
Submitted:
21 January 2026
Posted:
22 January 2026
You are already at the latest version
Abstract
The objective was to detect T. gondii and various protozoan oocysts in the feces of sheltered cats in Thailand using two molecular approaches, Sanger sequencing and un-targeted Next-generation sequencing (NGS). A total of 166 fecal samples from shelter cats, in 26 samples, Toxoplasma gondii oocyst–like structures were detected. The harvested oocysts were grouped into nine pooled samples. DNA was extracted from all pooled samples and tested using quantitative PCR (qPCR), employing coc1 and coc2 primers, which are commonly used to amplify Apicomplexan DNA. The sequenced of the qPCR products were analyzed by Sanger sequencing. Sequence from all pooled samples had similarity to Cystoisospora spp. To further characterize the oocyst species, NGS was performed. Bioinformatic analysis was conducted using a de novo assembler to generate scaffolds, which were then aligned against a custom database of coccidian whole-genome references. This analysis revealed the presence of T. gondii DNA in three pooled samples. In addition, DNA from other protozoan parasites—including Eimeria spp., Cystoisospora spp., Besnoitia besnoiti, Hammondia hammondi, and Cryptosporidium parvum—was also detected. These findings indicate that T. gondii is circulating among shelter cats, many of which were formerly stray. Moreover, NGS sequencing provided more comprehensive information on the diversity of coccidian species in cat feces compared with Sanger sequencing of PCR-amplified targets.
Keywords:
1. Introduction
2. Materials and Methods
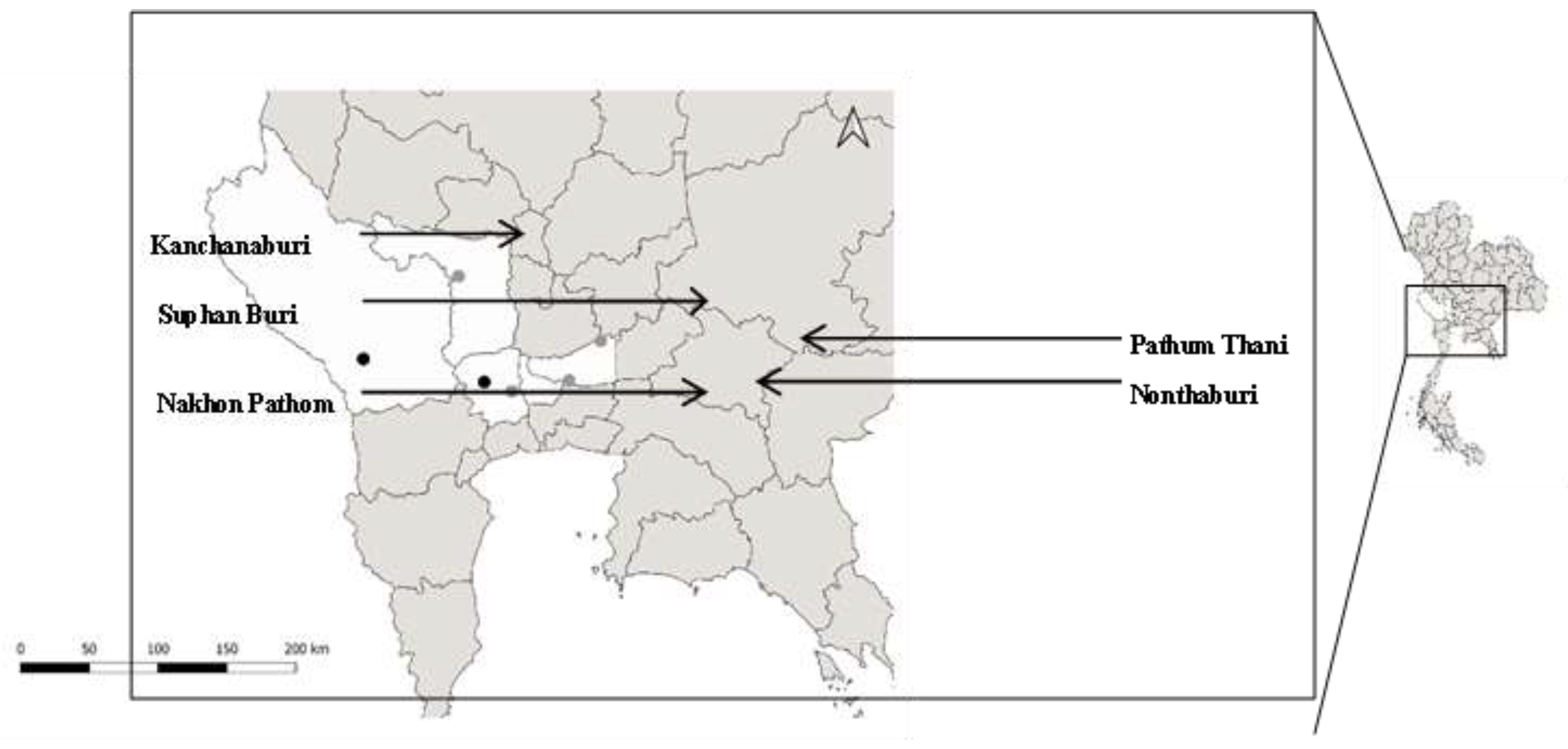
2.1. Fecal Sample and Oocyst Collection
2.2. Simple Floatation Technique
2.3. DNA Extraction from Oocyst
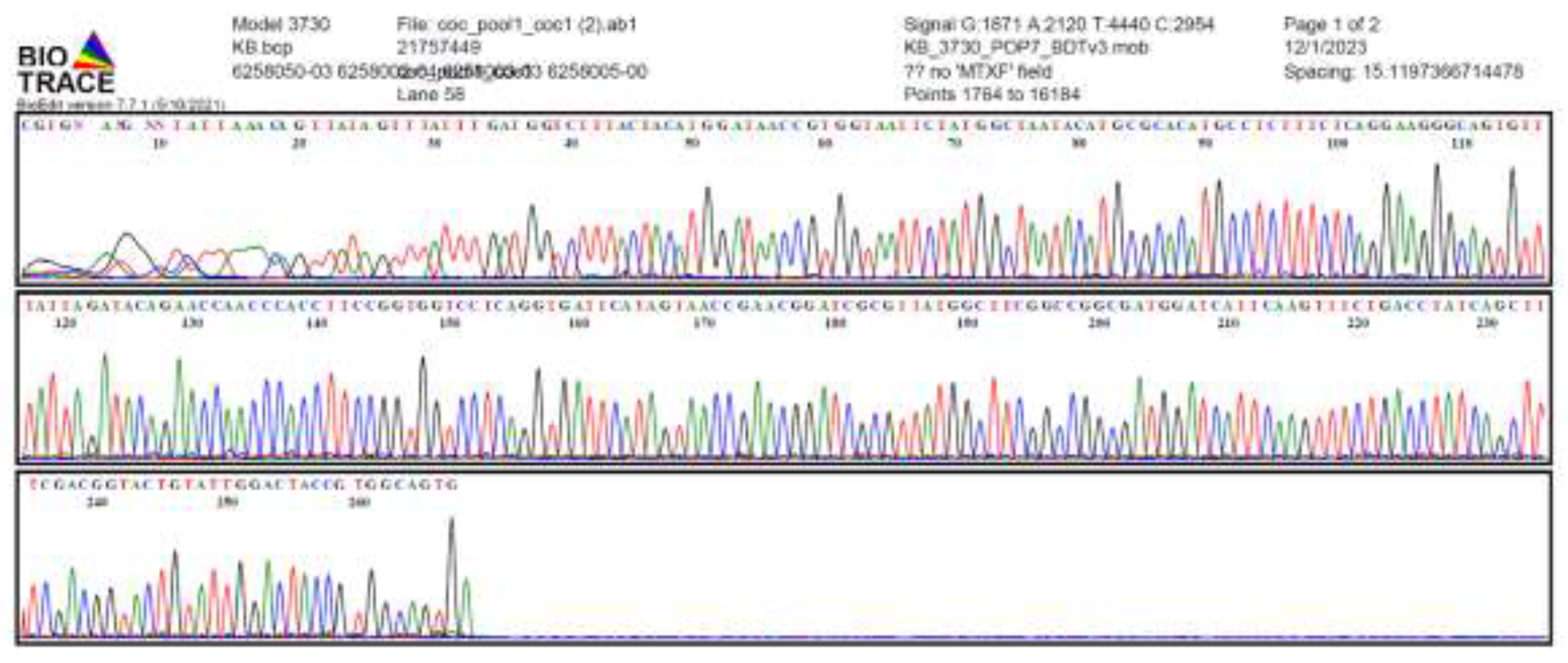
2.4. qPCR and Sanger Sequencing
2.5. DNA Library Preparation and Illumina Sequencing
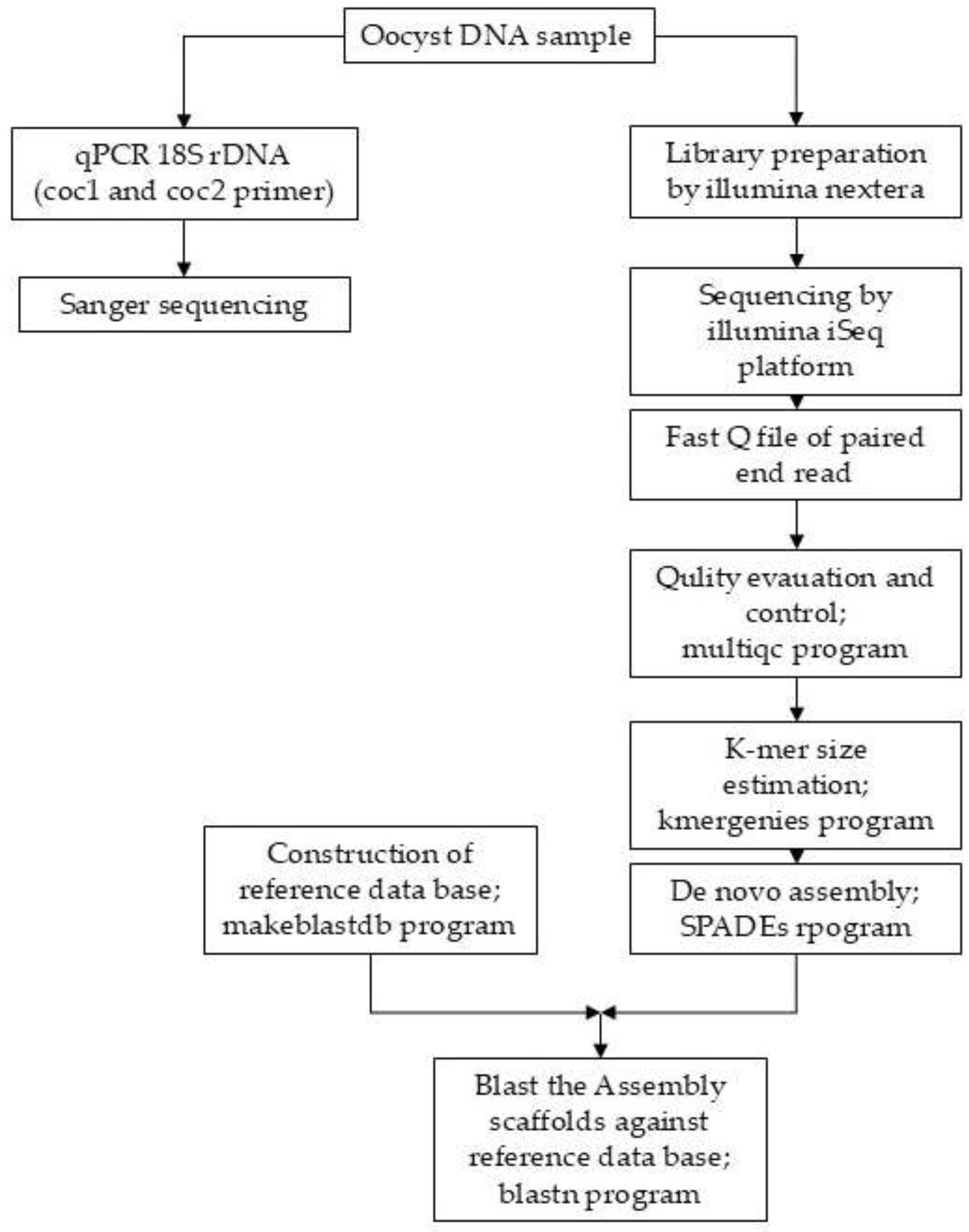
2.6. Bioinformatics Workflow
2.7. The Prevalence of T. gondii in Shelter Cats
3. Results
3.1. qPCR and Sanger Sequencing
3.2. Un-Targeted Next Generation Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robert-Gangneux, F.; Darde, M. L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Torrey, E. F.; Yolken, R. H. Toxoplasma oocysts as a public health problem. Trends Parasitol. 2013, 29, 380–384. [Google Scholar] [CrossRef]
- Dubey, J. P. The history and life cycle of Toxoplasma gondii. In Toxoplasma gondii, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar]
- Sroka, J.; Karamon, J.; Dutkiewicz, J.; Wojcik-Fatla, A; Cencek, T. Optimization of flotation, DNA extraction and PCR methods for detection of Toxoplasma gondii oocysts in cat faeces. Ann. Agric. Environ. Med. 2018, 25, 680–685. [Google Scholar] [CrossRef]
- Chemoh, W.; Sawangjaroen, N.; Nissapatorn, V.; Sermwittayawong, N. Molecular investigation on the occurrence of Toxoplasma gondii oocysts in cat feces using TOX-element and ITS-1 region targets. Vet. J. 2016, 215, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L. F.; Gajadhar, A. A. Detection and differentiation of coccidian oocysts by real-time PCR and melting curve analysis. J. Parasitol. 2011, 97, 725–730. [Google Scholar] [CrossRef]
- Suwancharoen, C.; Phuangsri, C.; Prakhammin, K.; Japa, O. Molecular detection and dense granule antigen 6 genotyping of feline Toxoplasma gondii in Phayao, Thailand. Vet. World 2022, 15, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Zulpo, D. L.; Sammi, A. S.; Dos Santos, J. R.; Sasse, J. P.; Martins, T. A.; Minutti, A. F.; Cardim, S. T.; de Barros, L. D.; Navarro, I. T.; Garcia, J. L. Toxoplasma gondii: A study of oocyst re-shedding in domestic cats. Vet. Parasitol. 2018, 249, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Miura, A. C.; Barros, L. D.; Minutti, A. F.; Martins, T. A.; Sasse, J. P.; Nino, B. S. L.; Garcia, J. L. Evaluation of quantitative polymerase chain reaction for the detection of Toxoplasma gondii oocysts shed by cats. Rev. Bras. Parasitol. Vet. 2021, 30, e016621. [Google Scholar] [CrossRef]
- Amouei, A.; Sarvi, S.; Sharif, M.; Aghayan, S. A.; Javidnia, J.; Mizani, A.; Moosazadeh, M.; Shams, N.; Hosseini, S. A.; Hosseininejad, Z.; Nayeri Chegeni, T.; Badali, H.; Daryani, A. A systematic review of Toxoplasma gondii genotypes and feline: Geographical distribution trends. Transbound. Emerg. Dis. 2020, 67, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Paparini, A.; Gofton, A.; Yang, R.; White, N.; Bunce, M.; Ryan, U. M. Comparison of Sanger and next generation sequencing performance for genotyping Cryptosporidium isolates at the 18S rRNA and actin loci. Exp. Parasitol. 2015, 151, 21–27. [Google Scholar] [CrossRef]
- Gad, M.; Fawzy, M. E.; Al-Herrawy, A. Z.; Abdo, S. M.; Nabet, N.; Hu, A. PacBio next-generation sequencing uncovers Apicomplexa diversity in different habitats. Sci. Rep. 2023, 13, 15063. [Google Scholar] [CrossRef]
- Moreno, Y.; Moreno-Mesonero, L.; Amorós, I.; Pérez, R.; Morillo, J. A.; Alonso, J. L. Multiple identification of most important waterborne protozoa in surface water used for irrigation purposes by 18S rRNA amplicon-based metagenomics. Int. J. Hyg. Environ. Health. 2018, 221, 102–111. [Google Scholar] [CrossRef]
- Dulanto Chiang, A.; Dekker, J. P. From the pipeline to the bedside: advances and challenges in clinical metagenomics. J. Infect. Dis. 2020, 221, S331–S340. [Google Scholar] [CrossRef]
- Mthethwa, N. P.; Amoah, I. D.; Reddy, P.; Bux, F.; Kumari, S. A review on application of next-generation sequencing methods for profiling of protozoan parasites in water: Current methodologies, challenges, and perspectives. J. Microbiol. Methods 2021, 187, 106269. [Google Scholar] [CrossRef] [PubMed]
- Feldmeyer, B.; Wheat, C. W.; Krezdorn, N.; Rotter, B.; Pfenninger, M. Short read Illumina data for the de novo assembly of a non-model snail species transcriptome (Radix balthica, Basommatophora, Pulmonata), and a comparison of assembler performance. BMC Genomics 2011, 12, 317. [Google Scholar] [CrossRef]
- Wylezich, C.; Belka, A.; Hanke, D.; Beer, M.; Blome, S.; Höper, D. Metagenomics for broad and improved parasite detection: a proof-of-concept study using swine faecal samples. Int. J. Parasitol. 2019, 49, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L. F.; Gajadhar, A. A. Detection and differentiation of coccidian oocysts by real-time PCR and melting curve analysis. J. Parasitol. 2011, 97, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J. P.; Lindsay, D. S.; Lappin, M. R. Toxoplasmosis and other intestinal coccidial infections in cats and dogs. Vet. Clin. North Am. Small Anim. Pract. 2009, 39, 1009–1034. [Google Scholar] [CrossRef]
- Lindsay, D. S.; Dubey, J. P. Neosporosis, toxoplasmosis, and sarcocystosis in ruminants: an update. Vet. Clin. North Am. Food Anim. Pract. 2020, 36, 205–222. [Google Scholar] [CrossRef]
- Almeria, S.; Cinar, H. N.; Dubey, J. P. Cyclospora cayetanensis and cyclosporiasis: an update. Microorganisms 2019, 7, 317. [Google Scholar] [CrossRef]
- Abbas, I.; Al-Araby, M.; Elmishmishy, B.; El-Alfy, E. S. Gastrointestinal parasites of cats in Egypt: high prevalence high zoonotic risk. BMC Vet. Res. 2022, 18, 420. [Google Scholar] [CrossRef]
- Jittapalapong, S.; Inparnkaew, T.; Pinyopanuwat, N.; Kengradomkij, C.; Sangvaranond, A.; Wongnakphet, S. Gastrointestinal parasites of stray cats in Bangkok metropolitan areas, Thailand. Agric. Nat. Resour. 2007, 41, 69–73. [Google Scholar]
- Jittapalapong, S.; Inpankaew, T.; Pinyopanuwat, N.; Chimnoi, W.; Kengradomkij, C.; Wongnarkpet, S.; Maruyama, S; Lekkla, A.; Sukthana, Y. Epidemiology of Toxoplasma gondii infection of stray cats in Bangkok, Thailand. Southeast Asian J. Trop. Med. Pub. Health. 2010, 41, 13–18. [Google Scholar]
- Kengradomkij, C.; Kamyingkird, K.; Pinyopanuwat, N.; Chimnoi, W.; Jittapalapong, S.; Inpankaew, T. Seroprevalence of Toxoplasma gondii from stray cats residing in temples, Bangkok, Thailand. J. Trop. Med. Parasitol. 2018, 41, 8–14. [Google Scholar]
- Miura, A. C.; Barros, L. D.; Minutti, A. F.; Martins, T. A.; Sasse, J. P.; Nino, B. S. L.; Garcia, J. L. Evaluation of quantitative polymerase chain reaction for the detection of Toxoplasma gondii oocysts shed by cats. Rev. Bras. Parasitol. Vet. 2021, 30, e016621. [Google Scholar] [CrossRef]
- Ramakrishnan, C.; Maier, S.; Walker, R. A.; Rehrauer, H.; Joekel, D. E.; Winiger, R. R.; Basso, W.U.; Grigg, M.E.; Hehl, A. B.; Deplazes, P.; Smith, N. C. An experimental genetically attenuated live vaccine to prevent transmission of Toxoplasma gondii by cats. Sci. Rep. 2019, 9, 1474. [Google Scholar] [CrossRef]
- Ho, M. S.; Barr, B. C.; Marsh, A. E.; Anderson, M. L.; Rowe, J. D.; Tarantal, A. F.; Hendrickx, A. G.; Sverlow, K.; Dubey, J. P.; Conrad, P. A. Identification of bovine Neospora parasites by PCR amplification and specific small-subunit rRNA sequence probe hybridization. J. Clin. Microbiol. 1996, 34, 1203–1208. [Google Scholar] [CrossRef]
- EpiTools Online Epidemiological Calculator. Available online: https://epitools.ausvet.com.au/ppvariablepoolsize (accessed on 13 October 2024).
- Mosquera, J. D.; Valle, C. A.; Nieto-Claudin, A.; Fessl, B.; Lewbart, G. A.; Deresienski, D.; Bouazzi, L.; Zapata, S; Villena, I; Poulle, M. L. Prevalence of Toxoplasma gondii in Galapagos birds: Inference of risk factors associated with diet. Plos One 2023, 18, e0287403. [Google Scholar] [CrossRef]
- Williams, C. J.; Moffitt, C. M. A critique of methods of sampling and reporting pathogens in populations of fish. J. Aquat. Anim. Health 2001, 13, 300–309. [Google Scholar] [CrossRef]
- Herrmann, D. C.; Pantchev, N.; Vrhovec, M. G.; Barutzki, D.; Wilking, H.; Frohlich, A.; Luder, C. G.; Conraths, F. J.; Schares, G. Atypical Toxoplasma gondii genotypes identified in oocysts shed by cats in Germany. Int. J. Parasitol. 2010, 40, 285–292. [Google Scholar] [CrossRef]
- Fehlberg, H. F.; Maciel, B. M.; Albuquerque, G. R. Identification and discrimination of Toxoplasma gondii, Sarcocystis spp., Neospora spp., and Cryptosporidium spp. by right-resolution melting analysis. Plos One 2017, 12, e0174168. [Google Scholar] [CrossRef]
- Udonsom, R.; Buddhirongawatr, R.; Nishikawa, Y.; Fereig, R. M.; Jirapattharasate, C. Toxoplasma gondii prevalence and risk factors in owned domestic cats from Nakhon Pathom Province, Thailand. Vet. Integr. Sci. 2021, 19, 557–566. [Google Scholar] [CrossRef]
- Jitsamai, W.; Khrutkham, N.; Hunprasit, V.; Chandrashekar, R.; Bowman, D.; Sukhumavasi, W. Prevalence of endoparasitic and viral infections in client-owned cats in metropolitan Bangkok, Thailand, and the risk factors associated with feline hookworm infections. Vet. Parasitol. Reg. Stud. Reports 2021, 25, 100584. [Google Scholar] [CrossRef]
- Soe, B. K.; Hlaing, K. S.; Naing, T. W.; Thaw, Z. H.; Myint, A. W. The first study on the prevalence of gastrointestinal parasites in owned and shelter cats in Yangon, Myanmar. Vet. World 2023, 16, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Blake, D. P.; Alias, H.; Billington, K. J.; Clark, E. L.; Mat-Isa, M. N.; Mohamad, A. F. H.; Mohd-Amin, M.R.; Tay, Y.L.; Smithe, A.L.; Tomley, F.M.; Wan, K. L. EmaxDB: Availability of a first draft genome sequence for the apicomplexan Eimeria maxima. Mol. Biochem. Parasitol. 2012, 184, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Fang, H.; Ohlson, T.; Frey-Skött, J.; Elofsson, A. Using evolutionary information for the query and target improves fold recognition. Proteins: Struct., Funct., Bioinf. 2004, 54, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L. A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef]



| Sample | Number of fecal samples | Location | Sanger sequencing Coccidia identification |
Un – targeted NGS (Illumina sequencing) |
|||
|---|---|---|---|---|---|---|---|
| Total reads (bp) |
Total scaffolds (N50) |
Coccidia identification |
|||||
| Pool 1 | 3 | Muang district, Kanchanaburi province |
Cystoisospora spp. | 129,493 | 604 (230) |
Cystoisospora spp, Besnoitia besnoiti, Hammondia hammondi, Toxoplasma gondii, Eimeria spp. |
|
| Pool 2 | 3 | Muang district, Kanchanaburi province |
Cystoisospora spp. | 88,157 | 191 (473) |
Cystoisospora spp., Toxoplasma gondii, |
|
| data | |||||||
| Pool 3 | 3 | Nakhon Chai Sri district, Nakhon Pathom province |
Cystoisospora spp. | 2,704,357 | 38,873 (112) |
Cystoisospora spp., Besnoitia besnoiti, Eimeria spp. |
|
| Pool 4 | 3 | Don Toom district, Nakhon Pathom province |
Cystoisospora spp. | 319,560 | 1,761 (208) |
ND | |
| Pool 5 | 2 | Don Chae Dee district, Suphan Buri province |
Cystoisospora spp. | 34,536 | 155 (230) |
Cystoisospora spp., | |
| Pool 6 | 3 | Muang district, Kanchanaburi province |
Cystoisospora spp. | 84,529 | 429 (213) |
ND | |
| Pool 7 | 3 | Muang district, Kanchanaburi province |
Cystoisospora spp. | 74,826 | 506 (208) |
Cystoisospora spp., | |
| Pool 8 | 3 | Don Toom district, Nakhon Pathom province |
Cystoisospora spp. | 153,529 | 873 (212) |
Cystoisospora spp., Cryptosporidium parvum, Besnoitia besnoiti, Eimeria spp. |
|
| Pool 9 | 3 | Don Toom district, Nakhon Pathom province |
Cystoisospora spp. | 124,896 | 628 (218) |
Toxoplasma gondii, Eimeria spp. |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



