Submitted:
13 January 2026
Posted:
14 January 2026
You are already at the latest version
Abstract
Phytopathogens threaten natural ecosystems and global food security. Horticultural trade is the main long-distance pathway causing the spread of these organisms and disease outbreaks worldwide. Most inspections for disease symptoms are conducted visually but this is insufficient given the large number of plants and the prevalence of asymptomatic infections. Therefore, there is increasing interest in the use of high-throughput sequencing (HTS) and environmental DNA (eDNA) for plant health surveillance. Many studies have used these technologies to detect phytopathogens, but fewer have done so in horticultural settings. Furthermore, much work has focused on the molecular and bioinformatic approaches for this work, with relatively little attention given to sample collection. This systematic review therefore provides an overview of the available sampling methods and their target plant pathogens, with a particular focus on the utility of these sampling methods in horticultural nurseries. It highlights some striking gaps in the literature and opportunities for further research, for example, the detection of bacterial phytopathogens using eDNA has received little attention despite having considerable potential as a surveillance and/or diagnostic tool.
Keywords:
eDNA
; environmental DNA
; high‐throughput sequencing
; horticulture
; phytopathogen surveillance
; molecular detection
; next generation sequencing
; plant pathogens
1. Introduction
Plant Trade, Biosecurity Risk, and the Need for eDNA Pathogen Surveillance in Horticultural Nurseries
Healthy plants are important to many areas of life, such as agriculture, ecosystem services, and the natural environment in a world where the global population reached 8 billion in 2022 and is expected to reach 9 billion in 2037 [1]. Native and introduced plant pathogens, or phytopathogens, are disease causing organisms which infect plants and cause a variety of symptoms, including reduced growth and plant death (necrosis). The damage caused by these pathogens can be further exacerbated by climate change [2,3]. More specifically, in horticulture plant pathogens threaten the global market that is expected to reach US$ 40.24 billion by 2026 [4]. Within this sector, ornamental plant trade has been growing steadily for many years [5,6]. These numbers highlight the need for maximizing agricultural efficiency and minimizing agricultural losses due to disease to meet the nutritional needs of the growing population.
In addition to native pathogens, foreign pathogens pose a huge threat to natural ecosystems globally [7]. Native plant communities have co-evolved with local pathogenic microorganisms and viruses. These ecosystems usually reach a balanced state in which the pathogens cause a manageable amount of damage to their host plants and wider ecosystems [8]. However, if a pathogen is introduced from another region of the world, it can wreak havoc on these ecosystems as the native plant species may not have evolved sufficient immunity [8].
Movement of live plant material, timber, and seeds facilitates the entry of phytopathogens into new geographical locations [2,9,10,11,12]. Many phytopathogens are spread by ornamental plants. For example, powdery mildews caused by Oidium spp., Erysiphe azaleae, and Podosphaera spiraeae, bacterial leaf spots caused by Pseudomonas spp., a grey mold caused by Botrytis cinerea, root rots caused by Phytophthora spp., and die-back caused by Botryosphaeria parva have all been detected on ornamental plants in northern Italy [13]. It is very difficult to eradicate invasive pathogens once they have established in the environment [8], therefore, reliable, early pathogen detection methods are crucial to prevent outbreaks. Early detection of plant pathogens in horticultural nurseries is also key to reducing their associated economic losses and the quantities of fungicides and pesticides applied.
The import of infected plant material into nurseries is believed to be re-sponsible for ca. 69% of newly introduced phytopathogen outbreaks worldwide [12]. Normally, plants are visually inspected for disease symptoms, which is not practical, and often not effective, for large shipments [8]. Additionally, many plants may be asymptomatic carriers of pathogens [12,14]. For example, species of the bacterial phytopathogen Pectobacterium are known to be spread via the ornamental plant trade, and asymptomatic infection is common [14]. One study found that 62% of asymptomatic horticultural plant samples were carrying pectinolytic bacteria, most of which were from the genus Pectobacterium [14]. Some examples of highly destructive phytopathogens which have been introduced via plant trade include Phytophthora ramorum [8], Dutch elm disease (Ophiostoma spp.) [15], and Ash dieback (Hymenoscyphus fraxineus) [16]. The economic cost of the Ash dieback outbreak in the UK, when accounting for safety felling, replanting, and loss of ecosystem services, is estimated to be £15 billion [16]. These numbers highlight the risk associated with the plant trade.
The horticulture sector produces a wide range of plant products such as food plants (e.g., fruits, vegetables, nuts) and ornamental plants (e.g., flowers, grasses, Christmas trees, outdoor/hardy ornamentals, house plants, seeds, bulbs) [4]. The demand for ornamental plants for commercial real estate construction is considered the main driver of this market [5]. The global export value of live plants and bulbs was US$13 billion in the year 2022 [6]. The nursery and floriculture production market value stood at US$353.26 billion in 2024 and is expected to reach US$569.46 billion by 2029 [5]. The Netherlands alone exported over US$12 billion worth of ornamental plant products (including bulbs, live plants, cut flowers, and ornamental foliage) in 2022 [6].
Reliable, early pathogen detection methods, such as eDNA (environmental DNA) detection, are crucial in horticultural settings to reduce economic losses and to prevent the spread of plant pathogens within the horticulture industry and natural ecosystems. The methods used to detect pathogens depends on the type of pathogen and resources available. This review therefore discusses the life histories of major groups of phytopathogenic organisms with respect to the appropriate sampling matrices to detect them in horticulture (e.g., water samples to detect waterborne organisms). The various eDNA sample types (air, water, growing media, plant tissue) and the various sampling methods and tools which can be used for each type of sample are outlined and compared via an extensive literature review. The sampling methods use bespoke and/or commercially available tools. We particularly focus on how these surveillance and diagnostic methods may be applied in working horticultural nursery settings.
2. Materials and Methods
This review was conducted using key word searches in Scopus and PubMed. The search terms used were as follows: (“Environmental DNA” OR “eDNA” OR “high throughput sequencing” OR “HTS” OR “PacBio” OR “Illumina” OR “next generation sequencing” OR “NGS” OR “metabarcoding”) AND (“phytopathogen*” OR “pathogen” OR “disease” OR “oomyc*” OR “fung*” OR “virus” OR “viral” OR “bacteria*” OR “nematode*”) AND (“nurser*” OR “horticultur*”). A total of 934 unique documents were found in this way, of which 27 were deemed to be relevant, this decision was corroborated independently by two researchers to minimize bias. Relevant studies were those which fulfilled the following criteria: used HTS and/or eDNA based methodology to detect phytopathogens in commercial nurseries.
3. Results & Discussion
3.1. Challenges of Disease Surveillance in Horticulture
The facilities needed to grow the diverse range of horticultural products can vary widely, ranging from tree farms which resemble small forests, to highly specialized potted plant nurseries, to potato fields. Horticultural nurseries typically grow a wide variety of plant taxa making them much more complicated than a monoculture system [17]. Nurseries vary widely in size, from large nurseries which ship plants internationally to small, highly specialized nurseries. The irrigation systems employed are also variable; they may rely on manual hand-watering of plants, overhead sprayers, drip irrigation, microsprinklers, capillary beds, or a combination of these systems [17]. Irrigation water source is also variable, for example, rainwater, well water, and/or mains water can be used. Many nurseries recycle irrigation water, which is important for water conservation but introduces phytosanitary risks if the water is not properly treated [18]. Furthermore, nurseries employ a wide range of phytosanitary practices, for example, a specialized micropropagation nursery will need much more stringent biosecurity practices than a nursery specializing in hardy, outdoor plants. Therefore, the challenges facing these facilities, including the phytopathogens, are also diverse and require varied solutions.
Nurseries differ from other plant growing systems in several ways which render them risky from a biosecurity perspective. The warm, humid, well-irrigated conditions in many nurseries provide ideal conditions for pathogens to thrive. For example, Phytophthora species are extremely well adapted to these environments, their thick-walled resting spores can survive unseen for long periods of time in soil or plant material and their motile zoospores are free-swimming and spread readily in water [7]. The presence of many different types of plants in a small area can make disease detection challenging [12] and facilitate the spread of pathogens from a coevolved, asymptomatic carrier to a plant with no natural immunity [8]. Nurseries may also facilitate the hybridization of closely related phytopathogenic species, which would never otherwise have come into contact [8]. These hybrids are often more aggressive and have wider host ranges than their parent taxa [8]. Hybrid Phytophthora alni has killed large numbers of riparian alder trees across Europe [7]. A survey of ornamental nursery stock in 2015 isolated a hybrid oomycete which contained genetic material from four Phytophthora species and a Phytopythium species [19].
3.2. Potential of High-Throughput Sequencing and Environmental DNA for Plant Pathogen Surveillance
High-throughput sequencing (HTS) can detect nucleic acids of any organism present in a sample and help identify it without any prior knowledge of what may be present [20]. The European and Mediterranean Plant Protection Organization has published guidelines regarding the use of HTS in plant health diagnostics [21], highlighting the real regulatory potential of this technology. There are numerous studies which have used HTS for detecting and diagnosing diseases in plants and this technology has been applied in many sectors, including horticulture (e.g., [22,23,24]), agriculture (e.g., [25,26,27,28]), and forestry (e.g., [29]). These methods can be helpful as they can detect pathogens even when there are no symptoms present. For example, HTS was used to identify fifteen viruses and three viroids associated with fig mosaic disease [30]. This is significant as mixed infections involve synergistic and antagonistic interactions between viruses, and symptoms may be exacerbated depending on the species present [30]. These studies frequently detect species which have not been found in a region before (e.g., [30,31,32]) and often also uncover novel taxa (e.g., [30,31]).
Environmental DNA (eDNA) is defined as the genetic material left by organisms in the environment [33]. eDNA can be extracted from environmental samples such as air, soil, and water, and can be used to assess the presence of species [33]. eDNA-based studies are becoming increasingly routine for analyzing the composition of biological communities and for monitoring of unwanted and/or invasive species. Several recent studies have developed analysis protocols for using eDNA to detect and monitor invasive species such as carp [34], bivalves [35], and mice [36]. A curated genomic database of notifiable animal viruses in Australia has been developed for surveillance purposes [37]. eDNA tools may also be useful for monitoring invasive species in hard to survey regions, such as Antarctica [38]. These examples are non-exhaustive but illustrate the extent to which eDNA-based methods may be useful for the surveillance of invasive species.
The potential utility of eDNA-based methods for diagnosis and surveillance in plant nurseries and other horticultural settings is huge. These methods have many advantages over traditional identification methods. When combined with a fit for purpose sampling strategy, they potentially allow for much more thorough and efficient detection of disease than visual inspections. Widely used molecular methods, such as species-specific real-time PCR testing are limited to detecting a single target species. Other identification methods such as enzyme-linked immunosorbent assay (ELISA) have their own issues. For example, when using ELISA to detect Phytophthora taxa, there is huge potential for a false positive result due to a cross-reaction with Pythium species which are approximately 10 to 100 times more numerous than Phytophthora species in most samples [39]. A 2025 study of Phytophthora diversity in British plant nurseries detected 85 unique Phytophthora species or complexes using eDNA sampling, including several taxa which had not been previously detected in that region [22].
The application of eDNA sequencing methods for surveillance in horticultural settings, such as commercial nurseries, requires robust, repeatable, and practically applicable sampling and testing methods for results to be meaningful. Much attention is given to the molecular methods and bioinformatic pipelines for eDNA applications in general, with good reason as both have been found to impact the outcome of a study [25,29,40]. However, the eDNA sampling method also has an important role in the accuracy of surveillance data [41,42]. In comparison with the laboratory- and computer-based components of eDNA analysis, consistent sampling methods are more difficult to achieve. Dickie et al. (2018) reviewed 75 independent studies using eDNA in terrestrial and freshwater ecosystems and found that only 5% of studies were replicable for various reasons. Only 10% of the studies used clearly defined sampling methods and 92% of papers did not clearly define the total area that plots are taken to represent. Sampling for eDNA studies differs considerably from sampling for other ecological studies. For example, there is usually a time-lag between sampling and obtaining results, the sampling is often difficult to repeat in a consistent manner, and there is also risk of contamination which is potentially highly problematic [42]. It is, therefore, clear that sampling methods require close attention for eDNA to be harnessed to the best extent possible.
There are many factors which must be considered when selecting an eDNA sample type and sampling method, including many which are somewhat unique to horticultural settings. eDNA can be extracted from various sampling matrices: air, water, growing media, and/or plant material. The type of sample taken will impact what organisms are detected—for example, Phytophthora capsici disperses via water, and only rarely by air [43], so a water sample is far more likely to detect P. capsici than an air sample. Logistical considerations such as cost, accessibility, sample stability, and sample storage are also important, particularly if the methods must be applied repeatedly, as would be the case for surveillance purposes. Also, the layout of horticultural facilities is extremely variable; for example, plants may be grown indoors or outdoors, irrigation and drainage systems take many forms, and the plant species being grown will impact what pathogens are present. Consequently, eDNA sampling for surveillance in horticultural settings involves all the standard issues associated with this kind of work, along with several extra issues unique to these settings.
3.3. Optimizing Sampling Matrices for Contrasting Phytopathogenic Groups
The life histories of pathogens have considerable practical consequences for detecting, controlling, and avoiding disease outbreaks [44]. In horticultural settings, it is crucial to understand how different pathogen groups are spread and what areas may act as reservoirs for the pathogens to survive in the absence of susceptible host plants. These factors are important when deciding what kind of sampling is appropriate to detect a given pathogen/group of pathogens using eDNA. Certain groups and sampling matrices are represented more frequently than others in the literature. This is reflected in the results of the systematic review conducted as part of this paper, as plant material is the most used sample matrix. The sampling matrix results for the detection of phytopathogens in horticultural nurseries from the systematic review are summarized in Figure 1.
These results show that sampling of plant material is more developed/common than any other type of sampling matrix for pathogen detection in nurseries, although, this is largely due to phytopathogenic viruses being exclusively detected in plant tissue. Only one study employed air sampling, which was successfully used to detect fungal phytopathogens [45]. Bacterial phytopathogens are very under-represented in the literature with no studies conducted to detect them in nursery settings found here. Studies are distributed across the other phytopathogenic groups as follows; ten studies of viruses, nine of fungi, and nine of oomycetes. These results illustrate striking gaps in the literature regarding the use of HTS and/or eDNA to detect phytopathogens in nurseries. These gaps represent huge opportunities to develop eDNA testing for phytopathogen surveillance. For example, eDNA water sampling in nursery settings has been used to successfully detect oomycetes (e.g., [7,22,46,47]) but has not been applied to other phytopathogenic groups. The methods used in these studies have already been developed and shown to work for one phytopathogen group, so with further research they could be successfully adapted to study others.
3.3.1. Phytopathogen Life Cycle and eDNA Detection
The impact of life cycle of several major phytopathogen groups on their detection in eDNA samples was investigated. In cases where no examples of certain phytopathogens were found in studies with commercial nurseries using eDNA-based methods, the wider literature has been used to predict appropriate sampling methods. A more comprehensive description of oomycete phytopathogens, and the relevant eDNA sample matrices, is given compared to other phytopathogen groups. This is because there are many important oomycete pathogens present in nursery systems and comparatively more research has been done on the detection of oomycete phytopathogens from environmental samples (other than plant tissue). Also, as the oomycetes consist of three distinct categories which are transmitted in different ways, this group clearly illustrates the need for careful selection of sample matrix when detecting phytopathogens.
Oomycetes
The oomycetes are a group of eukaryotic organisms in the Stramenophile clade that superficially resemble filamentous fungi [48] and includes many important phytopathogens from the highly diverse groups Phytophthora, Pythium, and downy mildews [48]. Many oomycete species exist in the soil and often overwinter as resting spores (oospores or chlamydospores). Pythium and Phytophthora species thrive in wet soil conditions which allow their asexual, flagellate zoospores to disperse [49]. Oomycetes spread as waterborne spores (zoospores or resting spores in soil particles) or airborne spores (zoosporangia with zoospores) [50]. Insect vectors rarely play a role in oomycete transmission [51]. Thus, detecting oomycetes using eDNA requires careful consideration when deciding what kind of sample to take. A summary of the main oomycete groups and the optimal sampling matrix for their detection is shown in Table 1 (based on matrices in which they have been detected using eDNA or otherwise).
Phytophthora
There are more than 200 Phytophthora species [54] and they have been detected in water, soil, plant tissue, and air samples (Table 1). Phytophthora species mainly rely on free-swimming zoospores in water for dispersal, allowing them to readily move through water in the soil, and are therefore generally considered as soil-borne pathogens [55]. Parke and Grünwald (2012) developed a systems approach for the management of Phytophthora species in ornamental nurseries. This work identified both water and growing media as key mechanisms of Phytophthora spread in nurseries [17]. Soilborne Phytophthora species thrive in wet soil conditions and infect the roots and root collar region of the plant with motile biflagellate zoospores [56]. Phytophthora cinnamomi [55], Phytophthora sojae, Phytophthora ultimum, and Phytophthora parasitica [48] are all important soil-borne Phytophthora species. Many Phytophthora species, such as Phytophthora ramorum, Phytophthora kernoviae, and Phytophthora palmivora produce caducous sporangia [55] which infect shoots, leaves, bark, and fruit of susceptible host plants [56], and thus, plant material can be tested to detect Phytophthora spp. [57]. These sporangia may be spread by wind and wind-driven rain [55,56]. There are several examples of Phytophthora species detected by aerial sampling, including Phytophthora infestans in experimental potato fields [58] and several Phytophthora species in a woody plant nursery [55]. There are also many examples of Phytophthora species being detected in water. Phytophthora species produce flagellate, motile zoospores, which enables soilborne species, such as P. cinnamomi and P. sojae, to move through soil water [48]. P. cinnamomi DNA can be readily detected in dry soil for up to 90 days [59]. Indeed, several Phytophthora species have been detected in the growing media of ornamental plants from commercial nurseries [57,60]. Airborne Phytophthora species, such as P. infestans [55], are also spread by the movement of water including rain, running water, and splash [56]. Thus, water samples have the potential to detect both airborne and soilborne species, so may detect the widest variety of Phytophthora species if sampling is conducted in a robust manner.
Pythium
Many species of Pythium are important plant pathogens [49]. For example, Pythium ultimum is found in the soil [61] and is highly destructive, causing root rot and damping off on a wide range of hosts (>300 species), including many ornamentals [48,61]. Pythium species have been detected in both water samples (e.g., [46,49,52]) and soil samples (e.g., [50]). Pythium species produce motile zoospores, which can swim through water to host plants, and long-lived resting oospores, which persist in growing media for long periods [62]. Therefore, in nursery settings, Pythium species are primarily spread through irrigation water and the movement of potting mixtures. Pythium species do not produce caducous sporangia and are, therefore, not dispersed by air [62]. Indeed, there does not appear to be any significant examples of airborne Pythium species based on the available literature. Thus, samples of either water or growing media may be suitable for Pythium detection.
Downy Mildews
Most downy mildews have narrow host ranges and depend on their host for growth and reproduction [48]. Important species include Hyaloperonospora arabidopsidis and Plasmopara viticola [48]. Available methods for detecting downy mildews are limited by the fact that they are not culturable, thus, developing new methods for their detection is particularly useful [63]. They can survive in soil as quiescent oospores and initiate infection through roots but are predominantly spread via airborne sporangiospores which form the characteristic downy patches on the lower surface of host leaves [48]. Peronospora, the largest downy mildew genus, has been detected in air samples using microscopy but their molecular analyses failed to detect Peronospora in such air [53]. This study was focused on fungal species so it is possible that using molecular methods aimed at oomycete detection would allow for the detection of downy mildew species. Thus, air sampling may be particularly beneficial when monitoring for downy mildews. However, a recent study found that the soil microbiota was a better predictor of a downy mildew (Plasmopara viticola) disease incidence than leaf microbiota—despite the airborne nature of the pathogen [64]. If these effects are better understood, it may be possible to predict or detect downy mildews using soil samples in the future.
Viruses
Phytopathogenic viruses are small, obligate parasites consisting of a nucleic acid molecule in a protein coat (capsid) and are among the most significant threats to plant health globally [65,66]. As they do not possess their own metabolism, viruses can be compared to resting stages (e.g., spores and cysts) of other pathogenic groups [67]. Thus, viruses can persist in the environment for long periods of time if conditions allow, for example, Poliovirus has been found to persist in groundwater and drinking water for 550 days [67]. Phytopathogenic viruses are present in water [68]. Many have extremely wide host ranges and are readily transmitted between hosts [65]. Some important phytopathogenic viruses and their ornamental plant hosts include Chrysanthemum virus B and Tobacco mosaic virus on chrysanthemum plants, Prunus necrotic ringspot virus on rose plants, and Alfalfa mosaic virus on lavender [69]. Plant viruses are naturally transmitted via three main pathways: pollen, seed, and vector [70] and are commonly transmitted by vegetative propagation [69]. Therefore, any samples testing any of these matrices could theoretically detect viral eDNA. Based on the results of this review, plant material is the most common sample medium used for the detection of viral eDNA. There are many recent examples of virus detection using HTS in plant material available in the literature, for example, in detecting viruses in food crops [27,30,31,32,71,72,73,74,75,76,77,78], viruses infecting wild plants (e.g., Senecio coronatus) [79], and viruses infecting ornamental plants [80,81,82,83].
There is potential to use alternative sample types to plant tissue. Many highly phytopathogenic viruses are transmitted by insect vectors [70]. Tomato spotted wild virus (TSWV) is transmitted by thrips; Tomato yellow leaf curl virus (TYLCV) and African cassava mosaic virus (ACMV) are transmitted by whitefly; and Cucumber mosaic virus (CMV), Cauliflower mosaic virus (CaMV), Plum pox virus (PPV), and Potato virus Y (PVY) are transmitted by aphids [84]. Thus, samples which can detect insect vectors may be useful in plant virus surveillance. Valentin et al. (2018) outlined a protocol for detecting insect infestations using eDNA from crop surfaces [85]. This method has been successfully applied to detect the virus-transmitting insect species Bemisia argentifolii and Tetranychus urticae [86]. Flying insects could also be present in air samples and these could be tested for the presence of phytopathogenic viruses. Viral RNA has been extracted from Prunus pollen to successfully detect four viruses [87]. This work illustrates the potential utility of pollen sampling for plant virus surveillance. Viral RNA extracted from bee and pollen samples has been shown to detect different viruses than viral RNA extracted from leaf and flower samples [88] reflecting the fact that not all plant viruses are transmitted via pollen. This highlights the need for careful consideration of sampling method in accordance with the virus of interest.
In summary, plant material is the optimal substrate for detecting phytopathogenic viruses. This is because, at the time of writing, it is most developed method for detecting these organisms. As viruses require their host to survive and reproduce [66], their presence in environmental samples may not be as consistent or predictable as other phytopathogen groups, making plant tissue a more reliable sample medium for their detection. However, there is still potential to develop surveillance protocols based on air or water sampling. For example, fifteen different phytopathogenic viruses from the Virgaviridae family were detected in water samples (freshwater, wastewater, and sludge) in Mexico [68]. Different sampling methods will detect different types of viruses. Some phytopathogenic viruses are soil-borne [89,90] but this literature review did not find any eDNA/eRNA studies conducted to detect these in growing media in nursery settings. Further research may elucidate how robust and consistent non-living sample matrices are for detecting phytopathogenic viruses.
Fungi
Fungi are responsible for approximately 70% of all plant diseases, and over 19,000 species of phytopathogenic fungi have been identified worldwide [91]. Many important fungal pathogens infect ornamental plants. For example, Botrytis cinerea which infects roses and gerbera flowers [92], Fusarium spp. which infect a wide range of ornamental plant species [93,94], Mycosphaerella mori which has been found on ornamental fig trees [95], and Colletotrichum spp. infect several commercially important ornamental plant species [92,96,97]. Infected plant material can be tested to detect fungal phytopathogens. For example, Cladosporium sp., Phoma herbarum, and Alternaria alternata from pine needles [98], Heterobasidion occidentale from stems and/or ramets with leaves [99], and fungal grapevine trunk disease agents from vine cuttings [100,101].
Many fungal species produce huge numbers of spores for wind dispersal, including the causative agents of Ash dieback (Hymenoscyphus fraxineus) [102], rust diseases such as Chrysanthemum white rust (Puccinia horiana) [103], and powdery mildew diseases such as Erysiphe azaleae on Rhododendron plants [102,104,105,106]. Fungal spores are spread via wind, water, soil, and animal vectors [91]. eDNA from air samples has been used to detect many fungal species including Alternaria spp., Cladosporium spp., and Epicoccum spp. [53] and has been successfully applied in forest nurseries [45]. Fungal DNA from aerial sampling accurately reflects disease prevalence of Ash dieback at regional scales [29]. Therefore, it is reasonable to assume that sampling of air may be appropriate for detecting many fungal pathogens.
Fungal phytopathogens can be detected using eDNA from water samples. Aquatic fungal species eDNA has been detected in water samples (e.g., [107,108,109]) but there is limited information available about the presence of non-aquatic fungal species DNA in water. Certain fungal phytopathogens are spread via splash dispersal, for example, the causal agent of rice blast disease (Magnaporthe oryzae) [92]. Rainwater traps, consisting of a container fitted with a funnel and mesh to prevent debris, have been successfully used in forests to detect several fungal phytopathogens such as Alternaria infectoria, Botrytis caroliniana, Colletotrichum acerbum, Fusarium lunatum, Ramularia armoraciae, and Verticillium albo-atrum [110]. Although rainwater traps may not be practical in a nursery setting, particularly in indoor nurseries, the success of this study illustrates that fungal eDNA is present in water which has washed over infected plants. Thus, water sampling may be useful even for detecting species which are not spread via water.
Many fungi exist in the soil and phytopathogenic fungi are often soilborne, infecting roots and other belowground parts of the plant. For example, Fusarium and Neocosmospora spp. are soilborne fungi which cause disease in woody ornamentals such as Bougainvillea, Hebe, and Hibiscus spp. [96]. Molecular methods have been applied to detect phytopathogenic fungi in soil, such as using real-time PCR to detect Cryphonectria parasitica which causes chestnut blight [111]. Based on our literature search, it seems that little work has been done using phytopathogenic eDNA extracted directly from the growing medium, although, there are some studies which have used these methods to investigate fungal diversity in forest nurseries and have detected pathogenic taxa (e.g., [112,113]). Soil eDNA is useful for assessing fungal diversity and is a useful tool for monitoring environmental health [114]. Fungal eDNA from soil detects more total species than fruitbody inventories, however, IUCN red-listed species are missed more frequently by eDNA approaches. This is largely due to these species not being represented in the DNA reference database used [115]. Thus, eDNA extracted from soil and growing media samples may be used to detect fungal phytopathogens.
Bacteria
Many important phytopathogens are bacteria [116,117,118]. Phytopathogenic bacteria have a wide range of survival strategies and can colonize plant surfaces and internal plant tissue [119]. Based on the systematic literature review conducted for this work, there appears to be a huge gap in the literature regarding the use of eDNA technologies for the detection of bacterial phytopathogenic taxa in horticultural nurseries. No studies which used eDNA methods to detect bacterial phytopathogens in commercial nurseries have been found. This lack of knowledge will stifle the application of these powerful techniques in detecting bacterial plant pathogens. However, it is possible to make certain assumptions or inferences, based on current knowledge of bacterial plant pathogens, to guide the use of eDNA.
Phytopathogenic bacterial DNA can be isolated from infected tissue in adult plants [120,121,122] and seeds [123]. Bacterial phytopathogens are transmitted in several ways. Many bacteria species possess flagella, which allow them to move independently to some degree [124]. The plant growth promoting bacteria, Azospirillum brasilense and Pseudomonas fluorescens, have been shown to migrate up to 80mm in soil towards wheat seedlings in the presence of chemical attractants (chemotaxis) [125]. However, in the absence of these chemical attractants it is unlikely that bacteria can move such a distance [126]. Water and wind flow can aid passive movement of bacteria through and/or over soil [124]. Phytopathogenic bacteria may also be transmitted via animal vectors [118]. Many species of phytopathogenic bacteria are restricted to either the sieve tubes in the phloem or to the vessels in the xylem [127]. These can be transmitted by graft inoculation and are mostly vectored by insects which feed in the xylem (sharpshooters) or the phloem (psyllids, leafhoppers) [127]. Many insect groups can spread bacterial pathogens from plant-to-plant, including several flying insects. Ralstonia solanacearum, which causes bacterial wilt in ornamental species, is spread by water drops and several flying insects including bees, wasps, and flies [118]. It is possible to detect insect vectors using eDNA extracted from air samples. Thus, it may be possible to recover bacterial eDNA using air, soil, or water samples.
It is likely that water samples would provide useful eDNA results for bacteria, particularly in horticultural settings, for various reasons. There is evidence that they work as phytopathogenic bacterial eDNA from the genus Agrobacterium has been detected using water sampling in hydroponic systems [128]. Air sampling also has potential; eDNA of the phytopathogenic bacteria Bacillus altitudinus and Pantoea aurantiacum has been detected in air samples taken from polytunnels [129]. Aerial sampling may yield variable results as it is dependent on weather conditions and/or the flight paths of flying insect vectors. Also, in a horticultural greenhouse setting there may be limited wind and flying insects, making consistent aerial sampling throughout the facility difficult to achieve. Because bacteria have limited mobility in growing media, particularly if the growing medium is dry, the results obtained from a growing media sample may vary unpredictably depending on the heterogeneity of the media.
Watering and drainage regimes are static and predictable within plant nurseries. Therefore, water sampling may provide a more representative and meaningful insight into the presence of bacterial taxa in a nursery system than both air and growing media sampling. Water sampling of growing media leachate may provide a more suitable assessment of the bacterial species present in the growing media. More work is needed to gain insight into how the presence of bacteria in growing media translates into the presence of bacteria/bacterial DNA in water leachate from the growing media. HTS is extremely sensitive so it should be possible to detect even small amounts of bacteria in the leachate. Several phytopathogenic bacteria have been reported to be present in open water and/or irrigation systems, including Pseudomonas syringae complex (e.g., [130,131]), Ralstonia solanacearum complex (e.g., [132,133]), Erwinia carotovora (e.g., [134]), Dickeya sp. (e.g., [135]), Xanthomonas campestris complex (e.g., [136]), X. axonopodis complex (e.g., [137]), and Clavibacter michiganensis subsp. michiganensis (e.g., [138]). Regarding the detection of phytopathogenic bacteria in irrigation systems, there is a lack of clear guidelines about methodology [117].
3.3.2. eDNA Sample Matrices and Methods
There are several eDNA sampling methods available for each sample type (air, water, growing media, and plant material). These technologies are very powerful and can be used in novel ways. For example, honeybee pollen pellet contents have been used to detect several oomycete and fungal phytopathogens [147]. There are many innovative methods available, many of which provide excellent results but may not be practical for surveillance purposes in horticultural settings for various reasons. Such settings require low cost, non-destructive, and representative sampling methods which will not interfere in the running of the business. Logistical considerations associated with each sample medium are summarized in Table 2.
The EPPO has published guidelines about using HTS for plant health diagnostics [20] and the requirements for sampling protocols [21]. Owing to their ability to detect nucleic acids from any organism, HTS tests are prone to contamination [20]. Thus, selecting sampling methods that will minimize the risk of contamination is crucial. Samples should be selected appropriately based on prior knowledge of the distribution of the pest to be detected, and they should be representative of the material under examination [21]. Details such as the date and time of sampling, the equipment used, sampling location, sample description, and the name of the sample collector should be recorded [21]. Sampling for eDNA generally involves sample collection using traditional, widely available sampling tools; for example, collecting water in a sterile bottle. This is followed by processing the sample appropriately for eDNA extraction, for example, filtering the collected water using specialized eDNA filters. The available sampling methods, and the associated advantages and disadvantages are outlined here.
Water Sampling
Water sampling has several advantages over other substrates. Water is used in irrigation and is therefore present in regular volumes in most horticultural settings. Many plant pathogens are spread via water [22,148] giving it high potential for use in surveillance. Also, it is likely that many phytopathogens not spread by water could also be detected in irrigation run-through in nurseries as they may be washed off the plant and/or through the growing medium. Depending on the location from which they are taken, water samples can include a much larger catchment than plant material or growing media samples. For example, if water was sampled from a drain after watering a group of plants, the phytopathogens from all the plants could, in theory, be detected. However, the downside of this is that if a certain phytopathogen is detected in a water sample, it may not be possible to identify which plant—if any—is infected with the pathogen. The eradication of the pest may therefore require the destruction of healthy plants depending on what plants are present in the infected water sample catchment, which may be more expensive for growers. Incentives may be necessary for growers to cooperate with these surveillance measures.
Despite the abundance of water in nurseries, irrigation and drainage systems are so variable that it is extremely difficult to develop consistent and repeatable protocols for generalized sampling [17,149]. Run-off water from nursery plants often percolates directly into the ground, making it difficult to collect. Even if accessible drainage systems are present in the nursery, obtaining a sufficient volume of water may be difficult as most water will be used up by the plants. Some protocols require large volumes of water (e.g., [22]) which may not be realistic for long-term surveillance as this may require overwatering plants. Water samples usually require filtration on site for eDNA preservation purposes. On-site filtration allows for convenient transport and containment of samples which may contain pathogens, thus reducing the risk of spreading these organisms to other nurseries. However, on-site filtration can be time consuming and requires specialized equipment.
To maximize the utility of water sampling for eDNA purposes, clear guidelines must be developed. Water sampling for eDNA extraction generally involves collecting and subsequently filtering the water sample. Most eDNA water sampling tools were developed for use in larger bodies of water (e.g., rivers, lakes, reservoirs, etc.) than those present in nurseries. Therefore, some modification of standard sampling methods will almost always be necessary to make them suitable for use in nursery settings. Water taken from large waterbodies generally contains more diluted pathogen concentrations and lower debris content than nursery irrigation waste. Water taken from a nursery, may contain large amounts of debris such as growing media particles and fallen plant material. This means that protocols which have been developed in environments such as lakes, rivers, and reservoirs may call for a much larger volume of water (e.g., 5L to 10L; Table 2) than is practical and/or necessary for sampling from e.g., a nursery drain. The filtration step for water samples taken from nurseries is expected to be more laborious as filters are likely to become clogged much more rapidly due to the presence of debris, thus, some degree of prefiltering may be required to remove large particles. Considerations such as where within the system and when to take the sample are crucial [117]. The turbidity of the water to be taken is also important [117] as it has been found that turbid water can negatively affect eDNA recovery [150].
The practical method of physically taking the water sample is important to consider. Low-cost, easy, reliable, and replicable methods are required for surveillance. Samples can be collected in sterile containers on site and then subsequently filtered in a laboratory using sterile filter funnels and a vacuum pump [151]. Commercially available kits for collecting water samples for eDNA extraction, such as the Sylphium eDNA Sampling Set (SYL009), are based on manually syringing the water sample through a filter paper. These kits are easy to use and standardize but can take a long time to filter the water, can be difficult to use for large volumes of water, and have more limited choice of appropriate filter types. An alternative method for field filtration of samples involves pushing water through a filter using an adapted, knapsack sprayer. This method, developed by Scibetta et al. (2012) [152] to assess Phytophthora diversity, has been successfully applied in nursery settings [22]. This method allows for much larger volumes of water to be sampled than the commercially available syringe filters. The potential downsides of this method include higher initial costs and the potential for pathogen spread as the equipment will be reused.
The sample volume required is extremely important and there is a huge range in the volumes of water used across eDNA studies (Table 2). In general, sampling larger water volumes results in a higher yield of DNA [153]. Several studies have used as little as 15 mL of water to successfully detect target organisms such as mosquitos [154], frogs [155], and phytopathogenic viruses from the Virgaviridae family [68]. Sampling of larger water volumes, even when taken through larger pore sizes (64µm) than traditional sampling methods (0.45µm), has been shown to improve detection of aquatic plant and mollusc taxa [156]. However, this method is only applicable for taxa which have some proportion of their eDNA contained in large fragments, such as in pieces of tissue, faeces, or globs of mucous [156]. Thus, these methods are not generally appropriate for phytopathogen detection, nevertheless, these results illustrate the importance of sample volumes. Wittweer et al. [150] compared oomycete (the crayfish plague agent Aphanomyces astaci) eDNA recovery from water collected by a simple depth filtration of ~1L of water to a comparatively much more labour-intensive and costly dead-end ultrafiltration of ~100L of water. The results showed that the higher-volume more laborious method was slightly better at detecting oomycete eDNA. However, the simple depth method was deemed most cost-effective, easy-to-use, and fast, making it the superior method overall [150]. The most common volumes of water for detecting phytopathogens, based on the literature, range from 1L to 5L, although replicates are often taken followed by subsequent pooling of the samples. However, these studies have generally, though not exclusively, been conducted in larger water bodies than those present in a plant nursery, so it is possible that a smaller volume would suffice in nursery settings. A summary table of some example studies which have successfully detected relevant organisms in water, the volume of water used, and the filter types used, is shown in Table 3.
Water samples for eDNA extraction are normally filtered and the general consensus is that it is the most effective method for maximizing DNA recovery from samples [157]. Filtration should be conducted as soon as possible after sampling, and ideally within 24 hours [157]. Longmire’s lysis buffer is generally the optimal buffer for storing eDNA samples [153]. However, in water samples with high proportions of PCR inhibitors like tannins and humic acids, CTAB may be a better choice [153]. The type of filter used during the filtration step is important, and different filter types are appropriate for different target organisms [150]. Filters can be composed of several different materials (Table 2) including glass fiber, nylon, cellulose nitrate, polycarbonate, nitrocellulose, polyvinylidene fluoride, and mixed cellulose ester [39,150]. Filter pore size is also crucial to consider—smaller pore sizes will get clogged quicker, and larger pore sizes risk missing smaller particles of target organisms. Pre-filtering samples through mesh with large pores, to remove particulate matter, could help prevent clogging. However, it is possible that DNA may be concentrated around these particles, so this step may impact results. Hong et al. [39] compared the filtration of Phytophthora and Pythium taxa using several filter types with various pore sizes (ranging from 0.45-5.0µm), various thicknesses (ranging from 6-180µm), and composed of various materials. The results showed that polyvinylidene fluoride filters of pore size 5µm and thickness 125µm were the optimal choice both in terms of filtering time and recovery rates [39]. This pore size is consistent with current work detecting oomycetes in water samples for eDNA purposes as the most frequently used pore size is 3µm or 5 µm [49]. However, filtration using larger pore size, while time efficient, could potentially fail to detect smaller organisms like bacteria. In a similar study, Wang et al. [158] assessed various filter pore sizes (ranging from 0.1-0.45µm) for filtering freshwater bacterial communities. They found that a small but significant percentage of bacteria can pass through even the smallest pore size tested (0.1µm) and that an average of 50% of bacteria passed through the larger pore size (0.45µm) and in one case 87% of the bacteria passed through these pores [158]. These studies highlight the need to carefully select an appropriate filter for the target taxa based on the size of the cells or spores which are of interest.
It is possible to preserve eDNA samples without filtration, and it would be highly convenient if on-site filtration was not necessary when taking multiple samples in a busy nursery setting. Water samples may be preserved without filtration either by storing them at -80 °C or at room temperature with added Longmire’s solution at a ratio of 1:3 Longmire’s solution to sample volume [159]. It has been found that wild pig (Sus scrofa) DNA is detectable in water samples preserved in this way for 56 days post-sampling using both preservation methods, however, DNA concentration declined during this period and this decline was starker in the samples stored at room temperature [159]. DNA was not detected in any samples stored at room temperature without Longmire’s solution [159]. This work provides interesting results which indicate that it should be possible to filter samples off-site. However, this work did not attempt to assess the diversity present in the samples and how the microbial communities may change with time, as it was not relevant for their purposes. It is possible that certain taxa may be outcompeted by other opportunistic taxa in ways that do not occur in the environment. Thus, further research is needed to quantify the changes in the microbes present in unfiltered samples over time.
Growing Media Sampling
Growing media sampling has several key advantages for phytopathogen detection, particularly in a nursery context. Normally, a relatively large sample of growing media is taken and homogenized (e.g., in a sealed bag) and a subsample of this mixture is taken for subsequent testing (e.g., [25,50]). Most plants in nurseries are grown in some sort of growth media so it is easy to access and widely available. Furthermore, many important plant pathogens are found in the rhizosphere [25]. It can be difficult to identify the causal agents of root diseases visually in a timely manner as aboveground symptoms often appear at late stages and are not very indicative of the root pathogen [25]. Some phytopathogens, such as P. cinnamomi, have been found to produce resting structures which persist in the roots of asymptomatic non-host plants [163], and growing media sampling may be useful in detecting such pathogens.
Several studies have illustrated the utility of soil sampling for phytopathogen surveillance. eDNA collected from topsoil (0 to 30 cm depth) samples taken near symptomatic, infected potato roots, and sequenced using Oxford Nanopore’s MinION, has been able to successfully detect bacteria, fungi, chromists, and nematodes [25]. Rossmann et al. [50] compared DNA baiting methods to eDNA metabarcoding for the detection of plant pathogenic oomycetes (Phytophthora and Pythium). Soil samples, approximately 1 L in volume, were taken from the root zone of woody ornamental plants. These were further sub-sampled by taking 50 mL, homogenizing it, and then finally taking 250 mg for eDNA extraction [50]. The results illustrated the utility of eDNA methods as the metabarcoding provided sensitive and unbiased detection of all oomycetes detected by traditional baiting methods [50].
There are some disadvantages of growing media sampling. For example, as growing media is heterogeneous it may be difficult to take a large enough sample to be representative of the overall diversity present. If small sample sizes are taken, then many species may be missed due to heterogeneity in the growing medium [50]. In potted plants, soil may be tightly packed and difficult to remove, and it may contain a lot of root material. Taking a large volume of soil from near the roots of a potted plant is potentially very damaging to the health and saleability of the plant. Foliar pathogens may also be missed. Furthermore, growing media contains diverse microbial communities and, therefore, the data obtained may be difficult to analyze due to a high degree of non-pathogen DNA sequence ‘noise’ [25]. Microbial cell density and respective genome size (for example the bacterial genome is much smaller than the fungal genome) could also introduce a bias towards the detection of certain pathogens [25]. Soils also have high humic acid content, which can act as a PCR inhibitor and negatively impact DNA yields [153].
Air Sampling
Aerial dispersal is a common and well-documented mechanism by which pathogens spread [164]. eDNA extracted from air samples may be useful in detecting any pathogens which produce airborne spores, including many fungal and some oomycete phytopathogens. Air sampling has already been applied in several studies at different spatial and temporal scales. It has been shown that permanent aerobiological networks, which are widely used for monitoring pollen counts, can not only detect forest pathogenic fungi, but also provide insights into their abundance at a regional scale [29]. Airborne sporangia of Phytophthora infestans have, for example, been detected using a Burkard recorder spore-trap [58].
Detecting eDNA in air samples requires consideration of several factors including the sampler to be used, the placement location and height of the sampler, the volume of air to be collected, and the period over which the air should be collected. Air sampling can be achieved by different methods: filtration, cyclone, impingement, or impaction technologies. Filtration methods pass air through filter membranes using pumps [165]. Cyclone samplers use centrifugal forces to capture bioaerosols on the surface of the sampler [165]. Impingement sampling passes air through nozzles and collects bioaerosols in a liquid collection medium as the air passes into the liquid. Impaction sampling involves collecting air samples through nozzles and subsequently impacting them onto surfaces such as gelatine or agar plates [165]. Filtration based methods have been reported to recover higher yields of DNA than impaction- or impingement-based arial sampling methods [166]. Studies of DNA extracted from air tend to use either filtration-based technology (e.g., [165,167]) or cyclone-based technology (e.g., [168,169]). Commercially available air samplers include the filtration based MD8 air sampler (Sartorius) [170] and the cyclonic Coriolis air sampler (Bertin Technologies) [171]. Airborne fungal diversity has also been successfully assessed using eDNA extracted from volumetric samplers mounted with sticky tape [172,173].
Plant Material Sampling
Sampling and testing of plant material is the most common technique for detecting plant pathogens. These samples are those taken from any part of the plant, including leaf, flower, root, and stem. Some advantages of sampling plant tissue include ease of collection, a higher chance of detecting the pathogen if it is present, and lower cost. Plant material samples for eDNA extraction should ideally be frozen immediately using liquid nitrogen and stored at -70 °C, or alternatively dried in a sealed container with a 10-fold excess of silica gel [139]. Many studies have used HTS of plant material to identify pathogens with good results. Some recent studies have found viruses and viroids associated with fig mosaic disease in Iraq [30], tobacco ringspot virus and tomato ringspot virus in pawpaw trees in the USA [78], and viruses associated with grapevine in Mexico and India [31,32]. Samples can be targeted at symptomatic areas of the plant, so the causal agent is unlikely to be missed. A major disadvantage of sampling plant tissue for surveillance purposes is the relatively small catchment area which will only elucidate the pathogens present in the portion of the plant taken for analysis. Time and labor costs required to select infected material should also be considered. eDNA from asymptomatic leaves will not detect pathogens which are present on nearby leaves or other plants [78]. This is in stark contrast to air or water sampling, which may detect a wide range of pathogens present in the environment, which have been captured in the air or water. Many pathogens are present in the soil and infect plant roots [25], and these may be unlikely to be present in the aboveground parts of plants.
4. Conclusions
This systematic literature review has summarized the available literature and discussed the practical implications of different sampling methods for detecting phytopathogenic organisms (viruses, oomycetes, fungi, bacteria, and nematodes) using eDNA metabarcoding in horticultural settings. Further research investigating the optimal eDNA sample types to detect specific groups of phytopathogenic organisms will be crucial to allow these technologies to reach their potential. In some cases, the methods discussed have not been applied specifically to phytopathogens but rather to other taxa. The existing literature about eDNA from water samples, for example, is predominantly focused on invertebrate species like fish and amphibians [174]. Research on bacterial eDNA, and specifically phytopathogenic bacterial eDNA, detection is much less developed than for fungi, oomycetes, and viruses. This represents a striking gap in the literature which should be addressed before using eDNA based methods routinely for phytopathogen surveillance purposes. Emerging eDNA sampling technologies used in other fields (e.g., human health) may be applied to horticulture.
It is our opinion that, where appropriate, water testing should be optimized and employed for detecting many phytopathogens in horticultural nurseries. This is because run-off irrigation water has a much larger catchment than other sampling methods, as it could, in theory, provide insight into the pathogen risk status in the nursery. Metabarcoding of water samples is particularly suitable for high-risk pathogens of the genus Phytophthora as several studies have illustrated its utility already and water has been identified as being a high risk for its spread in ornamental nurseries [17]. However, the downside to this is that if a sample is positive for a certain phytopathogen, it may be very difficult to locate the infected plant(s) in the nursery. The development of a trace-back system (Figure 2), involving testing a series of key points in the drainage system may allow for the infected plant to be triangulated to a certain area of the nursery, and the appropriate phytosanitary measures to be applied. In this system, if the final drain is clear of certain phytopathogens, the entire nursery could be deemed at a low risk from these taxa. This, however, relies on a significant change in the physical organization of nurseries, in the types of samples collected by inspection services, and in the types of diagnostics routinely applied to plant pathogen surveys.
Further research is needed to link disease presence to phytopathogen eDNA presence in the environment. DNA can persist extracellularly in the environment for extended periods depending on the conditions, making it difficult to distinguish between dead and viable organisms [175]. RNA deteriorates rapidly after cell death, so can provide a more accurate representation of whether living pathogens are present [175]. Many phytopathogens may exist in the environment at low levels so, from a phytopathological perspective, linking the amount of eDNA present in the environment to disease presence is also crucial. If a regulated pathogen is detected in run-off water in a nursery, for example, it is essential that the diagnosis is robust and that a high level of certainty can be given that the pathogen is present. Failure to make these connections could result in huge losses for nursery owners if they were asked to destroy healthy plants. The legislation regarding regulated phytopathogens may need to be altered to allow for negligible levels of these pathogens to be present in the environment, but more work is certainly required to define these “negligible” levels. Testing run-off irrigation water from deliberately inoculated plants in experimental settings may bridge this knowledge gap, providing proof for what has been observed in numerous studies—that eDNA from infectious phytopathogens is present in the environment. It would also allow for comparison of eDNA detection to traditional visual inspection. Robust, repeatable methods are required for eDNA phytopathogen surveillance to become routine. This requires detailed protocols at all stages of testing; sampling, DNA extraction, and subsequent bioinformatic analyses. Work is needed to validate these methods so that the use of eDNA to detect phytopathogens can be accredited for widespread use.
Author Contributions
Conceptualization, K.D. and T.R.H.; methodology, K.D. and T.R.H; formal analysis, K.D.; investigation, K.D.; resources, T.R.H.; data curation, K.D. and C.M.; writing—original draft preparation, K.D.; writing—review and editing, T.R.H., R.OH, C.M., A.R., M.T.; visualization, K.D.; supervision, T.R.H. and R.OH.; project administration, T.R.H.; funding acquisition, T.R.H. and R.OH. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Agriculture, Food and the Marine, Ireland, grant number 2023RP998.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| HTS | High-throughput sequencing |
| eDNA | Environmental DNA |
| EPPO | European and Mediterranean Plant Protection Organisation |
| ELISA | Enzyme-linked immunosorbent assay |
References
- United Nations; Department of Economic and Social Affairs. World Population Prospects 2024: Summary of Results; Stylus Publishing, LLC, 2024. [Google Scholar]
- Morton, L.; Green, S.; MacKay, J. Pathogen impacts and implications for species diversification in UK forestry. Forestry: An International Journal of Forest Research 2025, cpaf019. [Google Scholar] [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nature Reviews Microbiology 2023, 21, 640–656. [Google Scholar] [CrossRef]
- Global Market Estimates. Horticulture Market Report. Available online: https://www.globalmarketestimates.com/market-report/horticulture-market-3414 (accessed on 8 August 2025).
- The Business Research Company, Nursery and Floriculture Global Market Report. Available online: https://www.thebusinessresearchcompany.com/report/nursery-and-floriculture-production-global-market-report (accessed on 24 September 2025).
- Hinsley, A.; Hughes, A.C.; van Valkenburg, J.; Stark, T.; van Delft, J.; Sutherland, W.; Petrovan, S.O. Understanding the environmental and social risks from the international trade in ornamental plants. BioScience 2025, 75, 222–239. [Google Scholar] [CrossRef]
- Green, S.; Cooke, D.E.; Dunn, M.; Barwell, L.; Purse, B.; Chapman, D.S.; Valatin, G.; Schlenzig, A.; Barbrook, J.; Pettitt, T. PHYTO-THREATS: Addressing threats to UK forests and woodlands from Phytophthora; identifying risks of spread in trade and methods for mitigation. Forests 2021, 12, 1617. [Google Scholar] [CrossRef]
- Brasier, C. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathology 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Rodoni, B. The role of plant biosecurity in preventing and controlling emerging plant virus disease epidemics. Virus research 2009, 141, 150–157. [Google Scholar] [CrossRef]
- Colunga-Garcia, M.; Haack, R.; Magarey, R.; Borchert, D. Understanding trade pathways to target biosecurity surveillance. NeoBiota 2013, 18, 103–118. [Google Scholar] [CrossRef]
- Pollard, C.R.; Marzano, M. On a handshake: business-to-business trust in the biosecurity behaviours of the UK live plant trade. Biological Invasions 2023, 25, 2531–2547. [Google Scholar] [CrossRef]
- Rani, A.; Donovan, N.; Mantri, N. The future of plant pathogen diagnostics in a nursery production system. Biosensors and Bioelectronics 2019, 145, 111631. [Google Scholar] [CrossRef]
- Gullino, M.; Bertetti, D.; Garibaldi, A. Fungal and bacterial diseases on ornamental trees, shrubs, hedges and climbing plants detected in the last 20 years in northern Italy. In Proceedings of the IV International Symposium on Woody Ornamentals of the Temperate Zone 1331, 2021; pp. 311–318. [Google Scholar]
- Smoktunowicz, M.; Jonca, J.; Stachowska, A.; May, M.; Waleron, M.M.; Waleron, M.; Waleron, K. The international trade of ware vegetables and ornamental plants—An underestimated risk of accelerated spreading of phytopathogenic bacteria in the era of globalisation and ongoing climatic changes. Pathogens 2022, 11, 728. [Google Scholar] [CrossRef]
- Valatin, G.; Price, C.; Green, S. Reducing disease risks to British forests: An exploration of costs and benefits of nursery best practices. Forestry: An International Journal of Forest Research 2022, 95, 477–491. [Google Scholar] [CrossRef]
- Hill, L.; Jones, G.; Atkinson, N.; Hector, A.; Hemery, G.; Brown, N. The £15 billion cost of ash dieback in Britain. Current Biology 2019, 29, R315–R316. [Google Scholar] [CrossRef]
- Parke, J.L.; Grünwald, N.J. A systems approach for management of pests and pathogens of nursery crops. Plant Disease 2012, 96, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Stewart-Wade, S.M. Plant pathogens in recycled irrigation water in commercial plant nurseries and greenhouses: their detection and management. Irrigation Science 2011, 29, 267–297. [Google Scholar] [CrossRef]
- Benavent-Celma, C.; McLaggan, D.; van West, P.; Woodward, S. Evidence of a Natural Hybrid Oomycete Isolated from Ornamental Nursery Stock. Journal of Fungi 2023, 9, 627. [Google Scholar] [CrossRef]
- EPPO Standard on Diagnostics. PM 7/151 (1) Considerations for the use of high throughput sequencing in plant health diagnostics. EPPO Bulletin 2022, 52, 619–642. [Google Scholar] [CrossRef]
- EPPO Standard on Diagnostics. PM 7/98 (5). Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity; EPPO Bull. 2021; 51, pp. 468–498.
- Green, S.; Cooke, D.E.; Barwell, L.; Purse, B.V.; Cock, P.; Frederickson-Matika, D.; Randall, E.; Keillor, B.; Pritchard, L.; Thorpe, P. The prevalence of Phytophthora in British plant nurseries; high-risk hosts and substrates and opportunities to implement best practice. Plant Pathology 2025, 74, 696–717. [Google Scholar] [CrossRef]
- Antonelli, C.; Biscontri, M.; Tabet, D.; Vettraino, A.M. The never-ending presence of Phytophthora species in Italian nurseries. Pathogens 2022, 12, 15. [Google Scholar] [CrossRef]
- Bačová, A.; Cooke, D.E.; Milenković, I.; Májek, T.; Nagy, Z.Á.; Corcobado, T.; Randall, E.; Keillor, B.; Cock, P.J.; Jung, M.H. Hidden Phytophthora diversity unveiled in tree nurseries of the Czech Republic with traditional and metabarcoding techniques. European Journal of Plant Pathology 2024, 170, 131–156. [Google Scholar] [CrossRef]
- Braley, L.E.; Jewell, J.B.; Figueroa, J.; Humann, J.L.; Main, D.; Mora-Romero, G.A.; Moroz, N.; Woodhall, J.W.; White, R.A., III; Tanaka, K. Nanopore sequencing with GraphMap for comprehensive pathogen detection in potato field soil. Plant Disease 2023, 107, 2288–2295. [Google Scholar] [CrossRef]
- Temple, C.; Blouin, A.G.; Tindale, S.; Steyer, S.; Marechal, K.; Massart, S. High Throughput Sequencing technologies complemented by grower’s perception highlight the impact of tomato virome in diversified vegetable farms 2023.2001. 2012.523758. bioRxiv 2023. [Google Scholar]
- Villamor, D.; Keller, K.; Martin, R.; Tzanetakis, I. Comparison of high throughput sequencing to standard protocols for virus detection in berry crops. Plant Disease 2022, 106, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Frey, J.E.; Frey, B.; Frei, D.; Blaser, S.; Gueuning, M.; Bühlmann, A. Next generation biosecurity: Towards genome based identification to prevent spread of agronomic pests and pathogens using nanopore sequencing. PLoS One 2022, 17, e0270897. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, J.; Husson, C.; Chancerel, E.; Fabreguettes, O.; Chandelier, A.; Fourrier-Jeandel, C.; Dupuy, N.; Dutech, C.; Ioos, R.; Robin, C.; et al. Combining permanent aerobiological networks and molecular analyses for large-scale surveillance of forest fungal pathogens: A proof-of-concept. Plant Pathology 2021, 70, 181–194. [Google Scholar] [CrossRef]
- Al-Kaeath, N.; Zagier, S.; Alisawi, O.; Fadhal, F.A.; Mahfoudhi, N. High-Throughput Sequencing Identified Multiple Fig Viruses and Viroids Associated with Fig Mosaic Disease in Iraq. Plant Pathol J 2024, 40, 486–497. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Stevens, K.; Aguilar-Molina, V.H.; Fernández-Cortés, J.M.; Chabacano León, V.M.; De Donato, M.; Sharma, A.; Erickson, T.M.; Al Rwahnih, M. High-Throughput Sequencing of Grapevine in Mexico Reveals a High Incidence of Viruses including a New Member of the Genus Enamovirus. Viruses 2023, 15, 1561. [Google Scholar] [CrossRef]
- Kishan, G.; Sharma, S.; Holkar, S.; Gupta, N.; Khan, Z.; Singh, S.; Baranwal, V. Diverse spectra of virus infection identified through high throughput sequencing in nursery plants of two Indian grapevine cultivars. 2023. [Google Scholar] [CrossRef]
- IUCN. Issues Brief: Environmental DNA. Available online: https://iucn.org/resources/issues-brief/environmental-dna (accessed on 24 September 2025).
- Kageyama, S.A.; Hoogland, M.R.; Tajjioui, T.; Schreier, T.M.; Erickson, R.A.; Merkes, C.M. Validation of a Portable eDNA Detection Kit for Invasive Carps. Fishes 2022, 7, 363. [Google Scholar] [CrossRef]
- Egeter, B.; Veríssimo, J.; Lopes-Lima, M.; Chaves, C.; Pinto, J.; Riccardi, N.; Beja, P.; Fonseca, N.A. Speeding up the detection of invasive bivalve species using environmental DNA: A Nanopore and Illumina sequencing comparison. Mol Ecol Resour 2022, 22, 2232–2247. [Google Scholar] [CrossRef]
- Piaggio, A.J.; Gierus, L.; Taylor, D.R.; Holmes, N.D.; Will, D.J.; Gemmell, N.J.; Thomas, P.Q. Building an eDNA surveillance toolkit for invasive rodents on islands: can we detect wild-type and gene drive Mus musculus? BMC Biol 2024, 22, 261. [Google Scholar] [CrossRef]
- Batovska, J.; Brohier, N.D.; Mee, P.T.; Constable, F.E.; Rodoni, B.C.; Lynch, S.E. The Australian Biosecurity Genomic Database: a new resource for high-throughput sequencing analysis based on the National Notifiable Disease List of Terrestrial Animals. Database (Oxford) 2024, 2024. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.J.; Shaw, J.D.; Suter, L.; Atalah, J.; Bergstrom, D.M.; Biersma, E.; Convey, P.; Greve, M.; Holland, O.; Houghton, M.J. An expert-driven framework for applying eDNA tools to improve biosecurity in the Antarctic. Management of Biological Invasions 2023, 14, 379–402. [Google Scholar] [CrossRef]
- Hong, C.; Richardson, P.A.; Kong, P. Comparison of membrane filters as a tool for isolating pythiaceous species from irrigation water. Phytopathology 2002, 92, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.A.; Lelwala, R.V.; Elliott, C.E.; Windell, C.; Fiorito, S.; Dinsdale, A.; Whattam, M.; Pattemore, J.; Barrero, R.A. Side-by-Side Comparison of Post-Entry Quarantine and High Throughput Sequencing Methods for Virus and Viroid Diagnosis. Biology (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Hanks, E.M.; Hooten, M.B.; Baker, F.A. Reconciling multiple data sources to improve accuracy of large-scale prediction of forest disease incidence. Ecol Appl 2011, 21, 1173–1188. [Google Scholar] [CrossRef]
- Dickie, I.A.; Boyer, S.; Buckley, H.L.; Duncan, R.P.; Gardner, P.P.; Hogg, I.D.; Holdaway, R.J.; Lear, G.; Makiola, A.; Morales, S.E.; et al. Towards robust and repeatable sampling methods in eDNA-based studies. Mol Ecol Resour 2018. [Google Scholar] [CrossRef]
- Granke, L.; Windstam, S.; Hoch, H.; Smart, C.; Hausbeck, M. Dispersal and movement mechanisms of Phytophthora capsici sporangia. Phytopathology 2009, 99, 1258–1264. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B.M. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. Isme j 2008, 2, 321–334. [Google Scholar] [CrossRef]
- Larsson, R.; Menkis, A.; Olson, Å. Temporal dynamics of airborne fungi in Swedish forest nurseries. Applied and Environmental Microbiology 2025, 91, e01306-01324. [Google Scholar] [CrossRef]
- Redekar, N.R.; Eberhart, J.L.; Parke, J.L. Diversity of Phytophthora, Pythium, and Phytopythium species in recycled irrigation water in a container nursery. Phytobiomes Journal 2019, 3, 31–45. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Fungi and oomycetes in the irrigation water of forest nurseries. Forests 2020, 11, 459. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol Plant Pathol 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Pettitt, T.; Wakeham, A.; Wainwright, M.; White, J. Comparison of serological, culture, and bait methods for detection of Pythium and Phytophthora zoospores in water. Plant Pathology 2002, 51, 720–727. [Google Scholar] [CrossRef]
- Rossmann, S.; Lysøe, E.; Skogen, M.; Talgø, V.; Brurberg, M.B. DNA metabarcoding reveals broad presence of plant pathogenic oomycetes in soil from internationally traded plants. Frontiers in Microbiology 2021, 12, 637068. [Google Scholar] [CrossRef] [PubMed]
- Judelson, H.S.; Blanco, F.A. The spores of Phytophthora: weapons of the plant destroyer. Nat Rev Microbiol 2005, 3, 47–58. [Google Scholar] [CrossRef]
- Presser, J.W.; Goss, E.M. Environmental sampling reveals that Pythium insidiosum is ubiquitous and genetically diverse in North Central Florida. Med Mycol 2015, 53, 674–683. [Google Scholar] [CrossRef]
- Banchi, E.; Ametrano, C.G.; Stanković, D.; Verardo, P.; Moretti, O.; Gabrielli, F.; Lazzarin, S.; Borney, M.F.; Tassan, F.; Tretiach, M. DNA metabarcoding uncovers fungal diversity of mixed airborne samples in Italy. PloS one 2018, 13, e0194489. [Google Scholar] [CrossRef]
- Abad, Z.G.; Burgess, T.I.; Bourret, T.; Bensch, K.; Cacciola, S.O.; Scanu, B.; Mathew, R.; Kasiborski, B.; Srivastava, S.; Kageyama, K.; et al. Phytophthora: taxonomic and phylogenetic revision of the genus. Stud Mycol 2023, 106, 259–348. [Google Scholar] [CrossRef]
- Migliorini, D.; Ghelardini, L.; Luchi, N.; Capretti, P.; Onorari, M.; Santini, A. Temporal patterns of airborne Phytophthora spp. in a woody plant nursery area detected using real-time PCR. Aerobiologia 2019, 35, 201–214. [Google Scholar] [CrossRef]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Abdelfattah, A.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Cooke, D.E.; Schena, L. Metabarcoding analysis of Phytophthora diversity using genus-specific primers and 454 pyrosequencing. Phytopathology 2016, 106, 305–313. [Google Scholar] [CrossRef]
- Meno, L.; Abuley, I.K.; Escuredo, O.; Seijo, M.C. Factors influencing the airborne sporangia concentration of Phytophthora infestans and its relationship with potato disease severity. Scientia Horticulturae 2023, 307, 111520. [Google Scholar] [CrossRef]
- Kunadiya, M.B.; Burgess, T.I.; A Dunstan, W.; White, D.; StJ. Hardy, G.E. Persistence and degradation of Phytophthora cinnamomi DNA and RNA in different soil types. Environmental DNA 2021, 3, 92–104. [Google Scholar] [CrossRef]
- Puertolas, A.; Bonants, P.J.; Boa, E.; Woodward, S. Application of real-time PCR for the detection and quantification of oomycetes in ornamental nursery stock. Journal of Fungi 2021, 7, 87. [Google Scholar] [CrossRef]
- Eggertson, Q.A.; Rintoul, T.L.; Lévesque, C.A. Resolving the Globisporangium ultimum (Pythium ultimum) species complex. Mycologia 2023, 115, 768–786. [Google Scholar] [CrossRef] [PubMed]
- Canna Research. Pythium—Pests and Diseases. Available online: https://www.canna.ca/articles/pythium-pests-diseases (accessed on 29 December 2025).
- Salcedo, A.F.; Purayannur, S.; Standish, J.R.; Miles, T.; Thiessen, L.; Quesada-Ocampo, L.M. Fantastic Downy Mildew Pathogens and How to Find Them: Advances in Detection and Diagnostics. Plants (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.; Pellan, L.; Jaswa, A.; Cambon, M.C.; Chataigner, A.; Bonnard, O.; Raynal, M.; Debord, C.; Poeydebat, C.; Labarthe, S.; et al. Revealing microbial consortia that interfere with grapevine downy mildew through microbiome epidemiology. Environmental Microbiome 2025, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Büttner, C.; von Bargen, S.; Bandte, M. Phytopathogenic viruses. In Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Springer, 2014; pp. 115–122. [Google Scholar]
- Clinic, Cleveland. Virus. Available online: https://my.clevelandclinic.org/health/body/24861-virus (accessed on 10 December 2025).
- Krauss, S.; Griebler, C.; Technikwissenschaften, D.A.d. Pathogenic microorganisms and viruses in groundwater; acatech, 2011. [Google Scholar]
- Lopez-Roblero, A.; Martínez Cano, D.J.; Diego-García, E.; Guillén-Navarro, G.K.; Iša, P.; Zarza, E. Metagenomic analysis of plant viruses in tropical fresh and wastewater. Environmental DNA 2024, 6, e416. [Google Scholar] [CrossRef]
- Mitrofanova, I.V.; Zakubanskiy, A.V.; Mitrofanova, O.V. Viruses infecting main ornamental plants: an overview. Ornamental Horticulture 2018, 24, 95–102. [Google Scholar] [CrossRef]
- Hamelin, F.M.; Allen, L.J.S.; Prendeville, H.R.; Hajimorad, M.R.; Jeger, M.J. The evolution of plant virus transmission pathways. Journal of Theoretical Biology 2016, 396, 75–89. [Google Scholar] [CrossRef]
- Cornejo-Franco, J.F.; Medina-Salguero, A.; Flores, F.; Chica, E.; Grinstead, S.; Mollov, D.; Quito-Avila, D.F. Exploring the virome of Vasconcellea x heilbornii: the first step towards a sustainable production program for babaco in Ecuador. European Journal of Plant Pathology 2020, 157, 961–968. [Google Scholar] [CrossRef]
- Fox, A.; Fowkes, A.; Skelton, A.; Harju, V.; Buxton-Kirk, A.; Kelly, M.; Forde, S.; Pufal, H.; Conyers, C.; Ward, R. Using high-throughput sequencing in support of a plant health outbreak reveals novel viruses in Ullucus tuberosus (Basellaceae). Plant Pathology 2019, 68, 576–587. [Google Scholar] [CrossRef]
- Sepúlveda, M.; Cardona, D.; García, Y.G.; Higuita, M.; Gutiérrez, P.A.; Marín, M. Virome analysis for identification of viruses associated with asymptomatic infection of purple passion fruit (Passiflora edulis f. edulis) in Colombia. The Journal of Horticultural Science and Biotechnology 2022, 97, 187–200. [Google Scholar] [CrossRef]
- Baráth, D.; Jaksa-Czotter, N.; Molnár, J.; Varga, T.; Balássy, J.; Szabó, L.K.; Kirilla, Z.; Tusnády, G.E.; Preininger, É.; Várallyay, É. Small RNA NGS revealed the presence of Cherry virus A and Little cherry virus 1 on apricots in Hungary. Viruses 2018, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Pais da Cunha, A.T.; Chiumenti, M.; Ladeira, L.C.; Abou Kubaa, R.; Loconsole, G.; Pantaleo, V.; Minafra, A. High throughput sequencing from Angolan citrus accessions discloses the presence of emerging CTV strains. Virology Journal 2021, 18, 62. [Google Scholar] [CrossRef] [PubMed]
- Zikeli, K.; Berwarth, C.; Born, U.; Leible, T.; Jelkmann, W.; Hagemann, M.H. Prevalence, genetic diversity, and molecular detection of the apple hammerhead viroid in Germany. Front Microbiol 2025, 16, 1592572. [Google Scholar] [CrossRef]
- Thapa, P.; Diksha, D.; Sharma, S.K.; Khan, Z.A.; Gupta, N.; Shimray, M.Y.; Prajapati, M.R.; Holkar, S.K.; Naik, S.; Saha, S.; et al. Understanding the dissemination of viruses and viroids identified through virome analysis of major grapevine rootstocks and RPA-based detection of prevalent grapevine virus B. Scientia Horticulturae 2024, 337, 113538. [Google Scholar] [CrossRef]
- Choi, J.; Osatuke, A.C.; Erich, G.; Stevens, K.; Hwang, M.S.; Al Rwahnih, M.; Fuchs, M. High-Throughput Sequencing Reveals Tobacco and Tomato Ringspot Viruses in Pawpaw. Plants 2022, 11, 3565. [Google Scholar] [CrossRef]
- Choi, D.; Meier, S.K.; Park, H.; Baek, S.; Choi, M.S.; Ingle, R.A.; Hahn, Y. Identification of a novel potyvirus from the nickel-hyperaccumulating plant Senecio coronatus in South Africa. South African Journal of Botany 2025, 181, 232–235. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Kim, H.; Yi, S.-I.; Lim, S.; Park, J.M.; Cho, H.S.; Kim, H.; Kwon, S.-Y.; Moon, J.S. First report of Impatiens flower break virus infecting Impatiens walleriana in South Korea. Plant Disease 2017, 101, 394–394. [Google Scholar] [CrossRef]
- Li, Y.; Xia, F.; Wang, Y.; Yan, C.; Jia, A.; Zhang, Y. Characterization of a highly divergent Sugarcane mosaic virus from Canna indica L. by deep sequencing. BMC Microbiol 2019, 19, 260. [Google Scholar] [CrossRef] [PubMed]
- Amoia, S.S.; Minafra, A.; Nicoloso, V.; Loconsole, G.; Chiumenti, M. A New Jasmine Virus C Isolate Identified by Nanopore Sequencing Is Associated to Yellow Mosaic Symptoms of Jasminum officinale in Italy. Plants 2022, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Harrigian, F.; LeBlanc, N.; Eriksen, R.L.; Bush, E.; Salamanca, L.R.; Salgado-Salazar, C. Uncovering the Fungus Responsible for Stem and Root Rot of False Indigo: Pathogen Identification, New Disease Description, and Genome Analyses. Plant Dis 2025, 109, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol Plant Pathol 2011, 12, 938–954. [Google Scholar] [CrossRef]
- Valentin, R.E.; Fonseca, D.M.; Nielsen, A.L.; Leskey, T.C.; Lockwood, J.L. Early detection of invasive exotic insect infestations using eDNA from crop surfaces. Frontiers in Ecology and the Environment 2018, 16, 265–270. [Google Scholar] [CrossRef]
- Lee-Rodriguez, J.; Ranger, Christopher M.; Leach, A.; Michel, A.; Reding, Michael E.; Canas, L. Using Environmental DNA to Detect and Identify Sweetpotato Whitefly Bemisia argentifolii and Twospotted Spider Mite Tetranychus urticae in Greenhouse-Grown Tomato Plants. Environmental DNA 2024, 6, e70026. [Google Scholar] [CrossRef]
- Beaver-Kanuya, E.; Harper, S.J. Detection and quantification of four viruses in Prunus pollen: Implications for biosecurity. J Virol Methods 2019, 271, 113673. [Google Scholar] [CrossRef]
- Smadi, M.; Lee, E.; Phelan, J.; Wang, A.; Bilodeau, G.J.; Pernal, S.F.; Guarna, M.M.; Rott, M.; Griffiths, J.S. Plant virus diversity in bee and pollen samples from apple (Malus domestica) and sweet cherry (Prunus avium) agroecosystems. Frontiers in Plant Science 2024, 15–2024. [Google Scholar] [CrossRef]
- Santala, J.; Samuilova, O.; Hannukkala, A.; Latvala, S.; Kortemaa, H.; Beuch, U.; Kvarnheden, A.; Persson, P.; Topp, K.; Ørstad, K.; et al. Detection, distribution and control of Potato mop-top virus, a soil-borne virus, in northern Europe. Annals of Applied Biology 2010, 157, 163–178. [Google Scholar] [CrossRef]
- Deb, M.; Anderson, J.M. Development of a multiplexed PCR detection method for Barley and Cereal yellow dwarf viruses, Wheat spindle streak virus, Wheat streak mosaic virus and Soil-borne wheat mosaic virus. J Virol Methods 2008, 148, 17–24. [Google Scholar] [CrossRef]
- Wang, Q.; Han, Y.; Yu, Z.; Tian, S.; Sun, P.; Shi, Y.; Peng, C.; Gu, T.; Li, Z. Fungi in Horticultural Crops: Promotion, Pathogenicity and Monitoring. Agronomy 2025, 15, 1699. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Aiello, D.; Polizzi, G.; Crous, P.W.; Sandoval-Denis, M. Soilborne diseases caused by Fusarium and Neocosmospora spp. on ornamental plants in Italy. Phytopathologia Mediterranea 2019, 58, 127–138. [Google Scholar]
- Gullino, M.L.; Daughtrey, M.L.; Garibaldi, A.; Elmer, W.H. Fusarium wilts of ornamental crops and their management. Crop Protection 2015, 73, 50–59. [Google Scholar] [CrossRef]
- Vlad, F.F.; Iacomi, B.M. Pathogenic mycobiota of ornamental plants from green area in the city of Bucharest, Romania. Scientific Papers. Series A. Agronomy 2024, 67. [Google Scholar]
- Guarnaccia, V.; Gilardi, G.; Martino, I.; Garibaldi, A.; Gullino, M.L. Species diversity in Colletotrichum causing anthracnose of aromatic and ornamental Lamiaceae in Italy. Agronomy 2019, 9, 613. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Manawasinghe, I.; Lin, Y.; Jayawardena, R.; McKenzie, E.; Hyde, K.; Xiang, M. Identification and characterization of Colletotrichum species associated with ornamental plants in Southern China. Mycosphere 2023, 14, 262–302. [Google Scholar] [CrossRef]
- Larsson, R.; Menkis, A.; Skogström, O.; Espes, C.; Brogren-Mohlin, E.-K.; Larsson, M.; Olson, Å. The development of foliar fungal communities of nursery-grown Pinus sylvestris seedlings. Scandinavian Journal of Forest Research 2023, 38, 513–528. [Google Scholar] [CrossRef]
- Bérubé, J.A.; Gagné, P.N.; Ponchart, J.P.; Phelan, J.; Varga, A.; James, D. Heterobasidion species detected using High Throughput Sequencing (HTS) methods on British Columbia nursery plants. Canadian Journal of Plant Pathology 2019, 41, 560–565. [Google Scholar] [CrossRef]
- Garcia, J.F.; Todd, C.; Rolshausen, P.E.; Cantu, D. Nursery origin and propagation stage influence the endophytic fungal communities inhabiting grapevine planting stocks. Phytobiomes Journal 2025, 9, 287–301. [Google Scholar] [CrossRef]
- Lade, S.B.; Štraus, D.; Oliva, J. Variation in fungal community in grapevine (Vitis vinifera) nursery stock depends on nursery, variety and rootstock. Journal of Fungi 2022, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Wise, K.; Mueller, D.; Buck, J.; Fowler, J. Quarantines and ornamental rusts; American Pathological Society: St. Paul, 2004. [Google Scholar]
- Timmermann, V.; Børja, I.; Hietala, A.M.; Kirisits, T.; Solheim, H. Ash dieback: pathogen spread and diurnal patterns of ascospore dispersal, with special emphasis on Norway. EPPO Bulletin 2011, 41, 14–20. [Google Scholar] [CrossRef]
- Soldi, E.; Tiley, A.; O’Hanlon, R.; Murphy, B.R.; Hodkinson, T.R. Ash dieback and other pests and pathogens of Fraxinus on the island of Ireland. In Proceedings of the Biology and Environment: Proceedings of the Royal Irish Academy, 2022; pp. 85–122. [Google Scholar]
- Liu, M.; Braun, U. Powdery mildews on crops and ornamentals in Canada: a summary of the phylogeny and taxonomy from 2000–2019. Canadian Journal of Plant Pathology 2022, 44, 191–218. [Google Scholar] [CrossRef]
- Matsuoka, S.; Sugiyama, Y.; Nagano, M.; Doi, H. Influence of DNA extraction kits on freshwater fungal DNA metabarcoding. PeerJ 2022, 10, e13477. [Google Scholar] [CrossRef]
- Congram, M.; Torres Vilaça, S.; Wilson, C.C.; Kyle, C.J.; Lesbarrères, D.; Wikston, M.J.H.; Beaty, L.; Murray, D.L. Tracking the prevalence of a fungal pathogen, Batrachochytrium dendrobatidis (chytrid fungus), using environmental DNA. Environmental DNA 2022, 4, 687–699. [Google Scholar] [CrossRef]
- French, L.C.; Jusino, M.A.; Chambers, R.M.; Skelton, J. Community structure and functional diversity of aquatic fungi are correlated with water quality: Insights from multi-marker analysis of environmental DNA in a coastal watershed. Environmental DNA 2024, 6, e576. [Google Scholar] [CrossRef]
- McLaughlin, K.E.; Belaouni, H.A.; McClure, A.; McCullough, K.; Craig, D.; McKeown, J.; Stevenson, M.A.; Carmichael, E.; Dalzell, J.; O’Hanlon, R.; et al. Using Environmental DNA as a Plant Health Surveillance Tool in Forests. Forests 2025, 16, 632. [Google Scholar] [CrossRef]
- Chandelier, A.; Massot, M.; Fabreguettes, O.; Gischer, F.; Teng, F.; Robin, C. Early detection of Cryphonectria parasitica by real-time PCR. European Journal of Plant Pathology 2019, 153, 29–46. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kowalkowski, W.; Kartawik, N.; Baranowska, M.; Barzdajn, W. The soil fungal communities in nurseries producing Abies alba. Baltic Forestry 2020, 26. [Google Scholar] [CrossRef]
- Baranowska, M.; Behnke-Borowczyk, J.; Barzdajn, W.; Szmyt, J.; Korzeniewicz, R.; Łukowski, A.; Memišević-Hodžić, M.; Kartawik, N.; Kowalkowski, W. Effects of nursery production methods on fungal community diversity within soil and roots of Abies alba Mill. Scientific Reports 2023, 13, 21284. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Mills, J.G.; Gellie, N.J.C.; Bissett, A.; Lowe, A.J.; Breed, M.F. High-throughput eDNA monitoring of fungi to track functional recovery in ecological restoration. Biological Conservation 2018, 217, 113–120. [Google Scholar] [CrossRef]
- Frøslev, T.G.; Kjøller, R.; Bruun, H.H.; Ejrnæs, R.; Hansen, A.J.; Læssøe, T.; Heilmann-Clausen, J. Man against machine: Do fungal fruitbodies and eDNA give similar biodiversity assessments across broad environmental gradients? Biological Conservation 2019, 233, 201–212. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol 2012, 13, 614–629. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Bartoli, C. Plant pathogenic bacteria in open irrigation systems: what risk for crop health? Plant Pathology 2015, 64, 757–766. [Google Scholar] [CrossRef]
- Orlovskis, Z.; Canale, M.C.; Thole, V.; Pecher, P.; Lopes, J.R.; Hogenhout, S.A. Insect-borne plant pathogenic bacteria: getting a ride goes beyond physical contact. Curr Opin Insect Sci 2015, 9, 16–23. [Google Scholar] [CrossRef]
- Martins, P.M.M.; Merfa, M.V.; Takita, M.A.; De Souza, A.A. Persistence in Phytopathogenic Bacteria: Do We Know Enough? Front Microbiol 2018, 9, 1099. [Google Scholar] [CrossRef]
- McEvoy, A.; O’Regan, F.; Fleming, C.C.; Moreland, B.P.; Pollock, J.A.; McGuinness, B.W.; Hodkinson, T.R. Bleeding canker of horse chestnut (Aesculus hippocastanum) in Ireland: incidence, severity and characterization using DNA sequences and real-time PCR. Plant Pathology 2016, 65, 1419–1429. [Google Scholar] [CrossRef]
- Yang, S.; Johnson, M.A.; Hansen, M.A.; Bush, E.; Li, S.; Vinatzer, B.A. Metagenomic sequencing for detection and identification of the boxwood blight pathogen Calonectria pseudonaviculata. Scientific Reports 2022, 12, 1399. [Google Scholar] [CrossRef]
- Keremane, M.; Singh, K.; Ramadugu, C.; Krueger, R.R.; Skaggs, T.H. Next Generation Sequencing, and Development of a Pipeline as a Tool for the Detection and Discovery of Citrus Pathogens to Facilitate Safer Germplasm Exchange. Plants 2024, 13, 411. [Google Scholar] [CrossRef]
- Ban, L.-P.; Li, J.-D.; Yan, M.; Gao, Y.-H.; Zhang, J.-J.; Moural, T.W.; Zhu, F.; Wang, X.-M. Illumina Sequencing of 18S/16S rRNA Reveals Microbial Community Composition, Diversity, and Potential Pathogens in 17 Turfgrass Seeds. Plant Disease 2021, 105, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; van Elsas, J.D. Mechanisms and ecological implications of the movement of bacteria in soil. Applied Soil Ecology 2018, 129, 112–120. [Google Scholar] [CrossRef]
- Bashan, Y. Migration of the rhizosphere bacteria Azospirillum brusilense and Pseudomonas fluorescens towards wheat roots in the soil. Microbiology 1986, 132, 3407–3414. [Google Scholar] [CrossRef]
- Covelli, J.M.; Althabegoiti, M.J.; López, M.F.; Lodeiro, A.R. Swarming motility in Bradyrhizobium japonicum. Research in Microbiology 2013, 164, 136–144. [Google Scholar] [CrossRef]
- Bové, J.M.; Garnier, M. Phloem-and xylem-restricted plant pathogenic bacteria. Plant Science 2003, 164, 423–438. [Google Scholar] [CrossRef]
- Picot, A.; Cobo-Díaz, J.F.; Pawtowski, A.; Donot, C.; Legrand, F.; Le Floch, G.; Déniel, F. Water Microbiota in Greenhouses With Soilless Cultures of Tomato by Metabarcoding and Culture-Dependent Approaches. Frontiers in Microbiology 2020, 11–2020. [Google Scholar] [CrossRef]
- Kozdrój, J.; Ropek, D.R.; Frączek, K.; Bulski, K.; Breza-Boruta, B. Microbial Indoor Air Quality Within Greenhouses and Polytunnels Is Crucial for Sustainable Horticulture (Malopolska Province, Poland Conditions). Sustainability 2024, 16, 10058. [Google Scholar] [CrossRef]
- Riffaud, C.-H.; Morris, C.E. Detection of Pseudomonas syringae pv. aptata in irrigation water retention basins by immunofluorescence colony-staining. European Journal of Plant Pathology 2002, 108, 539–545. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.; Vanneste, J.; Montarry, J.; Oakley, B.; Guilbaud, C.; Glaux, C. Inferring the evolutionary history of the plant pathogen Pseudomonas syringae from its biogeography in headwaters of rivers in North America, Europe, and New Zealand. MBio 2010, 1. [Google Scholar] [CrossRef]
- Elphinstone, J.; Stanford, H.; Stead, D. Detection of Ralstonia solanacearum in potato tubers, Solanum dulcamara and associated irrigation water. In Bacterial wilt disease: Molecular and ecological aspects; Springer, 1998; pp. 133–139. [Google Scholar]
- Singh, D.; Sinha, S.; Yadav, D.; Chaudhary, G. Detection of Ralstonia solanacearum from asymptomatic tomato plants, irrigation water, and soil through non-selective enrichment medium with hrp gene-based bio-PCR. Current microbiology 2014, 69, 127–134. [Google Scholar] [CrossRef]
- Harrison, M.; Franc, G.; Maddox, D.; Michaud, J.; McCarter-Zorner, N. Presence of Erwinia carotovora in surface water in North America. Journal of Applied Microbiology 1987, 62, 565–570. [Google Scholar] [CrossRef]
- Laurila, J.; Ahola, V.; Lehtinen, A.; Joutsjoki, T.; Hannukkala, A.; Rahkonen, A.; Pirhonen, M. Characterization of Dickeya strains isolated from potato and river water samples in Finland. European Journal of Plant Pathology 2008, 122, 213–225. [Google Scholar] [CrossRef]
- Atmatjidou, V.; Fynn, R.; Hoitink, H. Dissemination and transmission of. Xanthomonas campestris pv. begoniae in an ebb and flow irrigation system 1991. [Google Scholar]
- Gent, D.H.; Lang, J.M.; Bartolo, M.E.; Schwartz, H.F. Inoculum sources and survival of Xanthomonas axonopodis pv. allii in Colorado. Plant disease 2005, 89, 507–514. [Google Scholar] [CrossRef]
- Schuster, M.; Wagner, L. Control of bacterial canker and root knot of hydroponic tomatoes. 1972. [Google Scholar]
- Qiagen. FAQ-114 How should I store plant material for DNA isolation using the DNeasy Plant Kit? Available online: https://www.qiagen.com/us/resources/faq/114 (accessed on 08 December 2025).
- Sylphium Molecular Ecology. Complete eDNA sampling sets (20x). Available online: https://sylphium.com/webshop/product/syl009/ (accessed on 10 December 202).
- Merck, KGaA. Reinforced Membrane Filter. Millipore, filter diam. 47mm, hydrophilic. Available online: https://www.sigmaaldrich.com/IE/en/product/mm/rw1904700 (accessed on 10 December 2025).
- Cooper Pegler, NAD. Cooper Pegler CP15 Sprayer 15 LTR. Available online: https://www.nad.ie/product/cooper-pegler-cp15-sprayer-15-ltr/ (accessed on 10 December 2025).
- Sartorius, AG. Re-usable Syringe Filter Holder 25mm. Available online: https://shop.sartorius.com/ie/p/re-usable-syringe-filter-holder-25-mm/16517----------E (accessed on 10 December 2025).
- Sartorius, AG. MD8 Airport Portable Air Sampler. Available online: https://shop.sartorius.com/ie/p/md8-airport-portable-air-sampler/16757 (accessed on 10 December 2025).
- Chemical, Cayman. Coriolis Compact Air Sampler Item No. 38284. Available online: https://www.caymanchem.com/product/38284/coriolis-registered-compact-air-sampler?srsltid=AfmBOoo0GnVPEZEWh0SEMThxeTBDhAXn7IHyhECmLvLHrJPKBfEMicFH (accessed on 10 December 2025).
- Sartorius, AG. Disposable Gelatine Membrane Filters, Diameter 80mm, triple packed, Pack Size 10. Available online: https://shop.sartorius.com/ie/p/disposable-gelatine-membrane-filters-diameter-80mm-triple-packed-pack-size-10/Gelatine-Filter-Method (accessed on 10 December 2025).
- Tremblay, É.D.; Duceppe, M.O.; Thurston, G.B.; Gagnon, M.C.; Côté, M.J.; Bilodeau, G.J. High-resolution biomonitoring of plant pathogens and plant species using metabarcoding of pollen pellet contents collected from a honey bee hive. Environmental DNA 2019, 1, 155–175. [Google Scholar] [CrossRef]
- Redekar, N.R.; Bourret, T.B.; Eberhart, J.L.; Johnson, G.E.; Pitton, B.J.; Haver, D.L.; Oki, L.R.; Parke, J.L. The population of oomycetes in a recycled irrigation water system at a horticultural nursery in southern California. Water Research 2020, 183, 116050. [Google Scholar] [CrossRef]
- Paudel, K.P.; Pandit, M.; Hinson, R. Irrigation water sources and irrigation application methods used by US plant nursery producers. Water Resources Research 2016, 52, 698–712. [Google Scholar] [CrossRef]
- Wittwer, C.; Nowak, C.; Strand, D.A.; Vrålstad, T.; Thines, M.; Stoll, S. Comparison of two water sampling approaches for eDNA-based crayfish plague detection. Limnologica 2018, 70, 1–9. [Google Scholar] [CrossRef]
- Pettitt, T.R. Monitoring oomycetes in water: combinations of methodologies used to answer key monitoring questions. Frontiers in Horticulture 2023, 2, 1210535. [Google Scholar] [CrossRef]
- Scibetta, S.; Schena, L.; Chimento, A.; Cacciola, S.O.; Cooke, D.E. A molecular method to assess Phytophthora diversity in environmental samples. J Microbiol Methods 2012, 88, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.E.; Ferrante, J.A.; Meigs-Friend, G.; Ulmer, A. Improving eDNA yield and inhibitor reduction through increased water volumes and multi-filter isolation techniques. Scientific Reports 2019, 9, 5259. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Valentini, A.; Dejean, T.; Montarsi, F.; Taberlet, P.; Glaizot, O.; Fumagalli, L. Detection of Invasive Mosquito Vectors Using Environmental DNA (eDNA) from Water Samples. PLoS One 2016, 11, e0162493. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol Lett 2008, 4, 423–425. [Google Scholar] [CrossRef]
- Schabacker, J.C.; Amish, S.J.; Ellis, B.K.; Gardner, B.; Miller, D.L.; Rutledge, E.A.; Sepulveda, A.J.; Luikart, G. Increased eDNA detection sensitivity using a novel high-volume water sampling method. Environmental DNA 2020, 2, 244–251. [Google Scholar] [CrossRef]
- Hinlo, R.; Gleeson, D.; Lintermans, M.; Furlan, E. Methods to maximise recovery of environmental DNA from water samples. PLoS One 2017, 12, e0179251. [Google Scholar] [CrossRef]
- Wang, Y.; Hammes, F.; Boon, N.; Egli, T. Quantification of the filterability of freshwater bacteria through 0.45, 0.22, and 0.1 microm pore size filters and shape-dependent enrichment of filterable bacterial communities. Environ Sci Technol 2007, 41, 7080–7086. [Google Scholar] [CrossRef]
- Williams, K.E.; Huyvaert, K.P.; Piaggio, A.J. No filters, no fridges: a method for preservation of water samples for eDNA analysis. BMC Research Notes 2016, 9, 298. [Google Scholar] [CrossRef]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The use of genus-specific amplicon pyrosequencing to assess Phytophthora species diversity using eDNA from soil and water in Northern Spain. PLoS One 2015, 10, e0119311. [Google Scholar] [CrossRef]
- Ott, N.J.; Nouri, M.T.; Browne, G.T. Full-Length ITS Amplicon Sequencing Resolves Phytophthora Species in Surface Waters Used for Orchard Irrigation in California’s San Joaquin Valley. Plant Dis 2024, 108, 3562–3577. [Google Scholar] [CrossRef]
- Anteljević, M.; Rosić, I.; Medić, O.; Ranković, T.; Sunjog, K.; Kračun-Kolarević, M.; Kolarević, S.; Dreo, T.; Benčič, A.; Berić, T.; et al. Irrigation systems as reservoirs of diverse and pathogenic Pseudomonas syringae strains endangering crop health. Water Research X 2025, 28, 100380. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Survival of Phytophthora cinnamomi as oospores, stromata, and thick-walled chlamydospores in roots of symptomatic and asymptomatic annual and herbaceous perennial plant species. Fungal biology 2013, 117, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Li, X.; Yang, X.; Andrade, D.; Xue, L.; McKinney, N. Prediction of plant diseases through modelling and monitoring airborne pathogen dispersal. CABI Reviews 2010, 1–11. [Google Scholar] [CrossRef]
- Harnpicharnchai, P.; Pumkaeo, P.; Siriarchawatana, P.; Likhitrattanapisal, S.; Mayteeworakoon, S.; Ingsrisawang, L.; Boonsin, W.; Eurwilaichitr, L.; Ingsriswang, S. AirDNA sampler: An efficient and simple device enabling high-yield, high-quality airborne environment DNA for metagenomic applications. PLoS One 2023, 18, e0287567. [Google Scholar]
- Frankel, M.; Timm, M.; Hansen, E.W.; Madsen, A.M. Comparison of sampling methods for the assessment of indoor microbial exposure. Indoor Air 2012, 22, 405–414. [Google Scholar] [CrossRef]
- Clare, E.L.; Economou, C.K.; Bennett, F.J.; Dyer, C.E.; Adams, K.; McRobie, B.; Drinkwater, R.; Littlefair, J.E. Measuring biodiversity from DNA in the air. Current Biology 2022, 32, 693–700.e695. [Google Scholar] [CrossRef]
- Chen, W.; Radford, D.; Hambleton, S. Towards Improved Detection and identification of rust fungal pathogens in environmental samples using a metabarcoding approach. Phytopathology 2022, 112, 535–548. [Google Scholar] [CrossRef]
- Williams, R.H.; Ward, E.; McCartney, H.A. Methods for integrated air sampling and DNA analysis for detection of airborne fungal spores. Applied and environmental microbiology 2001, 67, 2453–2459. [Google Scholar] [CrossRef]
- Parks, S.R.; Bennett, A.M.; Speight, S.E.; Benbough, J.E. An assessment of the Sartorius MD8 microbiological air sampler. J Appl Bacteriol 1996, 80, 529–534. [Google Scholar] [CrossRef]
- Carvalho, E.; Sindt, C.; Verdier, A.; Galan, C.; O’Donoghue, L.; Parks, S.; Thibaudon, M. Performance of the Coriolis air sampler, a high-volume aerosol-collection system for quantification of airborne spores and pollen grains. Aerobiologia 2008, 24, 191–201. [Google Scholar]
- Banchi, E.; Ametrano, C.G.; Tordoni, E.; Stanković, D.; Ongaro, S.; Tretiach, M.; Pallavicini, A.; Muggia, L.; Verardo, P.; Tassan, F.; et al. Environmental DNA assessment of airborne plant and fungal seasonal diversity. Science of The Total Environment 2020, 738, 140249. [Google Scholar] [CrossRef]
- Tordoni, E.; Ametrano, C.G.; Banchi, E.; Ongaro, S.; Pallavicini, A.; Bacaro, G.; Muggia, L. Integrated eDNA metabarcoding and morphological analyses assess spatio-temporal patterns of airborne fungal spores. Ecological Indicators 2021, 121, 107032. [Google Scholar] [CrossRef]
- Roussel, J.-M.; Paillisson, J.-M.; Treguier, A.; Petit, E. The downside of eDNA as a survey tool in water bodies. Journal of Applied Ecology 2015, 823–826. [Google Scholar] [CrossRef]
- Pochon, X.; Zaiko, A.; Fletcher, L.M.; Laroche, O.; Wood, S.A. Wanted dead or alive? Using metabarcoding of environmental DNA and RNA to distinguish living assemblages for biosecurity applications. PloS one 2017, 12, e0187636. [Google Scholar] [CrossRef]
Figure 1.
Summary of the results obtained from the systematic literature review, illustrating the number of studies found using the contrasting horticultural sampling matrices (plant material, growing media, water, air) to detect different phytopathogenic groups.
Figure 1.
Summary of the results obtained from the systematic literature review, illustrating the number of studies found using the contrasting horticultural sampling matrices (plant material, growing media, water, air) to detect different phytopathogenic groups.
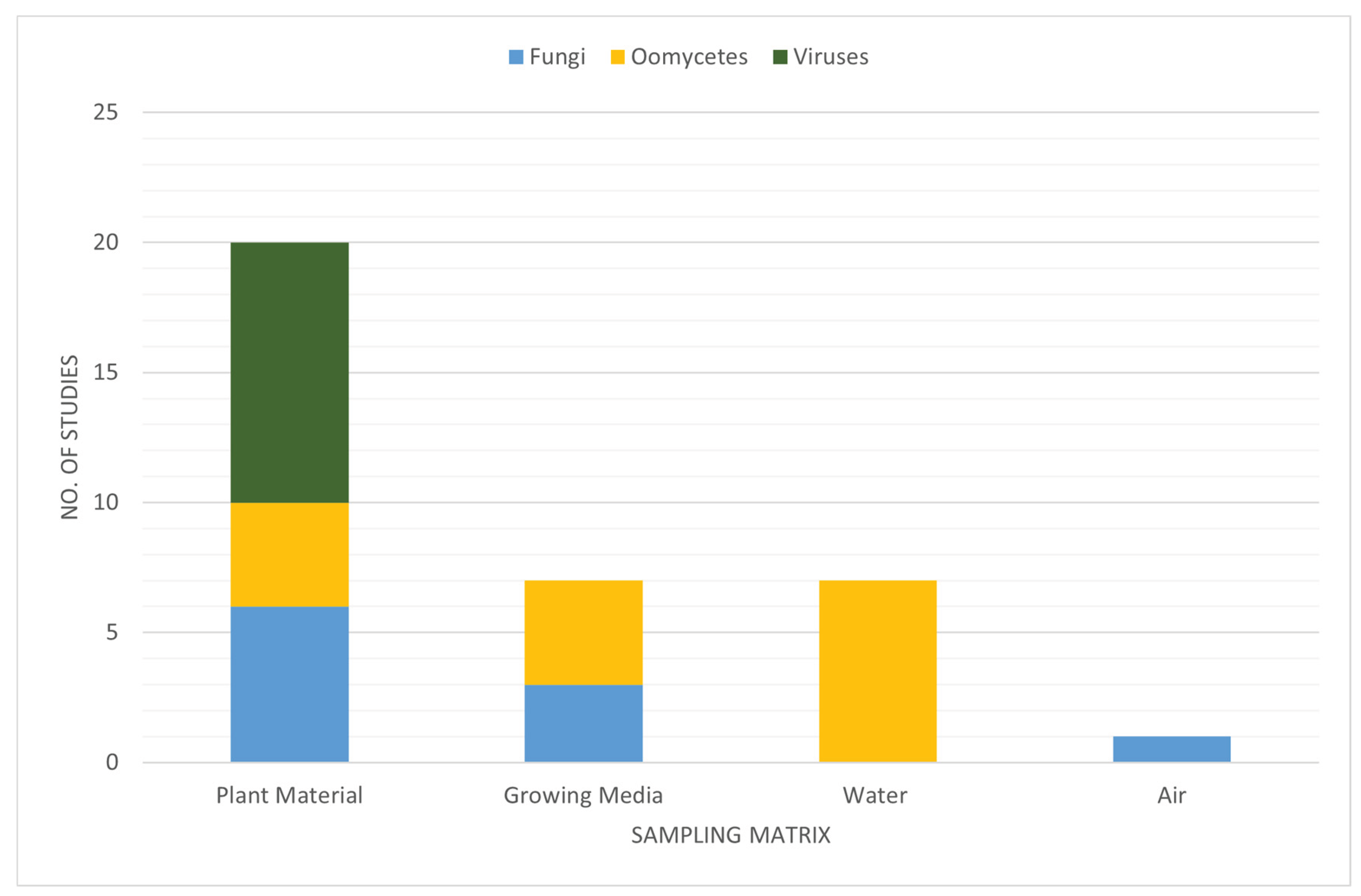
Figure 2.
Simple schematic of a water-testing “trace-back” system to locate infected plants within a nursery.
Figure 2.
Simple schematic of a water-testing “trace-back” system to locate infected plants within a nursery.
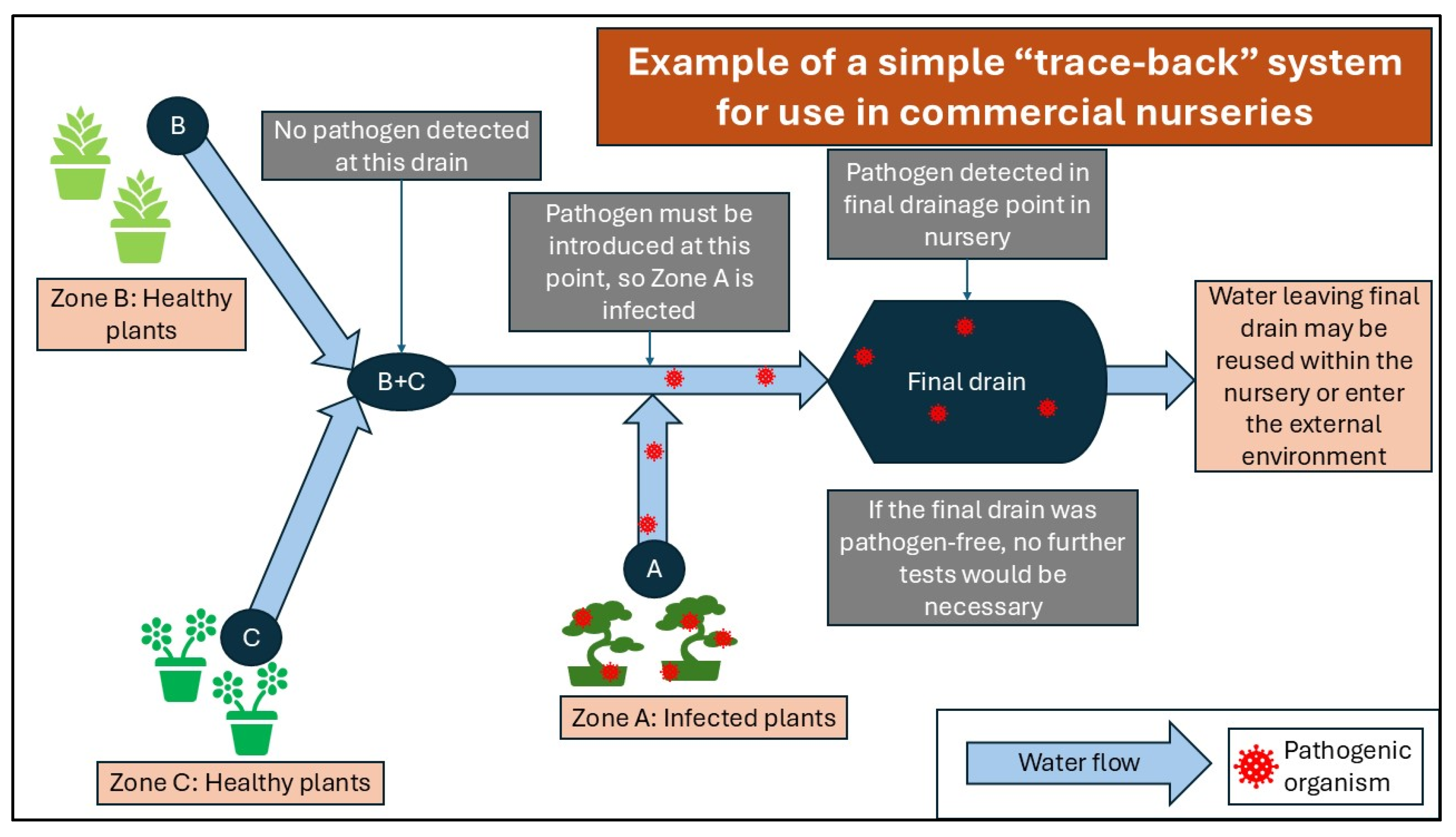

Table 1.
Summary of the main Oomycete groups and the appropriate sample media to detect them.
| Oomycete group | Recommended sample type | References |
|---|---|---|
| Pythium spp. | Growing media; water | [24,46,48,49,50,52] |
| Phytophthora spp. | Growing media; water; air | [22,24,46,48,50] |
| Downy mildew spp. | Air | [48,53] |
Table 2.
Logistical considerations associated with each sampling medium.
| Sample type | Organisms detectable | Cost of single set up | Consumables cost per sample (not incl. extraction or sequencing) | Requirements (sampling) | Requirements (storage) | References |
|---|---|---|---|---|---|---|
| Plant material | All | Negligible | Negligible | Sterile blade for taking portion of plant | Liquid nitrogen and -70 °C freezer OR silica gel in airtight container | [139] |
| Water | All | Varies depending on set up required, e.g., Knapsack Sprayer 15L* costs ca. 250-300EUR and re-usable syringe filter holders cost ca. 200EUR | From 1EUR per sample for consumables (i.e., filters and storage vials) OR ca. 15EUR for complete single-use kit | Appropriate filters, filter units, and pumps OR commercially available kit | Buffer solution & sterile container | [140,141,142,143] |
| Air | Fungi, viruses, oomycetes | e.g., Air samplers** cost 5,000-15,000EUR | Filters cost from ca. 20EUR per sample | Air sampler AND appropriate filters, filter units, and pumps | Buffer solution & sterile container | [144,145,146] |
| Growing media | Fungi, bacteria, oomycetes | Negligible | Negligible | Sterile shovel/spoon, bag for homogenizing | Buffer solution & sterile container | [25,50] |
*e.g., Cooper Pegler CP15 2000 Series **e.g., Sartorius MD8 Airport Air Sampler; Compact Air Sampler Coriolis (Bertin Technologies).
Table 3.
Summary of how various organisms have been successfully detected using water sampling.
| Pathogen group | Organism | Water volume | Filter used | Reference |
|---|---|---|---|---|
| Oomycetes | Phytophthora | 10L | 5µm cellulose acetate and cellulose nitrate mixture | [160] |
| Phytophthora, Pythium, Phytopythium | 1L | 5µm mixed cellulose ester filters | [46] | |
| Phytophthora | 3 x 5L | 1.2µm mixed cellulose ester filters | [22] | |
| Phytophthora | ~15L | Combined a 100µm nylon mesh filter, a 20µm nylon mesh filter, and a 5µm nitrocellulose filter | [161] | |
| Bacteria | Pseudomonas syringae | 1L | 0.45µm mixed cellulose ester filters | [162] |
| Fungi | Aquatic fungi | 1L | 0.7µm GF/F glass filters | [107] |
| Batrachochytrium dendrobatidis (fungal amphibian pathogen) | <500mL (average 270mL) | 0.22µm capsule filters | [108] | |
| Aquatic fungi | 3 x 2L | 5µm polyethersulfone (PES) membrane filters | [109] | |
| Viruses | Phytopathogenic viruses from the Virgaviridae family | 15mL | n/a | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



