Submitted:
20 January 2026
Posted:
20 January 2026
You are already at the latest version
Abstract
Climate change is making winters in temperate regions increasingly harsh and unpredictable, posing significant threats to honey bee (Apis mellifera) health. The gut microbiome, a distinct community of core bacterial species, is central to overwintering success by supporting immune function, nutrient assimilation, and pathogen resistance, but is highly sensitive to environmental stressors such as cold temperatures and dietary shifts. Stress-induced perturbations can reshape the composition and relative abundance of the gut microbiome in honey bees, leading to adverse effects on host health, physiological functions, and overwinter survival. Cold temperatures and additional stressors further destabilize the microbiome, compounding these effects. This review is the first to synthesize current knowledge on how extrinsic factors, such as diet, antibiotics, and pathogens, and intrinsic factors, including age and strain, influence the composition and function of the honey bee gut microbiota. Given the increasing severity of winter conditions under climate change, a deeper understanding of microbiome–host–environment interactions is essential for improving honey bee resilience. By integrating evidence on the microbiome’s roles in nutrient utilization, immune modulation, and pathogen defense, this review out-lines a framework to guide future research and management strategies aimed at mitigating overwintering losses and sustaining pollinator health in a changing global climate.
Keywords:
honey bee
; gut microbiome
; overwintering
; microbial dysbiosis
; cold stress
; pollinator health
; host-microbe interactions
1. Introduction
Climate change is an escalating global issue, altering temperature regimes, precipitation patterns, and seasonal dynamics across ecosystems [47]. Despite a general trend of increasing global temperature, North America continues to experience episodic, severe cold air outbreaks [6,18]. These disruptions are associated with increasingly extreme winter conditions, particularly in the eastern United States, and are projected to intensify during mid- to late winter [18,19]. Moreover, warming increases atmospheric moisture-holding capacity, contributing to heavier snowfall, intense rainfall, and more erratic weather events [6]. Over the past two decades, such climate-driven anomalies, especially prolonged cold extremes, have disrupted ecological dynamics and threatened biodiversity, with pollinators emerging as a particularly vulnerable group [9,14,69,83,91].
Anthropogenic climate change threatens pollinator populations through synergistic pathways, including phenological mismatches between accelerated plant flowering and pollinator activity [38,68], physiological stress from increased frequency of unseasonal frosts and prolonged cold snaps that disrupt diapause and elevate metabolic demands [58,94], enhanced virulence and expanded ranges of parasites (e.g., Varroa destructor) and pathogens (e.g., Nosema spp.) under cold- weather conditions [13,58], habitat degradation via climate-exacerbated disturbances like ice storms and flooding [79], and nutritional declines in pollen protein content [5,25]. These compounding stressors acutely impact managed pollinators, particularly the Western honey bee (Apis mellifera).
The Western honey bee is one of the most important pollinators that is integral to pollination in both managed agroecosystems and wild landscapes [44]. In the United States, the economic value attributed to honey bees' annual pollination services ranges from $18 to $20 billion annually (USDA-ARS, 2024), underscoring their vital role in sustaining the productivity of numerous nuts, vegetables, and fruits. Honey bee colonies experience a substantial decline, with the majority of losses occurring during the overwintering period. The extent of these losses is higher in the United States, averaging 40.4%, compared to 12.5% and 21.3% losses in Europe and Latin America, respectively [86]. During overwintering, colony losses are often attributed to limited foraging resources, nutritional deficiencies, and extremely low temperatures. Additionally, various pests and pathogens, including Aethina tumida (hive beetle), Ascosphaera apis (fungus), Varroa destructor (parasitic mite), Melissococcus plutonius (bacterium), and Paenibacillus larvae (bacterium), also negatively impact colony health during overwintering [3,39,40,75,77,88]. Infestation by Varroa destructor is one of the leading causes of winter colony decline, directly compromising bee health and serving as a vector for viruses such as deformed wing virus (DWV) [26,97].
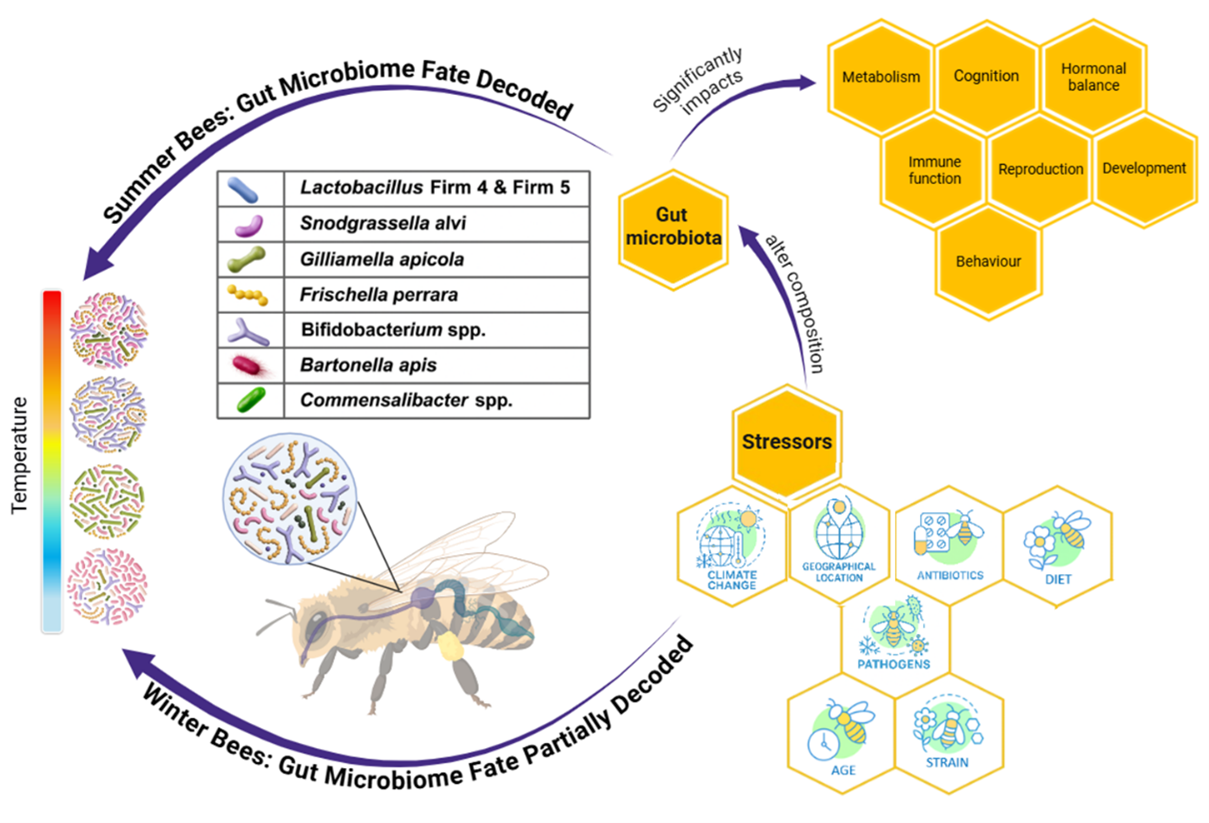
While genome-wide surveys have questioned the primacy of certain stressors, poor nutrition remains consistently linked to declining bee health. A key ecological factor influencing host nutrition is the “gut microbiome.” In general, the honey bee gut microbiota exhibits exceptionally low taxonomic diversity, with <10 bacterial phylotypes comprising >95% of the adult worker community (33, 54, 52, 62, 63). This highly conserved core microbiome, dominated by Gilliamella apicola, Snodgrassella alvi, Lactobacillus Firm-4 and Firm-5, and Bifidobacterium spp., persists across life stages, seasons, and environments, indicating strong host-specific selection [8,22,67]. Acquired horizontally via interactions (trophallaxis) rather than vertical transmission, these core taxa mediate essential functions [54,106]. Supplemental facultative taxa (e.g., Frischella perrara, Bartonella apis, and Commensalibacter sp.) occur variably across colonies and may provide auxiliary metabolic or defensive contributions [11,106]. Transient microbes (e.g., Lactobacillus kunkeei, Apibacter sp.) typically represent minor community components [23, 49, 54). The honey bee gut microbiota influence host physiology, development, and behavior; specifically, macromolecule digestion, nutrient absorption, endocrine signaling, detoxification, weight regulation, and pathogen resistance [84,106]. The gut microbiota of winter bees is abundant in Gilliamella apicola, Bartonella spp., Commensalibacter spp., and Snodgrassella spp., which helps with immunological regulation and energy metabolism under resource-limited conditions [50]. Environmental and developmental factors, including diet, age, climate, chemical exposure, antibiotics, and temperature stress, can alter the gut microbiome, thereby affecting immunity, metabolism, growth, and survival during winter [10]. Despite growing insights, major research gaps need to be addressed, particularly in understanding stress-induced dysbiosis, the interactive effects of multiple stressors on gut microbiome, and the functional consequences of microbial shifts in honeybees during overwintering. Specifically, we aim to examine how winter conditions influence the honey bee gut microbiome by synthesizing current knowledge on its interactions with overwintering physiology, dietary shifts, and antibiotic exposure, pathogens, and host factors such as age and genetic strain. While many of these factors independently affect gut microbial communities, their combined effects, particularly in the context of winter, remain poorly understood and have not been systematically explored. In this review, we examine extrinsic and intrinsic stressors, addressing them independently while synthesizing existing literature on the triadic interactions among winter conditions, gut microbes, and other environmental or biological stressors. In case no data is available on triadic interaction, we highlight critical knowledge gaps and outline targeted research questions and testable hypotheses to advance our understanding of the complex interplay between multiple stressors, gut microbiota, and honey bee health.
2. Functionality of the Gut Bacteria
2.1. Immune Modulation
Recent research has highlighted that specific gut microbes can enhance the honey bee’s immune response, offering protection against pathogens such as Serratia marcescens and Hafnia alvei. In particular, strains like Gilliamella apicola W8136 and Lactobacillus apis W8172 have been shown to reduce H. alvei infection and associated mortality. Transcriptomic analyses revealed that these beneficial bacteria induce distinct immune responses in the gut, with L. apis notably upregulating Toll pathway components such as PGRP-S3 and Spätzle, as well as stimulating the production of antibacterial peptides, including apidaecin, which exhibits strong inhibitory effects against pathogens [56]. The presence of Frischella perrara also plays a significant role in modulating gut immunity and maintaining epithelial homeostasis in the pyloric region of the honey bee gut. This symbiont elicits a strong immune response, most notably through the induction of a distinct “scab phenotype”, a melanized, darkly pigmented band that forms on the epithelial surface of the pylorus. Transcriptomic analyses of hive bees, both with and without the scab phenotype, reveal that F. perrara stimulates immune-related gene expression even under natural in-hive conditions where it coexists with other core gut bacterial species [31]. These findings underscore the strain-specific nature of gut microbes in regulating immunity in honey bees and highlight their critical role in host protection, particularly under winter stress when immune functions are otherwise suppressed.
2.2. Nutrient Utilization and Detoxification
Honey bee midgut primarily processes readily digestible nutrients such as sugars and amino acids derived from nectar and pollen, while more complex, refractory substances, including pollen coat polysaccharides and nitrogenous waste from the Malpighian tubules, transit to the hindgut for further microbial processing [34]. Gut bacteria play a fundamental role in the digestion and assimilation of nutrients essential for honey bee survival, particularly during overwintering when foraging ceases and bees rely entirely on stored resources such as honey and fermented pollen (bee bread). Within the hindgut, key bacterial taxa, including Gilliamella, Bifidobacterium, Bombilactobacillus, and Lactobacillus spp., contribute significantly to the degradation of complex carbohydrates and toxic sugars (e.g., mannose, arabinose, and rhamnose) via specialized enzymes such as pectin lyases and glycoside hydrolases [35,59,104]. In addition to carbohydrate metabolism, these microbes also play a crucial role in detoxifying plant secondary metabolites, such as flavonoids and cyanogenic glycosides, through deglycosylation [51,72]. Additionally, taxa such as Snodgrassella alvi and Gilliamella apicola harbor genes for urea metabolism and amino acid biosynthesis, suggesting a role in nitrogen recycling [60].
2.3. Protection Against Pathogens
Winter imposes heightened physiological stress on honey bee colonies, characterized by reduced foraging, clustering, and prolonged confinement, which collectively increase their vulnerability to pathogens. The gut microbiota mitigates these risks by enhancing immune responses, producing antimicrobial compounds, and maintaining gut homeostasis [54]. Core taxa such as Gilliamella, Snodgrassella, and Bifidobacterium are known to stimulate immune gene expression and produce short-chain fatty acids that inhibit the growth of opportunistic microbes [33]. However, cold stress and environmental perturbations, such as antibiotic exposure, can disrupt the gut microbiota, leading to dysbiosis and increased pathogen susceptibility, including infections by Nosema spp. and S. marcescens [85]. Bees colonized with a conventional gut microbiota, whether consisting of individual native strains or defined microbial consortia, demonstrate significantly greater survival when challenged with pathogens such as S. marcescens, H. alvei, and Escherichia coli compared to microbiota-depleted individuals [45, 55, 95). In larvae, which lack a mature core microbiota, certain non-core taxa such as Apilactobacillus kunkeei, present in both larvae and adults at low levels, may confer partial protection against Paenibacillus larvae and Melissococcus plutonius, the causative agents of American and European foulbrood, respectively [20,32]. Overall, these findings underscore the central role of the gut microbiome in maintaining honey bee health during overwintering, not only through metabolic support and nutrient recycling but also by strengthening immune function and pathogen resistance under physiologically challenging conditions.
The honey bee gut microbiome is highly sensitive to environmental and developmental stressors, particularly the temperature fluctuations and nutritional limitations characteristic of winter, often resulting in dysbiosis that compromises immune regulation, digestion, and detoxification functions [17,21,22,50,71,84]. As climate change drives more erratic winter conditions, the stability of the honey bee gut ecosystem is increasingly threatened. Such microbial dysbiosis, often subtle but consequential, may contribute to broader patterns of overwintering colony loss that are not fully explained by traditional factors alone. Winter constitutes a critical period for colony survival, and ecological and physiological challenges during this time may contribute to stagnation in the availability of managed honey bees. Among these challenges, disruptions to the gut microbiome have emerged as an underrecognized yet central factor influencing overwintering outcomes. Understanding how gut microbiota respond to winter-associated stressors is crucial not only for bee health but also for the beekeeping industry, where producers are increasingly turning to controlled indoor overwintering at 5–7 °C and 25% relative humidity to improve colony survival [28,76,96]. Despite this progress, it remains unclear how low temperatures, exacerbated by climate change, interact with other biotic and abiotic factors to influence gut microbial dynamics during winter. This review seeks to address these gaps by providing a foundational understanding of how climate change, particularly the trend toward increasingly harsh winters, impacts honey bee health via shifts in the gut microbiome.
3. Microbiome Dynamics in Honey Bees Adapted to Cold Season Spells
Honey bees exhibit distinct seasonal physiological and behavioral adaptations to survive winter conditions. These changes underline critical shifts in bee colony dynamics [17] and often involve seasonal variation in honey bee gut microbiota. Microbial variations in the honey bee gut have been reported to vary by month of the year [10,72,73]. Lactobacillus apis has been reported to be most abundant in October, while unculturable Lactobacillus species peaked in November. In spring, a notable increase in Lactobacillus, Bartonella, and Bifidobacterium spp. was observed, consistent with their roles in metabolism and immunity to pathogens such as Paenibacillus larvae and Melissococcus plutonius [98]. The presence of these bacteria on flowers supports the idea that spring foraging facilitates microbial acquisition [20,53].
Winter bees show increased gut bacterial loads with reduced microbial diversity and elevated levels of specific taxa. Gut microbes such as Gilliamella apicola, Bartonella spp., Commensalibacter spp., and Snodgrassella spp., which support digestion and immune function, are typically found at low densities in spring but become dominant during winter and have been associated with improved honey bee survival under harsh winter conditions [15,60,89]. During extreme cold, Bartonella spp. dominate the honey bee gut, leveraging their expanded metabolic capacity to convert waste products like lactate and ethanol into pyruvate, an energy source for themselves, the host, and other symbionts. Notably, Bartonella is the only gut bacterium capable of producing and secreting tryptophan and phenylalanine, whose metabolites are present in the bee gut despite the absence of relevant digestive enzymes in other microbes [60]. Prolonged cold exposure significantly alters the abundance of core gut bacteria, particularly Lactobacillus Firm-5, a taxon linked to carbohydrate metabolism and immune modulation [50]. Similarly, the densities of Snodgrassella alvi and Gilliamella apicola have been reported to increase significantly under prolonged overwintering conditions compared to the spring season [89]. These microbes are known to be associated with digestion and immunity in insects, suggesting a potential role in enhancing insect survival during harsh winter periods. Notably, indoor wintering (5–7 °C) mitigated some of these disruptions compared to natural colonies exposed to subzero temperatures, suggesting temperature thresholds for microbiome resilience.
Additionally, the bee microbiome has been reported to vary by commercial bee strains. Bolton™ bees show higher levels of Bartonella, Bifidobacterium, and Wolbachia, whereas Mann Lake™ bees have been reported to have a greater abundance of Commensalibacter [10]. Seasonal and geographic shifts in microbiota composition have also been reported to vary with colony management practices and underlying pathogen burdens. These findings highlight the complex interplay between microbial dynamics, bee physiology, and environmental stressors, mainly winter, in shaping honey bee resilience. Together, these studies underscore that winter temperature regimes directly shape microbial composition, with potential cascading effects on host nutrition, pathogen defense, and overwintering survival. Future work should explore whether microbial perturbations persist into spring, affecting colony recovery and pollination capacity.
4. Geographical and Seasonal Dynamics of the Honey Bee Gut Microbiome
The honey bee gut microbiome exhibited clear seasonal and geographical shifts shaped by environmental conditions. In temperate regions, Bifidobacterium abundance declined sharply from summer (15.2%) to winter (4.8%) [60], accompanied by reduced expression of carbohydrate-active enzymes, indicating microbial adjustment to honey-based diets. Large-scale surveys confirmed climate-related patterns, with psychrotolerant Lactobacillus strains (Firm-4 and Firm-5) more abundant in temperate colonies (32–38%) than in tropical ones (18–22%) [78,90].
Marked seasonal restructuring of the gut microbiome was evident across climates, but the trajectories differed by region. In subtropical hives, S. alvi and G. apicola declined steadily as temperatures dropped below 10 °C, whereas Canadian colonies exhibited an increase in Firm-4 abundance along with upregulation of glycogen fermentation pathways [7,17]. These contrasting patterns suggest region-specific microbial strategies to sustain host metabolism during cold stress. Functional analyses further supported this view: winter microbiomes were enriched in carbohydrate metabolism pathways (e.g., trehalose and glycogen breakdown), while summer microbiomes displayed greater capacity for amino acid biosynthesis [50]. Importantly, depletion experiments demonstrated that microbiome loss reduced cold tolerance, with winter bees showing significantly lower survival at low temperatures (45% vs. 78%) [50]. Collectively, these results suggest that gut microbial communities actively contribute to overwintering resilience through metabolic flexibility, though the precise mechanisms remain unresolved.
Despite these advances, major gaps remain in our understanding of microbiome-mediated cold adaptation. Most existing studies have focused on temperate and subtropical regions, while arctic and tropical alpine bees, facing distinct and extreme cold stress conditions, remain virtually unexplored. These extreme populations may harbor novel microbial taxa or symbiotic strategies that could revolutionize our understanding of cold resilience. Second, while correlations between microbiome shifts and winter survival are well-documented, the mechanistic links remain unclear. Does winter microbiome restructuring directly enhance host cold tolerance through specific metabolic outputs (e.g., cryoprotectant production), or are these changes merely secondary effects of dietary shifts? Controlled experiments using microbiota transplants (e.g., inoculating winter bees with summer-derived microbial communities) could help disentangle these relationships. Third, the recovery dynamics of winter-disrupted microbiomes in spring and their consequences for colony fitness are virtually unstudied. Given that early spring is a critical period for colony rebuilding and pollination services, understanding whether and how quickly microbial communities rebound could have major implications for hive management. Preliminary data suggest some core taxa like S. alvi may take weeks to recover to pre-winter levels, potentially creating a "microbial bottleneck" that affects colony health [7,50].
Addressing these questions through integrated field studies and manipulative experiments will not only advance fundamental knowledge of host-microbe interactions under climate stress but could also lead to practical innovations. For instance, identifying microbial taxa that promote cold tolerance might enable the development of probiotic supplements to boost overwintering success in managed colonies, a crucial need as climate change increases winter volatility. Similarly, understanding geographical variation in microbiome resilience could inform breeding programs or hive placement strategies to optimize pollinator health in different regions.
5. Modulators of the Honeybee Microbiome During Overwintering
5.1. Extrinsic Stressors in Honey Bees: Implications for Overwintering Survival and Gut Microbial Shifts
5.1.1. Cold Exposure and Dietary Modulation
Nutrition is a central factor of honeybee health, with extensive studies and recent reviews highlighting that a diverse pollen diet enhances both overall fitness and resistance to pathogens [12,26,29,66]. Since the gut microbiome plays a central role in nutrient metabolism, it is reasonable to hypothesize that dietary shifts may directly influence both the composition and functional capacity of the microbiome during overwintering. The overwintering period induces major physiological and behavioral changes in honey bee colonies, including the production of longer-lived workers, a dietary shift to stored pollen and supplemental feed, and an extended phase of reduced foraging activity. During winter, bees experience reduced pollen intake, and pollen diet reserves decline, and honey bees switch to a carbohydrate-based diet. This led to a shift in the gut microbiome, which could influence the health and safety of colonies during overwintering. A shift in the gut microbial community is determined by the type of carbohydrate consumed by honey bees during winter. Sucrose has been identified as a suitable overwintering food, as it supports the presence of beneficial bacteria such as Alphaproteobacteria, Bifidobacteriales, and Lactobacillaceae [99]
Bees fed with beebread exhibited greater microbiota stability and reduced mortality rates following N. ceranae infection compared to those provided with sugar syrup. Furthermore, the relative abundance of beneficial bacterial genera such as Lactobacillus and Snodgrassella was significantly higher in bees consuming beebread. In contrast, bees maintained on a sugar-only diet showed an increased prevalence of the genus Serratia (family Enterobacteriaceae), which is often associated with gut dysbiosis [46].
Furthermore, the pollen quantity and nutritional value are the key determinants of gut microbiota composition in honeybees. Pollen deprivation leads to marked disruptions in both the composition and metabolic function of the honey bee gut microbiome. Bees subjected to pollen starvation exhibit a significant decline in the relative abundance of core bacterial taxa, including Lactobacillus, Bombilactobacillus, Bifidobacterium, Gilliamella, and Snodgrassella. This microbial imbalance is accompanied by reduced expression of key fermentative enzymes, indicating a diminished capacity for nutrient metabolism and overall gut functionality [16,17,87]. A pollen-free diet, while increasing overall microbial load, has been shown to significantly reduce microbial diversity and evenness. This shift includes a marked decline in the relative abundance of beneficial, hive-associated bacterial taxa [81]. In parallel, studies have demonstrated that pollen substitutes can substantially alter microbial community composition when compared to natural pollen. Specifically, the relative abundance of core beneficial taxa such as Snodgrassella alvi and Gilliamella apicola was significantly lower in bees fed pollen substitutes [54,105]. In contrast, opportunistic and potentially pathogenic bacteria, including Frischella perrara and Lactobacillus kunkeei, increased in abundance under substitute diets [85]. These shifts suggest that pollen substitutes, while sustaining basic energy needs, fail to support the complex microbial communities necessary for optimal bee health. Moreover, feeding nurse bees aged pollen causes a reduction of Snodgrassella alvi in the honeybee gut, and an increase of Frischella perrara, a bacterium associated with impaired development, increased mortality, and a higher risk of Nosema infection [64].
Overall, it is clear that long-term use of pollen substitutes or nutritionally compromised pollen (e.g., aged pollen) leads to persistent dysbiosis in the honey bee gut microbiota, which in turn compromises host immunity and overall colony resilience. These findings underscore the critical importance of diet quality in maintaining a healthy and balanced gut microbiota in honey bees. Current research lacks comprehensive insights into how different dietary modulation, along with overwintering stress and associated gut microbiota alterations, impact bee physiology, immunity, gene expression, and behavior. Future studies in this area could facilitate the breeding or selection of honey bee strains with enhanced resilience to environmental and nutritional challenges. Key hypotheses for future research should focus on: (1) identifying specific microbial signatures, such as reduced or overrepresented taxa, that could serve as biomarkers of dietary stress, thereby enabling early detection and targeted intervention strategies in beekeeping practices; and (2) determining whether particular microbial taxa or diversity indices (e.g., low Snodgrassella alvi or high Frischella perrara) are consistently associated with dietary stress and colony health deterioration during overwintering.
5.1.2. Cold Exposure and Antibiotic Stress
In the United States, antibiotic treatment of bee colonies has been used extensively for more than 50 years to prevent foulbrood (Paenibacillus larvae), a bacterial illness of bee larvae [36]. Broad-spectrum activity of antibiotics can inadvertently disrupt the gut microbiota of both adult and larval honeybees, leading to poor development, higher mortality, and Nosema illness. Tetracycline, for instance, has been widely used in apiculture since the 1950s in some countries and has been shown to significantly reduce the abundance of core gut bacteria such as Snodgrassella alvi, Bifidobacterium, Lactobacillus, and Bombilactobacillus species. This microbial dysbiosis is associated with increased bee mortality and heightened susceptibility to opportunistic pathogens such as Serratia marcescens [85] Tylosin tartrate antibiotic treatment significantly reduces bacterial species richness and phylogenetic diversity in the honey bee gut microbiome; in particular, it decreases both the absolute abundance and strain-level diversity of beneficial core taxa such as Snodgrassella alvi and Bifidobacterium spp. Moreover, bees from tylosin-treated hives exhibited higher mortality when challenged with the opportunistic pathogen Serratia marcescens under controlled laboratory conditions [81]. Additionally, exposure to penicillin–streptomycin mixtures has been shown to increase honeybee susceptibility to Nosema ceranae infection and suppress the expression of key host antimicrobial peptides, including abaecin, defensin-1, and hymenoptaecin, under laboratory conditions [61].
Furthermore, cold stress exacerbates the negative effects of antibiotic treatment on the microbiome by impairing metabolic activity and weakening immune response [104]. While direct studies on the combined effects of antibiotics and cold stress on honey bees have not been conducted yet, the individual impacts suggest potential synergistic detriments. Cold stress can impair antioxidant defenses and development, while antibiotics disrupt gut microbiota and immune function. The intersection of these stressors could exacerbate health issues in honey bees, warranting further research into their combined effects. Understanding the combined effects of antibiotics and cold stress is crucial for developing strategies to maintain microbial homeostasis and prevent associated health disorders in honey bee colonies. Future research should address the following key questions: (1) What are the long-term ecological consequences of routine antibiotic use on microbial diversity in overwintering honey bees? (2) Can restoration of the gut microbiota, such as through probiotic supplementation, mitigate the negative impacts of antibiotic exposure and cold stress? (3) Does antibiotic use contribute to the emergence of antibiotic-resistant opportunistic pathogens in overwintering bees, thereby increasing the risk of winter colony collapse?
5.1.3. Cold Exposure and Pathogen Infestation
Changes in gut microbial structure during cold months may predispose colonies to winter loss, especially when combined with pathogen infestation. Honey bees are vulnerable to a wide range of pathogens and pests, including bacteria, fungi, parasitic mites, and beetles. Among bacterial pathogens, Paenibacillus larvae and Melissococcus plutonius are the causative agents of American foulbrood and European foulbrood, respectively, both highly contagious diseases that primarily affect bee larvae and pose significant threats to colony development and survival [39,40,77]. In addition to bacterial infections, honeybees are also susceptible to fungal pathogens such as Ascosphaera apis, which causes chalkbrood disease [3], as well as parasitic mites like Varroa destructor (Rosenkranz et al., 2010), and invasive pests including the small hive beetle (Aethina tumida) [75]. These biotic stressors contribute synergistically to colony health decline and are often implicated in broader syndromes such as colony collapse disorder.
The collective findings from the literature on the effects of various pathogens on honey bee physiological parameters and gut microbiome composition are summarized in Table 1. Overwintering losses in honey bee colonies have been strongly associated with the invasive ectoparasitic mite Varroa destructor and the array of viruses it vectors [43], most notably the deformed wing virus (DWV) [74], which has reached panzootic levels in bee populations infested by V. destructor. The gut microbiota modulates the virulence of a major honey bee viral pathogen, deformed wing virus (DWV) [30].
However, studies investigating the interactive effects of pathogens and winter-specific stressors on the gut microbiome remain limited. Despite this gap, the existing evidence provides a foundation to formulate several hypotheses that can inform and guide future research on the complex interplay between pathogen pressure, microbial
community dynamics, and overwintering stress in honey bees. From the available data, we propose three hypotheses that warrant investigation. First, we hypothesize that pathogen-induced dysbiosis, such as the reduction of Lactobacillus or Commensalibacter due to Nosema ceranae or viral infections, may be exacerbated under cold winter conditions, leading to compounded effects on immunity and nutrient assimilation. While Nosema infection alone decreases microbial diversity [46,57] cold stress further reduces beneficial taxa like Commensalibacter and Snodgrassella (Carlini et al., 2024), potentially pushing colonies past a critical threshold for survival. Finally, we hypothesize that the effectiveness of protective microbial strains (e.g., G. apicola W8136, L. apis W8172 [56] against opportunistic pathogens can be modulated by winter temperature and hive microclimate, potentially limiting their probiotic potential under suboptimal overwintering conditions. These hypotheses highlight the need for multifactorial studies integrating pathogen pressure, overwintering stress, microbial composition, and colony survival outcomes.
5.2. Intrinsic Stressors in Honey Bees: Implications for Overwintering Survival and Gut Microbial Shifts
5.2.1. Cold Stress and Honey Bee Age
As worker honey bees age, their gut microbiota transitions from a proteobacterial-dominated community (e.g., Snodgrassella) to one enriched in lactic acid bacteria (Lactobacillus spp.), reflecting changes in diet and metabolic needs [31,48]. Cold exposure exacerbates these age-related changes, as low temperatures reduce foraging activity, alter nutrient availability, and suppress immune function, potentially disrupting microbial symbionts critical for energy harvest and pathogen defense [70]. The interplay between aging, cold stress, and microbiome stability remains poorly understood in winter bees, which face unique survival pressures due to their extended lifespans and colony thermoregulation demands. Overwintering worker bees experience age-related shifts in their hindgut microbiota, with older individuals showing reduced microbial diversity when exposed to cold stress [65]. In particular, Lactobacillus carbohydrate metabolism spp., essential for carbohydrate metabolism declined sharply during winter, potentially limiting energy acquisition [50]. Cold exposure further intensified dysbiosis in aged bees, indicating a synergistic interaction between age and low temperature in destabilizing microbiome composition. Building on these findings, Anderson & Maes (2022) reported that both age and climate influence not only gut communities but also social microbiota, including those in the mandibular glands. Winter bees showed restructured microbial networks in these glands, accompanied by shifts in immune gene expression. Notably, younger winter bees retained more stable microbial communities, suggesting an adaptive mechanism for cold tolerance. Despite these insights, the causal links between cold-induced physiological aging, such as oxidative stress, and microbiome collapse in winter bees remain unexplored. Testing the following hypotheses could clarify the interplay between climate stress, aging, and microbial resilience.
(1) Does cold accelerate senescence in gut epithelial cells, disrupting microbial symbiosis?
Aged epithelial cells may exhibit impaired barrier function or nutrient secretion, compromising colonization resistance for core symbionts like Lactobacillus while permitting pathogen invasion (e.g., Serratia marcescens). Such mechanisms could mirror age-related dysbiosis in mammals but remain unexplored in insects.
(2) Could probiotic interventions offset winter dysbiosis in aged workers?
Additionally, specific bacterial strains may prime immune responses in age-specific cohorts of worker bees via activation of the Toll signaling pathway, thereby enhancing pathogen defense during periods of winter-associated immunosuppression. However, several practical challenges remain, including the strain-specific variability in immunomodulatory efficacy, reduced bacterial viability at low ambient temperatures, and the need for delivery methods compatible with winter cluster behavior. During this period, honey bee foraging activity is minimized, and oral uptake is significantly reduced, further complicating effective probiotic administration across a population comprising workers of varying ages.
Addressing these knowledge gaps could transform strategies for safeguarding honey bee populations against climate instability. Elucidating cold stress-induced microbiome aging may enable: predictive biomarkers using microbial signatures (e.g., Lactobacillus depletion or fungal proliferation) as early collapse indicators; targeted microbial therapies, including probiotic blends to restore cold-impaired functions (glycogen metabolism, oxidative stress mitigation) and prebiotics to nourish cryoprotective symbionts; and climate-adaptive beekeeping through microbiome-informed hive insulation, winter feeding, or breeding of cold-resilient colonies.
5.2.2. Cold Stress and Honey Bee Strain
The core microbial communities of honey bees exhibit notable interspecific variation, reflecting the influence of host biology and environment. For example, Apis cerana (Eastern honey bee) and Apis mellifera (Western honey bee), though closely related phylogenetically, harbor distinct gut microbiota compositions and functional profiles [55, 101). These differences are shaped by multiple factors, including host genetics, geographic distribution, and environmental conditions [102]. Recognizing such interspecific variation is critical for understanding host–microbe interactions and for developing strategies to strengthen bee health and resilience against colony losses.
Recent work by Zhang et al. (2023) [101] provides further insight by examining the gut microbiota of overwintering A. cerana and A. mellifera to investigate their mechanisms of cold tolerance. Using 16S rRNA gene amplicon sequencing and metagenomic approaches, they compared microbial diversity and community composition during the winter months. While both species shared a conserved core microbiota, including the genera Gilliamella, Lactobacillus, Bifidobacterium, and Snodgrassella, significant differences emerged in the relative abundance of specific bacterial groups. For instance, Bartonella and Commensalibacter were enriched in A. cerana, whereas Lactobacillus species dominated in A. mellifera [42,55,101,102].
These variations suggest species-specific adaptations that may contribute to differential cold tolerance and overwintering success. Functional analysis indicated that pathways related to amino acid metabolism, carbohydrate transport, and the tricarboxylic acid (TCA) cycle were enriched in both species, highlighting the metabolic adaptations that support overwintering survival. The study underscores the importance of gut microbiota in mediating physiological responses to environmental stressors and provides insights into the microbial factors influencing overwintering success in honey bees.
Moreover, the hindgut microbiota of overwintering honey bee workers from different climates also showed variability. The results indicated that the hindgut microbiota of workers overwintered in cold indoor environments remained stable and resistant to fungal proliferation, suggesting a robust microbiota in cold conditions. In contrast, workers overwintering in warm outdoor climates exhibited increased fungal abundance and changes in bacterial composition, indicating that warmer climates may compromise the stability of the hindgut microbiota [65]. While existing studies have begun to elucidate the differences in gut microbiota between A. cerana and A. mellifera, particularly concerning overwintering periods, several questions remain unanswered. Future research should focus on: (1) Longitudinal Studies: Investigating how gut microbiota composition changes throughout the entire overwintering period and correlating these changes with colony health and survival rates (2) Functional Analyses: Conducting metatranscriptomic and metabolomic studies to understand the functional roles of specific microbial taxa in supporting bee physiology during cold stress (3) Host-Microbe Interactions: Exploring how host genetics influence gut microbiota assembly and function, and how these interactions affect overwintering success [105].(4) Microbial Interventions: Assessing the potential of probiotic or prebiotic treatments to modulate gut microbiota and enhance overwintering resilience in different bee species shown in Wang et al., 2020 [99]. Addressing these questions will provide a comprehensive understanding of the gut microbiota's role in honey bee health and contribute to the development of strategies to mitigate colony losses during overwintering.
6. Gene Expression Under Cumulative Stress Response in Overwintering Honeybees
Cumulative stress during overwintering induces widespread transcriptional changes in honey bees that reflect their physiological adaptations to environmental adversity. Cold exposure, in particular, upregulates genes involved in metabolic regulation, thermogenesis, and immune function, including those related to fatty acid metabolism and mitochondrial activity [1]. Recent studies reveal that the combined effects of cold, poor nutrition, pathogens, and antibiotics further reshape the honey bee transcriptome, often suppressing key immune and detoxification pathways [24,92]. These gene expression changes provide critical insights into the molecular mechanisms underpinning honey bee resilience or vulnerability during overwintering. Table 2 summarizes key findings from the literature on gene expression responses to combined overwintering stressors.
These findings suggest that cold not only imposes direct metabolic constraints but also potentiates the negative effects of pathogens by impairing gene regulatory networks associated with immunity, stress tolerance, and longevity. This synergistic impact may highlight the complex interplay between environmental stressors and host-microbiome interactions during overwintering. Understanding how cold stress interacts with other factors to shape gut microbial dynamics and host gene expression is critical for unraveling the mechanisms that determine colony survival. Based on current evidence, several key questions emerge that warrant focused investigation:
- How does cold-induced disruption of the gut microbiome alter immune signaling pathways and antimicrobial peptide expression, thereby influencing susceptibility to opportunistic pathogens under combined pesticide and pathogen exposure?
- To what extent does the gut microbiome contribute to metabolic reprogramming and oxidative stress resilience by modulating host gene expression related to mitochondrial function and detoxification during cold stress?
- How do combined overwintering stressors drive gut microbial dysbiosis that impacts host detoxification gene networks, and what are the downstream effects on neural health and longevity in honey bees?
Addressing these hypotheses will advance our understanding of the molecular and microbial mechanisms underpinning honey bee resilience or vulnerability to winter stressors and inform strategies to mitigate colony losses in a changing climate.
Overall, this study highlights the key microbial players in winter honey bee colonies and their potential role in guiding microbiome-based interventions. Bartonella apis dominates winter microbiomes through its unique ability to metabolize hive-stored resources and synthesize essential amino acids [10,60], while Gilliamella apicola supports polysaccharide digestion and detoxification [50]. Psychrotolerant Lactobacillus strains from the Firm-4/Firm-5 clades (e.g., L. apis, L. mellifer) thrive under cold conditions, helping stabilize gut pH and suppress pathogens [7]. Declines in Snodgrassella alvi signal pollen scarcity and compromised gut barrier function [89]. Commensalibacter spp. contribute acetate for energy and antioxidant defense [101], and Bifidobacterium spp., although reduced by pollen deprivation, remain vital for sugar metabolism and immunity [60]. In contrast,

Table 2.
Gene expression responses in honey bees under combined overwintering stressors.
| Stressor Type | Cold Stress specific or Combined | Targeted Pathways / Systems | Key Genes Affected | Expression Response | Functional Consequences | Reference |
|---|---|---|---|---|---|---|
| Cold Stress | Cold Stress specific | Mitochondrial metabolism, immunity | Vitellogenin,Defensin-1, Cox5a | ↑ Upregulated | Enhanced immunity, metabolic reprogramming | [37] |
| Cold Stress specific | Antifreeze protein (AFP), protein lethal (2) (I(2)efl), vitellogenin (Vg) | ↑ Upregulated | Enhanced cold resistant ability | [82] | ||
| Pathogens (DWV, bacteria) | Cold + pathogen | Cellular immune function |
Defensin-1, Hymenoptaecin, Dorsal, eater |
↑ Upregulated | Enhanced immunity | [97] |
| Antibiotics (tetracycline) | Combined with Cold | Gut microbiota, immune modulation |
Defensin-1, AMPs, Lysozyme, PGRP-LC |
↓Suppressed AMPs ↑ Detox gene variability |
Microbiome disruption, weakened immunity, impaired digestion | [72] |
opportunistic taxa such as Frischella perrara and Enterobacteriaceae proliferate under stressors like cold, and malnutrition increasing winter mortality risks [49,65]. Collectively, these findings indicate that targeted microbiome-based strategies, including probiotics (Lactobacillus, Bifidobacterium), pollen supplementation, and controlled indoor overwintering, should be prioritized to enhance colony health and resilience during winter.
7. Conclusion
This review highlights the central role of the gut microbiome in mediating honey bee resilience to increasingly harsh overwintering conditions driven by climate change. Cold stress, compounded by poor nutrition, pathogens, antibiotics, and genetic variability, disrupts microbial homeostasis, suppresses immune and detoxification pathways, and elevates colony mortality. Although individual stressors have been well characterized, the synergistic effects of winter remain poorly understood. The gut microbiome represents a critical interface between environmental conditions and host physiology, and addressing this knowledge gap is essential for developing microbiome-informed strategies to enhance winter survival. Future research should integrate microbial composition with host gene expression and ecological outcomes. Microbial biomarkers, such as Bartonella/Gilliamella ratios and Snodgrassella alvi abundance, offer promising predictive tools, while engineered symbionts capable of chemical degradation or immune enhancement represent an innovative frontier. Grounding these approaches in a robust ecological framework will enable precision interventions to improve colony health, mitigate overwintering losses, and safeguard the essential ecosystem services provided by honey bees.
Author Contributions
Gagandeep Brar: Conceptualization (lead); writing – original draft (lead); Ramandeep Kaur: writing, review and editing (equal); Mandeep Kaur Gill: writing, review and editing (equal); Rupinderjeet Kaur: writing, review and editing (equal); Navjot Singh: writing, review and editing (equal). All authors read and approved the final version.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data generated.
Acknowledgments
We thank Dr. Arun Rajamohan for reviewing the final document.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: digital gene expression analysis of pollen's nutritive effects on healthy and varroa-parasitized bees. BMC Genomics 2011, 12, 496. [Google Scholar] [CrossRef]
- Anderson, K. E.; Maes, P. Social microbiota and social gland gene expression of worker honey bees by age and climate. Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Aronstein, K. A.; Murray, K. D. Chalkbrood disease in honey bees. J. Invertebr. Pathol. 2010, 103, S20–S29. [Google Scholar] [CrossRef] [PubMed]
- ARS Honey Bee Health. Available online: https://www.ars.usda.gov/oc/br/ccd/index/ (accessed on 9 June 2025).
- Navarro-Escalante, L.; Ashraf, A. H. M. Z.; Leonard, S. P.; Barrick, J. E. Protecting honey bees through microbiome engineering. Curr. Opin. Insect Sci. 2025, 72, 101416. [Google Scholar] [CrossRef]
- Blackport, R.; Fyfe, J. C. Amplified warming of North American cold extremes linked to human-induced changes in temperature variability. Nat. Commun. 2024, 15, 1–13. [Google Scholar] [CrossRef]
- Bleau, N.; Bouslama, S.; Giovenazzo, P.; Derome, N. Dynamics of the honeybee (Apis mellifera) gut microbiota throughout the overwintering period in Canada. Microorganisms 2020, 8, 1146. [Google Scholar] [CrossRef]
- Bonilla-Rosso, G.; Engel, P. Functional roles and metabolic niches in the honey bee gut microbiota. Curr. Opin. Microbiol. 2018, 43, 69–76. [Google Scholar] [CrossRef]
- Borg, Z. Climate Change and Bees: The Effects of a changing planet. The Best Bees Company. Available online: https://bestbees.com/climate-change-and-bees/ (accessed on 19 August 2024).
- Brar, G.; Ngor, L.; McFrederick, Q. S.; Torson, A. S.; Rajamohan, A.; Rinehart, J.; Singh, P.; Bowsher, J. H. High abundance of lactobacilli in the gut microbiome of honey bees during winter. Sci. Rep. 2025, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Brochet, S.; Quinn, A.; Mars, R. A.; Neuschwander, N.; Sauer, U.; Engel, P. Niche partitioning facilitates coexistence of closely related honey bee gut bacteria. eLife 2021, 10, e68583. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Budge, G. E.; Pietravalle, S.; Brown, M.; Laurenson, L.; Jones, B.; Tomkies, V.; Delaplane, K. S. Pathogens as predictors of honey bee colony strength in England and Wales. PLoS ONE 2015, 10, e0133228. [Google Scholar]
- Butolo, N. P.; Azevedo, P.; Alencar, L. D.; Malaspina, O.; Nocelli, R. C. F. Impact of low temperatures on the immune system of honeybees. J. Therm. Biol. 2021, 101, 103082. [Google Scholar] [CrossRef]
- Carlini, D. B.; Winslow, S. K.; Cloppenborg-Schmidt, K.; Baines, J. F. Quantitative microbiome profiling of honey bee (Apis mellifera) guts is predictive of winter colony loss in northern Virginia (USA). Sci. Rep. 2024, 14, 1–12. [Google Scholar] [CrossRef]
- Castelli, L.; Branchiccela, B.; Garrido, M.; Invernizzi, C.; Porrini, M.; Romero, H.; Santos, E.; Zunino, P.; Antúnez, K. Impact of nutritional stress on honeybee gut microbiota, immunity, and Nosema ceranae infection. Microb. Ecol. 2020, 80, 908–919. [Google Scholar] [CrossRef]
- Castelli, L.; Branchiccela, B.; Romero, H.; Zunino, P.; Antúnez, K. Seasonal dynamics of the honey bee gut microbiota in colonies under subtropical climate. Microb. Ecol. 2022, 83, 492–500. [Google Scholar] [PubMed]
- Cohen, J.; Francis, J. A.; Pfeiffer, K. Anomalous Arctic warming linked with severe winter weather in Northern Hemisphere continents. Commun. Earth Environ. 2024, 5, 1–14. [Google Scholar] [CrossRef]
- Cohen, J.; Pfeiffer, K.; Francis, J. A. Warm Arctic episodes linked with increased frequency of extreme winter weather in the United States. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Corby-Harris, V.; Maes, P.; Anderson, K. E. The bacterial communities associated with honey bee (Apis mellifera) foragers. PLoS ONE 2014, 9, e95056. [Google Scholar]
- Cormier, S. B.; Léger, A.; Boudreau, L. H.; Pichaud, N. Overwintering in North American domesticated honeybees (Apis mellifera) causes mitochondrial reprogramming while enhancing cellular immunity. J. Exp. Biol. 2022, 225, jeb244440. [Google Scholar] [CrossRef]
- Coulibaly, K. A. S.; Majeed, M. Z.; Sayed, S.; Yeo, K. Simulated climate warming influenced colony microclimatic conditions and gut bacterial abundance of honeybee subspecies Apis mellifera ligustica and A. mellifera sinisxinyuan. J. Apic. Sci. 2022, 66, 27. [Google Scholar] [CrossRef]
- Cox-Foster, D. L.; Conlan, S.; Holmes, E. C.; Palacios, G.; Evans, J. D.; Moran, N. A.; Lipkin, W. I. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef]
- Crailsheim, K.; Stolberg, E. Influence of diet, age and colony condition upon intestinal proteolytic activity and size of the hypopharyngeal glands in the honeybee (Apis mellifera L.). J. Insect Physiol. 1989, 35, 595–602. [Google Scholar] [CrossRef]
- Crowley, L. M.; Sadler, J. P.; Pritchard, J.; Hayward, S. A. L. Elevated CO2 impacts on plant-pollinator interactions: a systematic review and free air carbon enrichment field study. Insects 2021, 12, 512. [Google Scholar] [CrossRef]
- Dalmon, A.; Desbiez, C.; Coulon, M.; Thomasson, M.; Le Conte, Y.; Alaux, C.; Vallon, J.; Moury, B. Evidence for positive selection and recombination hotspots in Deformed wing virus (DWV). Sci. Rep. 2017, 7, 41045. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Chen, Y.; Rivera, R.; Carroll, M.; Chambers, M.; Hidalgo, G.; de Jong, E. W. Honey bee colonies provided with natural forage have lower pathogen loads and higher overwinter survival than those fed protein supplements. Apidologie 2016, 47, 186–196. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Graham, H.; Ahumada, F.; Smart, M.; Ziolkowski, N. The economics of honey bee (Hymenoptera: Apidae) management and overwintering strategies for colonies used to pollinate almonds. J. Econ. Entomol. 2019, 112, 2524–2533. [Google Scholar] [CrossRef]
- Dolezal, A. G.; Toth, A. L. Feedbacks between nutrition and disease in honey bee health. Curr. Opin. Insect Sci. 2018, 26, 114–119. [Google Scholar] [CrossRef]
- Dosch, C.; Manigk, A.; Streicher, T.; Tehel, A.; Paxton, R. J.; Tragust, S. The gut microbiota can provide viral tolerance in the honey bee. Microorganisms 2021, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Emery, O.; Schmidt, K.; Engel, P. Immune system stimulation by the gut symbiont Frischella perrara in the honey bee (Apis mellifera). Mol. Ecol. 2017, 26, 2576–2590. [Google Scholar] [CrossRef]
- Endo, A.; Salminen, S. Honeybees and beehives are rich sources for fructophilic lactic acid bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N. A. Functional and evolutionary insights into the simple yet specific gut microbiota of the honey bee from metagenomic analysis. Gut Microbes 2013, 4, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Kwong, W. K.; McFrederick, Q.; Anderson, K. E.; Barribeau, S. M.; Chandler, J. A.; Cornman, R. S.; Dainat, J.; De Miranda, J. R.; Doublet, V.; Emery, O.; Evans, J. D.; Farinelli, L.; Flenniken, M. L.; Granberg, F.; Grasis, J. A.; Gauthier, L.; Hayer, J.; Koch, H.; Dainat, B. The bee microbiome: impact on bee health and model for evolution and ecology of host-microbe interactions. mBio 2016, 7, e02164-15. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Martinson, V. G.; Moran, N. A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef]
- Evans, J. D. Diverse origins of tetracycline resistance in the honey bee bacterial pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Forrest, J. R. Plant–pollinator interactions and phenological change: what can we learn about climate impacts from experiments and observations? Oikos 2015, 124, 4–13. [Google Scholar] [CrossRef]
- Forsgren, E. European foulbrood in honey bees. J. Invertebr. Pathol. 2010, 103, S5–S9. [Google Scholar] [CrossRef]
- Genersch, E. American foulbrood in honeybees and its causative agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef] [PubMed]
- Gorrochategui-Ortega, J.; Muñoz-Colmenero, M.; Kovačić, M.; Filipi, J.; Puškadija, Z.; Kezić, N.; Parejo, M.; Büchler, R.; Estonba, A.; Zarraonaindia, I. A short exposure to a semi-natural habitat alleviates the honey bee hive microbial imbalance caused by agricultural stress. Sci. Rep. 2022, 12, 18832. [Google Scholar] [CrossRef]
- Lee, F. J.; Rusch, D. B.; Stewart, F. J.; Mattila, H. R.; Newton, I. L. G. Saccharide breakdown and fermentation by the honey bee gut microbiome. Environ. Microbiol. 2015, 17, 796–815. [Google Scholar] [CrossRef]
- Highfield, A. C.; Nagar, A. E.; Mackinder, L. C. M.; Noël, L. M. J.; Hall, M. J.; Martin, S. J.; Schroeder, D. C. Deformed wing virus implicated in overwintering honeybee colony losses. Appl. Environ. Microbiol. 2009, 75, 7212–7220. [Google Scholar] [CrossRef]
- Hill, D. B.; Webster, T. C. Apiculture and forestry (bees and trees). Agrofor. Syst. 1995, 29, 313–320. [Google Scholar] [CrossRef]
- Horak, R. D.; Leonard, S. P.; Moran, N. A. Symbionts shape host innate immunity in honeybees. Proc. R. Soc. B 2020, 287, 20201184. [Google Scholar] [CrossRef] [PubMed]
- Huang, S. K.; Ye, K. T.; Huang, W. F.; Ying, B. H.; Su, X.; Lin, L. H.; Li, J. H.; Chen, Y. P.; Li, J. L.; Bao, X. L.; Hu, J. Z. Influence of feeding type and Nosema ceranae infection on the gut microbiota of Apis cerana workers. mSystems 2018, 3, e00177-18. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S. L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M. I., Huang, K., Leitzell, K., Lonnoy, E., Matthews, J. B. R., Maycock, T. K., Waterfield, T., Yelekçi, O., Yu, R., Zhou, B., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Jones, J. C.; Fruciano, C.; Marchant, J.; Hildebrand, F.; Forslund, S.; Bork, P.; Hughes, W. O. H. The gut microbiome is associated with behavioral tasks in honey bees. Insectes Soc. 2018, 65, 419–429. [Google Scholar] [CrossRef]
- Romero, S.; Nastasa, A.; Chapman, A.; Kwong, W. K.; Foster, L. J. The honey bee gut microbiota: strategies for study and characterization. Environ. Microbiol. 2019, 28, 455–472. [Google Scholar] [CrossRef]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut microbiota structure differs between honeybees in winter and summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef]
- Kešnerová, L.; Mars, R. A. T.; Ellegaard, K. M.; Troilo, M.; Sauer, U.; Engel, P. Disentangling metabolic functions of bacteria in the honey bee gut. PLoS Biol. 2017, 15, e2003467. [Google Scholar] [CrossRef]
- Kwong, W. K.; Medina, L. A.; Koch, H.; Sing, K.-W.; Soh, E. J. Y.; Ascher, J. S.; Jaffé, R.; Moran, N. A. Dynamic microbiome evolution in social bees. Sci. Adv. 2017, 3, e1600513. [Google Scholar] [CrossRef]
- Killer, J.; Dubná, S.; Sedláček, I.; Švec, P. Lactobacillus apis sp. nov., from the stomach of honeybees (Apis mellifera), having an in vitro inhibitory effect on the causative agents of American and European foulbrood. Int. J. Syst. Evol. Microbiol. 2013, 64, 152–157. [Google Scholar] [CrossRef]
- Kwong, W. K.; Moran, N. A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W. K.; Mancenido, A. L.; Moran, N. A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef]
- Lang, H.; Duan, H.; Wang, J.; Zhang, W.; Guo, J.; Zhang, X.; Hu, X.; Zheng, H. Specific strains of honeybee gut Lactobacillus stimulate host immune system to protect against pathogenic Hafnia alvei. Microbiol. Spectr. 2022, 10, e01896-21. [Google Scholar] [CrossRef]
- Lau, E.; Maccaro, J.; McFrederick, Q. S.; Nieh, J. C. Exploring the interactions between Nosema ceranae infection and the honey bee gut microbiome. Sci. Rep. 2024, 14, 1–14. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: impact on honey bee populations and diseases. Rev. Sci. Tech. 2008, 27, 499–510. [Google Scholar]
- Lee, F. J.; Miller, K. I.; McKinlay, J. B.; Newton, I. L. G. Differential carbohydrate utilization and organic acid production by honey bee symbionts. FEMS Microbiol. Ecol. 2018, 94, fiy113. [Google Scholar] [CrossRef]
- Li, C.; Tang, M.; Li, X.; Zhou, X. Community dynamics in structure and function of honey bee gut bacteria in response to winter dietary shift. mBio 2022, 13, e01131-22. [Google Scholar] [CrossRef]
- Li, J. H.; Evans, J. D.; Li, W. F.; Zhao, Y. Z.; DeGrandi-Hoffman, G.; Huang, S. K.; Li, Z. G.; Hamilton, M.; Chen, Y. P. New evidence showing that the destruction of gut bacteria by antibiotic treatment could increase the honey bee’s vulnerability to Nosema infection. PLoS ONE 2017, 12, e0187505. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zheng, M.; Cai, M.; Zhang, Y.; Fan, Y.; Lin, Z.; Wang, Z.; Niu, Q.; Ji, T. Possible interactions between gut microbiome and division of labor in honey bees. Ecol. Evol. 2024, 14, e11707. [Google Scholar] [CrossRef] [PubMed]
- Smutin, D.; Lebedev, E.; Selitskiy, M.; Panyushev, N.; Adonin, L. Micro”bee”ota: honey bee normal microbiota as a part of superorganism. Microorganisms 2022, 10, 2359. [Google Scholar] [CrossRef]
- Maes, P. W.; Floyd, A. S.; Mott, B. M.; Anderson, K. E. Overwintering honey bee colonies: effect of worker age and climate on the hindgut microbiota. Insects 2021, 12, 224. [Google Scholar] [CrossRef]
- Maes, P. W.; Rodrigues, P. A. P.; Oliver, R.; Mott, B. M.; Anderson, K. E. Diet-related gut bacterial dysbiosis correlates with impaired development, increased mortality and Nosema disease in the honeybee (Apis mellifera). Mol. Ecol. 2016, 25, 5439–5450. [Google Scholar] [CrossRef]
- Meehan, D. E.; O'Toole, P. W. A review of diet and foraged pollen interactions with the honeybee gut microbiome. Microb. Ecol. 2025, 88, 54. [Google Scholar] [CrossRef] [PubMed]
- Harwood, G.; Salmela, H.; Freitak, D.; Amdam, G. Social immunity in honey bees: royal jelly as a vehicle in transferring bacterial pathogen fragments between nestmates. J. Exp. Biol. 2021, 224, jeb231076. [Google Scholar] [CrossRef]
- Memmott, J.; Craze, P. G.; Waser, N. M.; Price, M. V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Minaud, É.; Rebaudo, F.; Davidson, P.; Hatjina, F.; Hotho, A.; Mainardi, G.; Steffan-Dewenter, I.; Vardakas, P.; Verrier, E.; Requier, F. How stressors disrupt honey bee biological traits and overwintering mechanisms. Heliyon 2024, 10, e34390. [Google Scholar] [CrossRef]
- Mockler, B. K.; Kwong, W. K.; Moran, N. A.; Koch, H. Microbiome structure influences infection by the parasite Crithidia bombi in bumble bees. Appl. Environ. Microbiol. 2018, 84, e02335-17. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Olofsson, T. C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Saelao, P.; Borba, R. S.; Ricigliano, V.; Spivak, M.; Simone-Finstrom, M. Honeybee microbiome is stabilized in the presence of propolis. Biol. Lett. 2020, 16, 20200003. [Google Scholar] [CrossRef]
- Yun, J.-H.; Jung, M.-J.; Kim, P. S.; Bae, J.-W. Social status shapes the bacterial and fungal gut communities of the honey bee. Sci. Rep. 2018, 8, 2019. [Google Scholar] [CrossRef]
- Natsopoulou, M. E.; McMahon, D. P.; Doublet, V.; Frey, E.; Rosenkranz, P.; Paxton, R. J. The virulent, emerging genotype B of Deformed Wing Virus is closely linked to the overwinter loss of honeybee workers. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Neumann, P.; Elzen, P. J. The biology of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae): gaps in our knowledge of an invasive species. Apidologie 2004, 35, 229–247. [Google Scholar] [CrossRef]
- Owens, C. D. The thermology of wintering honey bee colonies. Technical Bulletins. 1971. Available online: https://ideas.repec.org/p/ags/uerstb/171857.html.
- Palacios, S.; Añon, G.; Arredondo, D.; Alarcón, M.; Zunino, P.; Campá, J.; Antúnez, K. Long-term monitoring of Paenibacillus larvae, causative agent of American Foulbrood, in Uruguay. J. Invertebr. Pathol. 2024, 207, 108186. [Google Scholar] [CrossRef]
- Papp, M.; Békési, L.; Farkas, R.; Makrai, L.; Judge, M. F.; Maróti, G.; Tőzsér, D.; Solymosi, N. Natural diversity of the honey bee (Apis mellifera) gut bacteriome in various climatic and seasonal states. PLoS ONE 2022, 17, e0273844. [Google Scholar] [CrossRef]
- Potts, S. G.; Biesmeijer, J. C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W. E. Global pollinator declines: trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Powell, J. E.; Carver, Z.; Leonard, S. P.; Moran, N. A. Field-realistic tylosin exposure impacts honey bee microbiota and pathogen susceptibility, which is ameliorated by native gut probiotics. Microbiol. Spectr. 2021, 9, e00103-21. [Google Scholar] [CrossRef] [PubMed]
- Powell, J. E.; Lau, P.; Rangel, J.; Arnott, R.; De Jong, T.; Moran, N. A. The microbiome and gene expression of honey bee workers are affected by a diet containing pollen substitutes. PLoS ONE 2023, 18, e0286070. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Wang, H.; Liu, Z.; Wang, Y.; Zhang, W.; Xu, B. Changes in cold tolerance during the overwintering period in Apis mellifera ligustica. J. Apic. Res. 2019, 58, 702–709. [Google Scholar] [CrossRef]
- Rajagopalan, K.; DeGrandi-Hoffman, G.; Pruett, M.; Jones, V. P.; Corby-Harris, V.; Pireaud, J.; Curry, R.; Hopkins, B.; Northfield, T. D. Warmer autumns and winters could reduce honey bee overwintering survival with potential risks for pollination services. Sci. Rep. 2024, 14, 1–12. [Google Scholar] [CrossRef]
- Raymann, K.; Coon, K. L.; Shaffer, Z.; Salisbury, S.; Moran, N. A. Pathogenicity of Serratia marcescens strains in honey bees. mBio 2018, 9, e01649-18. [Google Scholar] [CrossRef]
- Raymann, K.; Shaffer, Z.; Moran, N. A. Antibiotic exposure perturbs the gut microbiota and elevates mortality in honeybees. PLoS Biol. 2017, 15, e2001861. [Google Scholar] [CrossRef]
- Requier, F.; Leyton, M. S.; Morales, C. L.; Garibaldi, L. A.; Giacobino, A.; Porrini, M. P.; Rosso-Londoño, J. M.; Velarde, R. A.; Aignasse, A.; Aldea-Sánchez, P.; Allasino, M. L.; Arredondo, D.; Audisio, C.; Cagnolo, N. B.; Basualdo, M.; Branchiccela, B.; Calderón, R. A.; Castelli, L.; Castilhos, D.; Antúnez, K. First large-scale study reveals important losses of managed honey bee and stingless bee colonies in Latin America. Sci. Rep. 2024, 14, 1–18. [Google Scholar] [CrossRef]
- Ricigliano, V. A.; Anderson, K. E. Probing the honey bee diet-microbiota-host axis using pollen restriction and organic acid feeding. Insects 2020, 11, 291. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Rothman, J. A.; Carroll, M. J.; Meikle, W. G.; Anderson, K. E.; McFrederick, Q. S. Longitudinal effects of supplemental forage on the honey bee (Apis mellifera) microbiota and inter- and intra-colony variability. Microb. Ecol. 2018, 76, 814–824. [Google Scholar] [CrossRef]
- Martinson, V. G.; Danforth, B.; Minckley, R. L.; Rueppell, O.; Tingek, S.; Moran, N. A. A simple and distinctive microbiota associated with honey bees and bumble bees. Mol. Ecol. 2011, 20, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Shah, S. How climate change impacts winter weather. TIME. 13 February 2025. Available online: https://time.com/7222241/how-are-our-winters-impacted-by-climate-change.
- Simone-Finstrom, M.; Li-Byarlay, H.; Huang, M. H.; Strand, M. K.; Rueppell, O.; Tarpy, D. R. Migratory management and environmental conditions affect lifespan and oxidative stress in honey bees. Sci. Rep. 2016, 6, 32023. [Google Scholar] [CrossRef]
- Song, Y.; Fang, H.; Li, Y.; Zhang, X. Cold stress reshapes the honey bee gut microbiota. Front. Microbiol. 2022, 13, 832762. [Google Scholar]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Steele, M. I.; Motta, E. V. S.; Gattu, T.; Martinez, D.; Moran, N. A. The gut microbiota protects bees from invasion by a bacterial pathogen. Microbiol. Spectr. 2021, 9, e00394-21. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, N.; vanEngelsdorp, D.; Saegerman, C. Prioritizing changes in management practices associated with reduced winter honey bee colony losses for US beekeepers. Sci. Total Environ. 2020, 753, 141629. [Google Scholar] [CrossRef]
- Steinmann, N.; Corona, M.; Neumann, P.; Dainat, B. Overwintering is associated with reduced expression of immune genes and higher susceptibility to virus infection in honey bees. PLoS ONE 2015, 10, e0129956. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R. J.; Flaberg, E.; Szekely, L.; Olofsson, T. C. Symbionts as major modulators of insect health: lactic acid bacteria and honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Wang, H.; Liu, C.; Liu, Z.; Wang, Y.; Ma, L.; Xu, B. The different dietary sugars modulate the composition of the gut microbiota in honeybee during overwintering. BMC Microbiol. 2020, 20, 61. [Google Scholar] [CrossRef]
- Williams, J. R.; Swale, D. R.; Anderson, T. D. Comparative effects of technical-grade and formulated chlorantraniliprole on the survivorship and locomotor activity of the honey bee, Apis mellifera (L.). Pest Manag. Sci. 2020, 76, 2582–2588. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, A.; Huang, S. K.; Evans, J. D.; Cook, S. C.; Palmer-Young, E.; Chen, Y. P. Mediating a host cell signaling pathway linked to overwinter mortality offers a promising therapeutic approach for improving bee health. J. Adv. Res. 2023, 53, 99–114. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C. M. A. P.; Harris, H. M. B.; Mattarelli, P. A taxonomic note on the genus Lactobacillus: description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Nishida, A.; Kwong, W. K.; Koch, H.; Engel, P.; Steele, M. I.; Moran, N. A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. mBio 2016, 7, e01326-16. [Google Scholar] [CrossRef]
- Zheng, H.; Nishida, A.; Kwong, W. K.; Koch, H.; Engel, P.; Steele, M. I.; Moran, N. A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. mBio 2016, 7, e01326-16. [Google Scholar] [CrossRef]
- Zheng, H.; Powell, J. E.; Steele, M. I.; Dietrich, C.; Moran, N. A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. U.S.A 2017, 114, 4775–4780. [Google Scholar] [CrossRef]
- Zheng, H.; Steele, M. I.; Leonard, S. P.; Motta, E. V. S.; Moran, N. A. Honey bees as models for gut microbiota research. Lab Anim. 2018, 47, 317–325. [Google Scholar] [CrossRef] [PubMed]
Table 1.
Effects of various pathogens on honey bee physiological parameters and gut microbiome composition.
Table 1.
Effects of various pathogens on honey bee physiological parameters and gut microbiome composition.
| Factor/ Stress | Microbial Changes Observed | Core Genera Affected | Mechanism/Effect | Reference(s) |
|---|---|---|---|---|
| Nosema ceranae infection | Decrease in microbial diversity | ↓ Lactobacillus ↑ Snodgrassella |
Gut dysbiosis, weakened immunity | [46,57] |
| Hafnia alvei infection | Strain-specific protection against pathogen | Gilliamella apicola W8136, Lactobacillus apis W8172 (effective strains) | Certain strains help clear H. alvei infection; others ineffective | [56] |
| Cold Stress (Overwintering) | Reduced diversity, altered composition | ↑ Bartonella ↓ Commensalibacter |
Lower metabolism, limited foraging | [50,93] |
| Stable detection of Gilliamella, Bartonella, Snodgrassella, Lactobacillus, Frischella, Commensalibacter, and Bifidobacterium. | ↑Bartonella, and Bifidobacterium and then decreased in winter honey bees | Host metabolism and may affect the storage of energy | [10] | |
| Hive condition, i.e., winter survival or failure. | Lower microbial abundance and altered composition in failed hives | ↓ Commensalibacter ↓ Snodgrassella in non-surviving hives |
Microbial abundance and beta diversity strongly linked to winter survival | [15] |
| Environmental habitat (Anthropization) | ↑ Pantoea and Arsenophonus in agricultural ↑ Lactobacillus Commensalibacter and Snodgrasella in semi-natural |
Enviromental -linked microbiota shifts | [41] | |
| Overwintering climate (warm vs. cold) and worker age affect | In warm climates, worker bees exhibited reduced longevity and increased fungal abundance in the hindgut, along with shifts in bacterial communities. Conversely, Cold overwintering maintained a stable microbiota and longer worker lifespan. | ↑ fungi ↑Gilliamella spp |
Overwintering conditions have a significant influence on the gut microbiome and the health of honey bee colonies. Cold indoor environments may support microbiota stability and enhance colony survival during winter | [64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



