Submitted:
05 January 2026
Posted:
07 January 2026
You are already at the latest version
Abstract
The remarkable plasticity of plants is best exemplified by the capacity of their somatic cells to regenerate entire organs or the organism itself. The molecular and cellular events underlying this ability are complex and multifaceted. The initial phase, that is triggered either by wounding or exogenous hormone application and leads to cell cycle reactivation, is often called dedifferentiation. This review proposes that the dedifferentiation of mature somatic cells is a two-step process. It involves a transition into a transient senescence-like state. This leads to a critical cellular condition; in the absence of proliferative signals, cells become committed to programmed cell death. If, however, it is succeeded by a second hormonal rescue step, when phytohormones, mainly auxin and cytokinin, override the death pathway and activate cell division, callus arises. Callus is not an undifferentiated homogenous mass of cells, but an unorganised tissue with at least some cells having ground-tissue-like molecular identity with high developmental potential. Callus, though, might also form from pre-existing, competent populations of cells, e.g. pericycle cells, with no senescence-like intermitting state. It is also discussed whether this “one-step” callus formation pathway can be referred to as dedifferentiation.
Keywords:
plant cell and tissue culture
; protoplast
; callus
; plant regeneration
; cell division
; plant hormones
; senescence
; programmed cell death
; signaling
1. Introduction
The sessile nature of plants necessitates a remarkable capacity for adaptation and regeneration in response to environmental harms and mechanical damage [1,2]. The plant’s regenerative ability is based, beside the meristems, on their cellular plasticity, the capacity of differentiated somatic cells to alter their fate, and in some cases, display the potential to regenerate an organ or even the entire organism [3,4,5]. The initial process central to this phenomenon is cellular dedifferentiation, a term historically used to describe the reversion of a mature cell to a less specialized state within its own developmental lineage which allows its proliferation [6]. In plant biology, however, there is still uncertainty regarding the usage of this term. It is often broadly applied to any form of cellular reprogramming that leads to an increase in developmental potential, whether or not it is a reversal of development [7,8]. An example is the designation of callus as a mass of “dedifferentiated” cells. This might be correct if we consider dedifferentiation as the process leading to callus formation, but wrong to describe the developmental/genetic state of the callus cells as being “dedifferentiated” [7].
In this review, I will try to clarify how dedifferentiation might take place and how it relates to callus formation. Two primary experimental systems are instrumental in dissecting the mechanisms of plant cell dedifferentiation: the isolation and culture of protoplasts (a population of individual plant cells devoid of their walls) and the induction of callus (an unorganized ground tissue) by wounding or ectopic hormone levels [9,10,11]. This overview attempts to reconcile observations from these systems. I propose that the dedifferentiation of differentiated somatic cells, such as isolated mesophyll cells or cells surrounding a wound site, initiates with a senescence-like state that is followed by hormonal rescue leading to proliferation and/or redifferentiation. There is, however, a more direct callus formation pathway due to the overproliferation of pre-existing stem or stem cell-like cells that does not involve the senescence-like intermediate process. The applicability of the term dedifferentiation to describe this “one-step” callus formation pathway will be discussed.
2. Death Signals in the Reprogramming of Differentiated Cells at Wound Sites
The ability to regenerate lost or damaged tissues is a fundamental characteristic of multicellular life. It is essential for surviving injuries and maintaining organismal integrity. This capability is widespread across the tree of life, yet it differs dramatically among various taxa or even between different tissues of the same organism [1]. In the animal kingdom, organisms like planarians and hydra exhibit extraordinary regenerative potential. They are capable of rebuilding their entire body from small parts, whereas mammals, including humans, possess a more limited regeneration ability, largely restricted to specific tissues such as the liver and skin. Two fundamental, yet seemingly contradictory, processes profoundly contribute to these regeneration strategies: programmed cell death (PCD) and cellular proliferation [12].
Historically, cell death was viewed as a passive and terminal process, the inevitable conclusion of a cell’s life. However, over the past few decades it has been revealed that cell death is often an active, genetically controlled program that plays a crucial role in development and the maintenance of tissue homeostasis [13]. More surprisingly, studies in diverse organisms, from drosophila and hydra to zebrafish and mice, have shown that apoptosis, a form of programmed cell death (PCD) of animals, is involved in an essential and conserved mechanism for successful tissue repair and regeneration following injury [14,15]. The phenomenon is often termed as “apoptosis-induced proliferation” (AiP) designating a process where dying cells release mitogenic signals that stimulate the proliferation of neighbouring stem or progenitor cells to replace lost tissue [12,13]. In mice, for instance, apoptotic cells have been shown to activate a “phoenix rising” pathway, where the executioner caspases 3 and 7, key enzymes in the apoptotic cascade, trigger the release of growth signals like prostaglandin E2 (PGE2) to promote wound healing and liver regeneration [16]. This active communication from dying cells orchestrates the renewal of the collective tissue [12].
In contrast to most animals, plants possess a remarkable capacity for regeneration [17,18]. Many plant species can regenerate entire organs, such as leaves, stems, or roots, and in some cases, a whole plant can be reconstituted from a small piece of tissue or even a single cell [19]. Like animals, plants utilize PCD as a fundamental tool for development and for responding to environmental challenges [20]. PCD is essential for sculpting plant architecture, such as during xylem formation, and for defence strategies like the hypersensitive response, where localized cell death contains the spread of pathogens. Given the well-established role of PCD in initiating regeneration in animals, it is logical to hypothesize that a similar principle operates in plants. PCD is deeply integrated into plant defence, but its role as a pro-regenerative signal could create a complex regulatory challenge. The wound response in plants is inherently a balancing act between two potentially conflicting priorities: defence and regeneration [21]. Activating robust defence pathways is energetically costly and often involves growth inhibition, which is antithetical to the cell proliferation required for regeneration. Conversely, prioritizing regeneration might leave the plant vulnerable to infection at the wound site. The wound response is, therefore, can be considered as a multi-layered process that must simultaneously seal the injury to prevent water loss and pathogen entry and initiate repair and regeneration programs [21,22]. The cells that die at a wound site are not merely casualties of injury; they are the source of instructive signals to activate defence and regeneration. For example, when a cell in the root meristem dies, it loses turgor and collapses, creating a mechanical signal leading to elevated auxin accumulation and sensitivity in neighbouring cells, provoking their division [23]. The recognition of damage can also be mediated by the release of molecules from ruptured cells known as Damage-Associated Molecular Patterns (DAMPs) [24]. Wound cues also trigger waves of calcium ions (Ca2+), reactive oxygen species (ROS), and electrical signals that propagate from the wound site throughout the plant, alerting even distant tissues to the injury [22]. These early signals are crucial for activating context-dependent local and systemic downstream responses. Local responses often include the PCD of cells at the borderline of the wound [25,26,27]. It is plausible that the genetically controlled dismantling of these cells via PCD could release specific pro-regenerative signals, analogous to the “phoenix rising” pathway in mammals [16]. Indeed, recent work suggests that similar pathways exist in plants. It has been previously shown that when tomato plants are damaged, wounding triggers the activation of Ca2+-dependent metacaspases also involved in PCD [28]. This leads to the release of peptides that were proposed to bind and activate their extracellular receptors, initiating an immune-like response [29]. These peptides have recently been found to profoundly enhance plant regeneration via receptor-mediated induction of WOUND-INDUCED DEDIFFERENTIATION 1 (WIND1) [30,31], a master regulator of wound-induced cellular reprogramming in plants [32].
The complex network of phytohormones that orchestrate the subsequent cellular and physiological changes is central to the plant wound response. Jasmonic acid (JA), the master regulator of plant defence against herbivores and necrotrophic pathogens is rapidly synthesized upon wounding [22,33]. JA signalling activates massive transcriptional reprogramming that leads to the production of defence compounds and proteins. Besides defence reactions, JA, in concert with ethylene, is a central regulator of PCD and senescence [34,35] but it also triggers the regeneration pathway [36,37,38,39,40] but inhibits subsequent callus formation [41]. In parallel, the phytohormone auxin plays a pivotal role in guiding the regenerative process [5,36,42]. Auxin is a master coordinator of plant development, and its differential accumulation into local maxima is a common mechanism for initiating the formation of new organs. Following wounding, auxin transport is rerouted and its local synthesis is initiated, leading to its accumulation at the wound site, where it triggers cell division, cellular reprogramming, and the formation of new vascular connections or entire organs [5,42,43]. An increasing body of evidence also supports a profound and intricate connection between PCD and auxin signalling pathways [44]. This interplay is particularly evident in the context of wound healing and regeneration, where the death of some cells serves as a crucial signal to trigger the restorative division of others. At the heart of this regulatory network lies ETHYLENE RESPONSE FACTOR 115 (ERF115), an AP2/ERF transcription factor that has emerged as a pivotal integrator of cell death signals and hormonal cues to orchestrate regeneration among others activating WIND1 expression [26,40].
The function of ERF115 is rooted in its role as a guardian of the stem cell niche [45]. In the Arabidopsis root, ERF115 acts as a rate-limiting factor for the division of quiescent centre (QC) cells, which are responsible for maintaining the surrounding stem cells. Under normal conditions, ERF115 activity is tightly constrained by proteolysis via the APC/CCCS52A2 ubiquitin ligase, preventing uncontrolled proliferation and ensuring the longevity of the stem cell niche [46]. This homeostatic balance is disrupted upon injury. Cell death in or near the stem cell niche triggers a rapid and localized induction of ERF115 expression in the surviving adjacent cells, which then initiate divisions to replace the lost tissue [3,26]. This principle is observed across various contexts of cellular damage. For instance, wounding of the hypocotyl induces ERF115 expression within an hour [41]. DNA damage induced by the loss of TOPOISOMERASE1α function triggers PCD in stele stem cells; this cell death is partially compensated for by ERF115-mediated replenishment from neighbouring cells [47]. Similarly, mutations in genes essential for meristem maintenance, such as MEDIATOR18 and NRPB2, lead to spontaneous cell death in the root tip, which is invariably correlated with high expression of ERF115 in the surrounding tissue [48]. The close temporal and spatial association between cell death events and ERF115 induction suggests an unknown form of communication from the dying cell to its neighbouring cells.
The signals emanating from dying cells to activate ERF115 are multifaceted, involving both physical and chemical cues. The physical collapse of damaged cells creates changes in cellular pressure that are perceived by neighbours, spatially defining the zone of the regenerative response and ERF115 activation [23]. In the unwounded root meristem, ERF115 expression is limited to the quiescent centre (QC). However, as reactive oxygen species (ROS) levels increase ERF115 expression extends throughout the entire root meristem [49]. The wound hormone jasmonate (JA) was also shown as an inducer of ERF115 transcription, directly linking the plant’s defence and repair pathways [40]. Interestingly, the role of ERF115 is context-dependent. While it typically promotes regeneration, in response to aluminium toxicity, ERF115 participates in a pathway that promotes terminal differentiation and growth arrest following Al-induced cell death [50].
While auxin is not the primary inducer of ERF115 transcription, it is required to maintain high levels of ERF115 expression during the regeneration process [51]. In turn, ERF115 enhances the sensitivity of cells to auxin, partly by activating the expression of the key auxin response factor ARF5/MONOPTEROS. This mutual amplification ensures a robust and sustained response, granting regenerative competence to otherwise quiescent cells [51]. Auxin’s relationship with PCD is complex; it generally acts as a survival signal that suppresses cell death, but can also promote it under certain conditions [44,52,53]. In the context of regeneration, the most critical aspect of this relationship is how PCD alters local auxin dynamics. Dying cells can become a significant source of auxin. For example, the cyclic PCD of lateral root cap cells releases pulses of auxin that pattern the formation of new lateral roots [54]. Furthermore, the autolysis of cells releases tryptophan from protein breakdown, providing a substrate for local auxin biosynthesis, suggesting that many sites of cell death are also sites of auxin production [55]. Additionally, physical damage and the resulting cell death create an anatomical barrier that obstructs polar auxin transport, causing auxin to accumulate in the cells adjacent to the wound—much like rocks in a stream [51]. This localized auxin peak is essential for initiating the regenerative program [23,51,56] and engages in a powerful synergistic feedback loop with ERF115. To execute its function, ERF115 forms a heterodimeric complex with GRAS-domain transcription factors such as PHYTOCHROME A SIGNAL TRANSDUCTION 1 (PAT1) [26,41,57]. This complex then activates a downstream transcriptional cascade, including the DNA-BINDING ONE FINGER 3.4 (DOF3.4) transcription factor, which in turn promotes the expression of D3-type cyclins to drive regenerative cell divisions [58].
In summary, while a wound causes necrosis in damaged cells, it also initiates defence and cell death programs in nearby cell layers. These cells serve as an important source of local and systemic regenerative signals, which might save even themselves from dying by rerouting them for wound closure and/or regeneration. Similar phenomena can be observed in cultured leaf protoplasts freed from systemic signals.
3. The Senescence-like State of Protoplast-Derived Cells
The isolation of protoplasts from differentiated tissues like leaves provides a powerful system to study the initial events of cellular reprogramming in a synchronized population of single cells [9,59]. The process itself, involving enzymatic digestion of the cell wall and release into an artificial osmotic, is a profound stressor that in addition removes the cells from their native tissue context and hormonal cues [60,61,62]. The immediate response of these cells bears a striking resemblance to cellular senescence, a programmed developmental process preceding PCD [63,64,65,66]. Cells in both states exhibit similar structural changes, including widespread chromatin decondensation, disruption or shrinkage of the nucleolus, and condensation of ribosomal RNA gene clusters, indicative of reduced protein synthesis and a shift towards a quiescent state [59,63,65]. Furthermore, both processes are marked by a large-scale upregulation of genes from specific transcription factor (TF) families, including ANAC, WRKY, and bZIP, which are well-known master regulators of stress and senescence responses [59,65]. In agreement, transcriptome profiling of dedifferentiating protoplasts reveals a significant overlap with the gene expression patterns of senescing cells (Figure 1). More than two-thirds of the almost four thousand genes listed in the Arabidopsis senescence database [67] are expressed in freshly isolated leaf protoplasts [68]. These protoplasts strongly and transiently express the ERF115, PAT1, and ARF5 transcription factors (Table 1, [68]), underlying their similarity to cells acquiring cell death or wound-induced regeneration competence [26,51,57,69,70].
The senescence-like transient state was hypothesized to confer pluripotentiality to protoplast-derived cells [71,72]. However, at this stage the cells can only follow two pathways. If left in a hormone-free medium, they undergo cycles of chromatin condensation and decondensation before ultimately succumbing to programmed cell death [73]. Protoplast-derived cells, which are unable to divide and regenerate even in the presence of plant hormones are also subjected to programmed cell death [64]. These observations strongly support that the stress-induced, senescence-like state is a direct path to cell death unless the cell is “rescued” by specific proliferative signals inducing endogenous auxin synthesis [74]. Pluripotentiality of the proliferating cells is established only during proper culture conditions including the appropriate ratio of plant hormones [59,68,74]. Nevertheless, it has been demonstrated that the expression of genes associated with pluripotency, such as WOX5, PLT7, and BBM, appear at a very low frequency already in freshly isolated protoplasts. This random activation was considered as the consequence of stochastic changes in chromatin accessibility caused by isolation stress [68]. The expressions of these and other pluripotency markers were shown to get stabilized, however, only several days later in proliferating cells during microcallus formation. In addition, Arabidopsis mesophyll cell protoplasts’ regeneration was promoted by the ectopic expression of the WUSCHEL (WUS) and DORNRÖSCHEN (DRN; or ESR1) genes [68].
Seminal work using the tobacco protoplast system identified two functionally distinct phases of chromatin decondensation during dedifferentiation and cell cycle reactivation, respectively [22]. The first phase occurs rapidly during protoplast isolation, is independent of exogenous hormones, and corresponds to the loss of differentiated functions such as photosynthesis in the case of mesophyll protoplasts. This phase aligns perfectly with a stress-induced global opening of the chromatin that makes the genome accessible for the new transcriptional program of senescence [71]. This initial reprogramming involves marked changes in the expression of chromatin-associated genes, particularly those encoding histone variants, and are associated with epigenetic modifications [59,71,72,75,76]. Once a cell has entered the plastic, senescence-like state, its fate is largely dictated by the hormonal milieu. In this context the application of a protoplast culture medium typically rich in auxin and supplemented with cytokinin serves as a critical rescue signal [73,77]. However, it has recently been shown that exogenous auxin is insufficient, and the synthesis of endogenous auxin is required to accomplish the initial cell division of protoplast-derived cells [74]. The second phase of chromatin decondensation occurs only after protoplasts are supplied with phytohormones (in most cases, with auxin and cytokinin) and precedes endogenous auxin synthesis and the cell’s entry into S-phase [71,74,75,78]. The competent cells complete the mitotic cycle, thereby averting cell death but initiating the formation of a callus [59,68,71,74].
Callus formation, however, can be dissected from the senescence-like dedifferentiation process. The maintenance of the differentiated state in mature plant cells is an active process, partly enforced by epigenetic silencing mechanisms. For example, the POLYCOMB REPRESSIVE COMPLEX 2 (PRC2), which deposits repressive histone marks, plays a pivotal role in preventing the unscheduled dedifferentiation of mature somatic cells like root hairs. Loss of PRC2 function can lead to spontaneous reprogramming of single differentiated cells. However, since these cells remain under hormonal influence, they can directly regain cell division activity, leading to callus formation or even somatic embryogenesis (Ikeuchi et al. 2015). In contrast, reactivation of cell division in mesophyll cells requires histone deacetylation and the inhibition of this process prevents callus formation [74,79].
Therefore, the dedifferentiation of mesophyll protoplast-derived cells is a two-phase process: it starts with a senescence-like event that leads to the loss of differentiated functions, followed by cell proliferation and the formation of callus. However, can we consider this two-step process as dedifferentiation? To do so, we have to accept that the callus cells are the developmental predecessors of mesophyll cells.
4. The Formation and Nature of Callus Tissues
For decades, callus was described as a mass of “undifferentiated” or “dedifferentiated” cells. This view is increasingly being challenged by molecular data suggesting that callus is not a developmental void but rather a specific tissue state with a characteristic gene expression signature [7,11,41,80]. A comprehensive comparison of transcriptome datasets from Arabidopsis calli of various origins and ages has highlighted a core set of 21 consistently upregulated genes that may define this state [11].
If one accepts this gene set as a relevant molecular signature, it is highly informative of the nature of the callus tissue. It includes key transcription factors that are central to embryonic and post-embryonic development, particularly in the formation and maintenance of meristems and organ primordia [11]. Among these are AUXIN RESPONSE FACTOR 5/MONOPTEROS (ARF5/MP) and AINTEGUMENTA-LIKE 6/PLETHORA 3 (AIL6/PLT3), which are important regulators controlling ground, vascular, and stem cell identities, meristem formation, maintenance, and function, as well as the initiation and development of lateral organ primordia (e.g., [81,82,83,84,85,86,87,88,89]). Other core callus genes include the TFs SPATULA (SPT) and WRKY23, both of which are involved in regulating meristem size and auxin-dependent developmental processes [90,91,92,93]. The consistent expression of these and other meristem- and provascular-associated genes [11,80] suggests that calli, at least in their certain regions [80], represent a state characteristic for a developmentally plastic (embryonic, meristematic, primordial, provascular) ground tissue. In in vitro culture, this state is blocked from proceeding to organized development such as embryo- or organogenesis by the exogenous hormonal conditions of the culture medium, leading to overproliferation. The same may hold true for wound-induced in planta callus formation depending on hormonal gradients and sensitivities of the damaged tissues/organs. The (pre- or post) meristematic ground tissue nature of calli explains their high regenerative potential, as it provides a population of cells that can be readily induced to form new meristems and organs upon transfer to appropriate regeneration media.
If we accept that calli resemble the embryonic/meristematic ground tissue from which the internal tissues differentiate, callus formation from these tissues can be considered as de facto dedifferentiation in its developmental biology sense (stepping back in the developmental lineage). Interestingly, organ or embryo regeneration from epidermal tissues, often from the base cells of glandular trichomes, proceeds directly without callus formation [94,95,96]. However, protoplasts of epidermal cells can also divide and form cell colonies [97]. Since epidermal cells are not of ground tissue origin, this might not be considered as dedifferentiation in the classical sense, except the callus represents a very early, pre-meristematic, embryonic stage [98]. Another explanation can be, that during in vitro culture conditions cell differentiation cannot revert back from protoplastation-induced senescence and thus the cells acquire a new fate. Actually, they differentiate and do not dedifferentiate into callus. Extensive chromatin remodelling and stochastic gene expression are evoked by stress and exogenous hormones, which might open the avenue for this developmental shift [68].
The dramatic reprogramming of terminally differentiated cells via a senescence-like intermediate is not the only route to callus formation. A substantial body of evidence indicates that, particularly in response to exogenous auxin, callus can arise from specific, pre-existing populations of competent cells [8,10,17,99,100].
In Arabidopsis, auxin-induced callus from various explants, including roots and aerial organs, has been shown to originate from pericycle or pericycle-like cells [99,100,101]. This process hijacks the developmental program for lateral root formation. The signalling cascade involves the auxin-mediated degradation of repressor proteins, leading to the activation of ARF7 and ARF19, which in turn induce the expression of LATERAL ORGAN BOUNDARY (LBD) transcription factors [10,41,101]. In this scenario, the callus can be viewed as an over-proliferating, disorganized lateral root primordium [7,11], which forms from cells that are already endowed with a degree of developmental plasticity [8,100,102,103]. Therefore, auxin-induced callus formation in intact tissues not involving differentiated somatic cells might not be considered as a classic example of dedifferentiation.
5. Summary
The evidence reviewed here suggests that plant cell dedifferentiation is not a monolithic process but a collection of context-dependent strategies. I propose a model that integrates these observations. The two-step pathway — (1) stress/wound-induced entry into a state of senescence-like transient competence, followed by (2) hormonal rescue into a proliferative callus state — may represent the mechanism required to reprogram terminally differentiated cells that lack inherent plasticity, such as leaf mesophyll cells. The initial stress response, which shares features with developmental and stress-induced senescence, effectively erases the differentiated state at the chromatin and gene expression levels, creating a blank slate upon which a new developmental program can be written. Without the subsequent proliferative signals from exogenous and/or endogenous hormones (auxin and cytokinin), this state is a dead end, leading to cellular demise.
In contrast, the alternative pathway, which involves the direct activation of pre-existing competent cells like those of the pericycle, represents a more streamlined route to regeneration. These cells, by their nature and position, retain a higher degree of developmental potential and may not need to undergo the drastic and risky process of complete reprogramming via a senescence-like intermediate. Instead, they can be directly co-opted into a new developmental trajectory, such as lateral root formation, which can be diverted into less organised callus growth under the influence of exogenous and/or endogenous hormones, especially auxin and cytokinin.
These two pathways may not be mutually exclusive but could represent two ends of a continuum of cellular plasticity. The extent of reprogramming required likely depends on the starting cell type’s differentiation status and its proximity to stem cell niches. Wound-induced callus formation, for example, is triggered by stress and involves epigenetic reprogramming, but the responding cells must be primed for division, blurring the lines between the two models. This framework also has implications for other regenerative processes such as somatic embryogenesis. This process is often induced by stress and involves extensive genetic reprogramming, but it may follow different pathways of single or multiple cell origin [4,104].
6. Conclusions
The dedifferentiation of plant cells is a cornerstone of their developmental plasticity and regenerative capacity. I have proposed a model that distinguishes between two primary modes of reprogramming. The first is a two-step process where severe stress pushes a terminally differentiated cell into a transient, senescence-like state of competence, from which it can be rescued into a proliferative, pre-meristematic callus state by hormonal signals. The second is a more direct pathway where callus is initiated from pre-existing competent cells that are already poised for division and development. Recognizing that “dedifferentiation” encompasses distinct biological processes is essential for advancing our fundamental understanding of plant regeneration. This view beside clarifying long-standing terminology debates might also help improving plant tissue culture and biotechnological applications, which depend on the efficient and predictable manipulation of plant cell fate.
Acknowledgments
The publication was prepared inassociation with a research project supported by the National Research, Development and Innovation Office under the number NKFI-1 K 146364.
References
- Birnbaum, K.D.; Alvarado, A.S. Slicing across Kingdoms: Regeneration in Plants and Animals. Cell 2008, 132, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, F.; Canher, B.; Lanssens, F.; Bisht, A.; Stael, S.; De Veylder, L.; Heyman, J. Pars Pro Toto: Every Single Cell Matters. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Mironova, V.; Xu, J. A Single-Cell View of Tissue Regeneration in Plants. Curr. Opin. Plant Biol. 2019, 52, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Fehér, A. Somatic Embryogenesis - Stress-Induced Remodeling of Plant Cell Fate. Biochim. Biophys. Acta - Gene Regul. Mech. 2015, 1849. [Google Scholar] [CrossRef]
- Lup, S.D.; Tian, X.; Xu, J.; Pérez-Pérez, J.M. Wound Signaling of Regenerative Cell Reprogramming. Plant Sci. 2016, 250, 178–187. [Google Scholar] [CrossRef]
- Jopling, C.; Boue, S.; Belmonte, J.C.I. Dedifferentiation, Transdifferentiation and Reprogramming: Three Routes to Regeneration. Nat. Rev. Mol. Cell Biol. 2011, 12, 79–89. [Google Scholar] [CrossRef]
- Fehér, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in Plants and Animals: Dedifferentiation, Transdifferentiation, or Just Differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef]
- Jiang, F.; Zhu, J.; Liu, H.-L. Protoplasts: A Useful Research System for Plant Cell Biology, Especially Dedifferentiation. Protoplasma 2013, 250, 1231–1238. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of Induction and Repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef]
- Fehér, A. A Common Molecular Signature Indicates the Pre-Meristematic State of Plant Calli. Int. J. Mol. Sci. 2023, 24, 13122. [Google Scholar] [CrossRef] [PubMed]
- Kopeina, G.S.; Efimenko, A.Yu.; Tkachuk, V.A.; Zhivotovsky, B. Programmed Cell Death and Tissue Regeneration: A Link That Should Be Resolved. Physiol. Rev. 2025, 105, 2377–2412. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Y.; Steller, H. Live to Die Another Way: Modes of Programmed Cell Death and the Signals Emanating from Dying Cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 329–344. [Google Scholar] [CrossRef]
- Vriz, S.; Reiter, S.; Galliot, B. Cell Death: A Program to Regenerate. Curr. Top. Dev. Biol. 2014, 108, 121–151. [Google Scholar] [CrossRef] [PubMed]
- Guerin, D.J.; Kha, C.X.; Tseng, K.A.-S. From Cell Death to Regeneration: Rebuilding After Injury. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Li, F.; Huang, Q.; Chen, J.; Peng, Y.; Roop, D.R.; Bedford, J.S.; Li, C.-Y. Apoptotic Cells Activate the “Phoenix Rising” Pathway to Promote Wound Healing and Tissue Regeneration. Sci. Signal. 2010, 3, ra13–ra13. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant Regeneration: Cellular Origins and Molecular Mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Sugimoto, K.; Temman, H.; Kadokura, S.; Matsunaga, S. To Regenerate or Not to Regenerate: Factors That Drive Plant Regeneration. Curr. Opin. Plant Biol. 2019, 47, 138–150. [Google Scholar] [CrossRef]
- Ibáñez, S.; Carneros, E.; Testillano, P.S.; Pérez-Pérez, J.M. Advances in Plant Regeneration: Shake, Rattle and Roll. Plants 2020, 9, 897. [Google Scholar] [CrossRef]
- Daneva, A.; Gao, Z.; Durme, M.V.; Nowack, M.K. Functions and Regulation of Programmed Cell Death in Plant Development. Annu. Rev. Cell Dev. Biol. 2016, 32, 441–468. [Google Scholar] [CrossRef]
- Xu, D.; Yang, L. Regeneration and Defense: Unveiling the Molecular Interplay in Plants. New Phytol. 2025, 246, 2484–2494. [Google Scholar] [CrossRef] [PubMed]
- Vega-Muñoz, I.; Duran-Flores, D.; Fernández-Fernández, Á.D.; Heyman, J.; Ritter, A.; Stael, S. Breaking Bad News: Dynamic Molecular Mechanisms of Wound Response in Plants. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Hoermayer, L.; Montesinos, J.C.; Marhava, P.; Benková, E.; Yoshida, S.; Friml, J. Wounding-Induced Changes in Cellular Pressure and Localized Auxin Signalling Spatially Coordinate Restorative Divisions in Roots. Proc. Natl. Acad. Sci. 2020, 117, 15322–15331. [Google Scholar] [CrossRef]
- Heil, M.; Ibarra-Laclette, E.; Adame-Álvarez, R.M.; Martínez, O.; Ramirez-Chávez, E.; Molina-Torres, J.; Herrera-Estrella, L. How Plants Sense Wounds: Damaged-Self Recognition Is Based on Plant-Derived Elicitors and Induces Octadecanoid Signaling. PLOS ONE 2012, 7, e30537. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Somerville, C.R. Imaging Plant Cell Death: GFP-Nit1 Aggregation Marks an Early Step of Wound and Herbicide Induced Cell Death. BMC Plant Biol. 2005, 5, 4. [Google Scholar] [CrossRef]
- Heyman, J.; Cools, T.; Canher, B.; Shavialenka, S.; Traas, J.; Vercauteren, I.; Van den Daele, H.; Persiau, G.; De Jaeger, G.; Sugimoto, K.; et al. The Heterodimeric Transcription Factor Complex ERF115–PAT1 Grants Regeneration Competence. Nat. Plants 2016, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Iakimova, E.T.; Woltering, E.J. The Wound Response in Fresh-Cut Lettuce Involves Programmed Cell Death Events. Protoplasma 2018, 255, 1225–1238. [Google Scholar] [CrossRef]
- Fagundes, D.; Bohn, B.; Cabreira, C.; Leipelt, F.; Dias, N.; Bodanese-Zanettini, M.H.; Cagliari, A. Caspases in Plants: Metacaspase Gene Family in Plant Stress Responses. Funct. Integr. Genomics 2015, 15, 639–649. [Google Scholar] [CrossRef]
- Hander, T.; Fernández-Fernández, Á.D.; Kumpf, R.P.; Willems, P.; Schatowitz, H.; Rombaut, D.; Staes, A.; Nolf, J.; Pottie, R.; Yao, P.; et al. Damage on Plants Activates Ca2+-Dependent Metacaspases for Release of Immunomodulatory Peptides. Science 2019, 363, eaar7486. [Google Scholar] [CrossRef]
- Yang, W.; Zhai, H.; Wu, F.; Deng, L.; Chao, Y.; Meng, X.; Chen, Q.; Liu, C.; Bie, X.; Sun, C.; et al. Peptide REF1 Is a Local Wound Signal Promoting Plant Regeneration. Cell 2024, 187, 3024–3038.e14. [Google Scholar] [CrossRef]
- Geng, X.; Zhang, X.; Zhou, W. Triggering Regeneration: A New Recognized Local Wound Signal in Plants. Innov. Life 2025, 3, 100124–2. [Google Scholar] [CrossRef]
- Iwase, A.; Mitsuda, N.; Koyama, T.; Hiratsu, K.; Kojima, M.; Arai, T.; Inoue, Y.; Seki, M.; Sakakibara, H.; Sugimoto, K.; et al. The AP2/ERF Transcription Factor WIND1 Controls Cell Dedifferentiation in Arabidopsis. Curr. Biol. 2011, 21, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Sano, H.; Ohashi, Y. Jasmonic Acid in Wound Signal Transduction Pathways. Physiol. Plant. 1997, 101, 740–745. [Google Scholar] [CrossRef]
- Reinbothe, C.; Springer, A.; Samol, I.; Reinbothe, S. Plant Oxylipins: Role of Jasmonic Acid during Programmed Cell Death, Defence and Leaf Senescence. FEBS J. 2009, 276, 4666–4681. [Google Scholar] [CrossRef]
- Kim, J.; Chang, C.; Tucker, M.L. To Grow Old: Regulatory Role of Ethylene and Jasmonic Acid in Senescence. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Favero, D.S.; Sakamoto, Y.; Iwase, A.; Coleman, D.; Rymen, B.; Sugimoto, K. Molecular Mechanisms of Plant Regeneration. Annu. Rev. Plant Biol. 2019, 70, 377–406. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Rymen, B.; Sugimoto, K. How Do Plants Transduce Wound Signals to Induce Tissue Repair and Organ Regeneration? Curr. Opin. Plant Biol. 2020, 57, 72–77. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.-X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-Mediated Wound Signalling Promotes Plant Regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, W.; Gu, Z.; Wu, S.; E, Y.; Zhou, W.; Lin, J.; Xu, L. Roles of the Wound Hormone Jasmonate in Plant Regeneration. J. Exp. Bot. 2021, erab508. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Lozano-Torres, J.L.; Blilou, I.; Zhang, X.; Zhai, Q.; Smant, G.; Li, C.; Scheres, B. A Jasmonate Signaling Network Activates Root Stem Cells and Promotes Regeneration. Cell 2019, 177, 942–956.e14. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding Triggers Callus Formation via Dynamic Hormonal and Transcriptional Changes1[OPEN]. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar] [CrossRef]
- Maruyama, K.; Ikeuchi, M. Multifaceted Controls on Auxin Metabolism during Cellular Reprogramming and Organ Regeneration in Plants. J. Exp. Bot. 2026, 77, 141–148. [Google Scholar] [CrossRef]
- Omary, M.; Matosevich, R.; Efroni, I. Systemic Control of Plant Regeneration and Wound Repair. New Phytol. 2023, 237, 408–413. [Google Scholar] [CrossRef]
- Kacprzyk, J.; Burke, R.; Schwarze, J.; McCabe, P.F. Plant Programmed Cell Death Meets Auxin Signalling. FEBS J. 2022, 289, 1731–1745. [Google Scholar] [CrossRef]
- Heyman, J.; Cools, T.; Vandenbussche, F.; Heyndrickx, K.S.; Van Leene, J.; Vercauteren, I.; Vanderauwera, S.; Vandepoele, K.; De Jaeger, G.; Van Der Straeten, D.; et al. ERF115 Controls Root Quiescent Center Cell Division and Stem Cell Replenishment. Science 2013, 342, 860–863. [Google Scholar] [CrossRef]
- Geem, K.R.; Kim, H.; Ryu, H. SCFFBS1 Regulates Root Quiescent Center Cell Division via Protein Degradation of APC/CCCS52A2. Mol. Cells 2022, 45, 695–701. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Hong, J.H.; Gong, X.; Zhou, C.; Pérez-Pérez, J.M.; Xu, J. TOPOISOMERASE1α Acts through Two Distinct Mechanisms to Regulate Stele and Columella Stem Cell Maintenance. Plant Physiol. 2016, 171, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Raya-González, J.; Oropeza-Aburto, A.; López-Bucio, J.S.; Guevara-García, Á.A.; de Veylder, L.; López-Bucio, J.; Herrera-Estrella, L. MEDIATOR18 Influences Arabidopsis Root Architecture, Represses Auxin Signaling and Is a Critical Factor for Cell Viability in Root Meristems. Plant J. 2018, 96, 895–909. [Google Scholar] [CrossRef]
- Kong, X.; Tian, H.; Yu, Q.; Zhang, F.; Wang, R.; Gao, S.; Xu, W.; Liu, J.; Shani, E.; Fu, C.; et al. PHB3 Maintains Root Stem Cell Niche Identity through ROS-Responsive AP2/ERF Transcription Factors in Arabidopsis. Cell Rep. 2018, 22, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.B.; He, S.; Meyer, T.J.; Szurman-Zubrzycka, M.; Alfs, C.; Kwasniewska, J.; Pervis, A.; Gajecka, M.; Veerabahu, A.; Beaulieu, T.R.; et al. The Stem Cell Niche Transcription Factor ETHYLENE RESPONSE FACTOR 115 Participates in Aluminum-Induced Terminal Differentiation in Arabidopsis Roots. Plant Cell Environ. 2024, 47, 4432–4448. [Google Scholar] [CrossRef] [PubMed]
- Canher, B.; Heyman, J.; Savina, M.; Devendran, A.; Eekhout, T.; Vercauteren, I.; Prinsen, E.; Matosevich, R.; Xu, J.; Mironova, V.; et al. Rocks in the Auxin Stream: Wound-Induced Auxin Accumulation and ERF115 Expression Synergistically Drive Stem Cell Regeneration. Proc. Natl. Acad. Sci. U. S. A 2020, 117, 16667–16677. [Google Scholar] [CrossRef]
- Chang, X.; Riemann, M.; Liu, Q.; Nick, P. Actin as Deathly Switch? How Auxin Can Suppress Cell-Death Related Defence. PLOS ONE 2015, 10, e0125498. [Google Scholar] [CrossRef]
- Hong, J.H.; Savina, M.; Du, J.; Devendran, A.; Kannivadi Ramakanth, K.; Tian, X.; Sim, W.S.; Mironova, V.V.; Xu, J. A Sacrifice-for-Survival Mechanism Protects Root Stem Cell Niche from Chilling Stress. Cell 2017, 170, 102–113.e14. [Google Scholar] [CrossRef]
- Xuan, W.; Band, L.R.; Kumpf, R.P.; Van Damme, D.; Parizot, B.; De Rop, G.; Opdenacker, D.; Möller, B.K.; Skorzinski, N.; Njo, M.F.; et al. Cyclic Programmed Cell Death Stimulates Hormone Signaling and Root Development in Arabidopsis. Science 2016, 351, 384–387. [Google Scholar] [CrossRef]
- Sheldrake, A.R. The Production of Auxin by Dying Cells. J. Exp. Bot. 2021, 72, 2288–2300. [Google Scholar] [CrossRef]
- Matosevich, R.; Cohen, I.; Gil-Yarom, N.; Modrego, A.; Friedlander-Shani, L.; Verna, C.; Scarpella, E.; Efroni, I. Local Auxin Biosynthesis Is Required for Root Regeneration after Wounding. Nat. Plants 2020, 6, 1020–1030. [Google Scholar] [CrossRef]
- Ribeiro, C.; de Melo, B.P.; Lourenço-Tessutti, I.T.; Ballesteros, H.F.; Ribeiro, K.V.G.; Menuet, K.; Heyman, J.; Hemerly, A.; de Sá, M.F.G.; De Veylder, L.; et al. The Regeneration Conferring Transcription Factor Complex ERF115-PAT1 Coordinates a Wound-Induced Response in Root-Knot Nematode Induced Galls. New Phytol. 2024, 241, 878–895. [Google Scholar] [CrossRef] [PubMed]
- Bisht, A.; Eekhout, T.; Canher, B.; Lu, R.; Vercauteren, I.; De Jaeger, G.; Heyman, J.; De Veylder, L. PAT1-Type GRAS-Domain Proteins Control Regeneration by Activating DOF3.4 to Drive Cell Proliferation in Arabidopsis Roots. Plant Cell 2023, 35, 1513–1531. [Google Scholar] [CrossRef] [PubMed]
- Chupeau, M.-C.; Granier, F.; Pichon, O.; Renou, J.-P.; Gaudin, V.; Chupeau, Y. Characterization of the Early Events Leading to Totipotency in an Arabidopsis Protoplast Liquid Culture by Temporal Transcript Profiling. Plant Cell 2013, 25, 2444–2463. [Google Scholar] [CrossRef]
- Grosset, J.; Marty, I.; Chartier, Y.; Meyer, Y. mRNAs Newly Synthesized by Tobacco Mesophyll Protoplasts Are Wound-Inducible. Plant Mol. Biol. 1990, 15, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, A.K.; Roubelakis-Angelakis, K.A. The Generation of Active Oxygen Species Differs in Tobacco and Grapevine Mesophyll Protoplasts. Plant Physiol. 1999, 121, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Suzuki, K.; Kawasaki, H.; Watanabe, Y. Differential Responses of Brassica Napus and Petunia Hybrida to Leaf Protoplast Isolation Stress. Physiol. Plant. 2002, 114, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kawasaki, H.; Itho, Y.; Watanabe, Y. Senescence Development of Brassica Napus Leaf Protoplast during Isolation and Subsequent Culture. J. Plant Physiol. 1998, 152, 487–493. [Google Scholar] [CrossRef]
- Watanabe, M.; Setoguchi, D.; Uehara, K.; Ohtsuka, W.; Watanabe, Y. Apoptosis-like Cell Death of Brassica Napus Leaf Protoplasts. New Phytol. 2002, 156, 417–426. [Google Scholar] [CrossRef]
- Damri, M.; Granot, G.; Ben-Meir, H.; Avivi, Y.; Plaschkes, I.; Chalifa-Caspi, V.; Wolfson, M.; Fraifeld, V.; Grafi, G. Senescing Cells Share Common Features with Dedifferentiating Cells. Rejuvenation Res. 2009, 12, 435–443. [Google Scholar] [CrossRef]
- Grafi, G.; Chalifa-Caspi, V.; Nagar, T.; Plaschkes, I.; Barak, S.; Ransbotyn, V. Plant Response to Stress Meets Dedifferentiation. Planta 2011, 233, 433–438. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Li, S.; Tan, S.; Cao, J.; Wang, H.-L.; Luo, J.; Guo, H.; Zhang, Z.; Li, Z. Leaf Senescence Database v5.0: A Comprehensive Repository for Facilitating Plant Senescence Research. J. Mol. Biol. 2024, 436, 168530. [Google Scholar] [CrossRef]
- Xu, M.; Du, Q.; Tian, C.; Wang, Y.; Jiao, Y. Stochastic Gene Expression Drives Mesophyll Protoplast Regeneration. Sci. Adv. 2021. [Google Scholar] [CrossRef]
- Heyman, J.; Cools, T.; Vandenbussche, F.; Heyndrickx, K.S.; Van Leene, J.; Vercauteren, I.; Vanderauwera, S.; Vandepoele, K.; De Jaeger, G.; Van Der Straeten, D.; et al. ERF115 Controls Root Quiescent Center Cell Division and Stem Cell Replenishment. Science 2013, 342, 860–863. [Google Scholar] [CrossRef]
- Johnson, R.A.; Conklin, P.A.; Tjahjadi, M.; Missirian, V.; Toal, T.; Brady, S.M.; Britt, A.B. SUPPRESSOR OF GAMMA RESPONSE1 Links DNA Damage Response to Organ Regeneration. Plant Physiol. 2018, 176, 1665–1675. [Google Scholar] [CrossRef]
- Zhao, J.; Morozova, N.; Williams, L.; Libs, L.; Avivi, Y.; Grafi, G. Two Phases of Chromatin Decondensation during Dedifferentiation of Plant Cells: Distinction between Competence for Cell Fate Switch and a Commitment for S Phase. J. Biol. Chem. 2001, 276, 22772–22778. [Google Scholar] [CrossRef]
- Florentin, A.; Damri, M.; Grafi, G. Stress Induces Plant Somatic Cells to Acquire Some Features of Stem Cells Accompanied by Selective Chromatin Reorganization. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2013, 242, 1121–1133. [Google Scholar] [CrossRef]
- Grafi, G. How Cells Dedifferentiate: A Lesson from Plants. Dev. Biol. 2004, 268, 1–6. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Kawamura, A.; Suzuki, T.; Segami, S.; Maeshima, M.; Polyn, S.; De Veylder, L.; Sugimoto, K. Transcriptional Activation of Auxin Biosynthesis Drives Developmental Reprogramming of Differentiated Cells. Plant Cell 2022, 34, 4348–4365. [Google Scholar] [CrossRef]
- Williams, L.; Zhao, J.; Morozova, N.; Li, Y.; Avivi, Y.; Grafi, G. Chromatin Reorganization Accompanying Cellular Dedifferentiation Is Associated with Modifications of Histone H3, Redistribution of HP1, and Activation of E2F-Target Genes. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2003, 228, 113–120. [Google Scholar] [CrossRef]
- Avivi, Y.; Morad, V.; Ben-Meir, H.; Zhao, J.; Kashkush, K.; Tzfira, T.; Citovsky, V.; Grafi, G. Reorganization of Specific Chromosomal Domains and Activation of Silent Genes in Plant Cells Acquiring Pluripotentiality. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 230, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, T.; Miskolczi, P.; Ayaydin, F.; Mészáros, T.; Dudits, D.; Fehér, A. Exogenous Auxin and Cytokinin Dependent Activation of CDKs and Cell Division in Leaf Protoplast-Derived Cells of Alfalfa. 2000, Vol. 32, 129–141. [Google Scholar] [CrossRef]
- Fehér, A.; Ötvös, K.; Pasternak, T.P.; Szandtner, A.P. The Involvement of Reactive Oxygen Species (ROS) in the Cell Cycle Activation (G 0-to-G 1 Transition) of Plant Cells. 2007, Vol. 51, 823–826. [Google Scholar]
- Lee, K.; Park, O.-S.; Jung, S.-J.; Seo, P.J. Histone Deacetylation-Mediated Cellular Dedifferentiation in Arabidopsis. J. Plant Physiol. 2016, 191, 95–100. [Google Scholar] [CrossRef]
- Zhai, N.; Xu, L. Pluripotency Acquisition in the Middle Cell Layer of Callus Is Required for Organ Regeneration. Nat. Plants 2021, 7, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Berleth, T. The Arabidopsis Gene MONOPTEROS Encodes a Transcription Factor Mediating Embryo Axis Formation and Vascular Development. EMBO J. 1998, 17, 1405–1411. [Google Scholar] [CrossRef]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.-S.; Amasino, R.; Scheres, B. The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Schlereth, A.; Möller, B.; Liu, W.; Kientz, M.; Flipse, J.; Rademacher, E.H.; Schmid, M.; Jürgens, G.; Weijers, D. MONOPTEROS Controls Embryonic Root Initiation by Regulating a Mobile Transcription Factor. Nature 2010, 464, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Hofhuis, H.; Laskowski, M.; Du, Y.; Prasad, K.; Grigg, S.; Pinon, V.; Scheres, B. Phyllotaxis and Rhizotaxis in Arabidopsis Are Modified by Three PLETHORA Transcription Factors. Curr. Biol. 2013, 23, 956–962. [Google Scholar] [CrossRef]
- Horstman, A.; Willemsen, V.; Boutilier, K.; Heidstra, R. AINTEGUMENTA-LIKE Proteins: Hubs in a Plethora of Networks. Trends Plant Sci. 2014, 19, 146–157. [Google Scholar] [CrossRef]
- Krogan, N.T.; Marcos, D.; Weiner, A.I.; Berleth, T. The Auxin Response Factor MONOPTEROS Controls Meristem Function and Organogenesis in Both the Shoot and Root through the Direct Regulation of PIN Genes. New Phytol. 2016, 212, 42–50. [Google Scholar] [CrossRef]
- QingQing, L.; WenBin, N.; HanMa, Z. Progress on the functions of ARF5/MONOPTEROS(MP). Acta Bot. Boreali-Occident. Sin. 2016, 36, 197–203. [Google Scholar]
- Du, Y.; Scheres, B. PLETHORA Transcription Factors Orchestrate de Novo Organ Patterning during Arabidopsis Lateral Root Outgrowth. Proc. Natl. Acad. Sci. 2017, 114, 11709–11714. [Google Scholar] [CrossRef] [PubMed]
- Burkart, R.C.; Strotmann, V.I.; Kirschner, G.K.; Akinci, A.; Czempik, L.; Dolata, A.; Maizel, A.; Weidtkamp-Peters, S.; Stahl, Y. PLETHORA-WOX5 Interaction and Subnuclear Localization Control Arabidopsis Root Stem Cell Maintenance. EMBO Rep. 2022, 23, e54105. [Google Scholar] [CrossRef]
- Grunewald, W.; De Smet, I.; Lewis, D.R.; Löfke, C.; Jansen, L.; Goeminne, G.; Vanden Bossche, R.; Karimi, M.; De Rybel, B.; Vanholme, B.; et al. Transcription Factor WRKY23 Assists Auxin Distribution Patterns during Arabidopsis Root Development through Local Control on Flavonol Biosynthesis. Proc. Natl. Acad. Sci. U. S. A 2012, 109, 1554–1559. [Google Scholar] [CrossRef]
- Grunewald, W.; De Smet, I.; De Rybel, B.; Robert, H.S.; van de Cotte, B.; Willemsen, V.; Gheysen, G.; Weijers, D.; Friml, J.; Beeckman, T. Tightly Controlled WRKY23 Expression Mediates Arabidopsis Embryo Development. EMBO Rep. 2013, 14, 1136–1142. [Google Scholar] [CrossRef]
- Makkena, S.; Lamb, R.S. The bHLH Transcription Factor SPATULA Regulates Root Growth by Controlling the Size of the Root Meristem. BMC Plant Biol. 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Prát, T.; Hajný, J.; Grunewald, W.; Vasileva, M.; Molnár, G.; Tejos, R.; Schmid, M.; Sauer, M.; Friml, J. WRKY23 Is a Component of the Transcriptional Network Mediating Auxin Feedback on PIN Polarity. PLOS Genet. 2018, 14, e1007177. [Google Scholar] [CrossRef] [PubMed]
- Chlyah, A.; Van, M.T.T. Differential Reactivity in Epidermal Cells of Begonia Rex Excised and Grown in Vitro. Physiol. Plant. 1975, 35, 16–20. [Google Scholar] [CrossRef]
- Venverloo, C.J.; Koster, J.; Libbenga, K.R. The Formation of Adventitious Organs: IV. The Ontogeny of Shoots and Leaves From Epidermis Cells of Nautilocalyx Lynchii. Z. Für Pflanzenphysiol 1983, 109, 55–67. [Google Scholar] [CrossRef]
- Morinaka, H.; Coleman, D.; Sugimoto, K.; Iwase, A. Molecular Mechanisms of Plant Regeneration from Differentiated Cells: Approaches from Historical Tissue Culture Systems. Plant Cell Physiol. 2023, 64, 297–304. [Google Scholar] [CrossRef]
- Davey, M.R.; Freason, E.M.; Withers, L.A.; Power, J.B. Observations on the Morphology, Ultrastructure and Regeneration of Tobacco Leaf Epidermal Protoplasts. Plant Sci. Lett. 1974, 2, 23–27. [Google Scholar] [CrossRef]
- Javelle, M.; Vernoud, V.; Rogowsky, P.M.; Ingram, G.C. Epidermis: The Formation and Functions of a Fundamental Plant Tissue. New Phytol. 2011, 189, 17–39. [Google Scholar] [CrossRef]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis Xylem Pericycle Underlies Shoot Regeneration from Root and Hypocotyl Explants Grown in Vitro. Plant J. Cell Mol. Biol. 2009, 57, 626–644. [Google Scholar] [CrossRef]
- Sugimoto, K.; Jiao, Y.; Meyerowitz, E.M. Arabidopsis Regeneration from Multiple Tissues Occurs via a Root Development Pathway. Dev. Cell 2010, 18, 463–471. [Google Scholar] [CrossRef]
- Liu, H.; Wang, G.-C.; Feng, Z.; Zhu, J. Screening of Genes Associated with Dedifferentiation and Effect of LBD29 on Pericycle Cells in Arabidopsis Thaliana. Plant Growth Regul. 2010, 62, 127–136. [Google Scholar] [CrossRef]
- Xiao, W.; Molina, D.; Wunderling, A.; Ripper, D.; Vermeer, J.E.M.; Ragni, L. Pluripotent Pericycle Cells Trigger Different Growth Outputs by Integrating Developmental Cues into Distinct Regulatory Modules. Curr. Biol. 2020, 30, 4384–4398.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Umeda, M.; Kakimoto, T. Pericycle Cell Division Competence Underlies Various Developmental Programs. Plant Biotechnol. 2022, 39, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Fehér, A.; Bernula, D.; Gémes, K. The Many Ways of Somatic Embryo Initiation. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer International Publishing, 2016; pp. 23–37. ISBN 978-3-319-33705-0. [Google Scholar]
Figure 1.
Overlap among the Arabidopsis transcripts associated with leaf senescence [67] and expressed in freshly isolated leaf protoplasts [68].
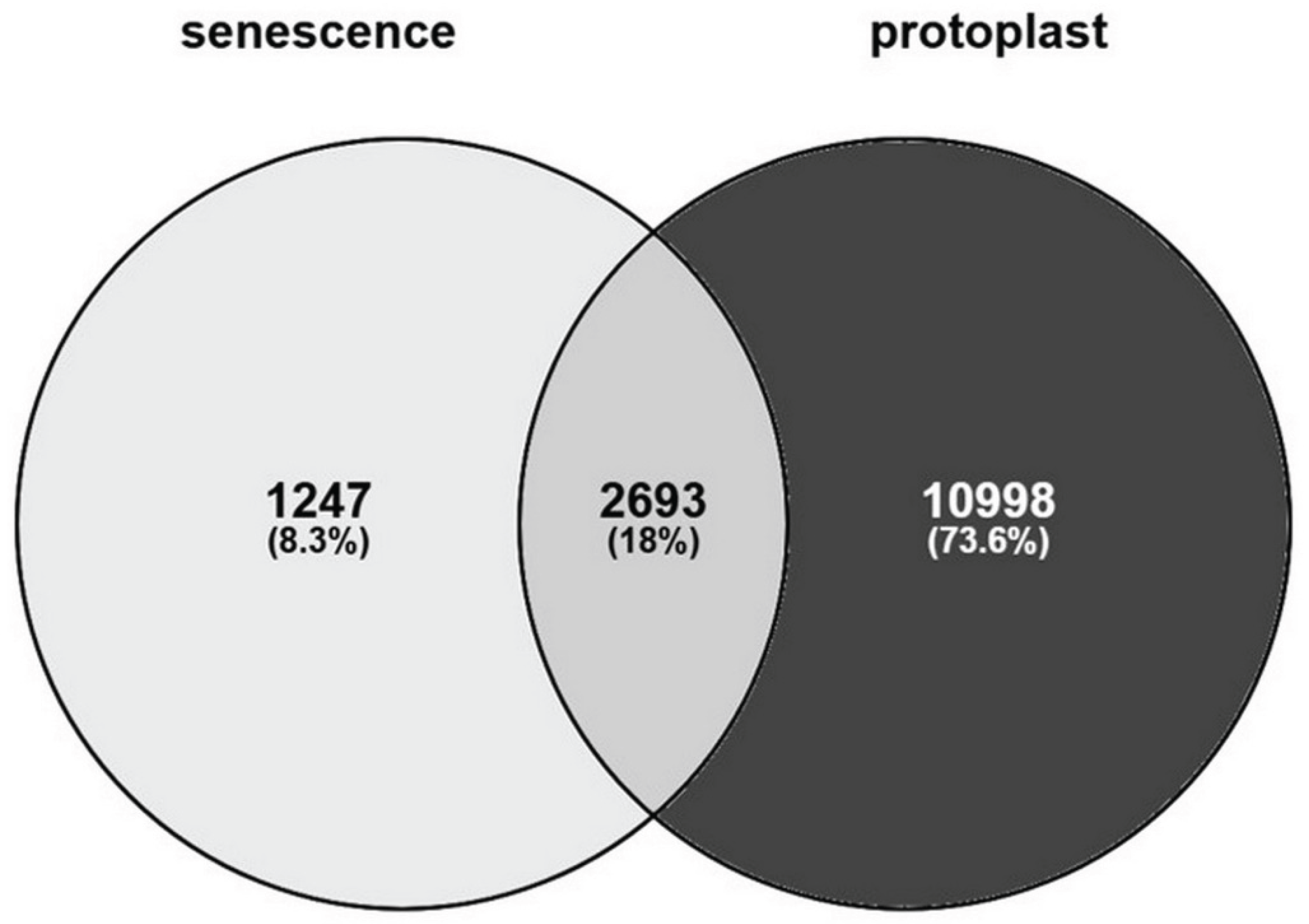

Table 1.
Differential expression of three regeneration-related transcription factors in freshly isolated and dividing 4-day-old Arabidopsis leaf protoplasts from the transcriptomic analysis of Xu et al. [68].
Table 1.
Differential expression of three regeneration-related transcription factors in freshly isolated and dividing 4-day-old Arabidopsis leaf protoplasts from the transcriptomic analysis of Xu et al. [68].
| protoplasts versus leaf cells | 4-day-old versus 0-day-old protoplasts | ||||||
| GID | log2FC | P value | FDR | log2FC | P value | FDR | |
| ERF115 | AT5G07310 | 11,45 | 1,8251E-37 | 0,00 | -1,31 | 0,00135717 | 0,00205361 |
| PAT1 | AT5G48150 | 0,75 | 8,1593E-06 | 1,3381E-05 | -2,18 | 3,7995E-35 | 4,3449E-34 |
| ARF5 | AT1G19850 | 6,59 | 1,169E-142 | 2,779E-140 | -0,79 | 2,7827E-05 | 4,9201E-05 |
log2FC – log2 value of fold change; P value – level of significance; FDR – fals discovery rate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



