
Submitted:
04 December 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
Background/Objectives: Multidrug-resistant (MDR) infections pose a significant public health threat by worsening patient outcomes, contributing to hospital outbreaks, and increasing the health and economic burden. Advanced genomic tools enhance the detection of resistance genes, virulence factors, and high-risk clones, thus improving the management of MDR infections. In the Autonomous Community of Aragon, the carbapenemase-producing Enterobacteriaceae (CPE) diversity and incidence have increased during the last years. This study analyses CPE trends at a tertiary hospital in Spain from 2021-2023, aiming to optimize personalised medicine. Methods: CPE isolates were the first isolate per patient, year, species, and carbapenemase from January 2021 to December 2023. Antibiotic susceptibility testing was performed by broth microdilution method. Whole genome sequencing (WGS) was performed using Illumina. The genomes were thoroughly analysed to determine their complete taxonomic classification, resistoma and plasmidome composition, and sequence typing using a suite of bioinformatics approaches. Results: Between 2021 and 2023, 0.4% of all isolates were CPE. The CPE rate tripled in 2022 and doubled again in 2023. The most common species was Klebsiella pneumoniae (51.8%) and the most common carbapenemase was blaoxa-48. WGS revealed concordant species identification and detailed carbapenemase distribution. Resistance rates to critical antibiotics such as carbapenems were variable but in most cases above 70%. Genetic diversity was observed in WGS and phylogenetic analyses, with plasmids often mediating carbapenemase dissemination. Conclusions: The increasing rate of CPE in healthcare settings highlights a critical public health challenge, with limited treatment options. Genomic characterization is essential to understand resistance mechanisms, aid therapy, limit outbreaks and improve precision medicine.
Keywords:
Carbapenem-Resistant Enterobacteriaceae
; Whole Genome Sequencing
; Drug Resistance
; Microbial
; Computational biology
1. Introduction
Infections caused by multidrug-resistant (MDR) microorganisms represent a significant global public health issue [1,2]. The isolation of MDR microorganisms from clinical or epidemiological samples often correlates with poorer prognoses and increased morbidity, particularly when other virulence factors are present [3]. Furthermore, many of these microorganisms have the ability to spread within communities and hospitals, leading to outbreaks and epidemics [4,5]. Accurate characterisation of resistance determinants, virulence factors, mobile genetic elements, and the clonal relationships among isolates can be achieved using high-throughput sequencing and bioinformatics tools. This enables the detection and tracking of high-risk clones, phenotypic correlation with antimicrobial susceptibility testing, and control of hospital outbreaks. These results complement those obtained by traditional methods and have proven useful in the management of infections and outbreaks caused by MDR microorganisms worldwide, leading to their increasingly wide spread use in clinical microbiology and public health increasingly widespread [6,7].
In 2017, the World Health Organization (WHO) published a priority list of pathogens to guide research and development of new antibiotics, categorising carbapenemase-producing Enterobacteriaceae (CPE) as critical priority [8]. CPE are characterized by causing difficult-to-treat infections, increased morbidity and mortality, and high transmissibility, thus necessitating their active surveillance. Additionally, their prevalence is high in certain countries, increasing the likelihood of transmission, particularly in hospital settings [9].
In 2021, as part of the National Plan against Antibiotic Resistance (PRAN) in Spain, the Network of Laboratories for the Surveillance of Resistant Microorganisms (RedLabRA) was created , with the primary objective of achieving comprehensive and high quality microbiological diagnostics [10]. This network integrates genomic sequencing in all cases of infection or colonisation by antibiotic-resistant microorganisms under surveillance in the National Health System (currently including Escherichia coli, Klebsiella pneumoniae, and Enterobacter cloacae complex producing carbapenemases).
In the Autonomous Community of Aragón, Spain, the isolation of CPE has been sporadic and was previously limited to OXA-48-producing K. pneumoniae, leading to hospital outbreaks that were identified and contained [11]. However, in recent years, both the number and diversity of bacterial species and carbapenemases have steadily increased, creating new paradigms for their surveillance, detection, treatment, and interdisciplinary communication. These findings may be related to the aftermath of the COVID-19 pandemic and the forced displacement caused by the war in Ukraine [12,13], which has altered our hospital’s medical care and referral system of military and civilian. In this sense, the Miguel Servet University Hospital (HUMS) has deemed it essential to monitor CPE and vancomycin-resistant Enterococcus due to the increasing incidence and impact of their spread in the community.
The integration of genomic data analysis into clinical microbiology, with a translational perspective, could yield results that add significant value to the care of patients with MDR microbial infections and advance the pursuit of personalized medicine [14].
In the ever-evolving landscape of global healthcare, the rise of multidrug resistant bacteria has become a pressing concern, posing a significant threat to public health and challenging the efficacy of conventional antimicrobial treatments. This research delves into the prevalence and characteristics of CPE in a tertiary hospital in Spain over the course of a three-year period, providing a comprehensive analysis of its typing and antimicrobial resistance determinants.
2. Results
2.1. Case Selection and Species Identification
From January 2021 to December 2023, a total of 38,145 Enterobacteriaceae were isolated in the clinical microbiology laboratory at HUMS. Approximately 97% of these (36,956 isolates), were obtained from clinical samples, with Escherichia coli (60.5%), Klebsiella pneumoniae (17.1%), Proteus mirabilis (6%), and Enterobacter cloacae complex (4.1%) being the most prevalent species. During this period, 112 CPE were identified, representing 0.4% of the total isolates. Besides, the distribution of both the total number of isolates and CPE was not uniform throughout the study period, as there was a consistent increase in the total number of enterobacteria and CPE. Specifically, the total number of enterobacteria increased by 18.1% in 2022 compared to 2021 and by 19.3% in 2023 compared to the previous year. However, the increase in CPE was greater than what could be explained solely by the increase in the total number of enterobacteria isolates, almost tripling in 2022 compared to 2021, representing 0.2% (p value <0.01) of the total isolates, and doubling again in 2023, representing 0.5% (p value <0.01) of total enterobacteria. Total isolations per species, year and presence of carbapenemase is presented in Table 1.
The criteria for selecting CPE isolates were the first isolate per patient, year, species, and type of carbapenemase. Consequently, the 112 CPE isolates corresponded to 93 unique patients, of whom 74.1% were male. Patients' age ranged from 0.6 to 90.7 years, with a median age of 51.2 years. Epidemiological samples, mainly rectal and perianal swabs, accounted for 68% of the isolates, while the remaining 32% were from clinical samples. Among the clinical samples, approximately 50% were urine, 25% were from surgical wounds, and the remaining 25% were from abscesses, tracheal exudates, other wounds types, ascitic fluid, and blood. Most isolates were obtained from hospitalised patients, with 17% from patients admitted to an intensive care unit. Moreover, 66% of the isolates were from HUMS, and the remaining 34% were from other hospitals for which HUMS serves as a reference centre. 110 ff the 112 CPE, had reported antibiotic susceptibility testing, and 91 were available for sequencing (81%). For the remaining 21 strains, there was no viable sample to re-culture for sequencing.
K. pneumoniae was the most common CPE species, accounting for 51.8% of the isolates, followed by E. coli (18.8%), Citrobacter spp (16.1%), Enterobacter cloacae complex (7.1%), Providencia stuartii (2.7%), Klebsiella oxytoca (1.8%), Proteus mirabilis (1%), and Serratia marcescens (1%). This species distribution was comparable in the subset of CPE isolates that underwent WGS, consisting of K. pneumoniae (n=53), E. coli(n=21), E. cloacae (n=7), Citrobacter spp (n=7), P. stuartii (n=3), K. oxytoca (n=2) and P. mirabilis (n=1)
WGS yielded concordant species identification compared to Matrix-assisted laser desorption ionization time of flight (MALDI-TOF), while providing enhanced resolution to distinguish closely related species such as K. oxytoca and K. michiganensis or within complexes such as in Citrobacter freundii complex, in which two isolates were reclassified as Citrobacter cronae and Citrobacter partucalensis.
2.2. Antibiotic Susceptibility
The antibiotic susceptibility analysis showed high rates of resistance among CPE isolates. Resistance to cephalosporins such as ceftazidime and cefepime was observed in 77.5% and 84.3% of isolates, respectively. For carbapenems, resistance to ertapenem, imipenem, and meropenem reached 92.1%, 76.4%, and 68.5%, respectively. Approximately three-quarters of the isolates were resistant to the fluoroquinolones ciprofloxacin and levofloxacin. Aminoglycoside resistance ranged from 52.8% for gentamicin to 64% for amikacin and 75.3% for tobramycin. Resistance to the "last-resort" antibiotics fosfomycin, tigecycline, and colistin was 39.3%, 33.3%, and 10.6%, respectively. Additionally, the new generation antibiotics ceftolozane-tazobactam, ceftazidime-avibactam, and cefiderocol showed non-susceptibility rates of 77.5%, 41.6%, and 31.7%, respectively. These resistance patterns were analysed considering the intrinsic resistance profiles of the Enterobacteriaceae species evaluated, such as the presence of chromosomal AmpC in some of them. The fosfomycin results were limited to E. coli and Klebsiella spp., and the tigecycline cutoff was based on the E. coli interpretation. A detailed description of the antibiotic susceptibility pattern by carbapenemase type in each isolate is shown in Figure 1.
2.3. Resistome
The resistome annotation results encompassed all resistance mechanisms detected in the genomes. However, the analysis focused only on those mechanisms involved in direct modification of the antibiotic molecule or its target site of action, excluding resistance mechanisms related to cell membrane permeability, active efflux, and lipopolysaccharide profile.
In total, 101 carbapenemase-encoding genes were identified from the 91 genomes sequenced. The most common carbapenemase found was OXA-48-like, represented by 50 isolates with different alleles of the blaoxa gene i.e blaOXA-48 (38 isolates), blaOXA-181 (5 isolates), blaOXA-244 (5 isolates), and blaOXA-484 (2 isolates). This was followed by NDM, with two alleles present, blaNDM-1 (15 isolates) and blaNDM-5 (9 isolates). The third most common was the VIM-type carbapenemase, mainly blaVIM-1 (13 isolates). The least frequent was the class A KPC carbapenemase, with a similar distribution between blaKPC-2 (7 isolates) and blaKPC-3 (6 isolates) alleles. Notably, ten strains were found to harbor dual carbapenemase genes, comprising blaNDM-1 + blaOXA-48 (7 isolates), blaNDM-1 + blaOXA-244 (2 isolates), and blaNDM-1 + blaKPC-2 (1 isolate).
Regarding other β-lactamases, extended-spectrum β-lactamase genes were detected in 47.3% of the isolates, with the CTX-M-type being the most common, including blaCTX-M-15, blaCTX-M-9, and blaCTX-M-55 alleles. Interestingly, the gene blaVEB-6 was detected in P. mirabilis. No mutations at positions 238 or 179 in the blaSHV gene, conferring resistance to third-generation cephalosporins [15] were observed. Because their epidemiological importance, the presence of class C β-lactamase genes in the genomes of E. coli and K. pneumoniae was evaluated, excluding the blaEC gene due to its potential chromosomal nature in E. coli. Several alleles of the blaCMY gene were detected in seven E. coli strains, and blaDHA-1 was found in one K. pneumoniae strain. Other β-lactamases detected in the isolates, mainly class A and D with lower hydrolytic profiles, included multiple alleles of blaSHV, blaTEM, and blaOXA genes. In particular, blaLEN-16 was associated with blaVIM-24 and blaCTX-M-9 in one isolate of Klebsiella variicola. Only three strains exhibited a carbapenemase gene as the sole β-lactamase in their genome, two Providencia stuartii blaNDM-5 and one Citrobacter amalonaticus blaVIM-1. Table 2 shows carbapenemases types by species.
Table 2 shows the distribution of carbapenemases according to the host microorganism. Total number of carbapenemases and Enterobacteriaceae ae showed in the last column and row, respectively.
Regarding fluoroquinolone resistance, 82.4% of the strains harboured genetic resistance determinants, including point mutations in the gyrA, gyrB, or parC genes, as well as the presence of the qnr gene, mainly multiple alleles of qnrB, and lower frequencies of qnrA, qnrD, and qnrS. The concurrent presence of at least one mutation in gyrA and parC, observed in 55 isolates, conferred phenotypic resistance in 100% of the cases. In contrast, the isolated presence of the qnr gene in 13 isolates resulted in variable levels of resistance and discrepant phenotypes, with some strains being ciprofloxacin-resistant but levofloxacin-sensitive. Additionally, six isolates had mutations in a single gene, either gyrA or gyrB, while 29 isolates had mutations in gyrA/gyrB, parC, and the simultaneous presence of qnr. The most common amino acid variant in GyrA was the substitution of serine for isoleucine at position 83, accounting for 43.9% of the cases, with less frequent changes to phenylalanine, leucine, and tyrosine at the same position, as well as other substitutions at position 87. Notably, the D87G substitution exhibited a fluoroquinolone-resistant phenotype, even in the absence of other resistance determinants. Substitutions in GyrB involved the change of glutamate to aspartate and serine to alanine at positions 463 and 466, respectively, the former being found only in the absence of gyrA mutations and conferring a resistant phenotype only when combined with the presence of the qnr gene. All substitutions in ParC corresponded to a change from serine to isoleucine at position 80, and no strains presented isolated mutations in the parC gene.
Genes encoding aminoglycoside-modifying enzymes were detected in approximately 90% of the isolates, with acetyltransferases being the most common type, followed by nucleotidyltransferases and phosphotransferases. More than one of these enzyme coding genes were found in two-thirds of the isolates, and 30% harboured three or more. The presence of the gene encoding AAC6'-Ib, an enzyme of epidemiological and clinical importance due to amikacin inactivation [16], was specifically investigated and detected in 56 isolates, 50% of which exhibited in vitro resistance to amikacin. The high diversity in the presence and combinations of genes of aminoglycoside-modifying enzymes made it challenging to correlate with specific in vitro susceptibility patterns to gentamicin, tobramycin, and amikacin, which were variable.
2.4. Typing
Multilocus sequence typing analysis revealed the presence of predominant sequence types among K. pneumoniae isolates, while the less common species exhibited greater genetic diversity. The most prevalent K. pneumoniae sequence type was ST147, followed by ST307 and ST395. Additionally, two isolates of the hypervirulent K. pneumoniae sequence type ST23 [17] were identified, which also harboured dual carbapenemase genes. In the case of E. coli, according to the Achtman MLST scheme [18], only two sequence types, ST2659 and ST167, were detected in two or more isolates, with four and two isolates, respectively. Interestingly, one K. michiganensis isolate showed a potential new sequence type, characterised by a G159T transversion in the mdh gene, representing a previously undescribed allele [19].
The cgMLST analysis of the K. pneumoniae isolates, using a more relaxed threshold of 15 different alleles to distinguish individual clusters given the temporal separation of the isolates [20,21,22], revealed a high degree of genetic diversity Figure 2. The analysis identified nine clusters containing at least two isolates, but only two of these clusters included four or more isolates. Notably, there was no clear clonal relationship between isolates expressing the same carbapenemase gene, with the exception of the blaOXA-181 variant, where the four identified strains showed a close genetic relationship. Interestingly, four of the strains harboring the dual carbapenemase genes blaNDM-1 and blaOXA-48 were grouped within the largest cluster, which was predominantly composed of the ST147 sequence type. In contrast, the E. coli analysis, using the same clustering threshold, identified only two clusters that directly corresponded to the observed sequence types Figure3. Importantly, these E. coli clusters exhibited a clear association between the isolates and the specific carbapenemase genes, with the main cluster carrying blaNDM-5 and the secondary cluster carrying the dual carbapenemase genes blaNDM-1 and blaOXA-244. Notably, two isolates harbouring blaOXA-484 were separated only by 35 alleles in the cgMLST and one of them was catalogued as a possible new ST due to a mismatch in the adk gene that needs to be confirmed by resequencing, being the closest match ST1722.
Phylogenetic analysis of the K. pneumoniae isolates revealed a distribution similar to the minimum spanning tree generated from cgMLST (Figure 4 and Figure 5). However, a group of strains, in particular those belonging to sequence type ST512 and producing KPC-3 carbapenemase as well as a clade of heterogeneous ST395 isolates expressing diverse carbapenemase types, exhibited a large divergence in their ancestry, taking into account that the reference genome is ST11. Notably, two isolates (KP835819 and KP963668) belonging to the same high-risk ST (395) and harbouring a blaoxa-48 carbapenemase clustered together, although the patients had no epidemiological link to each other and the isolates were collected more than six months apart, highlighting the benefits of cgMLST for epidemiological tracking of CPE. Phylogenetic analysis of the E. coli isolates demonstrated a clear separation between strains producing VIM-1 and NDM-1 metallo-β-lactamases compared to the rest of the cohort. In addition, the formation of phylogenetically related clades was observed, taht were not apparent in the cgMLST analysis, particularly among strains harboring blaKPC and blaOXA-484 genes. The significance of these phylogenetic relationships would be determined by the epidemiological context surrounding the strains.
2.5. Plasmids
The plasmid analysis focused on those associated with the presence of carbapenemase genes. Plasmids harbouring carbapenemase genes were identified in 93.4% of the isolates. For the blaOXA-48 gene, the predominant replicon type was IncL, with an average size of 50kb, of which 50% were predicted to be conjugative, while the remaining 50% were non-mobilizable. In contrast, the blaNDM-1 gene was primarily associated with IncF replicon type, which had an average size of 91.9kb, and 78.5% were conjugative, compared to 21.5% that were non-mobilizable. Overall, the most common plasmid replicon types were IncL and IncF, accounting for 60% of the samples. The average size of plasmids harbouring carbapenemase genes was 81Kb, ranging from 4.3Kb (blaOXA-48) to 342.9Kb (blaNDM-1), with 61.1% being conjugative, 4.7% mobilizable, and 34.1% non-mobilizable. Interestingly, more than one replicon was identified in 44.7% of the cases, and all plasmids larger than 80kb were predicted to be mobile.
Table 3 summarizes the molecular characterization of the CPE grouped by species.
In double carbapenemase carriers, the plasmid description refers to the one harbouring the metallobetalactamase gene. BLEE: Extended spectrum betalactamase.
3. Discussion
From 2021 onward, a consistent and progressive annual increase of approximately 20% in the total number of Enterobacteriaceae isolates was observed in our hospital. This upward trend appeared to be strongly correlated with an increase in the volume of laboratory samples, particularly those sourced from primary care settings. This rise followed the reactivation of clinical services after the gradual de-escalation of COVID-19 restrictions. Notably, this surge in isolates was not observed in epidemiological samples collected between 2021 and 2022. During this period, the majority of epidemiological samples were collected from intensive care units that remained operational despite the pandemic-related restrictions, potentially limiting the increase. However, the significant escalation of Enterobacteriaceae isolates observed in 2023 may probably be attributed to the implementation and strengthening of surveillance programmes focused on multidrug-resistant (MDR) organisms in the community [23]. These programmes, introduced in 2022, played a crucial role in enhancing the detection and surveillance of MDR pathogens, thereby contributing to the increased identification of Enterobacteriaceae isolates in the following year.
In addition to the overall increase in Enterobacteriaceae isolates, the rise in CPE isolates was particularly pronounced, even after adjusting for the general increase in reported isolates over the same period. This significant increase highlight a notable shift in the local epidemiology of CPE, which may be attributed to several factors, including the introduction of strains from high-prevalence regions and/or the enhanced surveillance efforts focused on multidrug-resistant (MDR) organisms within hospital settings [24,25,26]. The species distribution of CPE isolates also differed significantly from global patterns, with K. pneumoniae being the predominant species, followed by E. coli, Citrobacter spp., and E. cloacae complex. This deviation from global trends has been documented in other studies and may reflect species-specific traits that enhance the ability of these organisms to adapt to hospital environments [27,28]. In addition, the prevalence of carbapenemase production in these species may be facilitated by horizontal gene transfer mechanisms, that promote the spread of resistance traits, further contributing to the evolving local epidemiology of CPE [29,30,31].
Although the majority of CPE isolates were obtained from epidemiological samples, it is noteworthy that several patients developed infections requiring antibiotic treatment for the same CPE strains they initially carried [32,33]. However, this clinical progression is not fully captured in the data, as only the first isolate received in the laboratory was included in the analysis. This methodological limitation means that subsequent infections or isolates from the same patients, which could reflect the development of clinically significant infections or changes in resistance profiles, were not accounted for. Consequently, the data may not fully reflect the dynamic nature of CPE colonization and infection in the patient population over time.
Consistent with findings from previous studies [34,35], a high level of resistance to the major classes of antibiotics commonly used in clinical practice was observed, particularly against third- and fourth-generation cephalosporins [36,37], fluoroquinolones [38], and aminoglycosides [39]. This pattern of multidrug resistance is likely attributed to several interrelated mechanisms, including the hydrolytic activity of carbapenemases that, target and degrade other beta-lactams, the co-occurrence of genes encoding extended-spectrum beta-lactamases (ESBLs), and aminoglycoside-modifying enzymes on either the same or separate plasmids. Additionally, mutations in the gyrA and parC genes, which are selected by prior fluoroquinolones exposure, contribute to resistance [40]. Interestingly, the relatively low rate of meropenem resistance observed in our CPE isolates may be explained by the high prevalence of class D carbapenemases. These enzymes show variable sensitivity to meropenem, which, although effective against certain carbapenemases, remains an unsuitable therapeutic option due to its reduced efficacy when facing these enzymes, despite showing low minimum inhibitory concentrations (MICs) [41]. Resistance to new-generation cephalosporins was also notable, with a particularly high rate of resistance to ceftolozane-tazobactam. This resistance is probably due to the limited activity of ceftolozane-tazobactam in isolates expressing carbapenemases, despite its potent anti-Pseudomonas activity [42]. In contrast, resistance to ceftazidime-avibactam was found to be consistent with the prevalence of metallo-beta-lactamases, as avibactam is effective against class A and D carbapenemases but lacks activity against class B enzymes [43]. In contrast to the findings reported by Lasarte-Monterrubio et al. in other isolates from Spanish hospitals, the high rates of cefiderocol-resistant strains observed in this study is remarkable. This is particularly noteworthy given that cefiderocol, a recent addition to the antibacterial armamentarium, is especially intended to treat multidrug-resistant strains infections, including CPE. Mutations in genes that regulate iron transport and the expres-sion of the carbapenemase VIM-1 have been proposed as factors contributing to cefidero-col resistance [44]. However, due to limited knowledge of resistance determinants and the need for strain-specific analysis, these factors were not included in the resistome analysis, though further investigation in future studies would be very valuable.
Despite fosfomycin and tigecycline exhibited lower resistance rates compared to other antibiotics, more than one-third of isolates were non-susceptible to these agents. These antibiotics, along with colistimethate, have limited therapeutic efficacy and a higher risk of adverse effects [45], underscoring the need for tailored treatment strategies when managing CPE infections. Given these challenges, antibiotic susceptibility testing and carbapenemase characterization remain essential to guide appropriate and effective therapeutic interventions in the management of CPE infections.
The high prevalence of K. pneumoniae producing OXA-48 carbapenemase observed in this study is consistent with both local and national epidemiological data, as reported by the Spanish Society of Infectious Diseases and Clinical Microbiology [46]. However, an emerging concern is the increasing diversity of carbapenemase types over time, including several alleles of blaOXA-48 as well as class A and B carbapenemases. This trend highlights the evolving complexity of resistance mechanisms within the local epidemiology of multidrug-resistant organisms. Notably, no single carbapenemase type was found to be exclusive to a particular species, suggesting a broader dissemination of these resistance traits across different bacterial species. This widespread distribution of carbapenemase-producing strains further emphasises the need for active surveillance and comprehensive strategies to mitigate the spread of resistance.
The identification of key sequence types (STs) in Klebsiella pneumoniae, particularly high-risk clones such as ST147 and ST307, is consistent with previous reports of global outbreaks and the spread of carbapenemase-producing strains [47]. These STs are well-documented in the spread of carbapenemase resistance, highlighting the global nature of this problem [48,49]. Notably, two strains belonging to the hypervirulent ST23 clone were found to exhibit dual carbapenemase production, specifically blaNDM-1 and blaOXA-48, located on separate plasmids. This is the first reported instance of such an association in ST23, demonstrating the evolving complexity of resistance mechanisms and the need for enhanced genomic surveillance programmes in healthcare settings. The emergence of multidrug-resistant hypervirulent strains is of particular concern, as these isolates are associated with increased morbidity and mortality, representing a significant public health threat [50,51].
Interestingly, in contrast to the findings of other epidemiological surveillance studies on carbapenem-resistant Escherichia coli, which commonly identify ST131, ST38, and ST405 as predominant clones [52,53], our study revealed a different pattern. The isolates in this study exhibited marked heterogeneity in sequence types, with ST2659 being the most prevalent. Recently described in Algeria and Nepal [54,55], ST2659 is associated with blaNDM alleles and belongs to clonal complex 38, a group that has been linked to the global dissemination of carbapenemase-producing bacteria. The presence of ST2659 in this context further emphasises the dynamic and evolving nature of carbapenemase-producing bacteria, and the global spread of resistance determinants across different bacterial species and geographical regions.
Our study underlines the widespread presence of plasmids harboring carbapenemase genes, with 93.4% of isolates carrying such plasmids. The absence of plasmid identification in the remaining isolates probably reflects the limitations of only short-read sequencing for draft assembly. These plasmids may have been split into multiple fragments in smaller contigs during assembly or may be present in low copy numbers within the isolates. It is plausible that the combination of current data with long-read sequencing could facilitate the accurate characterisation of the mobile elements carrying the carbapenemase genes in these isolates. The predominant replicon types for blaOXA-48 and blaNDM-1 were IncL and IncF, respectively, with different mobilisation characteristics. Notably, 50% of blaOXA-48 associated plasmids were non-mobilizable, whereas a significant 78.5% of blaNDM-1 plasmids were conjugative. The identification of multiple replicons in nearly half of the cases highlights the complexity of carbapenemase gene transfer. The high mobility of plasmids larger than 80kb suggests the potential for rapid horizontal gene transfer, which may contribute to the spread of resistance in clinical settings.
4. Materials and Methods
4.1. Strains Included in the Study and Criteria of Selection of CPE
Data on the number, species, and antibiotic susceptibility profiles of Enterobacteriaceae isolates obtained at HUMS from January 2021 to December 2023 were extracted from the laboratory's electronic records (HORUS).
Antibiotic susceptibility testing was performed by broth microdilution method, using the MicroScan™ WalkAway semi-automated system (Beckman Coulter) with the exception of cefiderocol testing that was performed by Kirby-Bauer method. Results were interpreted according to the The European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines [56]. Inconclusive results were confirmed by Kirby-Bauer or MIC test strip methods (Liofilchem®).
All CPE isolates were recovered from human clinical or epidemiological samples submitted to the HUMS Microbiology Laboratory as part of routine diagnostic procedures.
Screening for carbapenemase production was performed in all strains that presented a MIC of meropenem and or ertapenem >=0.125 mg/L [57]. CPE in clinical samples were identified either by immunochromatographic assays NG-Test® CARBA 5 (NG-Biotech Laboratories) or genotypically by isothermal amplification techniques (Eazyplex® AmplexDiagnostics GmbH) or FilmArray® (BioFire Diagnostics), depending on the specific case and in accordance with laboratory protocols. All strains with a positive molecular result for carbapenemase detection were considered CPE, regardless of their minimum inhibitory concentration to carbapenems.
Epidemiological samples were initially screened for CPE using chromogenic selective media Brilliance™ CRE (Oxoid Limited), followed by a lateral flow assay NG-Test® CARBA 5 (NG-Biotech Laboratories) or real-time polymerase chain reaction (Xpert® Carba-R, Cepheid) confirmation of positive isolates.
Species-level identification was conducted using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight (MALDI-TOF) (Bruker Daltronik GmbH) from solid medium cultures, following manufacturer instructions.
4.2. WGS of CPE Isolates
All identified CPE isolates were candidate for WGS. To increase the available biomass, a subculture was prepared on Columbia blood. Genomic DNA was then extracted from the subculture using a magnetic capture-based method with a MagCore® system (RBC Bioscience), following the manufacturer's protocol, yielding 60 µL of eluate. The extracted genomic DNA was used to construct sequencing libraries using the Nextera XT™ Illumina® kit. Sample quantity and quality was verified at each step of the library preparation process using Qubit™ (Thermo Fisher Scientific, Inc) fluorometric quantification and Bioanalyzer™ (Agilent Technologies, Inc) analysis. Sequencing was performed on an Illumina® MiSeq™ instrument using MiSeq V2 300 reagent cartridges, employing a 150-base paired-end protocol and an expected average sequencing depth of greater than 30x.
De novo assembly was performed using Unicycler v0.5.0 [58], and the structural and functional quality of the assemblies was evaluated using QUAST v5.2.0 [59] and BUSCO v5.6.1 [60], respectively. Samples were rejected based on the following quality criteria: fewer than 800,000 reads with a quality score below 28 on the Phred scale, more than 10% Ns, N50 less than 30,000, or a genome size outside the range of 5.5 Mb ± 1.5 Mb. The presence of contaminants in the raw reads and assembled genomes was assessed using Mash v2.3 [61] and GUNC v1.0 [62], respectively. Finally, the assembly graphs were manually inspected using Bandage v0.8.1 [63]. Draft assemblies are under BioProject accession: PRJNA1190923
Species-level identification was verified using GAMBIT v1.0.1 [64] and the online PubMLST species identification service (https://pubmlst.org/species-id). Multi-locus sequence typing (MLST) was performed using mlst v2.23.0 [65], except for K. pneumoniae, which was analyzed separately. MLST and annotation of antimicrobial resistance and virulence genes for Klebsiella sp was performed with kleborate v2.4.1 [66]. Structural annotation was performed with prokka v1.14.6 [67], functional annotation with sma3s v2 [68] (Uniref90 database), and resistance mechanisms were annotated using rgi v6.0.3 [69] (CARD v3.2.9 database) and abricate v1.0.1 [70,71] (ResFinder database). Resistance mechanisms with coverage >80% and identity >95% were considered.
The phylogenetic analysis was based on the alignment of single nucleotide variants (SNVs) identified between the studied samples and the NCBI reference genome assemblies GCF_000240185.1 and GCF_000005845.2 for K. pneumoniae and for E. coli, respectively. Mapping, variant calling, and coreSNV calculation were performed using snippy v4.6.0 [72]. Recombination events were managed using the gubbins software v2.3 [73]. Phylogenetic inference and reconstruction were conducted using a maximum likelihood approach, with 1000 ultrafast bootstrap replicates and 1000 approximate likelihood-ratio tests. The substitution models employed were TVM+F+ASC+R3 for E. coli and TVM+F+ASC+G4 for K. pneumoniae, implemented in iqtree v2.2.6 [74,75,76].
Multilocus sequence typing of the core genome (gcMLST) was carried out for E. coli and K. pneumoniae using chewBBACA v3.3.1 [77] and the core genome schemes available from the website https://cgmlst.org/ncs website. Training data files were generated using prodigal v2.6.3 [78] with the same reference sequences as those employed in the phylogenetic analysis. The results of the cgMLST analysis were then visualised by constructing a minimum spanning tree, created using the MSTreeV2 algorithm within the grapetree v2.2 software [79].
The plasmidome was annotated by homologous sequence comparisons using the abricate v1.0.1 [70,80] software (PlasmidFinder database). Reconstruction and assembly of the plasmidome were conducted using MOB-suite v3.1.8 [81] and plasmidSPAdes v3.15.5 [82]. Further plasmidome annotation was performed using bakta v1.8.2 [83] (DB light v5.0 database) and rgi 6.0.3 [69], with visualization in the proksee software [84].
Nucleotide identity analyses were carried out employing fastANI v1.34 [85].
Inferential statistical analysis (Z-test to evaluate the difference between proportions) was performed using the prop_test function of the rstatix package in R v4.4.2.
5. Conclusions
The escalating prevalence of CPE in healthcare settings is an immediate and pressing public health challenge. This trend is particularly alarming given the limited therapeutic options available to treat CPE infections, which are often resistant to second- and third-line therapies. In light of our findings, the increase in CPE prevalence, from 0.2% to 0.5% of total Enterobacteriaceae isolates between 2021 and 2023, signals the need for ongoing and systematic surveillance to understand and address the dynamics of resistance. In this context, genomic characterisation, in particular by WGS, is essential. This work also highlights the valuable information that WGS of CPE in a tertiary hospital setting could provide, including an in-depth understanding of the genetic determinants of resistance, high-resolution species identification and delineation of strain-level relationships, which in turn can be used by clinicians and public health authorities to guide therapy and take action to prevent or mitigate potential outbreaks.
Author Contributions
Conceptualization, CT, AR and AT.; methodology, AT, CT, AR, AL and AM; software, AT and LF; validation, CT, AR, AM, AL, BF, CL and RM; formal analysis, AT, LF, MA, CT, AR, ML, LC and RM; investigation, AT, LF, MA, AL and AM; resources, AR, AM and AL; data curation, AT, LF, MA and RM; writing—original draft preparation, AT, CT, AR, AL, AM, ML, LC; writing—review and editing, AT, CT, AR, ML, LC; visualization, AT, ML, LC; supervision, AR, CT; project administration, AT, AR. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the local institutional ethics committee “Comité de Ética de la Investigación de la Comunidad Autónoma de Aragón (CEICA)”, with protocol code PI24/155 and approval date 17/04/2024 in record Nº 08/2024. The study was conducted in accordance with local regulations (Spain): The project is designed in compliance with the requirements of Law 14/2007, of 3 July, on Biomedical Research, and the applicable ethical principles.
Informed Consent Statement
Exemption of informed consent for the collection of retrospective data was accepted by the local institutional ethics committee “Comité de Ética de la Investigación de la Comunidad Autó-noma de Aragón (CEICA)” protocol code PI24/155 and approval date 17/04/2024 in record Nº 08/2024.
Data Availability Statement
Data were submitted to GenBank on 11/26/2024 under ID SUB14891856. Approval and assignment of accession numbers are pending. If necessary, a complete list of GenBank and/or BioSample accessions can be provided in a supplementary table. BioProject accession: PRJNA1190923
Acknowledgments
We would like to thank the Microbiology Laboratory of HUMS, especially the laboratory technicians, for their assistance in processing samples for clinical diagnostic purposes. We would also like to extend our gratitude to the Infectious Diseases Department for their close collaboration with the laboratory in the treatment of infections caused by CPE. Finally, we would like to thanks Sandra Martínez of the PhD program on Biomedical and Biotechnological Sciences of the “Universidad de la Rioja” for her valuable and insightful perspective on the bioinformatic methods.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Van Duin D, Doi Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence. 2017 May 19;8(4):460–9.
- Bonomo RA, Burd EM, Conly J, Limbago BM, Poirel L, Segre JA, et al. Carbapenemase-Producing Organisms: A Global Scourge. Clin Infect Dis. 2018 Apr 3;66(8):1290–7. [CrossRef]
- Kim D, Park BY, Choi MH, Yoon EJ, Lee H, Lee KJ, et al. Antimicrobial resistance and virulence factors of Klebsiella pneumoniae affecting 30 day mortality in patients with bloodstream infection. J Antimicrob Chemother [Internet]. 2018 Oct 5 [cited 2024 Nov 23]; Available from: https://academic.oup.com/jac/advance-article/doi/10.1093/jac/dky397/5116214. [CrossRef]
- Wielders CCH, Schouls LM, Woudt SHS, Notermans DW, Hendrickx APA, Bakker J, et al. Epidemiology of carbapenem-resistant and carbapenemase-producing Enterobacterales in the Netherlands 2017–2019. Antimicrob Resist Infect Control. 2022 Dec;11(1):57. [CrossRef]
- van Duin D, Paterson D. Multidrug Resistant Bacteria in the Community: Trends and Lessons Learned. Infect Dis Clin North Am. 2016 Jun;30(2):377–90. [CrossRef]
- Manageiro V, Cano M, Furtado C, Iglesias C, Reis L, Vieira P, et al. Genomic and epidemiological insight of an outbreak of carbapenemase-producing Enterobacterales in a Portuguese hospital with the emergence of the new KPC-124. J Infect Public Health. 2024 Mar;17(3):386–95. [CrossRef]
- Regad M, Lizon J, Alauzet C, Roth-Guepin G, Bonmati C, Pagliuca S, et al. Outbreak of carbapenemase-producing Citrobacter farmeri in an intensive care haematology department linked to a persistent wastewater reservoir in one hospital room, France, 2019 to 2022. Eurosurveillance [Internet]. 2024 Apr 4 [cited 2024 Nov 23];29(14). Available from: https://www.eurosurveillance.org/content/10.2807/1560-7917.ES.2024.29.14.2300386. [CrossRef]
- Prioritization of pathogens to guide discovery, research and development of new antibiotics for drug-resistant bacterial infections, including tuberculosis. Geneva: World Health Organization; 2017(WHO/EMP/IAU/2017.12).
- European Centre for Disease Prevention and Control. Risk assessment on the spread of carbapenemase-producing Enterobacteriaceae (CPE) : through patient transfer between healthcare facilities, with special emphasis on cross-border transfer [Internet]. LU: Publications Office; 2011 [cited 2024 Nov 23]. Available from: https://data.europa.eu/doi/10.2900/59034.
- Agencia Española de Medicamentos y Productos Sanitarios. Plan Nacional Resistencia Antibióticos PRAN [Internet]. PRAN. [cited 2024 Oct 31]. Available from: https://www.resistenciaantibioticos.es/es.
- Gómara M, López-Calleja AI, Iglesia BMPV, Cerón IF, López AR, Pinilla MJR. Detection of carbapenemases and other mechanisms of enzymatic resistance to β-lactams in Enterobacteriaceae with diminished susceptibility to carbapenems in a tertiary care hospital. Enfermedades Infecc Microbiol Clínica. 2018 May;36(5):296–301.
- Petrosillo N, Petersen E, Antoniak S. Ukraine war and antimicrobial resistance. Lancet Infect Dis. 2023 Jun;23(6):653–4. [CrossRef]
- Pallett SJC, Boyd SE, O’Shea MK, Martin J, Jenkins DR, Hutley EJ. The contribution of human conflict to the development of antimicrobial resistance. Commun Med. 2023 Oct 25;3(1):153. [CrossRef]
- Watkins RR. Antibiotic stewardship in the era of precision medicine. JAC-Antimicrob Resist. 2022 Jun 21;4(3):dlac066. [CrossRef]
- Tsang KK, Lam MMC, Wick RR, Wyres KL, Bachman M, Baker S, et al. Diversity, functional classification and genotyping of SHV β-lactamases in Klebsiella pneumoniae [Internet]. Genomics; 2024 [cited 2024 Nov 24]. Available from: http://biorxiv.org/lookup/doi/10.1101/2024.04.05.587953.
- Ramirez MS, Tolmasky ME. Aminoglycoside modifying enzymes. Drug Resist Updat. 2010 Dec;13(6):151–71.
- Wang S, Ding Q, Zhang Y, Zhang A, Wang Q, Wang R, et al. Evolution of Virulence, Fitness, and Carbapenem Resistance Transmission in ST23 Hypervirulent Klebsiella pneumoniae with the Capsular Polysaccharide Synthesis Gene wcaJ Inserted via Insertion Sequence Elements. Garcia-Solache MA, editor. Microbiol Spectr. 2022 Dec 21;10(6):e02400-22.
- Laukkanen-Ninios R, Didelot X, Jolley KA, Morelli G, Sangal V, Kristo P, et al. Population structure of the Yersinia pseudotuberculosis complex according to multilocus sequence typing. Environ Microbiol. 2011 Dec;13(12):3114–27. [CrossRef]
- Herzog KAT, Schneditz G, Leitner E, Feierl G, Hoffmann KM, Zollner-Schwetz I, et al. Genotypes of Klebsiella oxytoca Isolates from Patients with Nosocomial Pneumonia Are Distinct from Those of Isolates from Patients with Antibiotic-Associated Hemorrhagic Colitis. J Clin Microbiol. 2014 May;52(5):1607–16. [CrossRef]
- Zhou H, Liu W, Qin T, Liu C, Ren H. Defining and Evaluating a Core Genome Multilocus Sequence Typing Scheme for Whole-Genome Sequence-Based Typing of Klebsiella pneumoniae. Front Microbiol [Internet]. 2017 Mar 8 [cited 2024 Nov 24];8. Available from: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00371/full. [CrossRef]
- Nakamura A, Takahashi H, Arai M, Tsuchiya T, Wada S, Fujimoto Y, et al. Molecular subtyping for source tracking of Escherichia coli using core genome multilocus sequence typing at a food manufacturing plant. Eppinger M, editor. PLOS ONE. 2021 Dec 23;16(12):e0261352. [CrossRef]
- Higgins PG, Prior K, Harmsen D, Seifert H. Development and evaluation of a core genome multilocus typing scheme for whole-genome sequence-based typing of Acinetobacter baumannii. Gao F, editor. PLOS ONE. 2017 Jun 8;12(6):e0179228.
- Gobierno de Aragón. IRAS-PROA Aragón [Internet]. [cited 2024 Oct 31]. Available from: https://www.aragon.es/-/iras.
- Endale H, Mathewos M, Abdeta D. Potential Causes of Spread of Antimicrobial Resistance and Preventive Measures in One Health Perspective-A Review. Infect Drug Resist. 2023 Dec 8;16:7515–45. [CrossRef]
- Goryluk-Salmonowicz A, Popowska M. Factors promoting and limiting antimicrobial resistance in the environment – Existing knowledge gaps. Front Microbiol. 2022 Sep 20;13:992268. [CrossRef]
- Symochko L, Pereira P, Demyanyuk O, Pinheiro MNC, Barcelo D. Resistome in a changing environment: Hotspots and vectors of spreading with a focus on the Russian-Ukrainian War. Heliyon. 2024 Jun;10(12):e32716. [CrossRef]
- Logan LK, Weinstein RA. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J Infect Dis. 2017 Feb 15;215(suppl_1):S28–36. [CrossRef]
- Yan JJ. Metallo-beta-lactamase-producing Enterobacteriaceae isolates in a university hospital in Taiwan: prevalence of IMP-8 in Enterobacter cloacae and first identification of VIM-2 in Citrobacter freundii. J Antimicrob Chemother. 2002 Oct 1;50(4):503–11. [CrossRef]
- Da Silva G, Domingues S. Insights on the Horizontal Gene Transfer of Carbapenemase Determinants in the Opportunistic Pathogen Acinetobacter baumannii. Microorganisms. 2016 Aug 23;4(3):29. [CrossRef]
- Gottig S, Gruber TM, Stecher B, Wichelhaus TA, Kempf VAJ. In Vivo Horizontal Gene Transfer of the Carbapenemase OXA-48 During a Nosocomial Outbreak. Clin Infect Dis. 2015 Jun 15;60(12):1808–15. [CrossRef]
- Michaelis C, Grohmann E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms. Antibiotics. 2023 Feb 4;12(2):328. [CrossRef]
- Willems RPJ, Van Dijk K, Vehreschild MJGT, Biehl LM, Ket JCF, Remmelzwaal S, et al. Incidence of infection with multidrug-resistant Gram-negative bacteria and vancomycin-resistant enterococci in carriers: a systematic review and meta-regression analysis. Lancet Infect Dis. 2023 Jun;23(6):719–31. [CrossRef]
- Budhram DR, Mac S, Bielecki JM, Patel SN, Sander B. Health outcomes attributable to carbapenemase-producing Enterobacteriaceae infections: A systematic review and meta-analysis. Infect Control Hosp Epidemiol. 2020 Jan;41(1):37–43. [CrossRef]
- Suay-García B, Pérez-Gracia MT. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics. 2019 Aug 19;8(3):122. [CrossRef]
- Sabtcheva S, Stoikov I, Ivanov IN, Donchev D, Lesseva M, Georgieva S, et al. Genomic Characterization of Carbapenemase-Producing Enterobacter hormaechei, Serratia marcescens, Citrobacter freundii, Providencia stuartii, and Morganella morganii Clinical Isolates from Bulgaria. Antibiotics. 2024 May 16;13(5):455.
- Dilagui I, Loqman S, Lamrani Hanchi A, Soraa N. Antibiotic resistance patterns of carbapenemase-producing Enterobacterales in Mohammed VI University Hospital of Marrakech, Morocco. Infect Dis Now. 2022 Sep;52(6):334–40. [CrossRef]
- Eshetie S, Unakal C, Gelaw A, Ayelign B, Endris M, Moges F. Multidrug resistant and carbapenemase producing Enterobacteriaceae among patients with urinary tract infection at referral Hospital, Northwest Ethiopia. Antimicrob Resist Infect Control. 2015 Dec;4(1):12. [CrossRef]
- Boueroy P, Chopjitt P, Hatrongjit R, Morita M, Sugawara Y, Akeda Y, et al. Fluoroquinolone resistance determinants in carbapenem-resistant Escherichia coli isolated from urine clinical samples in Thailand. PeerJ. 2023 Nov 8;11:e16401. [CrossRef]
- Almaghrabi R, Clancy CJ, Doi Y, Hao B, Chen L, Shields RK, et al. Carbapenem-Resistant Klebsiella pneumoniae Strains Exhibit Diversity in Aminoglycoside-Modifying Enzymes, Which Exert Differing Effects on Plazomicin and Other Agents. Antimicrob Agents Chemother. 2014 Aug;58(8):4443–51. [CrossRef]
- Ching C, Orubu ESF, Sutradhar I, Wirtz VJ, Boucher HW, Zaman MH. Bacterial antibiotic resistance development and mutagenesis following exposure to subinhibitory concentrations of fluoroquinolones in vitro: a systematic review of the literature. JAC-Antimicrob Resist. 2020 Sep 30;2(3):dlaa068. [CrossRef]
- Oteo J, Ortega A, Bartolomé R, Bou G, Conejo C, Fernández-Martínez M, et al. Prospective Multicenter Study of Carbapenemase-Producing Enterobacteriaceae from 83 Hospitals in Spain Reveals High In Vitro Susceptibility to Colistin and Meropenem. Antimicrob Agents Chemother. 2015 Jun;59(6):3406–12. [CrossRef]
- Karlowsky JA, Lob SH, Siddiqui F, Akrich B, DeRyke CA, Young K, et al. In vitro activity of ceftolozane/tazobactam against multidrug-resistant Pseudomonas aeruginosa from patients in Western Europe: SMART 2017-2020. Int J Antimicrob Agents. 2023 May;61(5):106772. [CrossRef]
- Torres-Castillo LC, Fandiño C, Ramos MP, Ramos-Castaneda JA, Rioseco ML, Juliet C. In vitro activity of ceftazidime-avibactam against Gram-negative strains in Chile 2015–2021. J Glob Antimicrob Resist. 2023 Dec;35:143–8. [CrossRef]
- Lasarte-Monterrubio C, Guijarro-Sánchez P, Vázquez-Ucha JC, Alonso-Garcia I, Alvarez-Fraga L, Outeda M, et al. Antimicrobial Activity of Cefiderocol against the Carbapenemase-Producing Enterobacter cloacae Complex and Characterization of Reduced Susceptibility Associated with Metallo-β-Lactamase VIM-1. Antimicrob Agents Chemother. 2023 Apr 13;67(5):e01505-22. [CrossRef]
- Spapen H, Jacobs R, Van Gorp V, Troubleyn J, Honoré PM. Renal and neurological side effects of colistin in critically ill patients. Ann Intensive Care. 2011 May 25;1:14. [CrossRef]
- Cañada-García JE, Moure Z, Sola-Campoy PJ, Delgado-Valverde M, Cano ME, Gijón D, et al. CARB-ES-19 Multicenter Study of Carbapenemase-Producing Klebsiella pneumoniae and Escherichia coli From All Spanish Provinces Reveals Interregional Spread of High-Risk Clones Such as ST307/OXA-48 and ST512/KPC-3. Front Microbiol. 2022 Jun 30;13:918362.
- Zou H, Shen Y, Li C, Li Q. Two Phenotypes of Klebsiella pneumoniae ST147 Outbreak from Neonatal Sepsis with a Slight Increase in Virulence. Infect Drug Resist. 2022 Jan;Volume 15:1–12. [CrossRef]
- Peirano G, Chen L, Kreiswirth BN, Pitout JDD. Emerging Antimicrobial-Resistant High-Risk Klebsiella pneumoniae Clones ST307 and ST147. Antimicrob Agents Chemother. 2020 Sep 21;64(10):e01148-20.
- Lowe M, Kock MM, Coetzee J, Hoosien E, Peirano G, Strydom KA, et al. Klebsiella pneumoniae ST307 with blaOXA-181, South Africa, 2014–2016. Emerg Infect Dis. 2019 Apr;25(4):739–47.
- Gálvez-Silva M, Arros P, Berríos-Pastén C, Villamil A, Rodas PI, Araya I, et al. Carbapenem-resistant hypervirulent ST23 Klebsiella pneumoniae with a highly transmissible dual-carbapenemase plasmid in Chile. Biol Res. 2024 Mar 12;57(1):7. [CrossRef]
- Biedrzycka M, Izdebski R, Urbanowicz P, Polańska M, Hryniewicz W, Gniadkowski M, et al. MDR carbapenemase-producing Klebsiella pneumoniae of the hypervirulence-associated ST23 clone in Poland, 2009–19. J Antimicrob Chemother. 2022 Nov 28;77(12):3367–75. [CrossRef]
- Heljanko V, Tyni O, Johansson V, Virtanen JP, Räisänen K, Lehto KM, et al. Clinically relevant sequence types of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae detected in Finnish wastewater in 2021–2022. Antimicrob Resist Infect Control. 2024 Jan 30;13(1):14. [CrossRef]
- Boutzoukas AE, Komarow L, Chen L, Hanson B, Kanj SS, Liu Z, et al. International Epidemiology of Carbapenemase-Producing Escherichia coli. Clin Infect Dis. 2023 Aug 22;77(4):499–509. [CrossRef]
- Yaici L, Haenni M, Saras E, Boudehouche W, Touati A, Madec JY. blaNDM-5 -carrying IncX3 plasmid in Escherichia coli ST1284 isolated from raw milk collected in a dairy farm in Algeria. J Antimicrob Chemother. 2016 Sep;71(9):2671–2.
- Shrestha B, Tada T, Shimada K, Shrestha S, Ohara H, Pokhrel BM, et al. Emergence of Various NDM-Type-Metallo-β-Lactamase-Producing Escherichia coli Clinical Isolates in Nepal. Antimicrob Agents Chemother. 2017 Dec;61(12):e01425-17. [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters [Internet]. 2023. Available from: https://www.eucast.org/eucast_news/news_singleview?tx_ttnews%5Btt_news%5D=518&cHash=2509b0db92646dffba041406dcc9f20c.
- EUCAST_detection_of_resistance_mechanisms_170711.pdf [Internet]. [cited 2024 Nov 27]. Available from: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf.
- Wick RR, Judd LM, Gorrie CL, Holt KE. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. Phillippy AM, editor. PLOS Comput Biol. 2017 Jun 8;13(6):e1005595. [CrossRef]
- Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: quality assessment tool for genome assemblies. Bioinformatics. 2013 Apr 15;29(8):1072–5.
- Seppey M, Manni M, Zdobnov EM. BUSCO: Assessing Genome Assembly and Annotation Completeness. In: Kollmar M, editor. Gene Prediction [Internet]. New York, NY: Springer New York; 2019 [cited 2024 Nov 24]. p. 227–45. (Methods in Molecular Biology; vol. 1962). Available from: http://link.springer.com/10.1007/978-1-4939-9173-0_14. [CrossRef]
- Ondov BD, Starrett GJ, Sappington A, Kostic A, Koren S, Buck CB, et al. Mash Screen: high-throughput sequence containment estimation for genome discovery. Genome Biol. 2019 Dec;20(1):232. [CrossRef]
- Orakov A, Fullam A, Coelho LP, Khedkar S, Szklarczyk D, Mende DR, et al. GUNC: detection of chimerism and contamination in prokaryotic genomes. Genome Biol. 2021 Dec;22(1):178.
- Wick RR, Schultz MB, Zobel J, Holt KE. Bandage: interactive visualization of de novo genome assemblies. Bioinformatics. 2015 Oct 15;31(20):3350–2. [CrossRef]
- Lumpe J, Gumbleton L, Gorzalski A, Libuit K, Varghese V, Lloyd T, et al. GAMBIT (Genomic Approximation Method for Bacterial Identification and Tracking): A methodology to rapidly leverage whole genome sequencing of bacterial isolates for clinical identification. Chen CC, editor. PLOS ONE. 2023 Feb 16;18(2):e0277575. [CrossRef]
- Seemann T. mlst [Internet]. GitHub. 2022 [cited 2024 Oct 31]. Available from: https://github.com/tseemann/mlst.
- Lam MMC, Wick RR, Watts SC, Cerdeira LT, Wyres KL, Holt KE. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat Commun. 2021 Jul 7;12(1):4188. [CrossRef]
- Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014 Jul 15;30(14):2068–9.
- Casimiro-Soriguer CS, Muñoz-Mérida A, Pérez-Pulido AJ. Sma3s: A universal tool for easy functional annotation of proteomes and transcriptomes. PROTEOMICS. 2017 Jun;17(12):1700071. [CrossRef]
- Alcock BP, Huynh W, Chalil R, Smith KW, Raphenya AR, Wlodarski MA, et al. CARD 2023: expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023 Jan 6;51(D1):D690–9. [CrossRef]
- Seemann T. ABRicate [Internet]. GitHub. 2020 [cited 2024 Oct 31]. Available from: https://github.com/tseemann/abricate.
- Zankari E, Hasman H, Cosentino S, Vestergaard M, Rasmussen S, Lund O, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012 Nov 1;67(11):2640–4.
- Seemann T. Snippy [Internet]. GitHub. 2020 [cited 2024 Oct 31]. Available from: https://github.com/tseemann/snippy.
- Croucher NJ, Page AJ, Connor TR, Delaney AJ, Keane JA, Bentley SD, et al. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015 Feb 18;43(3):e15–e15.
- Nguyen LT, Schmidt HA, Von Haeseler A, Minh BQ. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol Biol Evol. 2015 Jan;32(1):268–74.
- Kalyaanamoorthy S, Minh BQ, Wong TKF, Von Haeseler A, Jermiin LS. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods. 2017 Jun;14(6):587–9. [CrossRef]
- Hoang DT, Chernomor O, Von Haeseler A, Minh BQ, Vinh LS. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol Biol Evol. 2018 Feb 1;35(2):518–22.
- Silva M, Machado MP, Silva DN, Rossi M, Moran-Gilad J, Santos S, et al. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb Genomics [Internet]. 2018 Mar 1 [cited 2024 Nov 24];4(3). Available from: https://www.microbiologyresearch.org/content/journal/mgen/10.1099/mgen.0.000166.
- Hyatt D, Chen GL, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010 Dec;11(1):119. [CrossRef]
- Zhou Z, Alikhan NF, Sergeant MJ, Luhmann N, Vaz C, Francisco AP, et al. GrapeTree: visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018 Sep;28(9):1395–404. [CrossRef]
- Carattoli A, Zankari E, García-Fernández A, Voldby Larsen M, Lund O, Villa L, et al. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob Agents Chemother. 2014 Jul;58(7):3895–903.
- Robertson J, Nash JHE. MOB-suite: software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb Genomics [Internet]. 2018 Aug 1 [cited 2024 Nov 24];4(8). Available from: https://www.microbiologyresearch.org/content/journal/mgen/10.1099/mgen.0.000206. [CrossRef]
- Antipov D, Hartwick N, Shen M, Raiko M, Lapidus A, Pevzner PA. plasmidSPAdes: assembling plasmids from whole genome sequencing data. Bioinformatics. 2016 Nov 15;32(22):3380–7. [CrossRef]
- Schwengers O, Jelonek L, Dieckmann MA, Beyvers S, Blom J, Goesmann A. Bakta: rapid and standardized annotation of bacterial genomes via alignment-free sequence identification: Find out more about Bakta, the motivation, challenges and applications, here. Microb Genomics [Internet]. 2021 Nov 30 [cited 2024 Nov 24];7(11). Available from: https://www.microbiologyresearch.org/content/journal/mgen/10.1099/mgen.0.000685. [CrossRef]
- Grant JR, Enns E, Marinier E, Mandal A, Herman EK, Chen C yu, et al. Proksee: in-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023 Jul 5;51(W1):W484–92.
- Jain C, Rodriguez-R LM, Phillippy AM, Konstantinidis KT, Aluru S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat Commun. 2018 Nov 30;9(1):5114. [CrossRef]
Figure 1.
Heatmap of antibiotic susceptibility pattern by isolate. Heatmap showing susceptibility profiles and carbapenemase of each isolate. Green: Susceptible, Red: Non-susceptible, Gray: Not Available, Yellow: Area of Technical Uncertainty. ID: Sample identification number CAZ: Ceftazidime, CFP: Cefepime, ERT: Ertapenem, IMI: Imipenem, MER: Meropenem, ATM: Aztreonam, CTO: Ceftolozane-Tazobactam, CZA: Ceftazidime-Avibactam, CFD: Cefiderocol, CIP: Ciprofloxacin, LEV: Levofloxacin, GEN: Gentamicin, TOB: Tobramycin, AMK: Amikacin, COL: Colistin, FOT: Fosfomycin, TIG: Tigecyclin.
Figure 1.
Heatmap of antibiotic susceptibility pattern by isolate. Heatmap showing susceptibility profiles and carbapenemase of each isolate. Green: Susceptible, Red: Non-susceptible, Gray: Not Available, Yellow: Area of Technical Uncertainty. ID: Sample identification number CAZ: Ceftazidime, CFP: Cefepime, ERT: Ertapenem, IMI: Imipenem, MER: Meropenem, ATM: Aztreonam, CTO: Ceftolozane-Tazobactam, CZA: Ceftazidime-Avibactam, CFD: Cefiderocol, CIP: Ciprofloxacin, LEV: Levofloxacin, GEN: Gentamicin, TOB: Tobramycin, AMK: Amikacin, COL: Colistin, FOT: Fosfomycin, TIG: Tigecyclin.
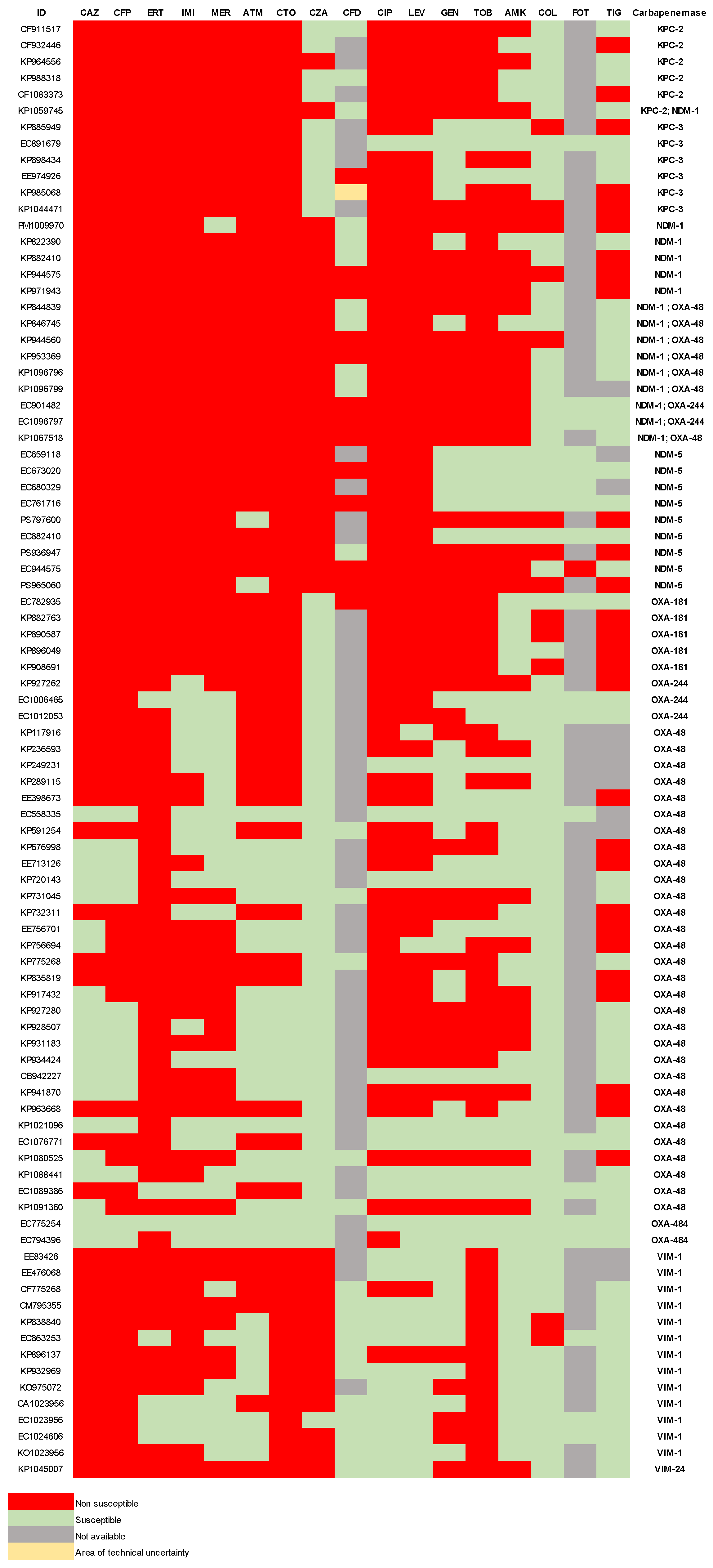
Figure 2.
MST of the K. pneumoniae cgMLST. Minimum spanning tree reconstructed from the allele matrix of the cgMLST of K. pneumoniae species complex. Circles represent isolates, connected by solid lines with lengths proportional to the allelic distances between them. The allelic difference between each pair is indicated as an integer on the corresponding connecting line. Isolates with 15 or fewer allelic differences are grouped within the same circle, depicted as a pie chart, where the circle's size reflects the number of isolates it contains. Dashed lines indicate allelic differences greater than 50. The number in brackets in the legend indicates the absolute frequency of the observation.
Figure 2.
MST of the K. pneumoniae cgMLST. Minimum spanning tree reconstructed from the allele matrix of the cgMLST of K. pneumoniae species complex. Circles represent isolates, connected by solid lines with lengths proportional to the allelic distances between them. The allelic difference between each pair is indicated as an integer on the corresponding connecting line. Isolates with 15 or fewer allelic differences are grouped within the same circle, depicted as a pie chart, where the circle's size reflects the number of isolates it contains. Dashed lines indicate allelic differences greater than 50. The number in brackets in the legend indicates the absolute frequency of the observation.
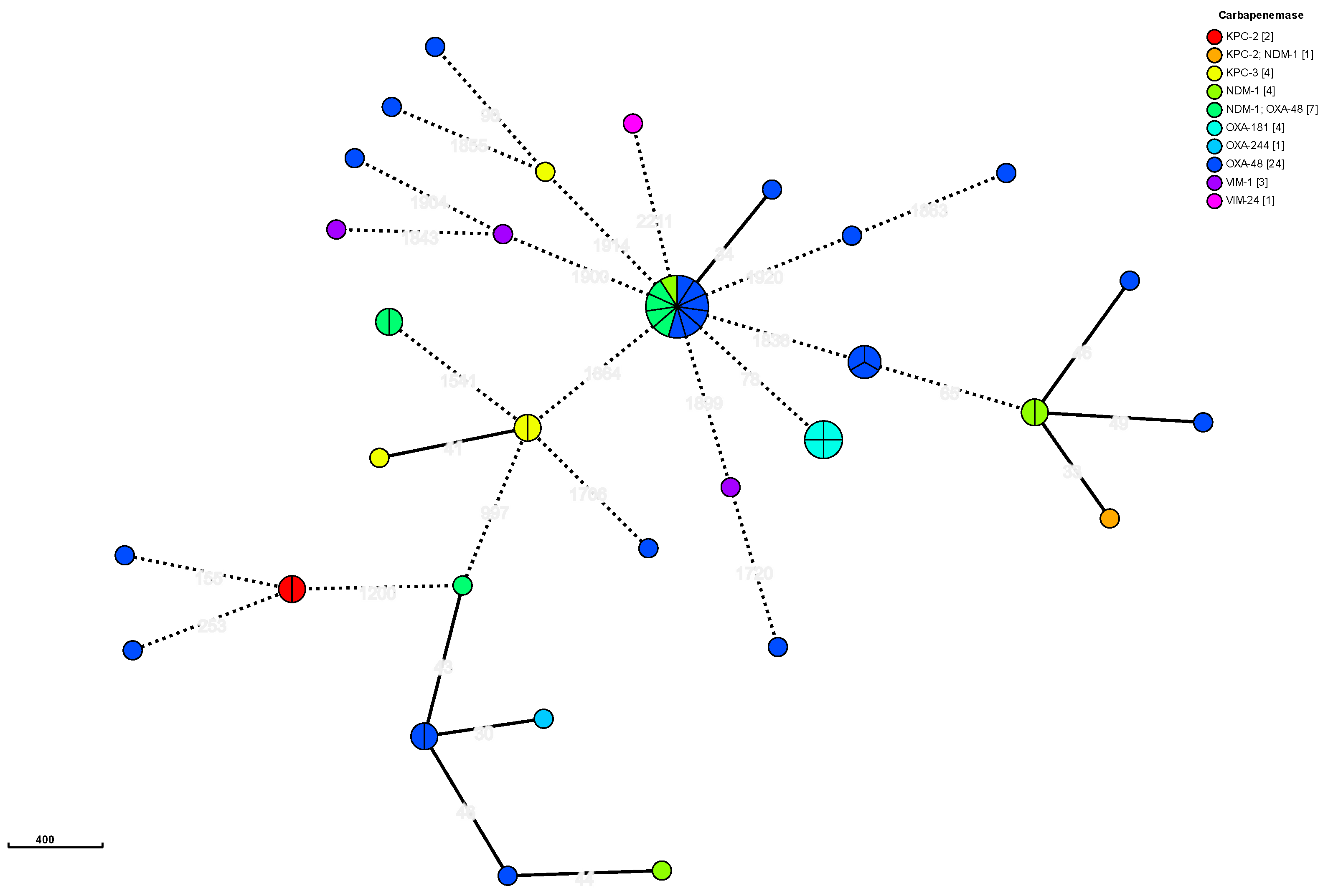
Figure 3.
MST of the E. coli cgMLST. Minimum spanning tree reconstructed from the allele matrix of the cgMLST of E. coli. Follows the same schematics as in Figure 2.
Figure 3.
MST of the E. coli cgMLST. Minimum spanning tree reconstructed from the allele matrix of the cgMLST of E. coli. Follows the same schematics as in Figure 2.
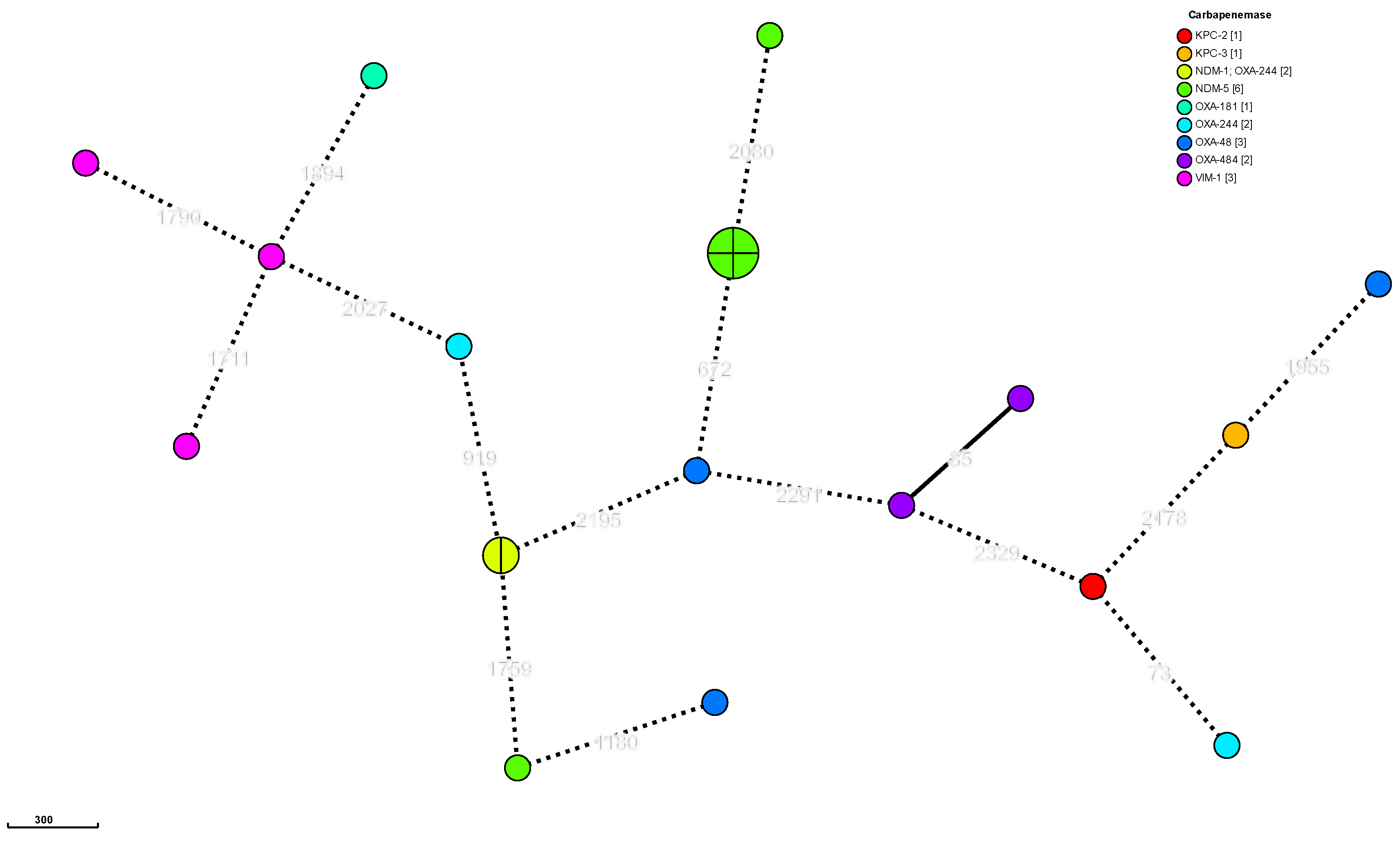
Figure 4.
Phylogenetic analysis of carbapenem-producing K. pneumoniae. Phylogenetic analysis of K. pneumoniae sensu stricto. The cladogram depicts the relationships among isolates. Inner shading indicates the ST of each isolate. Isolates without shading represent unique STs. The outermost circle highlights the carbapenemase associated with each isolate. Bootstrap values exceeding 70 are displayed in white-background boxes on the respective branches.
Figure 4.
Phylogenetic analysis of carbapenem-producing K. pneumoniae. Phylogenetic analysis of K. pneumoniae sensu stricto. The cladogram depicts the relationships among isolates. Inner shading indicates the ST of each isolate. Isolates without shading represent unique STs. The outermost circle highlights the carbapenemase associated with each isolate. Bootstrap values exceeding 70 are displayed in white-background boxes on the respective branches.
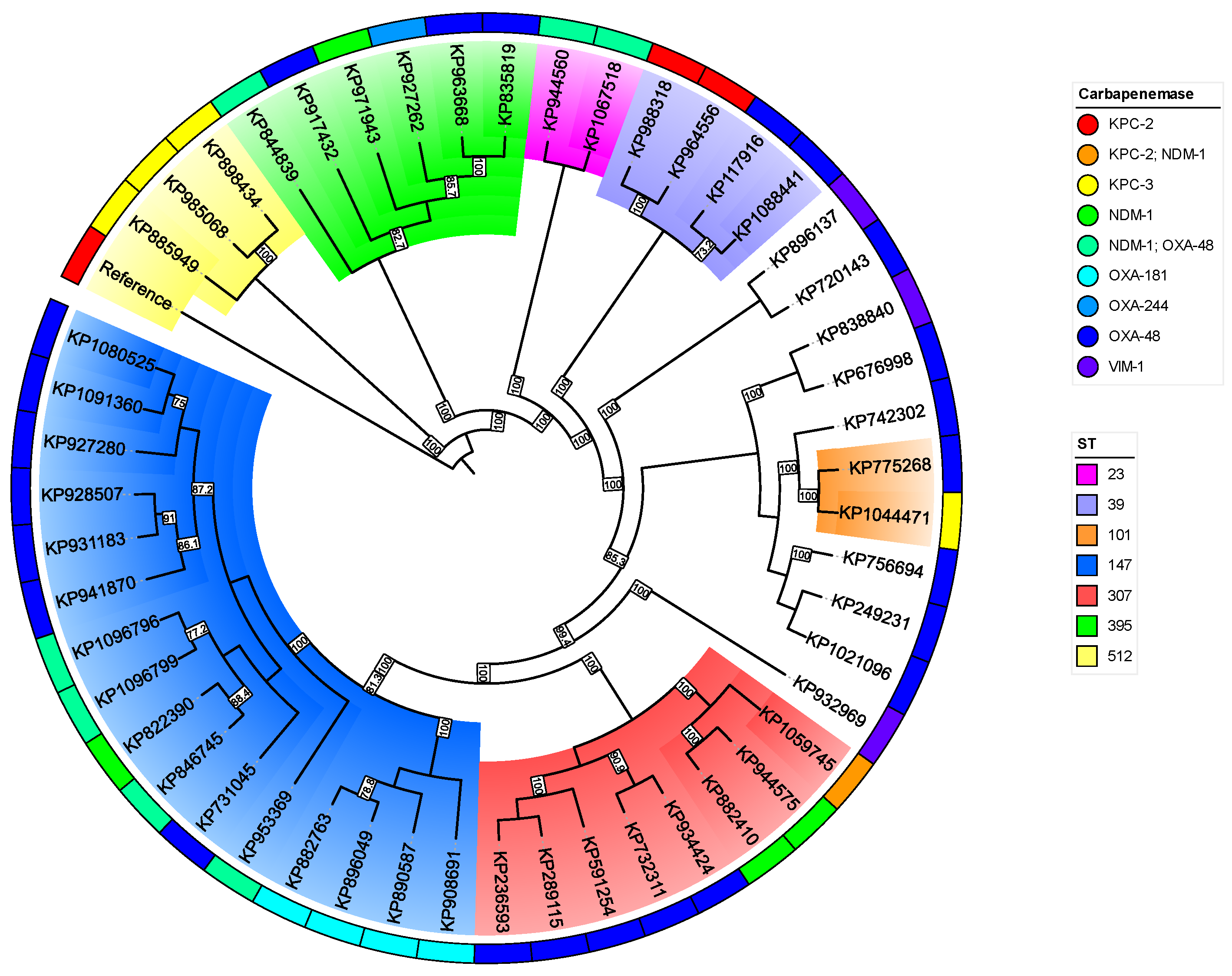
Figure 5.
Phylogenetic analysis of carbapenem-producing E. coli. Phylogenetic analysis of E. coli. The cladogram depicts the relationships among isolates and follows the same schematics as in Figure 4.
Figure 5.
Phylogenetic analysis of carbapenem-producing E. coli. Phylogenetic analysis of E. coli. The cladogram depicts the relationships among isolates and follows the same schematics as in Figure 4.
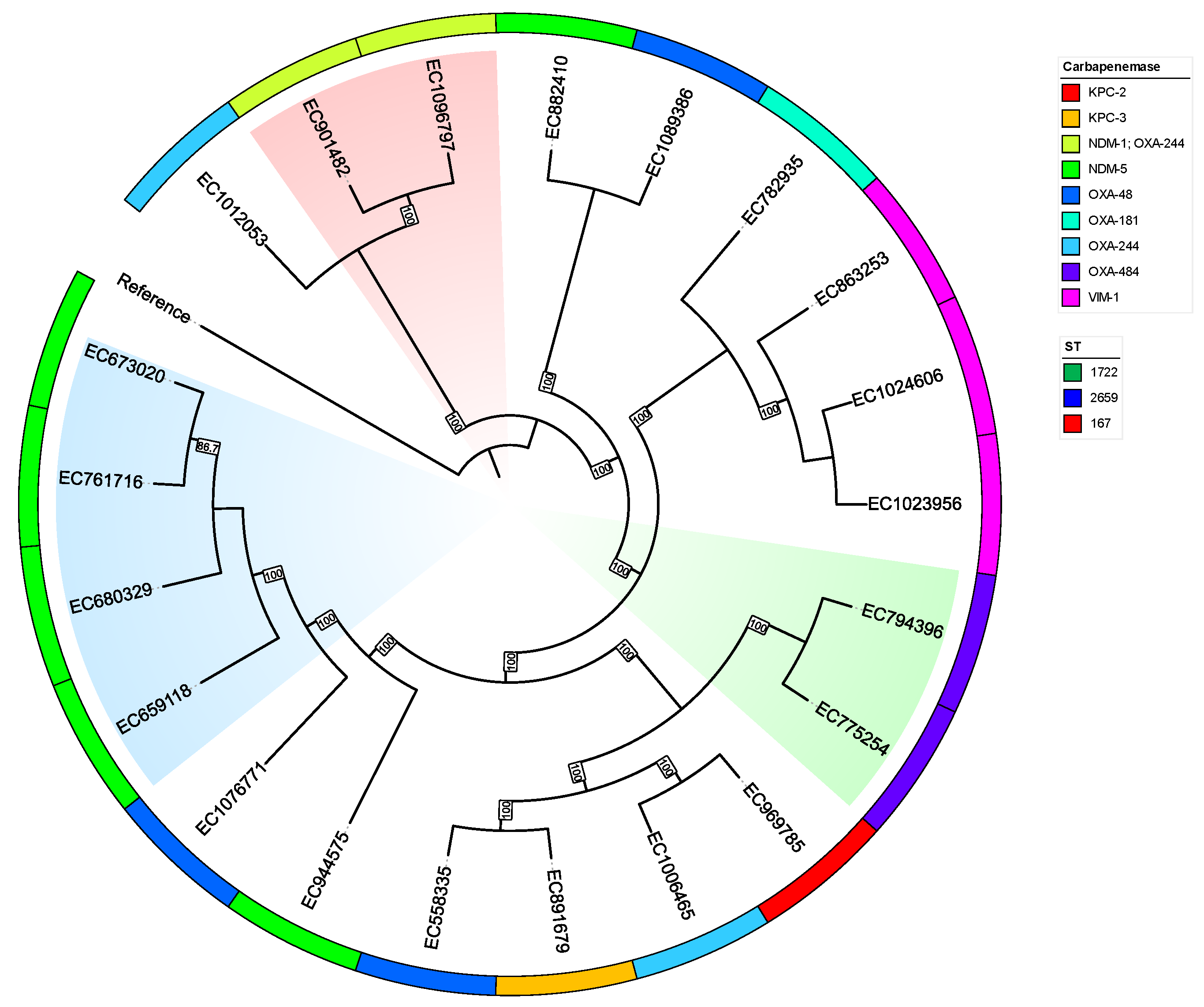

Table 1.
Distribution of Enterobacteriaceae isolation.
| Species | 2021 |
2021 carba |
2022 |
2022 carba |
2023 |
2023 carba |
Total |
| E. coli | 6629 | 0 | 7590 | 7 | 8870 | 14 | 23089 |
| K. pneumoniae | 1646 | 5 | 2102 | 14 | 2784 | 40 | 6532 |
| P. mirabilis | 649 | 0 | 741 | 0 | 891 | 1 | 2281 |
| E. cloacae | 424 | 3 | 542 | 2 | 579 | 3 | 1545 |
| K. oxytoca | 324 | 0 | 389 | 0 | 405 | 2 | 1118 |
| Citrobacter spp | 280 | 5 | 357 | 7 | 455 | 7 | 1092 |
| S. marcescens | 205 | 0 | 217 | 0 | 246 | 1 | 668 |
| M. morganii | 177 | 0 | 261 | 0 | 284 | 0 | 722 |
| K. aerogenes | 142 | 0 | 162 | 0 | 179 | 0 | 483 |
| P. stuartii | 42 | 0 | 60 | 1 | 70 | 2 | 172 |
| Others | 110 | 0 | 126 | 0 | 207 | 0 | 443 |
| Total | 10628 | 13 | 12547 | 31 | 14970 | 70 | 38189 |
Total isolation of Enterobacteriaceae by year and presence of carbapenemases. Carba: Isolations with carbapenemases.
Table 2.
Distribution of carbapenemases by species.
| Carbapenemase/Species | K. pneumoniae complex | E. coli | E. cloacae complex | Citrobacter spp | P. stuartii | K. oxytoca | P. mirabilis | Total |
|---|---|---|---|---|---|---|---|---|
| KPC-2 | 2 | 1 | 0 | 3 | 0 | 0 | 0 | 6 |
| KPC-3 | 4 | 0 | 2 | 0 | 0 | 0 | 0 | 6 |
| NDM-1 | 4 | 0 | 0 | 0 | 0 | 0 | 1 | 5 |
| NDM-5 | 0 | 6 | 0 | 0 | 3 | 0 | 0 | 9 |
| VIM-1 | 3 | 3 | 2 | 3 | 0 | 2 | 0 | 13 |
| VIM-24 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| OXA-48 | 24 | 3 | 3 | 1 | 0 | 0 | 0 | 31 |
| OXA-181 | 4 | 1 | 0 | 0 | 0 | 0 | 0 | 5 |
| OXA-244 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 3 |
| OXA-484 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| NDM-1 + OXA-48 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 7 |
| NDM-1 + OXA-244 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| NDM-1 + KPC-2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Total | 51 | 20 | 7 | 7 | 3 | 2 | 1 | 91 |
Table 3.
Genomic characterization summary of CPE.
| CPE | Carbapenemases | ST | cgMLST cluster | BLEE | AmpC | gyrA mutation | parC mutation | Aminoglycosides modifing genes | Plasmid_replicons (carbapenemase) | Predicted_mobility | Mean plasmid size |
|---|---|---|---|---|---|---|---|---|---|---|---|
| K. pneumoniae | OXA-48 | 13,15,39,101, 147,307,346, 395,405,685,4872 |
Yes | CTX-M-15 | 0 | S83I,S83F,D87A | S80I | Several | IncL/M,IncR | Variable | 46443 |
| OXA-181 | 147 | Yes | CTX-M-15 | 0 | S83I | S80I | Several | rep_cluster_1195 | Mainly non-mobilizable | 40801 | |
| OXA-244 | 395 | No | CTX-M-15 | 0 | S83I | S80I | Several | IncL/M | Conjugative | 87763 | |
| KPC-2 | 39 | Yes | CTX-M-15 | 0 | S83I,D87N | S80I | Several | IncFIB,IncFII,IncX3,ColRNAI_rep_cluster_1857 | Mainly conjugative | 94778 | |
| KPC-3 | 101,512 | Yes | 0 | 0 | S83I,S83Y,D87N | S80I | Several | IncFIB,IncFII,IncHI1B,IncR | Conjugative | 63166 | |
| NDM-1 | 147,307,395 | Yes | CTX-M-15 | 0 | S83I | S80I | Several | IncFIB,IncFII,IncHI1B | Conjugative | 278576 | |
| VIM-1 | 2,685,844,387 | No | 0 | DHA-1 | 0 | 0 | Several | IncL/M | Conjugative | 71362 | |
| KPC-2 + NDM-1 | 307 | No | 0 | 0 | S83I | S80I | aph(3')-VI only | IncFIB,IncHI1B,rep_cluster_1254 | Conjugative | 332589 | |
| NDM-1 + OXA-48 | 23,147,395 | Yes | CTX-M-15 | 0 | S83I | S80I | Several | IncFIB,IncFII,IncHI1B | Variable | 101820 | |
| E. coli | OXA-48 | 38,127,1598 | No | CTX-M-15 | 0 | S83L | 0 | Several | IncFIA,IncFII | non-mobilizable | 21541 |
| OXA-181 | 410 | No | 0 | CMY-4 | S83L,D87N | S80I | Several | IncX3 | non-mobilizable | 27916 | |
| OXA-244 | 44,13730 | No | CTX-M-15 | CMY-132 | S83L,D87N | S80I | aadA5 only | ND | ND | ND | |
| OXA-484 | 1722 | No | 0 | 0 | 0 | 0 | aadA2 only | rep_cluster_1195 | non-mobilizable | 14615 | |
| KPC-2 | 131 | No | 0 | CMY-132 | S83L | 0 | Several | IncFIB,IncFII,rep_cluster_2183 | Conjugative | 84857 | |
| KPC-3 | 135 | No | 0 | 0 | 0 | 0 | APH(3') only | IncX3 | Conjugative | 58796 | |
| NDM-5 | 46,405,2659 | No | CTX-M-55 | CMY-42 | D87N, S83L | S80I | aadA2 only | IncFIA,IncFIC,IncFIB | Mainly conjugative | 105572 | |
| VIM-1 | 29,327,539 | No | 0 | 0 | 0 | 0 | Several | IncL/M,IncL/M, IncI-gamma/K1 | Variable | 49209 | |
| NDM-1 + OXA-244 | 167 | Yes | CTX-M-15 | 0 | D87N, S83L | S80I | AAC(3)-Iid only | IncC,rep_cluster_1254 | Conjugative | 210764 | |
| Citrobacter spp | OXA-48 | 225 | ND | 0 | CYM-101 | 0 | 0 | 0 | IncL/M | Conjugative | 61961 |
| KPC-2 | 21259 | ND | CTX-M-15,CTX-M-9 | CMY-106,CYM-159 | S83I | S80I | Several | IncP,IncU | Monilizable | 29296 | |
| VIM-1 | 488493563 | ND | CTX-M-9 | CMY-2,CMY-48 | 0 | 0 | Several | IncL/M,IncY | Mainly conjugative | 57471 | |
| Enterobacer spp | OXA-48 | 13,24 | ND | CTX-M-15 | ACT-1,MIR-5 | S83I | 0 | 0 | IncL/M | Conjugative | 64843 |
| KPC-3 | 51 | ND | 0 | ACT-40 | 0 | 0 | Several | IncN | Conjugative | 50662 | |
| VIM-1 | 108198 | ND | CTX-M-9 | ACT-55 | 0 | 0 | Several | IncL/M,IncHI2A | Conjugative | 152182 | |
| Klebsiella spp | VIM-1 | 59 | ND | CTX-M-9 | 0 | S463A | 0 | Several | IncL/M,IncR | Conjugative | 174918 |
| VIM-24 | 4365 | ND | CTX-M-9 | 0 | S463A | 0 | Several | IncHI2A,rep_cluster_1088 | Conjugative | 289821 | |
| Providencia stuartii | NDM-5 | 11,23 | ND | 0 | CMY-16 | D87G | 0 | Several | IncC,rep_cluster_1254 | Variable | 78040 |
| Proteus mirabilis | NDM-1 | 446 | ND | VEB-6 | 0 | S463A | 0 | Several | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



