Submitted:
14 January 2026
Posted:
15 January 2026
You are already at the latest version
Abstract
Genetically encoded biosensors represent a cutting-edge class of biosensors due to real-time monitoring and programmability in living cell. However, the development of eukaryotic genetically encoded biosensors for new analytes is constrained by the shortage of signal–receptor pairs. Bacterial biosensors have been transferred to eukaryote to expand the signal detection space, having achieved remarkable success. However, due to the significant differences between eukaryotic and prokaryotic gene expression systems, optimizing bacterial biosensors has proven challenging. Successful cases indicate that developing orthogonal signal–receptor pairs directly from eukaryotic systems may offer a viable solution. Indeed, the potential of filamentous fungi—a highly diverse group of organisms that share conserved as well as specific signaling and metabolic pathways with yeast or mammalian cells—has been largely overlooked in biosensor development. In this review, we systematically examine sensing systems in filamentous fungi and summarize their signal recognition receptors, signal transduction pathways,responsive transcription factors and describe potential mining strategies for sensing elements from filamentous fungi.
Keywords:
genetically-encoded biosensor
; filamentous fungi
; synthetic biology
; transcription factor-based biosensor
; GPCR
; photoreceptor
; sensing element
1. Introduction
Genetically encoded biosensors are a class of biomolecular devices capable of transducing biological recognition events into detectable signals [1,2]. When compared to traditional biosensors like electrochemical biosensors, they display unique advantages. As an intrinsic part of living cells, genetically encoded biosensors enable real-time and noninvasive detection of small molecules or signals within cells, thereby facilitating continuous and dynamic monitoring [3,4]. Moreover, through subcellular localization design, high spatial resolution within cells can be achieved [5]. Genetically encoded biosensors can be stably inherited, ensuring sustainability at low cost once successfully designed and constructed [6]. Generally, a genetically encoded biosensor primarily comprises a signal recognition and transduction module, along with a signal output module [7]. Signal recognition elements are critical for the development of biosensors. In prokaryotes, signal recognition is typically mediated by allosteric transcription factors (TFs) [8], aptamers [9,10] and Two-Component Systems (TCS)[11]. In fungi, signal recognition elements include membrane-bound G-protein coupled receptors (GPCRs) and ion channels, TCS as well as zinc cluster TFs [12,13,14]. Currently, genetically encoded biosensors are majorly constructed based on TFs, as TF-based biosensors offer the versatility to regulate diverse downstream target genes. Indeed, TF-based biosensors serve as a core component in synthetic biology, with broad applications in high-throughput screening and selection, directed evolution, dynamic regulation of metabolic pathways, and the construction of synthetic genetic circuits [1,15,16]. Recently, synthetic design and construction based on filamentous fungal GPCRs and photoreceptors have been reported [17,18] (Figure 1). These strategies are fundamental to application scenarios in biomanufacturing, medical diagnostics, living material and environmental monitoring. With the increasing complexity and refinement of biomanufacturing processes, coupled with the growing demand for point-of-care diagnostics in the biomedical applications, there is an urgent need for more effective process control strategies and an expanded detectable signal space. The rapid development of biosensors represents a powerful tool for advancing the frontiers in these areas.
1. Limitations of Existing Yeast Biosensors for Synthetic Design
The commonly used yeast biosensors can be classified into endogenous induction system and heterologous bacterial-derived biosensors. Each category has inherent limitations and developmental bottlenecks for synthetic design.
The intensive used induction systems in metabolic engineering are dominated by endogenous biosensors in yeast, including the galactose-responsive GAL system, the copper-inducible PCUP1 system, the phosphate-responsive PPHO5 system and the methionine-regulated PMET3 system [20]. The advantages of these systems are evident, including ease of molecular operation and/or high inducibility. However, the portability and application scenarios of these native systems are constrained by complex signal transduction cascades, coupling with host growth requirements and off-target effects (non-orthogonality) [21,22,23] (Figure 2). In past decades, to overcome the limitation of endogenous biosensors, bacteria-derived biosensors were continually transferred to yeast and optimized through directed evolution [24,25]. Engineering prokaryotic regulators is an effective strategy to enrich the toolbox of yeast biosensors. However, progress is constrained by the limited number of identified bacterial biosensors and host compatibility issues such as growth toxicity, lowly inducibility and slow induction speed [26,27,28]. Although the strategy that fusing strong ADs to bacterial TFs may offer a solution, they may exhibit high leakiness, especially when multiple binding sites are present in tandem in the promoter [29]. Except the directed evolution, no rational strategy has been developed for optimization of bacterial transcriptional activators-derived biosensors, as the activation mechanism is different to that of eukaryotes.
In summary, the non-orthogonal endogenous induction systems, the underperformed bacterial-derived biosensors and the shortage of characterized signal-receptor pairs hindered the development of genetically encoded biosensor for synthetic design.
2. Emerging Opportunity: Mining Response Elements from Filamentous Fungi
Fungi serve as important models in eukaryotes for studying environmental perception and signal transduction. As ecological decomposers, filamentous fungi exhibit remarkable adaptability and the ability to sense a wide array of signals, including pheromones, nutrients, gases, light, host organisms, and mechanical stress [12].
Filamentous fungi have evolved to possess a rich repertoire of signal receptors, including GPCRs, Zinc clusters TF, photoreceptors and TCSs. First, GPCRs mediate the detection of nutrients, pheromones, and various types of stress. There are over 15,000 GPCRs have been predicted in Ascomycota, while only three GPCRs- Ste2, Ste3 and Gpr1, have been characterized in yeast [30]. In other words, species in Ascomycota have an average of over 30 GPCR. Second, it has been reported that fungi lack nuclear receptors, while zinc cluster TFs may represent their functional analogs [14]. Filamentous fungi possess a unique capacity of utilizing some nutrients like pentoses and biosynthesizing secondary metabolites when compared to yeast. Like nuclear receptor, increasing evidences showed that fungal zinc cluster TFs are small-molecule sensors [31,32,33]. For example, in ascomycetous fungi, a network of regulators has been reported to control biomass degradation and catabolic pathways for the resulting monomers, which are often the inducers of these regulators activity[31,34]. Moreover, filamentous fungi are renowned for drug discovery due to rich genetic resource of secondary metabolites biosynthesis [35]. Their pathway genes are clustered and found to often include a specialized transcriptional activator that drives high-level expression of key metabolic enzymes [36]. Most of these metabolic TFs belong to the Zn(II)2Cys6 family and their DBDs exhibit remarkable sequence recognition specificity. This specificity is dictated by the following molecular mechanisms: small molecule-induced activation (e.g., proline induced DNA-binding of PrnA[37]); signal-induced homo- or heterodimerization[32]. Moreover, S. cerevisiae natively lacks metabolic pathways and regulatory networks for utilizing certain carbon sources like pentose (e.g., xylose,arabinose and rhamnose) or synthesizing secondary metabolites, which indicates that heterologous reconstruction of these pathway-specific TF-mediated induction systems could be suitable for synthetic design. Third, unlike yeast are blind, filamentous fungi especially Ascomycota species are capable of perceiving a broad spectrum of light. Taken Botrytis cinerea as an example, there are at least eleven potential photoreceptors to perceive near-UV, blue, green, red and far-red light [38]. Filamentous fungi hence provide prolific genetic components for synthetic optogenetic tools. Lastly, filamentous fungi have been reported to possess far more histidine kinases (HKs) than yeasts. Only one HK-Sln1, has been identified in S. cerevisiae, while at least 10 HKs genes are predicted in filamentous fungus like Neurospora crassa, Cochliobolus heterostrophus, Gibberella moniliformis and Botryotinia fuckeliana[39].
Apart from the rich repertoire of signal receptors and potential orthogonality from unique phenotypes, mining biosensors from filamentous fungi has following advantages. First, many Zinc clusters TFs act as transcriptional activators. Beyond larger dynamic range, gene circuits built with activators are more concise and compact than those constructed with repressors. Second, molecular genetics between filamentous fungi and budding yeast are conserved [40,41], including gene expression machinery, environmental response and stress adaptation mechanisms, transporter, signaling transduction and primary metabolic pathways. In other words, when transfer a response system from filamentous fungi to S. cerevisiae, the signal-specific components would be enough to implement functionality, as evidenced by xylose and arabinose sensor XlnR and AraR[27]. Second, ease of molecular manipulation. Based on our experiences, there is no requirement for codon optimization to heterogeneously express a filamentous fungal derived gene in S. cerevisiae. However, this conservation in signaling pathways would be a double-edged sword, as it may also interfere with the endogenous systems of yeast, thereby undermining the orthogonality required for synthetic design. Therefore, more works need to be done in future to quantify this interference.
3. Native Biosensor in Filamentous Fungi
As key decomposers, these fungi possess the ability to detect and degrade complex plant biomass, where they sense breakdown products (monomeric nutrients) and activate intricate signaling cascades to induce downstream gene expression for further catabolism. Moreover, to adapt with fluctuations in pH, temperature, light, moisture, and oxygen availability, filamentous fungi have evolved specialized sensors—including transmembrane receptors, TFs and phytochromes—that detect these environmental cues and initiate adaptive survival strategies. Consequently, filamentous fungi naturally harbor a rich repertoire of small-molecule and environmental-factor biosensors. A thorough understanding of these signaling mechanisms provides the foundation for cross-species transfer and the rational engineering of these native components into synthetic biological devices.
3.1. Small Molecule-Induction Systems
Small molecules are sensed either through cell membrane receptors or after being transported into the cell via transporters, where they may also directly bind to intracellular receptors like zinc cluster TFs[32,42,43]. For the former, this recognition primarily relies on the cAMP-PKA signaling pathway to transmit the signal to specific TFs, thereby completing the signal output. Since receptors for most small molecules remain unidentified, they will not be discussed in detail here. We focus on the cAMP-PKA signaling pathway and the reported TFs that respond to specific small molecules.
3.1.1. Signaling Pathway of Small Molecules
Small molecules sensed by filamentous fungi include carbon sources, amino acids, volatile organic compounds (VOCs), quorum-sensing molecules and pheromone. The specific signal transduction pathways and the resulting biological function vary depending on the type of signal molecules and organisms, though their core upstream components share evolutionary conservation with yeast—most notably the cAMP-PKA pathway (Figure 3)[44]. When extracellular small molecules exceed a concentration threshold, membrane-localized GPCRs detect these ligands, triggering dissociation of the Gα subunit (e.g., Aspergillus GpaB[45]) or Gβγ. The free Gα-GTP then diffuses to adenylate cyclase (ACYA[46]), catalyzing production of the cAMP and subsequent activation of protein kinase A (PKA). On the other hand, pheromone peptides-triggered Gβγ dimer releasing initiates the MAPK cascade. These complex regulatory networks ultimately converge on the phosphorylation of specific effector TFs (Figure 3). Presumably, the conservation of cAMP-PKA signaling could provides a foundation for simplified cross-species biosensor transfer across fungal species.
3.1.2. Small-Molecule Responsive TFs
Many small-molecule responsive TFs have been identified in filamentous fungi (Table 1). Most of them are involved in nutrition utilization [34] and community communication. For example, different type of carbon sources including monomer of plant biomass degradation and amino acids; fungal species-specific GPCR-mediated pheromone for sexual reproduction; volatile compounds and small molecules for inter- or intra-species communication (i.e., quorum sensing). Based on information of these identified sequences of TF protein and DNA binding motif, they are expected to be applied for biosensor development in other eukaryotic species.
3.2. Environmental Cues Responsive Systems
Environmental signals are initially perceived by receptor proteins—such as GPCRs, ion channels or HKs—located on the cell membrane. These signals are then transduced through distinct signaling pathways like MAPK pathways, amplified via cascades, and ultimately lead to alterations in the activity of TFs, thereby driving physiological adaptations to environmental perturbations. Accordingly, the following sections will discuss key components—including GPCRs, signal transduction pathways, and TFs—in the context of different environmental stimuli.
3.2.1. Responsive GPCR to Environmental Cues
GPCRs represent the largest class of cell surface receptors in eukaryotes, and GPCR signaling serves as a primary mechanism for eukaryotic cells to perceive and respond to environmental cues. In filamentous fungi, GPCRs exhibit remarkable diversity and can be classified into 17 major categories encompassing over 15,000 members based on predicted structural similarity and homology[30]. These receptors are involved in sensing nutrients, hormones, light, pH, temperature, and inter-population signaling. Selected GPCRs, along with their potential ligands, physiological functions, and species distribution, are summarized in Table 2.
Despite their abundance, the vast majority of fungal GPCRs remain poorly characterized. For example, while GPCRs have been systematically predicted in several model filamentous fungi[72,73], critical information regarding their activating ligands, activation mechanisms, and associated signaling components remains largely unknown. Moreover, structural studies on fungal GPCRs are even more scarce. A notable exception is the recent resolution of the yeast Ste2 receptor, which revealed significant structural differences compared to mammalian GPCRs—particularly in the position of transmembrane helix H4 and the conformation of the G protein-binding site. This discovery highlights the potential of targeting fungal-specific GPCRs for the development of antifungal agents[74]. Furthermore, structural studies on GPCR-ligand interactions will facilitate the engineering of filamentous fungal GPCRs, thereby enriching the synthetic biology toolbox.
3.2.2. Signaling Pathway of Environmental Cues
Environmental signals such as oxidative stress, pH, temperature, light, water availability (or osmolarity) and even electromagnetic fields collectively shape the living conditions of organisms. Environmental perturbations influence growth, development and secondary metabolism biosynthesis. Filamentous fungi have evolved diverse survival strategies and signaling pathways to sense these perturbations and adapt to fluctuating environments (Figure 4)[12]. The core signaling pathways responding to environmental cues in filamentous fungi include the TOR-MAPK, HOG-MAPK, CWI-MAPK, Ca2+ signaling pathway, and HK-MAPK pathways[40]. These MAPK pathways are highly interconnected. For example, the HOG/MAPK signaling pathway is involved in sensing water availability, mechanical force, oxidative stress, pH, thermal, or even light [104,105]. On the other hand, a single environmental signal is able to trigger multiple signaling pathways. For instance, osmotic stress can trigger the HOG-MAPK, CWI-MAPK, Ca2+pathway, and HK-MAPK pathway simultaneously[41]. In account of the complexity of signaling networks involved in environmental cues response, the discussion focuses only on the well-characterized pathways in filamentous fungi, including pH, oxidative stress, light, and temperature.
The Rim/Pal pathway is a fungal-specific and well-studied signaling pathway that responds to extracellular pH changes[106]. This pathway is conserved in filamentous fungi, at least within the Ascomycota phylum, and shares similarities with S.cerevisiae. As the Rim101 ortholog in yeast, PacC in Aspergillus species serves as the central TF mediating pH signaling and requires a two-step proteolytic processing to become functional [107].
Environmental cues such as oxygen levels, mechanical damage, and different nutrient types and abundance trigger intracellular oxidative stress responses. Current research indicates that oxidative stress signaling in S. cerevisia and Aspergillus species are highly conserved and involve both the canonical HK-Hog1 pathway and non-canonical pathways mediated by SrrA (Skn7 ortholog) and NapA (Yap1 ortholog) [108]. These pathways differ in their reactive oxygen species (ROS, e.g., H2O2, O•−) sensors and downstream TFs. The effector transcription factor for the HK-Hog1 pathway is AtfA (AP-1 ortholog). Hence, these TFs are functional across species of yeast and filamentous fungi[109].
Unlike S. cerevisiae, which lacks light-sensing capability, filamentous fungi generally are capable of perceiving light and can distinguish different wavelengths. Several photoreceptors for specific light spectra have been characterized (Table 3). These include cryptochrome CryA for near-UV light, WC-1 and Vivid for blue light,opsin for green light and phytochrome FphA for red light[110]. These photoreceptors exhibit distinct subcellular localizations (e.g., opsin on the plasma membrane, WC-1 in the nucleus, and FphA in the cytoplasm). Except WCC complex (WC-1 and WC-2), the downstream signal transduction mechanisms of these photoreceptors remain poorly understood. Triggered by blue light, the flavin-cystine adduct was formed in LOV domain to expose activation domain and stabilize WC-1 and WC-2 dimer, which thereby translocates to the nucleus and functions as a TF. The Hog1-Atf1 pathway has been reported to participate in FphA-mediated red light signaling[110]. The signaling mechanisms of cryptochromes (e.g., CryA) in response to blue light remain largely unexplored in fungi. In Arabidopsis thaliana, cryptochrome CryA directly binds the TF- CIB1 upon blue light stimulation to activate transcription initiation[111]. This example may provide insights for studying CryA signaling mechanisms in fungi. Green light receptors like NOP-1 have been identified in filamentous fungi. Green light induces photoisomerization of retinal from all-trans to 13-cis, which activate the coupled pump activity. The guanylyl cyclase effector domain in rhodopsin may couple light sensing with the production of the second messenger cGMP [112].
Microorganisms inherently exist in mechanically dynamic environments, where cell wall integrity (CWI) is crucial for maintaining osmotic homeostasis and resisting external mechanical forces. Membrane proteins Wsc1 and Mid2 function as mechanosensors; their extracellular domains undergo spring-like extension and compression, thereby initiating intracellular signaling. Additionally, the calcium channel proteins Mid1 and Cch1 are capable of perceiving mechanical stress and membrane damage. Downstream signaling pathways—including the CrzA-Ca²⁺ pathway, the CWI-MAPK cascade, and the SmuSH pathway—act in coordination to mediate responses to sustained mechanical pressure[113]. By contrast, research on fungal sensors for temperature stimuli—both heat and cold—remains notably limited.
3.2.3. Responsive TFs to Environmental Cues
A set of transcription factors (TFs) responsive to specific environmental cues have been identified (Table 4). How do these TFs perceive their cognate signals? What are their regulons or DNA-binding motifs? Most importantly, do they hold potential for development as biosensors? This review examines the identified TFs in light of these questions.
NapA, AtfA, MsnA (ortholog of yeast Msn2/4), and SrrA (ortholog of yeast Skn7) are TFs involved in the oxidative stress response. Upon H₂O₂ stimulation, NapA is oxidized by the peroxiredoxin PrxA in the cytoplasm, leading to its translocation into the nucleus to initiate transcription. NapA can be inactivated by thioredoxin TrxA reduction in both the cytoplasm and nucleus [150]. AtfA, a bZIP-type TF, interacts with SakA, the terminal kinase of the high-osmolarity glycerol (HOG) mitogen-activated protein kinase (MAPK) pathway, thereby serving as a key regulator in response to osmotic and oxidative stress [151]. MsnA, involved in various stress responses including oxidative stress, may interact with VelB; however, its upstream signaling components remain unclear. It contains multiple potential phosphorylation and ubiquitination sites, and its DNA-binding motif has recently been resolved [152]. SrrA, a Skn7-type response regulator containing an HSF-like DNA-binding domain and a receiver domain, is essential for H₂O₂ tolerance [153]. Nevertheless, information regarding its upstream regulators, downstream DNA-binding motifs, and target genes remains limited. SrbA, a basic helix-loop-helix (bHLH) TF belonging to the sterol regulatory element-binding protein (SREBP) family, has a well-defined DNA-binding motif [154]. The ΔsrbA mutant exhibits impaired growth under hypoxic conditions. Under low oxygen, full-length SrbA located in the endoplasmic reticulum undergoes proteolytic cleavage in a multi-factor dependent process, releasing its N-terminal bHLH domain into the nucleus to function as a transcriptional activator [155].
PacC is a well-studied TF responsive to alkaline pH, and its DNA-binding sequence has been biochemically validated [156]. The upstream regulation of PacC involves the membrane-associated proteins PalI, PalH, and PalF.
HsfA is the master regulator of thermal stress adaptation. Its activity is modulated by phosphorylation, although the specific upstream regulatory kinases remain unidentified. Despite functional conservation across eukaryotes, HsfA homologs exhibit variations in structure, post-translational modifications, and interacting partners [157]. In contrast to the extensive research on heat-responsive TFs, regulators of cold adaptation in fungi are poorly understood. Recently, the transcription factor Scaffold5.t61 was demonstrated to respond to low temperatures and regulate the synthesis of high-quality red pigment via modulation of glutamate metabolism [158].
Deciphering metal ion regulatory networks provides valuable insights for designing biosensors to monitor physiological or environmental metal ions and for developing bioremediation strategies. Ca²⁺ and Zn²⁺ are essential divalent cations involved in diverse biological processes. The primary regulator of calcium homeostasis is CrzA, which has been identified in Aspergillus [159]. In A. fumigatus, ZafA is the zinc-responsive TF responsible for zinc homeostasis and can functionally complement Zap1 in S. cerevisiae. Notably, Cd²⁺ can mimic the repressive effect of Zn²⁺ on ZafA. Cu²⁺-responsive TFs are widely reported in fungi including filamentous fungi. Mac1 is activated under copper-limiting conditions, whereas trans-activator AceA is induced by high copper concentrations [160,161]. However, specific responsive TFs for other biologically relevant divalent ions such as Mn²⁺, Co²⁺, and Ni²⁺ remain unidentified, although Ca²⁺-CrzA signaling has been implicated in Mn²⁺ sensing.
In fungi, key TFs involved in mechanosensing and cell wall integrity include Rlm1 and CrzA. Rlm1is a critical component of the CWI MAPK pathway. Upon mechanical stress or cell wall damage, the CWI pathway is activated, leading to phosphorylation of Rlm1 at specific sites (e.g., Ser427 and Thr435) by the Slt2 (Mpk1), which enhances its transcriptional activity [162]. A recent study revealed the DNA binding sequence of Rlm1 and showed that Rlm1 in F. graminearum can be activated by mycotoxins such as deoxynivalenol (DON) [163]. CrzA, a key effector of Ca²⁺-calcineurin signaling, translocates from the cytoplasm to the nucleus upon dephosphorylation in response to calcium stress, promoting the expression of cell wall biosynthesis-related genes. This nuclear translocation can be inhibited by cyclosporine A [164].
4. Mining Strategies of Sensing Elements in Filamentous Fungi
4.1. Genome Mining
- (1)
- Sequence-Based Genome Mining
The availability of rich genomic resources from diverse species provide an opportunity to discover natural sensing elements. For identified sensing proteins, conserved-domain alignment can be used to retrieve homologues with potentially superior performance. For example, XylR from Bacillus licheniformis have the largest dynamic range than other 5 homologues [173]. Moreover, the numerous predicted [174] and identified gene clusters, including biosynthetic gene clusters (BGCs) or catabolic gene clusters, from filamentous fungi could provide a platform for biosensor mining. In prokaryotes, it is common that the TF in a gene cluster bind the primary substrate degraded by the co-clustered enzymes, which implement feedforward activation regulation to switch on the catabolic pathway. Indeed, by leveraging this principle,16 metabolite-inducible systems, a new resveratrol-responsive biosensor and the erythritol induction systems were discovered in Cupriavidus necator H16[175], Novosphingobium aromaticivorans [176] and E. coli [177], respectively. Likewise, in white-rot fungi, aromatic compounds (e.g, guaiacol) induce the heterodimerization of TFs- TH8421 and TH4300, activating laccase genes to degrade these compounds [32]. Therefore, it is reasonable that there are more likewise examples showing the feedback or feedforward regulatory systems in biosynthetic or catabolic metabolism, thereby many metabolite-responsive systems are expected to exist in filamentous fungi. In future, a high-throughput mining strategy could be envisioned, involving in silico prediction of TF-ligand (substrate/product) pairs based on fungal gene clusters, and modular construction of native or synthetic TF-promoter pair for prototype test in desired chassis (Figure 5).
- (2)
- Structure-guided genome mining
As protein structure is often more evolutionarily conserved than sequence[178]. Ligand-binding-pocket-centric search can uncover remote homologues. Prior to DALI [179] and TM-align [180] , Foldseek [181] is presently the most widely used tool for rapid structural comparison and serve as a powerful complement to sequence-based mining.
4.2. Transcriptome-Guided Mining
Transcriptome-guided mining has been described [182]. In this way, stress and 1-butanol biosensor have been developed [182,183]. Briefly, this strategy involves defining the specific conditions of interest under a signal stimulus. Relevant transcriptome data under these conditions are systematically retrieved from databases or generated experimentally. Condition-specific and upregulated TFs are then identified as candidates. The information of TF binding sites (TFBS) can be retrieved from published literature or mapped from ChIP-seq datasets. When TFBSs are unavailable, fusing the TF to a well-characterized DBD [27] or clone the native, signal-responsive promoter as a putative output promoter can be alternative strategies. Finally, the TF and its cognate promoter are modularly assembled and engineered in selected host.
4.3. Deciphering Protein-Ligand Interaction Landscapes
Recently, the TF-ligand interaction landscape has been mapped in E.coli[184]. This methodology can be adapted for filamentous fungi species. It involves identifying candidate binding ligands for TFs/enzymes through affinity purification coupled with precision mass spectrometry. The pool of candidate ligands is then refined using AI-driven molecular docking. Finally, the binding affinity and functional induction are validated using standard compounds. This approach is anticipated to enable the batch development of a number of small-molecule inducible systems from filamentous fungi.
4.4. AI-Assisted Ligand Binding Site (LBS) Prediction
Various AI-driven tools have been developed for LBS prediction [185]. These tools can perform an initial screening against proteomes. By incorporating prior knowledge, the range of potential ligands can be progressively narrowed. Subsequently, AI tools can be employed to predict ligand binding pose[185]. This workflow aims to significantly reduce the experimental burden prior to validation.
5. Concluding Remark and Future Perspectives
As key components of signal sensing, GPCRs, TFs, and photoreceptors serve as fundamental genetic parts in biosensor development. Due to the inherent limitations of prokaryotic components, the development of fungal-derived components orthogonal to yeast from filamentous fungi should receive greater attention, as it can not only compensate for the shortcomings of bacterial components but also leverage the advantages of eukaryotic systems.
The application scenarios of biosensors are extremely broad, such as in biomedical and materials science. Biosensor can be integrated to facilitate drug discovery and screening, intelligent drug delivery, therapeutic drug monitoring,point-of-care drug/pathogen diagnostics and living material fabrication. For example, filamentous fungi are well known for producing key pharmaceuticals, including penicillin, griseofulvin, lovastatin, cyclosporine, and ergometrine. Monitoring their pharmacokinetics become necessary due to potential issues with dosage and toxic side effects. The native molecular responsive elements-based living biosensors could provide a safter and faster solution for monitoring.
Despite the growing demand of biosensors, research on identification of signal-receptor pairs lag far behind. Particularly, in filamentous fungi, most of genetic resources are still “dark matter”. Therefore, the pressing challenge is to identify the ligands of a large number of orphan GPCRs, zinc cluster TFs and photoreceptors. The established modular construction tools, AI tools and yeast synthetic GPCR system could provide effective and efficient solutions to deorphanize these sensing elements, which are poised to significantly accelerate advancement in synthetic biology.
Author Contributions
Conceptualization, S.G. and Y.C.; Writing—Original Draft Preparation, S.G.; Writing—Review and Editing, S.G., Y.G., S.S., Z.L., Y.C.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by start-up fund of Wuxi Taihu University [2025THQD024]. The funders had no role in study design, data collection and analysis, publication decisions, or manuscript preparation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Qin, L.; Liu, X.; Xu, K.; Li, C. Mining and design of biosensors for engineering microbial cell factory. Curr Opin Biotechnol 2022, 75, 102694. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Lei, Q.Y. Metabolite sensing and signaling in cell metabolism. Signal Transduct Target Ther 2018, 3, 30. [Google Scholar] [CrossRef]
- Newman, R.H.; Fosbrink, M.D.; Zhang, J. Genetically encodable fluorescent biosensors for tracking signaling dynamics in living cells. Chem Rev 2011, 111, 3614–3666. [Google Scholar] [CrossRef]
- Zhang, C.; Wei, Z.H.; Ye, B.C. Imaging and tracing of intracellular metabolites utilizing genetically encoded fluorescent biosensors. Biotechnol J 2013, 8, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Mo, G.C.; Ross, B.; Hertel, F.; Manna, P.; Yang, X.; Greenwald, E.; Booth, C.; Plummer, A.M.; Tenner, B.; Chen, Z.; et al. Genetically encoded biosensors for visualizing live-cell biochemical activity at super-resolution. Nat Methods 2017, 14, 427–434. [Google Scholar] [CrossRef]
- Huang, C.W.; Lin, C.; Nguyen, M.K.; Hussain, A.; Bui, X.T.; Ngo, H.H. A review of biosensor for environmental monitoring: principle, application, and corresponding achievement of sustainable development goals. Bioengineered 2023, 14, 58–80. [Google Scholar] [CrossRef]
- Wang, B.; Barahona, M.; Buck, M. A modular cell-based biosensor using engineered genetic logic circuits to detect and integrate multiple environmental signals. Biosens Bioelectron 2013, 40, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.D.; Garruss, A.S.; Moretti, R.; Chan, S.; Arbing, M.A.; Cascio, D.; Rogers, J.K.; Isaacs, F.J.; Kosuri, S.; Baker, D.; et al. Engineering an allosteric transcription factor to respond to new ligands. Nat Methods 2016, 13, 177–183. [Google Scholar] [CrossRef]
- Tombelli, S.; Minunni, M.; Mascini, M. Analytical applications of aptamers. Biosens Bioelectron 2005, 20, 2424–2434. [Google Scholar] [CrossRef]
- Sefah, K.; Phillips, J.A.; Xiong, X.; Meng, L.; Van Simaeys, D.; Chen, H.; Martin, J.; Tan, W. Nucleic acid aptamers for biosensors and bio-analytical applications. Analyst 2009, 134, 1765–1775. [Google Scholar] [CrossRef]
- Beier, D.; Gross, R. Regulation of bacterial virulence by two-component systems. Curr Opin Microbiol 2006, 9, 143–152. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Xue, C.; Idnurm, A.; Rutherford, J.C.; Heitman, J.; Cardenas, M.E. Sensing the environment: lessons from fungi. Nat Rev Microbiol 2007, 5, 57–69. [Google Scholar] [CrossRef]
- Prole, D.L.; Taylor, C.W. Identification and analysis of cation channel homologues in human pathogenic fungi. PLoS One 2012, 7, e42404. [Google Scholar] [CrossRef]
- Naar, A.M.; Thakur, J.K. Nuclear receptor-like transcription factors in fungi. Genes Dev 2009, 23, 419–432. [Google Scholar] [CrossRef] [PubMed]
- D'Ambrosio, V.; Jensen, M.K. Lighting up yeast cell factories by transcription factor-based biosensors. FEMS Yeast Res 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Kondo, A.; Ishii, J. Engineering of Synthetic Transcriptional Switches in Yeast. Life (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Meng, F.; Shaw, W.M.; Kam, Y.K.K.; Ellis, T. Engineering yeast multicellular behaviors via synthetic adhesion and contact signaling. Cell 2025, 188, 4936–4949 e4914. [Google Scholar] [CrossRef]
- Qiao, L.; Niu, L.; Wang, Z.; Dai, D.; Tang, S.; Ma, X.; Deng, Z.; Yu, G.; Zhou, Y.; Yan, T.; et al. Optogenetic-Controlled iPSC-Based Vaccines for Prophylactic and Therapeutic Tumor Suppression in Mice. Adv Sci (Weinh) 2025, 12, e16115. [Google Scholar] [CrossRef] [PubMed]
- Krueger, D.; Izquierdo, E.; Viswanathan, R.; Hartmann, J.; Pallares Cartes, C.; De Renzis, S. Principles and applications of optogenetics in developmental biology. Development 2019, 146. [Google Scholar] [CrossRef]
- Gligorovski, V.; Sadeghi, A.; Rahi, S.J. Multidimensional characterization of inducible promoters and a highly light-sensitive LOV-transcription factor. Nat Commun 2023, 14, 3810. [Google Scholar] [CrossRef]
- Acar, M.; Becskei, A.; van Oudenaarden, A. Enhancement of cellular memory by reducing stochastic transitions. Nature 2005, 435, 228–232. [Google Scholar] [CrossRef]
- Endalur Gopinarayanan, V.; Nair, N.U. A semi-synthetic regulon enables rapid growth of yeast on xylose. Nat Commun 2018, 9, 1233. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Zhao, H. Design and engineering of intracellular-metabolite-sensing/regulation gene circuits in Saccharomyces cerevisiae. Biotechnol Bioeng 2016, 113, 206–215. [Google Scholar] [CrossRef]
- Skjoedt, M.L.; Snoek, T.; Kildegaard, K.R.; Arsovska, D.; Eichenberger, M.; Goedecke, T.J.; Rajkumar, A.S.; Zhang, J.; Kristensen, M.; Lehka, B.J.; et al. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat Chem Biol 2016, 12, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Snoek, T.; Chaberski, E.K.; Ambri, F.; Kol, S.; Bjorn, S.P.; Pang, B.; Barajas, J.F.; Welner, D.H.; Jensen, M.K.; Keasling, J.D. Evolution-guided engineering of small-molecule biosensors. Nucleic Acids Res 2020, 48, e3. [Google Scholar] [CrossRef] [PubMed]
- Stanton, B.C.; Nielsen, A.A.; Tamsir, A.; Clancy, K.; Peterson, T.; Voigt, C.A. Genomic mining of prokaryotic repressors for orthogonal logic gates. Nat Chem Biol 2014, 10, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Du, J.; Li, D.; Xiong, J.; Chen, Y. Versatile xylose and arabinose genetic switches development for yeasts. Metab Eng 2025, 87, 21–36. [Google Scholar] [CrossRef]
- Naseri, G.; Koffas, M.A.G. Application of combinatorial optimization strategies in synthetic biology. Nat Commun 2020, 11, 2446. [Google Scholar] [CrossRef]
- Belli, G.; Gari, E.; Piedrafita, L.; Aldea, M.; Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Res 1998, 26, 942–947. [Google Scholar] [CrossRef]
- Liu, Z.; Salamov, A.; Grigoriev, I.V. Identification and classification of fungal GPCR gene families 2011. 2025.625238. bioRxiv 2024, 2024. [Google Scholar]
- Pardee, K.; Necakov, A.S.; Krause, H. Nuclear Receptors: Small Molecule Sensors that Coordinate Growth, Metabolism and Reproduction. Subcell Biochem 2011, 52, 123–153. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, X.; Wu, K.; Liu, S.; Li, X.; Zhu, C.; Xiao, Y.; Fang, Z.; Liu, J. Two Zn(2)Cys(6)-type transcription factors respond to aromatic compounds and regulate the expression of laccases in the white-rot fungus Trametes hirsuta. Appl Environ Microbiol 2024, 90, e0054524. [Google Scholar] [CrossRef]
- Zhang, Y.; Cortez, J.D.; Hammer, S.K.; Carrasco-Lopez, C.; Garcia Echauri, S.A.; Wiggins, J.B.; Wang, W.; Avalos, J.L. Biosensor for branched-chain amino acid metabolism in yeast and applications in isobutanol and isopentanol production. Nat Commun 2022, 13, 270. [Google Scholar] [CrossRef]
- Kerkaert, J.D.; Huberman, L.B. Regulation of nutrient utilization in filamentous fungi. Appl Microbiol Biotechnol 2023, 107, 5873–5898. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: regulation, function and drug discovery. Nat Rev Microbiol 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Mozsik, L.; Iacovelli, R.; Bovenberg, R.A.L.; Driessen, A.J.M. Transcriptional Activation of Biosynthetic Gene Clusters in Filamentous Fungi. Front Bioeng Biotechnol 2022, 10, 901037. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.; Cubero, B.; Cecchetto, G.; Scazzocchio, C. PrnA, a Zn2Cys6 activator with a unique DNA recognition mode, requires inducer for in vivo binding. Mol Microbiol 2002, 44, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. How light affects the life of Botrytis. Fungal Genet Biol 2017, 106, 26–41. [Google Scholar] [CrossRef]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryot Cell 2003, 2, 1151–1161. [Google Scholar] [CrossRef]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and Role of Fungal Secondary Metabolites. Annu Rev Genet 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Banuett, F. Signalling in the yeasts: an informational cascade with links to the filamentous fungi. Microbiol Mol Biol Rev 1998, 62, 249–274. [Google Scholar] [CrossRef] [PubMed]
- Kraakman, L.; Lemaire, K.; Ma, P.; Teunissen, A.W.; Donaton, M.C.; Van Dijck, P.; Winderickx, J.; de Winde, J.H.; Thevelein, J.M. A Saccharomyces cerevisiae G-protein coupled receptor, Gpr1, is specifically required for glucose activation of the cAMP pathway during the transition to growth on glucose. Mol Microbiol 1999, 32, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Sze, J.Y.; Woontner, M.; Jaehning, J.A.; Kohlhaw, G.B. In vitro transcriptional activation by a metabolic intermediate: activation by Leu3 depends on alpha-isopropylmalate. Science 1992, 258, 1143–1145. [Google Scholar] [CrossRef] [PubMed]
- Lengeler, K.B.; Davidson, R.C.; D'Souza, C.; Harashima, T.; Shen, W.C.; Wang, P.; Pan, X.; Waugh, M.; Heitman, J. Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 2000, 64, 746–785. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, K.; Qin, Q.; Lin, G.; Hu, T.; Xu, Z.; Wang, S. G Protein alpha Subunit GpaB is Required for Asexual Development, Aflatoxin Biosynthesis and Pathogenicity by Regulating cAMP Signaling in Aspergillus flavus. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Yang, K.; Qin, Q.; Liu, Y.; Zhang, L.; Liang, L.; Lan, H.; Chen, C.; You, Y.; Zhang, F.; Wang, S. Adenylate Cyclase AcyA Regulates Development, Aflatoxin Biosynthesis and Fungal Virulence in Aspergillus flavus. Front Cell Infect Microbiol 2016, 6, 190. [Google Scholar] [CrossRef]
- van Peij, N.N.; Visser, J.; de Graaff, L.H. Isolation and analysis of xlnR, encoding a transcriptional activator co-ordinating xylanolytic expression in Aspergillus niger. Mol Microbiol 1998, 27, 131–142. [Google Scholar] [CrossRef]
- Battaglia, E.; Hansen, S.F.; Leendertse, A.; Madrid, S.; Mulder, H.; Nikolaev, I.; de Vries, R.P. Regulation of pentose utilisation by AraR, but not XlnR, differs in Aspergillus nidulans and Aspergillus niger. Appl Microbiol Biotechnol 2011, 91, 387–397. [Google Scholar] [CrossRef]
- Ogawa, M.; Kobayashi, T.; Koyama, Y. ManR, a novel Zn(II)2Cys6 transcriptional activator, controls the beta-mannan utilization system in Aspergillus oryzae. Fungal Genet Biol 2012, 49, 987–995. [Google Scholar] [CrossRef]
- Yuan, X.L.; Roubos, J.A.; van den Hondel, C.A.; Ram, A.F. Identification of InuR, a new Zn(II)2Cys6 transcriptional activator involved in the regulation of inulinolytic genes in Aspergillus niger. Mol Genet Genomics 2008, 279, 11–26. [Google Scholar] [CrossRef]
- Pardo, E.; Orejas, M. The Aspergillus nidulans Zn(II)2Cys6 transcription factor AN5673/RhaR mediates L-rhamnose utilization and the production of α-L-rhamnosidases. Microbial cell factories 2014, 13, 161. [Google Scholar] [PubMed]
- Kunitake, E.; Tani, S.; Sumitani, J.; Kawaguchi, T. A novel transcriptional regulator, ClbR, controls the cellobiose- and cellulose-responsive induction of cellulase and xylanase genes regulated by two distinct signaling pathways in Aspergillus aculeatus. Appl Microbiol Biotechnol 2013, 97, 2017–2028. [Google Scholar] [CrossRef]
- Alazi, E.; Niu, J.; Kowalczyk, J.E.; Peng, M.; Aguilar Pontes, M.V.; van Kan, J.A.; Visser, J.; de Vries, R.P.; Ram, A.F. The transcriptional activator GaaR of Aspergillus niger is required for release and utilization of d-galacturonic acid from pectin. FEBS Lett 2016, 590, 1804–1815. [Google Scholar] [CrossRef] [PubMed]
- Makita, T.; Katsuyama, Y.; Tani, S.; Suzuki, H.; Kato, N.; Todd, R.B.; Hynes, M.J.; Tsukagoshi, N.; Kato, M.; Kobayashi, T. Inducer-dependent nuclear localization of a Zn(II)(2)Cys(6) transcriptional activator, AmyR, in Aspergillus nidulans. Biosci Biotechnol Biochem 2009, 73, 391–399. [Google Scholar] [CrossRef]
- Zhao, Q.; Gao, L.; Xu, N.; Zhang, X.; Qin, Y.; Qu, Y.; Liu, G. An l-fucose-responsive transcription factor cross-regulates the expression of a diverse array of carbohydrate-active enzymes in Trichoderma reesei. Plos Genetics 2025, 21, e1011815. [Google Scholar] [CrossRef] [PubMed]
- Arentshorst, M.; Reijngoud, J.; van Tol, D.J.C.; Reid, I.D.; Arendsen, Y.; Pel, H.J.; van Peij, N.; Visser, J.; Punt, P.J.; Tsang, A.; et al. Utilization of ferulic acid in Aspergillus niger requires the transcription factor FarA and a newly identified Far-like protein (FarD) that lacks the canonical Zn(II)(2)Cys(6) domain. Front Fungal Biol 2022, 3, 978845. [Google Scholar] [CrossRef]
- Choi, H.; Cho, S.H.; Park, J.H.; Seok, Y.J. Fructose-responsive regulation by FruR in Faecalibacterium prausnitzii for its intestinal colonization. Commun Biol 2025, 8, 426. [Google Scholar] [CrossRef]
- Mira, N.P.; Henriques, S.F.; Keller, G.; Teixeira, M.C.; Matos, R.G.; Arraiano, C.M.; Winge, D.R.; Sa-Correia, I. Identification of a DNA-binding site for the transcription factor Haa1, required for Saccharomyces cerevisiae response to acetic acid stress. Nucleic Acids Res 2011, 39, 6896–6907. [Google Scholar] [CrossRef]
- Punt, P.J.; Strauss, J.; Smit, R.; Kinghorn, J.R.; van den Hondel, C.A.; Scazzocchio, C. The intergenic region between the divergently transcribed niiA and niaD genes of Aspergillus nidulans contains multiple NirA binding sites which act bidirectionally. Mol Cell Biol 1995, 15, 5688–5699. [Google Scholar] [CrossRef]
- Keller, S.; Macheleidt, J.; Scherlach, K.; Schmaler-Ripcke, J.; Jacobsen, I.D.; Heinekamp, T.; Brakhage, A.A. Pyomelanin formation in Aspergillus fumigatus requires HmgX and the transcriptional activator HmgR but is dispensable for virulence. PLoS One 2011, 6, e26604. [Google Scholar] [CrossRef]
- Long, N.; Orasch, T.; Zhang, S.; Gao, L.; Xu, X.; Hortschansky, P.; Ye, J.; Zhang, F.; Xu, K.; Gsaller, F.; et al. The Zn2Cys6-type transcription factor LeuB cross-links regulation of leucine biosynthesis and iron acquisition in Aspergillus fumigatus. PLoS Genet 2018, 14, e1007762. [Google Scholar] [CrossRef]
- Dzikowska, A.; Kacprzak, M.; Tomecki, R.; Koper, M.; Scazzocchio, C.; Weglenski, P. Specific induction and carbon/nitrogen repression of arginine catabolism gene of Aspergillus nidulans--functional in vivo analysis of the otaA promoter. Fungal Genet Biol 2003, 38, 175–186. [Google Scholar] [CrossRef]
- Natorff, R.; Sienko, M.; Brzywczy, J.; Paszewski, A. The Aspergillus nidulans metR gene encodes a bZIP protein which activates transcription of sulphur metabolism genes. Mol Microbiol 2003, 49, 1081–1094. [Google Scholar] [CrossRef]
- Wong Sak Hoi, J.; Dumas, B. Ste12 and Ste12-like proteins, fungal transcription factors regulating development and pathogenicity. Eukaryot Cell 2010, 9, 480–485. [Google Scholar] [CrossRef]
- Kulmburg, P.; Mathieu, M.; Dowzer, C.; Kelly, J.; Felenbok, B. Specific binding sites in the alcR and alcA promoters of the ethanol regulon for the CREA repressor mediating carbon catabolite repression in Aspergillus nidulans. Mol Microbiol 1993, 7, 847–857. [Google Scholar] [CrossRef]
- Miyamoto, K.; Murakami, T.; Kakumyan, P.; Keller, N.P.; Matsui, K. Formation of 1-octen-3-ol from Aspergillus flavus conidia is accelerated after disruption of cells independently of Ppo oxygenases, and is not a main cause of inhibition of germination. PeerJ 2014, 2, e395. [Google Scholar] [CrossRef]
- Ahn, C.H.; Lee, S.; Cho, E.; Kim, H.; Chung, B.; Park, W.; Shin, J.; Oh, K.B. A farnesoic acid-responsive transcription factor, Hot1, regulates yeast-hypha morphogenesis in Candida albicans. FEBS Lett 2017, 591, 1225–1235. [Google Scholar] [CrossRef]
- Hernday, A.D.; Lohse, M.B.; Fordyce, P.M.; Nobile, C.J.; DeRisi, J.L.; Johnson, A.D. Structure of the transcriptional network controlling white-opaque switching in Candida albicans. Mol Microbiol 2013, 90, 22–35. [Google Scholar] [CrossRef]
- Lee, K.; Hahn, J.S. Interplay of Aro80 and GATA activators in regulation of genes for catabolism of aromatic amino acids in Saccharomyces cerevisiae. Mol Microbiol 2013, 88, 1120–1134. [Google Scholar] [CrossRef]
- Tian, X.; He, G.J.; Hu, P.; Chen, L.; Tao, C.; Cui, Y.L.; Shen, L.; Ke, W.; Xu, H.; Zhao, Y.; et al. Cryptococcus neoformans sexual reproduction is controlled by a quorum sensing peptide. Nat Microbiol 2018, 3, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Potter, C.J.; Tasic, B.; Russler, E.V.; Liang, L.; Luo, L. The Q system: a repressible binary system for transgene expression, lineage tracing, and mosaic analysis. Cell 2010, 141, 536–548. [Google Scholar] [CrossRef]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global survey of canonical Aspergillus flavus G protein-coupled receptors. mBio 2014, 5, e01501-01514. [Google Scholar] [CrossRef]
- Cabrera, I.E.; Pacentine, I.V.; Lim, A.; Guerrero, N.; Krystofova, S.; Li, L.; Michkov, A.V.; Servin, J.A.; Ahrendt, S.R.; Carrillo, A.J.; et al. Global Analysis of Predicted G Protein-Coupled Receptor Genes in the Filamentous Fungus, Neurospora crassa. G3 (Bethesda) 2015, 5, 2729–2743. [Google Scholar] [CrossRef]
- Velazhahan, V.; Ma, N.; Vaidehi, N.; Tate, C.G. Activation mechanism of the class D fungal GPCR dimer Ste2. Nature 2022, 603, 743–748. [Google Scholar] [CrossRef]
- Hoffman, C.S. Glucose sensing via the protein kinase A pathway in Schizosaccharomyces pombe. Biochem Soc Trans 2005, 33, 257–260. [Google Scholar] [CrossRef]
- Li, L.; Borkovich, K.A. GPR-4 is a predicted G-protein-coupled receptor required for carbon source-dependent asexual growth and development in Neurospora crassa. Eukaryot Cell 2006, 5, 1287–1300. [Google Scholar] [CrossRef]
- de Souza, W.R.; Morais, E.R.; Krohn, N.G.; Savoldi, M.; Goldman, M.H.; Rodrigues, F.; Caldana, C.; Semelka, C.T.; Tikunov, A.P.; Macdonald, J.M.; et al. Identification of metabolic pathways influenced by the G-protein coupled receptors GprB and GprD in Aspergillus nidulans. PLoS One 2013, 8, e62088. [Google Scholar] [CrossRef]
- Xue, C.; Bahn, Y.S.; Cox, G.M.; Heitman, J. G protein-coupled receptor Gpr4 senses amino acids and activates the cAMP-PKA pathway in Cryptococcus neoformans. Mol Biol Cell 2006, 17, 667–679. [Google Scholar] [CrossRef]
- Jung, M.G.; Kim, S.S.; Yu, J.H.; Shin, K.S. Characterization of gprK Encoding a Putative Hybrid G-Protein-Coupled Receptor in Aspergillus fumigatus. PLoS One 2016, 11, e0161312. [Google Scholar] [CrossRef]
- Brown, N.A.; Dos Reis, T.F.; Ries, L.N.; Caldana, C.; Mah, J.H.; Yu, J.H.; Macdonald, J.M.; Goldman, G.H. G-protein coupled receptor-mediated nutrient sensing and developmental control in Aspergillus nidulans. Mol Microbiol 2015, 98, 420–439. [Google Scholar] [CrossRef]
- Chung, K.S.; Won, M.; Lee, S.B.; Jang, Y.J.; Hoe, K.L.; Kim, D.U.; Lee, J.W.; Kim, K.W.; Yoo, H.S. Isolation of a novel gene from Schizosaccharomyces pombe: stm1+ encoding a seven-transmembrane loop protein that may couple with the heterotrimeric Galpha 2 protein, Gpa2. J Biol Chem 2001, 276, 40190–40201. [Google Scholar] [CrossRef]
- Blumer, K.; Reneke, J.; Thorner, J. The STE2 gene product is the ligand-binding component of the alpha-factor receptor of Saccharomyces cerevisiae. Journal of Biological Chemistry 1988, 263, 10836–10842. [Google Scholar] [CrossRef]
- Seo, J.A.; Han, K.H.; Yu, J.H. The gprA and gprB genes encode putative G protein-coupled receptors required for self-fertilization in Aspergillus nidulans. Mol Microbiol 2004, 53, 1611–1623. [Google Scholar] [CrossRef]
- Kim, H.; Borkovich, K.A. A pheromone receptor gene, pre-1, is essential for mating type-specific directional growth and fusion of trichogynes and female fertility in Neurospora crassa. Molecular microbiology 2004, 52, 1781–1798. [Google Scholar] [CrossRef]
- Xue, C.; Wang, Y.; Hsueh, Y.P. Assessment of constitutive activity of a G protein-coupled receptor, CPR2, in Cryptococcus neoformans by heterologous and homologous methods. Methods Enzymol 2010, 484, 397–412. [Google Scholar] [CrossRef]
- McClelland, C.M.; Fu, J.; Woodlee, G.L.; Seymour, T.S.; Wickes, B.L. Isolation and characterization of the Cryptococcus neoformans MATa pheromone gene. Genetics 2002, 160, 935–947. [Google Scholar] [CrossRef]
- Nakayama, N.; Miyajima, A.; Arai, K. Nucleotide sequences of STE2 and STE3, cell type-specific sterile genes from Saccharomyces cerevisiae. EMBO J 1985, 4, 2643–2648. [Google Scholar] [CrossRef]
- Kim, H.; Borkovich, K.A. Pheromones are essential for male fertility and sufficient to direct chemotropic polarized growth of trichogynes during mating in Neurospora crassa. Eukaryot Cell 2006, 5, 544–554. [Google Scholar] [CrossRef]
- Moore, T.D.; Edman, J.C. The alpha-mating type locus of Cryptococcus neoformans contains a peptide pheromone gene. Mol Cell Biol 1993, 13, 1962–1970. [Google Scholar] [CrossRef]
- Lara-Martinez, D.; Tristan-Flores, F.E.; Cervantes-Montelongo, J.A.; Silva-Martinez, G.A. Fungal Stress Responses and the Importance of GPCRs. J Fungi (Basel) 2025, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Li, N.; Li, J.; Trail, F.; Dunlap, J.C.; Townsend, J.P. Light sensing by opsins and fungal ecology: NOP-1 modulates entry into sexual reproduction in response to environmental cues. Mol Ecol 2018, 27, 216–232. [Google Scholar] [CrossRef]
- Garcia-Martinez, J.; Brunk, M.; Avalos, J.; Terpitz, U. The CarO rhodopsin of the fungus Fusarium fujikuroi is a light-driven proton pump that retards spore germination. Sci Rep 2015, 5, 7798. [Google Scholar] [CrossRef]
- Bieszke, J.A. Characterization of the NOP-1 opsin photoreceptor in the filamentous fungus Neurospora crassa; The University of Texas Graduate School of Biomedical Sciences at Houston, 2002. [Google Scholar]
- Herranz, S.; Rodriguez, J.M.; Bussink, H.J.; Sanchez-Ferrero, J.C.; Arst, H.N., Jr.; Penalva, M.A.; Vincent, O. Arrestin-related proteins mediate pH signaling in fungi. Proc Natl Acad Sci U S A 2005, 102, 12141–12146. [Google Scholar] [CrossRef]
- Gao, J.; Xu, X.; Huang, K.; Liang, Z. Fungal G-Protein-Coupled Receptors: A Promising Mediator of the Impact of Extracellular Signals on Biosynthesis of Ochratoxin A. Front Microbiol 2021, 12, 631392. [Google Scholar] [CrossRef]
- Gehrke, A.; Heinekamp, T.; Jacobsen, I.D.; Brakhage, A.A. Heptahelical receptors GprC and GprD of Aspergillus fumigatus Are essential regulators of colony growth, hyphal morphogenesis, and virulence. Appl Environ Microbiol 2010, 76, 3989–3998. [Google Scholar] [CrossRef]
- Kuo, C.Y.; Tay, R.J.; Lin, H.C.; Juan, S.C.; Vidal-Diez de Ulzurrun, G.; Chang, Y.C.; Hoki, J.; Schroeder, F.C.; Hsueh, Y.P. The nematode-trapping fungus Arthrobotrys oligospora detects prey pheromones via G protein-coupled receptors. Nat Microbiol 2024, 9, 1738–1751. [Google Scholar] [CrossRef]
- Hu, X.; Hoffmann, D.S.; Wang, M.; Schuhmacher, L.; Stroe, M.C.; Schreckenberger, B.; Elstner, M.; Fischer, R. GprC of the nematode-trapping fungus Arthrobotrys flagrans activates mitochondria and reprograms fungal cells for nematode hunting. Nat Microbiol 2024, 9, 1752–1763. [Google Scholar] [CrossRef]
- Ballou, E.R.; Avelar, G.M.; Childers, D.S.; Mackie, J.; Bain, J.M.; Wagener, J.; Kastora, S.L.; Panea, M.D.; Hardison, S.E.; Walker, L.A.; et al. Lactate signalling regulates fungal beta-glucan masking and immune evasion. Nat Microbiol 2016, 2, 16238. [Google Scholar] [CrossRef]
- Affeldt, K.J.; Brodhagen, M.; Keller, N.P. Aspergillus oxylipin signaling and quorum sensing pathways depend on g protein-coupled receptors. Toxins (Basel) 2012, 4, 695–717. [Google Scholar] [CrossRef]
- Kulkarni, R.D.; Thon, M.R.; Pan, H.; Dean, R.A. Novel G-protein-coupled receptor-like proteins in the plant pathogenic fungus Magnaporthe grisea. Genome Biol 2005, 6, R24. [Google Scholar] [CrossRef]
- Brown, N.A.; Schrevens, S.; van Dijck, P.; Goldman, G.H. Fungal G-protein-coupled receptors: mediators of pathogenesis and targets for disease control. Nat Microbiol 2018, 3, 402–414. [Google Scholar] [CrossRef]
- Shang, J.; Shang, Y.; Tang, G.; Wang, C. Identification of a key G-protein coupled receptor in mediating appressorium formation and fungal virulence against insects. Sci China Life Sci 2021, 64, 466–477. [Google Scholar] [CrossRef]
- Fuchs, B.B.; Mylonakis, E. Our paths might cross: the role of the fungal cell wall integrity pathway in stress response and cross talk with other stress response pathways. Eukaryotic cell 2009, 8, 1616–1625. [Google Scholar] [CrossRef]
- Yu, Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat Microbiol 2016, 1, 16019. [Google Scholar] [CrossRef]
- Cornet, M.; Gaillardin, C. pH signaling in human fungal pathogens: a new target for antifungal strategies. Eukaryot Cell 2014, 13, 342–352. [Google Scholar] [CrossRef]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Penalva, M.A.; Arst, H.N., Jr. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid- and alkaline-expressed genes by ambient pH. EMBO J 1995, 14, 779–790. [Google Scholar] [CrossRef]
- Zadrąg-Tęcza, R.; Maślanka, R.; Bednarska, S.; Kwolek-Mirek, M. Response mechanisms to oxidative stress in yeast and filamentous fungi. Stress response mechanisms in fungi: theoretical and practical aspects 2018, 1–34. [Google Scholar]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.P.; Papon, N. Oxidative stress response pathways in fungi. Cell Mol Life Sci 2022, 79, 333. [Google Scholar] [CrossRef]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat Rev Microbiol 2019, 17, 25–36. [Google Scholar] [CrossRef]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef]
- Avelar, G.M.; Schumacher, R.I.; Zaini, P.A.; Leonard, G.; Richards, T.A.; Gomes, S.L. A rhodopsin-guanylyl cyclase gene fusion functions in visual perception in a fungus. Curr Biol 2014, 24, 1234–1240. [Google Scholar] [CrossRef]
- Mishra, R.; Minc, N.; Peter, M. Cells under pressure: how yeast cells respond to mechanical forces. Trends in Microbiology 2022, 30, 495–510. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science 2002, 297, 815–819. [Google Scholar] [CrossRef]
- Guerrero, M.; Ruiz, C.; Romero, A.; Robeson, L.; Ruiz, D.; Salinas, F. The N-Terminal Region of the BcWCL1 Photoreceptor Is Necessary for Self-Dimerization and Transcriptional Activation upon Light Stimulation in Yeast. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Tang, Y.; Zhu, P.; Lu, Z.; Qu, Y.; Huang, L.; Zheng, N.; Wang, Y.; Nie, H.; Jiang, Y.; Xu, L. The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Qi, Y.; Sun, X.; Ma, L.; Wen, Q.; Qiu, L.; Shen, J. Identification of two Pleurotus ostreatus blue light receptor genes (PoWC-1 and PoWC-2) and in vivo confirmation of complex PoWC-12 formation through yeast two hybrid system. Fungal Biol 2020, 124, 8–14. [Google Scholar] [CrossRef]
- Krobanan, K.; Liang, S.W.; Chiu, H.C.; Shen, W.C. The Blue-Light Photoreceptor Sfwc-1 Gene Regulates the Phototropic Response and Fruiting-Body Development in the Homothallic Ascomycete Sordaria fimicola. Appl Environ Microbiol 2019, 85. [Google Scholar] [CrossRef]
- Casas-Flores, S.; Rios-Momberg, M.; Bibbins, M.; Ponce-Noyola, P.; Herrera-Estrella, A. BLR-1 and BLR-2, key regulatory elements of photoconidiation and mycelial growth in Trichoderma atroviride. Microbiology (Reading) 2004, 150, 3561–3569. [Google Scholar] [CrossRef]
- Purschwitz, J.; Muller, S.; Kastner, C.; Schoser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr Biol 2008, 18, 255–259. [Google Scholar] [CrossRef]
- Schuhmacher, L.; Heck, S.; Pitz, M.; Mathey, E.; Lamparter, T.; Blumhofer, A.; Leister, K.; Fischer, R. The LOV-domain blue-light receptor LreA of the fungus Alternaria alternata binds predominantly FAD as chromophore and acts as a light and temperature sensor. J Biol Chem 2024, 300, 107238. [Google Scholar] [CrossRef]
- Sanz, C.; Rodriguez-Romero, J.; Idnurm, A.; Christie, J.M.; Heitman, J.; Corrochano, L.M.; Eslava, A.P. Phycomyces MADB interacts with MADA to form the primary photoreceptor complex for fungal phototropism. Proc Natl Acad Sci U S A 2009, 106, 7095–7100. [Google Scholar] [CrossRef]
- Schwerdtfeger, C.; Linden, H. VIVID is a flavoprotein and serves as a fungal blue light photoreceptor for photoadaptation. EMBO J 2003, 22, 4846–4855. [Google Scholar] [CrossRef]
- Foley, B.J.; Stutts, H.; Schmitt, S.L.; Lokhandwala, J.; Nagar, A.; Zoltowski, B.D. Characterization of a Vivid Homolog in Botrytis cinerea. Photochem Photobiol 2018, 94, 985–993. [Google Scholar] [CrossRef]
- Schmoll, M.; Franchi, L.; Kubicek, C.P. Envoy, a PAS/LOV domain protein of Hypocrea jecorina (Anamorph Trichoderma reesei), modulates cellulase gene transcription in response to light. Eukaryot Cell 2005, 4, 1998–2007. [Google Scholar] [CrossRef]
- Losi, A.; Gartner, W. A light life together: photosensing in the plant microbiota. Photochem Photobiol Sci 2021, 20, 451–473. [Google Scholar] [CrossRef]
- Bieszke, J.A.; Braun, E.L.; Bean, L.E.; Kang, S.; Natvig, D.O.; Borkovich, K.A. The nop-1 gene of Neurospora crassa encodes a seven transmembrane helix retinal-binding protein homologous to archaeal rhodopsins. Proc Natl Acad Sci U S A 1999, 96, 8034–8039. [Google Scholar] [CrossRef]
- Panzer, S.; Brych, A.; Batschauer, A.; Terpitz, U. Opsin 1 and Opsin 2 of the Corn Smut Fungus Ustilago maydis Are Green Light-Driven Proton Pumps. Front Microbiol 2019, 10, 735. [Google Scholar] [CrossRef]
- Ahrendt, S.R.; Medina, E.M.; Chang, C.A.; Stajich, J.E. Exploring the binding properties and structural stability of an opsin in the chytrid Spizellomyces punctatus using comparative and molecular modeling. PeerJ 2017, 5, e3206. [Google Scholar] [CrossRef]
- Kihara, J.; Tanaka, N.; Ueno, M.; Arase, S. Cloning and expression analysis of two opsin-like genes in the phytopathogenic fungus Bipolaris oryzae. FEMS Microbiol Lett 2009, 295, 289–294. [Google Scholar] [CrossRef]
- Fan, Y.; Solomon, P.; Oliver, R.P.; Brown, L.S. Photochemical characterization of a novel fungal rhodopsin from Phaeosphaeria nodorum. Biochim Biophys Acta 2011, 1807, 1457–1466. [Google Scholar] [CrossRef]
- Panzer, S.; Zhang, C.; Konte, T.; Brauer, C.; Diemar, A.; Yogendran, P.; Yu-Strzelczyk, J.; Nagel, G.; Gao, S.; Terpitz, U. Modified Rhodopsins From Aureobasidium pullulans Excel With Very High Proton-Transport Rates. Front Mol Biosci 2021, 8, 750528. [Google Scholar] [CrossRef]
- Adam, A.; Deimel, S.; Pardo-Medina, J.; García-Martínez, J.; Konte, T.; Limón, M.C.; Avalos, J.; Terpitz, U. Protein activity of the Fusarium fujikuroi rhodopsins CarO and OpsA and their relation to fungus–plant interaction. International Journal of Molecular Sciences 2018, 19, 215. [Google Scholar] [CrossRef]
- Avelar, G.M.; Glaser, T.; Leonard, G.; Richards, T.A.; Ulrich, H.; Gomes, S.L. A Cyclic GMP-Dependent K+ Channel in the Blastocladiomycete Fungus Blastocladiella emersonii. Eukaryot Cell 2015, 14, 958–963. [Google Scholar] [CrossRef]
- Broser, M.; Spreen, A.; Konold, P.E.; Schiewer, E.; Adam, S.; Borin, V.; Schapiro, I.; Seifert, R.; Kennis, J.T.M.; Bernal Sierra, Y.A.; et al. NeoR, a near-infrared absorbing rhodopsin. Nat Commun 2020, 11, 5682. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Chen, C.H.; Belden, W.J.; Madeti, C.; Roenneberg, T.; Merrow, M.; Loros, J.J.; Dunlap, J.C. Genetic and molecular characterization of a cryptochrome from the filamentous fungus Neurospora crassa. Eukaryot Cell 2010, 9, 738–750. [Google Scholar] [CrossRef]
- Garcia-Esquivel, M.; Esquivel-Naranjo, E.U.; Hernandez-Onate, M.A.; Ibarra-Laclette, E.; Herrera-Estrella, A. The Trichoderma atroviride cryptochrome/photolyase genes regulate the expression of blr1-independent genes both in red and blue light. Fungal Biol 2016, 120, 500–512. [Google Scholar] [CrossRef]
- Guzman-Moreno, J.; Flores-Martinez, A.; Brieba, L.G.; Herrera-Estrella, A. The Trichoderma reesei Cry1 protein is a member of the cryptochrome/photolyase family with 6–4 photoproduct repair activity. PLoS One 2014, 9, e100625. [Google Scholar] [CrossRef]
- Bayram, O.; Biesemann, C.; Krappmann, S.; Galland, P.; Braus, G.H. More than a repair enzyme: Aspergillus nidulans photolyase-like CryA is a regulator of sexual development. Mol Biol Cell 2008, 19, 3254–3262. [Google Scholar] [CrossRef]
- Castrillo, M.; Garcia-Martinez, J.; Avalos, J. Light-dependent functions of the Fusarium fujikuroi CryD DASH cryptochrome in development and secondary metabolism. Appl Environ Microbiol 2013, 79, 2777–2788. [Google Scholar] [CrossRef]
- Cohrs, K.C.; Schumacher, J. The Two Cryptochrome/Photolyase Family Proteins Fulfill Distinct Roles in DNA Photorepair and Regulation of Conidiation in the Gray Mold Fungus Botrytis cinerea. Appl Environ Microbiol 2017, 83. [Google Scholar] [CrossRef]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutierrez, G.; Miralles-Duran, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal cryptochrome with DNA repair activity reveals an early stage in cryptochrome evolution. Proc Natl Acad Sci U S A 2015, 112, 15130–15135. [Google Scholar] [CrossRef]
- Bonomi, H.R.; Toum, L.; Sycz, G.; Sieira, R.; Toscani, A.M.; Gudesblat, G.E.; Leskow, F.C.; Goldbaum, F.A.; Vojnov, A.A.; Malamud, F. Xanthomonas campestris attenuates virulence by sensing light through a bacteriophytochrome photoreceptor. EMBO Rep 2016, 17, 1565–1577. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Cai, Y.; Wu, L.F.; Song, T. Bacteriophytochrome from Magnetospirillum magneticum affects phototactic behavior in response to light. FEMS Microbiol Lett 2020, 367. [Google Scholar] [CrossRef]
- Xue, P.; El Kurdi, A.; Kohler, A.; Ma, H.; Kaeser, G.; Ali, A.; Fischer, R.; Krauss, N.; Lamparter, T. Evidence for weak interaction between phytochromes Agp1 and Agp2 from Agrobacterium fabrum. FEBS Lett 2019, 593, 926–941. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Curr Biol 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Igbalajobi, O.; Yu, Z.; Fischer, R. Red- and Blue-Light Sensing in the Plant Pathogen Alternaria alternata Depends on Phytochrome and the White-Collar Protein LreA. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Arreguin, J.A.; Cabrera-Ponce, J.L.; Leon-Ramirez, C.G.; Camargo-Escalante, M.O.; Ruiz-Herrera, J. Analysis of the photoreceptors involved in the light-depending basidiocarp formation in Ustilago maydis. Arch Microbiol 2020, 202, 93–103. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Noh, B.; Vierstra, R.D.; Loros, J.; Dunlap, J.C. Genetic and molecular analysis of phytochromes from the filamentous fungus Neurospora crassa. Eukaryot Cell 2005, 4, 2140–2152. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Guo, L.; Luo, Y.; Li, J.; Zhou, Y.; Wang, J.; Huang, X.; Tan, X.; Fu, M.; Yu, B.; et al. Peroxiredoxin PrxA and thioredoxin TrxA mediate the redox signal to the transcription factor NapA in the fungus Aspergillus nidulans. Int J Biol Macromol 2025, 310, 143434. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.; Pocsi, I.; Hamari, Z.; Leiter, E.; Emri, T.; Miskei, M.; Olah, J.; Toth, V.; Hegedus, N.; Prade, R.A.; et al. AtfA bZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans. Mol Genet Genomics 2010, 283, 289–303. [Google Scholar] [CrossRef]
- Bastakis, E.; Gerke, J.; Ozkan, S.; Harting, R.; Lienard, T.; Sasse, C.; Xylakis, E.S.; Aden, M.; Strohdiek, A.; Heinrich, G.; et al. Molecular circuit between Aspergillus nidulans transcription factors MsnA and VelB to coordinate fungal stress and developmental responses. PLoS Genet 2025, 21, e1011578. [Google Scholar] [CrossRef]
- Hagiwara, D.; Asano, Y.; Marui, J.; Furukawa, K.; Kanamaru, K.; Kato, M.; Abe, K.; Kobayashi, T.; Yamashino, T.; Mizuno, T. The SskA and SrrA response regulators are implicated in oxidative stress responses of hyphae and asexual spores in the phosphorelay signaling network of Aspergillus nidulans. Biosci Biotechnol Biochem 2007, 71, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.; Barker, B.M.; Carey, C.C.; Merriman, B.; Werner, E.R.; Lechner, B.E.; Dhingra, S.; Cheng, C.; Xu, W.; Blosser, S.J.; et al. ChIP-seq and in vivo transcriptome analyses of the Aspergillus fumigatus SREBP SrbA reveals a new regulator of the fungal hypoxia response and virulence. PLoS Pathog 2014, 10, e1004487. [Google Scholar] [CrossRef]
- Vaknin, Y.; Hillmann, F.; Iannitti, R.; Ben Baruch, N.; Sandovsky-Losica, H.; Shadkchan, Y.; Romani, L.; Brakhage, A.; Kniemeyer, O.; Osherov, N. Identification and Characterization of a Novel Aspergillus fumigatus Rhomboid Family Putative Protease, RbdA, Involved in Hypoxia Sensing and Virulence. Infect Immun 2016, 84, 1866–1878. [Google Scholar] [CrossRef] [PubMed]
- Tilburn, J.; Sarkar, S.; Widdick, D.; Espeso, E.; Orejas, M.; Mungroo, J.; Penalva, M.; Arst, H., Jr. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid-and alkaline-expressed genes by ambient pH. The EMBO journal 1995, 14, 779–790. [Google Scholar] [CrossRef]
- Veri, A.O.; Robbins, N.; Cowen, L.E. Regulation of the heat shock transcription factor Hsf1 in fungi: implications for temperature-dependent virulence traits. FEMS Yeast Res 2018, 18. [Google Scholar] [CrossRef]
- Zhou, J.; Long, H.; Guo, Y.; Lu, J.; Wang, N.; Liu, H.; Zhou, X.; Cai, M. Glutamate-related nitrogen metabolism regulates cold-adaptive synthesis of red pigment in polar fungus Geomyces sp. WNF-15A. J Biotechnol 2025, 404, 121–131. [Google Scholar] [CrossRef]
- Hagiwara, D.; Kondo, A.; Fujioka, T.; Abe, K. Functional analysis of C2H2 zinc finger transcription factor CrzA involved in calcium signaling in Aspergillus nidulans. Curr Genet 2008, 54, 325–338. [Google Scholar] [CrossRef]
- Cai, Z.; Du, W.; Zeng, Q.; Long, N.; Dai, C.; Lu, L. Cu-sensing transcription factor Mac1 coordinates with the Ctr transporter family to regulate Cu acquisition and virulence in Aspergillus fumigatus. Fungal Genet Biol 2017, 107, 31–43. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Inarrairaegui, A.; Ugalde, U. Copper Resistance in Aspergillus nidulans Relies on the P(I)-Type ATPase CrpA, Regulated by the Transcription Factor AceA. Front Microbiol 2017, 8, 912. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.S.; Sobering, A.K.; Romeo, M.J.; Levin, D.E. Regulation of the yeast Rlm1 transcription factor by the Mpk1 cell wall integrity MAP kinase. Mol Microbiol 2002, 46, 781–789. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Zhang, C.; Chen, L.; Yin, Y.; Chen, Y.; Liu, Z.; Ma, Z. The Mgv1-Rlm1 axis orchestrates SAGA and SWI/SNF complexes at target promoters. Nucleic Acids Res 2025, 53. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.N.A.; Rocha, M.C.; de Castro, P.A.; Silva-Rocha, R.; Silva, R.N.; Freitas, F.Z.; de Assis, L.J.; Bertolini, M.C.; Malavazi, I.; Goldman, G.H. The Aspergillus fumigatus CrzA Transcription Factor Activates Chitin Synthase Gene Expression during the Caspofungin Paradoxical Effect. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Martinez, A.E.; Cano-Dominguez, N.; Aguirre, J. Yap1 homologs mediate more than the redox regulation of the antioxidant response in filamentous fungi. Fungal Biol 2020, 124, 253–262. [Google Scholar] [CrossRef]
- Peng, S.; Hu, L.; Ge, W.; Deng, J.; Yao, L.; Li, H.; Xu, D.; Mo, H. ChIP-Seq Analysis of AtfA Interactions in Aspergillus flavus Reveals Its Involvement in Aflatoxin Metabolism and Virulence Under Oxidative Stress. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Leach, M.D.; Farrer, R.A.; Tan, K.; Miao, Z.; Walker, L.A.; Cuomo, C.A.; Wheeler, R.T.; Brown, A.J.; Wong, K.H.; Cowen, L.E. Hsf1 and Hsp90 orchestrate temperature-dependent global transcriptional remodelling and chromatin architecture in Candida albicans. Nat Commun 2016, 7, 11704. [Google Scholar] [CrossRef] [PubMed]
- Martins-Santana, L.; Paula, R.G.; Silva, A.G.; Lopes, D.C.B.; Silva, R.D.N.; Silva-Rocha, R. CRZ1 regulator and calcium cooperatively modulate holocellulases gene expression in Trichoderma reesei QM6a. Genet Mol Biol 2020, 43, e20190244. [Google Scholar] [CrossRef]
- de Castro, P.A.; Chen, C.; de Almeida, R.S.; Freitas, F.Z.; Bertolini, M.C.; Morais, E.R.; Brown, N.A.; Ramalho, L.N.; Hagiwara, D.; Mitchell, T.K.; et al. ChIP-seq reveals a role for CrzA in the Aspergillus fumigatus high-osmolarity glycerol response (HOG) signalling pathway. Mol Microbiol 2014, 94, 655–674. [Google Scholar] [CrossRef]
- Xu, H.; Fang, T.; Omran, R.P.; Whiteway, M.; Jiang, L. RNA sequencing reveals an additional Crz1-binding motif in promoters of its target genes in the human fungal pathogen Candida albicans. Cell Commun Signal 2020, 18, 1. [Google Scholar] [CrossRef]
- Moreno, M.A.; Ibrahim-Granet, O.; Vicentefranqueira, R.; Amich, J.; Ave, P.; Leal, F.; Latge, J.P.; Calera, J.A. The regulation of zinc homeostasis by the ZafA transcriptional activator is essential for Aspergillus fumigatus virulence. Mol Microbiol 2007, 64, 1182–1197. [Google Scholar] [CrossRef]
- Singhal, V.; Rathore, V. Effects of Zn2+ and Cu2+ on growth, lignin degradation and ligninolytic enzymes in Phanerochaete chrysosporium. World Journal of Microbiology and Biotechnology 2001, 17, 235–240. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Young, E.M.; Jones, T.S.; Densmore, D.; Voigt, C.A. Genetic circuit design automation for yeast. Nat Microbiol 2020, 5, 1349–1360. [Google Scholar] [CrossRef]
- Hannigan, G.D.; Prihoda, D.; Palicka, A.; Soukup, J.; Klempir, O.; Rampula, L.; Durcak, J.; Wurst, M.; Kotowski, J.; Chang, D.; et al. A deep learning genome-mining strategy for biosynthetic gene cluster prediction. Nucleic Acids Res 2019, 47, e110. [Google Scholar] [CrossRef]
- Hanko, E.K.R.; Paiva, A.C.; Jonczyk, M.; Abbott, M.; Minton, N.P.; Malys, N. A genome-wide approach for identification and characterisation of metabolite-inducible systems. Nat Commun 2020, 11, 1213. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, H.; Ang, E.L. A New Biosensor for Stilbenes and a Cannabinoid Enabled by Genome Mining of a Transcriptional Regulator. ACS Synth Biol 2020, 9, 698–705. [Google Scholar] [CrossRef]
- Ba, F.; Ji, X.; Huang, S.; Zhang, Y.; Liu, W.Q.; Liu, Y.; Ling, S.; Li, J. Engineering Escherichia coli to Utilize Erythritol as Sole Carbon Source. Adv Sci (Weinh) 2023, 10, e2207008. [Google Scholar] [CrossRef]
- Illergård, K.; Ardell, D.H.; Elofsson, A. Structure is three to ten times more conserved than sequence—a study of structural response in protein cores. Proteins: Structure, Function, and Bioinformatics 2009, 77, 499–508. [Google Scholar] [CrossRef]
- Holm, L.; Rosenstrom, P. Dali server: conservation mapping in 3D. Nucleic Acids Res 2010, 38, W545–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Skolnick, J. TM-align: a protein structure alignment algorithm based on the TM-score. Nucleic Acids Res 2005, 33, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- van Kempen, M.; Kim, S.S.; Tumescheit, C.; Mirdita, M.; Lee, J.; Gilchrist, C.L.M.; Soding, J.; Steinegger, M. Fast and accurate protein structure search with Foldseek. Nat Biotechnol 2024, 42, 243–246. [Google Scholar] [CrossRef]
- Dahl, R.H.; Zhang, F.; Alonso-Gutierrez, J.; Baidoo, E.; Batth, T.S.; Redding-Johanson, A.M.; Petzold, C.J.; Mukhopadhyay, A.; Lee, T.S.; Adams, P.D.; et al. Engineering dynamic pathway regulation using stress-response promoters. Nat Biotechnol 2013, 31, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Choi, Y.W.; Zhao, H.; Tan, M.H.; Ang, E.L. Discovery and engineering of a 1-butanol biosensor in Saccharomyces cerevisiae. Bioresour Technol 2017, 245, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Kotelnikov, S.; Egbert, M.E.; Ofaim, S.; Stevens, G.C.; Phanse, S.; Saccon, T.; Ignatov, M.; Dutta, S.; Istace, Z.; et al. Ligand interaction landscape of transcription factors and essential enzymes in E. coli. Cell 2025, 188, 1441–1455 e1415. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.; Kim, D.; Kim, B.; Choi, J.; Lee, J. Recent advances in AI-driven protein-ligand interaction predictions. Curr Opin Struct Biol 2025, 92, 103020. [Google Scholar] [CrossRef]
Figure 1.
Examples of filamentous fungus-derived genetically encoded biosensors applied in synthetic biology. Illustration of application of GPCR and photoreceptor were modified from [17,18,19].
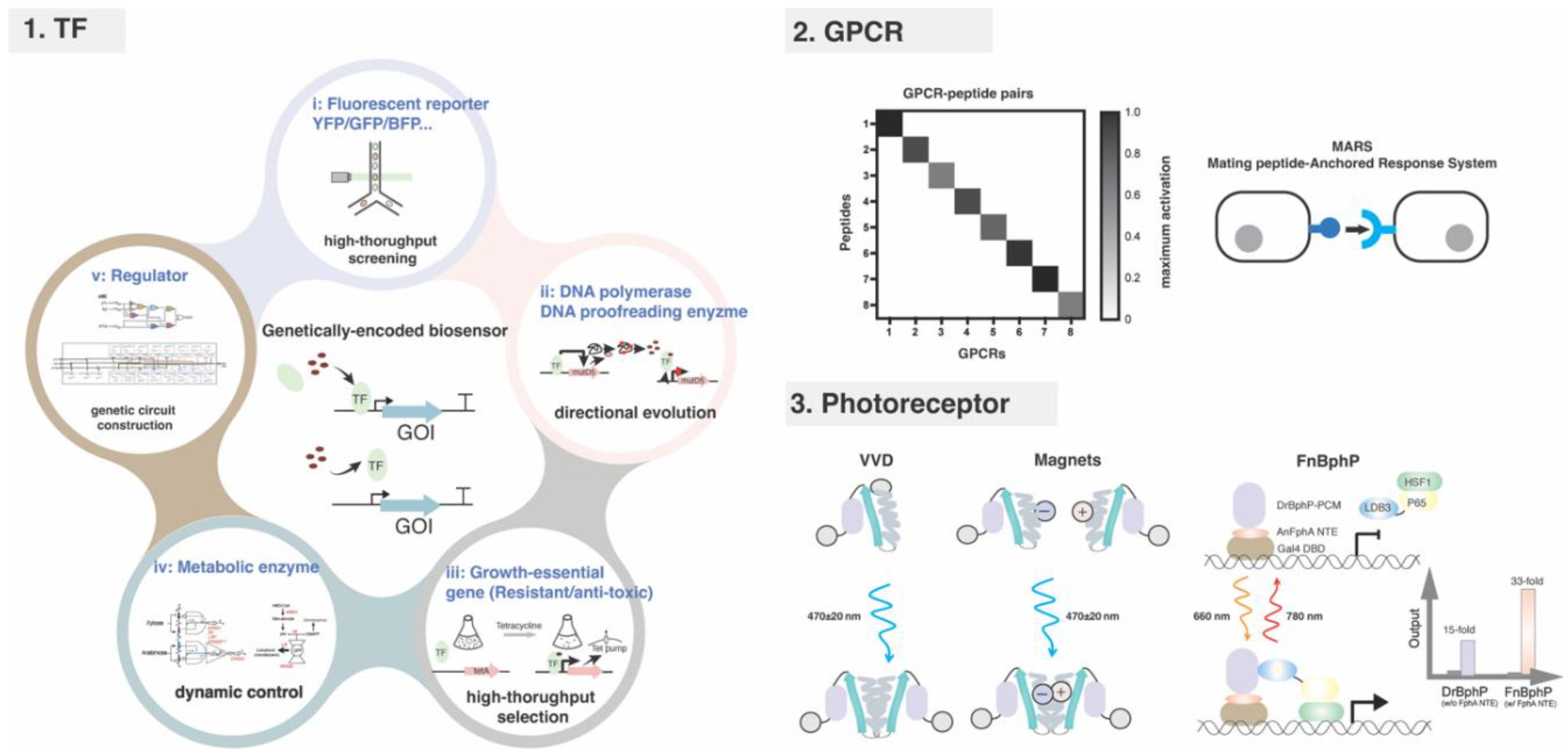
Figure 2.
Limitations of endogenous and bacterial-derived biosensors.
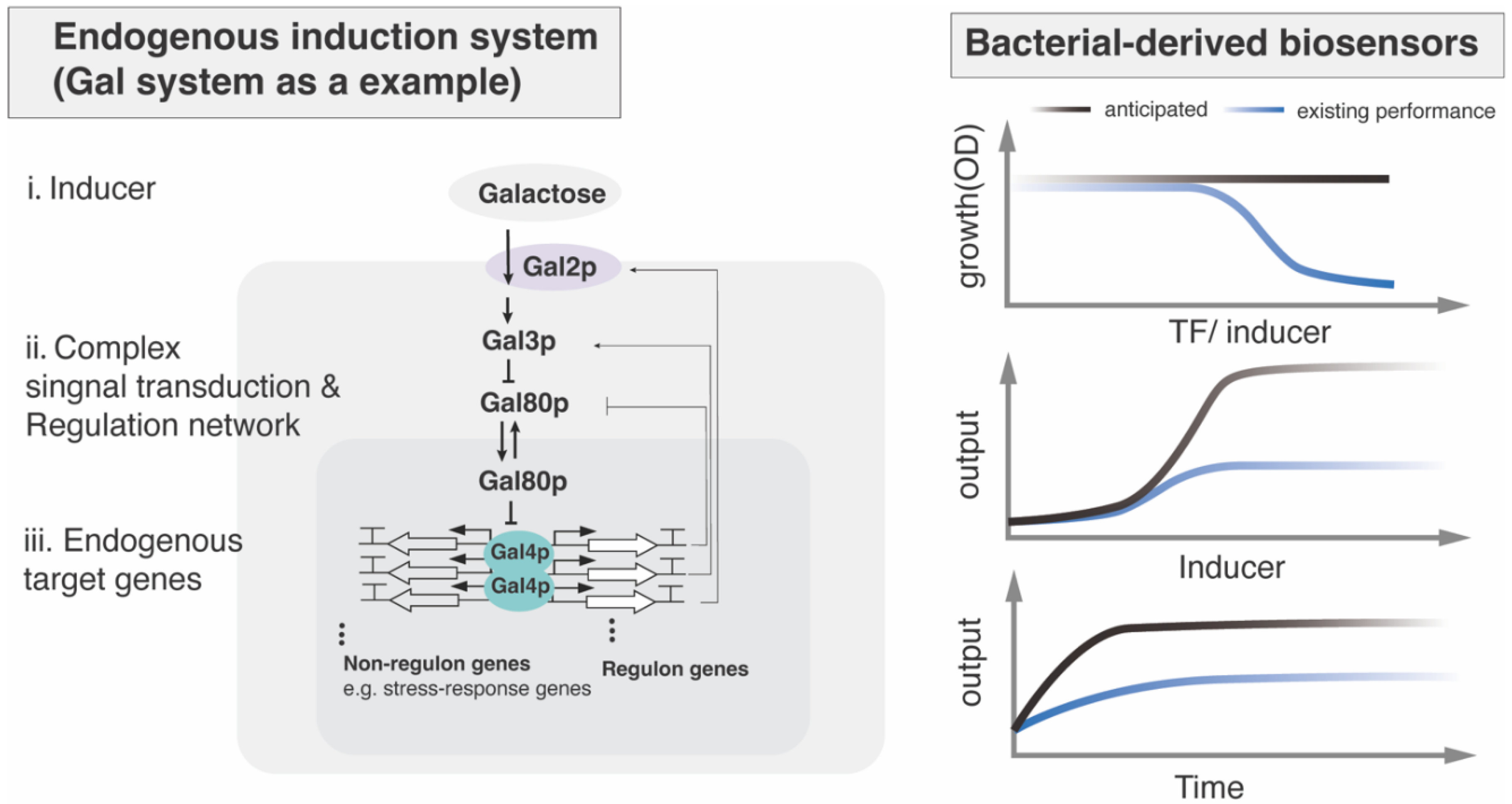
Figure 3.
Small-molecule signaling pathway in filamentous fungi.
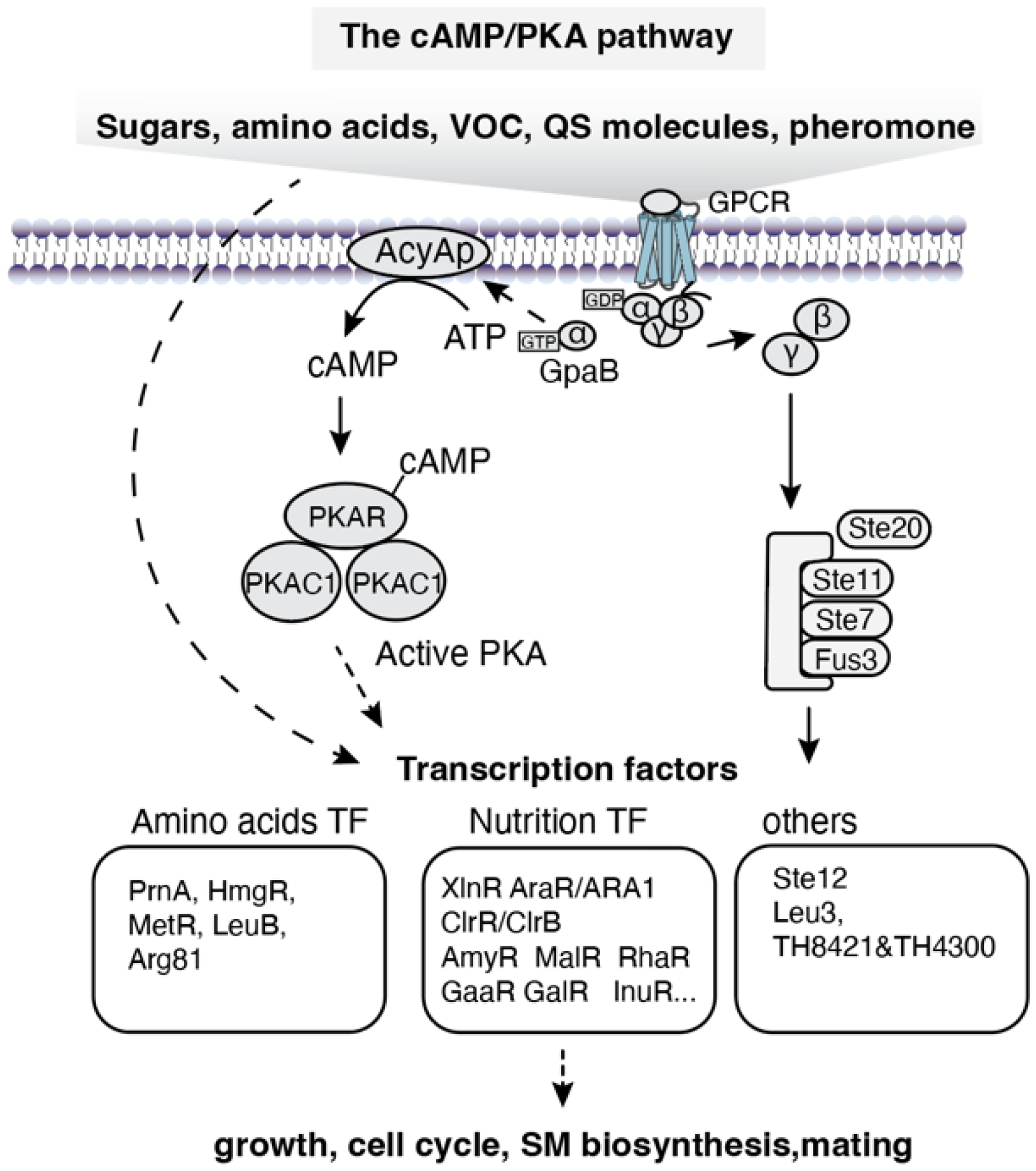
Figure 4.
Signaling pathway of environmental cues in filamentous fungi.
Figure 5.
Schematic illustration of gene cluster-guided biosensor mining and development.
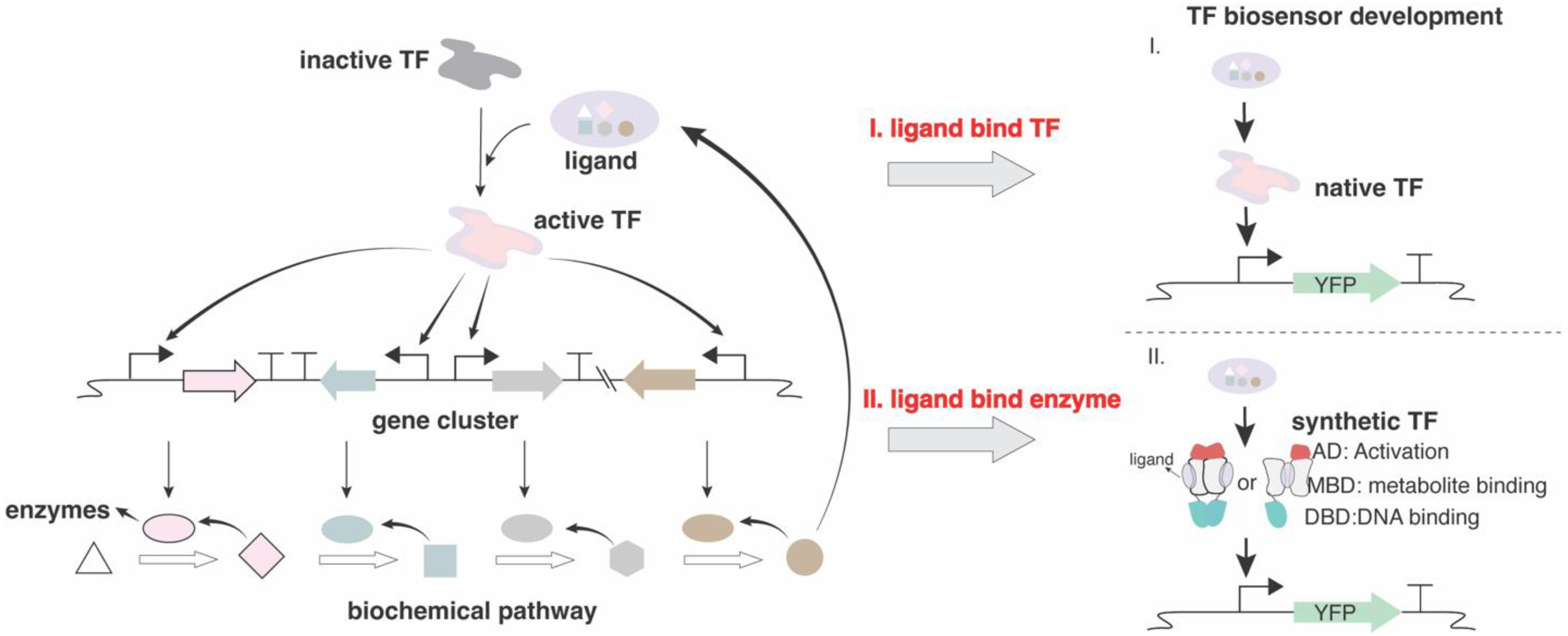

Table 1.
Small molecules responsive TFs summary.
| Inducer | Responsive TF | UAS | Species | Ref |
|---|---|---|---|---|
| Plant biomass degradation monomer | ||||
| xylose | XlnR | GGCTAAA |
Aspergillus spp.; Penicillium spp.. |
[47] |
| arabinose | AraR | N/A* | Aspergillus spp. | [48] |
| maltose | MalR | N/A | A. oryzae | [49] |
| sucrose/inulin | InuR | CGG-X8-CGG | A. niger | [50] |
| rhamnose | RhaR | CGG-X11-CCG | A. nidulans | [51] |
| cellobiose | ClbR | CGG or CCG | A. aculeatus | [52] |
| D-galacturonic acid | GaaR | TCC-X1-CCAAT | A. niger | [53] |
| isomaltose | AmyR | CGG-X8-CGG or CGGAAATTTAA | A. nidulans | [54] |
| L-fucose | FUR1 | CCGACGG | T. reesei | [55] |
| β-mannan | ManR | CAGAAT | A. oryzae | [49] |
| ferulic acid | FarA | CCTCGG | A. niger | [56] |
| d-Fructose | FruR | TGAWWGWTTT | F. prausnitzii | [57] |
| l-acetic acid | Haa1 | SMGGSG | S. cerevisiae | [58] |
| NO3- or NO2- | NirA | CTCCGHGG | Aspergillus spp. | [59] |
| Amino Acids | ||||
| proline | PrnA | CCGG-N-CCGG | A. nidulans | [37] |
| tyrosine | HmgR | N/A | A. fumigatus | [60] |
| valine/leucine/isoleucine | LeuB | CCG-X4-CGG | Aspergillus spp. | [61] |
| arginine | ARCA | CTGCACTTAGAG | Aspergillus nidulans | [62] |
| methionine | MetR | ATGRYRYCAT | Aspergillus nidulans | [63] |
| Pheromone | ||||
| pheromone | Ste12, SteA | TGAAACA |
Aspergillus spp. C.albicans |
[64] |
| Volatile Compounds (VOC) | ||||
| aromatic compounds(o-toluidine, guaiacol) | TH8421/TH4300 | TH8421: CGG-X10-CCG,CGG-X5-CGG; TH4300:CGG-X6-CGG |
T. hirsuta | [32] |
| acetaldehyde | AlcR | half site:TGCGG | A. nidulans | [65] |
| 1-octen-3-ol | N/A | N/A | A. flavus | [66] |
| Quorum Sensing Molecule (QSM) | ||||
| Farnesoic acid | Hot1 | TTAATAATCAAAAACAATTTAATCGT | C. albicans | [67] |
| Oxylipin | NosA | N/A | A. nidulans | |
| Farnesol | Czf1/Efg1(APSES TF) | CZF1: TTWRSCGCCG; Efg1:TGCAT |
Candida spp. | [68] |
| Isoamyl alcohol | Aro80 | CCG-X7-CCG | C. albicans | [69] |
| 1-dodecanol | Sfl1 | AGAA-X-TTCT | C. albicans | |
| QSP1 (QS peptide) | Cqs2 | N/A | C. neoformans | [70] |
| Other | ||||
| Quinic acid | QF-QS | GGRTAARYRYTTATCC | N. crassa | [71] |
| N/A*: Not Available | ||||
Table 2.
Environmental cues responsive GPCR summary.
| GPCR | Signal | Physiological role/ evidence | Species | Ref |
|---|---|---|---|---|
| Nutrition | ||||
| Gpr1, Git3, GPR-4, GprC, GprD | carbon source | sense the carbon source and further affect growth and metabolism |
S. cerevisiae, S. pombe, N. crassa, Aspergillus spp. |
[42,75,76,77] |
| CnGpr4 | methionine | upstream of the cAMP–PKA pathway and regulates methionine-induced mating and contributes to capsule formation | C. neoformans | [78] |
| AfGprK, GPR-7 (NCU09883) | pentose | ΔgprK mutant is restricted on the medium when pentose is the sole carbon source |
A. fumigatus, N. crassa |
[79] |
| GprH | tryptophan, glucose | The absence of GprH results in a reduction in cAMP levels and PKA activity upon adding glucose or tryptophan to starved cells | A. nidulans | [80] |
| Stm1, GPR-5 (NCU00300), GPR-6 (NCU09195),GprR | nitrogen source (e.g., arginine, ornithine) | expression was induced by N starvation signal thorough Gpa2(Gα) |
S. pombe, N. crassa, A. flavus |
[72,73,81] |
| Hormone | ||||
| Ste2, GprA, PRE-1 (NCU00138), Cpr2, Ste3a |
α-factor pheromone | mediate cell cycle arrest and cell fusion with the opposite mating type |
S. cerevisiae, A. nidulans, N. crassa, C. neoformans |
[82,83,84,85,86] |
| Ste3, GprB, PRE-2(NCU05758), Ste3α | a-factor pheromone | mediate cell cycle arrest and cell fusion with the opposite mating type |
S. cerevisiae, A. nidulans, N. crassa, C. neoformans |
[83,87,88,89] |
| Gpr-12 | abscisic acid (ABA) | could be involved in signaling related to abscisic acid | N. crassa | [73,90] |
| Light | ||||
| NOP-1, CarO | green light (534&561 nm) | light controlled sexual development |
N. crassa, F. fujikuroi |
[91,92] |
| ORP-1 (NCU01735) | N/A | N/A | N. crassa | [91,93] |
| Oxidative stress | ||||
| GprH | N/A | loss of gprH rendered the fungus more resistant to H2O2 | A. flavus | [72] |
| pH | ||||
| PalH/Rim21, GprM, GprR | alkalne pH | palH GPCR-arrestin(Rim8) signaling; null mutant of GprM and GprR result in remarkable morphogentically alteration |
A. flavus | [72,94] |
| GprD, GprF, GprG, GprK, GprM | acid pH | null mutant of these GPCRs were more resistatnt to acidic pH | A. flavus | [72] |
| Osmotic stress | ||||
| GprK, GprM, GprR | null mutant of gprK, gprM, and gprR were more sensitive than the wild-type to the hyperosmotic condition | A. flavus | [72] | |
| Temperature | ||||
| GprC, GprD, GprF, GprG, GprO | thermal stress | the expression of these GPCRs were significantly different at three temperatures (20◦C, 28◦C, and 37◦C); The growth defect of null mutant of GprC or GprD was found to be temperature dependent |
A. flavus, A. fumigatus |
[95,96] |
| Inter-communication signal | ||||
| Gpr1, Gpr2, Gpr3, GprC |
ascaroside | trigger nematode-trapping |
A.oligospora, A. flagrans |
[97,98] |
| CaGpr1 | l-lactic acid | Involved in Lactate signalling and regulates fungal β-glucan masking and immune evasion | Candida albicans | [99] |
| GprC, GprD | linoleic acid derivates (e.g., 9-HODE, 13-HODE) | quorum-sensing receptors | Aspergillus spp. | [72,100] |
| GprO, GprP | 13(S)-HpODE | oxylipin sensing | A. flavus | [72] |
| Pth11, MrGpr8 (Class X); GPR-15~39 | hydrophobic surfaces | the CFEM domain is essential for sensing the hydrophobic surfaces and fungal virulence to plant and insect | Magnaporthe spp., Neurospora crassa | [73,101,102,103] |
Table 3.
Photoreceptors in fungi.
| Photoreceptor | Protein domain | Chromophore | Species | Ref |
|---|---|---|---|---|
| LOV | ||||
| WC-1,WC-2 | LOV, PAS, GATA DBD,AD | FMN, FAD | N. crassa | [114] |
| BcWCL1, BcWCL2 | LOV, PAS, GATA DBD,AD | FMN, FAD | B. cinerea | [115] |
| FaWC1, FaWC2 | FaWC1: LOV, PAS, NLS, ZnF, Poly-Q FaWC2: PAS, NLS, ZnF |
FAD(FaWC1) | F. asiaticum | [116] |
| PoWC-1, PoWC-2 | PoWC1: PAS, ZnF, LOV PoWC2: PAS, GATA-ZnF |
FAD(PoWC-1) | P. ostreatus | [117] |
| SfWC-1 | LOV, PAS, GATA-ZnF, Poly-Q, NLS | FAD | S. fimicola | [118] |
| BLR-1, BLR-2 | BLR-1: LOV, PAS, GATA-ZnF,NLS, Poly-Q PoWC2: PAS, GATA-ZnF |
FMN | T. atroviride | [119] |
| LreA, LreB | LreA: LOV, PAS, GATA-ZnF,NLS LreB: PAS, GATA-ZnF |
FAD (LreA) | A. ndiulans | [120] |
| LreA | LOV, PAS, GATA-ZnF, NLS | FAD,FMN | A. alternata | [121] |
| MadA,MadB | MadA: LOV, PAS, GATA-ZnF, MAPK MadB: PAS, GATA-ZnF, WRKY |
FAD (MadA) | P. blakesleeanus | [122] |
| Vivid | N/C-cap, LOV | FAD,FMN |
N. crassa, B cinerea, T reesei |
[123,124,125] |
| BCLOV3, BCLOV4 | BCLOV3: short-LOV BCLOV4: RGS, LOV |
FAD,FMN | B. cinerea | [126] |
| Opsin/Rhodopsin | ||||
| NOP-1 | Seven transmembrane, helix retinal-binding protein, low pump activity | Retinal (11-cis-Retinal) | N. crassa | [127] |
| CarO | Seven transmembrane, Green light-driven proton pump |
all-trans-retinal |
F. fujikuroi | [92] |
| Opsin-1,Opsin-2 | Green Light-Driven Proton Pumps | U. maydis | [128] | |
| Opsin | Seven transmembrane, pump activity |
9- cis-retinal isomer | S. punctatus | [129] |
| BcOPs (BOP1, BOP2) | Seven transmembrane | all-trans-retinal | B. cinerea | [38] |
| Opsin-1,Opsin-2 | B. oryzae | [130] | ||
| PhaeoRD1,PhaeoRD2 | Seven transmembrane; pump activity |
P. nodorum | [131] | |
| ApOps1, ApOps2, ApOps3 | A. pullulans | [132] | ||
| OpsA | no pump activity | F. fujikuroi | [133] | |
| Rhodopsin-guanylyl cyclases (RhoGCs) | ||||
| BeGC1 | Rhodopsin domain, Guanylyl cyclase domain | retinal | B. emersonii | [134] |
| RGC1,RGC2,RGC3(NeoR) | R. globosum | [135] | ||
| Cryptochromes | ||||
| Neurospora CRY | photosensing;FAD binding domain; PHR;CCE | FAD, MTHF | N. crassa | [136] |
| Cry1 | N-terminal DNA photolyase, FAD-binding domain, C-terminal extension domain |
FAD | T. atroviride | [137] |
| Cry1 | FAD | T. reesei | [138] | |
| CryA | The PHR domain, FAD-binding domain, C-terminal extension domain |
FAD | A. nidulans | [139] |
| CryD | FAD binding domain, The photolyase domain |
FAD, MTHF | F. fujikuroi | [140] |
| BcCRY1 ,BcCRY2 | FAD | B. cinerea | [141] | |
| CryA | FAD, MTHF | P. blakesleeanus | [142] | |
| Phytochromes | ||||
| XccBphP | PAS2,GAF, PHY,PAS9 |
bilin,biliverdin, phycoerythrobilin, phytochromobilin |
X. campestris | [143] |
| MmBphP | PAS,GAF,PHY | M. magneticum | [144] | |
| Agp1,Agp2 | Agp1;PAS,GAF,PHY,HisK,ATPase Agp2:PAS,GAF,PHY,HWE-HK |
A. fabrum | [145] | |
| FphA | P2,GAF,PHY,HKD,RRD | A. nidulans | [146] | |
| FphA | A. alternata | [147] | ||
| Phy1 | PAS,GAF,PHY,HKD,RRD | U. maydis | [148] | |
| PHY-1,PHY-2 | PLD,GAF,PHY,HKD,RRD | N. crassa | [149] | |
| BcPHY-1,BcPHY-2,BcPHY-3 | PAS,GAF,PHY,HK,ATP,RRD | B. cinerea | [38] | |
Table 4.
Environmental cues responsive TFs summary.
| Inducer | Responsive TF | UAS | Species | Ref |
|---|---|---|---|---|
| Oxidative stress | ||||
| ROS: NADPH | NapA | (T/TT)ACTAA/TKASTAA | Filamentous fungal species | [165] |
| AtfA | DRTGTTGCAA | A. flavus | [166] | |
| MsnA | GCTGAGTCAGC | A. nidulans | [152] | |
| Low oxygen | SrbA | (A/G)TCA(T/C/G)(C/G)CCAC(T/C) | Aspergillus spp. | [154] |
| pH | ||||
| Alkaline pH | PacC | GCCARG | A. nidulans | [107] |
| Temperature | ||||
| Thermal stress | Hsf1 | TTCnnGAAnnTTC |
C. albicans; Aspergillus spp. |
[167] |
| Cold | Scaffold5.t61 | N/A | Geomyces sp. WNF-15A | [158] |
| metal ion | ||||
| Ca2+ | CrzA | GDGGCKBNB; A[GT][CG]CA[AC][AG]; GGAGGC(G/A)C(T/A)G |
T. reesei; A. fumigatus; C. albicans |
[168,169,170] |
| Zn2+ | ZafA | CAAGGT | A. fumigatus | [171] |
| Cu2+ | AceA | H(T)HNNGCTGD | P. chrysosporium | [172] |
| Mechanical Forces | ||||
| Ca2+ | Crz1 | G[T/G]GGC[T/A]G[T/G]G | Aspergillus spp. | [159] |
| Rlm1 | TGATGCTGTTGATGT,TGCTATTTTTGG | F. graminearum | [163] | |
| Water Availability | ||||
| Hog1 signaling | AtfA | DRTGTTGCAA | A. flavus | [166] |
| Electromagnetic Fields | ||||
| Not any fungi was identified to be able to sense electromagnetic Fields | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



