
Submitted:
22 January 2026
Posted:
22 January 2026
You are already at the latest version
Abstract
The skeletal muscle tissue of dairy cows plays a crucial role in energy metabolism and is the site of β-hydroxybutyrate (BHBA) metabolism. However, the regulatory mechanism of BHBA in muscle differentiation remains unclear. This study used bovine skeletal muscle satellite cells (BMSCs) to investigate the effects of BHBA on the differentiation of bone marrow mesenchymal stem cells and to explore the regulatory roles of LNC297, novel-miR-145, and GAS7 in this process. The results of dual-luciferase reporter gene assays and miRNA pull-down experiments verified the targeting relationship between LNC297, novel-miR-145, and GAS7. BHBA inhibits the differentiation of BMSCs in a dose-dependent manner. Both LNC297 and GAS7 promote the differentiation of BMSCs and attenuate the inhibitory effect of BHBA. In contrast, novel-miR-145 inhibits BMSCs differentiation and enhances the inhibitory effect of BHBA. Mechanistically, LNC297 acts as a competing endogenous RNA (ceRNA) or molecular sponge for novel-miR-145, thereby upregulating GAS7 expression and promoting its differentiation function. Impaired muscle development in ketosis dairy cows is associated with the accumulation of high concentrations of BHBA in muscle tissue. LNC297 may promote muscle differentiation through the novel-miR-145/GAS7 axis, thereby alleviating muscle damage in ketosis cows. These findings provide new insights into the mechanisms of muscle development disorders in ketosis dairy cows.
Keywords:
Ketosis cow
; skeletal muscle
; differentiation
; LNC297
; novel-miR-145/GAS7
1. Introduction
In late pregnancy and early postpartum, dairy cows often enter a state of negative energy balance (NEB) due to factors such as increased fetal size leading to uterine compression, hormonal changes, and insufficient energy intake [1]. At this stage, dairy cows mobilize stored body fat, which is converted into triglycerides (TG). These TGs are then broken down into non-esterified fatty acids (NEFA). When NEFAs undergo incomplete oxidation in the liver, a large amount of ketone bodies is produced and released into the bloodstream [2]. Ketone bodies include acetoacetate, β-hydroxybutyrate, and acetone. When the body experiences energy deficiency, they can be used as fuel and transported to extrahepatic tissues for oxidation [3]. Previous studies have shown that BHBA is the most abundant ketone body in the human body. It is not only an important energy substrate for peripheral tissues but is also considered to promote resistance against oxidative and inflammatory stress, as well as maintain metabolic health [4]. In addition, BHBA might trigger cellular responses, adapting to the environment by enhancing mitochondrial activity and strengthening mechanisms against oxidative stress [5]. BHBA can improve insulin resistance in type2 diabetic mice through the HCAR2/Ca2+/cAMP/PKA/Raf/ERK1/2/PPARγ pathway [6]. In studies on dairy cows, if excessive BHBA is produced from NEFA due to insufficient metabolic adaptation, and this production exceeds the cow's metabolic capacity, the concentration of BHBA in the blood may exceed 1.2 mM, thereby triggering ketosis [7]. BHBA might also serve as an alternative energy source to supply energy to muscles and maintain their specific physiological functions [8,9]. Existing evidence suggests that elevated levels of BHBA in the blood during ketosis in dairy cows not only reduce milk production and reproductive performance but also increase the risk of prenatal diseases, such as mastitis and displaced abomasum [7,10]. Our recent study showed that the expression levels of MYH4 and MYH7 proteins in the muscle tissue of ketosis dairy cows within one week after calving were reduced, suggesting that the muscle fibers of ketosis cows may have sustained damage [11]. However, whether the muscle tissue damage observed in ketosis dairy cows is directly caused by elevated concentrations of BHBA still requires further experimental validation.
Long non-coding RNAs (lncRNAs) are nucleotide sequences longer than 200 base pairs. Initially, they were considered transcriptional noise due to their lack of protein-coding potential. However, increasing evidence has shown that lncRNAs are involved in various biological processes, including cell proliferation, differentiation, and apoptosis [12,13,14]. The growth and development of skeletal muscle begin in the embryonic mesoderm. Mesenchymal stem cells derived from the embryonic mesoderm differentiate into myoblasts, which then proliferate and differentiate to form myotubes. These myotubes further mature into muscle fibers. Ultimately, the muscle fibers grow and develop into mature skeletal muscle [15,16]. This process relies on the coordinated regulation of coding genes and non-coding RNAs, such as muscle regulatory factors (MRFs) [17], paired box proteins 3/7 (Pax3/7), and the myocyte enhancer factor 2 (MEF2) family[18]. LncRNAs are distributed in different organelles and regulate gene expression at the epigenetic, transcriptional, and post-transcriptional levels. For example, lncRNAs function as "molecular scaffolds," "decoys," "guiding molecules," and "signaling molecules" to activate or repress transcription [19]. For example, LncRNA H19promotes the differentiation of bovine skeletal muscle satellite cells by inhibiting the Sirt1/FoxO1 pathway [20].
MicroRNA(miRNA) is a small non-coding RNA molecule, 21 to 23 nucleotides in length, classified as a highly conserved class of endogenous short non-coding RNAs [21]. miRNAs are universally present in all eukaryotes and act as post-transcriptional regulatory factors of protein-coding mRNAs. They mainly function by pairing with complementary sequences in the 3' UTR region of target genes, thereby promoting mRNA degradation or inhibiting its translation [22]. For example, miR-103 and miR-139 affect the differentiation of bovine skeletal muscle-derived satellite cells by regulating the gene expression of cyclin E1 (CCNE1) and dihydrofolate reductase (DHFR), respectively [23]. In addition, lncRNA might act as a competitive endogenous RNA (ceRNA) to sequester miRNA from its target mRNA and participate in the regulation of muscle development [24,25]. Previous studies have shown that lncRNA-MEG3 promotes the differentiation of bovine skeletal muscle by interacting with miRNA-135 and MEF2C, thereby enhancing the expression of genes such as MHC and MYOG [26].
Our previous sequencing data indicated that, during the early stages of calving, muscle fiber formation in ketosis dairy cows is impaired. We also observed an increase in the expression of an unnamed lncRNA, MSTRG29792.1 (LNC297), a decrease in the expression of a newly identified miRNA(novel-miR-145), and a reduction in the expression of the downstream target gene GAS7. However, it remains unclear whether the impaired muscle fibers in ketosis dairy cows are related to the accumulation of high concentrations of BHBA in muscle tissue, and whether LNC297, novel-miR-145, and GAS7 regulate myocyte differentiation and the underlying regulatory mechanisms. Additionally, the potential regulation of muscle function under the influence of BHBA is not well understood. In this study, we found that BHBA inhibits BMSCs differentiation, LNC297 acts as a molecular sponge for novel-miR-145, inhibiting its expression and promoting the expression of GAS7, thus enhancing BMSCs differentiation and alleviating the differentiation inhibition induced by BHBA. This could serve as a potential target for the treatment of ketosis in dairy cows.
2. Materials and Methods
2.1. BMSCs Culture and Differentiation
All animal experiments in the present study involving animals were performed under the direction of the Institutional Animal Care and Use Committee from the College of Animal Science and Technology, Sichuan Agricultural University, China. (Certification No. SYXK2019-187; Approval time: 2019/1/29). The bovine muscle satellite cells (BMSCs) were derived from the right hindlimb muscle tissue of a newly born Chinese Holstein dairy calf. The calf was euthanized using electric shock, and then muscle tissue was obtained using sterile dissection tools. The tissue was placed in PBS containing 4% penicillin-streptomycin (Gibco, Thermo, USA) and quickly transferred into a UV-sterilized biosafety cabinet. The muscle tissue was minced in PBS containing 2% penicillin-streptomycin and the fascia and blood vessels were removed. The minced muscle tissue was then digested with 0.1% Type II collagenase (Solarbio, Beijing, China) diluted in DMEM/F12 basal medium (Gibco, Thermo, USA) at a tissue-to-digestive enzyme ratio of 1:5. The digestion process was carried out in a 37°C water bath for 30 min, with gentle shaking every5 minutes. The digestion was stopped once no obvious tissue clumps remained. After digestion, 10%fetal bovine serum (FBS, Gibco, Thermo, USA) was added to terminate the digestion process. The resulting cell suspension was then filtered through a 70 μm filter and the filtered cell suspension was seeded into a T25 cell culture flask for incubation in a 37°C, 5% CO2 cell culture incubator. The growth medium was replaced every 2 d. When the cell density reached 70%-80%, passaging was performed. The cells were then passaged to the second generation and cryopreserved in liquid nitrogen for future use. Furthermore, when the second-generation BMSCs reached approximately 85% confluence, the growth medium was replaced with F12 differentiation medium containing 2% horse serum (Solarbio, Beijing, China) and cultured for 5 d. The differentiation status of muscle cells was monitored daily under a microscope.
2.2. Plasmid Construction and Cell Transfection
The LNC297 overexpression plasmid (pc-LNC297), short interfering RNAs of GAS7 (si-GAS7), and mimic and inhibitor of novel-miR-145 were synthesized by Beijing Qingke Biotechnology Co., Ltd. In the differentiation experiment, the second-generation BMSCs were cultured in a 6-well plate. When the cell density exceeded 80%, mimic, inhibitor, mimic NC, inhibitor NC, pc-LNC297, and siRNA were transfected into BMSCs according to the instructions of Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). After 6 hours of transfection, the medium was replaced with differentiation medium containing 2% horse serum for 3 d of culture. The sequence information is shown in Table S1.
2.3. Target Gene Prediction and Luciferase Reporter Gene Assay
This study used a dual-luciferase reporter system to explorethe interactions between novel-miR-145 and GAS7, as well as between GAS7 and LNC297. The miRanda and Targetscan (http://www.targetscan.org/vert_71/) databases were employed to predict the target genes of novel-miR-145 and their binding to the 3' UTR of these target genes. Potential binding sites between novel-miR-145 and LNC297 were predicted using DNASTAR and Lasergene v7.1 (based on sequence information). The luciferase reporter plasmids (wild type (WT) and mutant of the target sequence) were constructed by Qingke Biotechnology Co., Ltd. (Beijing, China). Next, 293T cells were seeded in a 24-well cell culture plate (NEST Biotechnology, Wuxi, China). When the cell density reached approximately 70%, novel-miR-145mimic or negative control was co-transfected with specific WT or mutant plasmids according to the Lipofectamine 3000 reagent manual. After 48 h, luciferase activity was measured using the Duo-Lite TM luciferase detection system from Vazyme (Nanjing, China).
2.4. Real-Time Fluorescence Quantitative Analysis
Total RNA from the cells was extracted according to the manufacturer's instructions of the RNAiso reagent kit (Takara, Beijing, China). The purity of the total RNA was assessed by measuring the A260/A280 and A260/A230 ratios using a NanoDrop 2000 UV-Vis spectrophotometer (Thermo, MA, USA). In addition, the integrity of the total RNA was assessed by 1.0% agarose gel electrophoresis.The total RNA and miRNA were reverse transcribed into first-strand cDNA using the PrimeScript RT reagent kit (Takara, Beijing, China) and the SYBR® PrimeScript™ miRNA RT-PCR Kit (Takara, Beijing, China), respectively, according to the manufacturer’s instructions. The qPCR analysis was performed using the CFX96 system (Bio-Rad, Hercules, CA, USA). The β-actin and U6 were used as reference genes for qPCR, with a reaction volume of 10 μL. The relative expression levels of each gene were analyzed using the 2-ΔΔCt method. The mRQ 3' primer from the Mir-X™ miRNA First-Strand Synthesis Kit (Takara, Beijing, China) was used as the reverse primer for miRNA measurement, and the primer information is shown in Table S2.
2.5. Immunofluorescence
The BMSCs were seeded into 24-well cell culture plates. When the cell density reached approximately 60%-70%, they were removed. After washing three times with PBS, 4% cell fixative (Servicebio, Wuhan, China) was added to fix the cells for 15 min. After fixation, the cells were washed three times with PBS, followed by the addition of 0.2% permeabilization solution (Servicebio, Wuhan, China) for 10 min. Afterward, the cells were washed three times with PBS, each time for 5 min. Add blocking solution (Servicebio, Wuhan, China) and incubate on a shaker for1 h. Then wash with PBS three times, each time for 5 min. Add 200 μL of the primary antibody diluted with antibody dilution buffer (Servicebio, Wuhan, China) to each well and incubate overnight at 4°C on a shaker. Afterward, add the fluorescent secondary antibody [FITC-conjugated Goat anti-Rabbit IgG (H+L)] and incubate for 2 h at room temperature in the dark. Then, add 250 μL of DAPI (Beyotime, Shanghai, China) for staining for 15 min. Finally, wash with PBS twice, each time for 5 min, then add a small amount of PBS and place the cell culture plate under an inverted fluorescence microscope to capture images of green fluorescence (positive cells, excitation wavelength =488 nm) and blue fluorescence (nucleus, excitation wavelength = 358 nm) in the same field of view.
2.6. RNA Pull-Down
The RNA pull-down Kit (GeneCreate, Wuhan, China) was used to verify the targeting relationship between novel-miR-145 and LNC297. The biotin-labeled mimic of novel-miR-145 was synthesized by Qingke Biotechnology Co., Ltd. The BMSCs were seeded into 10 cm culture dishes. When the cell density reached 60%, transfection was performed using Lipofectamine 3000 according to the manufacturer’s instructions. In the experimental group, the biotin-labeled miRNA probe and pc-LNC297 were co-transfected, while the control group was transfected with mimic NC and empty plasmid. After 48 h of culture, the cells were collected by trypsin digestion and washed twice with precooled PBS. The following steps were performed according to the manufacturer’s instructions for the RNA pull-down Kit. In brief, the collected cells were lysed, magnetic beads were incubated with the probes, RNA was extracted, the first strand of cDNA was synthesized, and qPCR was performed to verify whether novel-miR-145 binds to LNC297.
2.7. Western Blot Analysis
Total protein from cells was extracted using a total protein extraction kit (Solarbio, Beijing, China), and the protein concentration was measured using a protein assay kit (Bestbio, Shanghai, China). Only samples with protein concentrations greater than 10 mg/mL were used for subsequent experiments. In brief, the protein samples were prepared by mixing total protein with SDS-PAGE loading buffer (Biosharp, Shanghai, China) in 4:1 ratio, and then the samples were denatured at 99°C for 10 min. Denatured proteins were separated using a10% one-step gel electrophoresis prep kit (ORISCIENCE, Chengdu, China). The separated gel was used for protein transfer to a 0.22 μm PVDF membrane via a wet transfer method (electrophoresis conditions: 400 mA, 60 min). The membrane was gently incubated at room temperature for 1.5 hours in Tris-buffered saline with Tween (TBST) containing 5.0% skimmed milk (Solarbio, Shanghai, China). The membrane was then incubated overnight at 4°C with the corresponding primary antibody on a shaking platform. After primary antibody incubation, the membrane was washed three times with TBST for 5 min each. The membrane was then incubated for 1.5 h with the secondary antibody [Goat anti-Rabbit or anti-Mouse IgG H&L (HRP), Zen Bioscience, Chengdu, China]. Finally, the membrane was washed three times with TBST. Immunoreactions were detected using ECL chemiluminescence reagent (HAKATA, Shanghai, China), and images were captured using the Touch Imager Pro device from e-BLOT Biotech (Shanghai, China).WB grayscale values were statistically calculated using Image J (v 1.53, Bethesda, MD, USA). Specific information about the primary antibody is provided in Table S3.
2.8. Cell Culture, BHBA and NAHB Treatment and Transfection
BMSCs were cultured in growth medium containing 10% FBS in a cell incubator at 37°C with 5% CO2. For BHBA (773861, Sigma, USA) treatment, when the BMSCs reached 85-90% confluence, differentiation induction medium containing 2% horse serum was added and cultured for 60 h. The differentiation medium was then replaced with medium containing 0 mM,1.2 mM, 2.4 mM, or 4.8 mM BHBA and cultured for 12 h. For NAHB (54965, Sigma, USA) treatment, after60 h of differentiation, the differentiation medium was replaced with medium containing 4.8 mM NAHB and cultured for 12 h. To investigate the co-treatment of LNC297, novel-miR-145, and GAS7 with BHBA, BMSCs were transfected and induced to differentiate as described in method 2.2, and after 60 h of differentiation, the differentiation medium was replaced with induction differentiation medium containing 4.8 mM BHBA and cultured for an additional 12 h.
2.9. Statistical Analysis
Statistical analysis was performed using GraphPad (v10.4.0) software, with intergroup differences assessed by Student’s t-test. In addition, multiple comparisons were performed using one-way or two-way ANOVA analysis. When * P < 0.05, the differences were significant. All data are presented as means ± SEM.
3. Results
3.1. BHBA Inhibits the Differentiation of BMSCs
The BMSCs in this study were obtained from the hind limb skeletal muscle of neonatal Chinese Holstein cows. To verify their identity as BMSCs, Desmin muscle-specific antibody was used. Desmin is a unique intermediate filament protein in muscle cells, widely present in skeletal muscle, cardiac muscle, and smooth muscle cells[27]. the results show that the cells used in this study are BMSCs (Figure 1A). Previous studies have indicated that both the acidic BHBA and the neutral sodium salt (NAHB) might be used in in vitro experiments with 3-hydroxybutyrate. To investigate which substance better reflects the effect of 3-hydroxybutyrate on muscle differentiation in a ketosis cow model, BMSCs were treated with 4.8 mM BHBA and NAHB for 12 h, respectively. The Western blot results revealed that BHBA significantly inhibited the expression of MYOG, MYOD1, MYH7, and GAS7, while NAHB, except for inhibiting the expression of MYOG, had no significant effect on the protein expression of other genes (Figure 1B, C). The qPCR results show that BHBA inhibits the differentiation of BMSCs, promotes the expression of LNC297, and inhibits the expression of GAS7, while NAHB shows the opposite results. Therefore, this result suggests that BHBA is more suitable for further progressed, the expression of differentiation marker genes first increased and then decreased, with the best differentiation observed on day 3. The qPCR results indicated that GAS7 and LNC297 were positively correlated with BMSCs differentiation, while novel-miR-145 was negatively correlated with differentiation (Figure 1E-G). To further explore the effect of BHBA on BMSCs differentiation, BHBA concentrations of 0 mM, 1.2 mM, 2.4 mM, and 4.8 mM were applied to BMSCs for12 h on the third day of differentiation. The results showed that the higher the concentration, the more significant the inhibitory effect of BHBA on BMSCs differentiation. Additionally, LNC297, GAS7, andnovel-miR-145 also exhibited dose-dependent effects (Figure 1H-J). The above results suggest that BHBA inhibits BMSCs differentiation in a dose-dependent manner, and GAS7, LNC297, and novel-miR-145 might influence BMSCs differentiation.
3.2. Novel-miR-145 Enhances the Inhibitory Effect of BHBA on BMSCs Differentiation
To further explore the function of novel-miR-145, the downstream target genes were predicted using the Targetscan and miRanda databases. A total of 597 target genes were predicted in both databases (Figure 2A). GO and KEGG analyses were performed on these 597 target genes. These target genes are located in the mitochondrial outer membrane, endoplasmic reticulum-Golgi intermediate compartment, organelle outer membrane, and other regions. They are involved in biological processes such as tRNA metabolic process, regulation of phospholipase activity, transcription by RNA polymerase III, ncRNA processing, and others. They possess molecular functions like oxidoreductase activity, acting on the CH-NH group of donors, lysine N-methyltransferase activity, and protein-lysine N-methyltransferase activity. KEGG results showed that these genes are significantly enriched in pathways such as Calcium signaling pathway, Hypertrophic cardiomyopathy, and Viral myocarditis (Figure 2B, C). The differentiation of muscle cells is an essential process for muscle tissue generation, ensuring the structural integrity, functional characteristics, and repair ability of muscles. To explore the role of novel-miR-145 in BMSCs differentiation, we transfected the mimic, mimic NC, Inhibitor, and Inhibitor NC ofnovel-miR-145 into BMSCs and then cultured them in differentiation medium for 3 days. The results showed that the expression of novel-miR-145 was significantly upregulated in the mimic group and significantly downregulated in the Inhibitor group, indicating successful transfection. Furthermore, when the expression of novel-miR-145 was increased, the expression of muscle differentiation marker genes decreased, while inhibiting its expression showed the opposite trend. Additionally, the expression of GAS7 was inversely correlated with the expression of novel-miR-145, further suggesting that GAS7 might be a downstream target gene of novel-miR-145 (Figure 2D, E, H). To further verify whether novel-miR-145 could inhibit the effect of BHBA on muscle cell differentiation, we added 4.8 mM BHBA to the Inhibitor group and co-treated for 12 hours. The results showed that when the expression of novel-miR-145 was inhibited, the inhibitory effect of BHBA on muscle cell differentiation was relieved (Figure 2F, G, I). These results suggest that novel-miR-145 inhibits BMSCs differentiation and enhances the inhibitory effect of BHBA on BMSCs differentiation. Moreover, GAS7 may be a downstream target gene of novel-miR-145.
3.3. LNC297 and GAS7 Alleviate the Inhibitory Effect of BHBA on the Differentiation of BMSCs
To explore the role of LNC297 in the differentiation of BMSCs, after overexpressingLNC297, the expression of differentiation marker genes was increased, indicating that LNC297promotes BMSCs differentiation. Furthermore, BHBA inhibited this differentiation, but after co-treatment with LNC297 overexpression and BHBA, differentiation-related genes were significantly upregulated, further suggesting that LNC297can regulate the myogenic differentiation process (Figure 3A, B). The expression of GAS7 increased during differentiation. To further investigate its impact on BMSCs differentiation, we used interfering RNA to reduce the expression of GAS7. WB results showed successful inhibition of GAS7 expression, while the expression of differentiation-related genes decreased, and differentiation was suppressed. After co-treatment with BHBA, the differentiation of BMSCs was further inhibited (Figure 3C-F). The qPCR results were consistent with the WB results. Additionally, after overexpressing LNC297, the expression level of novel-miR-145 was downregulated (Figure3G-I). The above results suggest that LNC297 and GAS7 promote BMSCs differentiation and can alleviate the inhibitory effect of BHBA on BMSCs differentiation, which might help alleviate muscle tissue damage in bovine ketosis. Moreover, LNC297 may regulate the expression of novel-miR-145 to exert its effect.
3.4. LNC297 Promotes the Differentiation of BMSCs Through the Novel-miR-145/GAS7 Pathway
The dual luciferase reporter assay confirmed the targeting relationship between novel-miR-145 and GAS7 (Figure 4A, B). The results of the miRNA pull-down experiment show that LNC297 targets and binds with novel-miR-145 (Figure 4C). Additionally, the dual luciferase reporter assay indicates that LNC297 has a targeting relationship with GAS7 (Figure 4D). To further verify the regulatory effect of novel-miR-145 on GAS7, co-transfection experiments with mimic and siGAS7 were performed. Western blot results show that when mimic and siGAS7 were co-transfected, the expression levels of MYOG, MYOD1, MYH7, and GAS7 were the lowest, suggesting that novel-miR-145 targets and inhibits the expression of GAS7, thereby suppressing the differentiation of BMSCs (Figure 4E, F). Similarly, the results of co-transfection of mimic and siGAS7 followed by 12-hour BHBA treatment were consistent with the above results (Figure 4G, H). To verify the mechanism of action of LNC297 and novel-miR-145, this study co-transfected overexpressed LNC297 (pc-LNC297) and the overexpression mimic of novel-miR-145 into BMSCs. The results showed that LNC297 could alleviate the inhibitory effect of novel-miR-145 on the differentiation of BMSCs. Additionally, LNC297 inhibited the expression of novel-miR-145 and promoted the expression of GAS7 (Figure 4 I-P). In conclusion, LNC297 can target and competitively bind to novel-miR-145 to promote the expression of GAS7, thereby promoting the differentiation of BMSCs and alleviating the inhibition of muscle differentiation under BHBA stress.
4. Discussion
During pregnancy and parturition—especially in late gestation and the early postpartum period-dairy cows often experience negative energy balance (NEB) due to insufficient energy intake and increased energy expenditure. During this time, cows mobilize their body fat reserves to meet their energy demands [28]. However, merely relying on fat mobilization for energy supply is insufficient to meet the energy demands of mammary gland development, fetal growth, early postpartum milk protein synthesis, and amino acid requirements. Skeletal muscle tissue, which serves as the most readily available and largest reservoir of amino acids, plays a vital role under conditions of protein and/or energy negative balance by providing amino acids that can be used for gluconeogenesis and energy production [29]. Previous studies have shown that during NEB, the muscle tissue weight of dairy cows’ decreases, indicating a reduction in muscle tissue production. This is primarily because cows with ketosis break down muscle tissue to provide the necessary amino acids and other substances [30,31]. skeletal muscle mobilization in transition dairy cows is relatively limited, and most studies have been conducted within feeding trials. In-depth investigations into the underlying mechanisms are still lacking [32]. BHBA is one of the ketone bodies metabolized in the liver as a result of the mobilization of fat tissue in cows with ketosis to compensate for insufficient energy intake. It is an important biochemical indicator for determining whether a cow has ketosis. BHBA plays a significant role in regulating follicular growth and exacerbating LPS-induced inflammation in bovine endometrial epithelial cells by activating the oxidative stress/NF-κB signaling pathway [33,34].
However, BHBA might serve as an energy substrate for oxidation in muscle tissue, but the specific mechanisms and whether it affects muscle formation are still unclear. To more accurately reflect the accumulation of BHBA in muscle in an in vitro model, this study uses BHBA as the experimental drug. Both the acidic form of BHBA and its sodium salt form. However, previous studies have shown that NAHB, as a metabolic substrate, might enhance the energy efficiency of myogenic cells by stimulating the expression of G protein-coupled receptor 109a (GPR109a), thereby upregulating 3-HB transportersMCT1 and CD147, utilizing the enzymeOXCT1 and phosphorylated AMPK to increase ATP production. As a signaling molecule, NAHB activates GPR109a, promoting calcium influx, improving calcium homeostasis, and increasing the expression of calcium-related proteins such as CAMKK2.This signaling process activates calcium-calmodulin-dependent phosphatase (CaN), promoting the translocation of NFAT to the cell nucleus and gene expression, thereby stimulating the proliferation and differentiation of myogenic cells. This is inconsistent with the metabolic pattern of ketosis in dairy cows [35]. The results of this study also confirm that NAHB promotes myocyte differentiation, consistent with previous BHBA treatment inhibits muscle differentiation, which aligns with the findings from our previous studies [11]. Therefore, BHBA is more suitable for exploring the metabolic pattern of 3-hydroxybutyricacid in the muscle tissue of cows with ketosis. Additionally, the low concentrations of BHBA (1.2mM-2.4mM) have little effect on the differentiation of BMSCs, possibly because low concentrations of BHBA are important metabolic products of ketone bodies, serving as an energy source for cells and providing ATP to support normal cellular function. Especially under conditions of glucose scarcity (such as during fasting or intense exercise), ketone bodies act as an alternative energy source, maintaining the cell's energy balance and thus not interfering with normal cellular differentiation processes. However, high concentrations of BHBA, especially when cows are affected by ketosis, may lead to metabolic acidosis, which is the accumulation of acidic substances in the blood. This acidosis inhibits the normal function of skeletal muscle satellite cells, hindering their differentiation. This metabolic stress causes damage to cells.
Muscle tissue constitutes 40% of bodyweight in mammals and plays a vital role in regulating metabolism and maintaining internal balance [36]. The formation of skeletal muscle begins with the determination of muscle cell fate, a process regulated by the myogenic transcription factors Pax7 and MYOD1. Subsequently, the expression of numerous genes establishes the structure and function of the muscle [37,38]. A key step in this process is the fusion of mononucleated muscle cells to form multinucleated muscle fibers. This study found that as BMSCs differentiate, the differentiation effect of MYH7, MYOD1, and MYOG is most prominent on day3. Therefore, day 3 of differentiation was chosen as the time point to investigate the mechanism of BHBA's effect on differentiation.
In addition, when damaged, muscle-derived progenitor cells in adult muscle tissue are activated and fuse to generate new muscle fibers [39]. Therefore, BHBA will affect the differentiation of muscle tissue in ketosis-affected cows. However, we speculate that after BHBA stress, the muscle tissue repair mechanisms may be initiated. For example, when muscle is damaged, satellite cells are activated and begin to proliferate. The proliferating satellite cells enter the cell cycle, differentiate into myoblasts, and subsequently fuse to form new muscle fibers, aiding in the repair of the damaged muscle tissue [40]. Most mammalian genomes transcribe a large amount of non-coding RNA (ncRNA), such as microRNA, circular RNA, and long non-coding RNA. Among these, long non-coding RNA is the most functionally diverse category and plays an important role in the regulation of development, differentiation, and maintenance of cell specificity [41]. For example, Lnc-MEG8 targets and binds to miR-22-3p, thereby reducing the inhibitory effects of miR-22-3p on the key myogenic factors MYF5 and MYOG, which promotes myotube formation [42]. Through previous studies, we identified an unannotated long non-coding RNA, LNC297, whose expression level increases during differentiation. Additionally, after BHBA treatment, the expression of LNC297 rises with the increasing concentration of BHBA, indicating its important role in the differentiation process of BMSCs and its potential involvement in regulating muscle formation in ketosis-affected cows. Furthermore, we found that overexpression of LNC297 significantly upregulates the expression levels of muscle differentiation marker genes such as MYOG, MYOD1, and MYH7. After combined treatment with BHBA, the expression of muscle differentiation-related genes still showed an upward trend, suggesting that LNC297 can promote BMSCs differentiation and partially alleviate the inhibitory effects of BHBA on muscle differentiation.
This study found that novel-miR-145 significantly inhibits the differentiation ability of BMSCs, and under conditions of co-treatment with BHBA, the differentiation of BMSCs is further inhibited, indicating that the two may have a synergistic inhibitory effect in regulating muscle differentiation. Existing research has shown that miR-145 plays an important role as a negative regulatory factor in various cell types, particularly in myogenesis, where miR-145 interferes with the process of cellular differentiation into muscle by targeting muscle-specific genes or affecting the expression of Myogenic Regulatory Factors (such as MYOD, MYOG) [43,44]. The novel-miR-145 newly discovered in this study belongs to the miR-145 family and is likely involved in regulating important biological processes such as cell proliferation, differentiation, and apoptosis through similar mechanisms. In particular, both miR-145 and novel-miR-145 may play analogous roles in the regulation of muscle cell differentiation, cancer biology, and the proliferation of vascular smooth muscle cells. In this study, it was found that the expression of novel-miR-145 is downregulated as differentiation progresses, and it also shows a downward trend under BHBA treatment. Therefore, it is initially speculated that novel-miR-145 inhibits BMSCs differentiation. The results from Western blotting and qPCR also confirmed that novel-miR-145 suppresses the expression of muscle differentiation marker genes, thereby inhibiting BMSCs differentiation. Furthermore, after co-treatment with BHBA, the expression of muscle differentiation-related genes further showed a downregulation trend, suggesting that novel-miR-145 might promote BMSCs differentiation and exacerbate the inhibitory effects of BHBA on muscle differentiation.
On the other hand, BHBA is one of the main products of ketone metabolism and plays a complex role in muscle metabolism and energy homeostasis. Although some studies have reported that BHBA can promote cell antioxidant capacity and survival under certain conditions, under the conditions of this study, BHBA, whether administered alone or in combination with novel-miR-145, exhibited an inhibitory effect on BMSCs differentiation. This suggests that BHBA might amplify the inhibitory effects mediated by novel-miR-145 by affecting metabolic pathways and interfering with muscle cell differentiation signals (such as mTOR, AMPK, Wnt). Notably, BHBA may regulate the expression or stability of miRNAs, or influence the efficiency of miRNA target gene expression by altering the intracellular metabolic state, thereby enhancing the functionality of novel-miR-145. This observation indicates a potential complementary or intersecting relationship between the two, providing new clues for understanding the coordinated regulation of muscle cell fate by miRNAs and metabolites. However, the specific mechanisms of action still require further experimental validation.
miRNA exerts its functions through various mechanisms. Typically, miRNAs inhibit gene expression by binding to the 3' UTR of target mRNAs. This interaction involves specific complementary pairing between the miRNA and its target mRNA, often centered around a particular "seed sequence." Such binding can lead to the repression of mRNA translation or its degradation, ultimately influencing the protein products and thereby regulating biological processes including cell differentiation, apoptosis, and proliferation [45]. In this study, the online prediction tools TargetScan and miRanda were utilized to identify the target genes of novel-miR-145. Subsequently, GO and KEGG analyses were conducted to perform functional enrichment analysis of these target genes, allowing us to infer the potential functions of novel-miR-145. The results indicated that these target genes were significantly enriched in pathways such as Hypertrophic Cardiomyopathy, Arrhythmogenic Right Ventricular Cardiomyopathy, Calcium Signaling Pathway, and Viral Myocarditis. These pathways have been recognized as being critical for muscle cell differentiation [46,47]. Additionally, this study identified a key target gene, GAS7, which is involved in the PI3K/AKT signaling pathway, a pathway closely associated with muscle development [48,49]. The results of this study indicate that the overexpression of novel-miR-145 leads to a decrease in both the mRNA and protein levels of GAS7. Furthermore, the dual-luciferase assay revealed that there is a targeted relationship between novel-miR-145 and GAS7. Additionally, Western blot and quantitative results demonstrated that GAS7 promotes the differentiation of BMSCs. Therefore, based on these findings, we conclude that novel-miR-145 exerts an inhibitory effect on the differentiation of BMSCs by directly targeting GAS7.
In recent years, an increasing body of evidence has shown that long non-coding RNAs (lncRNAs), as ceRNAs interacting with microRNAs (miRNAs), play a significant role in the development and differentiation of muscle and fat by regulating the expression of their target mRNAs [50]. The interaction between lncRNA, miRNA, and mRNA plays acritical role in various biological processes during development and disease. It has been recognized as a common regulatory mechanism in the functional control of lncRNAs [51,52]. The results of this study indicate that LNC297 binds to novel-miR-145 and inhibits its expression, leading to the upregulation of the GAS7 gene, thereby regulating the differentiation of BMSCs. This study also explored the targeted relationship between LNC297 and GAS7; however, whether LNC297 directly regulates GAS7 under BHBA stress to further alleviate the inhibitory effects on muscle differentiation still requires further experiments for validation.
5. Conclusions
In summary, this study established that BHBA can inhibit the differentiation of BMSCs and developed an in vitro BHBA stress model, providing a theoretical basis for exploring the developmental mechanisms of muscle tissue in ketosis cows. Additionally, it identified LNC297 as a key factor in BMSCs differentiation that can alleviate the impairment of muscle differentiation under high BHBA stress. This research is the first to investigate the overexpression of LNC297 in BMSCs to promote differentiation, effectively enhancing the muscle differentiation process when co-treated with BHBA. Furthermore, it revealed a novel interaction between LNC297 and novel-miR-145. We provided evidence that LNC297 regulates the expression of GAS7 in BMSCs by acting as a sponge for novel-miR-145, thereby exerting its regulatory effects. This study enriches the understanding of long non-coding RNAs and competitive endogenous RNAs in the development of BMSCs and offers new targets for the treatment of ketosis cows and other metabolic syndromes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This research was funded by the Sichuan innovation team of national modern agricultural industry technology system, grant number SCCXTD-2026-13; Sichuan key research and development project on innovative breeding materials, methods, and new breeds of high-quality cattle, Key R&D Program of Sichuan Province, grant number 2021YFYZ0001; Major Livestock and Poultry Germplasm Resources (Breeding Breakthrough Project), Key R&D Program of Sichuan Province, grant number 2021YFYZ0007.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Institution Review Board Statement
All animal experiments in the present study involving animals were performed under the direction of the Institutional Animal Care and Use Committee from the College of Animal Science and Technology, Sichuan Agricultural University, China. (Certification No. SYXK2019-187; Approval time: 2019/1/29).
Abbreviations
| BHBA | β-hydroxybutyrate |
| NAHB | Sodium β-hy297. |
| BMSCs | Bovine skeletal muscle satellite cells |
| NEB | Negative energy balance |
| LNC297 | LncRNA MSTRG 29792.14 |
| ceRNA | Competing endogenous RNA |
| lncRNAs | Long non-coding RNAs |
| NEFA | Non-esterified fatty acids |
References
- Bell A W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation[J]. J Anim Sci, 1995, 73(9): 2804-2819.
- Adewuyi A A, Gruys E, Van Eerdenburg F J. Non esterified fatty acids (NEFA) in dairy cattle. A review[J]. Vet Q, 2005, 27(3): 117-126. [CrossRef]
- Kolb H, Kempf K, Röhling M, et al. Ketone bodies: from enemy to friend and guardian angel[J]. BMC Med, 2021, 19(1): 313.
- Chen L, Deng H, Cui H, et al. Inflammatory responses and inflammation-associated diseases in organs[J]. Oncotarget, 2018, 9(6): 7204-7218.
- Møller N. Ketone Body, 3-Hydroxybutyrate: Minor Metabolite - Major Medical Manifestations[J]. J Clin Endocrinol Metab, 2020, 105(9): dgaa370.
- Zhang Y, Li Z, Liu X, et al. 3-Hydroxybutyrate ameliorates insulin resistance by inhibiting PPARγ Ser273 phosphorylation in type 2 diabetic mice[J]. Signal Transduct Target Ther, 2023, 8(1): 190. [CrossRef]
- Duffield T F, Lissemore K D, Mcbride B W, et al. Impact of hyperketonemia in early lactation dairy cows on health and production[J]. J Dairy Sci, 2009, 92(2): 571-580.
- Palmer B F, Clegg D J. Starvation Ketosis and the Kidney[J]. Am J Nephrol, 2021, 52(6): 467-478.
- Sun X, Zhang B, Sun K, et al. Liver-Derived Ketogenesis via Overexpressing HMGCS2 Promotes the Recovery of Spinal Cord Injury[J]. Adv Biol (Weinh), 2024, 8(2): e2300481.
- Dong Z, Sun X, Tang Y, et al. β-hydroxybutyrate impairs monocyte function via the ROS-NLR family pyrin domain-containing three inflammasome (NLRP3) pathway in ketotic cows[J]. Front Vet Sci, 2022, 9: 925900.
- Tang T, Zhou J, Shao J, et al. Complementary Muscle Metabolomics and Proteomics of Muscle in Cows With Post-Calving Ketosis[J]. J Vet Intern Med, 2025, 39(6): e70214. [CrossRef]
- Tan Y T, Lin J F, Li T, et al. LncRNA-mediated posttranslational modifications and reprogramming of energy metabolism in cancer[J]. Cancer Commun (Lond), 2021, 41(2): 109-120.
- Zhang W, Zhao J, Deng L, et al. INKILN is a Novel Long Noncoding RNA Promoting Vascular Smooth Muscle Inflammation via Scaffolding MKL1 and USP10[J]. Circulation, 2023, 148(1): 47-67.
- Yu J, Wang W, Yang J, et al. LncRNA PSR Regulates Vascular Remodeling Through Encoding a Novel Protein Arteridin[J]. Circ Res, 2022, 131(9): 768-787. [CrossRef]
- Hernández-Hernández J M, García-González E G, Brun C E, et al. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration[J]. Semin Cell Dev Biol, 2017, 72: 10-18. [CrossRef]
- Bailey P, Holowacz T, Lassar A B. The origin of skeletal muscle stem cells in the embryo and the adult[J]. Curr Opin Cell Biol, 2001, 13(6): 679-689.
- Braun T, Gautel M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis[J]. Nat Rev Mol Cell Biol, 2011, 12(6): 349-361.
- Naya F J, Olson E. MEF2: a transcriptional target for signaling pathways controlling skeletal muscle growth and differentiation[J]. Curr Opin Cell Biol, 1999, 11(6): 683-688.
- Sun W, Yang Y, Xu C, et al. Regulatory mechanisms of long noncoding RNAs on gene expression in cancers[J]. Cancer Genet, 2017, 216-217: 105-110. [CrossRef]
- Xu X, Ji S, Li W, et al. LncRNA H19 promotes the differentiation of bovine skeletal muscle satellite cells by suppressing Sirt1/FoxO1[J]. Cell Mol Biol Lett, 2017, 22: 10.
- Chen K, Rajewsky N. The evolution of gene regulation by transcription factors and microRNAs[J]. Nat Rev Genet, 2007, 8(2): 93-103. [CrossRef]
- Sun K, Lai E C. Adult-specific functions of animal microRNAs[J]. Nat Rev Genet, 2013, 14(8): 535-548.
- Zhou S, Li S, Zhang W, et al. MiR-139 promotes differentiation of bovine skeletal muscle-derived satellite cells by regulating DHFR gene expression[J]. J Cell Physiol, 2018, 234(1): 632-641. [CrossRef]
- Song C, Yang Z, Jiang R, et al. lncRNA IGF2 AS Regulates Bovine Myogenesis through Different Pathways[J]. Mol Ther Nucleic Acids, 2020, 21: 874-884. [CrossRef]
- Cheng X, Li L, Shi G, et al. MEG3 Promotes Differentiation of Porcine Satellite Cells by Sponging miR-423-5p to Relieve Inhibiting Effect on SRF[J]. Cells, 2020, 9(2): 449.
- Liu M, Li B, Peng W, et al. LncRNA-MEG3 promotes bovine myoblast differentiation by sponging miR-135[J]. J Cell Physiol, 2019, 234(10): 18361-18370. [CrossRef]
- Delort F, Segard B D, Hakibilen C, et al. Alterations of redox dynamics and desmin post-translational modifications in skeletal muscle models of desminopathies[J]. Exp Cell Res, 2019, 383(2): 111539.
- Valergakis G E, Oikonomou G, Arsenos G, et al. Phenotypic association between energy balance indicators and reproductive performance in primiparous Holstein cows[J]. Vet Rec, 2011, 168(7): 189.
- Schäff C, Börner S, Hacke S, et al. Increased muscle fatty acid oxidation in dairy cows with intensive body fat mobilization during early lactation[J]. J Dairy Sci, 2013, 96(10): 6449-6460.
- Hatfield C, Tulley W, Hall R, et al. A Prospective Cohort Study on the Periparturient Muscle Tissue Mobilisation in High Producing Dairy Cows[J]. Animals (Basel), 2022, 12(14): 1772. [CrossRef]
- Bell A W, Burhans W S, Overton T R. Protein nutrition in late pregnancy, maternal protein reserves and lactation performance in dairy cows[J]. Proc Nutr Soc, 2000, 59(1): 119-126.
- Doepel L, Lapierre H, Kennelly J J. Peripartum performance and metabolism of dairy cows in response to prepartum energy and protein intake[J]. J Dairy Sci, 2002, 85(9): 2315-2334.
- Missio D, Fritzen A, Cupper Vieira C, et al. Increased β-hydroxybutyrate (BHBA) concentration affect follicular growth in cattle[J]. Anim Reprod Sci, 2022, 243: 107033. [CrossRef]
- Yang W, Wang F, Liu J, et al. β-Hydroxybutyrate aggravates LPS-induced inflammatory response in bovine endometrial epithelial cells by activating the oxidative stress/NF-κB signaling pathway[J]. Int Immunopharmacol, 2025, 154: 114609.
- Qiu X, Wu W, Zhang S, et al. 3-Hydroxybutyrate Promotes Myoblast Proliferation and Differentiation through Energy Metabolism and GPR109a-Mediated Ca(2+)-NFAT Signaling Pathways[J]. J Proteome Res, 2025, 24(4): 2063-2080.
- Liu L, Ding C, Fu T, et al. Histone methyltransferase MLL4 controls myofiber identity and muscle performance through MEF2 interaction[J]. J Clin Invest, 2020, 130(9): 4710-4725. [CrossRef]
- Weintraub H, Davis R, Tapscott S, et al. The myoD gene family: nodal point during specification of the muscle cell lineage[J]. Science, 1991, 251(4995): 761-766.
- Seale P, Sabourin L A, Girgis-Gabardo A, et al. Pax7 is required for the specification of myogenic satellite cells[J]. Cell, 2000, 102(6): 777-786.
- Fang J, Sia J, Soto J, et al. Skeletal muscle regeneration via the chemical induction and expansion of myogenic stem cells in situ or in vitro[J]. Nat Biomed Eng, 2021, 5(8): 864-879. [CrossRef]
- Yang F, Chi R, Zhang R, et al. A Novel ceRNA Axis LOC121818100/Novel-miR-400/SSRP1 Regulated Muscle Growth and Injury Repair in Sheep[J]. J Cachexia Sarcopenia Muscle, 2025, 16(3): e13836. [CrossRef]
- Neguembor M V, Jothi M, Gabellini D. Long noncoding RNAs, emerging players in muscle differentiation and disease[J]. Skelet Muscle, 2014, 4(1): 8.
- Ma X, La Y, Wang T, et al. Lnc-MEG8 regulates yak myoblast differentiation via the miR-22-3p/RTL1 axis[J]. BMC Genomics, 2024, 25(1): 1146.
- Yeh Y T, Wei J, Thorossian S, et al. MiR-145 mediates cell morphology-regulated mesenchymal stem cell differentiation to smooth muscle cells[J]. Biomaterials, 2019, 204: 59-69.
- Wang Y S, Li S H, Guo J, et al. Role of miR-145 in cardiac myofibroblast differentiation[J]. J Mol Cell Cardiol, 2014, 66: 94-105.
- Szatkowski C, Vallet J, Dormishian M, et al. Prokineticin receptor 1 as a novel suppressor of preadipocyte proliferation and differentiation to control obesity[J]. PLoS One, 2013, 8(12): e81175. [CrossRef]
- Qiu Y, Xu J, Chen Y, et al. Parkin plays a crucial role in acute viral myocarditis by regulating mitophagy activity[J]. Theranostics, 2024, 14(13): 5303-5315. [CrossRef]
- Field J T, Chapman D, Hai Y, et al. The mitophagy receptor BNIP3L/Nix coordinates nuclear calcium signaling to modulate the muscle phenotype[J]. Autophagy, 2025, 21(7): 1544-1555.
- Liu W F, Zhang Q W, Quan B, et al. Gas7 attenuates hepatocellular carcinoma progression and chemoresistance through the PI3K/Akt signaling pathway[J]. Cell Signal, 2023, 112: 110908.
- Li Y, Liu Z, Yan H, et al. Polygonatum sibiricum polysaccharide ameliorates skeletal muscle aging and mitochondrial dysfunction via PI3K/Akt/mTOR signaling pathway[J]. Phytomedicine, 2025, 136: 156316. [CrossRef]
- Song C, Wang J, Ma Y, et al. Linc-smad7 promotes myoblast differentiation and muscle regeneration via sponging miR-125b[J]. Epigenetics, 2018, 13(6): 591-604. [CrossRef]
- Liu X, Hou L, Huang W, et al. The Mechanism of Long Non-coding RNA MEG3 for Neurons Apoptosis Caused by Hypoxia: Mediated by miR-181b-12/15-LOX Signaling Pathway[J]. Front Cell Neurosci, 2016, 10: 201.
- Kong L, Zhuo Y, Xu J, et al. Identification of long non-coding RNAs and microRNAs involved in anther development in the tropical Camellia oleifera[J]. BMC Genomics, 2022, 23(1): 596. [CrossRef]
Figure 1.
BHBA inhibits the differentiation of BMSCs. (A) Immunofluorescence staining of BMSCs. microscopy images showing the localization of Desmin (green) in BMSCs. The nuclei were counterstained with DAPI (blue) (100×). (B, C) On the third day of BMSCs differentiation, 12 h after treatment with 4.8 mM BHBA and NAHB, Western blot analysis and gray value detection were conducted (n=3). (D, G and J) The relative expression levels of GSA7, novel-miR-145, LNC297 and myogenic differentiation-related genes (n=9). (E, F) Western blot analysis and gray value detection were conducted (n=3). (H, I) Western blot analysis and gray value detection were conducted (n=3). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
Figure 1.
BHBA inhibits the differentiation of BMSCs. (A) Immunofluorescence staining of BMSCs. microscopy images showing the localization of Desmin (green) in BMSCs. The nuclei were counterstained with DAPI (blue) (100×). (B, C) On the third day of BMSCs differentiation, 12 h after treatment with 4.8 mM BHBA and NAHB, Western blot analysis and gray value detection were conducted (n=3). (D, G and J) The relative expression levels of GSA7, novel-miR-145, LNC297 and myogenic differentiation-related genes (n=9). (E, F) Western blot analysis and gray value detection were conducted (n=3). (H, I) Western blot analysis and gray value detection were conducted (n=3). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
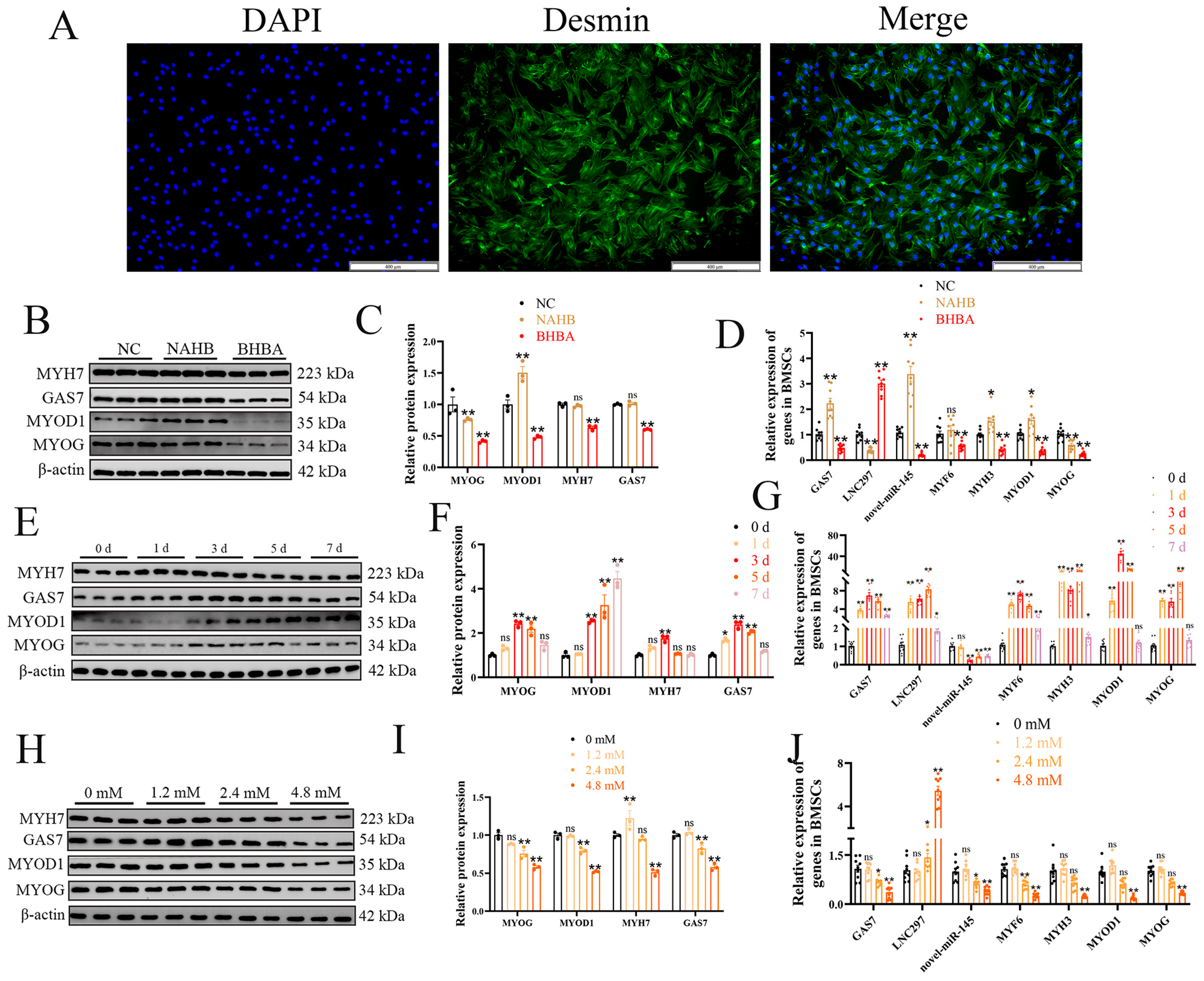
Figure 2.
novel-miR-145 enhances the inhibitory effect of BHBA on BMSCs differentiation. (A) Venn diagram of the number of target gene predictions from the Targetscan and miRanda databases. (B) GO analysis. (C) KEGG analysis of the differentially expressed genes only listed significant enrichment pathways. (D, E) After overexpressing and inhibiting the expression of novel-miR-145, WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (F, G) After inhibiting the expression of novel-miR-145, and co-treating with BHBA for 12 h, WB analysis of GAS7 and differentiation-related genes and grayscale value detection (n=3). (H, I) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
Figure 2.
novel-miR-145 enhances the inhibitory effect of BHBA on BMSCs differentiation. (A) Venn diagram of the number of target gene predictions from the Targetscan and miRanda databases. (B) GO analysis. (C) KEGG analysis of the differentially expressed genes only listed significant enrichment pathways. (D, E) After overexpressing and inhibiting the expression of novel-miR-145, WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (F, G) After inhibiting the expression of novel-miR-145, and co-treating with BHBA for 12 h, WB analysis of GAS7 and differentiation-related genes and grayscale value detection (n=3). (H, I) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
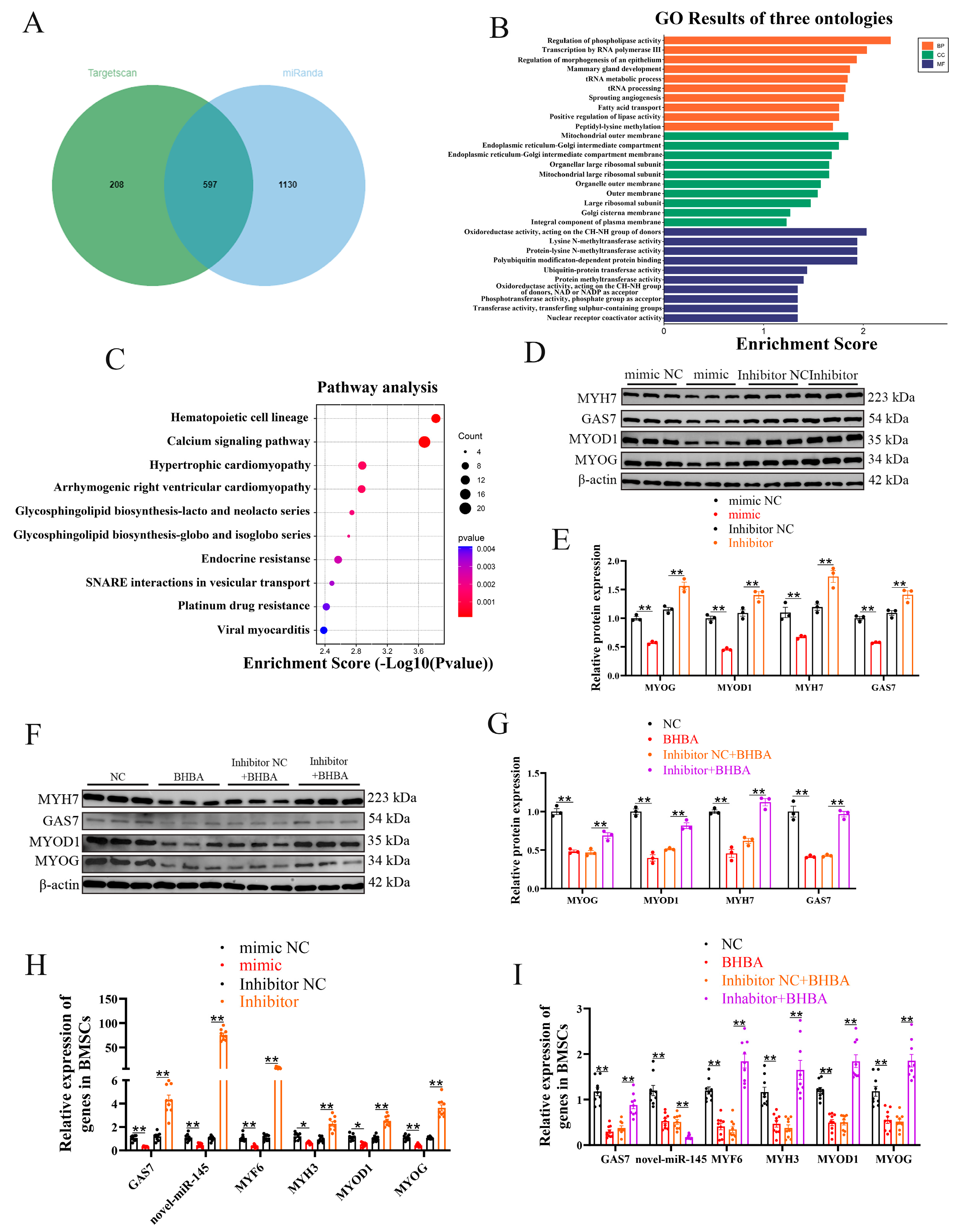
Figure 3.
LNC297 and GAS7 alleviate the inhibitory effect of BHBA on the differentiation of BMSCs. (A-F) WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (G-I) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
Figure 3.
LNC297 and GAS7 alleviate the inhibitory effect of BHBA on the differentiation of BMSCs. (A-F) WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (G-I) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
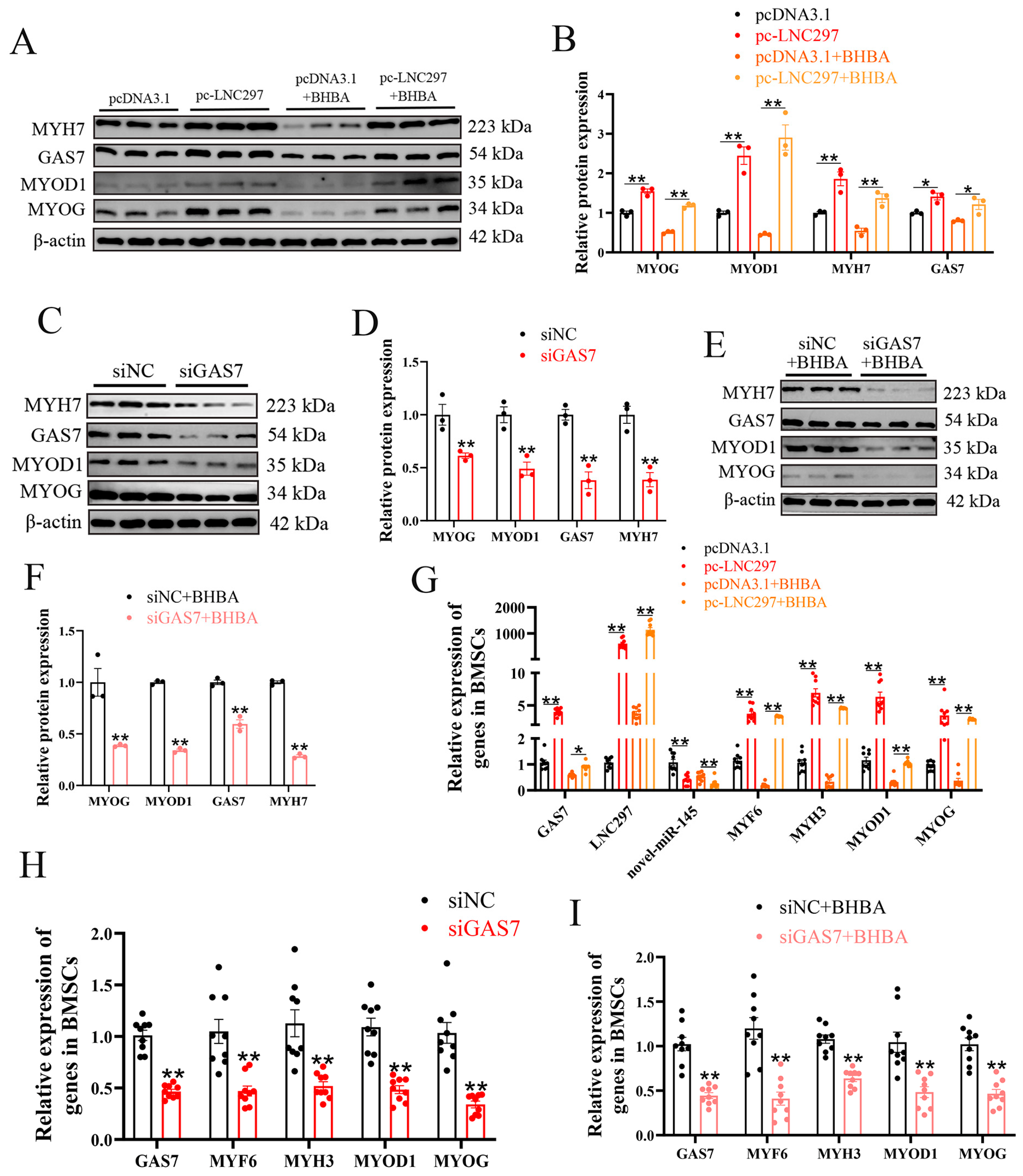
Figure 4.
LNC297 promotes the differentiation of BMSCs through the novel-miR-145/GAS7 pathway. (A) The predicted binding site of gene GAS7 with novel-miR-145. (B) Luciferase assay was conducted by co-transfecting GAS7 WT and mutant plasmids with novel-miR-145 mimic and NC into 293T cells, with the WT + NC group serving as the control (n = 6). (C) The miRNA pull-down experiment results show that LNC297 binds to novel-miR-145. Biotin-labeled novel-miR-145 or control miRNA (mimic NC) was transfected into BMSCs, followed by affinity capture and PCR detection of LNC297 expression. (D) Luciferase assay was conducted by co-transfecting GAS7 WT and mutant plasmids with pc-LNC297 and pcDNA3.1 into 293T cells, with the WT + NC group serving as the control (n = 6). (E-L) WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (M-P) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
Figure 4.
LNC297 promotes the differentiation of BMSCs through the novel-miR-145/GAS7 pathway. (A) The predicted binding site of gene GAS7 with novel-miR-145. (B) Luciferase assay was conducted by co-transfecting GAS7 WT and mutant plasmids with novel-miR-145 mimic and NC into 293T cells, with the WT + NC group serving as the control (n = 6). (C) The miRNA pull-down experiment results show that LNC297 binds to novel-miR-145. Biotin-labeled novel-miR-145 or control miRNA (mimic NC) was transfected into BMSCs, followed by affinity capture and PCR detection of LNC297 expression. (D) Luciferase assay was conducted by co-transfecting GAS7 WT and mutant plasmids with pc-LNC297 and pcDNA3.1 into 293T cells, with the WT + NC group serving as the control (n = 6). (E-L) WB analysis of GAS7 and differentiation-related genes, as well as grayscale value detection (n=3). (M-P) The relative expression levels of GSA7, novel-miR-145 and myogenic differentiation-related genes (n=9). The data presented as means ± SEM. * P < 0.05; ** P < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



