | Keratinocytes, macrophages |
| Resolution / proliferative transition | Macrophage polarization (M1 → M2-like) | Polyphenols, triterpenoids | Macrophages |
| Proliferative phase | TGF-β/Smad signaling | Triterpenoids, saponins | Fibroblasts |
| Proliferative phase | VEGF-mediated angiogenic signaling | Polyphenols, curcuminoids | Endothelial cells (direct); fibroblasts/macrophages (indirect) |
| Multiple phases | Nrf2–Keap1 redox signaling | Flavonoids, phenolic acids, terpenoids | Keratinocytes, fibroblasts, macrophages |
| Re-epithelialization | ERK1/2, PI3K/Akt | Diverse phytochemicals | Keratinocytes |
1 Mechanistic effect summarized in this table are derived from preclinical in vivo and mechanistically supported in vitro wound healing models. Reported pathway modulation reflects context and phase dependent observations and should not be interpreted as uniform or sustained activation across all stages of skin repair.
Table 2.
Dermopharmaceutical Formulation Strategies for Plant Derived Wound Healing Agents: Mechanistic Rationale and Translational Constraints.
Table 2.
Dermopharmaceutical Formulation Strategies for Plant Derived Wound Healing Agents: Mechanistic Rationale and Translational Constraints.
| Formulation Type | Mechanistic Implications | Translational and Regulatory Limitations | ||
| Primary Delivery Rationale | Relevant Phytochemical Classes | |||
| Hydrogels* | Local retention; moist wound environment; sustained release | Polysaccharides, hydrophilic polyphenols | Indirect support of cell migration and matrix remodeling via microenvironment modulation | Many benefits attributable to carrier properties rather than extract pharmacodynamics |
| Nanoemulsions | Improved solubility and dispersion of lipophilic compounds | Terpenoids, essential oils, curcuminoids | Increased local exposure in superficial wound layers, potentially enhancing early-phase signaling | Penetration metrics often not linked to improved tissue quality or long-term outcomes |
| Liposomes | Encapsulation and protection of unstable phytochemicals | Polyphenols, flavonoids | Altered cellular uptake and pathway engagement | Stability, scalability, and batch reproducibility remain challenging |
| Solid lipid nanoparticles | Controlled release and improved stability | Lipophilic phytochemicals | Prolonged residence time at the wound surface | Risk of overexposure and off target pathway activation |
| Composite systems | Combination of retention and enhanced delivery | Mixed phytochemical profiles | Multimodal modulation of inflammatory and redox pathways | Increased formulation complexity complicates regulatory approval |
| Conventional topical formulations | Simplicity and regulatory familiarity | Broad | Supportive delivery without targeted modulation | Limited control over exposure and pathway specificity |
1 Reported mechanistic implications are based on preclinical wound healing studies and reflect the combined contribution of phytochemical activity and delivery system associated effects. Improvements in formulation performance do not necessarily correspond to enhanced intrinsic bioactivity or clinically translatable therapeutic benefit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



