Submitted:
03 January 2026
Posted:
05 January 2026
You are already at the latest version
Abstract
As sedentary lifestyles are becoming more common, related health conditions including obesity, diabetes, heart disease, etc. are also becoming more prevalent. Physical exercise and activity are proven to be important for improving health, however, less common yet just as important for overall health is using performance enhancing techniques. While there are a wide variety of performance enhancing techniques, this paper focuses on 4 of the most common: Cold-Water-Immersion (CWI), Heat Stress, Blood-Flow-Restriction (BFR), and sleep. Research has shown CWI decreases hypertrophy in response to resistance exercise and has been shown to increase some markers of endurance adaptations but has not shown long-term physiological benefits. Heat Stress has shown to improve both hypertrophy in response to resistance exercise and mitochondrial biogenesis in response to endurance exercise. BFR training has not been shown to contribute to significant improvements in exercise adaptations compared to traditional higher intensity resistance and endurance exercise. Finally, sleep deprivation has shown to significantly decrease hypertrophy and mitochondrial health, indicating sufficient sleep is very important to obtaining exercise adaptations and health benefits. All together, these results suggest the importance of performance enhancing techniques for overall skeletal muscle health.
Keywords:
cold-water-immersion
; heat stress
; blood-flow-restriction
; sleep
; resistance-exercise
; endurance-exercise
; mitochondria
; hypertrophy
; skeletal-muscle
; metabolism
1. Introduction
Skeletal muscle tissue comprises approximately 40% of the body’s mass making it the most abundant tissue in the human body [1]. It also contains approximately 50-75% of all body proteins [2]. Healthy skeletal muscle and regular physical activity are key factors of overall health. Skeletal muscle tissues release myokine signals throughout the body which improve bone health and inflammation. Myokine signals also regulate hippocampal function in the brain, reducing depression symptoms, and improving cognitive function and neurogenesis [3]. These myokines are often released in response to physical exercise. Physical activity has been shown to lower the risk for many health problems including heart disease, type 2 diabetes, and several cancers [4]. In fact, heart failure and cancer, two leading causes of death in the US, are associated with extensive loss of muscle, strength, and metabolic function [5]. Additionally, physical exercise is associated with improved circadian rhythm and sleep which is also critical for overall health [6]. Physical exercise and muscle health is especially important in aging people at risk of sarcopenia (loss of muscle mass and strength) as loss of muscle mass is associated with an increased risk of falls, decreased independence, and death. Skeletal muscle health reduces the risk of both muscular and skeletal injuries, improves psychological wellbeing, and increases functional strength, all improving quality of life.
Different types of exercise can be performed to improve muscle health leading to more beneficial future health outcomes. The two main types of exercise this review will cover are endurance exercise (EE) and resistance exercise (RE). EE is defined as long-duration, low intensity exercise that causes a sustained elevated heart rate. EE primarily creates adaptations by increasing oxidative capacity and ATP production in muscle cells, typically through improving mitochondria concentration, efficiency, and quality [7]. RE is performed by high-intensity movements that require near-maximal force output and primarily creates adaptations by stimulating hypertrophy and increasing muscle strength through anabolic pathways [8].
An important factor in healthy muscle is a high concentration of healthy mitochondria. Mitochondria are the organelles responsible for producing ATP, the source of energy used by skeletal muscle and are required to supply energy for muscle function. Markers of healthy mitochondria are consistent balanced rates of mitochondrial death (mitophagy) and synthesis (biogenesis). Balanced turnover of mitochondria is important to remove damaged mitochondria and replace them with high-quality, more efficient ones. Failure to remove dying mitochondria reduces energy availability and can contribute to disease [9]. High O2 consumption also signals active, healthy mitochondria. Over long periods of acute training, muscle tissue adapts to produce more mitochondria with higher electron transport chain (ETC; protein complex that converts energy to ATP) densities.
To maximize these beneficial adaptations from exercise, the muscles and systems involved must be adequately rested and recovered before further exercise bouts. Adequate recovery prevents muscle breakdown and reduces injury risk. Optimized recovery is especially beneficial to groups like athletes or individuals with high training loads, those with limited time to train, and aging or clinical populations. This review will discuss and compare the viability and usefulness for maximizing training adaptations of four different recovery methods popularized by the wellness space: sleep, Blood Flow Restriction (BFR), cold water immersion (CWI)/cold plunge, and heat intervention/sauna. These all have or have had prominent places in fitness culture, with controversy surrounding their benefits and tradeoffs.
To evaluate the effectiveness of these recovery strategies I will measure how they affect markers of mitochondrial health in skeletal muscle and markers related to hypertrophy (mTOR, p70S6K, MAFbx, MuRF1/TRIM63, MSTN activity). EE stimulates activity of PGC1-alpha and TFAM genes, which increase mtDNA content and oxidative enzyme activity [10]. It also increases the percentage of type I muscle fibers which have a higher mitochondrial concentration than type II muscle fibers [7]. RE promotes skeletal muscle health and strength by increasing activation of mTOR and p70S6K transcription (genes associated with increased muscle hypertrophy) [11] and decreasing activity of MAFbx and MuRF1/TRIM63 (genes associated with muscle atrophy) [12]. This review will consider an effective recovery method to be one that is also associated with amplification of one or both of these pathways.
This review is based on studies performed on a mix of human, rat, and mouse models and protocols for both RE and EE. The benefits of animal-based studies are that they are often easier to control and measure markers with more invasive testing but the drawbacks are that they are based on animal physiology and it can be harder to simulate RE protocols. Thus, I’ve included a mixture of both animal and human studies for a broader, more robust analysis.
2. Results
2.1. Cold Water Immersion
Cold water immersion (CWI) involves submerging specific limbs or the whole body (typically for periods of 5-20 minutes) into cold water (typically ~8-15℃) [13]. The popularity of CWI/cold plunging/cold showering has grown significantly in recent years and CWI has become especially popular in wellness related communities on the internet [14]. Its popularity is reflected by changes in the global cold plunge market, which is expected to reach $500 billion by 2028, at a 6.5% compound annual growth rate [15]. Numerous viral videos about CWI claim that it has beneficial effects on mood and the immune system, and speeds up recovery for athletes [16].
These claims about speeding up recovery are partially supported by current literature. Post exercise CWI has been shown to be effective in recovering muscle strength and decreasing muscle soreness, inflammation, damage, and perceptions of fatigue [13]. This could increase total training adaptations over time through enhancing the quality of recovery and thus subsequent workouts. Although these are notable benefits for reducing fatigue, CWI also comes with significant drawbacks. CWI has been found to attenuate long term muscle mass and strength gains by reducing the phosphorylation of p70S6K, a key gene in the mTOR pathway that contributes to protein synthesis, for 48 hours after exercise. It also reduced satellite cell activation (stem cells that differentiate into muscle cells), further decreasing hypertrophy after RE [17]. Several other studies have found that repeated use of CWI after EE is associated with increased activation of several genes associated with increasing activation of mitochondrial biogenesis pathways, including PGC1-α, p38, and AMPK [18,19,20]. However, one meta-analysis examining recovery after various forms of resistance and endurance exercise found that CWI had no significant effect on improving endurance performance after 24 and 48 hours [21]. A potential difference between these studies that could explain the different results is that participants of the first study performed recreational team sports, typically not achieving maximum aerobic effort. However, the majority of studies included in the meta-analysis used EE protocols that reached maximum aerobic effort. Another study of 30 males performing high intensity sprint sessions found that although CWI can improve activation of PGC1-α, CWI did not produce improvement in endurance performance compared to a placebo [22]. Because of these discrepancies, there is not sufficient data to support the idea that repeated CWI can improve long-term adaptations to endurance training. To improve datasets, experiments must be done on humans over a long-period of time, comparing CWI and non-CWI groups’ improvement on endurance fitness tests to better determine whether CWI after EE leads to beneficial molecular and cellular changes.
2.2. Heat Stress
Heat stress interventions (HSI), including sauna use or hot water immersion, involve increasing the body's temperature for a controlled period of time. Many forms of this practice have existed in numerous cultures around the world for centuries, but the practice has become popular in the fitness and wellness communities as of recently [23]. Advocates of heat stress practices claim it can reduce the risk of cardiovascular diseases, improve muscle recovery, and increase cardiovascular fitness.
A study of middle distance runners found that those who underwent a sauna bathing protocol showed significant improvements in both a running heat tolerance test and temperature exercise test compared to a non-sauna bathing control group [24]. HSI has also been found to increase activation of Heat Shock Proteins (HSPs), MTORC1, and PGC1-α. This leads to increased muscle mass through increased satellite cell proliferation and differentiation, and increased activation of the mTOR pathway [22]. It has also been found to inhibit autophagy and protein denaturation, which are major contributors to muscular atrophy by reducing mitochondrial function and causing weakness and increased mortality risk. HSI prevents autophagy and denaturation, increasing overall hypertrophy. These benefits apply to endurance adaptations as well. Through increasing activation of PGC1-α, mitochondrial homeostasis is increased.
HSI also improves markers of skeletal muscle rejuvenation. Protocols of heating muscle resulted in improved macrophage activity, satellite cell proliferation, and muscle fiber rejuvenation after skeletal muscle injury. These benefits could apply to uninjured muscle as well because both RE and EE can induce muscle damage after exercise [25,26]. This can allow for a greater number of training sessions to be performed at a higher quality, increasing long-term adaptations.
2.3. Blood Flow Restriction
Blood flow restriction (BFR) training is the practice of applying a specialized tourniquet cuff to the proximal portion of an exercising extremity to restrict arterial inflow and venous outflow. The pressure used is typically 40-80% of an individual's limb occlusion pressure (LOP). BFR is typically applied either during active exercise or while resting in between sets and is used for both resistance and endurance exercise [27]. The practice originated in Japan in the 1960s and was developed initially as Kaatsu training by Dr. Yoshiaki Sato as he used the technique to rehabilitate a fracture in his leg. The practice gained public notoriety and shortly began being researched more significantly [28]. Since the 1960s its popularity has grown significantly in rehab and clinical populations, and strength training communities. One of the most popular claims about BFR is that it simulates conditions associated with high-intensity training within muscles, allowing the same training adaptations to be achieved with lower weight or endurance activity intensity [27]. This can be beneficial because if less intensity is used, the training is safer for at-risk populations such as the elderly or injured, and aims in lessening systemic fatigue.
One group did a study of 18 patients who had undergone unilateral ACL reconstruction. They divided these patients into BFR, SHAM-BFR, and control groups. Muscle biopsies found that change in quadriceps cross-sectional area (CSA) was significantly larger for the BFR group (4.9%) compared to SHAM-BFR group (1.3%). The same was true for change in peak torque and fatigue of knee extensors between BFR (14%, 6%) and SHAM-BFR (-1%, 2%). Both of these changes are signs of hypertrophy. A greater increase in these performances indicate greater activation of hypertrophy. mRNA expression of myostatin was reduced after both BFR and SHAM-BFR training [29]. Another study of BFR on patients undergoing cast immobilization found that a BFR group had a smaller increase in MURF1 expression after immobilization compared to a control group and the BFR group also had a smaller decrease in CSA of vastus lateralis and thigh flexor muscles. MURF1 and CSA decrease are signs of muscle atrophy. Attenuation of increase in MURF1 expression and CSA decrease show that BFR training is effective at reducing muscle atrophy [30].
A study comparing low-intensity BFR (30% 1 repetition max (RM)) to traditional high-intensity (70% 1RM) resistance training in ACL reconstruction patients found that the BFR group had significantly greater reduction in peak knee extensor and knee flexor torque loss. Self-reported function, Y-balance performance, and knee joint pain was also significantly improved in the BFR training group compared to the high-intensity training group [31].
Another study of 18 men found that a group who performed BFR walk-training 2 times/day, 6 days/week, for 3 weeks had a significant increase in thigh muscle CSA and maximum isometric strength in the BFR walking group compared to the control group [32].
Another study of 12 men compared gastrocnemius oxygenation, capillary lactate, and perceived exertion across 5 different exercise protocols (including a non-BFR high intensity protocol, intermittent and continuous BFR protocols at low intensity, and a low intensity non-BFR protocol). Researchers found that perceived exertion was greatest in the high-intensity protocol while muscle oxygenation was the same between high-intensity and intermittent BFR. Intermittent BFR protocols also had increased blood lactate concentration compared to the continuous BFR protocol [33].
A study of 24 older women undergoing 16 weeks of resistance exercise training targeting elbow flexor muscles found that the group undergoing low-intensity training with BFR had greater percent increase in CSA and isokinetic strength than those who only underwent low-intensity exercise without BFR, and had similar results to a high-intensity group [34]. These results show that BFR can be an effective alternative to high-intensity training that produces similar training adaptations.
Another study of 24 men compared expression of genes involved in hypoxia and angiogenesis when performing low-intensity, low-intensity + BFR, and high-intensity RE. Scientists found that after performing 8 weeks of bilateral knee extension exercise, the low-intensity BFR group had the greatest activation of HIF-1α and HIF-1β, VEGF, and nNOS and iNOS genes. These genes are associated with hypoxia and angiogenesis, indicating BFR training is beneficial for improving oxygen distribution, muscle growth, and mitochondrial efficiency [35].
2.4. Sleep
The scientific community and health and wellness communities agree that sleep is essential for overall health. The National Sleep Foundation has set guidelines recommending 7-9 hours of sleep per night for adults [36]. Studies have shown that those getting less than 7 hours of sleep each night were at significantly higher rates of all-cause mortality, and especially mortality related to cancer and cardiovascular issues [37]. Sufficient sleep is also critical for waking cognition and emotional regulation [38]. Evidence is clear that sleep is essential to overall health and quality of life. Fitness communities also frequently cite the importance of sufficient sleep for muscle growth and endurance adaptations [39,40]. However, the mechanisms for how sleep benefits exercise adaptations are frequently misunderstood or unknown.
A study of the effect of sleep deprivation on mice found that after 21 days of chronic sleep deprivation, PGC1-α expression was reduced by a statistically significant amount (Figure 1) [41]. Another study tested gene changes in an inducible skeletal muscle-specific Bmal1 knockout mouse model, which has a disrupted circadian rhythm. Results showed that circadian metabolic genes involved in substrate utilization and storage were differentially expressed in the Bmal1 knockout mice compared to control mice. There was also a significant shift from fast to slow type muscle fibers with lower ability to quickly produce energy in the Bmal1 knockout model [42]. Therefore, a healthy circadian rhythm, and thus sleep, are critical for maintaining muscle fiber type and mitochondrial oxidative levels.
Another study on the importance of sleep was done on a human cohort. Participants underwent a “muscle damage” protocol, which included 240 eccentric contractions on the knee extensor muscle. Then, participants were either subjected to sleep deprivation or allowed full nights’ sleep. Results showed that the group that experienced total sleep deprivation had significantly elevated IL-6, IGF-1, and cortisol compared to a group that received three nights of regular sleep [43]. Cortisol and IL-6 can both contribute to muscle wasting to meet energy needs under stressful conditions. Another study in humans compared uninjured groups subjected to either total sleep deprivation or normal sleep. These results showed that the sleep deprived group had 18% reduced protein synthesis, 21% increased plasma cortisol, and 24% decreased plasma testosterone. Although no difference was found in markers of protein degradation, these results still show how sleep is necessary for long term muscle growth [44].
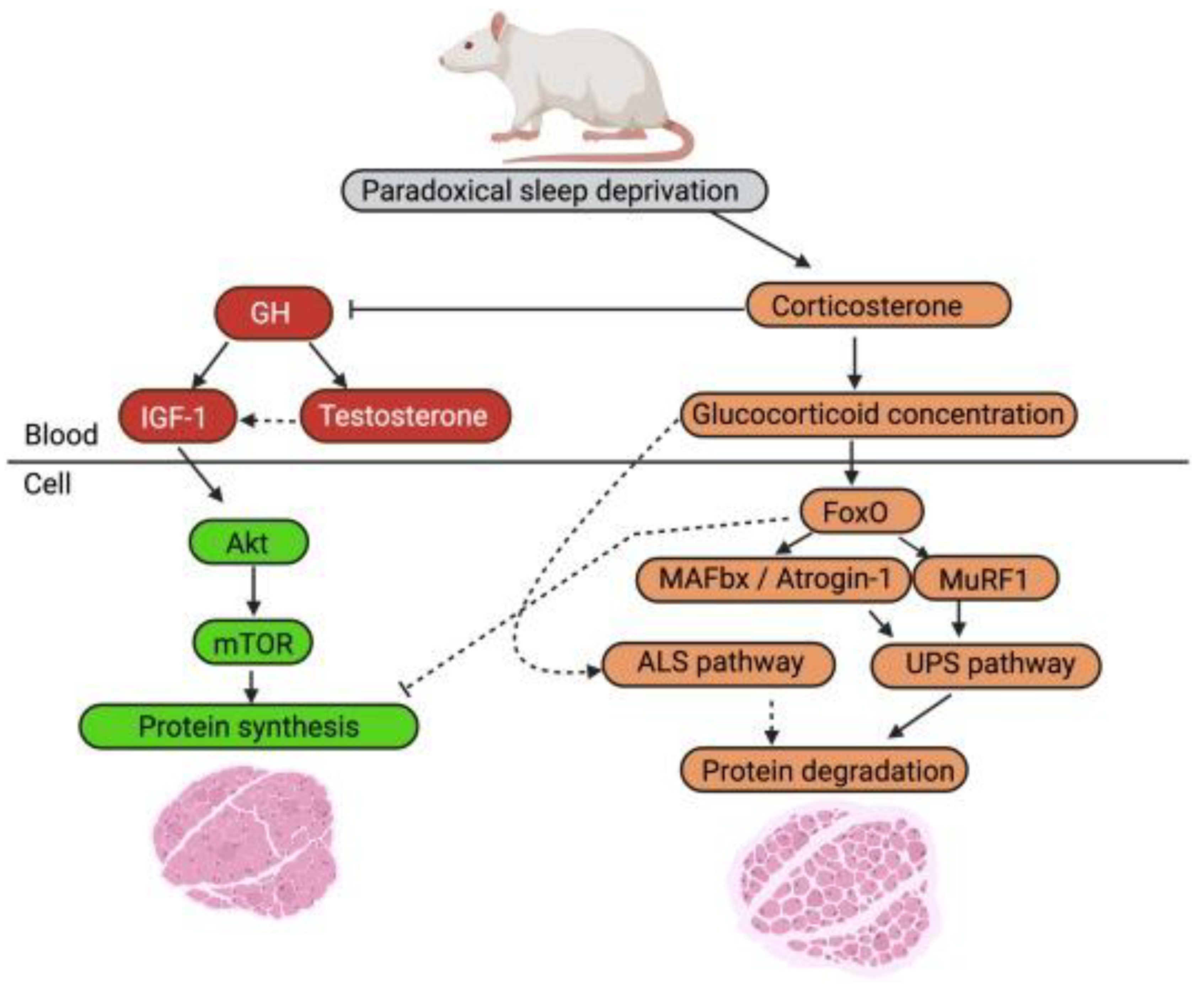
A review of the effects of sleep deprivation on muscle atrophy in rat models found that restricted sleep was linked to muscle atrophy, reduced muscle weight, reduced CSA, and decreased body mass [6]. A proposed mechanism responsible for this sleep-deprivation induced muscle atrophy includes an increase in the corticosterone pathway and a reduction in the testosterone pathway, leading to increased muscle atrophy and decreased protein synthesis (Figure 2) [6]. This model explains the molecular signalling connection between sleep deprivation and loss of muscle.
Another potential effect of sleep deprivation is neuronal misfiring in the peripheral nervous system. One study looked at the effects of rapid eye movement (REM) sleep deprivation in rats [45]. REM sleep is a stage of the sleep cycle where although the brain is active, it is important for brain health and recovery. Results showed an increased amount of mitochondrial DNA in the hippocampal region of the brain, as well as increased manic-like behaviors [45]. This disruption to the brain’s environment could potentially cause abnormal signaling to be sent to other parts of the body. It is possible that abnormal signaling from the central nervous system caused by sleep deprivation could influence peripheral nerve signaling, and thus lead to unintentional, repeated activation of skeletal muscle outside of exercise. This has the potential to increase oxidative stress and increase protein degradation, preventing muscle recovery and adaptations. Currently, there is very little literature related to improper motor neuron signaling and muscle damage. Future research investigating a causality between these would benefit our understanding of how motor neuron signaling affects muscle damage and long term exercise adaptations.
3. Discussion
Healthy skeletal muscle tremendously improves health outcomes and quality of life. Resistance and endurance exercise is a critical factor for developing healthy skeletal muscle and performance enhancing recovery techniques can be used to increase the beneficial adaptations that exercise induces. This section summarizes the effects and most useful applications of each performance enhancing method covered in this review.
Based on the negative effects on RE adaptations and slight positive benefits on EE adaptations, CWI is not recommended as a regular part of training for people lifting to increase muscle mass or improve health. It can potentially be used occasionally by athletes to decrease fatigue symptoms like muscle soreness before important competitions but is not recommended for general recovery to promote long term exercise adaptations.
Heat Stress has proven benefits to satellite cell proliferation, hypertrophy, and mitochondrial homeostasis, as well as inhibition of autophagy and protein denaturation. Proven long term physiological benefits on exercise performance also support use of heat stress as a viable method to supplement both resistance and endurance training. Post-exercise sauna or hot tub use is recommended for endurance athletes and individuals looking to improve hypertrophy.
The limited benefits of BFR to hypertrophy, angiogenesis, and mitochondrial biogenesis markers compared to traditional high-intensity training, as well as the practical difficulty of implementing BFR into training make it a less desirable method for improving muscle health and fitness. BFR will be primarily useful for individuals unable to do traditional high-intensity training. This includes elderly individuals with greater injury risk, individuals recovering from serious injuries, and athletes who want to avoid muscle damage before competitions.
Beyond its non-exercise related necessity, sleep is a critical component of recovery and adaptation. Its influence on protein synthesis and degradation pathways makes sleep optimization an important factor for resistance and endurance adaptations and recovery. Ensuring sufficient sleep each night is recommended for all individuals to improve overall health, hypertrophy, mitochondrial function, and exercise performance.
4. Conclusions
Muscle is critically important for the health of all individuals. Performance enhancers are beneficial to all but especially important for optimizing health and performance of athletes, individuals with limited training time, and clinical and elderly populations. This paper looked into these four performance enhancers: CWI, Heat Stress, BFR, and sleep. Heat Stress and sleep are the most promising for both increasing hypertrophy and mitochondrial biogenesis. BFR is primarily recommended for individuals with limited ability to train with high intensity. CWI is not recommended as a regular part of training. More research on the long-term effects of CWI on both resistance and endurance performance tests should be done to verify long term physiological and molecular changes. Further research into the effects of sleep deprivation on molecular pathways linked to exercise adaptations should also be conducted to confirm how sleep interacts with these processes.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CWI | Cold water immersion |
| BFR | Blood flow restriction |
| EE | Endurance exercise |
| RE | Resistance exercise |
| ATP | Adenosine Triphosphate |
| ETC | Electron transport chain |
| mTOR | Mechanistic Target of Rapamycin |
| P70S6K | p70 ribosomal S6 kinase |
| MAFbx | Muscle Atrophy F-box |
| MuRF1 | Muscle RING Finger 1 |
| TRIM63 | Tripartite Motif Containing 63 |
| MSTN | Myostatin |
| PGC1-α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| p38 | p38 mitogen-activated protein kinase |
| AMPK | AMP-activated protein kinase |
| HSI | Heat stress intervention |
| HSP | Heat shock protein |
| LOP | Limb occlusion pressure |
| CSA | Cross sectional area |
References
- Karagounis, L.G.; Hawley, J.A. Skeletal muscle: increasing the size of the locomotor cell. The International Journal of Biochemistry & Cell Biology 2010, 42(9), 1376–1379. [Google Scholar] [CrossRef]
- Wr, F.J.O. Skeletal Muscle: A Brief Review of Structure and Function. Calcified tissue international. 1 March 2015. Available online: https://pubmed.ncbi.nlm.nih.gov/25294644/.
- Pedersen, B.K. Physical activity and muscle–brain crosstalk. Nature Reviews Endocrinology 2019, 15(7), 383–392. [Google Scholar] [CrossRef]
- Kruk, J. Physical activity in the prevention of the most frequent chronic diseases: an analysis of the recent evidence. Asian Pacific journal of cancer prevention: APJCP 2007, 8(3), 325–338. Available online: https://pubmed.ncbi.nlm.nih.gov/18159963/.
- Wolfe, R.R. The underappreciated role of muscle in health and disease. The American Journal of Clinical Nutrition 2006, 84(3), 475–482. [Google Scholar] [CrossRef]
- Morrison, M.; Halson, S.L.; Weakley, J.; Hawley, J.A. Sleep, circadian biology and skeletal muscle interactions: Implications for metabolic health. Sleep Medicine Reviews 2022, 66, 101700. [Google Scholar] [CrossRef]
- Booth, F.W.; Ruegsegger, G.N.; Toedebusch, R.G.; Yan, Z. Endurance Exercise and the Regulation of Skeletal Muscle Metabolism. Progress in Molecular Biology and Translational Science 2015, 135, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Bandy, W.D.; Lovelace-Chandler, V.; McKitrick-Bandy, B. Adaptation of Skeletal Muscle to Resistance Training. Journal of Orthopaedic & Sports Physical Therapy 1990, 12(6), 248–255. [Google Scholar] [CrossRef]
- Hood, D.A.; Memme, J.M.; Oliveira, A.N.; Triolo, M. Maintenance of Skeletal Muscle Mitochondria in Health, Exercise, and Aging. Annual Review of Physiology 2019, 81(1), 19–41. [Google Scholar] [CrossRef]
- Kang, C.; Ji, L.L. Role of PGC-1α signaling in skeletal muscle health and disease. Annals of the New York Academy of Sciences 2012, 1271(1), 110–117. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.; Nunes, E.A.; Currier, B.S.; Mcleod, J.C.; Thomas, A.C.Q.; Phillips, S.M. An Evidence-based Narrative Review of Mechanisms of Resistance Exercise-induced Human Skeletal Muscle Hypertrophy. Medicine & Science in Sports & Exercise 2022, 54(9). [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS Journal. 2013, 280(17), 4294–4314. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.C.; Fyfe, J.J. Post-exercise Cold Water Immersion Effects on Physiological Adaptations to Resistance Training and the Underlying Mechanisms in Skeletal Muscle: A Narrative Review. Frontiers in Sports and Active Living 2021, 3. [Google Scholar] [CrossRef]
- Outdoor Swimwear And Accessories | Outdoor Swimmer Shop. Outdoor Swimmer Shop. Published 2025. Available online: https://www.outdoorswimmershop.com/ (accessed on 2 December 2025).
- The growing global demand for ice baths and cold plunges. Brassmonkey.co. Published 2025. Available online: https://www.brassmonkey.co/en-us/blogs/journal/the-growing-global-demand-for-ice-baths-and-cold-plunges (accessed on 2 December 2025).
- CI What is Cold Plunging and Why Are Celebrities Obsessed With It? Longevity.Technology - Latest News, Opinions, Analysis and Research. 18 April 2024. Available online: https://longevity.technology/news/what-is-cold-plunging-and-why-are-celebrities-obsessed-with-it/ (accessed on 2 December 2025).
- Roberts, L.A.; Raastad, T.; Markworth, J.F.; et al. Post-exercise cold water immersion attenuates acute anabolic signalling and long-term adaptations in muscle to strength training. The Journal of Physiology 2015, 593(18), 4285–4301. [Google Scholar] [CrossRef]
- Ihsan, M.; Markworth, J.F.; Watson, G.; et al. Regular postexercise cooling enhances mitochondrial biogenesis through AMPK and p38 MAPK in human skeletal muscle. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2015, 309(3), R286–R294. [Google Scholar] [CrossRef] [PubMed]
- Ihsan, M.; Watson, G.; Choo, H.C.; et al. Postexercise Muscle Cooling Enhances Gene Expression of PGC-1α. Medicine & Science in Sports & Exercise 2014, 46(10), 1900–1907. [Google Scholar] [CrossRef] [PubMed]
- Chung, N.; Park, J.; Lim, K. The effects of exercise and cold exposure on mitochondrial biogenesis in skeletal muscle and white adipose tissue. Journal of Exercise Nutrition & Biochemistry 2017, 21(2), 39–47. [Google Scholar] [CrossRef]
- Moore, E.; Fuller, J.T.; Buckley, J.D.; et al. Impact of Cold-Water Immersion Compared with Passive Recovery Following a Single Bout of Strenuous Exercise on Athletic Performance in Physically Active Participants: A Systematic Review with Meta-analysis and Meta-regression. Sports Medicine 2022, 52(7). [Google Scholar] [CrossRef]
- Normand-Gravier, T.; Solsona, R.; Dablainville, V.; et al. Effects of thermal interventions on skeletal muscle adaptations and regeneration: perspectives on epigenetics: a narrative review. European Journal of Applied Physiology 2024, 125. [Google Scholar] [CrossRef]
- Bromley, F. Sauna culture around the world: seven traditions, one ritual. olivemagazine. 22 May 2025. Available online: https://www.olivemagazine.com/travel/sauna-culture-around-the-world-seven-traditions-one-ritual/ (accessed on 2 December 2025).
- Kirby, N.V.; Lucas, S.J.E.; Armstrong, O.J.; Weaver, S.R.; Lucas, R.A.I. Intermittent post-exercise sauna bathing improves markers of exercise capacity in hot and temperate conditions in trained middle-distance runners. European Journal of Applied Physiology 2020, 121(2), 621–635. [Google Scholar] [CrossRef]
- Damas, F.; Libardi, C.A.; Ugrinowitsch, C. The development of skeletal muscle hypertrophy through resistance training: the role of muscle damage and muscle protein synthesis. European Journal of Applied Physiology 2017, 118(3), 485–500. [Google Scholar] [CrossRef]
- Leite, C.D.F.; Zovico, P.V.C.; Rica, R.L.; et al. Exercise-Induced Muscle Damage after a High-Intensity Interval Exercise Session: Systematic Review. International Journal of Environmental Research and Public Health 2023, 20(22), 7082–7082. [Google Scholar] [CrossRef]
- Blood Flow Restriction. Owens Recovery Science. Available online: https://owensrecoveryscience.com/blood-flow-restriction/.
- Freitas, E.D.S.; Karabulut, M.; Bemben, M.G. The Evolution of Blood Flow Restricted Exercise. Frontiers in Physiology 2021, 12, 747759. [Google Scholar] [CrossRef]
- Kacin, A.; Drobnič, M.; Marš, T.; et al. Functional and molecular adaptations of quadriceps and hamstring muscles to blood flow restricted training in patients with ACL rupture. Scandinavian Journal of Medicine & Science in Sports 2021, 31(8), 1636–1646. [Google Scholar] [CrossRef]
- Kakehi, S.; Tamura, Y.; Kubota, A.; et al. Effects of blood flow restriction on muscle size and gene expression in muscle during immobilization: A pilot study. Physiological Reports 2020, 8(14). [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.; Rosenblatt, B.; Haddad, F.; et al. Comparing the Effectiveness of Blood Flow Restriction and Traditional Heavy Load Resistance Training in the Post-Surgery Rehabilitation of Anterior Cruciate Ligament Reconstruction Patients: A UK National Health Service Randomised Controlled Trial. Sports medicine (Auckland, NZ) 2019, 49(11), 1787–1805. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kearns, C.F.; Sato, Y. Muscle size and strength are increased following walk training with restricted venous blood flow from the leg muscle, Kaatsu-walk training. Journal of Applied Physiology 2006, 100(5), 1460–1466. [Google Scholar] [CrossRef]
- Corvino, R.B.; Rossiter, H.B.; Loch, T.; Martins, J.C.; Caputo, F. Physiological responses to interval endurance exercise at different levels of blood flow restriction. European Journal of Applied Physiology 2017, 117(1), 39–52. [Google Scholar] [CrossRef]
- Takarada, Y.; Takazawa, H.; Sato, Y.; Takebayashi, S.; Tanaka, Y.; Ishii, N. Effects of resistance exercise combined with moderate vascular occlusion on muscular function in humans. Journal of Applied Physiology 2000, 88(6), 2097–2106. [Google Scholar] [CrossRef]
- Volga Fernandes, R.; Tricoli, V.; Garcia Soares, A.; Haruka Miyabara, E.; Saldanha Aoki, M.; Laurentino, G. Low-Load Resistance Exercise with Blood Flow Restriction Increases Hypoxia-Induced Angiogenic Genes Expression. Journal of Human Kinetics 2022, 84, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Hirshkowitz, M.; Whiton, K.; Albert, S.M.; et al. National Sleep Foundation’s updated sleep duration recommendations: final report. Sleep Health 2015, 1(4), 233–243. [Google Scholar] [CrossRef]
- Gallicchio, L.; Kalesan, B. Sleep duration and mortality: a systematic review and meta-analysis. Journal of Sleep Research 2009, 18(2), 148–158. [Google Scholar] [CrossRef] [PubMed]
- Worley, S.L. The Extraordinary Importance of Sleep: The Detrimental Effects of Inadequate Sleep on Health and Public Safety Drive an Explosion of Sleep Research. Pharmacy and Therapeutics 2018, 43(12), 758–763. Available online: https://pmc.ncbi.nlm.nih.gov/articles/PMC6281147/. [PubMed]
- Washtenaw Community College Fitness Center Staff. How Quality Sleep Impacts Your Fitness Journey. The Health & Fitness Center at Washtenaw Community College. Published March 15, 2024. Available online: https://www.wccfitness.org/blog/how-quality-sleep-impacts-your-fitness-journey/?scrlybrkr=46b27ade (accessed on 2 December 2025).
- Thrive Community Fitness. The Importance of Sleep In an Active Lifestyle. Thrivecf.com. Published 2020. Available online: https://www.thrivecf.com/blog.aspx?post=5408&title=The-Importance-of-Sleep-In-an-Active-Lifestyle (accessed on 2 December 2025).
- Zhang, Y.M.; Wang, Y.T.; Wei, R.M.; et al. Mitochondrial antioxidant elamipretide improves learning and memory impairment induced by chronic sleep deprivation in mice. Brain and behavior 2024, 14(5), e3508. [Google Scholar] [CrossRef] [PubMed]
- Hodge, B.A.; Wen, Y.; Riley, L.A.; et al. The endogenous molecular clock orchestrates the temporal separation of substrate metabolism in skeletal muscle. Skeletal Muscle 2015, 5(1). [Google Scholar] [CrossRef]
- Dáttilo, M.; Antunes, H.K.M.; Galbes, N.M.N.; et al. Effects of Sleep Deprivation on Acute Skeletal Muscle Recovery after Exercise. Medicine & Science in Sports & Exercise 2020, 52(2), 507–514. [Google Scholar] [CrossRef]
- Lamon, S.; Morabito, A.; Arentson-Lantz, E.; et al. The effect of acute sleep deprivation on skeletal muscle protein synthesis and the hormonal environment. Physiological Reports 2021, 9(1). [Google Scholar] [CrossRef]
- Kim, S.A.; Kim, S.; Park, H.J. REM-Sleep Deprivation Induces Mitochondrial Biogenesis in the Rat Hippocampus. In Vivo (Athens, Greece) 2022, 36(4), 1726–1733. [Google Scholar] [CrossRef]
Figure 1.
Effects of sleep deprivation on peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). Western blotting and reverse transcription-quantitative polymerase chain reaction assays were used to determine the effects of sleep deprivation on PGC-1α in c57L/6J mice. There is a significant difference between protein levels between Control and SD in saline solution (p < .05). Graph and data adapted from Yue-Ming et al., Brain and Behavior. 2024.
Figure 1.
Effects of sleep deprivation on peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). Western blotting and reverse transcription-quantitative polymerase chain reaction assays were used to determine the effects of sleep deprivation on PGC-1α in c57L/6J mice. There is a significant difference between protein levels between Control and SD in saline solution (p < .05). Graph and data adapted from Yue-Ming et al., Brain and Behavior. 2024.
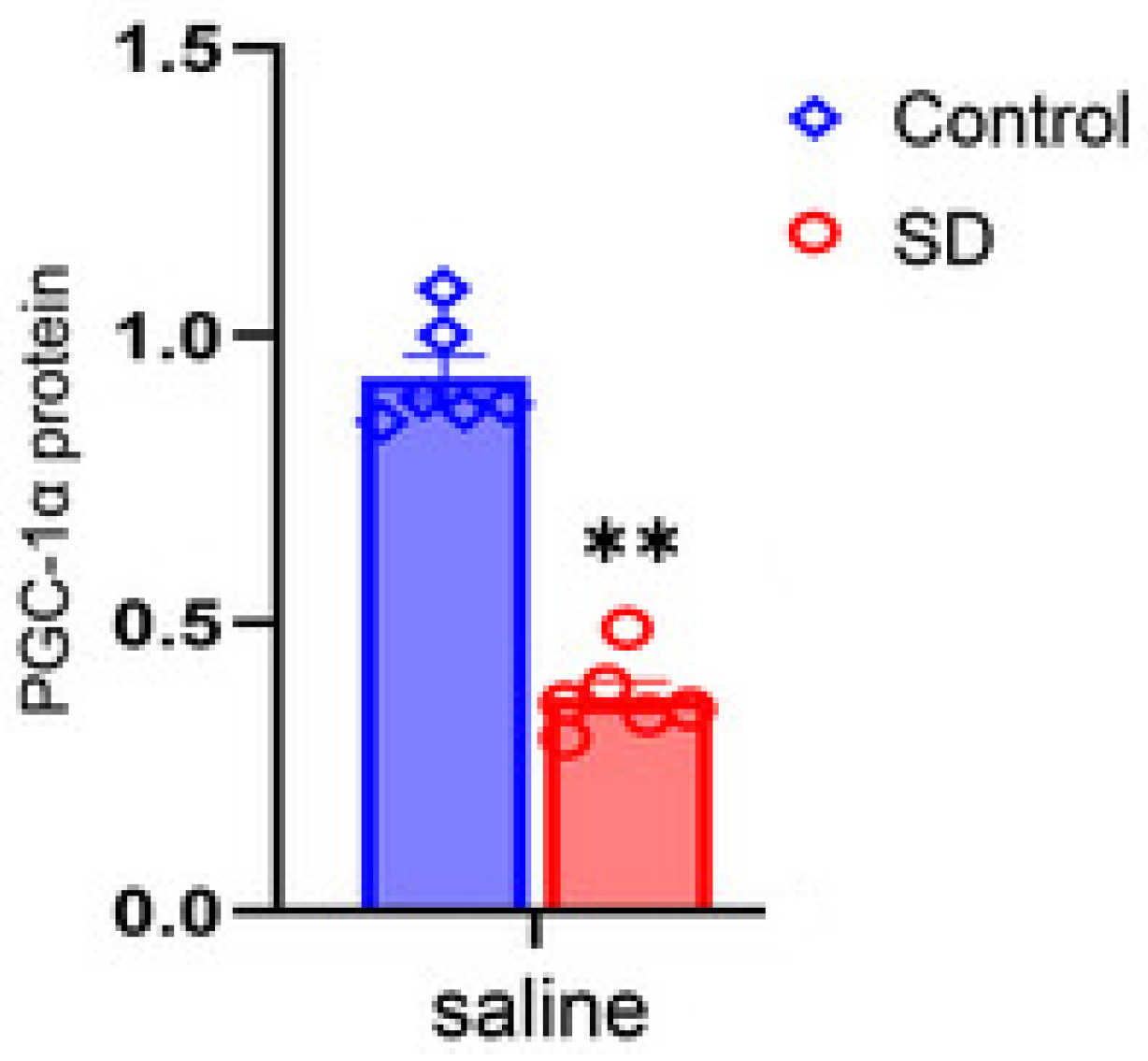
Figure 2.
Sleep deprivation of rodent’s effect on cellular pathways related to rates of skeletal muscle protein turnover. (Left side) Elevation of corticosterone from sleep deprivation may inhibit the activation of mTOR pathway proteins, decreasing rate of protein synthesis. (Right side) Increased corticosterone levels after sleep deprivation can cause increased activation of FoxO, MAFbx, and MuRF1 resulting in increased protein degradation. Graph and data adapted from Morison et al., Sleep Medicine Reviews. 2022.
Figure 2.
Sleep deprivation of rodent’s effect on cellular pathways related to rates of skeletal muscle protein turnover. (Left side) Elevation of corticosterone from sleep deprivation may inhibit the activation of mTOR pathway proteins, decreasing rate of protein synthesis. (Right side) Increased corticosterone levels after sleep deprivation can cause increased activation of FoxO, MAFbx, and MuRF1 resulting in increased protein degradation. Graph and data adapted from Morison et al., Sleep Medicine Reviews. 2022.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



