1. Introduction
Porphyromonas gingivalis is a key intracellular pathogen in periodontitis, capable of invading human gingival epithelial cells and persisting within tissues, contributing to the chronic inflammatory characteristic of the disease [
1,
2,
3,
4]. Intracellular persistence of
P. gingivalis is facilitated by its ability to evade host defenses and modulate local host immune responses [
1,
5,
6,
7,
8,
9], contributing to
P. gingivalis’ association with oral and several systemic diseases [
4]. The increasing rate of antibiotic resistance and the fact that conventional antibiotics may not always be successful against intracellular pathogens highlight the need for alternative strategies that promote pathogen elimination [
10]. Natural products are often considered safer for human consumption than synthetic drugs and, therefore, represent ideal candidates for therapeutic treatments [
11].
Frankincense oil can be extracted from
Boswellia serrata, a medicinal plant widely recognized for its anti-inflammatory properties [
11]. The
B. serrata extract contains bioactive boswellic acids with diverse biological activities, including antimicrobial effects against oral pathogens [
11,
12,
13]. We have previously reported that the
B. serrata extract eliminates
P. gingivalis planktonic cell growth and reduces
P. gingivalis biofilm formation [
13]. Other groups have shown that different compounds from
B. serrata inhibit other oral pathogens, such as
Streptococcus mutans and
Enterococcus faecalis [
12]. Boswellic acids and several of their acetyl compositions are well-documented to have antibacterial activity against Gram-positive bacteria and can synergize with antibiotics, making them potential therapeutic agents [
14].
Reactive oxygen species (ROS)-mediated killing is a canonical host defense mechanism employed by immune cells and epithelial cells, and leveraged in various antimicrobial strategies [
15,
16,
17,
18]. ROS production plays a central role in epithelial antimicrobial activity, directly damaging intracellular pathogens and promoting their clearance [
19]. Boswellic acid compounds have been shown to induce ROS production in host cells [
20,
21,
22,
23]. Mechanistic studies demonstrate that 11-keto-boswellic acids stimulate ROS synthesis in polymorphonuclear leukocytes via NADPH oxidase and MAPK signaling in a Ca²⁺-dependent manner [
24], providing direct evidence that boswellic acids can drive host oxidative responses. However,
B. serrata and related boswellic acids can also exhibit antioxidant effects in non-immune cells, indicating that responses may vary depending on the study model [
13].
Our recent studies demonstrate that
B. serrata extracts inhibit the growth of
P. gingivalis and biofilm formation, as well as reduce intracellular infection in human gingival epithelial cells, without exhibiting cytotoxic effects [
13]. These findings suggest that bioactive
B. serrata compounds may act by modulating host responses to limit intracellular survival of
P. gingivalis, although the precise mechanisms underlying this effect remain to be elucidated.
We hypothesize that bioactive compounds from B. serrata restrict P. gingivalis infection in human gingival epithelial cells by enhancing host antimicrobial responses. Here, we demonstrated that treatment with B. serrata compounds reduced intracellular bacterial load and increased ROS production, while ROS scavenging reversed the antimicrobial effect. Understanding the cellular mechanism and identifying the bioactive compounds from B. serrata involved in this effect may reveal novel therapeutic targets and support the development of strategies for managing periodontal infections.
3. Discussion
Frankincense has been used for centuries in traditional medicine across regions of Africa, India, and the Middle East, where
Boswellia species are found [
25]. Historically valued for their anti-inflammatory properties,
Boswellia extracts have more recently gained attention in biomedical research, with accumulating evidence supporting their therapeutic potential across a range of inflammatory and neoplastic conditions [
11]. Clinical and preclinical studies have reported beneficial effects of frankincense preparations in various diseases, including arthritis, inflammatory bowel disease, asthma, and brain tumors [
11,
26,
27,
28]. In addition to their anti-inflammatory activity, frankincense-derived compounds have been associated with anti-thrombotic, anti-ulcer, and anti-diabetic effects, largely attributed to bioactive constituents present in whole extracts [
11]. Among these constituents, boswellic acids are present in the four major bioactive fractions of pentacyclic triterpenic acids, β-boswellic acid (BA), acetyl-β-boswellic acid (ABA), 11-keto-β-boswellic acid (KBA), and 3-O-acetyl-11-keto-β-boswellic acid (AKBA), being the most extensively studied in therapeutic contexts [
25,
29].
Beyond immunomodulation,
B. serrata extracts have demonstrated antimicrobial activity against fungi as well as Gram-positive and Gram-negative bacteria, including
Staphylococcus epidermidis, Enterococcus faecalis, and Escherichia coli [
14]. Regarding oral pathogens, boswellic extracts have been reported to inhibit bacterial growth, with efficacy dependent on compound composition and concentration [
12]. Our group has previously shown that
B. serrata extract reduces the survival of
P. gingivalis under planktonic conditions and inhibits
P. gingivalis biofilm formation [
13]. Antimicrobial activities have been attributed, at least in part, to terpenoid-mediated disruption of bacterial membranes, inhibition of protein synthesis, and impairment of ATP production [
14,
30].
Despite evidence supporting growth inhibition and direct antimicrobial effects, prior studies have not elucidated mechanisms by which
Boswellia extracts influence host–pathogen interactions during infection. In this study, we demonstrate for the first time that whole
B. serrata extract and its bioactive compound AKBA modulate both bacterial internalization and intracellular persistence of
P. gingivalis in human gingival epithelial cells. Notably, treatment with
B. serrata enhanced bacterial uptake during early times of infection, while subsequently promoting efficient intracellular clearance at later times of infection through a ROS-dependent mechanism (
Figure 5).
The increase in bacterial internalization at early times of infection may reflect the immunomodulatory properties of boswellic acids. Similarly to other phytochemicals, such as curcumin and catechins, boswellic acids may prime epithelial cells by enhancing uptake or activation pathways that precede antimicrobial responses [
31,
32]. Previous studies have suggested that boswellic acids regulate intracellular vesicle trafficking, glycosylation, and molecular transport, thereby influencing the function of the Golgi apparatus and autophagy-related pathways [
33,
34]. Our data provide the first evidence that
B. serrata extracts directly affect the internalization of both bacterial pathogens and inert particulate latex beads.
Cellular mechanisms that restrict intracellular pathogen infections are highly context-dependent and vary according to the cell type and microbial features. In study models using professional immune cells, pathogen clearance involves various processes, including phagocytosis, inflammasome activation, ROS and nitric oxide production, vesicular fusion, endosomal degradation, autophagy, and proteasomal pathways [
35]. As part of the innate immune response, gingival epithelial cells retain the capacity to deploy antimicrobial mechanisms in response to infection [
36,
37].
Previous studies have shown that 11-keto-boswellic acids can stimulate ROS generation in neutrophils via NADPH oxidase and MAPK signaling, indicating their ability to activate host oxidative defenses [
24]. Conversely, in some non-immune cell types, AKBA has been reported to attenuate oxidative stress through Nrf2 activation, underscoring the context-dependent effects of boswellic acids [
38,
39,
40,
41]. In our model, oxidative stress emerged as a critical mediator of intracellular bacterial clearance, as NAC completely abrogated the antimicrobial effects of
B. serrata extracts. Importantly, we demonstrated that AKBA was a key compound in the elimination of intracellular
P. gingivalis, which also involved ROS-dependent mechanisms, as evidenced by the blocking of its bacterial-killing effects by NAC. These findings are consistent with prior studies demonstrating the central role of ROS in reducing intracellular pathogens and the capacity of natural products to potentiate host antimicrobial responses [
14,
18].
Taken together, our data support a dual mechanism of action for the whole B. serrata extract and AKBA: an early step characterized by enhanced bacterial uptake in the initial times of infection, likely reflecting epithelial activation, followed by ROS-mediated intracellular killing at later times of infection. This biphasic response suggests that Boswellia compounds reinforce epithelial defense mechanisms by coupling host metabolic and oxidative pathways, leading to pathogen elimination. Importantly, these findings provide new insights into how natural compounds can simultaneously modulate host immunity while limiting microbial persistence in oral epithelial infections.
From a clinical perspective, these results support the potential use of B. serrata–derived bioactive compounds as potential therapeutics in periodontal disease, where both antimicrobial activity and host modulation are desirable. Future studies should investigate additional bioactive components within frankincense extracts, explore ROS-independent mechanisms, and assess interactions with immune cells to further define the therapeutic framework for leveraging natural compounds in host-directed antimicrobial strategies.
4. Materials and Methods
4.1. Boswellia Serrata Extract and Compounds
B. serrata extract USP reference standard (cat #1076250, Sigma Aldrich-ST. Louis, MO, USA) was resuspended in dimethyl sulfoxide (DMSO; Sigma Aldrich-ST. Louis, MO, USA) and used in our experiments, as we previously described [
13]. Purified compounds from
B. serrata were 3-O-Acteyl-11-keto-β-boswellic acid (AKBA - cat# PHL89169, Sigma Aldrich-St. Louis, MO, USA) and β-Boswellic acid (BA - cat # 80342, Sigma Aldrich-St. Louis, MO, USA). The compounds were resuspended in DMSO at 128mg/mL for
B. Serrata, 10 mg/mL for AKBA, and 50 mg/mL for BA. The compounds were stored at 4 °C until use.
4.2. Bacterial Strain and Cell Culture
P. gingivalis (ATCC
® 33277) was from American Type Culture Collection (ATTC—Manassas, Virginia, Washington, DC, USA) and grown as previously described [
7,
13]. Briefly,
P. gingivalis was grown anaerobically at 37 °C for approximately 7 days in Brucella agar plates (Anaerobe systems, cat# AS-141—Morgan Hill, CA, USA). Isolated and pure colonies were collected from agar plates and inoculated into supplemented BHI broth at 37 °C under anaerobic conditions for approximately 48 h prior to the experiments. Freshly grown bacteria were used in experiments after being collected in log phase and quantified by optical density at 600 nm using the SpectraMax iD3 microplate reader (Molecular Devices—Ramsey, MN, USA).
Immortalized human gingival keratinocytes (HPV-16GM), referred to as human gingival epithelial cells, were obtained from Applied Biological Materials (ABM, cat#T0717 - Richmond, CA, USA) and maintained as we previously described [
7,
13]. Briefly, human gingival epithelial cells were grown and maintained in keratinocyte-serum-free medium (KSFM) supplemented with 30 µg/mL of bovine pituitary extract, 0.2 ng/mL of human recombinant epidermal growth factor, 100 U/mL of penicillin, and 100 µg/mL of streptomycin (Gibco -Gaithersburg, MO, USA). The cells were grown in a humidified incubator at 37 °C, 5% CO2, and quantified using trypan blue (Sigma-Aldrich - St. Louis, MO, USA) exclusion before each experiment.
4.5. Quantification of P. gingivalis Infection
The antibiotic protection assays were performed to quantify intracellular
P. gingivalis in human gingival epithelial cells, as we previously described [
7,
13]. Briefly, 3 x 10
5 human gingival epithelial cells were seeded in 6-well plates (Costar, Corning-Glendale, AZ, USA) and incubated overnight in supplemented KSFM media without antibiotics. The cells were infected with
P. gingivalis at an MOI of 100 in the presence or absence of
B. serrata extracts or its compounds AKBA and BA at concentrations of 16 µg/mL, 4 µg/mL, and 32 µg/mL, respectively, in OptiMEM (Gibco, Gaithersburg, MO, USA) for 2 h, at 37 °C, 5% CO
2. Then, the cells were washed three times with sterile prewarmed PBS. To remove extracellular bacteria, the cells were treated with metronidazole (200 µg/mL) and gentamicin (300 µg/mL) in OptiMEM for an additional 1 h at 37 °C and 5% CO2. Then, the cells were washed three times with prewarmed PBS and maintained in OptiMEM for up to 5 h, or for a total of 9 h. After 5 h or 9 h of infection, sterile distilled water was added to the wells and incubated at room temperature for 20 min to lyse the cells. Each cell lysate was plated onto a Brucella Blood Agar Plate (Anaerobe Systems, Morgan Hill, CA, USA) and incubated under anaerobic conditions for 10 days at 37 °C. The colony-forming units (CFU) were then counted and quantified.
For the detection of P. gingivalis infection using immunofluorescence microscopy, 1 × 105 human gingival epithelial cells were seeded onto a 13 mm coverslip in a 24-well plate and incubated overnight at 37 °C and 5% CO2. The experiment followed the description above for the “colony-forming unit assays”, in which the cells were infected for 2 h in the presence or absence of B. serrata extract or its compounds AKBA and BA at concentrations of 16 µg/mL, 4 µg/mL, and 32 µg/mL, respectively. Then, the cells were treated with antibiotics for an additional 1 h and incubated in OptiMEM at 37 °C, 5% CO2 for up to 5 h, 7 h, or 9 h. In some experiments, we pretreated the cells with NAC (10 µM) 1 h before infection. At the end of the experiment, infected cells on the coverslips were washed three times with sterile PBS, then fixed with 4% formaldehyde. The cells were washed three times with PBS, permeabilized, and blocked with a solution of 0.2% Triton X-100 (Sigma Aldrich-St. Louis, MO, USA) in 5% Goat Serum/PBS (Sigma Aldrich-St. Louis, MO, USA) for 40 min, at RT. The cells were incubated with a primary rabbit polyclonal antibody anti-P. gingivalis (cat# ANT0085, Diatheva – Cartoceto, Italy) at a concentration of 1:50 prepared in 0.05% Triton X-100 in 5% Goat Serum/PBS at 4 °C, for 3 h. After three washes with PBS, secondary goat anti-rabbit IgG (cat# A11012, Invitrogen—Carlsbad, CA, USA) at a concentration of 1:200 was prepared in 0.05% Triton X-100 in 5% Goat Serum/PBS, and was added into the wells for incubation for 2 h, protected from light, at RT. The coverslips were washed three times and mounted onto a slide using Vectashield Hardset Antifade Mounting Medium with DAPI (Cat # H-1500, Vector Laboratories, Burlingame, CA, USA). Images were then acquired using a Nikon Eclipse 50i fluorescence microscope with an Infinity 3 camera (Nikon Instruments, Melville, NY, USA) and the Lumenera Infinity Analyze 6.3 software (Teledyne Lumenera, Ottawa, ON, Canada), or a THUNDER imager 3D assay microscope (Leica). The fluorescence intensity was quantified using the software ImageJ and expressed as arbitrary units.
4.6. Latex Bead Uptake Assay
For the detection of inert particle internalization using fluorescence microscopy, 1 × 105 human gingival epithelial cells were seeded overnight onto a 13 mm coverslip in a 24-well plate at 37 °C and 5% CO2. The cells were incubated with 0.5 μm or 1 μm green fluorescent latex beads (Cat # F-7895, Thermo Fisher Scientific, MA-USA) for 2 h at a cell/bead ratio of 1:100 in the presence or absence of 16 µg/mL of B. serrata extracts. Then, the cells were washed three times with sterile PBS and incubated in OptiMEM for an additional 3 h at 37 °C and 5% CO2, for a total of 5 h. The cells were fixed with 4% formaldehyde solution for 30 min. The coverslips were washed three times and mounted onto a slide using Vectashield Hardset Antifade Mounting Medium with DAPI (Cat # H-1500, Vector Laboratories, Burlingame, CA, USA) and Vectashield Hardset Antifade Mounting Medium with phalloidin (Cat # H-1600, Vector Laboratories, Burlingame, CA, USA). Images were then acquired using a Nikon Eclipse 50i fluorescence microscope with an Infinity 3 camera (Nikon Instruments, Melville, NY, USA) and the Lumenera Infinity Analyze 6.3 software (Teledyne Lumenera, Ottawa, ON, Canada), or a THUNDER imager 3D assay microscope (Leica). The percentage of bead-containing cells was quantified using ImageJ software and expressed as arbitrary units.
4.7. Measurement of ROS Production
For detection of ROS production using fluorescence microscopy, 1 × 105 human gingival epithelial cells were seeded overnight onto a 13 mm coverslip in a 24-well plate at 37 °C and 5% CO2. The experiment followed the description above for the “colony-forming units assay”, in which the cells were infected for 2 h in the presence or absence of B. serrata extracts, treated with antibiotics for an additional 1 h, and then incubated in OptiMEM for an additional 4 h, at 37 °C, 5% CO2 for a total of 7 h of infection. CellROX green fluorogenic probe was added to the wells 1 h before the end of the experiment. Hydrogen peroxide was used as a positive control. At the end of the experiment, the cells on the coverslips were washed three times with sterile PBS, then fixed with 4% formaldehyde for 30 min. The coverslips were washed three times and mounted onto a slide using Vectashield Hardset Antifade Mounting Medium with DAPI (Cat # H-1500, Vector Laboratories, Burlingame, CA, USA). Images were then acquired using a Nikon Eclipse 50i fluorescence microscope with an Infinity 3 camera (Nikon Instruments, Melville, NY, USA) and the Lumenera Infinity Analyze 6.3 software (Teledyne Lumenera, Ottawa, ON, Canada), or a THUNDER imager 3D assay microscope (Leica). The fluorescence intensity was quantified using ImageJ software and expressed as arbitrary units.
4.8. Statistics
Statistical analysis was performed using Prism GraphPad (GraphPad Software, version 9.5.1). The results are presented as standard deviation (SD) and were analyzed using an unpaired t-test with Welch’s correction for up to two conditions and one-way ANOVA followed by Tukey’s multiple comparison test for more than two analyzed conditions. Differences resulting in p < 0.05 were considered significant.
Author Contributions
Conceptualization, D.M.O, C.L.C.A.-d.-S. and A.C.A.M.S.; methodology, D.M.O, C.L.C.A.-d.-S. and A.C.A.M.S.; formal analysis, P.H.C., C.L.C.A.-d.-S., and A.C.A.M.S.; investigation, D.V., P.H.C., A.S., L.H., H.M.D., and S.D.; resources, C.L.C.A.-d.-S. and A.C.A.M.S.; writing—original draft preparation, D.V., P.H.C., C.L.C.A.-d.-S., and A.C.A.M.S.; writing—review and editing, C.L.C.A.-d.-S. and A.M.S.; supervision, A.C.A.M.S.; project administration, A.C.A.M.S.; funding acquisition, C.L.C.A.-d.-S. and A.C.A.M.S. All authors have read and agreed to the published version of the manuscript.
Funding:. This research was funded by intramural funds from the University of the Pacific, Arthur A. Dugoni, School of Dentistry, to CLCAS and A.C.A.M.S. (D30060 and Pilot grant C10219).
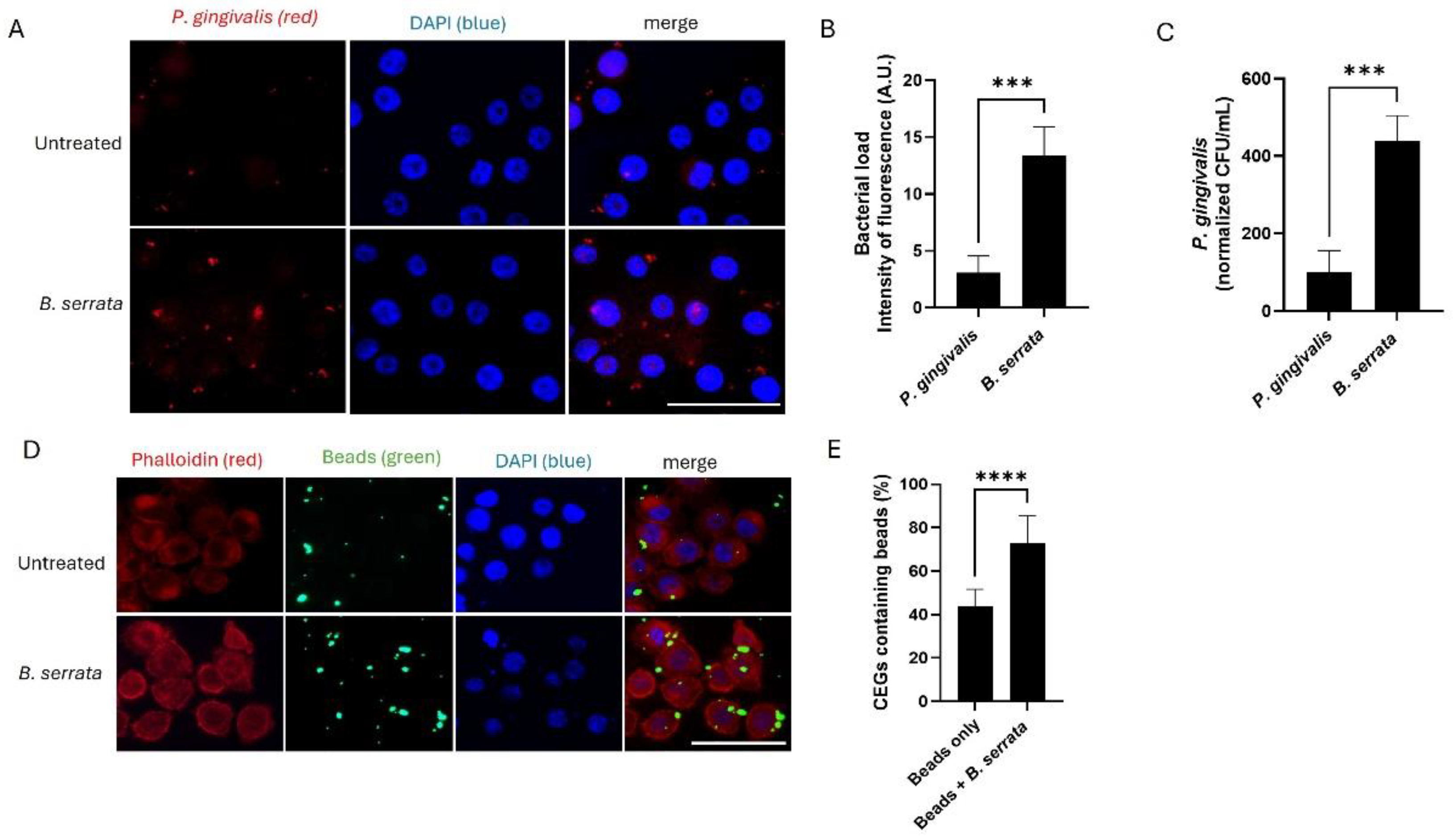
Figure 1.
B. serrata enhances bacterial internalization at early times of infection in human gingival epithelial cells. (A) Immunofluorescence micrograph showing intracellular P. gingivalis (red) infection in human gingival epithelial cells incubated with or without B. serrata extracts, at 5 h post-infection (scale bar = 50 µm). (B) Quantification of intracellular fluorescence intensity in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 5 h post-infection (** = p < 0.001, n = 3). (C) CFU quantification of viable P. gingivalis incubated with or without B. serrata extracts, at 5 h post-infection (*** = p < 0.001, n = 4). (D) Representative fluorescence micrograph of human gingival epithelial cells incubated with green fluorescent beads in the presence or absence of B. serrata extract at 5 h post-infection. (E) Percentage of bead-containing gingival epithelial cells at 5 h post-infection (**** = p < 0.0001, n = 4).
Figure 1.
B. serrata enhances bacterial internalization at early times of infection in human gingival epithelial cells. (A) Immunofluorescence micrograph showing intracellular P. gingivalis (red) infection in human gingival epithelial cells incubated with or without B. serrata extracts, at 5 h post-infection (scale bar = 50 µm). (B) Quantification of intracellular fluorescence intensity in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 5 h post-infection (** = p < 0.001, n = 3). (C) CFU quantification of viable P. gingivalis incubated with or without B. serrata extracts, at 5 h post-infection (*** = p < 0.001, n = 4). (D) Representative fluorescence micrograph of human gingival epithelial cells incubated with green fluorescent beads in the presence or absence of B. serrata extract at 5 h post-infection. (E) Percentage of bead-containing gingival epithelial cells at 5 h post-infection (**** = p < 0.0001, n = 4).
Figure 2.
B. serrata reduces the intracellular load of P. gingivalis in human gingival epithelial cells. (A) Immunofluorescence micrograph showing intracellular P. gingivalis at 7 h post-infection in untreated control and B. serrata–treated gingival epithelial cells (scale bar = 50 µm). (B) Quantification of intracellular intensity of fluorescence in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 7 h post-infection (* = p < 0.05, n = 3). (C) Immunofluorescence micrograph showing intracellular P. gingivalis at 9 h post-infection in untreated control and B. serrata–treated gingival epithelial cells (scale bar = 50 µm). (D) Quantification of intracellular intensity of fluorescence in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 9 h post-infection (*** = p < 0.001, n = 3). (E) CFU quantification of viable intracellular P. gingivalis in the presence or absence of B. serrata treatment at 9 h post-infection (**** = p <0.0001, n = 4).
Figure 2.
B. serrata reduces the intracellular load of P. gingivalis in human gingival epithelial cells. (A) Immunofluorescence micrograph showing intracellular P. gingivalis at 7 h post-infection in untreated control and B. serrata–treated gingival epithelial cells (scale bar = 50 µm). (B) Quantification of intracellular intensity of fluorescence in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 7 h post-infection (* = p < 0.05, n = 3). (C) Immunofluorescence micrograph showing intracellular P. gingivalis at 9 h post-infection in untreated control and B. serrata–treated gingival epithelial cells (scale bar = 50 µm). (D) Quantification of intracellular intensity of fluorescence in P. gingivalis-infected cells incubated with or without B. serrata extracts, at 9 h post-infection (*** = p < 0.001, n = 3). (E) CFU quantification of viable intracellular P. gingivalis in the presence or absence of B. serrata treatment at 9 h post-infection (**** = p <0.0001, n = 4).
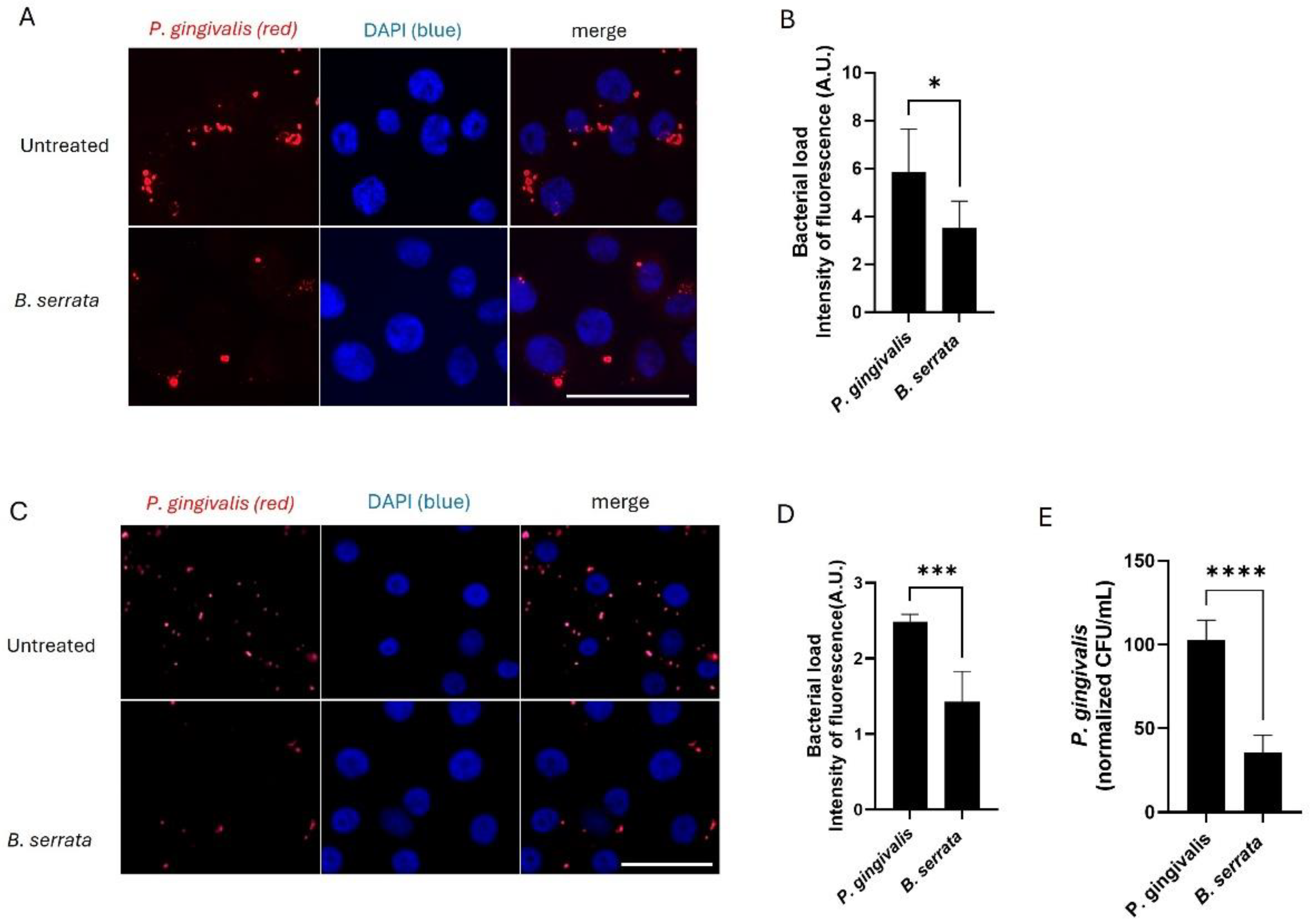
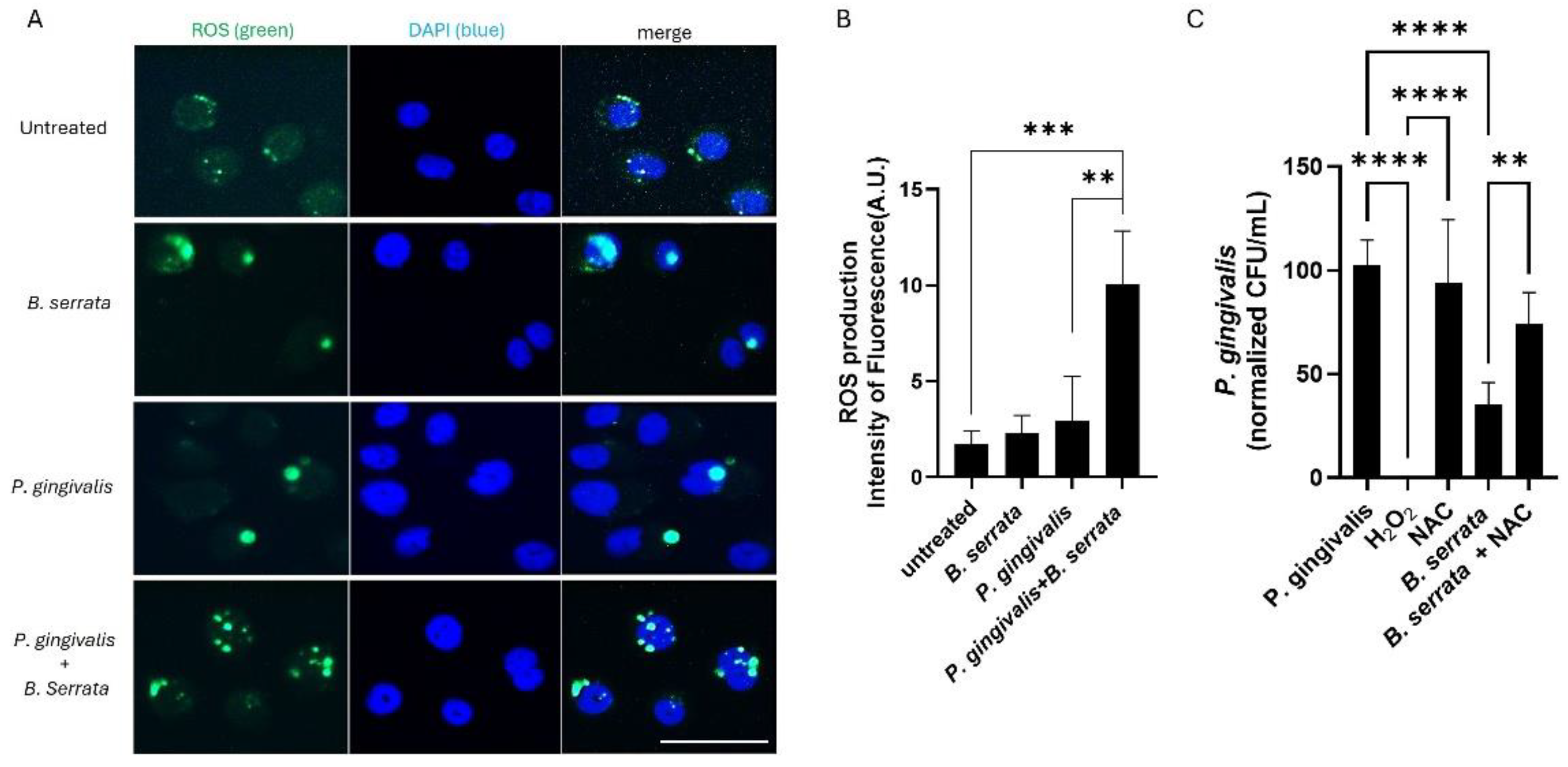
Figure 3.
B. serrata induces ROS production and restricts the intracellular survival of P. gingivalis in human gingival epithelial cells. (A) Representative fluorescence micrograph of ROS generation in human gingival epithelial cells infected or not with P. gingivalis and treated with or without B. serrata extracts. Detection of ROS using the CellROX fluorogenic probe was performed 7 h post-infection (scale bar = 50 µm). (B) Quantification of the green CellROX fluorogenic probe fluorescence intensity in B. serrata–treated cells compared with untreated controls (** = p < 0.005, *** = p < 0.001, n = 4). (C) CFU quantification of viable intracellular P. gingivalis treated with or without H2O2 or B. serrata, and pretreated or not with NAC, at 9 h post-infection. (** = p < 0.005, **** = p < 0.0001, n = 6).
Figure 3.
B. serrata induces ROS production and restricts the intracellular survival of P. gingivalis in human gingival epithelial cells. (A) Representative fluorescence micrograph of ROS generation in human gingival epithelial cells infected or not with P. gingivalis and treated with or without B. serrata extracts. Detection of ROS using the CellROX fluorogenic probe was performed 7 h post-infection (scale bar = 50 µm). (B) Quantification of the green CellROX fluorogenic probe fluorescence intensity in B. serrata–treated cells compared with untreated controls (** = p < 0.005, *** = p < 0.001, n = 4). (C) CFU quantification of viable intracellular P. gingivalis treated with or without H2O2 or B. serrata, and pretreated or not with NAC, at 9 h post-infection. (** = p < 0.005, **** = p < 0.0001, n = 6).
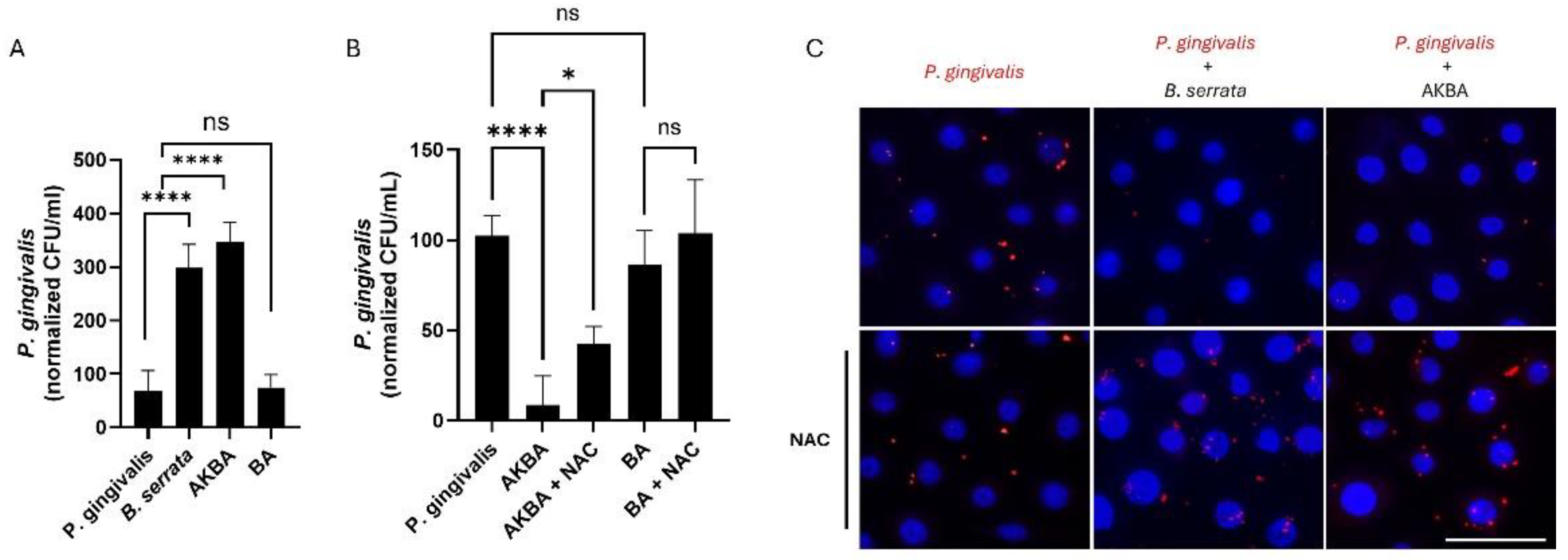
Figure 4.
AKBA promotes the killing of P. gingivalis through ROS production, reproducing the B. serrata phenotype. (A) CFU quantification of viable intracellular P. gingivalis treated with or without B. serrata extract or its bioactive components AKBA or BA at 5 h post-infection (**** = p < 0.0001, n = 6). (B) CFU quantification of viable intracellular P. gingivalis treated with or without AKBA or BA, and pretreated or not with NAC, at 9 h post-infection (* = p < 0.05, **** = p < 0.0001, n = 4). (C) Immunofluorescence micrograph showing intracellular P. gingivalis (red) in human gingival epithelial cells incubated with or without B. serrata extracts or AKBA, and pretreated or not with NAC, after 9 h of infection. (scale bar = 50 µm).
Figure 4.
AKBA promotes the killing of P. gingivalis through ROS production, reproducing the B. serrata phenotype. (A) CFU quantification of viable intracellular P. gingivalis treated with or without B. serrata extract or its bioactive components AKBA or BA at 5 h post-infection (**** = p < 0.0001, n = 6). (B) CFU quantification of viable intracellular P. gingivalis treated with or without AKBA or BA, and pretreated or not with NAC, at 9 h post-infection (* = p < 0.05, **** = p < 0.0001, n = 4). (C) Immunofluorescence micrograph showing intracellular P. gingivalis (red) in human gingival epithelial cells incubated with or without B. serrata extracts or AKBA, and pretreated or not with NAC, after 9 h of infection. (scale bar = 50 µm).
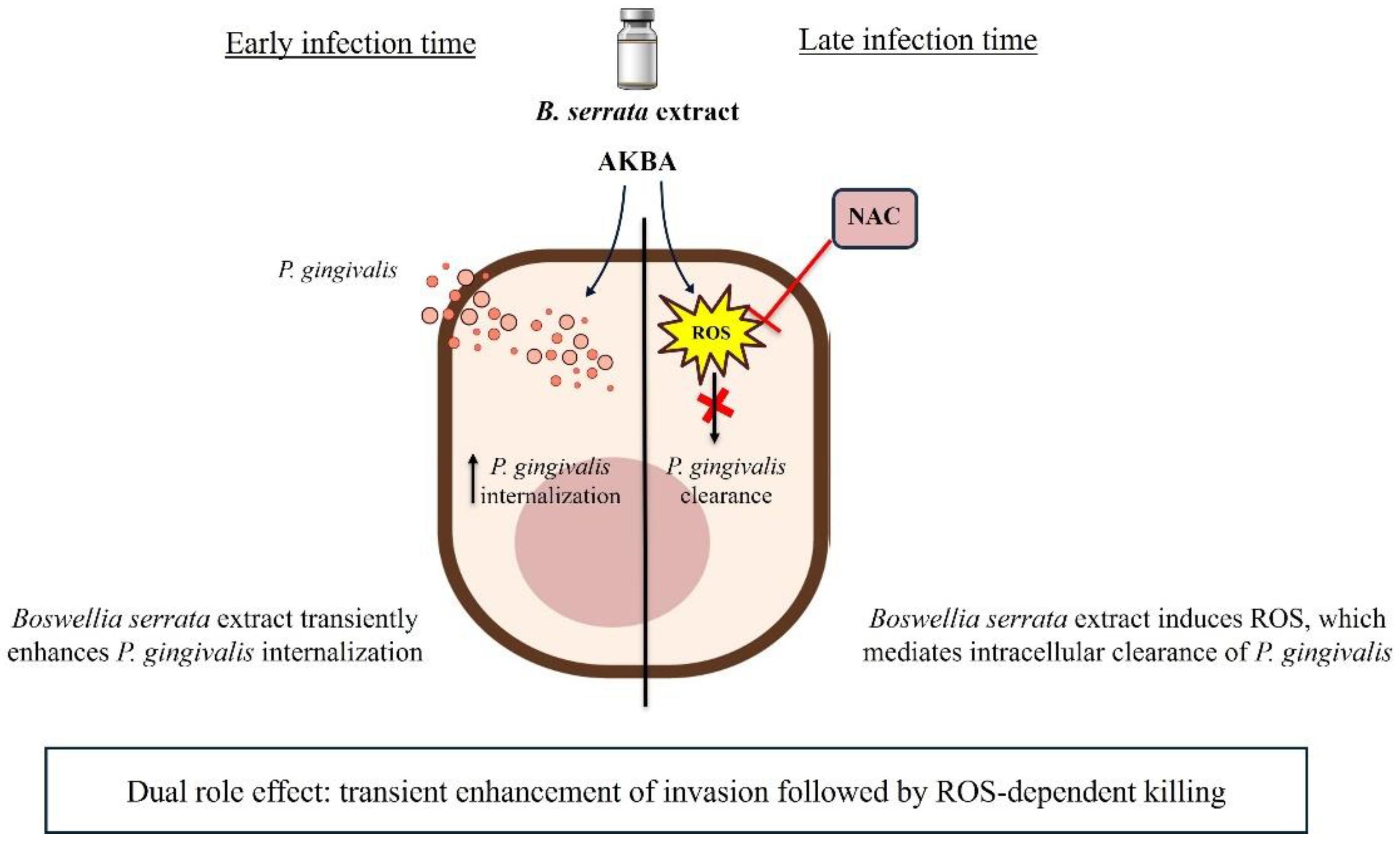
Figure 5.
Figure 5. Proposed scheme of the dual antibacterial effects of B. serrata extract and AKBA. Schematic representation illustrating the dual role by which Boswellia serrata exerts antimicrobial effects against P. gingivalis. At early times of infection, B. serrata and the AKBA bioactive compound enhance bacterial internalization by stimulating endocytic mechanisms and bacterial uptake. At later times of infection, B. serrata and its bioactive compound, AKBA, trigger robust ROS production, leading to intracellular bacterial killing. The combination of rapid bacterial internalization and host-mediated oxidative mechanisms highlights the multifaceted protective role of B. serrata in controlling P. gingivalis infection.
Figure 5.
Figure 5. Proposed scheme of the dual antibacterial effects of B. serrata extract and AKBA. Schematic representation illustrating the dual role by which Boswellia serrata exerts antimicrobial effects against P. gingivalis. At early times of infection, B. serrata and the AKBA bioactive compound enhance bacterial internalization by stimulating endocytic mechanisms and bacterial uptake. At later times of infection, B. serrata and its bioactive compound, AKBA, trigger robust ROS production, leading to intracellular bacterial killing. The combination of rapid bacterial internalization and host-mediated oxidative mechanisms highlights the multifaceted protective role of B. serrata in controlling P. gingivalis infection.