
Submitted:
02 March 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Background: Linezolid-non-susceptible Enterococcus faecalis (LNSEf) has emerged as a critical clinical concern worldwide; however, data from Latin American settings remain scarce. Methods: We investigated 392 non-duplicated E. faecalis isolates collected over 12 months in a Mexican tertiary care hospital. Antimicrobial susceptibility was assessed phenotypically, and 24 isolates with linezolid minimum inhibitory concentrations ≥4 µg/mL underwent whole-genome sequencing. Bioinformatic analyses included multilocus sequence typing (MLST), detection of linezolid resistance mechanisms (optrA, cfrA, 23S rRNA mutations), and phylogenetic reconstruction. Results: Of the 392 isolates, 6.12% demonstrated linezolid non-susceptibility, predominantly associated with plasmid- or chromosomally encoded optrA, with only two isolates harboring cfrA. No mutations were detected in 23S rRNA domain V or ribosomal proteins L3/L4. Clinically, LNSEf isolates were linked to immunosuppression, prior surgical procedures, and extended hospital stays. MLST revealed broad genetic diversity, indicating both clonal expansion and independent acquisitions of resistance. Despite retaining susceptibility to ampicillin, vancomycin, and daptomycin, most LNSEf isolates possessed extensive resistance to other classes of antibiotics, particularly aminoglycosides and fluoroquinolones. Conclusions: LNSEf poses a growing threat in this region, driven largely by optrA. Strengthened genomic surveillance, stringent infection control, and optimized antimicrobial stewardship are vital to contain its spread.
Keywords:
Enterococcus faecalis
; linezolid resistance
; optrA
; cfr
; multidrug-resistant bacteria
; genomic surveillance
1. Introduction
Linezolid, a key oxazolidinone antibiotic, is extensively used to manage severe infections caused by multidrug-resistant gram-positive bacteria, including Enterococcus faecalis and Enterococcus faecium [1,2]. The mechanism of action of linezolid involves the inhibition of bacterial protein synthesis by binding to the 23S ribosomal RNA of the 50S subunit, thereby preventing the formation of a functional 70S initiation complex and inhibiting the translation process [3,4]. Additionally, it competes with incoming substrates at the A-site of the ribosome, located near the catalytic center, further disrupting protein synthesis [4,5]. As a result, linezolid is primarily bacteriostatic against most Gram-positive bacteria, including enterococci and staphylococci, although it can be bactericidal against certain strains of streptococci [6,7].
Linezolid has been primarily used to treat infections caused by multidrug-resistant Gram-positive organisms, including methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant enterococci (VRE), and multidrug-resistant Mycobacterium tuberculosis (MDR-TB) [1,8]. It is approved for use in conditions such as nosocomial pneumonia, community-acquired pneumonia, complicated and uncomplicated skin and soft tissue infections, and infections caused by vancomycin-resistant Enterococcus faecium (VRE) [3]. Its unique mechanism of action and oral bioavailability make it a versatile option for both hospital and outpatient settings [1,9].
However, the emergence of linezolid-non-susceptible enterococci (LNSE) poses a significant challenge to clinical management and necessitates ongoing surveillance and a deeper understanding of resistance mechanisms [10,11,12,13]. Linezolid-non-susceptibility in Enterococcus faecalis is relatively rare but is emerging as a concern in various regions [10,11]. For instance, a study from the United States reported a low resistance rate of 2% among E. faecalis isolates [10]. Similarly, a study from Iran found a linezolid-non-susceptibility rate of 1.3% in E. faecalis isolates from urinary tract infections [11]. In contrast, a study from Korea reported a higher resistance rate of 18.8% among E. faecalis isolates, and in a tertiary hospital in China, the frequency of linezolid-non-susceptible E. faecalis was reported to be 22.61% among clinical isolates from urinary tract infections [12,13].
Epidemiological studies conducted in various regions have highlighted the sporadic nature of LRE outbreaks, suggesting that these cases often result from independent acquisition events rather than clonal dissemination [14,15]. For instance, in Chongqing, China, a study of low-level LRE strains revealed the presence of novel mutations and sequence types, indicating a diverse genetic landscape and emphasizing the importance of local surveillance [15]. Similarly, genomic epidemiology studies in China and other countries have shown the coexistence of multiple resistance mechanisms, underscoring the complexity of controlling the spread of LRE [16,17,18].
The occurrence of linezolid-non-susceptible Enterococcus faecalis (LNSEf) is driven by multiple mechanisms such as mutations in the 23S rRNA, alterations in ribosomal proteins L3 and L4, and acquisition of resistance genes like optrA, poxtA and cfr [10,19,20,21]. Mutations in the linezolid target sites, including the V domain of the 23S ribosomal RNA genes and the 50S ribosomal proteins L3 and L4, significantly affect linezolid susceptibility by altering the drug's binding affinity to the ribosome [22]. Mutations in the 23S rRNA, particularly at positions such as G2576, are well-documented mechanisms of resistance, as they directly impact the binding site of linezolid, reducing its efficacy [23]. Mutations in ribosomal proteins L3 and L4, although located further from the linezolid binding site, can also contribute to resistance [24]. These mutations may induce conformational changes in the ribosome that indirectly affect linezolid binding [24]. For instance, specific mutations in L3, such as R138V, have been associated with increased linezolid minimum inhibitory concentrations (MICs), indicating reduced susceptibility [24,25].
The optrA gene is a well-documented contributor to linezolid-non-susceptibility, as it encodes an ATP-binding cassette (ABC) transporter that actively effluxes linezolid out of the bacterial cell, thereby reducing its intracellular concentration and effectiveness [26,27]. Studies from China have shown a high prevalence of optrA in various settings, including hospital sewage and community environments, often associated with other antimicrobial resistance genes (ARGs), such as fexA and erm(A), which can co-transfer with optrA via mobile genetic elements like plasmids and transposons [16,27,28].
The poxtA gene, although less frequently discussed, also contributes to linezolid-non-susceptibility by encoding a protein that protects the ribosome from the action of oxazolidinones [29]. It has been identified in various Enterococcus species, including E. faecalis, and is often found on plasmids, facilitating its horizontal transfer [29,30]. The presence of poxtA, along with optrA, can lead to higher levels of resistance due to their combined effects on linezolid susceptibility [29,30,31]
The cfr gene affects linezolid susceptibility by encoding a methyltransferase that modifies the 23S rRNA at position A2503 [32]. This methylation alters the binding site of linezolid on the bacterial ribosome, thereby reducing the drug's ability to inhibit protein synthesis [33]. The cfr gene confers a multidrug-resistant phenotype, not only to oxazolidinones like linezolid but also to phenicols, lincosamides, pleuromutilins, and streptogramin A compounds [32]. The cfr gene is often located on mobile genetic elements such as plasmids and transposons, facilitating its horizontal transfer across different bacterial species and contributing to the dissemination of resistance [34].
This diversity in the regional prevalence of linezolid-non-susceptibility and resistance mechanisms highlights the importance of regional surveillance to monitor resistance patterns. Overall, while linezolid-non-susceptibility in E. faecalis is not yet widespread, it is an emerging concern that requires ongoing monitoring. In the present study, we aim to investigate the molecular epidemiology and mechanisms of linezolid-non-susceptibility in Enterococcus faecalis strains isolated from a Mexican tertiary care university hospital. By characterizing the genetic basis of resistance and analyzing the clonal relationships among isolates, we seek to provide insights into the local dynamics of linezolid-non-susceptible enterococci and contribute to the global understanding of this emerging threat.
2. Materials and Methods
2.1. Bacterial Strain Collection, Clinical Data Gathering, and Antimicrobial Susceptibility Testing
A total of 392 non-duplicated Enterococcus faecalis strains were obtained in the microbiology laboratory of our institution from June 2023 to June 2024. The strains were collected from various sample types, including tissue, urine, blood culture, surgical wound secretion, abscess, peritoneal fluid, bone, and cerebrospinal fluid (CSF). They were initially identified using the VITEK2 system (BioMérieux, Lyon, France). Antimicrobial susceptibility testing for ampicillin (AMP), ciprofloxacin (CIP), daptomycin (DAP), nitrofurantoin (NIT), gentamicin (GEN), streptomycin (STR), levofloxacin (LEV), linezolid (LNZ), benzylpenicillin (PEN), tetracycline (TET), and vancomycin (VA) was performed using AST-GP75 and P663 cards (BioMérieux). Results were interpreted following CLSI guidelines, considering linezolid susceptibility in Enterococcus faecalis (LSEf) at ≤2 µg/mL, intermediate resistance at 4 µg/mL, and resistance at ≥8 µg/mL. For the purposes of this study, the latter two categories were classified as linezolid-non-susceptible Enterococcus faecalis (LNSEf) [35].
Clinical records of patients with LNSEf isolates were reviewed, and sociodemographic, microbiological, and clinical data were systematically collected. The data included age, gender, admission diagnosis, occupation, prior antibiotic use within 90 days and 14 days, type of antibiotics used, previous hospitalizations, comorbidities, Charlson Comorbidity Index, surgical history within the last three months, type of surgery if applicable, use of vasopressor agents, ICU admission, initial empirical treatment, length of hospital stay, and in-hospital mortality. For each patient with a resistant E. faecalis isolate, two control patients with linezolid susceptible E. faecalis isolates, matched by age, gender, and clinical ward, were selected to compare and identify factors associated with LNSEf.
2.2. DNA Extraction, Library Construction, and Whole-Genome Sequencing
Genomic DNA was extracted from pure cultures of Enterococcus faecalis using the ZymoBIOMICS DNA Miniprep Kit (Zymo Research, USA), following the manufacturer’s instructions. DNA purity was assessed by spectrophotometry (Nanodrop One C, Thermo Fisher Scientific, USA), selecting samples with an A260/280 ratio between 1.8 and 2.0. DNA concentration was quantified using fluorometry (Qubit 4.0, Invitrogen, Thermo Fisher Scientific, USA) with the dsDNA High Sensitivity (HS) Assay. Extracted DNA was subsequently stored at −70 °C.
For whole-genome library preparation, the Illumina DNA Prep protocol (Illumina, USA) was used, which applies a bead-based transposome complex to fragment genomic DNA. The process was automated using the Biomek NGeniuS Next Generation Library Prep System (Beckman Coulter, USA), with a total DNA input of 20 ng and programmed using 8 cycles to amplify tagmented DNA using IDT indexes set A. Library quality was assessed using the Standard (S2) Cartridge Kit with a Qsep1 instrument (BioOptic Inc., Taiwan), which resulted in fragment sizes of approximately 600–700 bp. DNA concen-tration and dilution were further verified using the dsDNA HS Assay fluorometry (Qubit 4.0, Invitrogen, Thermo Fisher Scientific, USA). The prepared libraries were sequenced on an Illumina NextSeq 2K platform (Illumina, USA) in paired-end mode with 150 bp read lengths. Flow cell loading and sequencing were performed following Illumina protocols to ensure data quality and integrity.
2.3. Bioinformatic Analyses
The bioinformatic workflow was directed toward the genomic characterization of Enterococcus faecalis, emphasizing molecular typing, detection of antimicrobial resistance (AMR) genes—particularly for linezolid-non-susceptibility—and phylogenetic analysis.
Initial quality control (QC) of the sequencing data in FASTQ format, representing clinical isolates of E. faecalis, was performed using FastQC and MultiQC. These tools evaluated read quality metrics such as per-base sequence quality, GC content, and adapter contamination, ensuring the data met quality standards for downstream analyses (Table S1 of the supplementary materials). Taxonomic classification was conducted using Kraken2, with a reference database enabling the isolation of E. faecalis-specific reads. Non-target sequences were discarded, ensuring that the subsequent steps focused exclusively on the target organism. To enhance the quality of the data, trimmed reads were generated using fastp. This process removed low-quality bases and adapter sequences, resulting in high-quality datasets optimized for genome assembly.
Genome assembly was performed with SPAdes using the "careful" mode to reduce assembly errors. The resulting assemblies were evaluated using QUAST, with metrics including the number of contigs, total genome length (~2.8–3 Mbp for E. faecalis), N50, and GC content (~37%) (Table S2 of the supplementary materials). These metrics provided an assessment of the assembly completeness and contiguity.
The raw sequences have been made publicly available via the Zenodo platform at https://doi.org/10.5281/zenodo.14873829. The genome assemblies of the 24 LNSEf generated in this study have been deposited in GenBank under BioProject ID PRJNA1217060, with individual genome accessions JBLVLD000000000–JBLVKX000000000 (Table S2 of the supplementary materials).
AMR gene detection focused on identifying resistance determinants, including genes related to linezolid non-susceptibility (optrA, cfr, poxtA) and other antimicrobial classes. ARIBA was used with ResFinder and CARD databases to detect AMR genes directly from raw reads or assemblies. Additionally, LRE-Finder (version 1.0.0) was employed to identify optrA, cfr, cfrB, and poxtA genes, as well as common mutations in the V domain of the 23S rRNA (G2576T and G2505A) in LNSEf isolates.
All detected optrA sequences were blasted against the complete optrA gene sequence from plasmid pE349 (GenBank Accession No. NG_048023.1). Specific mutations associated with linezolid non-susceptibility were further analyzed using BWA-MEM for read mapping and bcftools for variant calling, ensuring precise detection of resistance-related genetic variations. To predict additional linezolid resistance genes such as cfrD, the Resistance Gene Identifier (RGI) (version 6.0.3) was used, utilizing reference data from CARD (version 4.0.0).
To validate the identity of the detected resistance genes, all sequences corresponding to optrA, cfr, and related genes were translated into amino acid sequences using Prodigal (version 2.6.3). These protein sequences were subsequently subjected to BLASTP (DIAMOND, version 2.1.10) analysis against the non-redundant protein (NR) database of NCBI, with stringent screening parameters of ≥ 99% sequence identity and an E-value threshold of < 1e-10 (Table S3 of the supplementary materials.) This approach ensured high-confidence matches and confirmed the presence of functional resistance determinants.
The localization of optrA genes was determined by performing BLASTN searches against the NCBI nt database using contigs containing optrA sequences. Plasmid-specific replicons were detected using PlasmidFinder (version 2.1, database 2023-12-04), and alignment results were analyzed to compare identity and coverage with known plasmid and chromosomal sequences.
Assembled contigs were analyzed utilizing the ABRicate tool (version 1.0.1) to identify virulence-associated genes. The virulence factor database (VFDB) was employed with a minimum identity threshold of 80% and a minimum gene coverage of 80%. Virulence genes were identified based on their sequence similarity to previously reported entries in the VFDB.
Molecular typing was performed using multilocus sequence typing (MLST) via the PubMLST database (https://pubmlst.org/organisms/enterococcus-faecalis), classifying isolates into sequence types (STs) based on the allelic profiles of seven housekeeping genes (gdh, gyd, pstS, gki, aroE, xpt, and yqiL).
Phylogenetic analysis for each Enterococcus faecalis isolates involved extracting core genome single nucleotide polymorphisms (SNPs) using Parsnp (v1.2), followed by SNP alignments generated with vcf2phylip (v2.0), and the construction of maximum likelihood (ML) phylogenetic trees with IQ-TREE (v2.4.0) using the GTR+G model and 1000 ultrafast bootstrap replicates. For optrA phylogenetic analysis, sequencing reads were aligned to the optrA reference sequence (NG_048023.1) using Bowtie2 (v2.5.1), and variants were identified with Snippy (v4.6.0), retaining only high-confidence SNPs (QUAL > 100, DP ≥ 10). Consensus sequences from each isolate were aligned with MAFFT (v7.526), and an ML phylogenetic tree was reconstructed using IQ-TREE (v2.4.0, GTR+G model, 1000 bootstrap replicates). Trees were visualized in iTOL (v6.8), elucidating evolutionary relationships and strain clustering.
2.4. Statistical Analysis
Demographic data are reported as simple relative frequencies. The normality of the data distribution was assessed via the Shapiro–Wilk test. Pearson’s chi-square test and Fisher’s exact test were used to compare proportions, as appropriate. For comparisons of quantitative variables, Student’s t tests and Wilcoxon‒Mann‒Whitney tests were used for normally and nonnormally distributed data, respectively.
Statistical analyses were conducted using Python (v3.9) for data processing and visualization, leveraging libraries such as pandas (v1.3.5), NumPy (v1.21.4), and Matplotlib (v3.5.0). Additionally, R software (v4.2.2) was utilized for statistical analyses, validation, and graphical representation.
3. Results
Of the 392 non-duplicated Enterococcus faecalis strains, we identified 26 isolates with linezolid MICs ≥ 4 µg/mL through automated phenotypic methods, which were subsequently subjected to whole-genome sequencing (WGS). After excluding isolates with inadequate readings or insufficient quality for downstream analysis, 24 isolates were selected for molecular typing, characterization of linezolid-non-susceptibility mechanisms, and phylogenetic tree construction. Additionally, 12 linezolid-susceptible isolates underwent WGS to assess genetic diversity, of which 3 were excluded due to insufficient quality for further analysis.
Finally, 48 linezolid-susceptible isolates were documented from clinical records (including the 12 previously described isolates that underwent WGS) for sociodemographic and clinical comparisons. The study workflow, which encompasses whole-genome sequencing, antimicrobial resistance characterization, and the sociodemographic and clinical profiling of patients with linezolid-susceptible and non-susceptible Enterococcus faecalis isolates, is depicted in Figure 1.
Among the 24 patients (7 women, 17 men) with linezolid-non-susceptible E. faecalis (LNSEf) isolates, the median age was 48 years (interquartile range [IQR] 31–57). Comorbidities were identified in 66.7% of patients (n=16), with a median Charlson comorbidity index of 1 (IQR 0–3). The most common comorbidities included diabetes mellitus (25.0%), hypertension (20.83%), immunosuppression (12.50%), and chronic kidney disease (8.33%). Overall, 66.7% of patients had undergone a surgical intervention, while 12.5% required admission to the intensive care unit (ICU). The median hospital stay was 14 days (IQR 11.25–20.25), and in-hospital mortality was 16.7% (n=4).
In terms of occupation, unemployed individuals represented the most common category, accounting for 7 cases (29.17%). Housewives comprised 5 cases (20.83%), followed by students with 4 cases (16.67%). Construction workers were affected in 2 cases (8.33%), while delivery workers, commerce workers, engineers, and couriers each accounted for 1 case (4.17%).
Sex-based comparisons revealed that women had a higher median Charlson comorbidity index (3 vs. 0; p = 0.059) and a significantly higher in-hospital mortality rate (75% vs. 25%; p = 0.040) than men. Conversely, men were significantly more likely to have undergone recent surgical intervention (87.5% vs. 12.5%; p = 0.045). These findings highlight notable sex-related differences in comorbidity burden and clinical outcomes among patients with linezolid-non-susceptible E. faecalis.
With respect to prior antibiotic exposure, 45.83% (n=11) of patients had received antibiotic treatment within the past 90 days, while 37.5% (n=9) had recent antibiotic exposure within the two weeks preceding sample collection. Regarding the specific antibiotics administered before resistance was detected, cefazolin plus gentamicin (CZ+GEN) and ciprofloxacin (CIP) were the most frequently used, each in 8.33% (n=2) of cases. Other antibiotics were used in single cases, each accounting for 4.17% of cases, including ceftriaxone plus clindamycin plus ciprofloxacin (CRO+CLI+CIP), ceftriaxone (CRO), meropenem plus ceftriaxone (MEM+CRO), ceftriaxone plus clindamycin (CRO+CLI), ceftriaxone plus metronidazole plus amikacin (CRO+MTZ+AN), cefotaxime (CTX), and ceftazidime plus vancomycin (CAZ+VA).

Table 1.
Sociodemographic and clinical characteristics of patients with linezolid-non-susceptible Enterococcus faecalis isolates distributed by sex.
Table 1.
Sociodemographic and clinical characteristics of patients with linezolid-non-susceptible Enterococcus faecalis isolates distributed by sex.
| Variable | Total (n = 24) |
Women (%) (n = 7) |
Men (%) (n = 17) |
p value |
|---|---|---|---|---|
| Age – median (IQR) | 48 (31-57) | 56 (53.5-62.25) | 43 (27-55) | 0.092 |
| Previous antibiotic use in 90 days – n (%) | 11 (45.8) | 3 (27.3) | 8 (72.7) | 1 |
| Previous Antibiotic in 14 days – n (%) | 9 (37.5) | 3 (33.3) | 6 (66.7) | 0.643 |
| Previous hospitalization in 90 days – n (%) | 13 (54.2) | 4 (30.8) | 9 (69.2) | 0.66 |
| Comorbidities – n (%) | 16 (66.7) | 6 (37.5) | 10 (62.5) | 0.1243 |
| Charlson comorbidity index – median (IQR) | 1 (0-3) | 3 (1.5-3.75) | 0 (0-2) | 0.059 |
| Diabetes Mellitus – n (%) | 6 (25.0) | 3 (50) | 3 (50) | 0.2786 |
| Hypertension – n (%) | 5 (20.8) | 3 (60) | 2 (40) | 0.0886 |
| Cardiovascular disease – n (%) | 0 (0) | 0 (0) | 0 (0) | 1 |
| Obesity – n (%) | 1 (4.2) | 1 (100) | 0 (0) | 0.260 |
| Immunosuppression – n (%) | 3 (12.5) | 1 (33.3) | 2 (66.7) | 1 |
| Chronic kidney disease – n (%) | 2 (8.3) | 0 (0) | 2 (100) | 1 |
| Surgical Intervention – n (%) | 16 (66.7) | 2 (12.5) | 14 (87.5) | 0.045 |
| Central line insertion – n (%) | 9 (37.5) | 3 (33.3) | 6 (66.7) | 0.643 |
| Urinary catheter – n (%) | 16 (66.7) | 5 (31.2) | 11 (68.8) | 0.621 |
| Need for vasopressors – n (%) | 7 (29.2) | 3 (42.9) | 4 (57.1) | 0.318 |
| ICU admission – n (%) | 3 (12.5) | 2 (66.7) | 1 (33.3) | 0.155 |
| Hospital stay – median (IQR) | 14 (11.25-20.25) | 14 (10.25-14.75) | 15 (12-23) | 0.343 |
| In-hospital mortality – n (%) | 4 (16.7) | 3 (75) | 1 (25) | 0.040 |
In the matched comparison of 24 patients with linezolid-non-susceptible E. faecalis (LNSEf) and 48 patients with linezolid-susceptible E. faecalis (LSEf)—matched by age, sex, and hospital department—the LNSEf group exhibited significantly higher rates of immunosuppression (12.5% vs. 0%; p = 0.0339) and prior surgical interventions (66.7% vs. 22.9%; p = 0.0008), along with a significantly longer median hospital stay (14 days [IQR: 11.25–20.25] vs. 9.5 days [IQR: 3.75–15]; p = 0.02).
Although prior hospitalization within the preceding 90 days (54.2% vs. 31.2%; p = 0.104) and the presence of any comorbidity (70.8% vs. 47.9%; p = 0.111) were more frequent in the LNSEf group, these differences did not reach statistical significance. Notably, in-hospital mortality rates were identical in both groups (16.7%; p = 1).
Overall, these findings suggest that LNSEf infection was associated with a greater burden of immunosuppression, a higher frequency of surgical interventions, and prolonged hospitalization, while other sociodemographic and clinical factors remain largely comparable. Further details are presented in Table 2.
The documented cases of LNSEf were distributed across various hospital departments. Traumatology, General Surgery, and Thorax and Cardiovascular Surgery each accounted for 12.5% (n=3). Cardiology and Adult Infectious Diseases departments represented 8.3% (n=2), with most cases concentrated in surgical units.
Regarding sample sources, urine was the most common site of isolation (29.2%, n=7). Surgical wound exudates, foot tissue samples, and blood cultures from central lines each accounted for 8.3% (n=2). The sociodemographic and clinical characteristics of patients with LNSEf are detailed in Table 3, while an extended complementary version is available in Table S3 of the supplementary materials.
With respect to the antimicrobial susceptibility profile, ampicillin, penicillin, nitrofurantoin, and vancomycin exhibited complete (100%) susceptibility. Similarly, daptomycin demonstrated 100% susceptibility according to Clinical and Laboratory Standards Institute (CLSI) criteria (MIC ≤ 4 µg/mL). In contrast, tetracycline resistance was notably high (91.7%), and significant rates of high-level aminoglycoside resistance were observed for gentamicin (66.7%) and streptomycin (50%). Levofloxacin resistance affected 62.5% of isolates, with an additional 4.2% displaying intermediate MIC values, while ciprofloxacin resistance reached 66.7%. The phenotypic and genotypic characteristics, along with hospital distribution, are presented in Table 3, and the overall resistance profile is illustrated in Figure S1 of the supplementary materials.
Genotypic analysis revealed 15 distinct sequence types (STs) among the 24 LNSEf isolates. ST585 was the most frequent (16.7%, n=4), followed by ST32 (12.5%, n=3). ST506 and ST476 were each identified in two isolates (8.3%). The remaining STs—ST101, ST283, ST287, ST202, ST480, ST376, ST179, ST16, ST69, ST415, and ST40—each accounted for 4.2% (n=1). Additionally, two isolates (8.3%) were classified as ‘Not Determined’ (ND).
The primary resistance mechanism was the optrA gene, present in all isolates. The cfrA gene, co-occurring with optrA, was detected in two isolates (8.3%). The most frequent optrA variants were G1879A, C1933T (45.8%) and T411G, T626G, G866A (29.2%). A smaller subset (8.3%) carried the T10G, T35A, C54T, A91G, A107G, T626G, C949T, A1966G profile. Two isolates exhibited an even more complex mutation set (including A134T, G1278A, A1331G, A1541C, C1933T) together with cfrA. No mutations were identified in 23S rRNA (G2505A, G2576U) or ribosomal proteins rplC (L3) and rplD (L4).
BLASTN analysis identified optrA in both chromosomal and plasmidic locations, with 11 isolates (45.8%) in each. In two isolates (8.3%), optrA was chromosomal while cfrA was plasmidic. The presence of optrA on plasmids suggests a high potential for horizontal transfer, whereas chromosomal integration may indicate stable inheritance within specific E. faecalis genetic backgrounds. To validate resistance determinants, optrA and cfrA protein sequences underwent BLASTP (DIAMOND) analysis against the NCBI non-redundant protein database. High sequence identity (≥99%) and significant E-values (<1e-10) confirmed their classification as functional ABC-F type ribosomal protection proteins. The identification of antibiotic resistance proteins using BLASTP analysis in clinical isolates is presented in Table S4 of the supplementary material.
Besides optrA and cfrA, various antimicrobial resistance (AMR) genes were identified. The most common were lsa(A) and fexA (100%), followed by erm(B) (95.8%) and erm(A) (87.5%), linked to macrolide resistance. dfrG (87.5%) conferred trimethoprim resistance. Aminoglycoside-modifying enzymes ant(6)-Ia and aph(3')-III were found in 83.3%, while tet(L) (75.0%) and tet(M) (8.3%) contributed to tetracycline resistance. Other notable genes included aac(6')-aph(2'') (58.3%), cat (45.8%), lnu(B) (37.5%), and fluoroquinolone resistance-associated mutations in parC (S80I) and gyrA (S83I). These findings underscore a multidrug-resistant profile that may limit treatment options.
Analysis using PlasmidFinder revealed a diverse set of plasmid replicons among the 24 LNSEF isolates. The most frequently detected replicon was repUS43, identified in 16 isolates. Other commonly found plasmid types included rep9b (5 isolates), rep7a (3 isolates), and rep1 and rep9c, each present in 2 isolates. Less frequently detected replicons included rep2, rep6, rep22, and rep18b, each observed in a single isolate. Notably, four isolates did not present any detectable plasmid replicons. The presence of multiple plasmid replicons suggests a potential role in the dissemination of antimicrobial resistance genes within Enterococcus faecalis populations.
Further analysis revealed that certain sequence types (STs) exhibited consistent optrA variants with uniform genomic localization in the isolates examined. For instance, both ST101 and ST202 carried the T10G, T35A, C54T, A91G, A107G, T626G, C949T, A1966G variant exclusively on plasmids, while ST16 and ST283 displayed the T626G, A1541C variant, also on plasmids. ST179, in contrast, consistently had the G1879A, C1933T variant integrated into the chromosome. Notably, however, G1879A, C1933T appears in multiple ST backgrounds and can be chromosomal or plasmid-borne in different isolates, highlighting the overall plasticity of optrA. These findings suggest that certain STs appear predisposed to harbor specific optrA variants in particular genomic contexts, which may influence the stability and transmissibility of linezolid resistance within Enterococcus faecalis populations.
Most E. faecalis isolates shared a core set of virulence factors involved in capsule formation (cps genes), biofilm production (pilus-associated ebp genes and srtC), and proteolytic activity (fsrA/B/C, gelE, sprE). In certain sequence types (e.g., ST585, ST179), the presence of cytolysin genes (cylA/B/M/S/I/R) further enhances their virulence potential. Additionally, elements such as asa1 and prgB/asc10 suggest an increased capacity for aggregation and plasmid transfer. Collectively, these findings underscore the multifaceted pathogenicity of E. faecalis, where adhesion, biofilm formation, exoenzyme production, and, in specific STs, cytolysin-mediated toxicity play key roles in its virulence. The complete set of virulence factors is presented in Table S4 of the supplementary material.
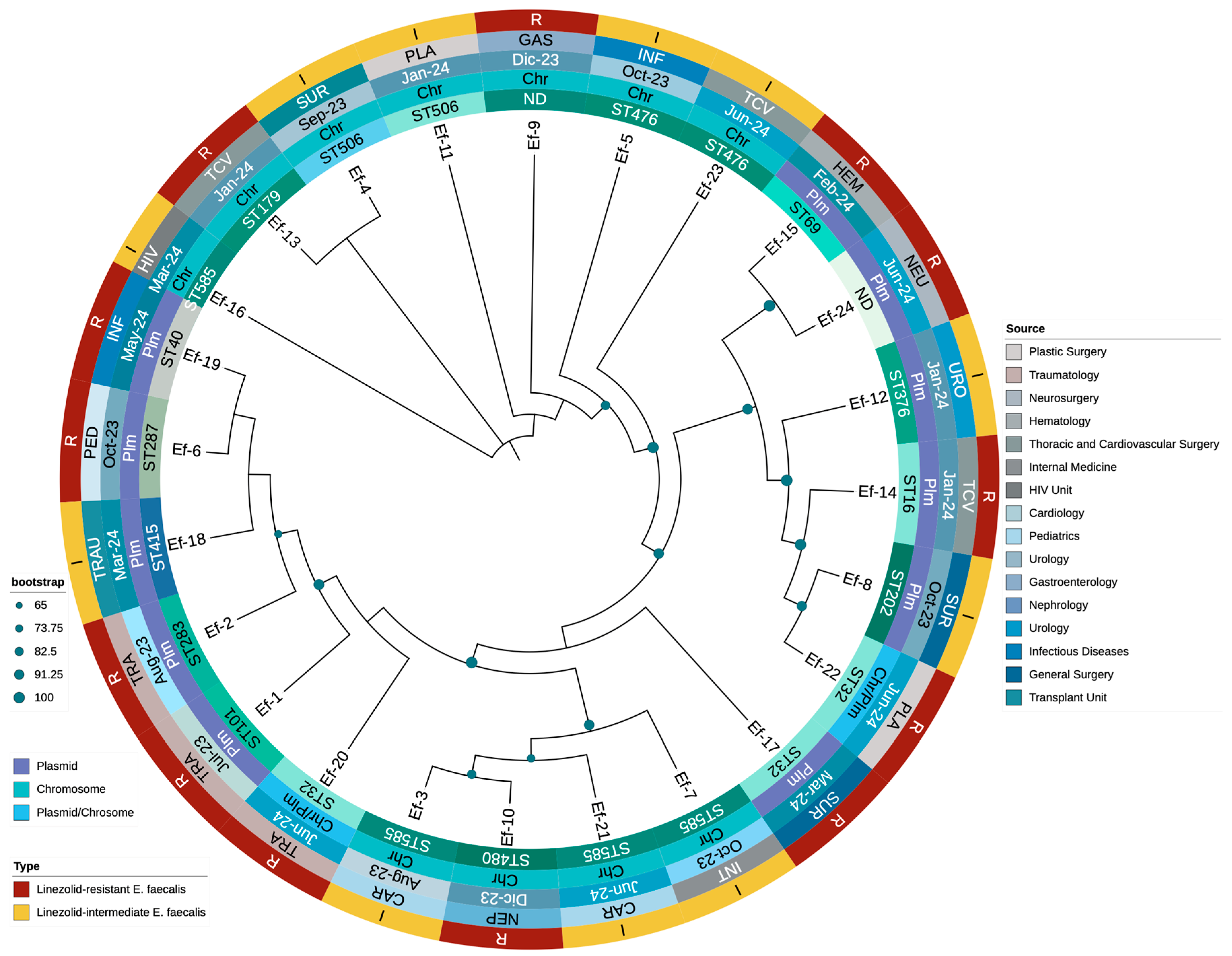
A phylogenetic tree was constructed using whole-genome sequences to explore the evolutionary relationships among the 24 LNSEF strains and 9 LSEF strains. The analysis revealed a diverse phylogenetic structure, with LNSEF isolates distributed across multiple sequence types (STs) and hospital departments, suggesting widespread dissemination. ST32, ST585, and ST476 appeared prominently, while ST101 was observed only once, confirming it is not a major lineage among the LNSEF isolates. Linezolid non-susceptible isolates were scattered across different clades, rather than clustering into a single lineage, suggesting independent emergence of resistance across diverse genetic backgrounds. The presence of the optrA gene was consistently associated with linezolid non-susceptibility, with two isolates also harboring cfrA, suggesting potential horizontal gene transfer. Interestingly, ST32 exhibited two distinct optrA variants: one plasmid-borne (T411G, T626G, G866A), and another chromosomally integrated (T10G, T35A, C54T, A91G, A107G, A134T, T626G, C949T, G1278A, A1331G, A1541C, C1933T), the latter accompanied by cfrA in a plasmid. Additionally, optrA remained chromosomal in all ST585 isolates, whereas ST506 and ST476 consistently carried optrA in the chromosome as well. These findings suggest that specific optrA variants may be preferentially associated with either chromosomal or plasmidic integration, depending on the genetic background. The phylogenetic structure highlights a concerning trend of increasing non-susceptible isolates from late 2023 to mid-2024, likely driven by selective pressure. Additionally, specific optrA mutations (G1879A, C1933T, A1441C, T411G) were detected in distinct STs, supporting the hypothesis of parallel evolution of resistance mechanisms. The association between phylogenetic distribution, resistance profiles, and hospital departments where isolates were identified is presented in Figure 2.
Finally, a phylogenetic tree was constructed using optrA sequences to investigate the evolutionary relationships among LNSEF isolates (Figure 2). The analysis revealed a structured yet diverse distribution of optrA sequences, with distinct clusters corresponding to specific sequence types (STs) and genomic localizations. Different optrA variants evolved independently across multiple genetic backgrounds rather than from a single ancestral strain. Notably, chromosomal optrA variants formed distinct clades separate from plasmid-borne ones, reinforcing the role of genomic context in shaping linezolid non-susceptibility. ST585 and ST506 consistently harbored chromosomal optrA, clustering together despite originating from different hospital units. Conversely, ST32 and ST476 showed a higher frequency of plasmid-associated optrA, suggesting increased horizontal gene transfer potential. Some isolates exhibited both chromosomal and plasmidic optrA variants, particularly within ST32, indicating recombination events or parallel resistance acquisition. This aligns with cfrA co-occurrence in optrA-harboring plasmids, supporting mobile genetic element-mediated dissemination. The diversity of optrA sequences also correlated with functional polymorphisms, with mutations (G1879A, C1933T, A1441C, T411G, T626G) clustering within specific STs. These mutations may modulate linezolid resistance levels, contributing to different phenotypic expressions of non-susceptibility. Overall, the phylogenetic structure highlights the complexity of optrA evolution in Enterococcus faecalis, with multiple independent acquisition events shaping resistance in the hospital setting. The relationship between optrA sequence diversity, genetic background, and resistance mechanisms is detailed in Figure 2.
4. Discussion
This study provides critical insights into the prevalence and mechanisms of linezolid non-susceptibility among Enterococcus faecalis isolates in a Mexican tertiary care university hospital. Among the 392 isolates analyzed, 6.12% (n=24) exhibited linezolid non-susceptibility, including 2.8% (n=11) with intermediate susceptibility and 3.32% (n=13) classified as resistant, primarily mediated by the optrA gene.
According to the medical literature, linezolid resistance in Enterococcus faecalis is generally low but not negligible, with a geographically variable prevalence[10,11,12,13,36,37,38]. Although our prevalence were higher than that reported in some developed countries, it has not yet reached the levels observed in certain regions of South Korea or China [12,13].
Historically, the primary mechanism of linezolid resistance across multiple species (e.g., Staphylococcus aureus, Enterococcus faecium) has been mutations in the domain V region of the 23S rRNA, most commonly G2576T in staphylococci [39,40,41,42]. However, in E. faecalis, the plasmid-borne optrA gene has emerged in recent years as the predominant driver of linezolid nonsusceptibility in many geographic regions [11,12,13,14,15].
Compared to other linezolid resistance mechanisms, such as point mutations in the 23S rRNA or the presence of cfr, the optrA gene provides a significant advantage due to its frequent localization on mobile genetic elements (plasmids or transposons), facilitating horizontal transfer between strains and even across different Enterococcus species [26]. This mobility allows optrA to disseminate rapidly in both clinical and community settings, often co-occurring with additional resistance genes (e.g., fexA) on the same plasmid, thereby expanding the multidrug resistance profile [11,12,13,14,15]. While ribosomal mutations remain chromosomal and non-transferable, and cfr, despite its potential for plasmid localization, appears less prevalent than optrA in E. faecalis isolates worldwide, optrA has emerged as the dominant reservoir for linezolid resistance [11,12,13,14,15].
Consistent with this trend, our data revealed a lower prevalence of cfrA-mediated resistance, with only two isolates carrying this gene, in contrast to the higher rates reported in studies from China and South Korea [12,13]. Additionally, the absence of 23S rRNA mutations differed from findings in European cohorts, suggesting regional variability in resistance mechanisms [43]. No mutations were identified in ribosomal proteins L3 and L4 or in the poxtA gene among our isolates. These findings underscore the predominance of alternative resistance mechanisms and emphasize the need for ongoing surveillance.
Clinical analysis revealed that LNSEF isolates were associated with higher rates of immunosuppression, prior surgical history, and prolonged hospital stays. This aligns with the medical literature, which identifies risk factors for linezolid-resistant enterococcal infections, including prior exposure to linezolid and other antibiotics, prolonged hospitalization, and underlying conditions such as gastrointestinal surgery, urogenital disorders, malignancies, diabetes, and polymicrobial infections [14,44]. These factors create selective pressure, facilitating the emergence and persistence of resistant strains [14,44].
However, none of the patients in our cohort had documented prior linezolid use, suggesting that resistance may spread through mechanisms beyond direct selective pressure from this agent. This finding is consistent with reports of horizontal gene transfer of linezolid-resistance determinants (e.g., optrA) among enterococci, as well as potential cross-resistance arising from prior exposure to other antibiotics [44]. The presence of resistant strains in patients without prior linezolid exposure underscores the need for stringent infection control measures and highlights the potential for resistant clones to circulate within healthcare environments, driven by multiple overlapping risk factors.
The phylogenetic analysis confirmed significant genetic diversity among LNSEF isolates, with 19 distinct STs identified. Notably, certain STs exhibited consistent optrA variants and genomic localization. ST101 and ST202 harbored optrA variants exclusively on plasmids, while ST16 and ST283 carried optrA in plasmids with identical mutations (T626G, A1541C). In contrast, ST179 consistently exhibited the G1879A, C1933T optrA variant in the chromosome. These findings suggest that specific optrA variants may be preferentially associated with either chromosomal or plasmidic integration, potentially influencing the stability and dissemination of linezolid resistance.
Additionally, whole-genome sequencing revealed that LNSEF isolates were widely distributed across hospital departments, supporting both clonal expansion and independent resistance acquisition. The clustering of ST32, ST585, ST101, and ST16 in multiple departments suggests potential nosocomial transmission and highlights the need for targeted infection control strategies.
The identification of diverse sequence types (STs), including ST585 and ST32, supports the polyclonal dissemination of LNSEF within our institution, consistent with reports of Enterococcus faecalis resistant to linezolid harboring optrA and cfr genes in hospital settings [45]. This widespread polyclonal distribution highlights the complexity of managing antibiotic resistance, as it facilitates the persistence and transmission of resistant strains across diverse bacterial populations and hospital environments [46]. The ability of E. faecalis to disseminate resistance through multiple genetic backgrounds poses a significant challenge for infection control, emphasizing the urgent need for enhanced surveillance and containment strategies.
The interplay between optrA polymorphisms, its genetic context (plasmid vs. chromosome), and the co-occurrence of additional resistance determinants, such as cfrA, collectively modulates the degree of linezolid resistance [47,48]. Our findings indicate that the presence of optrA alone does not uniformly confer high-level resistance; instead, resistance levels may be influenced by specific mutations within optrA, its genomic location, and the strain’s background (ST). Isolates harboring optrA exhibit MIC values ranging from 4 µg/mL (intermediate) to ≥8 µg/mL (resistant), consistent with reports that plasmid-borne optrA generally correlates with higher MICs [49]. However, emerging evidence suggests that certain chromosomal variants can also confer resistance ≥8 µg/mL, likely through additional genetic factors or specific sequence types that enhance gene expression [48].
Beyond reduced susceptibility to linezolid, our E. faecalis isolates exhibited extensive resistance to multiple antibiotic classes. High-level aminoglycoside resistance (HLAR), characterized by impaired synergy with gentamicin or streptomycin, correlated with the presence of aac(6')-aph(2''), ant(6)-Ia, and aph(3')-III genes. Fluoroquinolone resistance was associated with gyrA (S83I) and/or parC (S80I) mutations, while near-universal carriage of tet(L) and/or tet(M) explained the widespread resistance to tetracyclines. Additional resistance determinants, including erm(B), cat, fexA, and fosB, were identified as conferring resistance to macrolides, phenicols, and fosfomycin. Despite this extensive multidrug resistance profile, most isolates remained susceptible to ampicillin, penicillin, vancomycin, and daptomycin, reinforcing their continued role as viable therapeutic options. This observation aligns with prior reports showing that E. faecalis isolates carrying linezolid resistance determinants—such as optrA or cfr—frequently retain susceptibility to β-lactams, glycopeptides, and daptomycin [26,28,48]. The mechanisms underlying linezolid resistance (often ribosomal target modifications or efflux) do not typically confer cross-resistance to these alternative antibiotic classes, underscoring the importance of comprehensive susceptibility testing to guide effective treatment choices.
Several limitations should be acknowledged. First, during data collection, some variables were missing and had to be excluded from the analyses. While strict inclusion criteria were applied to ensure data quality, the retrospective nature of clinical and sociodemographic data collection inherently limits the ability to control for all potential confounders. Prospective studies are needed to further evaluate the robustness of factors associated with LNSEf acquisition. Second, confirmatory methods such as broth microdilution or E-tests were not incorporated, limiting the ability to assess resistance levels beyond MICs >8 µg/mL. However, all isolates with linezolid MICs >4 µg/mL harbored genetic determinants associated with resistance. Finally, the single-center nature of this study may limit the generalizability of the results to other settings, particularly those with differing patient demographics, healthcare resources, or antimicrobial resistance patterns.
Our findings emphasize the critical role of genomic surveillance and antimicrobial stewardship programs, particularly in high-risk hospital settings. The identification of novel optrA mutations offers opportunities for further research into their functional implications and potential as therapeutic targets. Future multicenter studies are needed to validate these observations and capture broader epidemiological trends.
Integrating genomic surveillance into routine clinical practice is essential to monitoring and controlling the spread of LNSEf. Strengthening surveillance efforts, implementing robust infection control measures, and optimizing antimicrobial stewardship strategies will be key to limiting the dissemination of linezolid-non-susceptible E. faecalis within healthcare settings.
5. Conclusions
In conclusion, our findings highlight the emerging clinical significance of linezolid-non-susceptible Enterococcus faecalis (LNSEf) within a Mexican tertiary care hospital, where 6.12% of E. faecalis isolates exhibited intermediate or full resistance to linezolid, primarily driven by optrA. The notable sequence type (ST) diversity, along with the presence of both plasmid- and chromosomally encoded optrA variants—occasionally co-occurring with cfr—reflects the complex evolutionary trajectories of these resistant strains. Notably, the absence of prior linezolid exposure in most patients underscores the potential for horizontal gene transfer and highlights the need for enhanced infection control measures. Clinically, LNSEf isolates were more frequently identified in patients with immunosuppression, recent surgical interventions, or prolonged hospital stays, suggesting a multifactorial risk profile beyond direct linezolid usage. Although most LNSEf isolates remained susceptible to ampicillin, vancomycin, and daptomycin, the co-occurrence of additional resistance determinants—such as aac(6’)-aph(2’’), erm(B), and tet(L)—illustrates a broader multidrug-resistant phenotype. These findings underscore the importance of ongoing genomic surveillance, stringent antimicrobial stewardship, and targeted infection control strategies to curb the dissemination of linezolid-non-susceptible enterococci and preserve the efficacy of critical antimicrobials.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1. Sequencing quality metrics for 24 linezolid non-susceptible E. faecalis Isolates. Table S2. Quality Metrics of Enterococcus faecalis Genome Assemblies. Table S3. Clinical and microbiological characteristics of patients with Enterococcus faecalis non-susceptible to linezolid. Table S4. Identification of Antibiotic Resistance Proteins Using BLASTP Analysis in Clinical Isolates. Table S5. Virulence factor profiles of E. faecalis isolates by clinical source and MLST . Figure S1. Antibiotic resistance profile of LNSEf isolates. Table S3. Virulence factor profiles of E. faecalis isolates by clinical source and MLST.
Author Contributions
Conceptualization, JB-R, PM-A, and LP-G; methodology, JCDA-J, VCV-C, BBA-C, and JB-R; software, JB-R, JCDA-J and VCV-C; validation, JB-R and JCDA-J; formal analysis, JB-R, PM-A, LP-G and JCDA-J; investigation, PM-A, AG-Q, CAG-Q, KG-P, EAR-M; resources, VCV-C, JCDA-J, PM-A, JB-R , AG-Q, BBA-C, and CAG-Q; data curation, JB-R, JCDA-J, AG-Q, BBA-C, CAG-Q, KG-P, EAR-M, and LP-G; writing—original draft preparation, PM-A, LP-G, KG-P and EAR-M; writing—review and editing, JB-R and JCDA-J; visualization, JB-R , PM-A and JCDA-J; supervision, JB-R , VCV-C and JCDA-J; project administration, JB-R and JCDA-J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study was approved by the “Comité de ética en investi-gación en ciencias de la salud del Centro Universitario de Tlajomulco, Universidad de Guadalajara” (ethical approval number CUTLAJO/DS/CEICS/020/24) and was conducted in accordance with the principles of the Declaration of Helsinki. All procedures were performed in compliance with national legislation and institutional guidelines.
Informed Consent Statement
As this study was conducted retrospectively and involved only deidentified data, informed consent was waived in accordance with the guidelines of the “Comité de ética en investigación en ciencias de la salud del Centro Universitario de Tlajomulco, Universidad de Guadalajara” (ethical approval number CUTLAJO/DS/CEICS/020/24) and relevant ethical regulations).
Data Availability Statement
The original data presented in this study are openly available through the Zenodo platform at https://doi.org/10.5281/zenodo.14873829. The genome assemblies of the 24 LNSEf strains generated in this study have been deposited in GenBank under BioProject ID PRJNA1217060, with individual genome accession numbers ranging from JBLVLD000000000 to JBLVKX000000000. Additionally, further documentation can be found in the supplementary materials.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABC | ATP-Binding Cassette |
| AMR | Antimicrobial Resistance |
| AMP | Ampicillin |
| ARGs | Antimicrobial Resistance Genes |
| AST | Antimicrobial Susceptibility Testing |
| ATP | Adenosine Triphosphate |
| CARD | Comprehensive Antibiotic Resistance Database |
| CAR | Cardiology |
| cgMLST | Core Genome Multilocus Sequence Typing |
| CIP | Ciprofloxacin |
| CLSI | Clinical and Laboratory Standards Institute |
| CSF | Cerebrospinal Fluid |
| DAP | Daptomycin |
| DNA | Deoxyribonucleic Acid |
| FASTQ | Format for Sequence Data |
| GAS | Gastroenterology |
| GEN | Gentamicin |
| HEM | Hematology |
| HIV | Human Immunodeficiency Virus |
| ICU | Intensive Care Unit |
| INF | Infectious Diseases |
| INT | Internal Medicine |
| LEV | Levofloxacin |
| LNZ | Linezolid |
| LNSEf | Linezolid-Non-Susceptible Enterococcus faecalis |
| LRE | Linezolid-Resistant Enterococcus |
| LSEf | Linezolid-Susceptible Enterococcus faecalis |
| MDR-TB | Multidrug-Resistant Tuberculosis |
| MIC | Minimum Inhibitory Concentration |
| ML | Maximum Likelihood |
| MLST | Multilocus Sequence Typing |
| MRSA | Methicillin-Resistant Staphylococcus aureus |
| NEP | Nephrology |
| NEU | Neurosurgery |
| NIT | Nitrofurantoin |
| NR | Non-Redundant Database |
| PCR | Polymerase Chain Reaction |
| PED | Pediatrics |
| PEN | Benzylpenicillin |
| PLA | Plastic Surgery |
| QC | Quality Control |
| rRNA | Ribosomal Ribonucleic Acid |
| RGI | Resistance Gene Identifier |
| SNP | Single Nucleotide Polymorphism |
| ST | Sequence Type |
| STR | Streptomycin |
| SUR | General Surgery |
| TCV | Thoracic and Cardiovascular Surgery |
| TET | Tetracycline |
| TRA | Traumatology |
| TRAU | Transplant Unit |
| URO | Urology |
| VA | Vancomycin |
| WHO | World Health Organization |
| WGS | Whole-Genome Sequencing |
| SUR | General Surgery |
| TCV | Thoracic and Cardiovascular Surgery |
| TET | Tetracycline |
| TRA | Traumatology |
| TRAU | Transplant Unit |
| URO | Urology |
| VA | Vancomycin |
| VFDB | Virulence Factor Database |
| WHO | World Health Organization |
| WGS | Whole-Genome Sequencing |
References
- Zahedi Bialvaei, A.; Rahbar, M.; Yousefi, M.; Asgharzadeh, M.; Samadi Kafil, H. Linezolid: A Promising Option in the Treatment of Gram-Positives. J Antimicrob Chemother 2017, 72, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Leach, K.L.; Brickner, S.J.; Noe, M.C.; Miller, P.F. Linezolid, the First Oxazolidinone Antibacterial Agent. Ann N Y Acad Sci 2011, 1222, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, S.M.; Farhadi, T.; Ganjparvar, M. Linezolid: A Review of Its Properties, Function, and Use in Critical Care. DDDT 2018, 12, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, J.A.; Kanyo, Z.F.; Wang, D.; Franceschi, F.J.; Moore, P.B.; Steitz, T.A.; Duffy, E.M. Crystal Structure of the Oxazolidinone Antibiotic Linezolid Bound to the 50S Ribosomal Subunit. J. Med. Chem. 2008, 51, 3353–3356. [Google Scholar] [CrossRef]
- Makarov, G.I.; Makarova, T.M. A Noncanonical Binding Site of Linezolid Revealed via Molecular Dynamics Simulations. J Comput Aided Mol Des 2020, 34, 281–291. [Google Scholar] [CrossRef]
- Alloush, H.; Salisbury, V.; Lewis, R.; Macgowan, A. Pharmacodynamics of Linezolid in a Clinical Isolate of Streptococcus Pneumoniae Genetically Modified to Express Lux Genes. The Journal of antimicrobial chemotherapy 2003, 52 3, 511–513. [Google Scholar] [CrossRef]
- Brauers, J.; Kresken, M.; Menke, A.; Orland, A.; Weiher, H.; Morrissey, I. Bactericidal Activity of Daptomycin, Vancomycin, Teicoplanin and Linezolid against STaphylococcus Aureus, Enterococcus Faecalis and Enterococcus Faecium Using Human Peak Free Serum Drug Concentrations. International Journal of Antimicrobial Agents 2007, 29, 322–325. [Google Scholar] [CrossRef]
- Nahid, P.; Mase, S.R.; Migliori, G.B.; Sotgiu, G.; Bothamley, G.H.; Brozek, J.L.; Cattamanchi, A.; Cegielski, J.P.; Chen, L.; Daley, C.L.; et al. Treatment of Drug-Resistant Tuberculosis. An Official ATS/CDC/ERS/IDSA Clinical Practice Guideline. Am J Respir Crit Care Med 2019, 200, e93–e142. [Google Scholar] [CrossRef]
- Welshman, I.R.; Sisson, T.A.; Jungbluth, G.L.; Stalker, D.J.; Hopkins, N.K. Linezolid Absolute Bioavailability and the Effect of Food on Oral Bioavailability. Biopharmaceutics & Drug Disposition 2001, 22, 91–97. [Google Scholar] [CrossRef]
- Gargis, A.S.; Spicer, L.M.; Kent, A.G.; Zhu, W.; Campbell, D.; McAllister, G.; Ewing, T.O.; Albrecht, V.; Stevens, V.A.; Sheth, M.; et al. Sentinel Surveillance Reveals Emerging Daptomycin-Resistant ST736 Enterococcus Faecium and Multiple Mechanisms of Linezolid Resistance in Enterococci in the United States. Frontiers in Microbiology 2022, 12. [Google Scholar] [CrossRef]
- Seyedolmohadesin, M.; Kouhzad, M.; Götz, F.; Ashkani, M.; Aminzadeh, S.; Bostanghadiri, N. Emergence of Lineage ST150 and Linezolid Resistance in Enterococcus Faecalis: A Molecular Epidemiology Study of UTIs in Tehran, Iran. Front Microbiol 2024, 15, 1464691. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Huh, H.J.; Song, D.J.; Shim, H.J.; Park, K.S.; Kang, C.-I.; Ki, C.-S.; Lee, N.Y. Resistance Mechanisms of Linezolid-Nonsusceptible Enterococci in Korea: Low Rate of 23S rRNA Mutations in Enterococcus Faecium. J Med Microbiol 2017, 66, 1730–1735. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, F.; Bai, B.; Lin, Z.; Xu, G.; Chen, Z.; Sun, X.; Zheng, J.; Deng, Q.; Yu, Z. Linezolid Resistance in Enterococcus Faecalis Associated With Urinary Tract Infections of Patients in a Tertiary Hospitals in China: Resistance Mechanisms, Virulence, and Risk Factors. Frontiers in Public Health 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Pan, H.; Lou, Y.; Wu, Z.; Zhang, J.; Huang, Y.; Yu, W.; Qiu, Y. Epidemiological Characteristics and Genetic Structure of Linezolid-Resistant Enterococcus Faecalis. IDR 2018, Volume 11, 2397–2409. [Google Scholar] [CrossRef]
- Hua, R.; Xia, Y.; Wu, W.; Yang, M.; Yan, J. Molecular Epidemiology and Mechanisms of 43 Low-Level Linezolid-Resistant Enterococcus Faecalis Strains in Chongqing, China. Ann Lab Med 2019, 39, 36–42. [Google Scholar] [CrossRef]
- Shen, W.; Hu, Y.; Liu, D.; Wang, Y.; Schwarz, S.; Zhang, R.; Cai, J. Prevalence and Genetic Characterization of Linezolid Resistance Gene Reservoirs in Hospital Sewage from Zhejiang Province, China. Sci Total Environ 2024, 955, 177162. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Xiao, S.; Han, L.; Wu, Q. Antimicrobial Resistance, Virulence Gene Profiles, and Molecular Epidemiology of Enterococcal Isolates from Patients with Urinary Tract Infections in Shanghai, China. Microbiol Spectr 2025, 13, e0121724. [Google Scholar] [CrossRef]
- Brenciani, A.; Cinthi, M.; Coccitto, S.N.; Massacci, F.R.; Albini, E.; Cucco, L.; Paniccià, M.; Freitas, A.R.; Schwarz, S.; Giovanetti, E.; et al. Global Spread of the Linezolid-Resistant Enterococcus Faecalis ST476 Clonal Lineage Carrying optrA. J Antimicrob Chemother 2024, 79, 846–850. [Google Scholar] [CrossRef]
- Shan, X.; Li, C.; Zhang, L.; Zou, C.; Yu, R.; Schwarz, S.; Shang, Y.; Li, D.; Brenciani, A.; Du, X.-D. poxtA Amplification and Mutations in 23S rRNA Confer Enhanced Linezolid Resistance in Enterococcus Faecalis. J Antimicrob Chemother 2024, 79, 3199–3203. [Google Scholar] [CrossRef]
- Jiang, L.; Xie, N.; Chen, M.; Liu, Y.; Wang, S.; Mao, J.; Li, J.; Huang, X. Synergistic Combination of Linezolid and Fosfomycin Closing Each Other’s Mutant Selection Window to Prevent Enterococcal Resistance. Front Microbiol 2020, 11, 605962. [Google Scholar] [CrossRef]
- Cavaco, L.M.; Bernal, J.F.; Zankari, E.; Léon, M.; Hendriksen, R.S.; Perez-Gutierrez, E.; Aarestrup, F.M.; Donado-Godoy, P. Detection of Linezolid Resistance Due to the optrA Gene in Enterococcus Faecalis from Poultry Meat from the American Continent (Colombia). Journal of Antimicrobial Chemotherapy 2017, 72, 678–683. [Google Scholar] [CrossRef]
- Long, K.S.; Vester, B. Resistance to Linezolid Caused by Modifications at Its Binding Site on the Ribosome. Antimicrob Agents Chemother 2012, 56, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Long, K.S.; Munck, C.; Andersen, T.M.B.; Schaub, M.A.; Hobbie, S.N.; Böttger, E.C.; Vester, B. Mutations in 23S rRNA at the Peptidyl Transferase Center and Their Relationship to Linezolid Binding and Cross-Resistance. Antimicrob Agents Chemother 2010, 54, 4705–4713. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, M.; Sawant, A.R.; Prashanth, K.; Sistla, S. Multiple Mechanisms of Linezolid Resistance in Staphylococcus Haemolyticus Detected by Whole-Genome Sequencing. J Med Microbiol 2023, 72. [Google Scholar] [CrossRef]
- Locke, J.B.; Morales, G.; Hilgers, M.; G C, K.; Rahawi, S.; José Picazo, J.; Shaw, K.J.; Stein, J.L. Elevated Linezolid Resistance in Clinical Cfr-Positive Staphylococcus Aureus Isolates Is Associated with Co-Occurring Mutations in Ribosomal Protein L3. Antimicrob Agents Chemother 2010, 54, 5352–5355. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A Novel Gene, optrA, That Confers Transferable Resistance to Oxazolidinones and Phenicols and Its Presence in Enterococcus Faecalis and Enterococcus Faecium of Human and Animal Origin. J Antimicrob Chemother 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Deng, Z.; Shen, Y.; Wei, W.; Xiang, Q.; Liu, Z.; Hanf, K.; Huang, S.; Lv, Z.; Cao, T.; et al. High Prevalence and Plasmidome Diversity of optrA-Positive Enterococci in a Shenzhen Community, China. Front Microbiol 2024, 15, 1505107. [Google Scholar] [CrossRef]
- Cai, J.; Schwarz, S.; Chi, D.; Wang, Z.; Zhang, R.; Wang, Y. Faecal Carriage of optrA-Positive Enterococci in Asymptomatic Healthy Humans in Hangzhou, China. Clin Microbiol Infect 2019, 25, 630.e1–630.e6. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, K.; Liu, Z.; Li, Y.; Xiao, X.; Du, X.-D.; Li, R.; Wang, Z. Genomic Insights into Linezolid-Resistant Enterococci Revealed Its Evolutionary Diversity and poxtA Copy Number Heterogeneity. Int J Antimicrob Agents 2023, 62, 106929. [Google Scholar] [CrossRef]
- Sun, W.; Liu, H.; Liu, J.; Jiang, Q.; Pan, Y.; Yang, Y.; Zhu, X.; Ge, J. Detection of optrA and poxtA Genes in Linezolid-Resistant Enterococcus Isolates from Fur Animals in China. Lett Appl Microbiol 2022, 75, 1590–1595. [Google Scholar] [CrossRef]
- Hou, J.; Xu, Q.; Zhou, L.; Chai, J.; Lin, L.; Ma, C.; Zhu, Y.; Zhang, W. Identification of an Enterococcus Faecium Strain Isolated from Raw Bovine Milk Co-Harbouring the Oxazolidinone Resistance Genes optrA and poxtA in China. Vet Microbiol 2024, 293, 110103. [Google Scholar] [CrossRef] [PubMed]
- LaMarre, J.; Mendes, R.E.; Szal, T.; Schwarz, S.; Jones, R.N.; Mankin, A.S. The Genetic Environment of the Cfr Gene and the Presence of Other Mechanisms Account for the Very High Linezolid Resistance of Staphylococcus Epidermidis Isolate 426-3147L. Antimicrob Agents Chemother 2013, 57, 1173–1179. [Google Scholar] [CrossRef]
- Shore, A.C.; Lazaris, A.; Kinnevey, P.M.; Brennan, O.M.; Brennan, G.I.; O’Connell, B.; Feßler, A.T.; Schwarz, S.; Coleman, D.C. First Report of Cfr-Carrying Plasmids in the Pandemic Sequence Type 22 Methicillin-Resistant Staphylococcus Aureus Staphylococcal Cassette Chromosome Mec Type IV Clone. Antimicrob Agents Chemother 2016, 60, 3007–3015. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Zhang, W.; Du, X.-D.; Krüger, H.; Feßler, A.T.; Ma, S.; Zhu, Y.; Wu, C.; Shen, J.; Wang, Y. Mobile Oxazolidinone Resistance Genes in Gram-Positive and Gram-Negative Bacteria. Clin Microbiol Rev 2021, 34, e0018820. [Google Scholar] [CrossRef]
- M100Ed35 | Performance Standards for Antimicrobial Susceptibility Testing, 35th Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 28 January 2025).
- Kent, A.G.; Spicer, L.M.; Campbell, D.; Breaker, E.; McAllister, G.A.; Ewing, T.O.; Longo, C.; Balbuena, R.; Burroughs, M.; Burgin, A.; et al. Sentinel Surveillance Reveals Phylogenetic Diversity and Detection of Linear Plasmids Harboring vanA and optrA among Enterococci Collected in the United States. Antimicrob Agents Chemother 2024, 68, e0059124. [Google Scholar] [CrossRef]
- Yang, W.; Li, X.; Chen, J.; Zhang, G.; Li, J.; Zhang, J.; Wang, T.; Kang, W.; Gao, H.; Zhang, Z.; et al. Multicentre Evaluation of in Vitro Activity of Contezolid against Drug-Resistant Staphylococcus and Enterococcus. J Antimicrob Chemother 2024, 79, 3132–3141. [Google Scholar] [CrossRef]
- Dadashi, M.; Sharifian, P.; Bostanshirin, N.; Hajikhani, B.; Bostanghadiri, N.; Khosravi-Dehaghi, N.; Belkum, A. van; Darban-Sarokhalil, D. The Global Prevalence of Daptomycin, Tigecycline, and Linezolid-Resistant Enterococcus Faecalis and Enterococcus Faecium Strains From Human Clinical Samples: A Systematic Review and Meta-Analysis. Frontiers in Medicine 2021, 8. [Google Scholar] [CrossRef]
- Quiles-Melero, I.; Gómez-Gil, R.; Romero-Gómez, M.P.; Sánchez-Díaz, A.M.; de Pablos, M.; García-Rodriguez, J.; Gutiérrez, A.; Mingorance, J. Mechanisms of Linezolid Resistance among Staphylococci in a Tertiary Hospital. J Clin Microbiol 2013, 51, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Mendes, R.E.; Streit, J.M.; Hogan, P.A.; Flamm, R.K. Five-Year Summary of In Vitro Activity and Resistance Mechanisms of Linezolid against Clinically Important Gram-Positive Cocci in the United States from the LEADER Surveillance Program (2011 to 2015). Antimicrob Agents Chemother 2017, 61, e00609-17. [Google Scholar] [CrossRef]
- Besier, S.; Ludwig, A.; Zander, J.; Brade, V.; Wichelhaus, T.A. Linezolid Resistance in Staphylococcus Aureus: Gene Dosage Effect, Stability, Fitness Costs, and Cross-Resistances. Antimicrob Agents Chemother 2008, 52, 1570–1572. [Google Scholar] [CrossRef]
- Alonso, M.; Marín, M.; Iglesias, C.; Cercenado, E.; Bouza, E.; García de Viedma, D. Rapid Identification of Linezolid Resistance in Enterococcus Spp. Based on High-Resolution Melting Analysis. J Microbiol Methods 2014, 98, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Linezolid-Resistant Enterococci in Polish Hospitals: Species, Clonality and Determinants of Linezolid Resistance. Eur J Clin Microbiol Infect Dis 2017, 36, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, G.; Li, J.; Chen, L.; Liu, H.; Bi, W.; Lu, H.; Zhou, T. A High Incidence and Coexistence of Multiresistance Genes Cfr and optrA among Linezolid-Resistant Enterococci Isolated from a Teaching Hospital in Wenzhou, China. Eur J Clin Microbiol Infect Dis 2018, 37, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lucas, C.; Fernández, J.; Vázquez, X.; de Toro, M.; Ladero, V.; Fuster, C.; Rodicio, R.; Rodicio, M.R. Detection of the optrA Gene Among Polyclonal Linezolid-Susceptible Isolates of Enterococcus Faecalis Recovered from Community Patients. Microbial Drug Resistance 2022, 28, 773–779. [Google Scholar] [CrossRef]
- Egan, S.A.; Shore, A.C.; O’Connell, B.; Brennan, G.I.; Coleman, D.C. Linezolid Resistance in Enterococcus Faecium and Enterococcus Faecalis from Hospitalized Patients in Ireland: High Prevalence of the MDR Genes optrA and poxtA in Isolates with Diverse Genetic Backgrounds. Journal of Antimicrobial Chemotherapy 2020, 75, 1704–1711. [Google Scholar] [CrossRef]
- He, T.; Shen, Y.; Schwarz, S.; Cai, J.; Lv, Y.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J.; et al. Genetic Environment of the Transferable Oxazolidinone/Phenicol Resistance Gene optrA in Enterococcus Faecalis Isolates of Human and Animal Origin. J Antimicrob Chemother 2016, 71, 1466–1473. [Google Scholar] [CrossRef]
- Li, D.; Wang, Y.; Schwarz, S.; Cai, J.; Fan, R.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J. Co-Location of the Oxazolidinone Resistance Genes optrA and Cfr on a Multiresistance Plasmid from Staphylococcus Sciuri. J Antimicrob Chemother 2016, 71, 1474–1478. [Google Scholar] [CrossRef]
- Yoon, S.; Son, S.H.; Kim, Y.B.; Seo, K.W.; Lee, Y.J. Molecular Characteristics of optrA-Carrying Enterococcus Faecalis from Chicken Meat in South Korea. Poult Sci 2020, 99, 6990–6996. [Google Scholar] [CrossRef]
Figure 1.
Study workflow for whole-genome sequencing and characterization of Enterococcus faecalis isolates.
Figure 1.
Study workflow for whole-genome sequencing and characterization of Enterococcus faecalis isolates.
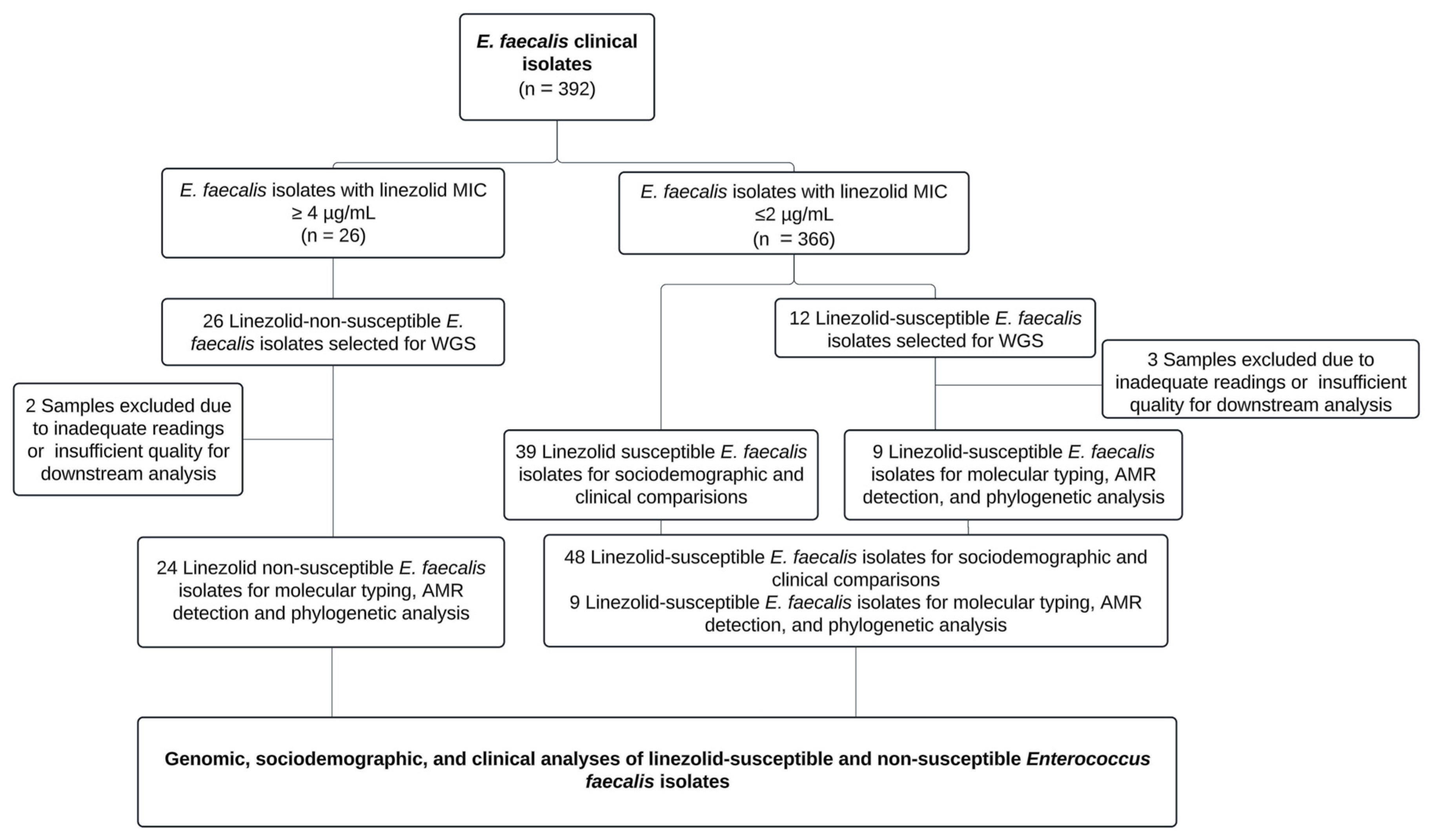
Figure 2.
Phylogenetic distribution, resistance profile, and clinical origin of Enterococcus faecalis isolates resistant and susceptible to linezolid during the study period.
Figure 2.
Phylogenetic distribution, resistance profile, and clinical origin of Enterococcus faecalis isolates resistant and susceptible to linezolid during the study period.
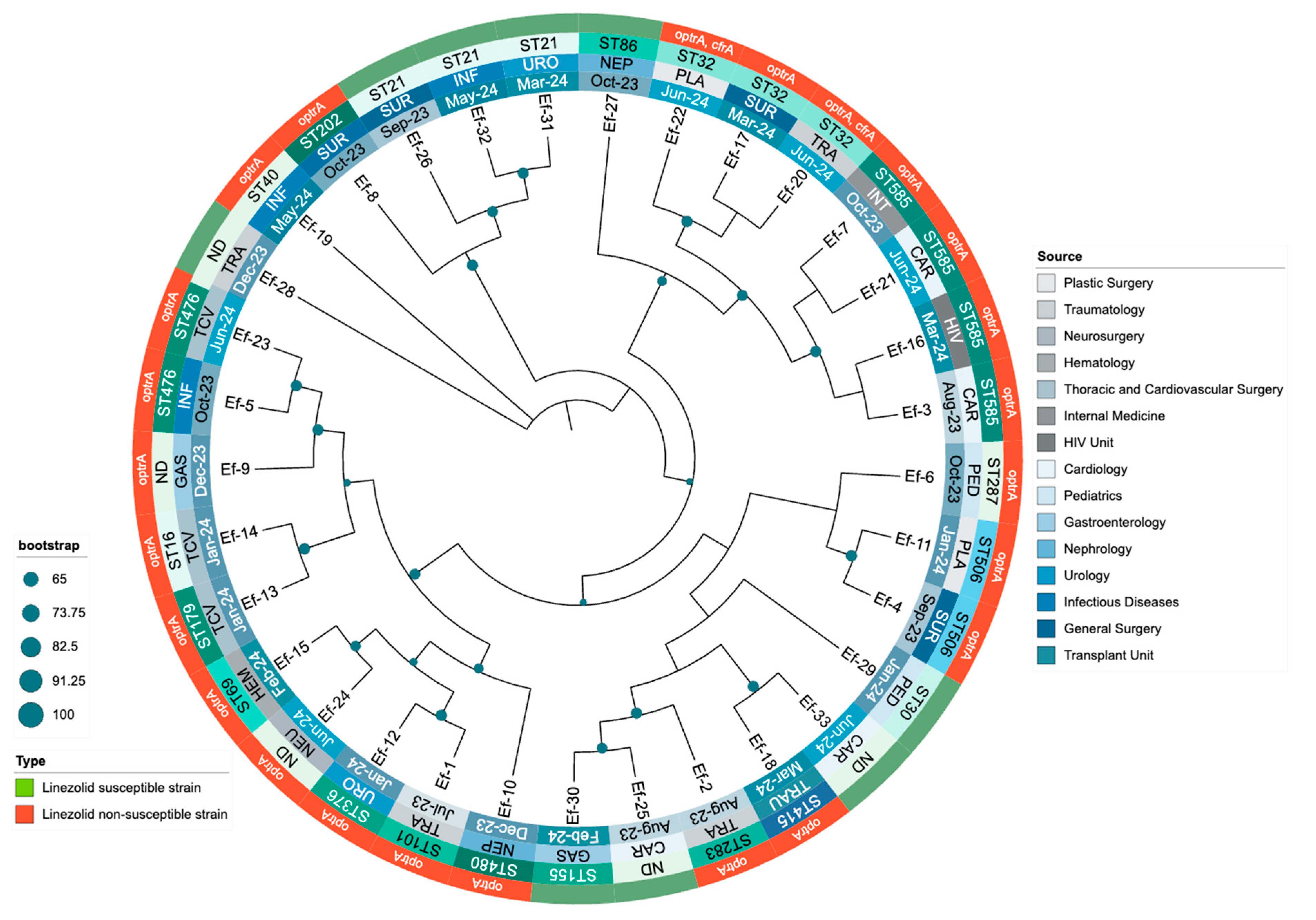
Table 2.
Sociodemographic and clinical characteristics of patients with linezolid-susceptible and linezolid-non-susceptible Enterococcus faecalis isolates.
Table 2.
Sociodemographic and clinical characteristics of patients with linezolid-susceptible and linezolid-non-susceptible Enterococcus faecalis isolates.
| Variable | Total (n = 72) |
LSEf (n = 48) |
LNSEf (n = 24) |
p value |
|---|---|---|---|---|
| Age – median (IQR) | 48 (31.25-57) | 48 (31.25-57.25) | 48 (31-57) | 0.9096 |
| Male sex – n (%) | 51 (70.8) | 34 (66.7) | 17 (70.8) | 0.928 |
| Previous antibiotic use in 90 days – n (%) | 34 (47.2) | 23 (47.9) | 11 (45.8) | 1 |
| Previous Antibiotic in 14 days – n (%) | 29 (40.3) | 20 (41.7) | 9 (37.5) | 0.932 |
| Previous hospitalization in 90 days – n (%) | 28 (38.9) | 15 (31.2) | 13 (54.2) | 0.104 |
| Comorbidities – n (%) | 40 (55.6) | 23 (47.9) | 17 (70.8) | 0.111 |
| Charlson comorbidity index – median (IQR) | 1 (0–3) | 1.5 (0–3) | 1 (0–3) | 0.905 |
| Diabetes Mellitus – n (%) | 15 (20.8) | 9 (19.6) | 6 (25) | 0.826 |
| Hypertension – n (%) | 15 (20.8) | 10 (20.8) | 5 (20.8) | 1 |
| Cardiovascular disease – n (%) | 0 (0) | 0 (0) | 0 (0) | 1 |
| Obesity – n (%) | 2 (2.8) | 1 (2.1) | 1 (4.2) | 1 |
| Immunosuppression – n (%) | 3 (4.2) | 0 (0) | 3 (12.5) | 0.033 |
| Chronic kidney disease – n (%) | 10 (13.9) | 8 (16.7) | 2 (8.3) | 0.478 |
| Surgical Intervention – n (%) | 27 (37.5) | 11 (22.9) | 16 (66.7) | <0.001 |
| Central line insertion – n (%) | 25 (34.8) | 15 (31.2) | 10 (41.7) | 0.540 |
| Urinary catheter – n (%) | 40 (55.6) | 23 (47.9) | 17 (70.8) | 0.111 |
| Need for vasopressors – n (%) | 14 (19.4) | 7 (14.6) | 7 (29.2) | 0.246 |
| ICU admission – n (%) | 11 (15.3) | 8 (16.7) | 3 (13) | 1 |
| Hospital stay – median (IQR) | 12 (4–17.25) | 9.5 (3.75–15) | 14 (11.25–20.25) | 0.02 |
| In-hospital mortality – n (%) | 12 (16.7) | 8 (16.7) | 4 (16.7) | 1 |
Table 3.
Phenotypic and genotypic profiles of 24 linezolid-non-susceptible Enterococcus faecalis isolates.
Table 3.
Phenotypic and genotypic profiles of 24 linezolid-non-susceptible Enterococcus faecalis isolates.
| Label | Date | Source | AMP | CIP | DAP | NIT | HLG | HLS | LEV | LNZ | PEN | TET | VA | MLST | Genes associated with linezolid resistance | Locations of linezolid resistance Genes | Other AMR genes | Other identified plasmids |
| Ef-1 | Jul-23 | Traumatology | <=2 | 1 | 4 | <=16 | SYN-S | SYN-S | 1 | >=8 | 2 | >=16 | 1 | ST101 | optrA (T10G, T35A, C54T, A91G, A107G, T626G, C949T, A1966G) | Plasmid | erm(B), fexA, lsa(A), NarA, NarB, tet(L) | repUS43 |
| Ef-2 | Aug-23 | Traumatology | <=2 | 1 | 1 | <=16 | SYN-S | SYN-R | 1 | >=8 | 2 | >=16 | <=0.5 | ST283 | optrA (T626G, A1541C) | Plasmid | ant(6)-Ia, aph(2'')-Ic, aph(3')-III, dfrG, erm(A), erm(B), fexA, lsa(A), tet(L), tet(M) | rep2, rep6, rep9b |
| Ef-3 | Aug-23 | Cardiology | <=2 | >=8 | 2 | <=16 | SYN-R | SYN-R | >=8 | 4 | 2 | >=16 | 1 | ST585 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(6)-Ia, ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E),, parC (S80I), str, tet(L) | rep7a, repUS43 |
| Ef-4 | Sep-23 | General Surgery | <=2 | 1 | 4 | <=16 | SYN-S | SYN-S | 1 | 4 | 1 | >=16 | 1 | ST506 | optrA (G1879A, C1933T) | Chromosome | ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lsa(A), tet(L) | - |
| Ef-5 | Oct-23 | Infectious Diseases | <=2 | >=8 | 2 | <=16 | SYN-R | SYN-S | >=8 | 4 | 2 | <=1 | 1 | ST476 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, gyrA (S83I), lsa(A), parC (S80I | - |
| Ef-6 | Oct-23 | Pediatrics | <=2 | 4 | 2 | <=16 | SYN-R | SYN-R | 4 | >=8 | 2 | >=16 | 1 | ST287 | optrA (G1879A, C1933T) | Plasmid | aac(6')-aph(2''), ant(6)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lsa(A), NarA, NarB, tet(L) | rep1, rep9b |
| Ef-7 | Oct-23 | Internal Medicine | <=2 | >=8 | 1 | <=16 | SYN-R | SYN-S | >=8 | 4 | 2 | >=16 | 1 | ST585 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, dfrG, erm(A), erm(B), fexA, lsa(A), parC (S80I), tet(L) | repUS43 |
| Ef-8 | Oct-23 | General Surgery | <=2 | 4 | 2 | <=16 | SYN-S | SYN-S | 4 | 4 | 2 | >=16 | 1 | ST202 | optrA (T10G, T35A, C54T, A91G, A107G, T626G, C949T, A1966G) | Plasmid | ant(9)-Ia, dfrG, erm(B), erm(B), fexA, fosB, lsa(A), NarA, NarB, tet(L) | - |
| Ef-9 | Dec-23 | Gastroenterology | <=2 | >=8 | 2 | <=16 | SYN-R | SYN-S | >=8 | >=8 | 1 | >=16 | 1 | ND | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, dfrG, erm(A), erm(B), fexA, fosB, gyrA (S83I), lsa(A), NarA, NarB, parC (S80I), tet(L) | - |
| Ef-10 | Dec-23 | Nephrology | <=2 | >=8 | 2 | <=16 | SYN-S | SYN-S | >=8 | >=8 | 2 | >=16 | 1 | ST480 | optrA ((G1879A, C1933T) | Chromosome | ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lsa(A), NarA, NarB, tet(L) | repUS43 |
| Ef-11 | Jan-24 | Plastic Surgery | <=2 | 1 | 2 | <=16 | SYN-R | SYN-R | 1 | 4 | 1 | >=16 | <=0.5 | ST506 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, dfrG, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E),tet(L) | repUS43 |
| Ef-12 | Jan-24 | Urology | <=2 | 4 | 2 | <=16 | SYN-R | SYN-R | 2 | 4 | 8 | >=16 | 1 | ST376 | optrA (T411G, T626G, G866A) | Plasmid | aac(6')-aph(2''), ant(6)-Ia, ant(6)-Ia, aph(3')-III, cat, dfrG, erm(A), fexA, lnu(B), lsa(A), lsa(E), tet(L) | repUS43 |
| Ef-13 | Jan-24 | Thoracic and Cardiovascular Surgery | <=2 | <=0.5 | 4 | <=16 | SYN-R | SYN-R | 1 | >=8 | 2 | >=16 | 1 | ST179 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E), tet(M) | repUS43 |
| Ef-14 | Jan-24 | Thoracic and Cardiovascular Surgery | <=2 | <=0.5 | 2 | <=16 | SYN-R | SYN-R | 1 | >=8 | 2 | >=16 | 1 | ST16 | optrA (T626G, A1541C) | Plasmid | aac(6')-aph(2''), aph(3')-III, dfrG, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E), tet(M)c | rep9b, repUS43 |
| Ef-15 | Feb-24 | Hematology | <=2 | >=8 | 2 | <=16 | SYN-S | SYN-S | >=8 | >=8 | 2 | >=16 | <=0.5 | ST69 | optrA (T411G, T626G, G866A) | Plasmid | cat, dfrG, erm(B), fexA, lsa(A), parC | rep22, repUS43 |
| Ef-16 | Mar-24 | HIV Unit | <=2 | >=8 | 1 | <=16 | SYN-R | SYN-R | >=8 | 4 | 2 | >=16 | <=0.5 | ST585 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, dfrG, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E), parC (S80I), str, tet(L) | rep7a, repUS43 |
| Ef-17 | Mar-24 | General Surgery | <=2 | >=8 | 4 | <=16 | SYN-S | SYN-S | >=8 | >=8 | 2 | >=16 | <=0.5 | ST32 | optrA (T411G, T626G, G866A) | Plasmid | ant(6)-Ia, aph(3')-III, erm(A), erm(B), fexA, gyrA (S83I), lsa(A), parC (S80I), tet(L) | repUS43 |
| Ef-18 | Mar-24 | Transplant Unit | <=2 | 4 | 2 | <=16 | SYN-S | SYN-S | 4 | 4 | 2 | >=16 | 2 | ST415 | optrA (T411G, T626G, G866A) | Plasmid | cat, dfrG, erm(A), erm(B), fexA, fosB3, lsa(A), NarA, NarB, tet(L) | rep1, rep9c |
| Ef-19 | May-24 | Infectious Diseases | <=2 | <=0.5 | 2 | <=16 | SYN-R | SYN-R | 0.5 | >=8 | 2 | >=16 | 1 | ST40 | optrA (T411G, T626G, G866A) | Plasmid | aac(6')-aph(2''), ant(6)-Ia, ant(6)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lnu(B), lsa(A), lsa(E), tet(L) | rep18b, rep9b, repUS43 |
| Ef-20 | Jun-24 | Traumatology | <=2 | 4 | 2 | <=16 | SYN-R | SYN-R | >=8 | >=8 | 2 | >=16 | 1 | ST32 | optrA (T10G, T35A, C54T, A91G, A107G, A134T, T626G, C949T, G1278A, A1331G, A1541C, C1933T ), cfrA | optrA Chromosome, cfrA plasmid | aadD, ant(9)-Ia, aph(2'')-Ic, aph(3')-III, bleO, cat, dfrG, erm(A), erm(B), fexA, fosB3, lnu(A), lsa(A), tet(L) | repUS43 |
| Ef-21 | Jun-24 | Cardiology | <=2 | >=8 | 1 | <=16 | SYN-R | SYN-S | >=8 | 4 | 2 | >=16 | 1 | ST585 | optrA (G1879A, C1933T) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, cat, dfrG, erm(A), fexA, lnu(B), lsa(A), lsa(E), parC (S80I), str, tet(L) | rep7a, repUS43 |
| Ef-22 | Jun-24 | Plastic Surgery | <=2 | 4 | 2 | <=16 | SYN-R | SYN-R | >=8 | >=8 | 2 | >=16 | 1 | ST32 | optrA (T10G, T35A, C54T, A91G, A107G, A134T, T626G, C949T, G1278A, A1331G, A1541C, C1933T), cfrA | optrA Chromosome, cfrA plasmid | aadD, ant(9)-Ia, aph(2'')-Ic, aph(3')-III, bleO, cat, dfrG, erm(A), erm(B), fexA, fosB3, lnu(A), lsa(A), tet(L) | repUS43 |
| Ef-23 | Jun-24 | Thoracic and Cardiovascular Surgery | <=2 | >=8 | 2 | <=16 | SYN-R | SYN-S | >=8 | 4 | 2 | <=1 | 1 | ST476 | optrA (T411G, T626G, G866A) | Chromosome | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, gyrA (S83I), lnu(B), lsa(A), lsa(E), parC (S80I) | rep9b, rep9c |
| Ef-24 | Jun-24 | Neurosurgery | <=2 | <=0.5 | 2 | <=16 | SYN-R | SYN-R | 0.5 | >=8 | 2 | >=16 | 1 | ND | optrA (T411G, T626G, G866A) | Plasmid | aac(6')-aph(2''), ant(6)-Ia, ant(9)-Ia, aph(3')-III, dfrG, erm(A), erm(B), fexA, lsa(A), parC (S80I), tet(L) | repUS43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



