Submitted:
09 January 2026
Posted:
13 January 2026
You are already at the latest version
Abstract
Melanogenesis is a highly complex process regulated by multiple signaling pathways that control melanin synthesis in melanocytes and its subsequent transfer to keratinocytes. This process is further influenced by an intricate network of interactions among various skin cell populations, including inflammatory cells, which release paracrine factors in response to internal and external stimuli, such as UV radiation. The aim of this study was to evaluate effectiveness a new cosmetic formulation Skin Glow Complex designed for the topical treatment of skin dyschromia. We investigated the potential benefits of the for-mulation in two major resident skin cell types, keratinocytes and fibroblasts subjected to UV irradiation. Additionally, its effects were tested in 3D human melanocyte spheroid model, that better mimics the skin's environment. Treatment with the new formulation prevented UV-induced reactive oxygen species (ROS) formation in keratinocytes. In dermal fibroblasts, the formulation decreased the expression of matrix metalloproteinases while simultaneously promoting cell proliferation and collagen synthesis. Finally, results obtained from the melanocyte spheroid model confirmed the formulation’s ability to reduce melanin production, reinforcing its potential use in the treatment of skin dyschromia. Overall, these findings indicate that the new product represents a promising natural option to support skin repair and counteract aging and UV-induced damage.
Keywords:
spheroid
; 3D culture
; keratinocyte
; melanocyte
; fibroblast
; skin repair
1. Introduction
Melanogenesis is the process by which melanocytes synthesize melanin within specialized organelles known as melanosomes and represents a fundamental defense mechanism against ultraviolet (UV) induced damage [1]. This process synthesizes two types of melanin: eumelanin and pheomelanin, which protect the skin from damaging UV radiation [1]. Melanocytes are neural crest derived cells located in the basal layer of the epidermis and functionally interact with keratinocytes through paracrine signaling, regulating pigment production and transfer [2]. Differences in skin pigmentation among individuals are not determined by melanocyte number, which remains relatively constant, but rather by melanosome characteristics, the type of melanin produced, and its distribution within keratinocytes. Disruptions in these processes may lead to pigmentary disorders influenced by both intrinsic factors, such as genetics and hormonal regulation, and extrinsic factors, particularly UV exposure [1].
Solar radiation includes UVA, UVB, and UVC wavelengths; however, only UVA and UVB can reach the Earth’s surface. UVA radiation penetrates deeply into the dermis, inducing oxidative stress and degradation of structural fibers, thereby contributing to photoaging and immediate, transient pigmentation. In contrast, UVB radiation primarily affects the epidermis, where it stimulates melanogenesis, causes erythema and DNA damage, and plays a key role in skin carcinogenesis. Excessive or prolonged exposure to UV radiation is therefore a major risk factor for the development of pigmentary alterations [3].
The term dyschromia refers to alterations in normal skin pigmentation and may present as hypopigmentation or hyperpigmentation [4]. Hypopigmented conditions are characterized by a partial or complete reduction in melanin production. Hyperpigmentation, more commonly acquired, may be localized or diffuse and generally results from increased melanin synthesis or abnormal pigment distribution. Major contributing factors include UV exposure, chronic inflammation, mechanical irritation, and hormonal stimulation [4].
Among acquired hyperpigmentary disorders, melasma is a chronic and recurrent condition that predominantly affects women and is exacerbated by sun exposure, typically involving the facial region [5,6,7,8]. Its pathogenesis is multifactorial and not yet fully understood [7,8]. Another common condition is post-inflammatory hyperpigmentation, which develops following cutaneous inflammation or trauma and tends to be more persistent in darker skin phototypes [9,10,11]. In these cases, melanin accumulation may be confined to the epidermis or deposited within the dermis because of basement membrane damage [12]. Up to date, many efforts have been made to apply several technologies, such as exosomes used, to the development of novel therapeutic strategies to achieve skin regeneration and repair [13]. Exosomes are nanosized extracellular vesicles (100-150 nm) that carry lipids, proteins, and nucleic acids, including growth factors and cytokines, and play a crucial role in intercellular communication [14]. They exhibit high stability, safety even at high doses, and the ability to penetrate the stratum corneum, making them particularly suitable for dermatological applications [15]. Among natural sources of exosomes, bovine colostrum represents an abundant and easily accessible reservoir with high isolation yields. Colostrum-derived exosomes contain elevated levels of growth factors (such as TGF-β, IGF, and FGF) and immunomodulatory cytokines, which synergistically contribute to skin regeneration processes [15,16,17]. The objective of this research was to evaluate the efficacy of a novel formulation, Skin Glow Complex (SGC), containing exosomes, growth factors, and cytokines purified from bovine colostrum, on different human skin cell lines, including keratinocytes, fibroblasts, and melanocytes. Given the central role of these cell types in maintaining skin structure, function, and pigmentation, we aimed to investigate the biological effects of SGC on key cellular processes involved in skin homeostasis and regeneration. Specifically, the study assessed the ability of SGC to modulate cellular responses associated with photoinduced damage, extracellular matrix remodeling, oxidative stress, and melanogenesis. By evaluating its effects in both conventional in vitro models and advanced three-dimensional melanocyte spheroid systems that more closely recapitulate the physiological skin microenvironment, this work seeks to provide mechanistic insight into the potential of colostrum-derived bioactive components as a cell-free, multi-target therapeutic strategy for skin regeneration and functional improvement.
2. Materials and Methods
For our research we purchased a novel product called Skin Glow Complex 100+ (SGC100+) contains exosomes purified from bovine colostrum passively loaded with growth factors and cytokines purified from bovine colostrum. In particular, this product includes 20 billion exosomes and 200 mg of growth factors. The product contains also hyaluronic acid and niacinamide.
2.1. Propagation and Maintenance of Cells
Normal adult human dermal fibroblasts (HDFa) (ThermoFisher Scientific) were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% FBS, streptomycin (0.3 mg mL–1) and penicillin (50 IU mL–1), and GlutaMAX (1 mM) using 75 cm2 polystyrene culture flasks. Cells were maintained in a humidified environment (37 °C and 5% CO2) and split every 2-3 days depending on cell confluence. Normal adult primary human epidermal keratinocytes (HEKa) (Invitrogen) were maintained in a humidified incubator with 5% CO2 at 37˚C in Epilife Medium according to the manufacturer's instructions and split every 2-3 days depending on cell confluence. The cells to be irradiated with UV-C were washed with PBS and then irradiated with UV-C for the necessary time using a UV-C lamp. Immediately after UV-C irradiation, the cells were cultured in their respective media.
2.2. UV-C Irradiation and Cell Proliferation Assay
Fibroblasts and Keratinocytes were harvested by using a trypsin-EDTA solution, counted with a hemocytometer and plated in 96-well plates (5×103 cells/well). The following day, the cells were irradiated with 4.5 mJ/cm2 of UV-C and cells were treated with SGC100+ at three different concentrations (0.1%, 0.5% and 1%) and incubated for 24 hours in a humidified environment (37 °C and 5% CO2). At the end of the treatment, MTT solution (1 mg/mL) was added to each well and cells were incubated for 2 hours in a humidified environment (37 °C and 5% CO2). Epoch Microplate Spectrophotometer (Bio Tek Instruments) was used to read the absorbance at 569 nm. Values were normalized with respect to control untreated cells and were expressed as the percent variation of cell proliferation assay.
2.3. Collagen Production
In HDFa cells, collagen synthesis was determined over 7 days in the presence or absence of SGC100+ at three different concentrations (0.1%, 0.5% and 1%). Secreted C-terminal propeptide of collagen type I (CICP) was measured in culture supernatant using an enzyme immunoassay kit (Metra Biosystem), according to the manufacturer’s instructions. The collagen synthesis was measured at 405 nm with Epoch Microplate Spectrophotometer (Bio Tek Instruments). Experiments were performed in duplicate.
2.4. Determination of MMP-2 Concentration
MMP-2 expression, constitutively secreted from fibroblasts, was determined with a commercially available quantitative ELISA test (Amersham Pharmacia Biotech). All procedures were performed according to the instructions of the manufacturer. Experiments were performed in duplicate.
2.5. Scratch-Wound Assay
HDFa cells were seeded in six-well plates (2.5 × 106 cells/well) and grown to confluence. A scratch was created using a sterile 1 mL pipette tip, and cells were washed with PBS to remove debris. Fresh medium, with or without the tested product at 1%, was then added. Untreated cells served as the control. Wound closure was monitored at 24 hours post-scratch.
2.6. ROS Estimation
Oxidative stress was measured by detecting intracellular ROS of HEKa. ROS was estimated by 2’,7’-dichlorofluorescein diacetate (Merck, Darmstadt, Germany), which reacts with intracellular oxidative ROS and is converted to dichlorofluorescein (DCF). The cells were seeded at a density of 2.5 ×103 cells/dish and treated with SGC100 at three different concentrations (0.1%, 0.5% and 1%) overnight. As a positive control, the cells were pretreated with 100 µM hydrogen peroxide for 30 min. The cells were irradiated with 0.7 mJ/cm2 of UV-C, and after 6 hours, the cells were incubated with DCF-DA at a concentration of 50 µM for 30 min. The cells were washed once with PBS and observed with a fluorescent microscope (Nikon, GeneTex). Images were acquired with the NIKON DS-Qi2 camera and analyzed with Image J software (version 1.53).
2.7. 3D Cultivation of Cell Lines B16F10 and Treatment with SCG100
Murine melanoma cell lines B16F10 (ATCC) were cultured in Dulbecco’s Modified Eagle Media (DMEM) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. According to [18] we used the hanging-drop method to generate melanocyte aggregates (spheroids). Methylcellulose was added to the cell culture medium at a concentration of 0.24% w/v to make a cell suspension containing an appropriate number of cells. An array of 25 μL drops of culture medium containing 1,000 cells was made on the lid of the culture dish. Then the lid was carefully flipped over and placed on the top of the 1× PBS-filled culture dish to minimize evaporation of the drop. After 72 hours in a hanging-drop culture, each aggregate was collected and transferred to each well of a ULA flat-bottom 96-well microplate (Corning). For experimental we used human melanocytes (CELL Applications). Three different concentrations tested (0.1%, 0.5% and 1%) of SGC100+ were diluted in a phenol red-free cell culture medium and was added to the cell culture medium when melanocyte aggregates were transferred to the well plate from hanging-drops at 72 hours after the cell seeding. Spheroids were exposed to depigmenting agents for 96 hours.
2.8. Measurement of Melanin Content
The culture medium in the well plate was read directly by a plate reader. We used a standard curve obtained from synthetic melanin (Sigma) dissolved in 1 N NH4OH, diluted in a culture medium. The absorbance was averaged from three wells, and each experiment was performed in triplicate. For comparison between samples, we used a depigmenting index (DI) as the percentage of decreased melanin in treated samples to an untreated control:
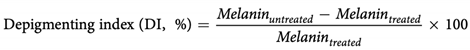
2.9. Statistical Analysis
The normality of the data was assessed using the Shapiro-Wilk test. After performing statistical analysis, one-way analysis of variance (ANOVA) was used to identify statistically significant differences among the various groups, followed by Tukey’s test for post-hoc analysis. The statistical software Past4Project (version 4.03) was utilized and the value of p < 0.01 (*) was considered statistically significant and p < 0.001 (**) was considered statistically highly significant. All data are presented as mean ± standard deviation (SD). Fluorescence intensity quantification of the acquired images was performed using Image J software (version 1.53).
3. Results and Discussion
3.1. Effects on Fibroblasts
Fibroblasts play a key role in skin elasticity by regulating the dermal microenvironment [19]. To evaluate the effect of SGC100+ on fibroblast growth, cells were treated with different concentrations (0.1%, 0.5% and 1%) and subsequently exposed to UV-C irradiation (Figure 1). In the absence of UV exposure, the tested concentrations significantly increased cell proliferation (178.4%, 210.9%, and 327.7% for the 0.1%, 0.5%, and 1% concentrations, respectively) (Figure 1). Even under UV irradiation, cells treated with SGC100+ maintained good proliferative activity (147.6%, 179.4%, and 247.3% for the 0.1%, 0.5%, and 1% concentrations, respectively) compared with the negative control, indicating a protective effect of the product (Figure 1). The expression of matrix metalloproteinase-2 (MMP2), an enzyme involved in collagen and ECM degradation, was then analyzed (Figure 2). Treatment with SGC100+ significantly reduced MMP2 expression levels, particularly at the 1% concentration. An increase in collagen production was also demonstrated in fibroblasts treated with SGC100+, particularly at a 1% concentration, compared to untreated controls (Figure 3). This further demonstrates SGC100+ components protective activity. Finally, the wound-scratch assay showed enhanced migratory capacity in HDFs treated with SGC100+ (Figure 4), in agreement with previous studies reporting a high content of proliferative and promigratory proteins in bovine colostrum [20]. Overall, these results indicate that SGC100+ promotes fibroblast survival, collagen production, and ECM reconstruction under UV-induced stress, thereby contributing to improved skin strength and elasticity. Dermal fibroblasts play a key role in maintaining the structural integrity of the skin, as they regulate the synthesis and remodeling of the extracellular matrix, including collagen and elastin, and dynamically respond to environmental stimuli such as UV radiation [19,21]. The dose-dependent increase in cell proliferation observed under basal conditions suggests that SGC100+ contains bioactive components capable of directly stimulating fibroblast growth mechanisms. Moreover, the ability of treated cells to maintain high proliferative activity even after UV-C exposure indicates a potential cytoprotective effect of the product, likely mediated by a reduction in oxidative damage and the activation of cellular survival pathways, attributable to the presence of various growth factors and bioactive peptides. Bovine colostrum containing bioactive peptides that activate the nuclear factor erytheroid-2-related factor 2 (Nrf2), a major regulator of redox balance, and plays a crucial role in the antioxidant defense system. Studies show colostrum components activate Nrf2 to increase antioxidant enzymes, protecting cells and tissues from damage, even suggesting therapeutic uses for Nrf2-activating peptides from colostrum [22,23]. Another relevant finding is the significant reduction in MMP2 expression following treatment with SGC100+, particularly at the 1% concentration. It is well established that UV irradiation induces the expression of matrix metalloproteinases (MMPs), thereby accelerating collagen degradation and contributing to photoaging. Therefore, the downregulation of MMP2 suggests a protective role of SGC100+ in preserving extracellular matrix integrity and counteracting UV-induced degenerative processes [24]. The wound-scratch assay results further demonstrate a significant increase in the migratory capacity of treated fibroblasts. Cell migration is an essential process for tissue repair and skin regeneration and is often impaired under conditions of oxidative stress and chronic inflammation [25]. The observed effect is consistent with previous studies reporting that bovine colostrum contains high levels of pro-proliferative and pro-migratory factors, such as IGF-1, TGF-β, and lactoferrin, which are known to promote tissue regeneration and wound healing [26]. Overall, these findings indicate that SGC100+ exerts a significant positive effect on the viability and functionality of human dermal fibroblasts. Indeed, it not only promotes fibroblast survival but also contributes to extracellular matrix reconstruction and the maintenance of dermal homeostasis under UV-induced stress conditions.
3.2. Effects on Keratinocytes
The proliferation of primary human keratinocytes (HEKa) was evaluated by MTT assay 24 hours after UV-C irradiation (4.5 mJ/cm²) and treatment with SGC100 at different concentrations (0.1%, 0.5%, and 1%). UV-C irradiation caused a significant reduction in cell viability compared to non-irradiated control cells, confirming the cytotoxic effect of UV-induced stress (Figure 5). Treatment with SGC100 showed a dose-dependent effect on cell proliferation. In particular, the 1% concentration resulted in a significant increase in cell viability compared with irradiated and with untreated control cells. Lower concentrations (0.1% and 0.5%) showed a weaker protective effect, but still an increase in proliferation compared with irradiated untreated cells (Figure 5). Overall, these data indicate that SGC100 can counteract the UV-C–induced reduction in cell growth, with the maximum effect observed at the 1% concentration. Intracellular oxidative stress was evaluated using the DCF-DA assay in HEKa keratinocytes irradiated with UV-C (0.7 mJ/cm²). UV-C irradiation induced a marked increase in DCF fluorescence, indicative of elevated production of reactive oxygen species (ROS), similarly to the positive control treated with hydrogen peroxide (Figure 6). Pre-treatment with SGC100 reduced intracellular ROS accumulation in a dose-dependent manner. In particular, the 1% concentration showed a marked reduction in DCF fluorescence, suggesting effective antioxidant activity and high cellular protection against UV-C–induced oxidative stress (Figure 6). Exposure to ultraviolet radiation represents one of the major stress factors for the skin, capable of impairing keratinocyte viability through the induction of cellular damage, oxidative stress, and alterations in proliferative mechanisms. In this study, UV-C irradiation caused a significant reduction in keratinocyte viability, confirming the high cytotoxicity of this type of radiation, which is widely documented in the literature for its ability to induce direct DNA damage and increase the production of reactive oxygen species (ROS) [27,28]. Treatment with SGC100 showed a clear dose-dependent protective effect on cell proliferation, with a significant recovery of viability particularly at the 1% concentration. The efficacy of SGC100+ is attributable to the synergistic action of bioactive molecules present in bovine colostrum, including growth factors, immunoglobulins, lactoferrin, and antioxidant peptides. The protective and regenerative effects of these biomolecules against UV-induced damage have been scientifically demonstrated, particularly in terms of reducing oxidative stress, enhancing DNA repair mechanisms, and promoting keratinocyte survival [29,30,31,32].
3.3. Effects on 3D Melanocyte Spheroids
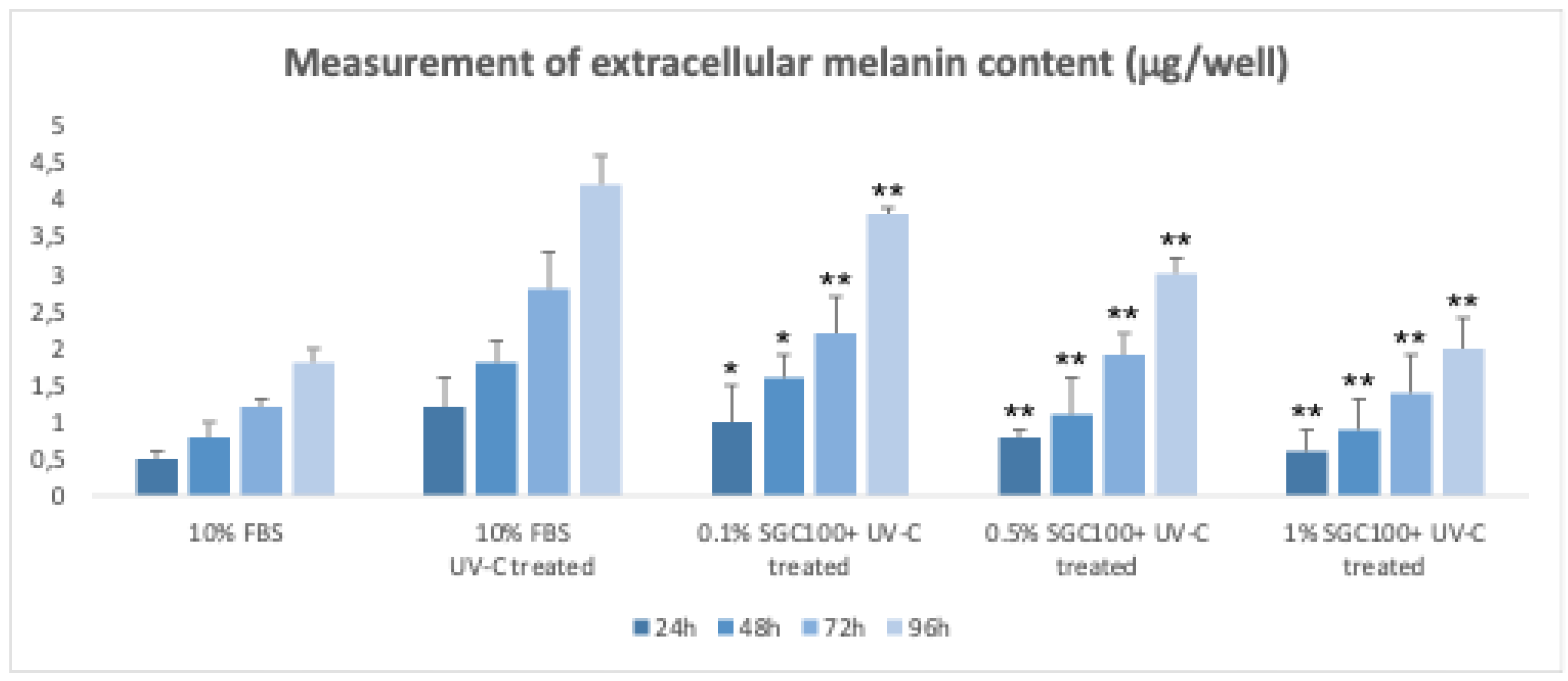
Murine B16F10 melanoma cells are commonly used as a model to study melanogenesis [18,31,33]. However, in two-dimensional (2D) cultures, these cells progressively lose their ability to produce melanin with continued passaging, making a pre-treatment with α-MSH necessary to induce pigmentation [18,31]. α-MSH stimulates melanogenesis by increasing intracellular cAMP levels but may also alter cellular status, for example by inhibiting cell proliferation [18,31]. To overcome these limitations, a three-dimensional (3D) culture model of B16 cells was developed using the hanging-drop method, which allows the spontaneous formation of cellular aggregates [18]. In this system, cells autonomously produce melanin without the need for α-MSH treatment [18]. After transferring the aggregates to ultralow attachment 96-well plates, melanin production and secretion progressively increase over time and are easily detectable by the naked eye in the culture medium. This makes the 3D model particularly suitable for non-invasive, high-throughput measurements of extracellular melanin. Using this system, the effects of different concentrations of SGC100+ on melanogenesis were evaluated. Treatment with 1% SGC100+ significantly reduced melanin production after UV stimulation (Figure 7). Moreover, SGC100+ demonstrated a protective effect against UV-induced damage by limiting the increase in melanin typically observed following irradiation (Figure 7). 3D cultures of B16 cells represent a more physiologically relevant model than 2D cultures, as they do not require exogenous stimuli for melanin production and allow effective evaluation of the anti-melanogenic and photoprotective activities of novel treatments such as SGC100+.
Melanin plays a central role in skin photoprotection by absorbing and dissipating ultraviolet (UV) radiation and limiting UV-induced oxidative stress. It is synthesized within melanocytes through a complex and tightly regulated process known as melanogenesis, which occurs in specialized organelles called melanosomes and involves key enzymes such as tyrosinase (TYR), tyrosinase-related protein 1 (TRP-1), and TRP-2/dopachrome tautomerase (DCT) [34,35]. At the molecular level, UV exposure stimulates melanogenesis primarily through activation of the melanocortin 1 receptor (MC1R) expressed on melanocytes. Binding of α-melanocyte-stimulating hormone (α-MSH) to MC1R triggers adenylate cyclase activation, resulting in increased intracellular cyclic AMP (cAMP) levels. This, in turn, activates protein kinase A (PKA), leading to phosphorylation of the transcription factor CREB and subsequent upregulation of microphthalmia-associated transcription factor (MITF), the master regulator of melanocyte differentiation and melanin synthesis [36,37,38]. Although melanin is essential for protecting the skin against UV damage, excessive or dysregulated melanin production can lead to hyperpigmentation disorders, including melasma and post-inflammatory hyperpigmentation [39]. In this context, three-dimensional (3D) melanocyte or melanoma culture models represent a more physiologically relevant system compared to conventional two-dimensional (2D) cultures, thanks to the absence of exogenous α-MSH stimulation. From a molecular perspective, compounds capable of reducing UV-induced melanogenesis may act through multiple mechanisms, including modulation of cAMP signaling, downregulation of MITF expression, inhibition of tyrosinase activity, or attenuation of oxidative stress via antioxidant effects [40,41]. In this regard, several studies have demonstrated that bioactive components present in colostrum can modulate the molecular pathways controlling pigmentation in vitro. Specifically, bovine colostrum extracts have been shown to reduce melanin production in α-MSH–stimulated B16 melanoma cells by regulating the MC1R–cAMP/CREB/MITF signaling axis, leading to decreased expression of key melanogenic enzymes such as tyrosinase and TRP-1.
4. Conclusions
In this study, we investigated the biological effects of Skin Glow Complex 100+ (SGC100+), a novel formulation containing bovine colostrum derived exosomes enriched with growth factors and cytokines, on key human skin cell types involved in skin structure, homeostasis, and pigmentation. Our findings demonstrate that SGC100+ exerts a pronounced pro-regenerative effect on dermal fibroblasts by promoting cell proliferation, enhancing collagen type I synthesis, reducing MMP-2 expression, and accelerating cell migration under both basal and UV-stressed conditions. These effects suggest a significant role of SGC100+ in preserving extracellular matrix integrity and counteracting the molecular mechanisms underlying photoaging, which are largely driven by UV-induced oxidative stress and matrix degradation [19,21,24]. The observed fibroblast responses are consistent with the presence of colostrum-derived bioactive factors known to activate antioxidant and cytoprotective pathways, including Nrf2 signaling, thereby reinforcing cellular resilience to environmental stressors [22,23]. Similarly, SGC100+ showed a protective and restorative effect on primary human keratinocytes exposed to UV-C radiation. Treatment significantly improved cell viability and markedly reduced intracellular ROS accumulation in a dose-dependent manner. These data highlight the antioxidant capacity of the formulation and its ability to mitigate UV-induced oxidative damage, a key driver of epidermal dysfunction, inflammation, and premature aging [27,28,29,30,31,32]. The combined stimulation of proliferation and reduction of oxidative stress underscores the potential of SGC100+ to support epidermal renewal and barrier function. Importantly, the use of a 3D melanocyte spheroid model allowed the evaluation of melanogenesis in a physiologically relevant system that does not require exogenous α-MSH stimulation. In this model, SGC100+ significantly reduced melanin production under both basal and UV-stimulated conditions, indicating a direct modulatory effect on melanocyte activity. Given the central role of UV-induced activation of the MC1R–cAMP–CREB–MITF axis in melanogenesis [36,37,38], the observed depigmenting effect may result from a combination of antioxidant activity and interference with key melanogenic signaling pathways, as previously reported for bovine colostrum–derived bioactive compounds [38,39]. These findings are particularly relevant in the context of acquired hyperpigmentation disorders, such as melasma and post-inflammatory hyperpigmentation, where excessive or dysregulated melanin production plays a pivotal pathogenic role [39]. Taken together, the results of this study support that SGC100+ acts is capable of simultaneously enhancing skin regeneration, protecting against UV-induced oxidative damage, remodeling the extracellular matrix, and modulating melanogenesis. The integration of exosomes, growth factors, and cytokines from bovine colostrum appears to confer synergistic biological effects across multiple skin compartments. Although further in vivo and clinical studies are required to confirm these findings, the present data provide strong mechanistic evidence supporting the potential application of SGC100+ in dermatological and cosmetic approaches aimed at skin regeneration, photo-protection, and the management of pigmentary disorders.
Author Contributions
“Conceptualization, M.V.B.; methodology, A.S. and M.V.B.; software, A.V.; validation, A.S. and M.V.B.; formal analysis, A.V.; data curation, M.V.B.; writing—original draft preparation, M.V.B.; writing—review and editing, A.S., A.V. and M.V.B.; supervision, M.V.B.; funding acquisition, M.V.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Byun, K. Melanogenesis Pathway in the Skin; Melanin, Kim, H.M., Eds.; Springer: Singapore, 2025; pp. 39–58. [Google Scholar]
- Yuan, X.H.; Jin, Z.H. Paracrine regulation of melanogenesis. Br J Dermatol 2018, 178(3), 632–639. [Google Scholar] [CrossRef]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New insights in photoaging, UVA induced damage and skin types. Exp Dermatol 2014, 23(1), 7–12. [Google Scholar] [CrossRef]
- Moumen, M.; McMichael, A.J. Atypical Dyschromia in Skin of Color. J Drugs Dermatol 2024, 23(2), 100–102. [Google Scholar] [CrossRef]
- Passeron, T.; Picardo, M. Melasma, a photoaging disorder. Pigment Cell Melanoma Res 2018, 31(4), 461–465. [Google Scholar] [CrossRef]
- Miao, F.; Wan, J.; Zhou, Y.; Shi, Y. Unraveling Melasma: From Epidermal Pigmentation to Microenvironmental Dysregulation. Biology 2025, 14, 1402. [Google Scholar] [CrossRef]
- Wawrzyńczak, A. Cosmetic and Pharmaceutic Products with Selected Natural and Synthetic Substances for Melasma Treatment and Methods of Their Analysis. Cosmetics 2023, 10, 86. [Google Scholar] [CrossRef]
- Maddaleno, A.S.; Camargo, J.; Mitjans, M.; Vinardell, M.P. Melanogenesis and Melasma Treatment. Cosmetics 2021, 8, 82. [Google Scholar] [CrossRef]
- Chaowattanapanit, S.; Silpa-Archa, N.; Kohli, I.; Lim, H.W.; Hamzavi, I. Postinflammatory hyperpigmentation: A comprehensive overview: Treatment options and prevention. J Am Acad Dermatol 2017, 77(4), 607–621. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Shieh, C.S.; Huang, T.L.; et al. Evaluating the Risk Factors of Post Inflammatory Hyperpigmentation Complications with Nd-YAG Laser Toning Using LASSO-Based Algorithm. Appl Sci 2020, 10, 2049. [Google Scholar] [CrossRef]
- Maghfour, J.; Olayinka, J.; Hamzavi, I.H.; Mohammad, T.F. A Focused review on the pathophysiology of post-inflammatory hyperpigmentation. Pigment Cell Melanoma Res 2022, 35(3), 320–327. [Google Scholar] [CrossRef]
- Shi, X.; Xia, X.; Xiao, Y.; et al. Increased melanin induces aberrant keratinocyte − melanocyte − basal − fibroblast cell communication and fibrogenesis by inducing iron overload and ferroptosis resistance in keloids. Cell Commun Signal 2025, 23, 141. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Fan, L.; et al. The Potential Application of Exosomes as Therapeutic Agents, Carriers, and Biomarkers in Skin Diseases. Int J Nanomedicine 2025, 20, 12627–12658. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367(6478), eaau6977. [Google Scholar] [CrossRef]
- Lei, L.; Zhou, S.; Zeng, L.; et al. Exosome-Based Therapeutics in Dermatology. Biomater Res 2025, 29, 0148. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.; Chisanga, D.; Liem, M.; et al. Bovine milk-derived exosomes from colostrum are enriched with proteins implicated in immune response and growth. Sci Rep 2017, 7(1), 5933. [Google Scholar] [CrossRef]
- Jabłońska, M.; Sawicki, T.; Żulewska, J.; Staniewska, K.; Łobacz, A.; Przybyłowicz, K.E. The Role of Bovine Milk-Derived Exosomes in Human Health and Disease. Molecules 2024, 29, 5835. [Google Scholar] [CrossRef]
- Chung, S.; Lim, G.J.; Lee, J.Y. Quantitative analysis of melanin content in a three-dimensional melanoma cell culture. Sci Rep 2019, 9, 780. [Google Scholar] [CrossRef]
- Boraldi, F.; Lofaro, F.D.; Bonacorsi, S.; Mazzilli, A.; Garcia-Fernandez, M.; Quaglino, D. The Role of Fibroblasts in Skin Homeostasis and Repair. Biomedicines 2024, 12(7), 1586. [Google Scholar] [CrossRef]
- Playford, R.J.; Cattell, M.; Marchbank, T. Marked variability in bioactivity between commercially available bovine colostrum for human use; implications for clinical trials. PLoS One 2020, 15(6), e0234719. [Google Scholar]
- Jipu, R.; Serban, I.L.; Goriuc, A.; Jipu, A.G.; Luchian, I.; Amititeloaie, C.; Tarniceriu, C.C.; Hurjui, I.; Butnaru, O.M.; Hurjui, L.L. Targeting Dermal Fibroblast Senescence: From Cellular Plasticity to Anti-Aging Therapies. Biomedicines 2025, 13, 1927. [Google Scholar] [CrossRef]
- Tonolo, F.; Folda, A.; Cesaro, L.; et al. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J Funct Foods 2020, 64, 103696. [Google Scholar] [CrossRef]
- Qin, D.D.; Hu, Z.; Han, S.; et al. Bioactive peptides alleviating oxidative stress-associated diseases by targeting the Nrf2 signaling pathway. Food Sci Hum Wellness 2025, 14, 9250253. [Google Scholar] [CrossRef]
- Kim, D.J.; Iwasaki, A.; Chien, A.L.; Kang, S. UVB-mediated DNA damage induces matrix metalloproteinases to promote photoaging in an AhR- and SP1-dependent manner. JCI Insight 2022, 7(9), e156344. [Google Scholar] [CrossRef]
- Radstake, W.E.; Gautam, K.; Van Rompay, C.; et al. Comparison of in vitro scratch wound assay experimental procedures. Biochem Biophys Rep 2023, 33, 101423. [Google Scholar] [CrossRef]
- Yalçıntaş, Y.M.; Duman, H.; López, J.M.M.; et al. Revealing the Potency of Growth Factors in Bovine Colostrum. Nutrients 2024, 16(14), 2359. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Photochem Photobiol 2015, 91(1), 140–55. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Richa Kumar, A.; Tyagi, M.B.; Sinha, R.P. Molecular mechanisms of ultraviolet radiation-induced DNA damage and repair. J Nucleic Acids 2010, 592980. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation - a review. Int J Cosmet Sci 2005, 27(1), 17–34. [Google Scholar] [CrossRef]
- Masaki, H. Role of antioxidants in the skin: anti-aging effects. J Dermatol Sci 2010, 58(2), 85–90. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Kim, H.; Kim, D.E.; et al. The Potential of Bovine Colostrum-Derived Exosomes to Repair Aged and Damaged Skin Cells. Pharmaceutics 2022, 14(2), 307. [Google Scholar] [CrossRef]
- Bolat, E.; Karagöz, Z.; Alves, J.L.d.B.; Neto, J.P.R.C.; Witkowska, A.M.; El-Seedi, H.; Lombardo, M.; Karav, S. The Potential Applications of Natural Colostrum in Skin Health. Cosmetics 2024, 11, 197. [Google Scholar] [CrossRef]
- Branquinho, M.S.; Silva, M.B.; Silva, J.C.; et al. A 2D and 3D melanogenesis model with human primary cells induced by tyrosine. J Biol Methods 2020, 7(3), e134. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol Rev 2004, 84(4), 1155–228. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J 1991, 5(14), 2902–9. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Ballotti, R. Cyclic AMP a Key Messenger in the Regulation of Skin Pigmentation. PCMR 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Levy, C.; Khaled, M.; Fisher, D.E. MITF: master regulator of melanocyte development and melanoma oncogene. Trends Mol Med 2006, 12(9), 406–14. [Google Scholar] [CrossRef]
- D'Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int J Mol Sci 2013, 14(6), 12222–48. [Google Scholar] [CrossRef]
- Passeron, T.; Krutmann, J.; Andersen, M.L.; Katta, R.; Zouboulis, C.C. Clinical and biological impact of the exposome on the skin. J Eur Acad Dermatol Venereol 2020, 34(4), 4–25. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: medicinal chemistry perspective of tyrosinase inhibitors. J Enzyme Inhib Med Chem 2017, 32(1), 403–425. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Guo, Z.; Tan, T.; Zhang, Y. Novel tyrosinase inhibitory peptide with free radical scavenging ability. J Enzyme Inhib Med Chem 2019, 34(1), 1633–1640. [Google Scholar] [CrossRef]
Figure 1.
HDFa cells survival at 24 h following treatment with and without UV-C exposure at three different concentrations of SGC100+ (0.1%, 0.5%, and 1%). Asterisks indicate the level of statistical highly significance compared to control samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated twice).
Figure 1.
HDFa cells survival at 24 h following treatment with and without UV-C exposure at three different concentrations of SGC100+ (0.1%, 0.5%, and 1%). Asterisks indicate the level of statistical highly significance compared to control samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated twice).
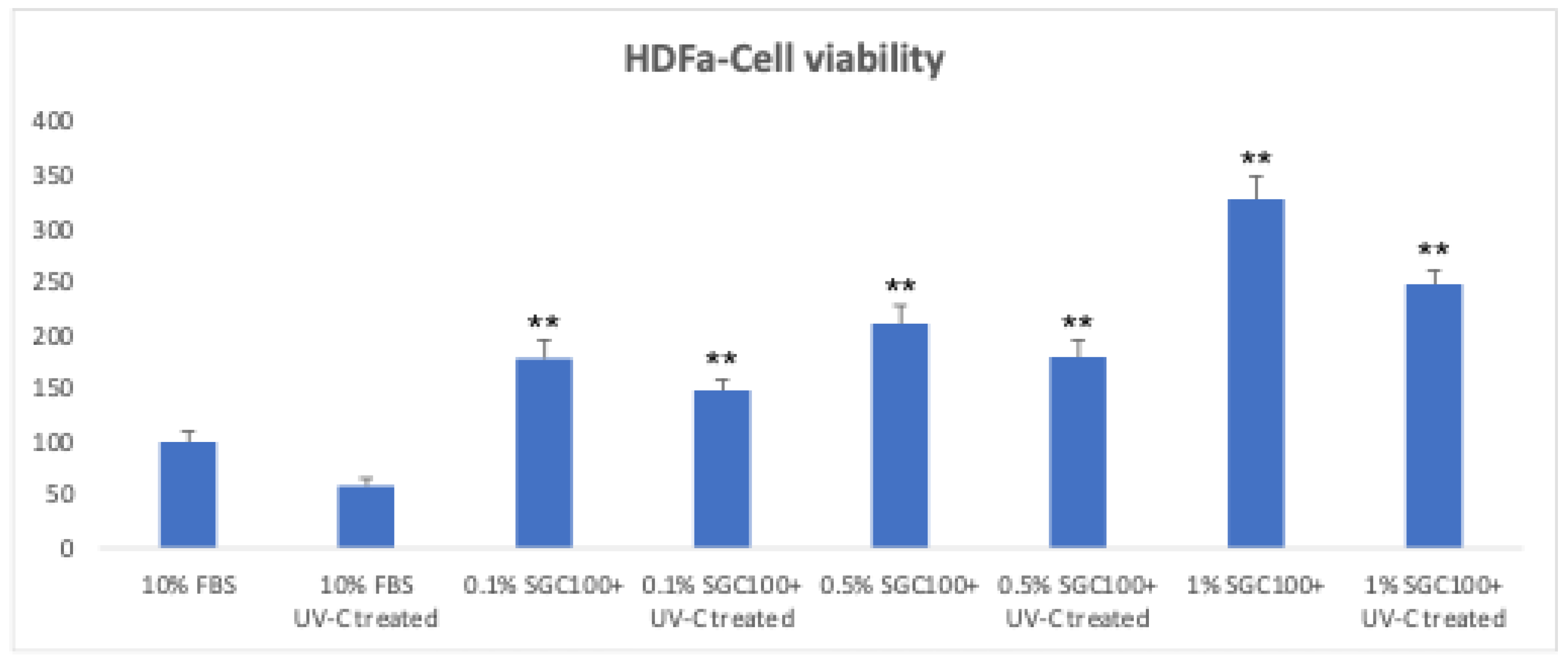
Figure 2.
Graphical representation of proenzyme MMP-2 production in HDFa cells. Data are presented as mean ± SEM. Vertical bars represent the standard error of the mean. Asterisks denote the level of statistical highly significance (p < 0.001).
Figure 2.
Graphical representation of proenzyme MMP-2 production in HDFa cells. Data are presented as mean ± SEM. Vertical bars represent the standard error of the mean. Asterisks denote the level of statistical highly significance (p < 0.001).
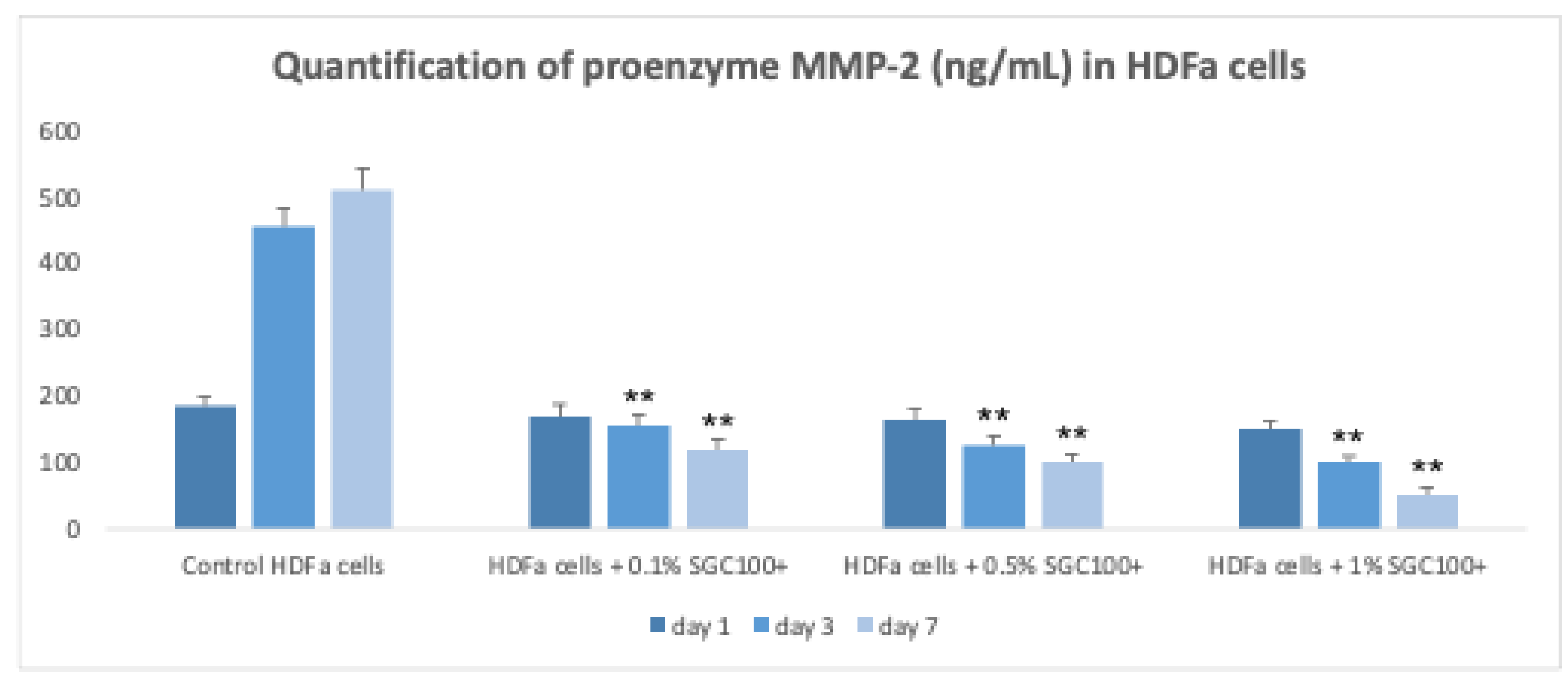
Figure 3.
Graphical representation of collagen production in HDFa cells. Data are plotted as mean ± SEM. Standard deviations were represented by vertical bars. Asterisks denote the degree of significance between results: * p < 0.01, ** p < 0.001.
Figure 3.
Graphical representation of collagen production in HDFa cells. Data are plotted as mean ± SEM. Standard deviations were represented by vertical bars. Asterisks denote the degree of significance between results: * p < 0.01, ** p < 0.001.
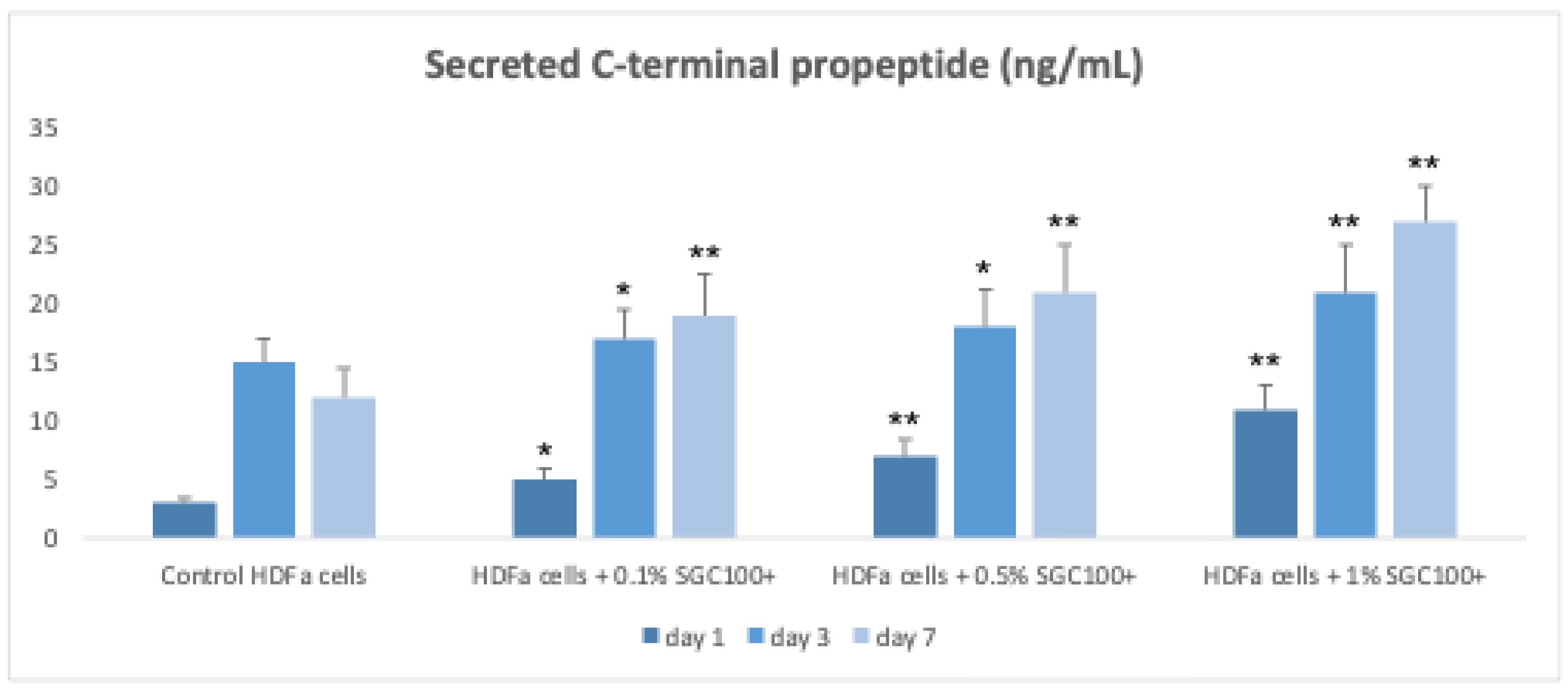
Figure 4.
Scratch assay on human fibroblasts treated with SGC100+ at three different concentrations for 24 hours. Untreated cells were used as the control (NC). Asterisks denote the degree of significance between treated and untreated samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated two times).
Figure 4.
Scratch assay on human fibroblasts treated with SGC100+ at three different concentrations for 24 hours. Untreated cells were used as the control (NC). Asterisks denote the degree of significance between treated and untreated samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated two times).
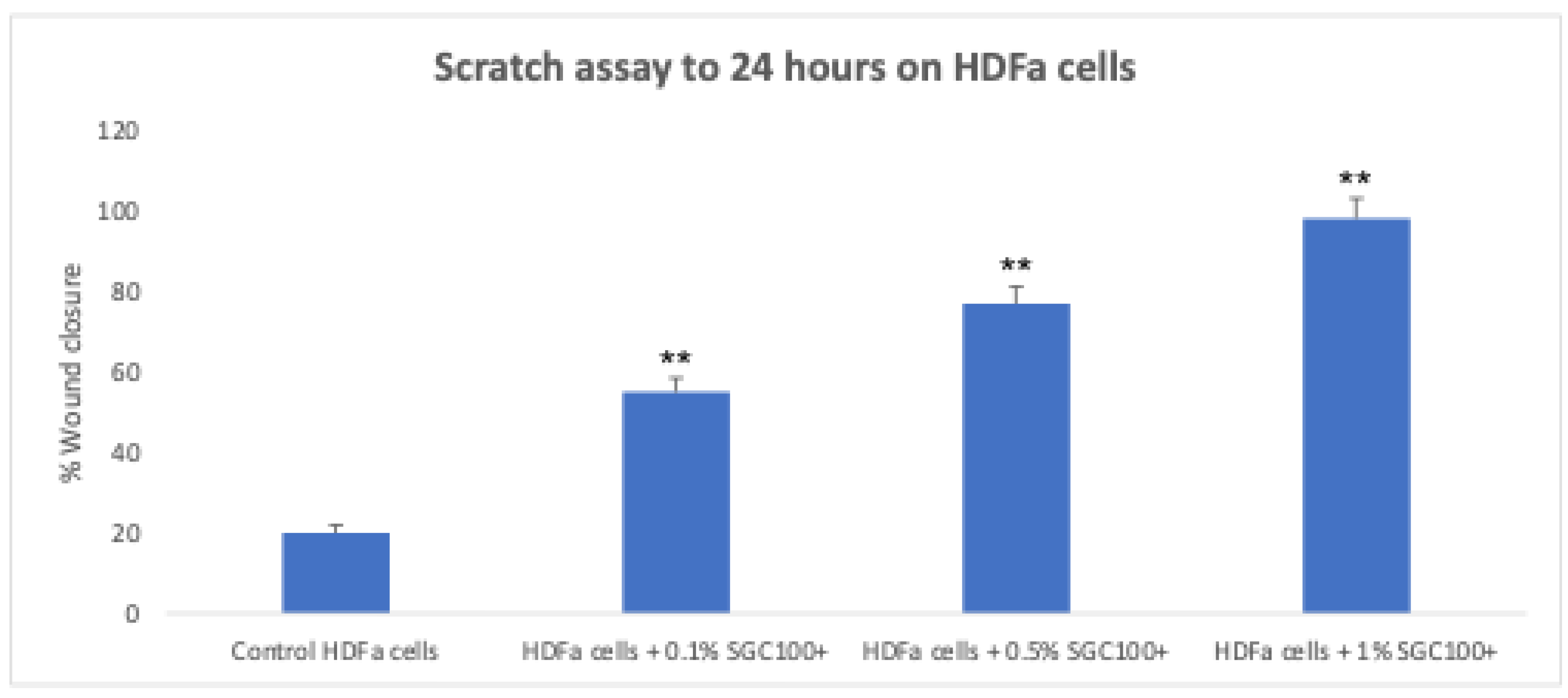
Figure 5.
HEKa cells survival at 24 h following treatment with and without UV-C exposure at three different concentrations of SGC100+ (0.1%, 0.5%, and 1%). Asterisks indicate the level of statistical highly significance compared to control samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated twice).
Figure 5.
HEKa cells survival at 24 h following treatment with and without UV-C exposure at three different concentrations of SGC100+ (0.1%, 0.5%, and 1%). Asterisks indicate the level of statistical highly significance compared to control samples (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated twice).
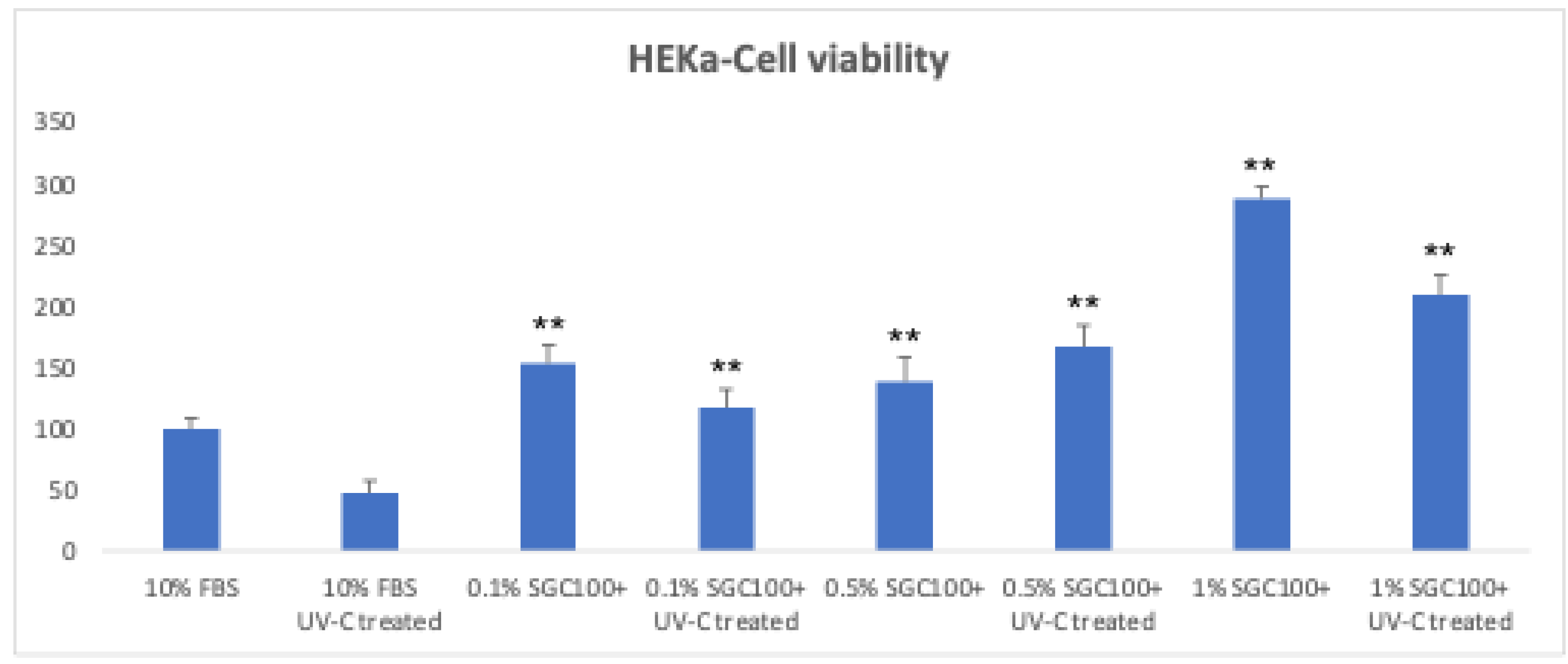
Figure 6.
Measured of intracellular ROS in HEKa. Asterisks denote the degree of significance between treated and untreated samples (negative control) (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated two times).
Figure 6.
Measured of intracellular ROS in HEKa. Asterisks denote the degree of significance between treated and untreated samples (negative control) (p < 0.001). Error bars represent the standard deviation of the mean (the experiment was repeated two times).
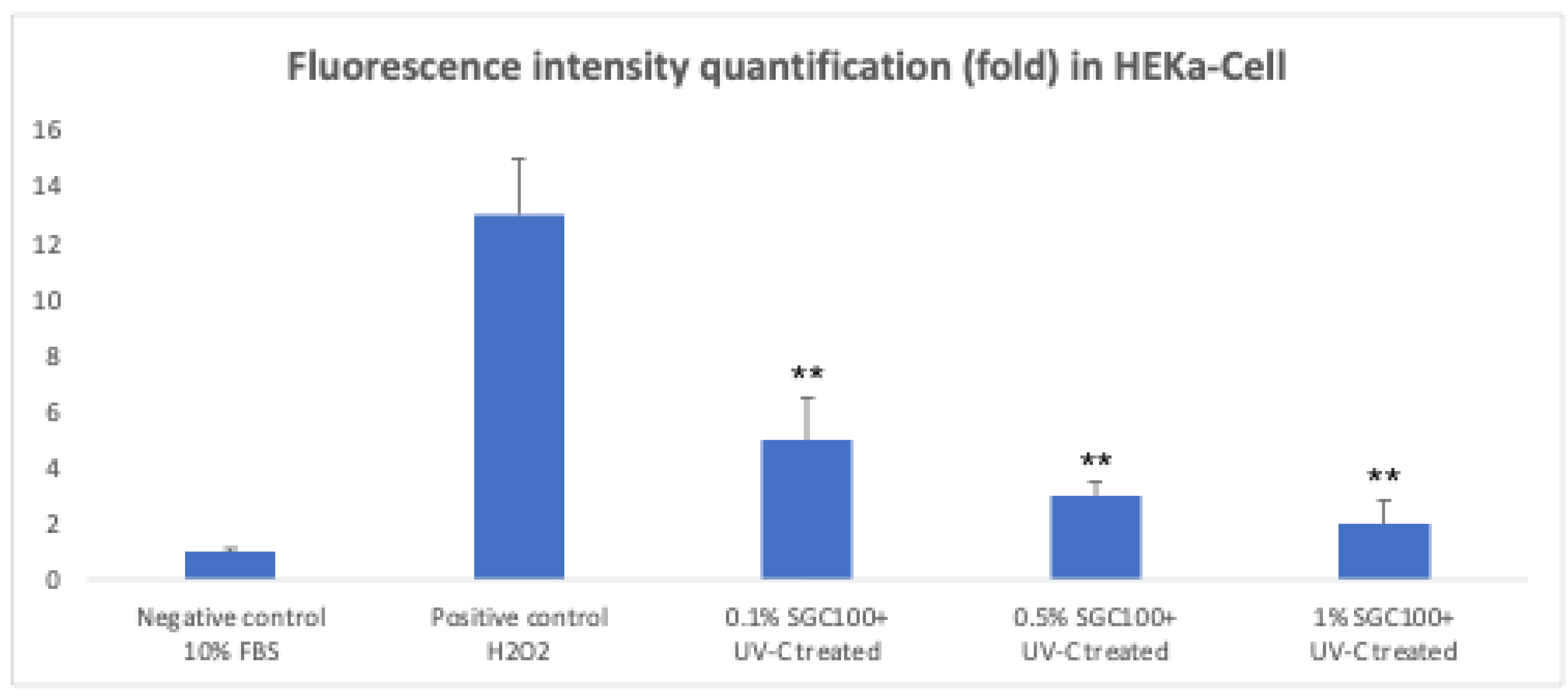
Figure 7.
Spontaneous melanin production in 3D cultures of melanocytes without the addition of melanin-synthesizing hormones. Asterisks denote the degree of significance between results: * p < 0.01, ** p < 0.001. Error bars represent the standard deviation of the mean (the experiment was repeated two times).
Figure 7.
Spontaneous melanin production in 3D cultures of melanocytes without the addition of melanin-synthesizing hormones. Asterisks denote the degree of significance between results: * p < 0.01, ** p < 0.001. Error bars represent the standard deviation of the mean (the experiment was repeated two times).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



