
Submitted:
21 January 2026
Posted:
22 January 2026
You are already at the latest version
Abstract
Plants exhibit complex internal dynamics in response to environmental conditions, yet whether these dynamics reflect structured affective regimes remains unclear. This study investigates whether internal plant signals encode information about affective states defined relationally by sustained environmental conditions. Valence and arousal were operationalised using temperature, humidity, and residualised light. Using only internal plant measurements—including bioelectrical activity and volatile gas emissions—we evaluated whether machine learning models could decode affective structure without access to environmental variables. Binary classification revealed that valence was reliably decoded over longer temporal windows, whereas arousal required shorter windows, suggesting distinct underlying timescales. Direct multiclass quadrant classification proved unstable, but an Echo State Network capturing temporal dependencies achieved improved performance. These results indicate that plant internal dynamics carry a learnable, temporally extended signature of environmentally defined affective regimes, supporting an interpretation of plant affect as embodied environmental engagement.
Keywords:
plants
; affective science
; neural network
1. Introduction
Plants silently compute the world’s crises (drought, toxins, climate shifts) through complex processes. Unlike electronic sensors, which provide direct streams of data, plants integrate diverse signals into emergent, nonlinear responses that encode more than the sum of their inputs. Interpreting these responses may allow us to make more holistic decisions about how we act in the world, ultimately guiding choices that benefit both humans and our environment.
The mechanisms by which plants experience and communicate environmental change through their own umwelt are increasingly well understood. Traits such as phototropism, chemical detection, and bioelectrical signaling represent deeply conserved strategies for sensing and adapting to environmental variability [1]. These processes manifest as measurable bioelectrical response, respiratory changes, and even physical motions that together form the basis of phytosensing [2,3]. Recent work has demonstrated that plant bioelectrical signals can be used to classify both environmental conditions and affective states in humans, suggesting that plant physiological dynamics encode rich, interpretable information beyond simple stress responses [4,5].
Plants continuously sense and respond to their environment through coupled electrical, physiological, and metabolic processes. Changes in light, temperature, and humidity effects electrical potentials across membranes, stomatal conductance, and respiration-photosynthesis balance, producing measurable signals in both bioelectrical activity and respiratory outputs [6,7,8]. These responses are increasingly understood not as isolated stress reactions, but as components of integrated signaling systems that coordinate whole-plant regulation across spatial and temporal scales [9,10].
In affective science, emotional states are commonly represented within a low-dimensional space defined by valence (positive–negative) and arousal (low–high activation) [11]. This dimensional framework can be repurposed as an operational description of internal state. Under this interpretation, valence corresponds to the relative favourability of environmental conditions for plant function, while arousal corresponds to the intensity of physiological activation required to respond to those conditions [12].
In this study, we investigate whether changes in a plant’s internal physiological state, specifically bioelectrical activity and respiration-related gas proxies, can be consistently mapped to externally defined environmental conditions framed within an emotional space. We focus on a tomato plant (Solanum lycopersicum) grown within a controlled chamber in which light, temperature, and humidity are actively manipulated. Internal state is monitored via sensors: electrical signals acquired through electrodes, and metabolic proxies measured as environmental CO2 (eCO2) and total volatile organic compounds (TVOC).
Rather than relying solely on statistics, we implement a recurrent computational readout in the form of an Echo State Network (ESN). The ESN is used both to (i) infer the plant’s current emotion-like state from environmental inputs and physiological signals, and (ii) predict future internal state trajectories under ongoing environmental conditions. This approach treats the plant–environment system as a dynamical process and tests whether a predictive internal representation of plant state emerges.
1.1. Plant Electrophysiology and Internal State
Plant cells maintain membrane potentials through ion transport, enabling the generation of electrical signals in response to environmental changes [13]. These include fast action potentials and slower variation potentials, both of which have been observed in response to changes in light, temperature, humidity, and mechanical disturbance [6,7]. These signals occur through vascular and cellular pathways, allowing for coordination of physiological responses [14].
Bioelectrical activity has therefore been proposed as a candidate indicator of internal plant state, reflecting both immediate stimulus detection and longer-timescale regulatory adjustments [10]. However, electrical signals alone are difficult to interpret without complementary context as similar physiological changes can arise from different drivers [15]. Therefore, changes in processes such as bioelectrical activity is best interpreted not as a direct encoding of specific stimuli, but as part of a malleable internal state shaped by environmental history and experience.
1.2. Respiration and Volatile Emissions as Metabolic Indicators
Environmental conditions also affect plant respiration and metabolism. Temperature influences respiratory rates, while humidity affects transpiration and stomatal behaviour, therefore shaping internal CO2 dynamics [8]. Plants also emit volatile organic compounds (VOCs) whose production can increase under stressors such as high light and elevated temperature, as well as under biotic challenges [16,17]. These signals evolve on slower timescales than electrical responses, complementing bioelectrical measurements.
In this work, eCO2 and TVOC are measured using a gas sensor. Within a controlled chamber they provide reproducible indicators of metabolic and chemical changes associated with environmental transitions.
1.3. Valence–Arousal as an Operational Framework
The valence–arousal model provides a common and interpretable means of describing complex affective states [11]. Applied here, valence tells us the ’favourability’ of an environmental condition for tomatoes (e.g. high humidity reducing water stress), while arousal shows the ’degree of physiological activation’ (e.g. elevated temperature increasing stress) [12]. In this study, valence and arousal are defined exclusively by environmental conditions, and are used as reference labels against which internal physiological dynamics are correlated.
1.4. Reservoir Computing as a Framework
Reservoir computing is a computational paradigm in which a dynamical system transforms input signals into a high-dimensional state space. A simple readout function is then trained to map these states onto desired outputs. In this study, an Echo State Network (ESN) is employed as the readout model to infer affective state from plant physiological data. The ESN state update can be shown as:
where is the time-based input, is the internal reservoir state, and W are the input and recurrent weights, the leak rate, and a nonlinear activation function.
In this work, the plant itself is not treated as a reservoir necessarily. Its physiological dynamics constitute a process whose internal states are captured through sensors and projected into a Echo State Network readout for inference [18]. The inputs therefore correspond to internal plant signals that reflect the plant’s relationship to environmental conditions.
An ESN was chosen based on the potential relationship dynamics of the system and the environment. Firstly, the physiological signals recorded from the plant are continuous data, temporally structured, and exhibit history-dependent responses to environmental changes. ESN’s are designed to retain short-term memory of inputs based on their recurrent dynamics which allows for sequence decoding and correlation [19]. Secondly, the relationship between the plant and its environment is potentially nonlinear. This means that the retention of recurrent weights allows for linear labelling to be placed on the output and readout layer [20]. Thirdly, ESN’s are robust to noise which occurs in biological sensing over long time periods [21].
The readout layer is trained to map reservoir states onto discrete affective states in valence–arousal space, enabling inference of the plant’s current affective state from its internal physiological dynamics:
where represents predicted conditions (e.g., changes in affective state) that are subsequently mapped onto dimensional emotion (e.g., valence or arousal state). Reservoir computing is particularly suited to biological time series data because it requires minimal training of internal dynamics, tolerates noise, and can exploit the natural complexity of physical systems [22,23,24,25]. Plants, as dynamical substrates, have these qualities: high-dimensional responses, nonlinear coupling, and environmental sensitivity.
1.5. Phytosensing & Discovery
Previous research has demonstrated that plants can act as environmental biosensors [2,26]. Their electrical, thermal, and morphological responses provide high-dimensional encodings of ambient conditions. Beyond electrophysiology, phytosensing encompasses stomatal conductance, volatile organic compound release, and photosynthetic fluorescence [27]. These multi-modal signals encode integrative states that are difficult to reduce to single abiotic variables, but which hold potential as ecological readouts of complex conditions.
1.5.1. Phytosensing in Posthumanism
From a posthumanist perspective, plants are not passive data sources but active mediators of environmental information. This work builds on frameworks by [28,29], and [30], which emphasise relationality, distributed agency, and the decentring of human primacy. Here, the plant is not merely instrumentalised; instead, the sensing system is conceptualised as a hybrid assemblage of plant, human, and machine, in which each participant contributes to environmental cognition.
While research in plant electrophysiology has demonstrated that plants register and encode environmental perturbations through electrical, hydraulic, and morphological signals [2,3,26,31], such studies have largely remained within the domain of physiological measurement and bio-inspired computation. However, a growing body of scholarship in science, technology and environmental humanities has begun to reconceptualise sensing itself as a distributed and more-than-human process. [32] has described how their study Program Earth develops infrastructure that embed environmental monitoring into living systems, while [33] experiments with plants as sensing partners to explore entangled modes of perception. Contemporary work in plant cognition has argued that plants exhibit integrative, memory-like, and adaptive signaling dynamics that warrant investigation using frameworks usually reserved for cognitive systems [34]. These perspectives challenge the framing of plants as passive biosensors. By bringing these literatures together, we position our study at the intersection of physiological science and posthumanist theory: the plant is both a reservoir computer encoding environmental dynamics and an active participant in a the ecology of sensing.
2. Materials and Methods
2.1. Proposed Approach
Our approach builds on plant electrophysiology and reservoir computing [2,26], while also engaging with more-than-human sensing frameworks [32,33], situating plants not only as data sources but as active participants in sensing assemblages. Our study employs bioelectrical and respiratory sensors to capture plant state trajectories. These are processed through reservoir computing models to infer the affective state of the subject plant, and make predictions as to the affective state based on known environmental conditions. The study does not treat this prediction as a passive output: instead, the readout layer is a site of translation between plant computation and human decision-making, embedding posthumanist principles into a practical sensing methodology.
2.2. Overview
A single tomato plant (Solanum lycopersicum) was monitored within a controlled chamber made for environmental sensing and bioelectrical recording. Environmental variables were actively manipulated using a heating pad (1 - B) and fans (1 - G), while internal plant signals were recorded continuously. All data was logged using a Raspberry Pi with environmental and bioelectrical sensors.
2.3. Environmental Sensing
Environmental variables were sampled at 0.5Hz, a rate quick enough to capture chamber dynamics. All sensors communicated via I2C to a Raspberry Pi 5, and pictured in Figure 1 and Figure 1:
- Light and infrared: LTR303 sensor positioned at base height (1 - D).
- Temperature and humidity: HDC302 sensor located near the plant stem at midheight (1- E).
- Gas sensors: SGP30 sensor measuring eCO2 and TVOC within the chamber (1 - C).
2.4. Biosensing
As well as environmental sensors, bioelectricty was measured via differential electrodes. An ADS1115 module was used for bioelectrical signals. Electrodes where placed in the plant tissue itself. The live electrode was placed near the canopy of the plant, with the ground being inserted more basally. Signals were sampled at 100Hz. Although slower than rates used in some electrophysiological studies, this frequency was sufficient to capture electrical fluctuations associated with physiological regulation while maintaining the ability to record over long durations [4,35].
The chamber was set near a window in December and January for circadian rhythm, and was subject to ambient temperatures that were optimal for tomato plant growth during the day’s the office was occupied (approx. 25 °C). The heater was turned on at random intervals to eliminate the potential for rhythmically primed response to temperature increases. The heater was on for between 15 and 45 minutes, with the length of time also being randomly assigned programmatically. If the temperature of the chamber reached a critical 36 °C level fans where instantiated to bring the temperature down while maintaining above 35 °C to ensure a heat stressful environment. Fans also activate every 24 hours to flux fresh air into the chamber to protect from over-saturation of VOC’s. Watering occurred once a week, where the chamber lid was removed during a period where the fans and heater were not engaged. The soil was then well saturated with water.
2.5. Valence and Arousal
Valence and arousal were determined by the environmental conditions of the chamber. It was determined that humidity was the main positive driver in the valence direction, whereby higher humidity resulted in higher valence, and lower arousal. Temperature was the positive driver in the arousal direction, whereby higher temperature resulted in high arousal and lower valence. Light was also taken into account. Light was determined to be a positive driver for both valence and arousal. However, due to the nature of light in the chamber, whereby light has the effect of increasing temperature and decreasing humidity, the residual effect of light after temperature and humidity’s effect on valence and arousal had been calculated was used.
Environmental variables were first normalised using z-scoring. Residualised light was obtained via linear regression of light against temperature and humidity:
where , , and are regression coefficients. The residual light was then defined as:
then represents light that is statistically independent of temperature and humidity within the chamber.
Valence V and arousal A were then defined as linear combinations of the residual light, temperature, and humidity:
With these equations, valence increases with residual light and humidity and decreases with temperature, while arousal increases with residual light and temperature and decreases with humidity. This structure reflects both the empirical coupling of environmental variables in the chamber and the intended interpretation of valence and arousal as orthogonal affective dimensions defined by environmental factors rather than inferred from internal signals.
Continuous valence and arousal signals were converted into binary affective labels to enable classification-based decoding. Valence and arousal signs were defined as
where denotes the indicator function. This yields four affective quadrants corresponding to the Cartesian product of valence and arousal signs:
| Quadrant | Valence | Arousal |
| 0 | − | − |
| 1 | − | + |
| 2 | + | − |
| 3 | + | + |
Importantly, these labels are defined by environmental variables and are never inferred from internal plant signals during training.
3. Results
Data was collected over a two week period in late December to early January in an office in Kyoto, Japan. Data was collected and stored through cloud-based InfluxDB database. This method allows for off-chip large scale data collection, as well as built in temporal formatting. IR levels below 1 were filtered out, and while this did not disclude night-time readings, it did eliminate the times when the light sensor failed to write to the database. Using only internal plant signals, we train an ESN to decode the plant’s location within the defined affective space. Importantly, the model never observes environmental variables directly. Successful decoding therefore implies that internal plant dynamics carry information predictive of the externally defined affective regime, and therefore the environment itself, and could be conducive to emotional states in relation to its subjective experience.
To test the viability at first, linear classifiers were trained to assess valence and arousal independently using internal plant signals only. Input features were gas measurements (TVOC and eCO2) and bioelectrical signals.
Time-series data were segmented into windows, and summary statistics (mean, standard deviation) were computed within each window. After testing, it was decided that binary classifiers would be trained separately for valence and arousal. Balanced accuracy was used as the primary metric to account for class imbalance. Quadrant predictions were then derived by combining the predicted valence and arousal results, rather than being learned directly. This decomposition preserves the separation of affective dimensions.
3.1. Environmental Affective Space
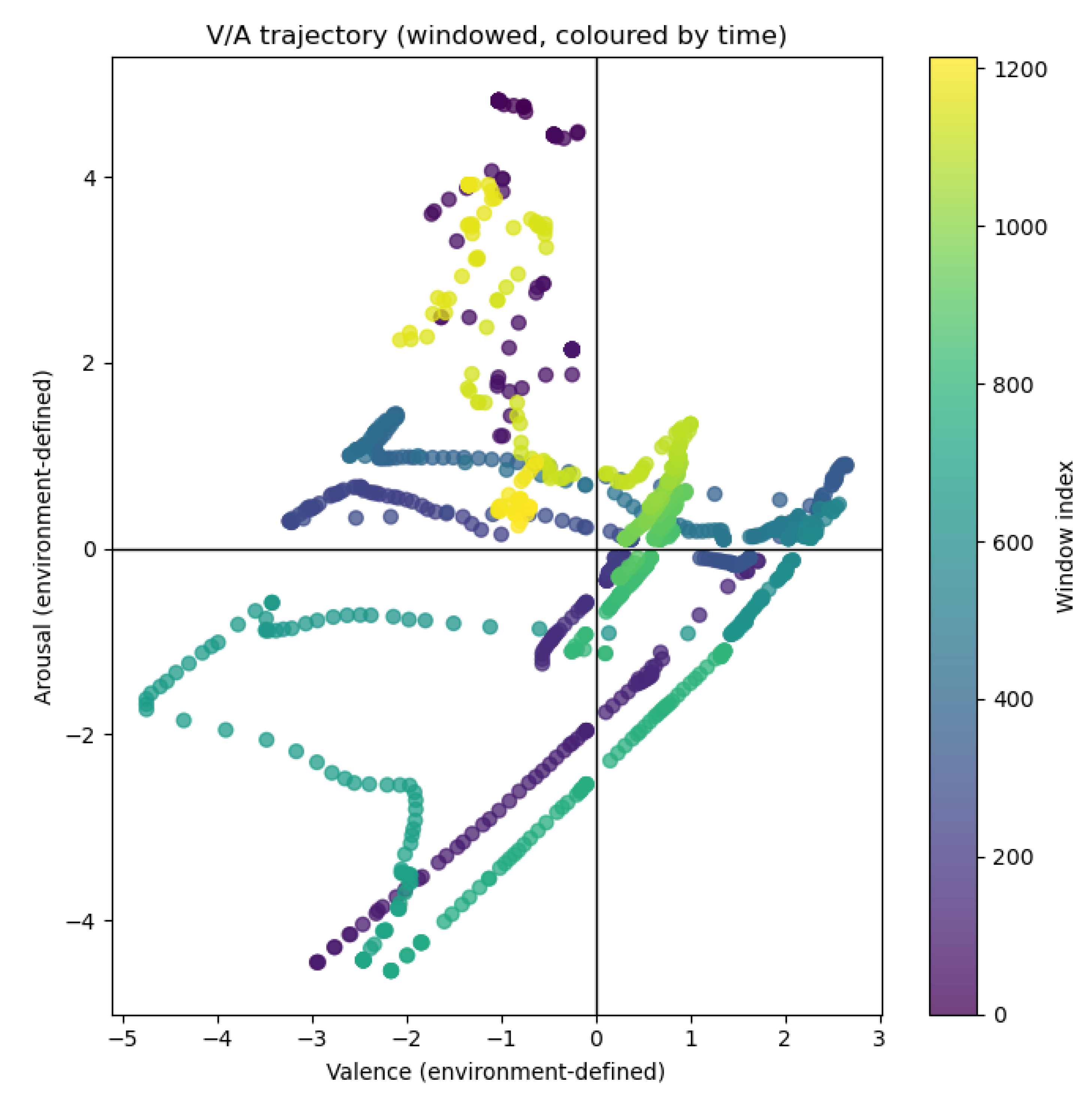
Environmental conditions occupied all four quadrants of the defined valence/arousal space. Valence and arousal labels were temporally stable, with regime durations typically spanning tens of minutes to several hours, providing a suitable target for decoding from internal plant signals. Figure 2 shows the trajectory of valence and arousal over 144 hours, with each window being 7 minutes long.
3.2. Classifying Valence from Internal Signals
Using only internal plant signals, binary valence classification exceeded majority-class baselines across multiple temporal window sizes. With a 20-minute window and 15-minute stride, the classifier achieved a mean accuracy of () and balanced accuracy of , compared to a majority baseline of . Confusion matrix revealed structured errors rather than random misclassifications, indicating that internal signals carried information predictive of the environmental valence regime.
Figure 3.
Confusion matrix for binary decoding of valence from internal plant signals. Diagonal dominance indicates decoding performance exceeding majority-class baselines.
Figure 3.
Confusion matrix for binary decoding of valence from internal plant signals. Diagonal dominance indicates decoding performance exceeding majority-class baselines.
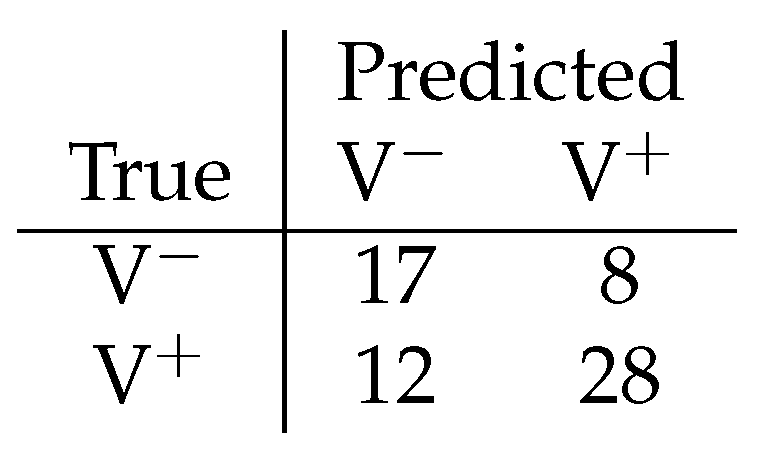
3.3. Classifying Arousal from Internal Signals
Arousal decoding proved more temporally sensitive than valence. When decoded with windows sizes similar to valence, the decoder provided near-baseline performance. However, windows of 1 minute () provided substantially improved balanced accuracy () , suggesting that arousal-related internal dynamics operate on faster timescales than valence-related dynamics. This difference is consistent with the interpretation of arousal as reflecting rapid physiological responsiveness rather than slow environmental appraisal.
Figure 4.
Confusion matrix for binary decoding of arousal from internal plant signals. Diagonal dominance indicates structured decoding performance exceeding majority-class baselines.
Figure 4.
Confusion matrix for binary decoding of arousal from internal plant signals. Diagonal dominance indicates structured decoding performance exceeding majority-class baselines.
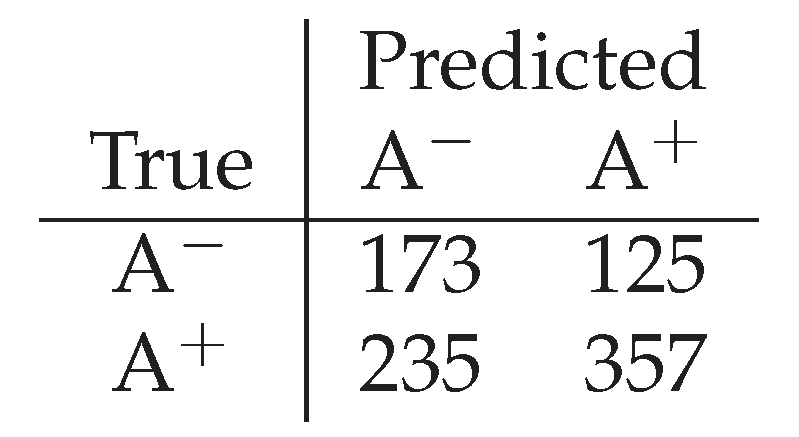
3.4. Quadrant Classification Through Combined Binary Classifier
Direct classification of affective quadrants from internal signals resulted in unstable performance and unstructured confusion matrices. However, breaking down the task into independent valence and arousal classifiers and combining them produced more interpretable results. However, quadrant classification accuracy remained modest (0.35–0.45), balanced accuracy improved relative to chance (0.37–0.48).
Performance varied substantially across cross-validation folds. Some folds lacked full quadrant coverage, inflating the variance and limiting macro-averaged metrics. These limitations give reason for a ESN decoding approach that preserves temporal continuity rather than relying on static windowed summaries.
3.5. Echo State Network Classification
ESN decoding substantially increased sample size () and stabilised performance across cross-validation folds. Quadrant classification accuracy reached , with balanced accuracy of , exceeding chance levels (0.25) and outperforming the window-based approaches. Misclassifications occurred primarily between adjacent quadrants rather than opposing affective states.
4. Discussion
This study explores the hypothesis that plants occupy affective states relative to their environmental conditions, and that these states are reflected in internal physiological dynamics that can be learned by machines. Affective state was defined not as a subjective mental property but as a relational construct arising from sustained environmental regimes. The internal signals of the plant were treated as an embodied record of these regimes, and machine learning was used to determine whether affective structure could be decoded without direct access to environmental variables. We seek to follow on the posthumanist message that affectation is not an inherently conscious notion, and that detection is not only conducted by machines, but that humans, machines and beings can form coherent assemblages capable of relating and expanding each members conscious existence.
Across all classification approaches, internal plant signals were found to contain information predictive of externally defined affective regimes. However, performance varied substantially depending on both the affective dimension considered and the temporal structure of the model.
Binary classification of valence consistently exceeded majority-class baselines and remained relatively stable across window sizes, indicating that valence-related information is distributed over longer temporal scales. This capturing of valence interestingly correlates with the claims of [36], whose work claims that valence is a mere mechanic to distinguish between ’advantage and harm’ and that this function should not be limited to conscious beings. In contrast, arousal decoding was highly sensitive to window duration, with performance improving markedly at short timescales. This asymmetry suggests that arousal reflects faster, more transient physiological dynamics, whereas valence corresponds to slower, more persistent internal states. This temporal dissociation is consistent with established valence–arousal models in affective science, in which arousal tracks rapid physiological mobilisation while valence reflects sustained environmental appraisal [37].
Direct multiclass classification of affective quadrants from static windowed features was unreliable, producing unstable performance and random confusion matrices. Decomposing the problem into independent valence and arousal classifiers improved interpretability and revealed structured errors, with misclassifications primarily occurring between adjacent quadrants. This supports the interpretation that affective structure is present in internal signals but not well captured by static, multiclass classification systems.
Echo State Network decoding further improved performance by preserving temporal continuity in internal plant dynamics. Compared to window-based classifiers, the ESN achieved higher average accuracy and reduced fold-to-fold variability, indicating that affective information is distributed across time rather than being localised to individual windows. Similar to binary quadrant classification, the misclassifications occurred primarily between adjacent quadrants as opposed to random affective state changes. This indicates that the ESN captured affective structure based on temporal dependencies at a much better than chance rate, and exceeding binary classifications. However, performance was far from perfect. This indicates that although temporal structure in internal plant dynamics is informative of affective regime, the mapping between internal state and environmental affect is neither deterministic nor fully captured by the present model.
Several limitations should be acknowledged. Firstly, the study focused on a single plant species under controlled indoor conditions, limiting generalisability, although the species chosen was a model species for experimentation in dynamics capturing. Secondly, classification performance varied across folds, reflecting both class imbalance and the inherently probabilistic relationship between internal plant dynamics and environmental regimes. Finally, while Echo State Networks captured temporal structure more effectively than static classifiers, they are simplified models of plant physiology and do not capture the full picture of biochemical and electrical signaling. Future work should explore multi-plant replication, nonlinear affective mappings, and richer temporal models to further characterise the structure and limits of plant affective dynamics.
Taken together, these results demonstrate that plants engage in structured affective regimes correlated with their environmental conditions, and that these regimes are learnable by machines based on internal dynamics. Rather than supporting a simplistic stimulus–response model, the findings point toward a temporally based, embodied form of affective organisation in plants that is probabilistic and history-dependent. While the present models are limited, the results nevertheless support the claim that plants occupy an affective space that can be inferred from their internal dynamics and used to characterise environmental conditions.
This study treats affection as a relational and functional construct of the environment. Valence and arousal are operationalised as structured responses to environmental regimes, defined in terms of environmental data and internal physiological states, without reference to mental states. This distinction is aligned with works such as [36] and [38], who state that affect is a non-conscious form of environmental engagement. In the context of plants, this view is consistent with arguments that plant behaviour reflects sophisticated adaptive regulation and internal integration without necessitating subjective awareness or consciousness [39]. From this perspective, affect doesn’t need to be conscious in order to be real, structured, or functionally significant. By separating affection from consciousness, this work contributes to a broader effort to understand affective organisation as a property of embodied systems operating across biological scales. This study has shown plants participate in affective regimes without needing to claim about subjective experience, just as many physiological and regulatory processes in animals operate affectively without entering awareness.
In this sense, affect in plants need not be understood as emotion, but as a temporally extended mode of environmental engagement that can be inferred, but not reduced, through internal state dynamics. The affective space constructed in this study is not exclusive to plants, but overlaps with dimensions humans themselves inhabit, suggesting the possibility of future human–plant interfaces grounded not in anthropomorphism, but in shared affective structure.
Author Contributions
Conceptualization, B.C., L.C. and O.W.; methodology, B.C. and L.C.; software, B.C.; validation, B.C. and L.C.; formal analysis, B.C.; investigation, B.C.; resources, B.C.; data curation, B.C.; writing—original draft preparation, B.C.; writing—review and editing, B.C. and L.C.; visualization, B.C. and L.C.; supervision, L.C.; project administration, O.W.; funding acquisition, O.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Calvo, P.; Gagliano, M.; Souza, G.M.; Trewavas, A. Plants are intelligent, here’s how. Annals of Botany 2020, 125, 11–28. [CrossRef]
- Volkov, A.G.; Ranatunga, D.R. Plants as environmental biosensors. Plant Signaling & Behavior 2006, 1, 105–115. [CrossRef]
- Aust, T.; et al. Automated Phytosensing: Ozone Exposure Classification Based on Plant Electrical Signals. arXiv preprint arXiv:2412.13312 2024. [CrossRef]
- Gloor, P.A. Plant Bioelectrical Signals for Environmental and Emotional State Classification. Biosensors 2025, 15, 744. [CrossRef]
- Kruse, J.A.; et al. Leveraging the Sensitivity of Plants with Deep Learning to Recognize Human Emotions. Sensors 2024, 24, 1917. [CrossRef]
- Fromm, J.; Lautner, S. Electrical signals and their physiological significance in plants. Plant, Cell & Environment 2007, 30, 249–257.
- Volkov, A.G. Plant Electrophysiology: Theory and Methods; Springer, 2006.
- Taiz, L.; Zeiger, E.; et al. Plant Physiology and Development, 6 ed.; Sinauer Associates, 2015.
- Choi, W.G.; Toyota, M.; Kim, S.J.; Hilleary, R.; Gilroy, S. Salt stress-induced Ca2+ waves are associated with rapid, long-distance root-to-shoot signalling in plants. Proceedings of the National Academy of Sciences 2016, 113, E2805–E2814.
- Trewavas, A. Plant Behaviour and Intelligence; Oxford University Press, 2014.
- Russell, J.A. A circumplex model of affect. Journal of Personality and Social Psychology 1980, 39, 1161–1178.
- Ahire, V.; Shah, K.; Khan, M.N.; Pakhale, N.; Sookha, L.R.; Ganaie, M.A.; Dhall, A. MAVEN: Multi-modal Attention for Valence-Arousal Emotion Network 2025. arXiv:2503.12623 [cs], . [CrossRef]
- Pereira, D.R.; Papa, J.P.; Saraiva, G.F.R.; Souza, G.M. Automatic classification of plant electrophysiological responses to environmental stimuli using machine learning and interval arithmetic. Computers and Electronics in Agriculture 2018, 145, 35–42. [CrossRef]
- Notaguchi, M.; Okamoto, S. Dynamics of long-distance signaling via plant vascular tissues. Frontiers in Plant Science 2015, 6. [CrossRef]
- Barbosa-Caro, J.C.; Wudick, M.M. Revisiting plant electric signaling: Challenging an old phenomenon with novel discoveries. Current Opinion in Plant Biology 2024, 79, 102528–102528. [CrossRef]
- Loreto, F.; Schnitzler, J.P. Isoprene emission in plants: a physiological and ecological perspective. Trends in Plant Science 2006, 11, 65–72.
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends in Plant Science 2010, 15, 176–184.
- Ebato, Y.; Nobukawa, S.; Sakemi, Y.; Nishimura, H.; Kanamaru, T.; Sviridova, N.; Aihara, K. Impact of time-history terms on reservoir dynamics and prediction accuracy in echo state networks. Scientific Reports 2024, 14. [CrossRef]
- Mochiutti, E.; Antonelo, E.A.; Camponogara, E. Physics-informed Echo State Networks for modeling controllable dynamical systems. Neurocomputing 2025, 639, 130251–130251. [CrossRef]
- Ma, Q.; Shen, L.; Cottrell, G.W. DeePr-ESN: A deep projection-encoding echo-state network. Information Sciences 2020, 511, 152–171. [CrossRef]
- Sheng, C.; Zhao, J.; Liu, Y.; Wang, W. Prediction for noisy nonlinear time series by echo state network based on dual estimation. Neurocomputing 2012, 82, 186–195. [CrossRef]
- Jaeger, H. Adaptive Nonlinear System Identification with Echo State Networks. Advances in Neural Information Processing Systems 2002, 15, 609–616.
- Maass, W.; Natschläger, T.; Markram, H. Real-Time Computing Without Stable States: A New Framework for Neural Computation Based on Perturbations. Neural Computation 2002, 14, 2531–2560. [CrossRef]
- Tanaka, G.; Yamane, T.; Héroux, J.B.; Nakane, R.; Kanazawa, N.; Takeda, S.; Numata, H.; Nakano, D.; Hirose, A. Recent advances in physical reservoir computing: A review. Neural Networks 2019, 115, 100–123. [CrossRef]
- Nakajima, K. Physical reservoir computing—an introductory perspective. Japanese Journal of Applied Physics 2020, 59, 060501. [CrossRef]
- Pieters, O.; De Swaef, T.; Stock, M.; Wyffels, F. Leveraging plant physiological dynamics using physical reservoir computing. Scientific Reports 2022, 12, 1–12. [CrossRef]
- Fromm, J.; Lautner, S. Electrical signals and their physiological significance in plants. Plant, Cell & Environment 2006, 30, 249–257. [CrossRef]
- Haraway, D. Simians, Cyborgs, and Women: The Reinvention of Nature; Routledge: New York, 1991.
- Barad, K. Meeting the Universe Halfway: Quantum Physics and the Entanglement of Matter and Meaning; Duke University Press: Durham, NC, 2007. [CrossRef]
- Braidotti, R. The Posthuman; Polity Press: Cambridge, 2013.
- Masi, E.; Ciszak, M.; Stefano, G.; Renna, L.; Azzarello, E.; Pandolfi, C.; Mugnai, S.; Baluška, F.; Arecchi, F.; Gallelli, A.; et al. Electrical signals and fast long-distance communication in plants. Plant Signaling & Behavior 2009, 4, 409–417. [CrossRef]
- Gabrys, J.; Hayles, N.; Krapp, P.; Raley, R.; Weber, S. Program Earth: Environmental Sensing Technology and the Making of a Computational Planet; University of Minnesota Press: Minneapolis, 2016. [CrossRef]
- Myers, N. Photosynthetic entanglements: Experiments in plant sensing. Configurations 2017, 25, 79–104. [CrossRef]
- Segundo-Ortin, M.; Calvo, P. Plant Sentience? Between Romanticism and Denial. Animal Sentience 2023, 8. [CrossRef]
- Maria, V.; Carolina, A.; Souza, G.M.; Xavier, J. Using plant electrical signals of water hyacinth ( Eichhornia crassipes ) for water pollution monitoring. Open Life Sciences 2025, 20, 20251120–20251120. [CrossRef]
- Lyon, P.; Kuchling, F. Valuing what happens: a biogenic approach to valence and (potentially) affect. Philosophical Transactions of the Royal Society B: Biological Sciences 2021, 376, 20190752. [CrossRef]
- Cui, X.; Tian, Y.; Zhang, L.; Chen, Y.; Bai, Y.; Li, D.; Liu, J.; Gable, P.; Yin, H. The role of valence, arousal, stimulus type, and temporal paradigm in the effect of emotion on time perception: A meta-analysis. Psychonomic Bulletin & Review 2022. [CrossRef]
- Dehaene, S. Consciousness and the brain : deciphering how the brain codes our thoughts; Penguin Books: New York, New York, 2014.
- Calvo, P.; Trewavas, A. Cognition and intelligence of green plants. Information for animal scientists. Biochemical and Biophysical Research Communications 2020. [CrossRef]
Figure 1.
Annotated experimental setup showing sensors, heater, fan and camera positions.

Figure 2.
Trajectory of environmental valence and arousal over a 144-hour period. Valence and arousal were computed from residualised light, temperature, and humidity, and are shown as a continuous affective trajectory. Points represent 7-minute windows, illustrating extended residence within affective regimes and gradual transitions between them.
Figure 2.
Trajectory of environmental valence and arousal over a 144-hour period. Valence and arousal were computed from residualised light, temperature, and humidity, and are shown as a continuous affective trajectory. Points represent 7-minute windows, illustrating extended residence within affective regimes and gradual transitions between them.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



