
Submitted:
08 January 2026
Posted:
09 January 2026
You are already at the latest version
Abstract
Background/Objectives: It is of upmost importance to study environmental bacteria, as these microorganisms remain poorly characterized regarding their diversity, antimicrobial resistance, and impact on the global ecosystem. This knowledge gap is particularly pronounced for marine bacteria. In this study, we aimed to isolate marine bacteria from different sources and to gain insights into the environmental bacterial resistome, an aspect that remains largely neglected. Methods: Bacteria were isolated from several marine sources using two different culture media, and their identification was based on 16S rRNA gene analysis. Whole-genome sequencing was performed for selected isolates belonging to novel taxa. Antimicrobial susceptibility to seven antibiotics was evaluated using the disk diffusion method. Results: A total of 171 bacterial isolates belonging to the phyla Pseudomonadota, Bacteroidota, Planctomycetota, Actinomycetota, and Bacillota were obtained from diverse marine samples. The most abundant group belonged to the class Alphaproteobacteria. Thirty isolates represented novel taxa, comprising 16 new species and one new genus. Despite the challenges associated with determining antibiotic resistance profiles in environmental bacteria, only one isolate (1.8%) was pan-susceptible, whereas 54 (98.2%) showed resistance to at least one of the tested antibiotics. Moreover, 33 isolates exhibited a multidrug-resistant phenotype. Genome analysis of four novel taxa revealed the presence of an incomplete AdeFGH efflux pump. Conclusions: This study highlights the high bacterial diversity in marine environments, the striking prevalence of antibiotic resistance, and the major methodological challenges in studying environmental bacteria. Importantly, it emphasizes the relevance of culturomics-based approaches for uncovering hidden microbial diversity and characterizing environmental resistomes.
Keywords:
1. Introduction
2. Results and Discussion
2.1. Bacterial Isolation from Marine Samples
2.2. Taxonomic Affiliation of the Isolates
2.3. Novel Taxa
2.4. Environmental Resistome and One Health Perspective
2.4.1. Challenges in Antimicrobial Resistance Testing of Environmental Isolates
2.4.2. Phenotypic Resistance Profiles
2.4.3. Genomic Analysis of AMR Genes
4. Materials and Methods
4.1. Sampling
4.2. Media and Culture Conditions
4.3. Sample Processing
4.4. Bacterial Identification by Molecular Analysis
4.5. Genome Sequencing, Assembly, and Analysis
4.6. Antibiotic Susceptibility Testing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Grosberg, R.K.; Vermeij, G.J.; Wainwright, P.C. Biodiversity in water and on land. Curr. Biol. 2012, 22, R900–R903. [Google Scholar] [CrossRef]
- Logares, R. Decoding populations in the ocean microbiome. Microbiome 2024, 12, 67. [Google Scholar] [CrossRef]
- Salazar, G.; Sunagawa, S. Marine microbial diversity. Curr. Biol. 2017, 27, R489–R494. [Google Scholar] [CrossRef] [PubMed]
- Zengler, K. Central role of the cell in microbial ecology. Microbiol. Mol. Biol. Rev. 2009, 73, 712–729. [Google Scholar] [CrossRef]
- Rodrigues, C.J.C.; de Carvalho, C.C.C.R. Cultivating marine bacteria under laboratory conditions: Overcoming the “unculturable” dogma. Front. Bioeng. Biotechnol. 2022, 10, 964589. [Google Scholar] [CrossRef] [PubMed]
- Asensio-Montesinos, F.; Molina, R.; Anfuso, G.; Manno, G.; Lo Re, C. Natural and Human Impacts on Coastal Areas. J. Mar. Sci. Eng. 2024, 12. [Google Scholar] [CrossRef]
- Glibert, P.M.; Burkholder, J.M. Causes of harmful algal blooms. In Harmful Algal Blooms: A Compendium Desk Reference; Shumway, S.A., Burkholder, J.M., Morton, S.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; pp. 1–39. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Li, X.-J.; Meng, N.; Yang, G.-P.; Zhang, G.; Zhuang, G.-C. Regulation and response of heterotrophic bacterial production to environmental changes in marginal seas of the Western Pacific Ocean. Glob. Planet. Change 2025, 245, 104678. [Google Scholar] [CrossRef]
- Castaño-Ortiz, J.M.; Serra-Compte, A.; Álvarez-Muñoz, D.; Barceló, D.; Rodríguez-Mozaz, S. Antibiotics in the marine environment: Occurrence and impact. In Contaminants of Emerging Concern in the Marine Environment: Current Challenges in Marine Pollution; Barceló, D., Ed.; Academic Press : Cambridge, MA, USA, 2023; pp. 73–104. [Google Scholar] [CrossRef]
- Selvarajan, R.; Obize, C.; Sibanda, T.; Abia, A.L.K.; Long, H. Evolution and emergence of antibiotic resistance in given ecosystems: Possible strategies for addressing the challenge of antibiotic resistance. Antibiotics 2022, 12(1), 28. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Antibiotic Resistance: Moving From Individual Health Norms to Social Norms in One Health and Global Health. Front. Microbiol. 2020, 11, 1914. [Google Scholar] [CrossRef]
- Muloi, D.M.; Jauneikaite, E.; Anjum, M.F.; Essack, S.Y.; Singleton, D.A.; et al. Exploiting genomics for antimicrobial resistance surveillance at One Health interfaces. Lancet Microbe 2023, 4, e1056–e1062. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.; Rahman, M.H.; Al-Surrayai, T.; Al-Dhumair, A.; Al-Hasan, M. A One-Health perspective of antimicrobial resistance (AMR): Human, animals and environmental health. Life 2025, 15, 1598. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antibiotic resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- Bonanno Ferraro, G.; Brandtner, D.; Franco, A.; Iaconelli, M.; Mancini, P.; et al. Global quantification and distribution of antibiotic resistance genes in oceans and seas: Anthropogenic impacts and regional variability. Sci. Total Environ. 2024, 955, 176765. [Google Scholar] [CrossRef]
- Sun, J.; Li, S.; Fan, C.; Cui, K.; Tan, H.; Qiao, L.; Lu, L. N-Acetylglucosamine promotes tomato plant growth by shaping the community structure and metabolism of the rhizosphere microbiome. Microbiol. Spectr. 2022, 10(3), e00358-22. [Google Scholar] [CrossRef]
- Kämpfer, P.; Falsen, E.; Lodders, N.; Langer, S.; Busse, H.J.; Schumann, P. Ornithinibacillus contaminans sp. nov., an endospore-forming species. Int. J. Syst. Evol. Microbiol. 2010, 60, 2930–2934. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-G.; Choi, D.H.; Hyun, S.; Cho, B.C. Oceanobacillus profundus sp. nov., isolated from a deep-sea sediment core. Int. J. Syst. Evol. Microbiol. 2007, 57, 409–413. [Google Scholar] [CrossRef]
- Paul, S.I.; Rahman, M.M.; Salam, M.A.; Khan, M.A.R.; Islam, M.T. Identification of marine sponge-associated bacteria of the Saint Martin’s island of the Bay of Bengal emphasizing on the prevention of motile Aeromonas septicemia in Labeo rohita. Aquaculture 2021, 545, 737156. [Google Scholar] [CrossRef]
- Deng, Y.; Mou, T.; Wang, J.; Su, J.; Yan, Y.; Zhang, Y.Q. Characterization of three rapidly growing novel Mycobacterium species with significant polycyclic aromatic hydrocarbon bioremediation potential. Front. Microbiol. 2023, 14, 1225746. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Jia, Y.; Ruth, N.; Qiao, C.; Wang, J.; Zhao, B.; Yan, Y. Biodegradation of phthalic acid esters by a newly isolated Mycobacterium sp. YC-RL4 and the bioprocess with environmental samples. Environ. Sci. Pollut. Res. 2016, 23, 16609–16619. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, P.; Karlson, U.; Stackebrandt, E.; Kroppenstedt, R.M. Mycobacterium frederiksbergense sp. nov., a novel polycyclic aromatic hydrocarbon-degrading Mycobacterium species. Int. J. Syst. Evol. Microbiol. 2001, 51, 1715–1722. [Google Scholar] [CrossRef]
- Kim, E.J.; Fathoni, A.; Jeong, G.T.; Jeong, H.D.; Nam, T.J.; Kong, I.S.; Kim, J.K. Microbacterium oxydans, a novel alginate- and laminarin-degrading bacterium for the reutilization of brown-seaweed waste. J. Environ. Manag. 2013, 130, 153–159. [Google Scholar] [CrossRef]
- Lenchi, N.; Anzil, A.; Servais, P.; Kebbouche-Gana, S.; Gana, M.L.; Llirós, M. Microbacterium algeriense sp. nov., a novel actinobacterium isolated from Algerian oil production waters. Int. J. Syst. Evol. Microbiol. 2020, 70(12), 6044–6051. [Google Scholar] [CrossRef]
- Vitorino, I.R.; Lage, O.M. The Planctomycetia: An overview of the currently largest class within the phylum Planctomycetes. Antonie van Leeuwenhoek 2022, 115, 169–201. [Google Scholar] [CrossRef]
- Venter, J.C.; Remington, K.; Heidelberg, J.F.; Halpern, A.L.; Rusch, D.; Eisen, J.A.; Wu, D.; Paulsen, I.; Nelson, K.E.; Nelson, W.; Fouts, D.E.; Levy, S.; Knap, A.H.; Lomas, M.W.; Nealson, K.; White, O.; Peterson, J.; Hoffman, J.; Parsons, R.; Baden-Tillson, H.; Pfannkoch, C.; Rogers, Y.H.; Smith, H.O. Environmental genome shotgun sequencing of the Sargasso Sea. Science 2004, 304, 66–74. [Google Scholar] [CrossRef]
- Yilmaz, P.; Yarza, P.; Rapp, J.Z.; Glöckner, F.O. Expanding the world of marine bacterial and archaeal clades. Front. Microbiol. 2016, 6, 1524. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Xia, X.; Suzuki, K.; Liu, H. Vertical profiles of bacteria in the tropical and subarctic oceans revealed by pyrosequencing. PLoS ONE 2013, 8, e79423. [Google Scholar] [CrossRef] [PubMed]
- Heip, C.; Hummel, H.; van Avesaath, P.; Appeltans, W.; Arvanitidis, C.; Aspden, R.; Austen, M.; Boero, F.; Bouma, T.J.; Boxshall, G.; Buchholz, R.; Crowe, T.; Delaney, A.; Deprez, T.; Emblow, C.; Féral, J.-P.; Gasol, J.; Nash, R. Marine Biodiversity and Ecosystem Functioning; Nash, R., Ed.; MarBEF Project: [s.l.], 2009. [Google Scholar] [CrossRef]
- Wiese, J.; Imhoff, J.F.; Horn, H.; Borchert, E.; Kyrpides, N.C.; Göker, M.; Klenk, H.P.; Woyke, T.; Hentschel, U. Genome analysis of the marine bacterium Kiloniella laminariae and first insights into comparative genomics with related Kiloniella species. Arch. Microbiol. 2020, 202, 815–824. [Google Scholar] [CrossRef]
- Montana, S.; Schramm, S.T.; Traglia, G.M.; Chiem, K.; Parmeciano Di Noto, G.; Almuzara, M.; Barberis, C.; Vay, C.; Quiroga, C.; Tolmasky, M.E.; et al. The genetic analysis of an Acinetobacter johnsonii clinical strain evidenced the presence of horizontal genetic transfer. PLoS ONE 2016, 11, e0161528. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Shah, J.; Ozongwu, C.; Pike, R. Incidence of Acinetobacter species other than A. baumannii among clinical isolates of Acinetobacter: evidence for emerging species. J. Clin. Microbiol. 2010, 48, 1445–1449. [Google Scholar] [CrossRef]
- van den Broek, P.J.; van der Reijden, T.J.; van Strijen, E.; Helmig-Schurter, A.V.; Bernards, A.T.; Dijkshoorn, L. Endemic and epidemic Acinetobacter species in a university hospital: an 8-year survey. J. Clin. Microbiol. 2009, 47, 3593–3599. [Google Scholar] [CrossRef]
- Yumoto, T.; Kono, Y.; Kawano, S.; Kamoi, C.; Iida, A.; Nose, M.; Sato, K.; Ugawa, T.; Okada, H.; Ujike, Y.; et al. Citrobacter braakii bacteremia-induced septic shock after colonoscopy preparation with polyethylene glycol in a critically ill patient: a case report. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 22. [Google Scholar] [CrossRef]
- Oyeka, M.; Antony, S. Citrobacter braakii bacteremia: case report and review of the literature. Infect. Disord. Drug Targets 2017, 17, 59–63. [Google Scholar] [CrossRef]
- Hirai, J.; Uechi, K.; Hagihara, M.; Sakanashi, D.; Kinjo, T.; Haranaga, S.; Fujita, J. Bacteremia due to Citrobacter braakii: a case report and literature review. J. Infect. Chemother. 2016, 22, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, L.; Zhou, H.; Yuan, M.; Hu, D.; Wang, Y.; Sun, H.; Xu, J.; Lan, R. Antimicrobial resistance and molecular characterization of Citrobacter spp. causing extraintestinal infections. Front. Cell. Infect. Microbiol. 2021, 11, 737636. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, D.; Liu, L.; Lan, R.; Hao, S.; Jin, W.; Sun, H.; Wang, Y.; Liang, Y.; Xu, J. Genetic diversity, multidrug resistance, and virulence of Citrobacter freundii from diarrheal patients and healthy individuals. Front. Cell. Infect. Microbiol. 2018, 8, 233. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.H.; Wang, N.Y.; Wu, A.Y.; Lin, C.C.; Lee, C.M.; Liu, C.P. Citrobacter freundii bacteremia: risk factors of mortality and prevalence of resistance genes. J. Microbiol. Immunol. Infect. 2018, 51, 565–572. [Google Scholar] [CrossRef]
- Miyoshi, S. Extracellular proteolytic enzymes produced by human pathogenic Vibrio species. Front. Microbiol. 2013, 4, 339. [Google Scholar] [CrossRef]
- Liu, R.; Qiu, L.; Yu, Z.; Zi, J.; Yue, F.; Wang, L.; Zhang, H.; Teng, W.; Liu, X.; Song, L. Identification and characterisation of pathogenic Vibrio splendidus from Yesso scallop (Patinopecten yessoensis) cultured in a low temperature environment. J. Invertebr. Pathol. 2013, 114, 144–150. [Google Scholar] [CrossRef]
- Jun, L.; Woo, N.Y.S. Pathogenicity of Vibrio in fish: an overview. J. Ocean Univ. Qingdao 2003, 2, 117–128. [Google Scholar] [CrossRef]
- Lv, H.; Sahin, N.; Tani, A. Methylotenera oryzisoli sp. nov., a lanthanide-dependent methylotrophic bacterium isolated from rice field soil. Int. J. Syst. Evol. Microbiol. 2020, 70, 2713–2718. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzeby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Dai, X. High salt cross-protects Escherichia coli from antibiotic treatment through increasing efflux pump expression. mSphere 2018, 3, e00095-18. [Google Scholar] [CrossRef]
- Godinho, O.; Calisto, R.; Øvreås, L.; Quinteira, S.; Lage, O.M. Antibiotic susceptibility of marine Planctomycetes. Antonie Van Leeuwenhoek 2019, 112, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Sharma, G.; Srinivas, T.N.; Anil Kumar, P. Aliiglaciecola coringensis sp. nov., isolated from a water sample collected from mangrove forest in Coringa, Andhra Pradesh, India. Antonie Van Leeuwenhoek 2014, 106, 1097–1103. [Google Scholar] [CrossRef]
- Yoo, J.H.; Han, J.E.; Lee, J.Y.; Jeong, S.W.; Jeong, Y.S.; Lee, J.Y.; Lee, S.Y.; Sung, H.; Tak, E.J.; Kim, H.S.; Kim, P.S.; Choi, J.W.; Kim, D.Y.; Jeong, I.C.; Gim, D.H.; Kang, S.M.; Bae, J.W. Parasphingorhabdus cellanae sp. nov., isolated from the gut of a Korean limpet, Cellana toreuma. Int. J. Syst. Evol. Microbiol. 2022, 72, 5470. [Google Scholar] [CrossRef]
- Han, X.Y.; Andrade, R.A. Brevundimonas diminuta infections and its resistance to fluoroquinolones. J. Antimicrob. Chemother. 2005, 55, 853–859. [Google Scholar] [CrossRef]
- World Health Organization. WHO medically important antimicrobials list (WHO MIA List) for human medicine; World Health Organization: Geneva, 2024; Available online: https://www.who.int/news/item/08-02-2024-who-medically-important-antimicrobial-list-2024.
- Wareth, G.; Dadar, M.; Ali, H.; Hamdy, M.E.R.; Al-Talhy, A.M.; Elkharsawi, A.R.; Tawab, A.A.A.E.; Neubauer, H. The perspective of antibiotic therapeutic challenges of brucellosis in the Middle East and North African countries: current situation and therapeutic management. Transbound. Emerg. Dis. 2022, 69, e1253–e1268. [Google Scholar] [CrossRef] [PubMed]
- Zeb, S.; Yasmin, H.; Malik, I.R.; et al. Antimicrobial resistant Brucella spp. prevail in raw milk and animal feces of different livestock farms. BMC Microbiol. 2025, 25, 231. [Google Scholar] [CrossRef]
- Tang, T.; Sun, X.; Dong, Y.; Liu, Q. Erythrobacter aureus sp. nov., a plant growth-promoting bacterium isolated from sediment in the Yellow Sea, China. 3 Biotech 2019, 9, 430. [Google Scholar] [CrossRef]
- Silva, S.G.; Paula, P.; da Silva, J.P.; Mil-Homens, D.; Teixeira, M.C.; Fialho, A.M.; Costa, R.; Keller-Costa, T. Insights into the antimicrobial activities and metabolomes of Aquimarina (Flavobacteriaceae, Bacteroidetes) species from the rare marine biosphere. Mar. Drugs 2022, 20, 423. [Google Scholar] [CrossRef]
- Dieterich, C.L.; Probst, S.I.; Ueoka, R.; Sandu, I.; Schäfle, D.; Molin, M.D.; Minas, H.A.; Costa, R.; Oxenius, A.; Sander, P.; Piel, J. Aquimarins, peptide antibiotics with amino-modified C-termini from a sponge-derived Aquimarina sp. bacterium. Angew. Chem. Int. Ed. 2022, 61, e202115802. [Google Scholar] [CrossRef]
- Offret, C.; Desriac, F.; Le Chevalier, P.; Mounier, J.; Jégou, C.; Fleury, Y. Spotlight on antimicrobial metabolites from the marine bacteria Pseudoalteromonas: chemodiversity and ecological significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef] [PubMed]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Perichon, B. Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Poirel, L.; Vuillemin, X.; Kieffer, N.; Mueller, L.; Descombes, M.C.; Nordmann, P. Identification of FosA8, a plasmid-encoded fosfomycin resistance determinant from Escherichia coli, and its origin in Leclercia adecarboxylata. Antimicrob. Agents Chemother. 2019, 63, e01403-19. [Google Scholar] [CrossRef] [PubMed]
- Godinho, O.; Klimek, D.; Jackiewicz, A.; Guedes, B.; Almeida, E.; Calisto, R.; Vitorino, I.R.; Santos, J.D.N.; González, I.; Lobo-da-Cunha, A.; Calusinska, M.; Quinteira, S.; Lage, O.M. Stieleria tagensis sp. nov., a novel member of the phylum Planctomycetota isolated from Tagus River in Portugal. Antonie Van Leeuwenhoek 2023, 116, 1209–1225. [Google Scholar] [CrossRef]
- Beaumont, N.J.; Austen, M.C.; Atkins, J.P.; Burdon, D.; Degraer, S.; Dentinho, T.P.; Derous, S.; Holm, P.; Horton, T.; van Ierland, E.; et al. Identification, definition and quantification of goods and services provided by marine biodiversity: implications for the ecosystem approach. Mar. Pollut. Bull. 2007, 54, 253–265. [Google Scholar] [CrossRef]
- Vignaroli, C.; Pasquaroli, S.; Citterio, B.; Di Cesare, A.; Mangiaterra, G.; Fattorini, D.; Biavasco, F. Antibiotic and heavy metal resistance in enterococci from coastal marine sediment. Environ. Pollut. 2018, 237, 406–413. [Google Scholar] [CrossRef]
- Vignaroli, C.; Luna, G.M.; Pasquaroli, S.; Di Cesare, A.; Petruzzella, R.; Paroncini, P.; Biavasco, F. Epidemic Escherichia coli ST131 and Enterococcus faecium ST17 in coastal marine sediments from an Italian beach. Environ. Sci. Technol. 2013, 47, 13772–13780. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Sellera, F.P.; Moura, Q.; Esposito, F.; Sabino, C.P.; Lincopan, N. Identification and genomic features of halotolerant extended-spectrum-beta-lactamase (CTX-M)-producing Escherichia coli in urban-impacted coastal waters, Southeast Brazil. Mar. Pollut. Bull. 2020, 150, 110689. [Google Scholar] [CrossRef]
- Alves, M.S.; Pereira, A.; Araujo, S.M.; Castro, B.B.; Correia, A.C.; Henriques, I. Seawater is a reservoir of multi-resistant Escherichia coli, including strains hosting plasmid-mediated quinolones resistance and extended-spectrum beta-lactamases genes. Front. Microbiol. 2014, 5, 426. [Google Scholar] [CrossRef]
- Al-Bahry, S.N.; Mahmoud, I.Y.; Al-Belushi, K.I.; Elshafie, A.E.; Al-Harthy, A.; Bakheit, C.K. Coastal sewage discharge and its impact on fish with reference to antibiotic resistant enteric bacteria and enteric pathogens as bio-indicators of pollution. Chemosphere 2009, 77, 1534–1539. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J. Planctomycetes diversity associated with macroalgae. FEMS Microbiol. Ecol. 2011, 78, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Lage, O.M.; Bondoso, J. Bringing Planctomycetes into pure culture. Front. Microbiol. 2012, 3, 405. [Google Scholar] [CrossRef] [PubMed]
- Lage, O.M.; Bondoso, J.; Viana, F. Isolation and characterization of Planctomycetes from the sediments of a fish farm wastewater treatment tank. Arch. Microbiol. 2012, 194, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, I.; Albuquerque, L.; Wiegand, S.; Kallscheuer, N.; da Costa, M.S.; Lobo-da-Cunha, A.; Jogler, C.; Lage, O.M. Alienimonas chondri sp. nov., a novel planctomycete isolated from the biofilm of the red alga Chondrus crispus. Syst. Appl. Microbiol. 2020, 43, 126083. [Google Scholar] [CrossRef] [PubMed]
- EUCAST; European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters, Version 10.0; EUCAST: Växjö, Sweden, 2020; Available online: https://www.eucast.org/clinical_breakpoints/.
- Girlich, D.; Poirel, L.; Nordmann, P. First isolation of the blaOXA-23 carbapenemase gene from an environmental Acinetobacter baumannii isolate. Antimicrob. Agents Chemother. 2010, 54, 578–579. [Google Scholar] [CrossRef]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes strains selected on ciprofloxacin or the disinfectant benzalkonium chloride exhibit reduced susceptibility to ciprofloxacin, gentamicin, benzalkonium chloride, and other toxic compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef]
- Cohen-Bazire, G.; Sistrom, W.R.; Stanier, R.Y. Kinetic studies of pigment synthesis by non-sulphur purple bacteria. J. Cell Comp. Physiol. 1957, 49, 25–68. [Google Scholar] [CrossRef] [PubMed]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Parte, A.C. LPSN—List of Prokaryotic Names with Standing in Nomenclature (bacterio.net), 20 years on. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; Edalatmand, A.; Petkau, A.; Syed, S.A.; Tsang, K.K.; et al. CARD 2023: expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- CLSI (Clinical and Laboratory Standards Institute). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100: Wayne, PA, USA, 2020. [Google Scholar]
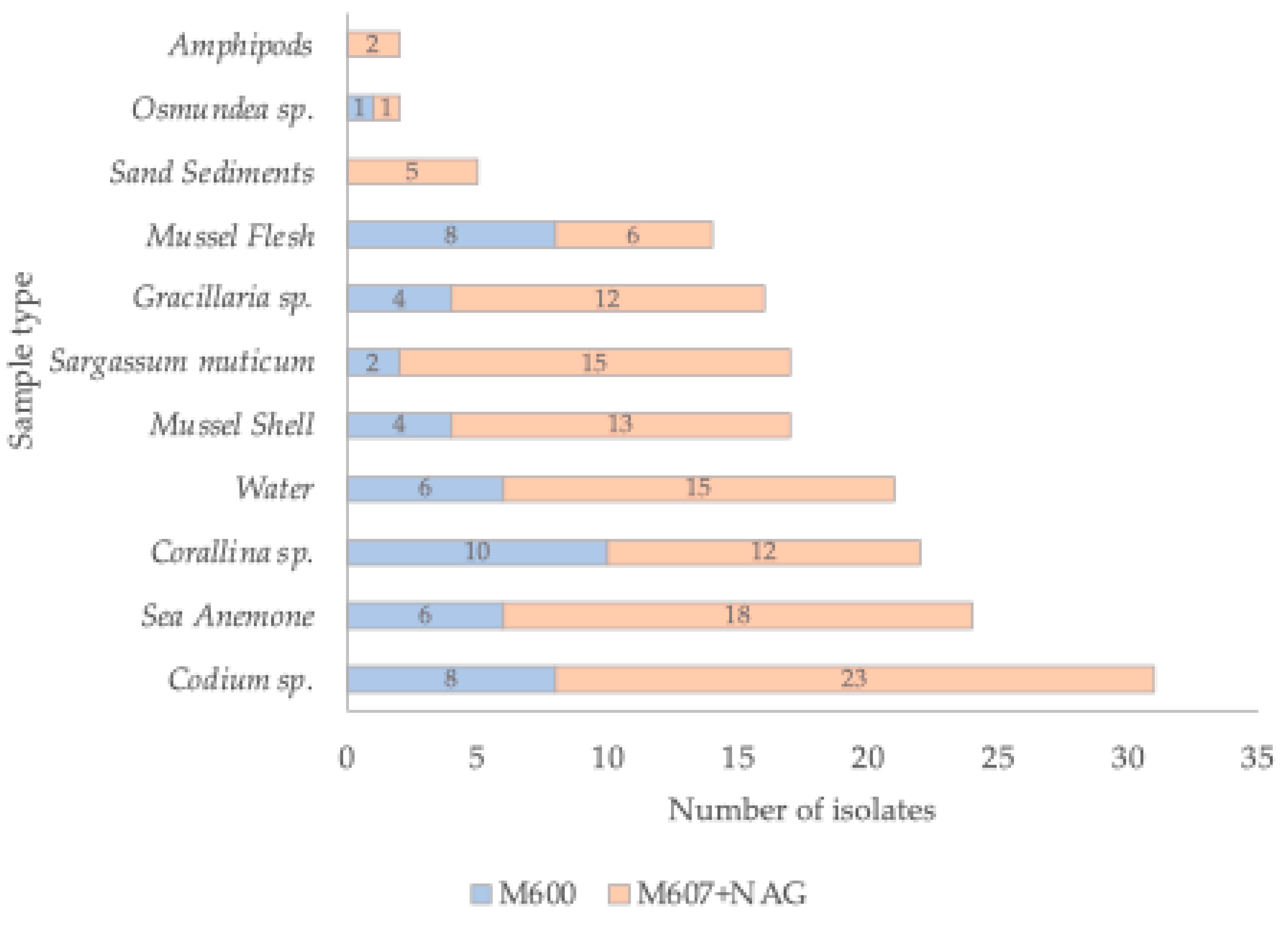
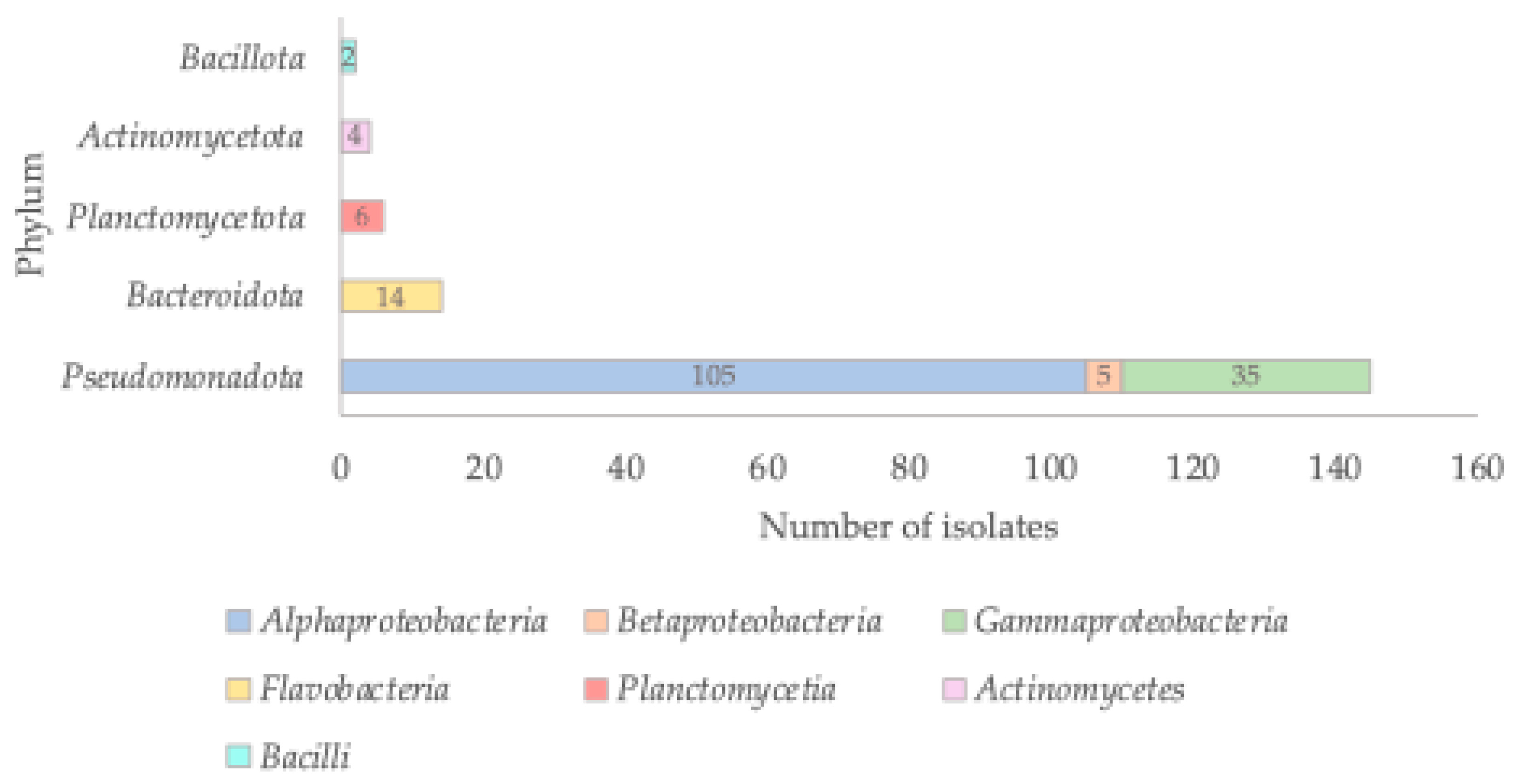
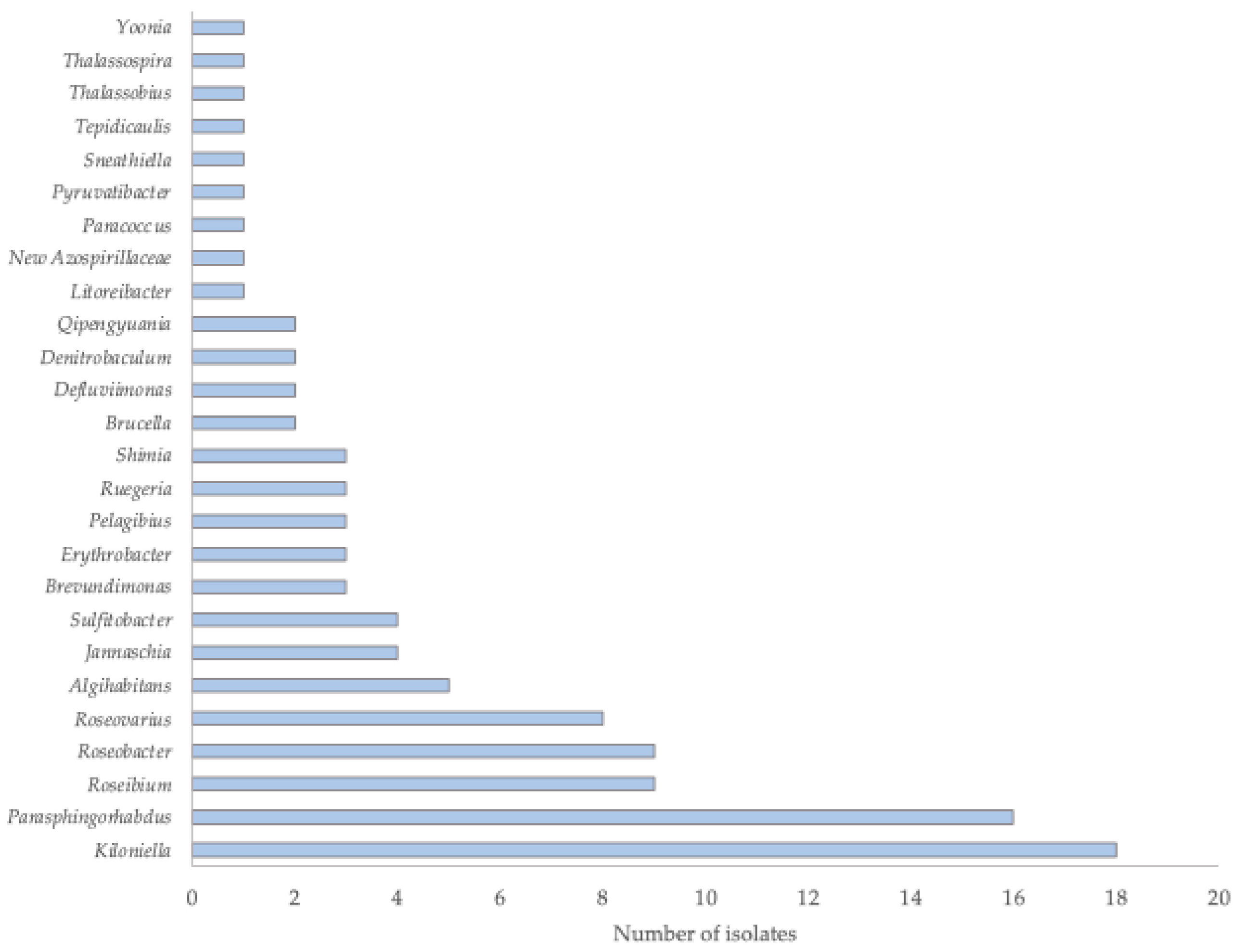
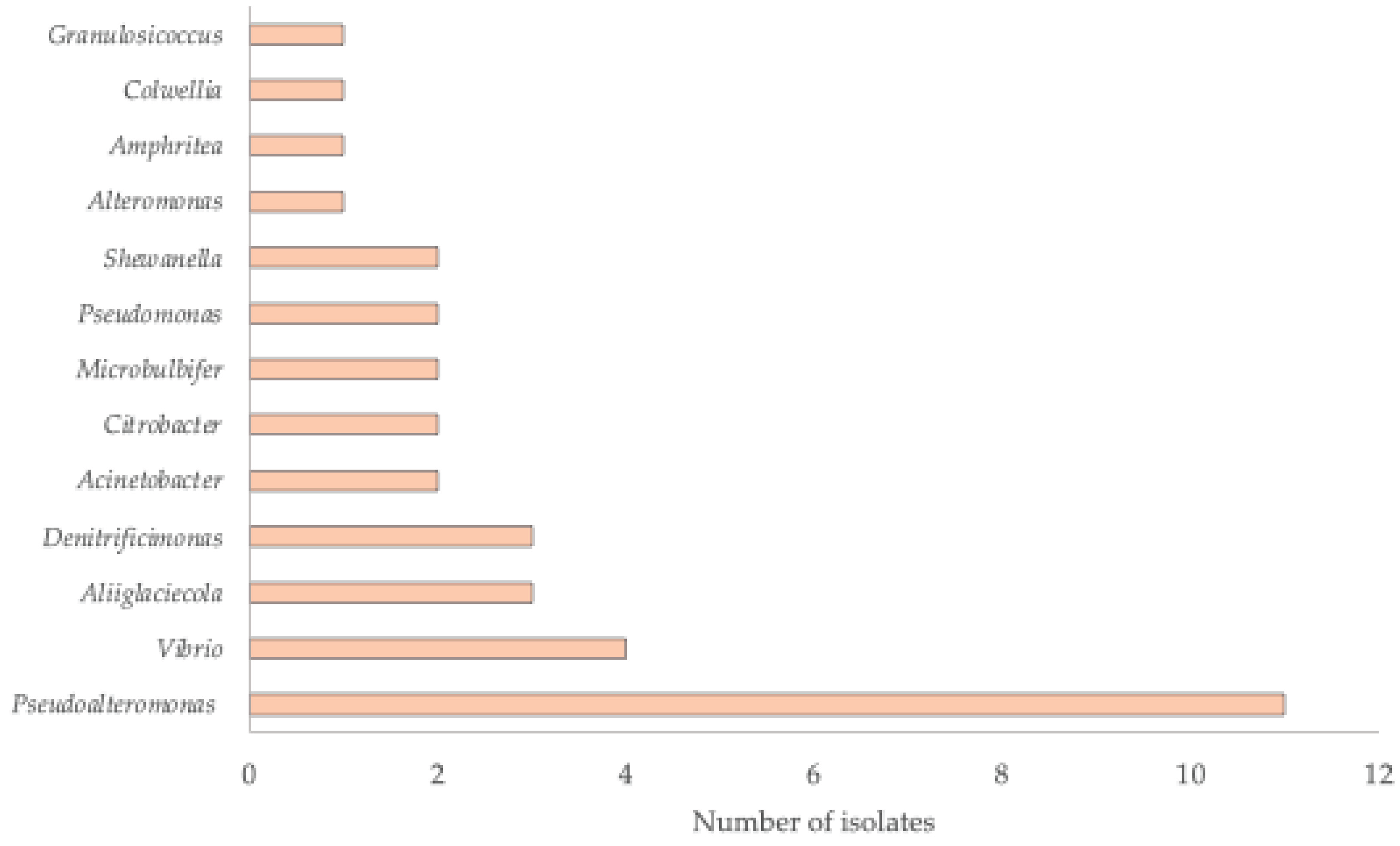

| Isolate | NCBI Accession Number | Sample Source | Phylum | Class | Closest taxa | % Identity * |
| PMO121_14 | PP079866 | Mussel Shell | Bacteroidota | Flavobacteriia | Aquimarina algiphila strain 9Alg 151 | 96.16 |
| PMO114_2.a | PP079845 | Codium sp. | Bacteroidota | Flavobacteriia | Muricauda koreensis strain ECD12 | 98.61 |
| PMO140_13 | PP079889 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Defluviimonas aestuarii strain BS14 | 97.22 |
| PMO140_20.1.b | PP079880 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Defluviimonas aestuarii strain BS14 | 97.38 |
| PMO121_15.2 | PP079843 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Jannaschia seosinensis strain CL-SP26 | 98.21 |
| PMO121_15.5 | PP079842 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Jannaschia seosinensis strain CL-SP26 | 98.16 |
| PMO140_2.r | PP079893 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Jannaschia seosinensis strain CL-SP26 | 97.48 |
| PMO140_6 | PP079883 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Jannaschia seosinensis strain CL-SP26 | 98.21 |
| PMO110_11 | PP079825 | Sea Anemone | Pseudomonadota | Alphaproteobacteria | Litoreibacter meonggei strain MA1-1 | 98.46 |
| PMO119_11 | OK103952 | Sea Anemone | Pseudomonadota | Alphaproteobacteria | Inquilinus ginsengisoli strain Gsoil 080 | 92.33 |
| PMO102_6.2 | PP079808 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Parasphingorhabdus litoris DSM 22379 | 98.4 |
| PMO123_3.2 | PP079787 | Codium sp. | Pseudomonadota | Alphaproteobacteria | Pyruvatibacter mobilis CGMCC 1.15125 | 98 |
| PMO121_3.1 | PP079785 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Qipengyuania seohaensis strain SW-135 | 98.57 |
| PMO114_18 | PP079776 | Codium sp. | Pseudomonadota | Alphaproteobacteria | Roseobacter cerasinus strain AI77 | 98.64 |
| PMO114_5.2 | PP079773 | Codium sp. | Pseudomonadota | Alphaproteobacteria | Roseobacter cerasinus strain AI77 | 98.31 |
| PMO132_8.1 | PP079774 | Codium sp. | Pseudomonadota | Alphaproteobacteria | Roseobacter cerasinus strain AI77 | 98.34 |
| PMO138_13.1 | PP079882 | Sea Anemone | Pseudomonadota | Alphaproteobacteria | Shimia thalassica strain CECT 7735 | 97.49 |
| PMO90_13 | OK103951 | Gracilaria sp. | Pseudomonadota | Alphaproteobacteria | Sneathiella aquamaris 216LB-ZA1-12 | 98.1 |
| PMO140_12 | PP079890 | Mussel Shell | Pseudomonadota | Alphaproteobacteria | Sulfitobacter marinus strain SW-265 | 98.14 |
| PMO127_3 | OK103955 | Gracilaria sp. | Pseudomonadota | Alphaproteobacteria | Tepidicaulis marinus strain MA2 | 95.45 |
| PMO138_12 | OK103953 | Sea Anemone | Pseudomonadota | Betaproteobacteria | Methylotenera oryzisoli strain La3113 | 95.32 |
| PMO138_15.2 | PP079900 | Sea Anemone | Pseudomonadota | Betaproteobacteria | Methylotenera oryzisoli strain La3113 | 95.49 |
| PMO138_17 | PP079899 | Sea Anemone | Pseudomonadota | Betaproteobacteria | Methylotenera oryzisoli strain La3113 | 95.33 |
| PMO138_18 | PP079898 | Sea Anemone | Pseudomonadota | Betaproteobacteria | Methylotenera oryzisoli strain La3113 | 94.92 |
| PMO138_2 | PP079903 | Sea Anemone | Pseudomonadota | Betaproteobacteria | Methylotenera oryzisoli strain La3113 | 95.56 |
| PMO133_10.1 | PP079870 | Corallina sp. | Pseudomonadota | Gammaproteobacteria | Aliiglaciecola litoralis strain Sd 2-38 | 95.95 |
| PMO87_21 | PP079871 | Corallina sp. | Pseudomonadota | Gammaproteobacteria | Aliiglaciecola litoralis strain Sd 2-38 | 96.19 |
| PMO87_4.a | PP079872 | Corallina sp. | Pseudomonadota | Gammaproteobacteria | Aliiglaciecola litoralis strain Sd 2-38 | 95.6 |
| PMO114_2.T | PP079869 | Codium sp. | Pseudomonadota | Gammaproteobacteria | Alteromonas alba strain 190 | 94.61 |
| PMO85_2 | PP079870 | Corallina sp. | Pseudomonadota | Gammaproteobacteria | Microbulbifer echini strain AM134 | 97.94 |
| Isolate | Sample Source | Closest species | % identity* | Phylum/Class | Inhibition zone diameter (mm) | ||||||
| AMP | CTX | IPM | VAN | TET | GEN | CIP | |||||
| PMO87_15.4.1 | Corallina sp. | Mycobacterium adipatum | 99.37 | Actinomycetota/Actinomycetia | 31 | NM | 40 | 10 | 0 | 0 | 42 |
| PMO133_7 | Corallina sp. | Microbacterium algeriense | 99.89 | Actinomycetota/Actinomycetia | 0 | 0 | 0 | 19 | 0 | 0 | 0 |
| PMO90_19.1 | Gracilaria sp. | Aquimarina algiphila | 99.15 | Bacteroidota/Flavobacteriia | ND | ND | ND | 21 | 16 | 10 | 49 |
| 132_20.2.3 | Codium sp. | Aquimarina macrocephali | 100 | Bacteroidota/Flavobacteriia | 10 | 22 | 0 | 20 | 0 | ND | 66 |
| PMO87_22 | Corallina sp. | Zobellia russellii | 99.92 | Bacteroidota/Flavobacteriia | 8 | 0 | 0 | 11 | 8 | 0 | 9 |
| PMO137_2 | Sediments | Novipirellula rosea | 99.77 | Planctomycetota/Planctomycetia | 0 | 0 | 0 | 0 | 0 | 0 | 34 |
| PMO137_6 | Sediments | Novipirellula rosea | 99.77 | Planctomycetota/Planctomycetia | 0 | 0 | 0 | 0 | 0 | 0 | 40 |
| PMO137_9 | Sediments | Novipirellula rosea | 99.78 | Planctomycetota/Planctomycetia | 0 | 0 | 0 | 0 | 0 | 0 | 32 |
| PMO112_11.1.l | Mussel Shell | Rubinisphaera brasiliensis | 100 | Planctomycetota/Planctomycetia | 0 | 0 | 0 | 0 | 32 | 0 | 0 |
| PMO100_1.1 | Sea Anemone | Algihabitans albus | 100 | Pseudomonadota/Alphaproteobacteria | 17 | 0 | 0 | 0 | 11 | 0 | 27 |
| PMO94_4 | Seawater | Brevundimonas bullata | 99.86 | Pseudomonadota/Alphaproteobacteria | 10 | 21 | 24 | 0 | 8 | 20 | 0 |
| PMO122_3.1 | Seawater | Brevundimonas fontaquae | 99.66 | Pseudomonadota/Alphaproteobacteria | 13 | 30 | 0 | ND | 9 | 0 | 0 |
| PMO87_15.1.b | Corallina sp. | Brucella rhizosphaerae | 99.68 | Pseudomonadota/Alphaproteobacteria | 0 | 26 | 0 | 0 | 0 | 0 | 17 |
| PMO140_13 | Mussel Shell | Defluviimonas aestuarii | 97.22 | Pseudomonadota/Alphaproteobacteria | 0 | 22 | 14 | 0 | 18 | 12 | 66 |
| PMO112_8.2 | Mussel Shell | Erythrobacter rubeus | 98.97 | Pseudomonadota/Alphaproteobacteria | NM | NM | 0 | 27 | 0 | 0 | 40 |
| PMO140_15.2 | Mussel Shell | Erythrobacter rubeus | 98.99 | Pseudomonadota/Alphaproteobacteria | 48 | 60 | 0 | 27 | 8 | 0 | 43 |
| PMO139_7.1 | Mussel Interior | Erythrobacter rubeus | 99.03 | Pseudomonadota/Alphaproteobacteria | NM | NM | NM | 21 | 0 | 0 | 44 |
| PMO121_15.5 | Mussel Shell | Jannaschia seosinensis | 98.16 | Pseudomonadota/Alphaproteobacteria | NM | NM | 14 | 15 | 7 | 15 | NM |
| PMO140_2.r | Mussel Shell | Jannaschia seosinensis | 97.48 | Pseudomonadota/Alphaproteobacteria | NM | NM | NM | 16 | 7 | 20 | NM |
| PMO102_1.1 | Mussel Shell | Parasphingorhabdus cellanae | 99.92 | Pseudomonadota/Alphaproteobacteria | 0 | 34 | 0 | 26 | 0 | 0 | 0 |
| PMO95_5.L.1 | Codium sp. | Parasphingorhabdus litoris | 99.6 | Pseudomonadota/Alphaproteobacteria | 0 | 31 | 0 | 22 | 0 | 0 | 0 |
| PMO114_1.a | Codium sp. | Parasphingorhabdus cellanae | 99.85 | Pseudomonadota/Alphaproteobacteria | 0 | 44 | 0 | 12 | 0 | 0 | 20 |
| PMO114_13 | Codium sp. | Parasphingorhabdus cellanae | 99.92 | Pseudomonadota/Alphaproteobacteria | 0 | 54 | 0 | 27 | 0 | 0 | 30 |
| PMO114_16.1 | Codium sp. | Parasphingorhabdus cellanae | 100 | Pseudomonadota/Alphaproteobacteria | 0 | 44 | 0 | 13 | 0 | 0 | 23 |
| PMO115_16.1 | Corallina sp. | Parasphingorhabdus cellanae | 99.05 | Pseudomonadota/Alphaproteobacteria | 0 | 35 | 0 | 29 | 0 | 0 | 16 |
| PMO96_5.1 | Corallina sp. | Parasphingorhabdus cellanae | 100 | Pseudomonadota/Alphaproteobacteria | NM | NM | NM | 36 | 0 | 0 | 28 |
| PMO95_8.1 | Codium sp. | Parasphingorhabdus cellanae | 99.92 | Pseudomonadota/Alphaproteobacteria | 28 | NM | NM | 25 | 0 | 8 | 19 |
| PMO95_13.2 | Codium sp. | Parasphingorhabdus cellanae | 99.84 | Pseudomonadota/Alphaproteobacteria | 10 | 22 | 30 | 32 | 8 | 0 | 15 |
| PMO123_1 | Codium sp. | Parasphingorhabdus cellanae | 99.19 | Pseudomonadota/Alphaproteobacteria | 0 | 47 | 0 | 27 | 0 | 0 | 0 |
| PMO123_2 | Codium sp. | Parasphingorhabdus cellanae | 99.07 | Pseudomonadota/Alphaproteobacteria | 0 | 50 | 0 | 22 | 0 | 0 | 0 |
| PMO95_11 | Codium sp. | Parasphingorhabdus cellanae | 99.69 | Pseudomonadota/Alphaproteobacteria | 20 | NM | NM | 26 | 0 | 0 | 0 |
| PMO95_10 | Codium sp. | Parasphingorhabdus cellanae | 100 | Pseudomonadota/Alphaproteobacteria | 24 | NM | NM | 26 | 7 | 0 | 28 |
| PMO140_9.1.2 | Mussel Shell | Parasphingorhabdus cellanae | 99.92 | Pseudomonadota/Alphaproteobacteria | 0 | 42 | 0 | 27 | 0 | 0 | 0 |
| PMO114_8.v | Codium sp. | Qipengyuania gelatinilytica | 98.75 | Pseudomonadota/Alphaproteobacteria | 46 | 56 | 0 | 32 | 9 | 11 | 43 |
| PMO110_1.1 | Sea Anemone | Roseovarius aestuarii | 99.28 | Pseudomonadota/Alphaproteobacteria | 0 | 40 | 14 | 0 | 9 | 0 | 48 |
| PMO111_2.1 | Mussel Interior | Ruegeria faecimaris | 98.96 | Pseudomonadota/Alphaproteobacteria | 39 | 52 | 17 | 11 | 8 | 0 | 46 |
| PMO90_13 | Gracilaria sp. | Sneathiella aquimaris | 98.1 | Pseudomonadota/Alphaproteobacteria | 0 | 32 | 46 | 0 | 9 | 11 | 12 |
| PMO122_2.a | Seawater | Yoonia maritima | 98.78 | Pseudomonadota/Alphaproteobacteria | NM | NM | NM | 18 | 10 | 26 | 48 |
| PMO94_2 | Seawater | Acinetobacter johnsonii | 100 | Pseudomonadota/Gammaproteobacteria | 22 | 23 | 40 | ND | 20 | 27 | 0 |
| PMO87_4.a | Corallina sp. | Aliiglaciecola litoralis | 95.6 | Pseudomonadota/Gammaproteobacteria | 16 | 27 | 25 | 18 | 8 | 13 | 25 |
| PMO87_21 | Corallina sp. | Aliiglaciecola litoralis | 96.19 | Pseudomonadota/Gammaproteobacteria | 13 | 41 | 26 | 18 | 21 | 20 | 32 |
| PMO133_10.1 | Corallina sp. | Aliiglaciecola litoralis | 95.95 | Pseudomonadota/Gammaproteobacteria | 0 | 40 | 0 | 21 | 8 | 15 | 31 |
| PMO120_1 | Mussel Interior | Citrobacter braakii | 99.92 | Pseudomonadota/Gammaproteobacteria | 6 | 32 | 15 | ND | 18 | 17 | 40 |
| PMO126_r.1 | Sargassum muticum | Citrobacter freundii | 100 | Pseudomonadota/Gammaproteobacteria | 10 | 29 | 15 | ND | 18 | 15 | 20 |
| PMO140_4 | Mussel Shell | Microbulbifer echini | 100 | Pseudomonadota/Gammaproteobacteria | 0 | 47 | 0 | 12 | 0 | 15 | 35 |
| PMO85_2 | Seawater | Microbulbifer okhotskensis | 99.05 | Pseudomonadota/Gammaproteobacteria | 0 | 21 | 33 | 0 | 20 | 18 | 40 |
| PMO113_2 | Seawater | Pseudoalteromonas prydzensis | 99.06 | Pseudomonadota/Gammaproteobacteria | 14 | 24 | 0 | 0 | 0 | 10 | 23 |
| PMO141_1.v | Invertebrate | Pseudoalteromonas translucida | 100 | Pseudomonadota/Gammaproteobacteria | 17 | 0 | 0 | 12 | 9 | 10 | 31 |
| PMO88_1 | Osmundea sp. | Pseudoalteromonas arctica | 99.78 | Pseudomonadota/Gammaproteobacteria | 20 | 25 | 25 | 0 | 13 | 11 | 24 |
| PMO87_5.2 | Corallina sp. | Pseudoalteromonas translucida | 100 | Pseudomonadota/Gammaproteobacteria | 18 | 25 | 21 | 0 | 12 | 10 | 24 |
| PMO85_3.1 | Seawater | Pseudoalteromonas carrageenovora | 100 | Pseudomonadota/Gammaproteobacteria | 21 | 37 | 29 | 10 | 13 | 11 | 35 |
| PMO103_8 | Seawater | Denitrificimonas caeni | 99.61 | Pseudomonadota/Gammaproteobacteria | 19 | 0 | 34 | 0 | 0 | 20 | 26 |
| PMO94_5 | Seawater | Denitrificimonas caeni | 99.35 | Pseudomonadota/Gammaproteobacteria | ND | ND | ND | 0 | 0 | 17 | 0 |
| PMO108_1.1.l | Gracillaria sp | Shewanella colwelliana | 100 | Pseudomonadota/Gammaproteobacteria | ND | ND | 0 | 0 | 0 | 10 | 32 |
| PMO138_9.1 | Sea Anemone | Stutzerimonas nitrititolerans | 99.75 | Pseudomonadota/Gammaproteobacteria | 0 | 18 | 0 | 0 | 0 | 20 | 22 |
| Component | M600 (per 1000 mL) | M607 + NAG (per 1000 mL) |
| Peptone Yeast Extract Tris-HCl Glucose (2.5%) Hutner’s Basal Salts* Vitamin Mix** NAG (5%) Agar Natural Seawater |
1 g 1 g 50 mL 40 mL 20 mL 10 mL – 16 g 880 mL |
0.25 g 0.25 g 50 mL 10 mL 20 mL 10 mL 10 mL 16 g 900 mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



