
Submitted:
03 January 2026
Posted:
05 January 2026
You are already at the latest version
Abstract
Recent findings suggest that the gut microbiome significantly influences cancer outcomes,
including responses to immune checkpoint inhibitor (ICI) treatments. Although
early research focused on gut bacteria, it is now understood that the
microbiome includes a bacteriome, virome, and mycobiome, all of which can
modulate host immunity. Some commensal bacteria enhance anti-tumor immune
responses and improve ICI efficacy, as demonstrated in both mice and patients.
Fecal microbiota transplants (FMT) from patients responding to ICI have
successfully reversed resistance in certain non-responders. In addition to
bacteria, gut fungi and viruses are gaining attention as further factors
influencing ICI effiectiveness and toxicity. Recent multi-omics studies across
cancer cohorts show that fungal and viral populations in the gut vary between
ICI responders and non-responders. Commensal fungi may shape anti-cancer
immunity by inducing inflammatory or tolerogenic pathways, while viral
components can stimulate innate immune sensors that promote tumor surveillance.
On the other hand, gut dysbiosis marked by expansion of pathobionts (including
opportunistic fungi) and reduction of beneficial microbes is linked to serious
immune-related adverse events (irAEs) such as ICI-induced colitis. This review
discusses the multi-kingdom gut microbiome – bacteria, fungi, and viruses – and
their interactions with the immune system in cancer therapy. We emphasize known
mechanisms linking these microbes to anti-tumor immunity, overview human studies
associating gut microbiome profiles with ICI outcomes and explore strategies to
modulate the microbiome to enhance ICI efficacy while reducing toxicity.
Understanding and utilizing the gut mycobiome and virome in conjunction with
the bacteriome could pave the way for new biomarkers and therapeutic adjuvants
in cancer immunotherapy.
Keywords:
mycobiome
; virome
; gut microbiome
; immune checkpoint inhibitor
; fecal microbiota transplant
; cancer
1. Introduction
Cancer continues to be a significant global health challenge. The most recent GLOBOCAN report estimates approximately 20 million new cancer cases and 9.7 million deaths in 2022, with around 53.5 million people living five years post-diagnosis; roughly 1 in 5 people will face a cancer diagnosis in their lifetime [1]. These numbers highlight the magnitude of the challenge and the necessity for improved, more tolerable therapies. In the United States, around 2,041,910 new cancer cases and 618,120 deaths are expected in 2025(2). Although mortality rates have consistently declined through 2022—preventing nearly 4.5 million deaths since 1991 through smoking reduction, earlier diagnosis, and improved treatment—substantial disparities remain [2].
Over the past decade, immuno-oncology has redefined clinical practice by utilizing the host's immune system instead of directly targeting or starving tumor cells [3,4,5,6]. Therapeutic classes currently include: (i) immune-checkpoint blockade against PD-1/PD-L1 and CTLA-4, with emerging pathways like LAG-3 becoming standard practice; (ii) adoptive cell therapies — CAR-T for hematologic malignancies and TILs/CAR-NK moving into solid tumors; (iii) therapeutic vaccination, including custom neoantigen and dendritic-cell approaches; (iv) oncolytic virotherapy (T-VEC); (v) cytokine/immune-modulating regimens [3,7,8,9,10]. Mechanistically, these approaches are complementary — disabling inhibitory circuits, providing tumor-reactive effector lymphocytes, priming de novo responses and readjusting the immune system environment [9]. Still, lasting benefits are heterogeneous and toxicity can be substantial, indicating tumor-intrinsic and microenvironmental resistance mechanisms. Consequently, biomarker-driven combinations and improvements in delivery and formulation represent active frontiers [5,11,12].
Immune checkpoint inhibitors (ICIs) – monoclonal antibodies that restore anti-tumor T cell activity by blocking inhibitory receptors like PD-1, PD-L1, CTLA-4 – have revolutionized the management of many malignancies [6]. However, only a subset of patients attains lasting remissions with ICIs, and others face considerable immune-related toxicities(6, 13, 14). This variability has led scientists to explore factors beyond tumor genetics that influence ICI outcomes, with the gut microbiome emerging as a key player [15,16]. The human gastrointestinal tract contains a vast and diverse microbial ecosystem, collectively referred to as the gut microbiota [17]. This intricate community of microorganisms has developed alongside the host and is essential for digestion, metabolism, and particularly in shaping the development and function of the immune system [17,18,19]. In recent years, cancer researchers have grown more aware that gut microbes can influence both cancer progression and the efficacy of cancer therapies [15,16].
Early studies in 2015 provided proof-of-concept that gut bacteria can influence cancer immunotherapy [20,21]. In mouse models, the depletion of commensals through antibiotics was shown to reduce the anti-tumor effectiveness of ICIs [15,20]. On the other hand, transferring gut microbiota from one animal to another could convey the level of ICI responsiveness [15,21]. Around the same time, divergent findings in mice from different facilities highlighted that variations in gut flora might account for inconsistencies in ICI treatment outcomes [15,20,21]. These discoveries ignited a wave of research on particular bacterial strains that enhance anti-cancer immunity. Certainly, both preclinical and clinical studies soon confirmed that patients who respond to ICIs possess a distinct gut microbial composition compared to non-responders [15,22,23,24]. Specific bacteria, like Bifidobacterium, Akkermansia, Faecalibacterium, Bacteroides spp, have consistently been linked to positive ICI responses across multiple cancers [15,23,24,25]. Notably, fecal microbiota transplants from ICI-responsive donors demonstrated the capacity to overcome resistance in some refractory patients [26,27,28] indicating a causal relationship between the gut microbiota and ICI effectiveness [15]. Additional studies showed that baseline microbiota characteristics could also anticipate susceptibility to ICI-induced toxicities such as colitis [29,30] suggesting the gut environment affects both the effectiveness of treatment and safety.
Initially centered on gut bacteria, the gut ecosystem also consists of commensal fungi – the gut mycobiome, viruses – gut virome, archaea and protists [17]. These additional microorganisms, while less abundant than bacteria in terms of biomass, can produce immunologically active molecules and interact with both the host and bacterial communities significantly. The roles of the virome and mycobiome in cancer and immunotherapy have been largely overlooked, thus remain unclear. Nonetheless, advanced metagenomic and multi-kingdom studies are beginning to reveal how fungi and viruses can also influence anti-tumor immunity and ICI outcomes [31,32,33]. Since ICIs activate the immune system broadly, it is likely that the entire gut microbiome, across multiple kingdoms, affects the balance of immunity and tolerance throughout treatment.
2. Literature search strategy
For this narrative review, we have gathered information from the specialized literature published in English over the past decade, with a focus on the interaction between the gut microbiome and cancer immunotherapy, particularly emphasizing the gut bacteriome, gut virome, and gut fungi. Databases such as PubMed, Scopus, and Web of Science were systematically queried using combinations of the following keywords: “gut microbiome”, “gut microbiota”, “gut bacteriome”, “gut virome”, “gut mycobiome”, “cancer immunotherapy”, and “immune checkpoint inhibitors”. The types of articles included are reviews (narrative and systematic), preclinical and clinical studies, as well as meta-analyses, all of which were selected based on their title and abstract, particularly those examining the relationship between gut microbiota components and cancer immunotherapy. Exclusion criteria included non-peer-reviewed sources, conference abstracts, and studies not directly related to immunotherapy.
3. Immune Checkpoint Blockade: Mechanism and Clinical Relevance
The immune surveillance hypothesis suggested that our immune system patrols for and eliminates new cancer cells, preventing tumor development [34]. Both the innate and adaptive branches of the immune system collaborate to identify and remove emerging tumors [35]. Innate immune cells like natural killer (NK) cells, macrophages, and dendritic cells form an initial barrier of defense by killing abnormal cells directly or by presenting tumor antigens to T cells [36]. Adaptive immune cells, particularly cytotoxic T lymphocytes (CTLs), can recognize tumor-derived antigens and launch a precise attack on cancerous cells [36]. In healthy immune surveillance, these components collaborate to prevent malignancies [37]. Despite this, any cancer that arises in a host has, by definition, successfully managed to evade or suppress the immune response that would typically eliminate it [37]. Tumors achieve this immune evasion via various mechanisms, such as creating an immunosuppressive tumor microenvironment and utilizing regulatory “brakes” of the immune system referred to as immune checkpoints [4].
Immune checkpoints are inhibitory pathways embedded into the immune system that ensure self-tolerance and prevent excessive harm to healthy tissues during immune responses [38]. Cancer cells frequently exploit these checkpoints to turn off anti-tumor T cells, enabling the tumor to grow unchecked [38]. This understanding led to the idea behind developing immune checkpoint inhibitor (ICI) therapies – by blocking these inhibitory checkpoint molecules, we can unleash T cells and restore the immune system’s capacity to attack cancer [39,40,41,42]. In essence, checkpoint blockade therapy seeks to tip the balance in favor of immune activation rather than suppression, thus overcoming a primary escape mechanism used by cancer.
In recent years, cancer therapy has been revolutionized by immunotherapy, which uses the immune system to fight tumors more effectively [4]. Significant advances in the field include chimeric antigen receptor (CAR) T-cell therapy, tumor-infiltrating lymphocyte (TIL) therapy, cancer vaccines, and particularly immune checkpoint inhibitors (ICIs)[3,4]. Among these, ICIs have caused a fundamental change in oncology by producing unmatched and long-lasting responses in several types of cancer. ICIs target regulatory checkpoint pathways – crucial for immune homeostasis – that tumors exploit to suppress anti-tumor immunity. By inhibiting checkpoints like CTLA-4 and PD-1/PD-L1, these treatments can “retrain” the immune system to oppose a stronger fight against cancer as shown in Figure 1.
ICIs have quickly transitioned from being experimental drugs to standard cancer treatments. They are currently approved for a broad range of malignancies, including melanoma, non-small cell lung cancer (NSCLC), renal cell carcinoma, bladder cancer, head and neck cancers, some colorectal cancers with mismatch-repair deficiency, Hodgkin lymphoma, among many others [43]. The clinical impact has been remarkable – for instance, advanced melanoma, once known to be fatal within a year, now has about half of patients surviving at 5 years in the era of combined checkpoint blockade [44]. Certain patients achieve full remissions that are sustained even after stopping treatment, suggesting the potential for cures in conditions once deemed uncurable [35]. Similar advancements have been observed in other cancers – for metastatic NSCLC, the addition of PD-1/PD-L1 inhibitors has greatly extended survival, with some patients achieving long-term remission [3]. Therefore, the expected outcomes of ICI therapy, although variable by cancer type, include a significant prolongation of survival for responders and a subset of long-term survivors that were not seen with previous therapies.
Nonetheless, despite these advancements, immune activation through CTLA-4 or PD-1 blockade by itself is not sufficient to manage tumor progression in all patients [4]. A significant number of patients either fail to respond to ICIs or show an initial response that is followed by a relapse [35]. Even in tumors with high immunogenicity such as melanoma, where long-term response rates approach 60%, roughly 25% of patients exhibit primary resistance – characterized by disease progression within ~6 months of starting ICI – and many others later develop acquired resistance despite of an initial response [44,45,46].Tumor-intrinsic factors, tumor-extrinsic factors, and host factors (germline genetics, microbiome) all converge to influence outcomes. Significant efforts are made to identify reliable predictive biomarkers and to develop innovative approaches – whether new checkpoints, combination regimens, or personalized adjustments – to improve efficacy [43,47,48]. Patient-specific factors, such as genetic makeup and gut microbiota composition, have recently been linked to responsiveness to ICI therapy [31,49,50,51]. Research indicates that the gut microbiome can influence systemic immunity – certain commensal bacteria profiles in patients correlate with improved anti-PD-1 therapy outcomes, whereas antibiotic disruption of the microbiome is linked to poorer outcomes [43]. In mouse models, fecal microbiota transplant (FMT) from ICI-responding patients into non-responding mice was able to convey anti-tumor effects, showing that commensal microbes can affect treatment effectiveness [15]. The gut microbiome is a highly modifiable factor, raising the possibility of manipulating microbiota (through diet, probiotics, or FMT) to improve patient responses in the future [40].
Immune checkpoint inhibitors (ICIs) can cause a variety of adverse effects, ranging from mild pharmacologic side effects to unique immune-related adverse events (irAEs) [4,6,52]. Typical general reactions include fatigue, rash, diarrhea, nausea, fever, pruritus, and musculoskeletal pain [39,53,54]. IrAEs can impact virtually any organ: skin and gut are commonly affected, followed by endocrine glands and liver, and less frequently the heart and lungs [55]. The profile of irAEs differs depending on the ICI target. CTLA-4 inhibitors often cause colitis, hypophysitis and dermatologic eruptions [56,57], whereas PD-1/PD-L1 inhibitors are more likely to lead to pneumonitis, thyroid dysfunction, arthralgias, and vitiligo [53,57]. Combination immunotherapy (CTLA-4 + PD-1 blockade) often increases toxicity for only a modest gain in efficacy [58]: in metastatic melanoma trials, adding a CTLA-4 antibody to PD-1 therapy roughly doubled the incidence of high-grade (≥3) irAEs compared with PD-1 alone [39,59,60]. Notably, thyroid immune-related events become more frequent with combined therapy [53,61,62]. These findings highlight that unleashing multiple immune “brakes” can produce synergistic anti-tumor effects at the cost of higher immune toxicity [4,39]. This baseline understanding of ICI toxicity provides a foundation for understanding how the host microbiome plays a role in further modulating these outcomes.
4. Microbiota: A General Overview
From birth, humans are colonized by a large community of microorganisms – the microbiota – especially on mucosal areas such as the gastrointestinal tract [17]. The gut microbiota is extremely diverse, containing thousands of species and multiple kingdoms of microbes that coexist in a symbiotic “superorganism” alongside the host [17,63]. Bacteria are predominant by cell count, with four major phyla (Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria) representing approximately 98% of the microbial residents of the gut [64]. These bacteria are crucial for fermenting dietary fiber to produce short-chain fatty acids (SCFAs) that nourish colon cells, for synthesizing vitamins and aiding in the metabolism of bile acids and other compounds, and for colonization resistance against pathogens by outcompeting or inhibiting them [65]. Beside bacteria, the gut harbors smaller populations of fungi – the mycobiome, a virome of viruses, mainly bacteriophages that infect bacteria [64], archaea and protists, all being part of the gut ecosystem [65]. Although less abundant, these non-bacterial members can influence community dynamics – fungi can bloom when antibiotics wipe out competing bacteria [66], leading to issues like Candida overgrowth [67], while bacteriophages can modulate bacterial populations [68]. Overall, the gut microbiome functions as a complex, interdependent community, and sustaining its balanced state of eubiosis is crucial for health, whereas disruptions to its composition (dysbiosis) can negatively impact host physiology( [69].
One of the most vital functions of the gut microbiota is its interaction with the immune system of the host [64]. The intestinal barrier – made of tightly joined epithelial cells, mucus secreted by goblet cells, antimicrobial peptides from Paneth cells and IgA from local plasma cells – serves as the primary line of defense that restricts microbes to the gut lumen [63,70]. It’s become clear that the presence of commensal microbes is essential for proper immune development: germ-free animals have underdeveloped gut lymphoid tissue, low baseline IgA levels, and fewer effector and memory T cells [71]. The microbiota provides continuous stimuli (such as microbial DNA, lipopolysaccharides, and metabolites) that calibrate the host immune set-point – effectively adjusting the balance between an active immune response and immune tolerance [72].
Considering the profound immunological impact, it is not surprising dysbiosis has been associated with multiple diseases [73,74]. External factors such as antibiotics, poor diet or infections can disturb the delicate microbial balance [75]. The loss of beneficial commensals or blooms of opportunistic organisms can disrupt the gut’s immune tolerance and barrier integrity [69]. Antibiotic-induced dysbiosis frequently increases gut permeability and allows microbial metabolites to enter the bloodstream, triggering chronic low-grade inflammation [64]. This type of systemic inflammation has been linked to metabolic disorders and a pro-tumorigenic environment [73]. Persistent dysbiosis is a known factor in conditions such as inflammatory bowel disease and colorectal cancer [73]. Patients diagnosed with colorectal cancer (CRC) have lower levels of butyrate-producing gut bacteria, indicating that diminished microbial production of anti-inflammatory metabolites such as butyrate may impair immune surveillance and promote tumor development [64,76]. Certain gut bacteria present in dysbiosis have been implicated in cancer risk: Helicobacter pylori infection can be a cause of gastric cancer [77], some strains of Escherichia coli produce a toxin causing DNA damage in colonic cells [78,79] and Fusobacterium nucleatum, which thrive in oral and gut dysbiosis, can be found in colorectal cancer tissue [80,81]. This insights have led to the microbiome being recognized as a factor in cancer’s “enabling characteristics” as a determinant of chronic inflammation, immune evasion, and even therapy resistance [78]. In summary, who inhabits our gut can shape our baseline cancer risk and tumor behavior.
Based on this, naturally the gut microbiota has emerged as an important player in determining how patients respond to cancer therapies, particularly the latest class of immunotherapies [82]. Clinicians noted more than a century ago that severe infections could sometimes result to tumor regressions – an observation utilized by William Coley in the 1890s [83]. Coley purposefully injected cancer patients with combinations of live or inactivated bacteria and at times observed remarkable tumor shrinkage accompanying the provoked fevers and immune activation. “Coley’s toxin” is recognized as the initial attempt of cancer immunotherapy and highlights the concept that microbial stimuli can activate the immune system better fight cancer [39,83,84]. In the contemporary age, current immune checkpoint inhibitors (ICIs) – drugs like anti-CTLA-4 or anti-PD-1/PD-L1 antibodies – have revolutionized oncology by achieving lasting remissions in a subset of patients. Nonetheless, a significant number of patients do not respond to ICIs, fact that led to intense research into finding out why [35]. In 2015, two landmark studies first pointed to the gut microbiome as possible key factor. Vetizou et al. (2015) showed that the anti-CTLA-4 antibody, ipilimumab, could only effectively manage tumors in mice that had certain Bacteroides species in their gut [20]. Germ-free or antibiotic-treated mice didn’t respond until Bacteroides fragilis was introduced [20]. However, Sivan et al. (2015) observed that the efficacy of anti-PD-L1 therapy in mice was increased by the presence of Bifidobacterium in the gut – mice from a colony with abundant Bifidobacterium had better tumor control compared to those lacking these bacteria and administering Bifidobacterium to the latter improved their outcomes [21]. Decades after Coley’s primitive bacterial adjuvant, we have circled back to understanding the powerful help that microbes can provide in the fight against cancer [83].
Considering that bacteria represent the largest and most studied part of the gut microbiome, it is in this area of research that most microbiome-oncology insights have been obtained. At this point, numerous preclinical and clinical studies have identified specific bacterial species that can boost or suppress anti-tumor immune responses. In this section, we outline key discoveries regarding the relationship between the gut bacteriome and the immune system in the context of cancer therapy, as well as the bacterial taxa associated with better or poorer results from ICIs.
Distinct microbiota in responders vs. non-responders: One of the first clues that gut bacteria influences immunotherapy outcomes came from comparing the stool of cancer patients who respond to ICIs and those who do not [85,86]. These studies frequently report microbial “signatures” linked to response [85]. In melanoma patients undergoing treatment with anti-PD-1 checkpoint inhibitors, responders had significantly greater gut microbial diversity and an abundance of Clostridiales/Firmicutes taxa, while non-responders often had microbiomes dominated by Bacteroidales [25,87,88]. Responders exhibited higher levels of Faecalibacterium prausnitzii and Ruminococcus, along with Akkermansia, whereas non-responders had more Bacteroides, which was linked with immunosuppressive cell populations in the blood [31,72,87]. In patients receiving anti-CTLA-4 treatment, small studies similarly indicate that those experiencing clinical improvements often had a gut microbiome enriched in Firmicutes, whereas those with poorer outcomes displayed an abundance of Bacteroidetes phylum bacteria [80,89,90]. Interestingly, it was found that Bacteroides caccae and Bacteroides uniformis were more prevalent in patients who responded positively to ICIs overall, implying that not all Bacteroides are “bad” – context matters [80,91,92]. However, a common trend observed in several studies is that pathobionts or dysbiotic indicators are often associated with non-response, whereas a diverse, commensal-rich ecosystem correlates with response [29,75,93]. These studies set the stage for exploring causality further.
Many specific bacterial taxa have been identified in various studies, and although we cannot cover them all here, some names appear repeatedly. On the “beneficial” side, many are well astablished commensals known for their good metabolic or immunologic roles: Akkermansia muciniphila [94,95,96,97,98] (a mucus-degrading bacterium that can improve gut barrier integrity and stimulate mucosal DCs), Faecalibacterium prausnitzii [98,99] (an anti-inflammatory butyrate producer), Bifidobacterium species [99,100,101] (fiber-fermenters frequently used as probiotics), and members of the Ruminococcaceae [51,102] and Lachnospiraceae [51,103,104] families are consistently linked to better ICI responses [80]. Another notable genus is Bacteroides – although an excess of Bacteroides overall can indicate dysbiosis, specific species like Bacteroides fragilis and Bacteroides thetaiotaomicron have demonstrated enhancement of anti-CTLA-4 therapy in mice and were associated with beneficial immunity in some patients [20,86,105,106].
Conversely, scientists have identified bacteria that could be detrimental to cancer immunotherapy. These are usually organisms linked to pro-inflammatory dysbiosis or opportunistic infections [29,75,107]. An abundance of Proteobacteria is frequently seen as warning sign in microbiome studies, as it may indicate an unbalanced state [91,102]. In patients treated with ICI, an overgrowth of Proteobacteria or certain species of Enterococcus and Streptococcus has been associated with poorer responses and increased immune-related side effects, such as colitis [72,102,106,108]. Bacteroidales members, despite containing some beneficial species, are often observed at higher levels in non-responders – possibly due to certain Bacteroides and Prevotella species that stimulate expansion of regulatory T cells and myeloid suppressor cells blunting anti-tumor immunity [72,92,108,109]. Bacteroides intestinalis serves as an example of a potential pathogen, recently associated with severe checkpoint inhibitor colitis in melanoma patients [90,110]. Likewise, Fusobacterium nucleatum, known for promoting colorectal cancer growth and evading immune defenses, could hinder immunotherapy if present in the tumor microenvironment [78,81].
Several studies have associated gut microbiota composition with the likelihood of developing ICI-induced colitis, a typical irAE [93,111]. In patients with metastatic melanoma receiving ipilimumab, baseline microbial profiles were strikingly different between those who developed severe colitis and those who did not [30,90]. Notably, beneficial anaerobes in the phylum Bacteroidetes were enriched in patients who did not experience colitis, suggesting a protective effect of these commensals [39,90]. Members of Bacteroidaceae, Rikenellaceae, Barnesiellaceae families were more prevalent at baseline in patients who did not develop ipilimumab colitis [90]. These bacteria are thought to promote mucosal regulatory pathways [39]. Consistent with this, preclinical mouse studies showed that Bacteroides fragilis could protect against CTLA-4–induced colitis and was actually necessary for optimal anti-CTLA-4 tumor efficacy [20,106]. In contrast, patients who lacked these Bacteroidetes and instead had high baseline levels of Gram-positive Firmicutes were more susceptible to colitis. The initial abundance of the Firmicutes phylum (especially Ruminococcaceae family members like Faecalibacterium) correlated with an increased risk of ipilimumab-induced colitis [39,112]. Intriguingly, the same Firmicutes-rich profile was also linked with improved tumor response to treatment [39,112,113]. Faecalibacterium prausnitzii – a butyrate-producing anti-inflammatory bacterium – was found at higher levels in patients who had positive clinical outcomes, although these patients also had higher rates of colitis [30,114]. This indicates a double-edged sword: a microbiome that strongly boosts T-cell activity might inhance ICI efficacy at the expense of lowering the threshold for intestinal autoimmunity.
Beyond single cohorts, larger studies have strengthened the microbiome–toxicity link. A recent prospective study involving 195 patients receiving anti-PD-1 ± anti-CTLA-4 therapy found that baseline dysbiosis characterized by the growth of opportunistic bacteria was associated with subsequent severe irAEs [29]. Patients who later developed grade ≥3 toxicities had a significantly higher relative abundance of a prespecified “pathobiont” group at the start of the treatment 8.2% compared to 4.8% in those without severe irAEs [29]. Moreover, when beginning ICI therapy, these patients showed an acute loss of beneficial Ruminococcaceae in their gut microbiome, a shift not seen in patients who tolerated treatment well [29]. By the time severe colitis or other irAEs manifested, levels of Ruminococcus had drastically decreased in the affected patients [29]. These findings suggest that a resilient, well-balanced microbiome that’s rich in fiber-degrading and SCFA-producing bacteria may protect against immune-mediated toxicity, while a microbiome tilted toward pathobionts primes the host for unchecked inflammation. In practical terms, broad-spectrum antibiotic exposure has been associated with poorer ICI outcomes, including not only reduced efficacy but possibly higher risk of colitis according to some reports [24,91,97,115,116]. The lack of commensals like Akkermansia muciniphila and Faecalibacterium correlates with both poor tumor response and a higher incidence of gastrointestinal irAEs [29,108,111]. This highlights the integral role of the microbiome in adjusting the balance between the host immune system’s aggression and tolerance also summarized in Table 1.
It’s important to understand that “good” and “bad” bacteria often rely on context. The overall impact of the microbiome on cancer immunity results from a complex balance: diversity and community structure likely influence outcomes as much as the presence or lack of any single species [80)].
6. Gut Mycobiome in Immunity and Cancer
6.1. Composition and Roles in Health and Disease
The gut mycobiome represents a small but important portion of the microbiota. Fungal cells are significantly less numerous than bacteria, around of 0.1% or less of gut microbes, yet they include various yeasts and molds such as Candida, Saccharomyces, Malassezia, and others [32,85,126,127,128]. Under healthy circumstances, these fungi are present at low levels and might even provide benefits. Commensal fungi can aid digestion through fermenting dietary fibers and contribute to immune system development [129]. Interestingly, mice raised completely germ-free and fungus-free exhibit more significant immune deficiencies compared to germ-free mice that are colonized with a few fungi, suggesting that initial exposure to fungal antigens helps “train” the immune system [126]. Therefore, a balanced mycobiome likely aids in maintaining immune homeostasis, while disturbances in fungal populations are linked disease. For instance, in inflammatory bowel disease (IBD) patients, the gut mycobiota frequently shifts toward increased levels of Candida and reduced diversity of harmless fungi like Saccharomyces [130] – changes believed to both partake in the cause of and exacerbate the chronic inflammation [126,131,132]. Another study found that Candida tropicalis was significantly increased in Crohn’s disease and formed abnormal biofilms alongside pathogenic bacteria on the gut lining [133], underlining how fungal and bacterial dysbiosis often go hand-in-hand [66].
Fungi have distinct cell wall components – β-glucans, mannans, chitin – that represent potent pathogen-associated molecular patterns (PAMPs) stimulating host immune response through pattern recognition receptors (PRRs) [134]. One key receptor is Dectin-1 (CLEC7A), a C-type lectin found on dendritic cells and macrophages that binds β-1,3-glucans selectively from fungal cell walls [135]. Engagement of Dectin-1 initiates intracellular signaling pathways that promote cytokine synthesis and facilitates the differentiation of Th17 and Th1 helper T cells [135,136] both critical for mucosal defense and integrity [137]. In such manner, colonization with Candida albicans can trigger IL-17-producing T cells that protect against pathogenic bacteria invasion – a symbiotic immune activation that benefits the host [130]. However, if fungal growth is not kept in check, overactivation of the immune system may occur.
6.2. Fungi in Cancer and the “Tumor Mycobiome”
Recent research indicate that fungi are not confined to the gut lumen – they can spread or inhabit in various tissues, including tumors [138,139,140,141]. Large-scale sequencing studies across thousands of human tumor samples have discovered detectable fungal DNA (and even whole cells microscopically) in many types of cancers, generally at low levels and with cancer-type-specific profiles [128,139,140]. One comprehensive study found that fungal signals were present in ~35% of samples across 17 cancer types, at an average of ~1 fungal cell per 10,000 tumor cells [128]. Intriguingly, the composition of these cancer-associated fungi varied by cancer: gastrointestinal (GI) tumors often had higher levels of Candida species, lung tumors had signatures of environmental molds like Aspergillus or Blastomyces and pancreatic tumors displayed a notable predominance of Malassezia yeasts [128,140,142,143]. Malassezia is a genus normally present on skin and within the sebaceous glands, but researchers discovered it can migrate up the pancreatic ductal tree [140]. In a mouse model of pancreatic ductal adenocarcinoma (PDA), gut fungi migrated into the pancreas and significantly sped up the tumor progression [140]. PDA-bearing mice exhibited a ~3000-fold increase in fungal abundance in their tumors, with Malassezia predominating over other fungi. More interestingly, eliminating the fungi with antifungals protected the mice from tumor growth, while reintroducing Malassezia – but not Candida or other fungi – restored the cancer acceleration [140]. The mechanism was traced to the immune system’s complement cascade: Malassezia cell wall glycans triggered the mannose-binding lectin (MBL) pathway of complement activation, creating an inflammatory environment that promoted tumor progression [140]. Deletion of MBL or complement C3 eliminated this pro-tumor effect, implicating complement as a key mediator of fungus-driven oncogenesis [140]. In another study, Candida in colon tumors was correlated with increased metastatic disease and alterations in cell adhesion molecules, indicating that these fungi might promote a more aggressive tumor phenotype [128].
Conversely, it may be that some of these associations reflect fungi opportunistically colonizing immunosuppressed tumor environments rather than actively promoting cancer. Determining cause versus effect is a difficult challenge. Nonetheless, the emerging idea of a “tumor mycobiome” has important implications: fungi and their metabolites within tumor microenvironments might modulate local immune responses [138]. Fungi in tumors could continuously activate PRRs, leading to either tumor-promoting inflammation or anergic tolerance, depending on context. The tumor mycobiome also presents potential biomarker opportunities – a study found that the presence of specific fungi in tumor tissues was predictive of reduced patient survival, raising the possibility that fungal DNA detection could serve as a prognostic indicator [128]. This is a novel area of research, so mechanistic understanding is limited, but it is becoming increasingly clear that fungi are yet another important player in the complex ecosystem of the tumor microenvironment.
6.3. Mycobiome Influence on Cancer Immunotherapy (ICI Efficacy)
Given that gut bacteria can affect patients’ responses to immune checkpoint inhibitor (ICI) therapies, it naturally raises the question of whether the gut mycobiome also influences immunotherapy outcomes. Although research is in its incipient phases, initial findings suggest that the mycobiome indeed contributes to the variance in ICI efficacy as summarized in Table 2. A recent multi-cohort study analyzed 862 fecal metagenomes from cancer patients receiving ICI therapy across several cohorts to identify microbial characteristics linked to treatment response [32]. Strikingly, fungal species emerged as important indicators of response: a machine learning model using gut fungal profiles could predict ICI responders vs. non-responders with an average area under the ROC curve (AUC) of 0.87(32). This performance was equivalent to, and independent of, bacterial markers. In fact, the integration of fungal markers with bacterial biomarkers led to an enhanced predictive accuracy (AUC ~0.89), highlighting that the mycobiome offers supplementary information beyond the bacteriome [32]. Specific fungi highlighted in that study include Schizosaccharomyces octosporus, a SCFA-producing fermentative yeast that was enriched in ICI responders’ guts [32]. SCFAs are known to strengthen anti-tumor immunity by enhancing T cell function [144,145], so this fungus might indirectly support the anti-cancer immune response through its metabolites. Another finding was that responders, as predicted by fungal markers, showed higher baseline levels of “exhausted” T cells in peripheral blood [32]. This could suggest that certain fungal components are causing a mild chronic T cell activation that primes the immune system. That alone could predict a more effective ICI outcome.
Additional clinical data comes from smaller cohort studies. In advanced hepatocellular carcinoma (HCC) patients receiving ICIs, one study reported that those who achieved lasting clinical benefit had a greater mycobiome α-diversity at baseline compared to non-responders [146]. This mirrored the pattern observed with bacterial diversity, however, the magnitude of differences in fungal communities was less pronounced than that of bacteria in this HCC cohort, and few specific fungal taxa showed significant association with outcomes [146]. Fungi may affect the response to immunotherapy primarily alongside bacteria instead of acting as the main contributors independently, interactions between the two, symbiotic or antagonistic, could modulate the net immunological outcome.
6.4. Fungal-Derived Immune Modulators (β-Glucans and Others)
Another way the mycobiome can affect cancer immunity is through isolated fungal components used as therapeutics. A notable example is β-glucan, the polysaccharide derived from yeast cell walls. In oncology, purified β-glucans, sourced from Saccharomyces cerevisiae or medicinal mushrooms, have shown immune-stimulating and tumor-suppressing properties [134,143,147,148]. Preclinical studies indicate that β-glucan can prime neutrophils and macrophages [136] to more effectively eliminate tumor cells, especially in the presence of tumor-targeting antibodies, resulting in an additional CR3-dependent cytotoxicity [147]. In the realm of checkpoint blockade immunotherapy, incorporating β-glucan has demonstrated promising synergistic outcomes in experiments [149,150,151]. A study conducted on mice [151] tested a particulate yeast β-glucan (WGP) together with anti–PD-1 and anti–PD-L1 antibodies. The combined treatment resulted in increased immune cell infiltration into tumors and a more favorable tumor microenvironment compared to administrating ICIs alone – specifically, more dendritic cells and effector T cells accumulated in the tumor, while gene expression indicated a shift toward Th1/CTL responses [151]. Tumor growth was significantly slower in mice receiving the β-glucan + ICI combination, suggesting enhanced anti-tumor efficacy [151]. Mechanistically, it appears β-glucan stimulation can diminish the immunosuppressive activity of myeloid-derived suppressor cells and Tregs through Dectin-1 signaling, while also activating antigen-presenting cells [152]. The overall effect is to “unleash” the immune system further when PD-1/PD-L1 blockade removes inhibitory signals on T cells. Exploratory clinical reports suggested that β-glucan supplementation might help overcome resistance to PD-1 inhibitors in patients. In another study, cancer patients who had progressed on PD-1 inhibitors experienced tumor shrinkage or disease stabilization after starting a daily yeast β-glucan supplement alongside continued anti-PD-1 therapy [153]. Currently, clinical trials are beginning to formally test β-glucans as immunotherapy adjuvants [152,154].
7. Gut Virome in Immunity and Cancer
7.1. Composition of the Gut Virome and Baseline Role
The gut virome refers to the collection of viruses found in the gastrointestinal tract [158]. This includes bacteriophages as well as viruses that infect human cells [159]. Phages are believed to be highly prevalent in the gut – estimates indicate that viral particles might equal or exceed bacterial cells in feces [160,161]. Most of these are double-stranded DNA phages, members of families like Caudovirales and single-stranded DNA (ssDNA) phages of the family Microviridae, which infect common gut bacteria [161]. Through infection and lysis cycles, phages regulate bacterial population size and turnover by reducing the numbers of commensal bacteria that have grown excessively dominant [160,162]. Phages also mediate horizontal gene transfer between bacteria by transduction, some improving bacterial fitness or virulence, such as toxin genes or antibiotic resistance factors, influencing the range of microbial molecules that engage with the host immune system [161].
Besides phages, the gut virome includes eukaryotic viruses that infect human cells, such as rotaviruses, noroviruses, adenoviruses, astroviruses, enteroviruses, and many others [163]. In healthy individuals, the majority of these infections are asymptomatic or cause only mild short-lived illness. A study by Kernbauer et al. (2014)[164] showed that an innocuous murine norovirus infection in germ-free mice was able to restore intestinal immune functions typically absent in germ-free animals. Germ-free mice have undeveloped gut immunity, but when these mice were mono-colonized with mouse norovirus (MNV), many of their intestinal immune defects were corrected – the virus effectively substituted for the “education” that commensal bacteria typically deliver [164]. This finding introduced the idea of “viral commensalism”: certain viruses can function as commensal-like agents that modulate the baseline immune state [164]. The presence of a harmless virus, mainly through shaping the bacteria population, keeps the immune system vigilant, maintaining structures such as gut-associated lymphoid tissue and priming innate interferon pathways, all without inducing disease symptoms [162]. Constantly, the body is exposed to dietary and environmental viruses. The presence of viral particles in the stool of most healthy individuals indicates that the immune system is consistently detecting and handling these exposures. Table 3 presents current evidence on viral influences in immunotherapy—spanning gut virome diversity, blood anellovirus burden, oncolytic virus combinations, latent virus reactivation, and bacteriophages—and outlines where associations with ICI efficacy or toxicity are established versus still emerging.
7.2. Virome Alterations in Cancer and Therapy: Lessons from Virome Depletion
Evidence for the direct influence of the virome on anti-cancer immunity came from a recent mouse study examining how disrupting the virome impacts tumor treatment [165]. Researchers treated tumor-bearing mice with a combination of antiviral drugs to broadly deplete the gut virome and then assessed the outcome of chemotherapy (5-fluorouracil) on colorectal cancer [165]. The antiviral cocktail substantially reduced both DNA and RNA viruses in the mice’s guts, without killing bacteria. This resulted in faster tumor growth, poorer survival and reduced efficacy of chemotherapy compared to control mice with an intact virome [165]. The antiviral-treated mice had fewer dendritic cells and CD8+ T-cells within tumors, suggesting an impaired immune response [165]. After the researchers restored the gut virome by conducting a fecal microbiota transplant (FMT) from normal mice into the antiviral-treated mice, the anti-tumor immunity was rescued and tumor management improved [165]. By investigating the mechanism, they discovered that antiviral treatment had specifically downregulated the TLR3–IRF3–IFN-β pathway in both the gut and tumor microenvironment [165]. In normal mice, a degree of TLR3 signaling likely occurs due to the virome during small episodes of viral replication or release of phage dsDNA/RNA fragments, resulting in the synthesis of IFN-β and chemokines that help recruit dendritic cells and T cells to the tumor. The researchers confirmed this by administering a synthetic TLR3 agonist (polyinosinic:polycytidylic acid, poly(I:C)) to virome-depleted mice, which effectively mimics viral dsRNA. Remarkably, poly(I:C) treatment restored dendritic cell and CD8+ T-cell infiltration into tumors and enhanced the efficacy of 5-FU chemotherapy in those mice [165]. This hightlights that that the immune dysfunction was due to the absence of viral-sensing stimulation, and reintroducing that signal restored anti-tumor immunity. Even though this particular study was in the context of chemotherapy, the principles are likely to extend to immunotherapy with ICIs [33].
7.3. Clinical Observations: Virome and Immune Checkpoint Inhibitors Therapy
Direct clinical data regarding the virome’s influence on cancer immunotherapy remains limited, but a few points are worth noting. One attempt to find a virome-based biomarker involved anelloviruses, which are ubiquitous, non-pathogenic DNA viruses that typically rise in titer when the immune system is suppressed. Pescarmona et al. (2021)(166) monitored torque teno virus (TTV) levels present in the blood of advanced melanoma patients receiving anti–PD-1 treatment, reasoning that TTV level could indicate the degree of immune competence, since elevated TTV implies weaker immune control. However, they found no significant difference in TTV viral load between responders and non-responders, and anti–PD-1 therapy did not consistently alter TTV levels [166]. Prior to treatment, TTV counts in melanoma patients were similar to healthy individuals and during therapy some patients experienced slight TTV rises or falls regardless of outcome [166]. This negative result suggests that a single plasma virus titer is unlikely to represent an informative biomarker for ICI effectiveness. The virome’s influence might be more complex – potentially involving tissue-localized effects or the combined presence of many virus types. It also shoes that not every virus modulates immunity in a way that affects tumor responses. In immunocompetent cancer patients on PD-1 blockers, the immune system is not globally suppressed (in fact it’s activated), so it’s perhaps unsurprising that TTV, a marker of severe immunosuppression, wasn’t relevant.
Another aspect of the virome’s relevance lies in immune-related adverse events (irAEs) from ICIs. Evidence suggests that latent viruses can be “unleashed” when checkpoints are blocked, resulting in tissue damage [167,168,169,170,171,172]. A notable example comes from ICI-induced hepatitis: A 2021 study conducted by Hutchinson et al. examined melanoma patients who developed hepatitis on combined anti–PD-1 + anti–CTLA-4 therapy. They found that these patients frequently had an expansion of effector memory CD4⁺ T-cells specific for viruses like Epstein-Barr virus (EBV) or cytomegalovirus (CMV)(173). Checkpoint blockade not only reactivated T cells against tumor antigens but also memory T cells targeting latent viral antigens, which then caused collateral liver damage. They also observed a seasonal timing – patients with pre-existing high CMV antibody titers and expanded CMV-specific T cells were more prone to hepatitis, suggesting subclinical CMV reactivation in the setting of metastatic disease [173]. Importantly, patients identified to be at risk who received PD-1 monotherapy or prophylactic anti-CMV medication had reduced incidence of hepatitis compared to those given full PD-1+CTLA-4 without preventive measures [173]. This study highlights how the virome can influence toxicity: the loss of PD-1-mediated tolerance enabled a “virus-specific immunopathology” to emerge. Similar mechanisms might underlie other irAEs – some cases of checkpoint-induced encephalitis have been linked to T cells targeting obscure viruses or even gut microbes cross-reacting with brain tissue [174]. Clinicians do sometimes check for viral triggers – interestingly, CMV colitis can flare in ICI patients on steroids, and distinguishing CMV colitis from autoimmune colitis is important for choosing the appropriate treatment [167,169,171]. Enhanced antiviral therapy could be a key strategy to ensure the safe administration of ICIs in infected patients [175].
On the other hand, chronic viruses might sometimes assist anti-tumor immunity. A multicenter study found that cancer patients, excluding those with liver cancer, who happened to have chronic hepatitis B virus (HBV) infection responded better to anti–PD-1 therapy compared to their HBV-negative counterparts [176]. These HBV+ patients had improved objective response rates and extended survival on immunotherapy [176]. Immune profiling indicated that HBV infection had led to a higher baseline frequency of exhausted CD8+ T cells (HBV-specific T cells) in these patients, which were reactivated with PD-1 blockade and contributed to anti-tumor effects[176]. This finding is quite intriguing – it implies that not all chronic viral infections are detrimental in cancer therapy. On the contrary, some might create an immune environment that is actually more responsive to checkpoint inhibition.
The concept of using viruses for cancer therapy is not new: oncolytic viruses are an active area of treatment development. T-VEC (talimogene laherparepvec), an engineered herpes simplex virus, is approved for treating melanoma – it infects and lyses tumor cells while promoting local inflammation. Additional oncolytic viruses currently undergoing trials for various tumors [177,178]. The effectiveness of these approaches further highlights that viruses can serve as potent tools to provoke an immune response against cancers. They essentially convert “cold” tumors into “hot” ones by releasing tumor antigens in the context of viral PAMPs and cell death.
The gut virome, although not intentionally introduced, might perform a mini oncolytic-like function if viral replication occasionally leads to bursts of interferon and cell debris release in the gut, systemically enhancing immune vigilance. However, most of the virome’s impact on immunity likely comes via indirect routes through bacteria [162]. There is evidence in mouse models studies that phage populations shift during cancer progression and in response to treatments [162]. However, distinguishing cause from effect is challenging in vivo due to the complexity of phage dynamics. This area will benefit from thorough preclinical studies.
8. Modulation Strategies for the Gut Microbiome in ICI Therapy
8.1. Dietary Interventions
Diet significantly influences the composition of the gut microbiota as shown in Table 4. Recent clinical studies show that high-fiber and plant-rich diets are associated with better ICI outcomes [186], while some other dietary patterns may have negative effects. For example, melanoma patients who consumed over 20g of dietary fiber daily experienced significantly longer progression-free survival on anti-PD-1 treatment than those eating less fiber. Each additional 5g of daily fiber intake corresponded to a 30% reduced risk of progression or death [186]. In the same study, patients on high-fiber diets who avoided probiotic supplements responded best to ICI, while those taking over-the-counter probiotics experienced worse outcomes [186]. A separate cohort study conducted in Poland found that high consumption of plant-based foods was linked with improved anti–PD-1 response, while high dairy consumption correlated with worse outcomes [187]. Intriguingly, that study also noted that patients with daily bowel movements (likely due to higher fiber intake) had better responses compared to those with less frequent stool passage, highlighting the link between diet, gut motility, and microbiome health [187]. Traditional high-fiber dietary patterns such as the Mediterranean diet, have been proposed as supportive measures during immunotherapy [78,188]. These diets increase gut production of short-chain fatty acids (SCFAs), which activate gut receptors (GPCR41/43) on immune cells and have been correlated with better anti–PD-1 outcomes [78]. In contrast, diets high in simple sugars and fats such as the “Western” diet, may promote microbial metabolites that are less favorable or potentially immunosuppressive [69,189,190,191]. Excessive sugar also supports Candida albicans, a commensal yeast that can overgrow with high-carb diets and antibiotic use, potentially skewing immunity toward inflammatory Th17 responses [130]. Therefore, a balanced diet that’s low in refined carbohydrates may help in keeping the gut mycobiome in check.
Fasting and ketogenic diets have been taken into consideration as well [98,188]. Intermittent fasting or calorie restriction eating can positively alter the microbiome and immune profile: a recent systematic review found that pairing ICIs with an intermittent fasting or fasting-mimicking diet enhanced tumor control in mouse models and was associated with higher levels of SCFA-producing bacteria in both mice and patients [98]. These bacteria are known for their immunomodulatory metabolites and shown a positive correlation with objective responses in patients [98,192,193,194]. Ketogenic diets, on the other hand, may result in a reduced fiber intake and potentially decrease microbiota diversity and SCFA production. Although keto diets are sometimes adopted in cancer for metabolic purposes, caution is advised [188,195]. Another example is the “IMmunoDiet” study in cancer patients [196]. Ongoing trials are testing whether high-fiber or Mediterranean diet interventions can directly improve ICI efficacy [195,197]. In clinical practice, nutritional guidance may become a standard component of immunotherapy care.
8.2. Prebiotics and Supplements
Prebiotics are nutrients, usually fibers or oligosaccharides, that specifically support healthy gut microbes. Supplementing cancer patients’ diets with prebiotics might enhance ICI efficacy by increasing beneficial bacteria and their metabolites as shown in Table 5. Common prebiotics include inulin and fructo-oligosaccharides (FOS), galacto-oligosaccharides, resistant starch, and pectins. Resistant starch supplementation is known to enhance the growth of starch-degrading Ruminococcaceae and related fermenters, increasing butyrate and SCAFs production in the colon [218]. Inulin/FOS promote growth of Bifidobacterium species, which have been associated with improved responses in melanoma and other cancers [98,219,220]. Another example are polyphenols in green tea, pomegranate, and berries which are metabolized by gut bacteria into bioactive compounds that may improve anti-tumor immunity [18,221]. These can increase the prevalence of beneficial taxa like Lactobacillus and Bifidobacterium simultaneously suppressing pathogenic bacteria and fungi – polyphenols have mild antimicrobial properties that spare commensals [18,69,198].
Postbiotics can be considered as dietary supplements. Butyrate or its derivatives could potentially be administered to replicate the effect of a fiber-rich microbiome in stimulating anti-tumor T cells and protecting the gut lining [144,222,223]. β-glucans, complex polysaccharides derived from fungal cell walls, are another supplement currently being studied [134]. When taken orally, particulate yeast β-glucan can be taken up by gut phagocytes and ultimately prime neutrophils in the bone marrow to destroy tumor cells more effectively, while also activating dendritic cells [134,136,224]. These immunostimulatory polysaccharides function primarily as oral adjuvants, triggering PRRs and promoting a Th1-dominated, anti-tumor immune response [69]. In a similar manner, certain bacterial TLR agonists may be regarded as “postbiotic” immune supplements – though these are more often delivered as drugs or intratumoral agents rather than oral supplements.
Another important “supplement” to consider is vitamin D. Adequate vitamin D levels has been correlated with a favorable microbiome environment and immune response. A study focused on advanced melanoma patients revealed that individuals with adequate 25(OH)D levels (from supplementation or sunlight) during anti–PD-1 therapy had significantly higher objective response rates (56% vs 36% ORR) and longer PFS (11.25 vs 5.75 months) compared to vitamin D–deficient patients [225]. Although that study did not directly analyze the microbiome, separate research has noted that vitamin D can shape the gut microbiota – patients low in vitamin D had lower abundance of Ruminococcaceae and Lachnospiraceae, which increased following vitamin D repletion [78,226]. The retrospective analysis found even better ICI outcomes and fewer colitis events in patients taking vitamin D supplements [225,226]. Given vitamin D’s role in mucosal immunity and maintaining barrier integrity, ensuring cancer patients are not deficient could be a simple step to potentially reduce dysbiosis and inflammation.
Short-Chain Fatty Acids (SCFAs): These are key players since they are messengers between microbiota and immune cells. Systemic butyrate and propionate levels have been correlated with better ICI responses – patients with higher serum butyrate had better anti–PD-1 outcomes in cancer studies [78,222]. SCFAs can bind to GPR41/43 on immune cells[78]; butyrate, in particular, influences dendritic cell and T-cell differentiation by promoting IL-10 and Tregs locally, while also enhancing CD8 memory and IFN-γ production systemically at low concentrations [69]. Therapeutically, can be provided through oral resistant starch (prebiotic) or encapsulated butyrate. There is also interest in using butyrate enemas for the treatment of checkpoint-induced colitis while maintaining anti-tumor immunity.
8.3. Probiotics and “Mycobiotics”
Probiotics are live microorganisms administered for health advantages. In cancer immunotherapy, probiotics have a complex role, yet to be fully understood [86,233,234,235]. The majority of probiotics available on the market contain strains of Lactobacillus or Bifidobacterium that are generally beneficial for gut health [100]. Despite that, unregulated use of generic probiotics hasn’t demonstrated clear benefit in ICI-treated patients [235]. An observational study found that melanoma patients taking over-the-counter probiotics had worse responses to PD-1 blockade, particulary if their dietary fiber intake was low [186]. One hypothesis is that these standard probiotics might not integrate well or could even reduce the diversity of the native microbiome if used carelessly. Overwhelming the gut with large amounts of exogenous Lactobacillus could outcompete certain commensals that are actually more crucial for anti-cancer immunity (such as Akkermansia or Faecalibacterium) or simply may not address the specific deficiencies of an individual’s microbiome [186]. The focus is therefore shifting to next-generation probiotics – choosing commensal bacteria recognized by research as enhancing ICI efficacy and delivering them in a targeted way. Consortia of multiple selected commensals - “VE800”, a defined 11-strain mix by Vedanta Biosciences used in NCT04208958 clinical trial, and MB097 (nine different species of gut commensal bacteria, all linked to positive CPI response in multiple clinical studies) used in NCT06540391 clinical trial. Early results from these trials are pending, but they represent an initial shift to a rational, personalized probiotic approach.
Beyond bacteria, probiotic fungi, also referred to as “mycobiotics”, are an emerging concept. The best-known example is Saccharomyces boulardii, a non-pathogenic yeast commonly used to prevent or treat antibiotic-associated diarrhea and Clostridioides difficile infection [236]. S. boulardii modulates gut immunity and releases enzymes that neutralize bacterial toxins [237]. Supporting this concept, a recent proof-of-principle study engineered S. cerevisiae var. boulardii to secrete miniature anti-PD-L1 proteins (Sb_haPD-1), showing that orally delivered yeast can transiently transit the gut, secrete functional ICIs in situ, be well tolerated, shift gut bacterial communities, reduce Treg prevalence in mesenteric nodes, and significantly lower distal small-intestinal tumor burden in an ICI-refractory colorectal cancer mouse model [238]. Although S. boulardii hasn’t been specifically studied in ICI patients, it is speculated that it could help maintain microbiome balance during therapy or antibiotic use and possibly lower the risk of checkpoint-induced colitis by outcompeting opportunistic Candida.
Personalized probiotic approaches will probably be necessary – using stool sequencing to identify which key beneficial microbes a patient lacks, followed by supplementing those. Irresponsible use of one-size-fits-all probiotics should be replaced by this tailored approach promises improved results as summarized in Table 6.
8.4. Fecal Microbiota Transplantation (FMT)
Fecal microbiota transplantation – the transfer of stool from a healthy donor to a patient – is the most straightforward method to modify the entire gut microbiome. FMT is already an established therapy for refractory C. difficile infection [241] as drawn in Figure 2., and now it’s currently being tested as an immunotherapy adjunct [15,242,243]. Evidence from both animal and human studies indicates that germ-free or antibiotic-treated mice have diminished responses to ICIs, which can be restored by transplanting feces from responder patients [242,244,245]. In humans, important trials in 2021 offered strong proof that FMT can influence ICI outcomes [26,28]. In melanoma patients unresponsive to anti–PD-1, FMT from elite responders combined with resuming anti–PD-1 therapy led to clinical responses in a subset of recipients [26,27,28]. A certain percentage of previously refractory patients experienced tumor regression or disease stabilization following FMT plus pembrolizumab, presumably due to engraftment of immunomodulatory microbes from the donors [26]. Bacteria taxa which may be responsible for FMT success in ICI treated patients is summarized in Table 7.
Overall, FMT is in its initial phase within oncology, but the early successes in melanoma have sparked hope [75,245]. Larger trials are underway to determine if FMT can consistently rescue patients who don’t respond to ICIs [242,246]. If validated, FMT or next-gen microbiome products inspired by FMT could become part of the immunotherapy arsenal.
FMT’s broad mechanism – resetting the gut ecosystem – could influence not just efficacy but also toxicity(111, 122). Interest exists in using FMT to address immune-related colitis with some case reports showing FMT can help ICI-associated colitis and steroid-refractory colitis in ICI patients by reintroducing regulatory microbes(93, 249, 250). This dual benefit, enhancing anti-tumor immunity while healing the gut lining, would make microbiome modulation quite unique in therapy.
9. Future Directions
The concept of considering the microbiome as part of cancer treatment is quickly gaining popularity. Oncologists might soon routinely seek the advice of microbiome specialists or nutritionists to personalize immunotherapy protocols. Much as we check PD-L1 or tumor genomics, we can also profile a patient’s gut microbiota before therapy. The ultimate goal is a true multi-disciplinary approach to cancer management – where microbiologists join oncologists in tumor boards to interpret microbiome data and recommend interventions. Although numerous questions persist – which species matter most? how to safely implement these ideas? how to integrate with existing treatments? – the advancements made so far are promising. Despite significant advances, substantial gaps remain in our understanding of the roles of the gut mycobiome and virome [68,126], the mechanisms that drive multi-kingdom interactions [32,66,146], and the optimal strategies for microbiome modulation in clinical practice [86,118,235]. There is also a need for standardized protocols, large-scale randomized trials, and integration of multi-omics data to develop certain biomarkers and interventions.
10. Conclusion
In conclusion, the gut bacteriome plays a key role in modulating the host immune system’s readiness to fight cancer. Insights gained from bacteria have established the paradigm that manipulating the microbiota could serve as a novel adjunct strategy in oncology – either by predicting which patients are likely to respond to immunotherapy or by actively altering the microbiome to improve outcomes. Current research is extending this investigation to the other constituents of the microbiome. The upcoming challenge is understanding how gut fungi and viruses, which live alongside bacteria, influence to the immunological environment in cancer patients receiving ICIs. Early evidence suggests that these too may have significant effects on T cell activation status and on which bacteria thrive or decline under therapy. Therefore, while bacteria have been at the forefront of microbiome-cancer research, a holistic view of the “oncobiome” is emerging, promising a deeper and more complete understanding of the host–microbe interactions in cancer immunotherapy.
Supplementary Materials
Not applicable.
Author Contributions
Conceptualization, A.L.A., G.G.P. and O.A.; writing—original draft preparation, .L.A., A.D.G., O.A., G.G.P., A.R.T.; writing—review and editing, A.L.A., A.D.G., O.A., G.G.P., A.R.T.; visualization, A.L.A.; All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [CrossRef]
- Siegel, R.L.; Kratzer, T.B.; Giaquinto, A.N.; Sung, H.; Jemal, A. Cancer statistics, 2025. CA Cancer J. Clin. 2025, 75, 10–45.
- Garg, P.; Pareek, S.; Kulkarni, P.; Horne, D.; Salgia, R.; Singhal, S.S. Next-generation immunotherapy: Advancing clinical applications in cancer treatment. J. Clin. Med. 2024, 13, 21. [CrossRef]
- Sun, Q.; Hong, Z.; Zhang, C.; Wang, L.; Han, Z.; Ma, D. Immune checkpoint therapy for solid tumours: Clinical dilemmas and future trends. Signal Transduct. Target. Ther. 2023, 8, 320. [CrossRef]
- Natanael, R.C.; Măruțescu, L.; Grădișteanu Pircalabioru, G. Immuno-oncology at the crossroads: Confronting challenges in the quest for effective cancer therapies. Int. J. Mol. Sci. 2025, 26, 1–20. [CrossRef]
- Adashek, J.J.; Moran, J.A.; Le, D.T.; Kurzrock, R. Lessons learned from a decade of immune checkpoint inhibition: The good, the bad, and the ugly. Cancer Metastasis Rev. 2025, 44, 43–58. [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742.
- Kong, X.; Zhang, J.; Chen, S.; Wang, X.; Xi, Q.; Shen, H.; et al. Immune checkpoint inhibitors: Breakthroughs in cancer treatment. Cancer Biol. Med. 2024, 21, 451–472. [CrossRef]
- Kciuk, M.; Yahya, E.B.; Mohamed Ibrahim Mohamed, M.; Rashid, S.; Iqbal, M.O.; Kontek, R.; et al. Recent advances in molecular mechanisms of cancer immunotherapy. Cancers 2023, 15, 10. [CrossRef]
- Hargrave, A.; Mustafa, A.S.; Hanif, A.; Tunio, J.H.; Hanif, S.N.M. Recent advances in cancer immunotherapy with a focus on FDA-approved vaccines and neoantigen-based vaccines. Vaccines 2023, 11, 11. [CrossRef]
- Jia, Y.; Liu, L.; Shan, B. Future of immune checkpoint inhibitors: Focus on tumor immune microenvironment. Ann. Transl. Med. 2020, 8, 1095. [CrossRef]
- Muthukutty, P.; Woo, H.Y.; Ragothaman, M.; Yoo, S.Y. Recent advances in cancer immunotherapy delivery modalities. Pharmaceutics 2023, 15, 2. [CrossRef]
- Johnson, D.B.; Nebhan, C.A.; Moslehi, J.J.; Balko, J.M. Immune-checkpoint inhibitors: Long-term implications of toxicity. Nat. Rev. Clin. Oncol. 2022, 19, 254–267. [CrossRef]
- Shen, X.; Yang, J.; Qian, G.; Sheng, M.; Wang, Y.; Li, G.; et al. Treatment-related adverse events of immune checkpoint inhibitors in clinical trials: A systematic review and meta-analysis. Front. Oncol. 2024, 14, 1391724. [CrossRef]
- Gazzaniga, F.; Dennis, L.K. The gut microbiome and cancer response to immune checkpoint inhibitors. J. Clin. Invest. 2025, 135, eXXXXX.
- Xie, J.; Liu, M.; Deng, X.; Tang, Y.; Zheng, S.; Ou, X.; et al. Gut microbiota reshapes cancer immunotherapy efficacy: Mechanisms and therapeutic strategies. iMeta 2024, 3, eXXX. [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836.
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; et al. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [CrossRef]
- Shao, T.; Hsu, R.; Rafizadeh, D.L.; Wang, L.; Bowlus, C.L.; Kumar, N.; et al. The gut ecosystem and immune tolerance. J. Autoimmun. 2023, 141, 103114. [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 2015, 350, 1084–1089. [CrossRef]
- Gharaibeh, R.; Jobin, C. Microbiota and cancer immunotherapy: In search of microbial signals. Gut 2018, 68, 385–388. [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.-L.; et al. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [CrossRef]
- Davar, D.; Dzutsev, A.K.; McCulloch, J.A.; Rodrigues, R.R.; Chauvin, J.-M.; Morrison, R.M.; et al. Fecal microbiota transplant overcomes resistance to anti–PD-1 therapy in melanoma patients. Science 2021, 371, 595–602. [CrossRef]
- Baruch, E.N.; Golan, T.; Wargo, J.A. Fecal microbiota transplantation as a means of overcoming immunotherapy-resistant cancers—Hype or hope? Ther. Adv. Med. Oncol. 2021, 13, 1–15. [CrossRef]
- Baruch, E.N.; Youngster, I.; Ben-Betzalel, G.; Ortenberg, R.; Lahat, A.; Katz, L.; et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science 2020, 371, 602–609. [CrossRef]
- Verheijden, R.J.; van Eijs, M.J.M.; Paganelli, F.L.; Viveen, M.C.; Rogers, M.R.C.; Top, J.; et al. Gut microbiome and immune checkpoint inhibitor toxicity. Eur. J. Cancer 2025, 216, 115221. [CrossRef]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Roux, K.L.; Monot, C.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [CrossRef]
- Lin, Y.; Xie, M.; Lau, H.C.; Zeng, R.; Zhang, R.; Wang, L.; et al. Effects of gut microbiota on immune checkpoint inhibitors in multi-cancer and as microbial biomarkers for predicting therapeutic response. Med 2025, 6, 100530. [CrossRef]
- Huang, X.; Hu, M.; Sun, T.; Li, J.; Zhou, Y.; Yan, Y.; et al. Multi-kingdom gut microbiota analyses define bacterial–fungal interplay and microbial markers of pan-cancer immunotherapy across cohorts. Cell Host Microbe 2023, 31, 1930–1943.e4. [CrossRef]
- Broecker, F.; Moelling, K. The roles of the virome in cancer. Microorganisms 2021, 9, 12.
- Swann, J.B.; Smyth, M.J. Immune surveillance of tumors. J. Clin. Invest. 2007, 117, 1137–1146. [CrossRef]
- Archilla-Ortega, A.; Domuro, C.; Martín-Liberal, J.; Muñoz, P. Blockade of novel immune checkpoints and new therapeutic combinations to boost antitumor immunity. J. Exp. Clin. Cancer Res. 2022, 41, 1–15. [CrossRef]
- Yang, Y.L.; Yang, F.; Huang, Z.Q.; Li, Y.Y.; Shi, H.Y.; Sun, Q.; et al. T cells, NK cells, and tumor-associated macrophages in cancer immunotherapy and the current state of the art of drug delivery systems. Front. Immunol. 2023, 14, 1199173. [CrossRef]
- Gaynor, N.; Crown, J.; Collins, D. Immune checkpoint inhibitors: Key trials and an emerging role in breast cancer. Semin. Cancer Biol. 2020, 65, 44–52. [CrossRef]
- Lee, L.; Gupta, M.; Sahasranaman, S. Immune checkpoint inhibitors: An introduction to the next-generation cancer immunotherapy. J. Clin. Pharmacol. 2016, 56, 157–169. [CrossRef]
- Esfahani, K.; Roudaia, L.; Buhlaiga, N.; Rincón, S.; Papneja, N.; Miller, W. A review of cancer immunotherapy: From the past, to the present, to the future. Curr. Oncol. 2019, 27, S87–S97. [CrossRef]
- Frankel, A.E.; Deshmukh, S.; Reddy, A.; Lightcap, J.; Hayes, M.; McClellan, S.; et al. Cancer immune checkpoint inhibitor therapy and the gut microbiota. Integr. Cancer Ther. 2019, 18, 1–10. [CrossRef]
- Faget, J.; Peters, S.; Quantin, X.; Meylan, E.; Bonnefoy, N. Neutrophils in the era of immune checkpoint blockade. J. Immunother. Cancer 2021, 9, e002242. [CrossRef]
- Choi, Y.; Seok, S.H.; Yoon, H.Y.; Ryu, J.H.; Kwon, I.C. Advancing cancer immunotherapy through siRNA-based gene silencing for immune checkpoint blockade. Adv. Drug Deliv. Rev. 2024, 204, 115306. [CrossRef]
- Elkrief, A.; Derosa, L.; Zitvogel, L.; Kroemer, G.; Routy, B. The intimate relationship between gut microbiota and cancer immunotherapy. Gut Microbes 2019, 10, 424–428. [CrossRef]
- Cartwright, D.; Kidd, A.; Ansel, S.; Ascierto, M.; Spiliopoulou, P. Oncogenic signalling pathways in cancer immunotherapy: Leader or follower in this delicate dance? Int. J. Mol. Sci. 2025, 26, 1–22. [CrossRef]
- Fenton, S.E.; Sosman, J.A.; Chandra, S. Resistance mechanisms in melanoma to immune-oncologic therapy with checkpoint inhibitors. Cancer Drug Resist. 2019, 2, 744–761.
- Hassel, J.C.; Zimmer, L.; Sickmann, T.; Eigentler, T.K.; Meier, F.; Mohr, P.; et al. Medical needs and therapeutic options for melanoma patients resistant to anti-PD-1-directed immune checkpoint inhibition. Cancers 2023, 15, 13. [CrossRef]
- Mihaila, R.I.; Gheorghe, A.S.; Zob, D.L.; Stanculeanu, D.L. The importance of predictive biomarkers and their correlation with the response to immunotherapy in solid tumors—Impact on clinical practice. Biomedicines 2024, 12, 9.
- Sankar, K.; Ye, J.; Li, Z.; Zheng, L.; Song, W.; Hu-Lieskovan, S. The role of biomarkers in personalized immunotherapy. Biomark. Res. 2022, 10, 1–15. [CrossRef]
- Yu, X.; Fang, L. The role of the gut microbiota in tumor, immunity, and immunotherapy. Front. Immunol. 2024, 15, 1–18.
- Lim, M.; Han, S.; Nam, Y.-D. Understanding the role of the gut microbiome in solid tumor responses to immune checkpoint inhibitors for personalized therapeutic strategies: A review. Front. Immunol. 2025, 15, 1–20. [CrossRef]
- Lee, P.-C.; Wu, C.-J.; Hung, Y.; Lin, C.-J.; Chao, Y.; Hou, M.; et al. Association of gut microbiota and metabolites with tumor response to immune checkpoint inhibitors in patients with unresectable hepatocellular carcinoma. J. Clin. Oncol. 2021, 39, eXXXX. [CrossRef]
- Das, S.; Johnson, D. Immune-related adverse events and anti-tumor efficacy of immune checkpoint inhibitors. J. Immunother. Cancer 2019, 7, 306. [CrossRef]
- Yin, Q.; Wu, L.; Han, L.; Zheng, X.; Tong, R.; Li, L.; et al. Immune-related adverse events of immune checkpoint inhibitors: A review. Front. Immunol. 2023, 14, 1–18. [CrossRef]
- Miao, Y.-D.; Quan, W.-X.; Tang, X.-L.; Shi, W.-W.; Li, Q.; Li, R.J.; et al. Uncovering the flip side of immune checkpoint inhibitors: A comprehensive review of immune-related adverse events and predictive biomarkers. Int. J. Biol. Sci. 2024, 20, 621–642. [CrossRef]
- Mallio, C.A.; Bernetti, C.; Cea, L.; Buoso, A.; Stiffi, M.; Vertulli, D.; et al. Adverse effects of immune-checkpoint inhibitors: A comprehensive imaging-oriented review. Curr. Oncol. 2023, 30, 4700–4723. [CrossRef]
- Di Dalmazi, G.; Ippolito, S.; Lupi, I.; Caturegli, P. Hypophysitis induced by immune checkpoint inhibitors: A 10-year assessment. Expert Rev. Endocrinol. Metab. 2019, 14, 381–398. [CrossRef]
- He, C.; Qu, T. Immune checkpoint inhibitor-related cutaneous adverse events. Asia Pac. J. Clin. Oncol. 2020, 16, e149–e153. [CrossRef]
- Som, A.; Mandaliya, R.; Alsaadi, D.; Farshidpour, M.; Charabaty, A.; Malhotra, N.; et al. Immune checkpoint inhibitor-induced colitis: A comprehensive review. World J. Clin. Cases 2019, 7, 405–418. [CrossRef]
- Guitton, R.; Laparra, A.; Chanson, N.; Champiat, S.; Danlos, F.-X.; Michot, J.-M.; et al. Immune-related adverse events occurring rapidly after a single dose of immune checkpoint blockade. J. Immunother. Cancer 2025, 13, eXXXX. [CrossRef]
- Jayathilaka, B.; Mian, F.; Franchini, F.; Au-Yeung, G.; Iqbal, M.J. Cancer- and treatment-specific incidence rates of immune-related adverse events induced by immune checkpoint inhibitors: A systematic review. Br. J. Cancer 2025, 132, 51–57. [CrossRef]
- Lu, D.; Yao, J.; Yuan, G.; Gao, Y.; Zhang, J.; Guo, X. Immune checkpoint inhibitor-related new-onset thyroid dysfunction: A retrospective analysis using the US FDA Adverse Event Reporting System. Oncologist 2022, 27, e126–e132. [CrossRef]
- Yamauchi, I.; Yabe, D. Best practices in the management of thyroid dysfunction induced by immune checkpoint inhibitors. Eur. Thyroid J. 2025, 14, 1–12. [CrossRef]
- Frankel, A.E.; Coughlin, L.A.; Kim, J.; Froehlich, T.; Xie, Y.; Frenkel, E.P.; et al. Metagenomic shotgun sequencing and unbiased metabolomic profiling identify specific human gut microbiota and metabolites associated with immune checkpoint therapy efficacy in melanoma patients. Neoplasia 2017, 19, 848–855. [CrossRef]
- Baiden-Amissah, R.; Tuyaerts, S. Contribution of aging, obesity, and microbiota on tumor immunotherapy efficacy and toxicity. Int. J. Mol. Sci. 2019, 20, 3586. [CrossRef]
- Yang, K.; Li, G.; Li, Q.; Wang, W.; Zhao, X.; Shao, N.; et al. Distribution of gut microbiota across intestinal segments and their impact on human physiological and pathological processes. Cell Biosci. 2025, 15, 47. [CrossRef]
- Zhang, M.; Zhang, Y.; Zhao, Z.; Deng, F.; Jiang, H.; Liu, C.; et al. Bacterial–fungal interactions: Mutualism, antagonism, and competition. Life 2025, 15, 8. [CrossRef]
- Perez, J.C. The interplay between gut bacteria and the yeast Candida albicans. Gut Microbes 2021, 13, 1979877. [CrossRef]
- Easwaran, M.; Abdelrahman, F.; El-Shibiny, A.; Venkidasamy, B.; Thiruvengadam, M.; Sivalingam, P.; et al. Exploring bacteriophages to combat gut dysbiosis: A promising new frontier in microbiome therapy. Microb. Pathog. 2025, 208, 108008. [CrossRef]
- Campbell, C.; Kandalgaonkar, M.; Golonka, R.; Yeoh, B.; Vijay-Kumar, M.; Saha, P. Crosstalk between gut microbiota and host immunity: Impact on inflammation and immunotherapy. Biomedicines 2023, 11, 2110. [CrossRef]
- Gierynska, M.; Szulc-Dabrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the intestinal barrier: The involvement of epithelial cells and microbiota—A mutual relationship. Animals 2022, 12, 2. [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [CrossRef]
- Gong, J.; Chehrazi-Raffle, A.; Placencio-Hickok, V.; Guan, M.; Hendifar, A.; Salgia, R. The gut microbiome and response to immune checkpoint inhibitors: Preclinical and clinical strategies. Clin. Transl. Med. 2019, 8, 9. [CrossRef]
- Shen, Y.; Fan, N.; Ma, S.X.; Cheng, X.; Yang, X.; Wang, G. Gut microbiota dysbiosis: Pathogenesis, diseases, prevention, and therapy. MedComm 2025, 6, e70168. [CrossRef]
- Thapa, R.; Magar, A.T.; Shrestha, J.; Panth, N.; Idrees, S.; Sadaf, T.; et al. Influence of gut and lung dysbiosis on lung cancer progression and their modulation as promising therapeutic targets: A comprehensive review. MedComm 2024, 5, eXXX. [CrossRef]
- Elkrief, A.; Routy, B.; Daillère, L.; Bolte, L.; Wargo, J.; McQuade, J.; et al. Gut microbiota in immuno-oncology: A practical guide for medical oncologists with a focus on antibiotics stewardship. ASCO Educ. Book 2025, 45, e472902. [CrossRef]
- Xu, L.; Hu, B.; He, J.; Fu, X.; Liu, N. Intratumor microbiome-derived butyrate promotes chemo-resistance in colorectal cancer. Front. Pharmacol. 2024, 15, 1510851. [CrossRef]
- Duan, Y.; Xu, Y.; Dou, Y.; Xu, D. Helicobacter pylori and gastric cancer: Mechanisms and new perspectives. J. Hematol. Oncol. 2025, 18, 10. [CrossRef]
- Gamrath, L.; Pedersen, T.B.; Møller, M.V.; Volmer, L.; Holst-Christensen, L.; Vestermark, L.; et al. Role of the microbiome and diet for response to cancer checkpoint immunotherapy: A narrative review of clinical trials. Curr. Oncol. Rep. 2025, 27, 45–58. [CrossRef]
- Wang, Y.; Fu, K. Genotoxins: The mechanistic links between Escherichia coli and colorectal cancer. Cancers 2023, 15, 4. [CrossRef]
- Buder-Bakhaya, K.; Hassel, J.C. Biomarkers for clinical benefit of immune checkpoint inhibitor treatment—A review from the melanoma perspective and beyond. Front. Immunol. 2018, 9, 1474. [CrossRef]
- Ranjbar, M.; Salehi, R.; Haghjooy Javanmard, S.; Rafiee, L.; Faraji, H.; Jafarpor, S.; et al. The dysbiosis signature of Fusobacterium nucleatum in colorectal cancer—Cause or consequences? A systematic review. Cancer Cell Int. 2021, 21, 194. [CrossRef]
- Khoudia, D.; Mbaye, B.; Ndir, S.; Favier, A.; Bercovici, M.; Jouvin, M.; et al. Coupling culturomics and metagenomics sequencing to characterize the gut microbiome of patients with cancer treated with immune checkpoint inhibitors. Gut Pathog. 2025, 17, 1–15.
- Carlson, R.D.; Flickinger, J.C., Jr.; Snook, A.E. Talkin’ toxins: From Coley’s to modern cancer immunotherapy. Toxins 2020, 12, 4. [CrossRef]
- Yu, X.; Li, W.; Li, Z.; Wu, Q.; Sun, S. Influence of microbiota on tumor immunotherapy. Int. J. Biol. Sci. 2024, 20, 2264–2294. [CrossRef]
- Zhu, X.; Hu, M.; Huang, X.; Li, L.; Lin, X.; Shao, X.; et al. Interplay between gut microbial communities and metabolites modulates pan-cancer immunotherapy responses. Cell Metab. 2025, 37, 806–823.e6. [CrossRef]
- Li, W.; Zhang, K.; Liu, Y.; Qiu, L.; Hong, Z. Gut microbiota shapes cancer immunotherapy responses. NPJ Biofilms Microbiomes 2025, 11, 1–14.
- Li, S.; Liu, J.; Ma, Y.; Zhang, W. Gut microbiota affects PD-L1 therapy and its mechanism in melanoma. Cancer Immunol. Immunother. 2025, 74, 1–14.
- Jiminez, V.; Yusuf, N. Role of the microbiome in immunotherapy of melanoma. Cancer J. 2023, 29, 70–74. [CrossRef]
- Usyk, M.; Pandey, A.; Hayes, R.; Moran, U.; Pavlick, A.; Osman, I.; et al. Bacteroides vulgatus and Bacteroides dorei predict immune-related adverse events in immune checkpoint blockade treatment of metastatic melanoma. Genome Med. 2021, 13, 1–15. [CrossRef]
- Dubin, K.; Callahan, M.K.; Ren, B.; Khanin, R.; Viale, A.; Ling, L.; et al. Intestinal microbiome analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat. Commun. 2016, 7, 10391. [CrossRef]
- Hu, W.; Zhang, Y.; Wang, Y.; Liu, X.; Sun, T.; Xu, J. Exploring fecal microbiota signatures associated with immune response and antibiotic impact in NSCLC: Insights from metagenomic and machine learning approaches. Front. Cell. Infect. Microbiol. 2025, 15, 1–18. [CrossRef]
- Hu, W.; Xu, J.; Sun, T.; Zhang, Y.; Fang, Y.; Li, H.; et al. Exploring gut microbiome markers in predicting efficacy of immunotherapy against lung cancer. J. Clin. Oncol. 2024, 42, eXXXX.
- Schwartz, D.J.; Rebeck, O.N.; Dantas, G. Complex interactions between the microbiome and cancer immune therapy. Crit. Rev. Clin. Lab. Sci. 2019, 56, 567–585. [CrossRef]
- Fan, S.; Jiang, Z.; Zhang, Z.; Xing, J.; Wang, D.; Tang, D. Akkermansia muciniphila: A potential booster to improve the effectiveness of cancer immunotherapy. J. Cancer Res. Clin. Oncol. 2023, 149, 13477–13494. [CrossRef]
- Li, L.; McAllister, F. Too much water drowned the miller: Akkermansia determines immunotherapy responses. Cell Rep. Med. 2022, 3, 100642. [CrossRef]
- Grenda, A.; Iwan, E.; Chmielewska, I.; Krawczyk, P.; Giza, A.; Bomba, A.; et al. Presence of Akkermansiaceae in gut microbiome and immunotherapy effectiveness in patients with advanced non-small cell lung cancer. AMB Express 2022, 12, 1–10. [CrossRef]
- Derosa, L.; Routy, B.; Thomas, A.M.; Iebba, V.; Zalcman, G.; Friard, S.; et al. Intestinal Akkermansia muciniphila predicts clinical response to PD-1 blockade in patients with advanced non-small-cell lung cancer. Nat. Med. 2022, 28, 315–324. [CrossRef]
- Somodi, C.; Dora, D.; Horvath, M.; Szegvari, G.; Lohinai, Z. Gut microbiome changes and cancer immunotherapy outcomes associated with dietary interventions: A systematic review of preclinical and clinical evidence. J. Transl. Med. 2025, 23, 756. [CrossRef]
- Chan, M.A.; Gallegos, A.A.; Ortega, J.A.; Sánchez, E.; Mayora, X.; Yocupicio, O.; et al. The interplay between the gut microbiome and immunotherapy in gastrointestinal malignancies: Mechanisms, clinical implications, and therapeutic potential. Int. J. Med. Sci. Clin. Res. Stud. 2025, 5, 1–15.
- Xu, M.; Li, X.; Lau, H.C.; Yu, J. The gut microbiota in cancer immunity and immunotherapy. Cell. Mol. Immunol. 2025, 22, 1012–1031 . [CrossRef]
- Pei, B.; Peng, S.; Huang, C.; Zhou, F. Bifidobacterium modulation of tumor immunotherapy and its mechanism. Cancer Immunol. Immunother. 2024, 73, 94. [CrossRef]
- Yi, Z.; Wang, T.; Tang, X.; Hu, Y.; Zhang, H.; Tang, D.; et al. Gut microbiome affects the response to anti-PD-1 immunotherapy in patients with hepatocellular carcinoma. J. Immunother. Cancer 2019, 7, 1–12.
- Lee, P.-C.; Wu, C.-J.; Hung, Y.; Lin, C.-J.; Chen-Ta, C.; Lin, I.-C.; et al. Gut microbiota and metabolites associate with outcomes of immune checkpoint inhibitor–treated unresectable hepatocellular carcinoma. J. Immunother. Cancer 2022, 10, eXXXX.
- Mao, J.; Wang, D.; Jiang, L.; Xu, Y.; Li, J.; Shi, Y.; et al. Gut microbiome is associated with the clinical response to anti-PD-1 based immunotherapy in hepatobiliary cancers. J. Immunother. Cancer 2021, 9, eXXXX.
- Peng, K.; Li, Y.; Yang, Q.; Yu, P.; Zeng, T.; Lin, C.; et al. The therapeutic promise of probiotic Bacteroides fragilis (BF839) in cancer immunotherapy. Front. Microbiol. 2025, 16, 1523754. [CrossRef]
- Wang, Z.; Zhang, S.-J.; Liu, L.; Huang, Z.; He, D.; Hu, Y. The gut microbiota modulates responses to anti–PD-1 and chemotherapy combination therapy and related adverse events in patients with advanced solid tumors. Front. Oncol. 2022, 12, 1–15.
- Gupta, H.; Fatima, K.; Kaur, J. Understanding crosstalk between the gut and liver microbiome: Pathogenesis to therapeutic approaches in liver cancer. Cancer Cell Int. 2025, 25, 1–20. [CrossRef]
- Joy, S.; Cortes, E.G.; Otsuji, T.; Chen, H.; Dy, G.; Segal, B.; et al. Fluctuations in gut microbiome composition during immune checkpoint inhibitor therapy. World J. Oncol. 2023, 14, 178–187.
- Zhang, M.; Liu, J.; Xia, Q. Role of gut microbiome in cancer immunotherapy: From predictive biomarker to therapeutic target. Exp. Hematol. Oncol. 2023, 12, 1–15. [CrossRef]
- Wargo, J.A.; Andrews, M.C.; Duong, C.P.M.; Gopalakrishnan, V.; Iebba, V.; Chen, W.-S.; et al. Gut microbiota signatures are associated with toxicity to combined CTLA-4 and PD-1 blockade. Nat. Med. 2020, 27, 1432–1441. [CrossRef]
- Liu, X.; Tang, H.; Zhou, Q.; Zeng, Y.; Lu, B.; Chen, D.; et al. Gut microbiota composition in patients with advanced malignancies experiencing immune-related adverse events. Front. Immunol. 2023, 14, 1–14. [CrossRef]
- He, C.; Li, M.; Li, B.; Zhang, H.; Qiu, D.; Fang, X.; et al. Relating gut microbiome and its modulating factors to immunotherapy in solid tumors: A systematic review. Front. Oncol. 2020, 11, 1–18.
- Usyk, M.; Huilin, R.B.; Kostic, R.; Gopalakrishnan, A.; Ling, H.; Osman, I.; et al. Gut microbiome is associated with recurrence-free survival in patients with resected Stage IIIB-D or Stage IV melanoma treated with immune checkpoint inhibitors. bioRxiv 2024, preprint.
- Zhou, G.; Zhang, N.; Meng, K.; Pan, F. Interaction between gut microbiota and immune checkpoint inhibitor-related colitis. Front. Immunol. 2022, 13, 1001623. [CrossRef]
- Velikova, T.; Krastev, B.; Lozenov, S.; Gencheva, R.; Peshevska-Sekulovska, M.; Nikolaev, G.; et al. Antibiotic-related changes in microbiome: The hidden villain behind colorectal carcinoma immunotherapy failure. Int. J. Mol. Sci. 2021, 22, 1–20. [CrossRef]
- Elkrief, A.; Derosa, L.; Kroemer, G.; Zitvogel, L.; Routy, B. The negative impact of antibiotics on outcomes in cancer patients treated with immunotherapy: A new independent prognostic factor? Ann. Oncol. 2019, 30, 1–8. [CrossRef]
- Yang, Y.; Shi, X. Big lessons from the little Akkermansia muciniphila in hepatocellular carcinoma. Front. Immunol. 2025, 16, 1524563. [CrossRef]
- Barragan-Carrillo, R.; Zengin, Z.; Pal, S. Microbiome modulation for the treatment of solid neoplasms. J. Clin. Oncol. 2025, 43, 2734–2738. [CrossRef]
- Yin, H.; Lu, Y.; Pan, G.; Ye, K.; Ma, Y.; Han, X.; et al. The commensal consortium of the gut microbiome is associated with favorable responses to anti-programmed death protein 1 (PD-1) therapy in thoracic neoplasms. Cancer Biol. Med. 2021, 18, 1040–1052.
- Xu, Y.; Pan, G.; Shi, W.; Zhang, X.; Hu, X.; Chen, X.-J. Gut microbiota as a prognostic biomarker for unresectable hepatocellular carcinoma treated with anti-PD-1 therapy. Front. Genet. 2024, 15, 1–12.
- Zhang, Y.; Chen, S.; Zou, H.; Han, Z.; Xie, T.; Zhang, B.; et al. Correlation of the gut microbiome and immune-related adverse events in gastrointestinal cancer patients treated with immune checkpoint inhibitors. Front. Cell. Infect. Microbiol. 2023, 13, 1–14. [CrossRef]
- Liu, X.; Tang, H.; Zhou, Q.; Zeng, Y.; Lu, B.; Chen, M.; et al. Gut microbiome contributes to the development of immune checkpoint inhibitor-related colitis. J. Crohns Colitis 2023, 17, 1–12. [CrossRef]
- Sakai, K.; Sakurai, T.; Velasco, M.D.D.; Nagai, T.; Chikugo, T.; Ueshima, K.; et al. Intestinal microbiota and gene expression reveal similarity and dissimilarity between immune-mediated colitis and ulcerative colitis. Front. Oncol. 2021, 11, 1–16. [CrossRef]
- Pi, Z.; Chen, S.; Kou, Y.; Wang, Z.; Jiang, R.; Han, H.; et al. The gut microbiome is associated with clinical response to anti–PD-1/PD-L1 immunotherapy in gastrointestinal cancer. Cancer Immunol. Res. 2020, 8, 1251–1261.
- Sun, Y.-C.; Lee, P.-C.; Kuo, Y.; Wang, W.-K.; Chang, C.-C.; Chen, L.; et al. An exploratory study for the association of gut microbiome with efficacy of immune checkpoint inhibitor in patients with hepatocellular carcinoma. J. Hepatocell. Carcinoma 2021, 8, 809–822.
- Iliev, I.D.; Cadwell, K. Effects of intestinal fungi and viruses on immune responses and inflammatory bowel diseases. Gastroenterology 2020, 159, 1–15. [CrossRef]
- Zong, Z.; Zhou, F.; Zhang, L. The fungal mycobiome: A new hallmark of cancer revealed by pan-cancer analyses. Signal Transduct. Target. Ther. 2023, 8, 50. [CrossRef]
- Dohlman, A.B.; Klug, J.; Mesko, M.; Gao, I.H.; Lipkin, S.M.; Shen, X.; et al. A pan-cancer mycobiome analysis reveals fungal involvement in gastrointestinal and lung tumors. Cell 2022, 185, 3807–3822.e12. [CrossRef]
- Liu, H.Y.; Li, S.; Ogamune, K.J.; Ahmed, A.A.; Kim, I.H.; Zhang, Y.; et al. Fungi in the gut microbiota: Interactions, homeostasis, and host physiology. Microorganisms 2025, 13, 1. [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [CrossRef]
- Mukherjee, P.K.; Sendid, B.; Hoarau, G.; Colombel, J.-F.; Poulain, D.; Ghannoum, M.A. Mycobiota in gastrointestinal diseases. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 77–87.
- Chehoud, C.; O’Toole, P.W.; Cotter, P.D.; Scanlan, P.D. Forgotten fungi—The gut mycobiome in human health and disease. FEMS Microbiol. Rev. 2017, 41, 479–511. [CrossRef]
- Hoarau, G.; Mukherjee, P.K.; Gower-Rousseau, C.; Hager, C.; Chandra, J.; Retuerto, M.A.; et al. Bacteriome and mycobiome interactions underscore microbial dysbiosis in familial Crohn's disease. mBio 2016, 7, e01250-16. [CrossRef]
- Briard, B.; Fontaine, T.; Kanneganti, T.-D.; Gow, N.A.R.; Papon, N. Fungal cell wall components modulate our immune system. Cell Surf. 2021, 7, 100067. [CrossRef]
- Marakalala, M.J.; Kerrigan, A.M.; Brown, G.D. Dectin-1: A role in antifungal defense and consequences of genetic polymorphisms in humans. Mamm. Genome 2011, 22, 55–65. [CrossRef]
- Camilli, G.; Tabouret, G.; Quintin, J. The complexity of fungal β-glucan in health and disease: Effects on the mononuclear phagocyte system. Front. Immunol. 2018, 9, 673. [CrossRef]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [CrossRef]
- Wang, Y.; Wang, Y.; Zhou, Y.-H.; Feng, Y.; Sun, T.; Xu, J. Tumor-related fungi and crosstalk with gut fungi in the tumor microenvironment. Cancer Biol. Med. 2024, 21, 977–994. [CrossRef]
- Narunsky-Haziza, L.; Sepich-Poore, G.D.; Livyatan, I.; Asraf, O.; Martino, C.; Nejman, D.; et al. Pan-cancer analyses reveal cancer-type-specific fungal ecologies and bacteriome interactions. Cell 2022, 185, 3789–3806.e17. [CrossRef]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [CrossRef]
- Kozlov, M. Do fungi lurking inside cancers speed their growth? Nature 2022, 604, 1–3. [CrossRef]
- Zhang, L.; Zhou, D.D.; Feng, J.; Liao, Z.J.; Shu, X.L.; Yang, R.M.; et al. Intestinal fungal signatures and their impact on immune checkpoint inhibitor efficacy: A multi-cohort meta-analysis. NPJ Biofilms Microbiomes 2025, 11, 188. [CrossRef]
- Carrillo-Serradell, L.; Liu-Tindall, J.; Planells-Romeo, V.; Aragon-Serrano, L.; Isamat, M.; Gabaldon, T.; et al. The human mycobiome: Composition, immune interactions, and impact on disease. Int. J. Mol. Sci. 2025, 26, 15. [CrossRef]
- Yan, Y.; Ye, M.; Song, Y.; Xu, W.; Zhang, X.; Liu, Y.; et al. Gut microbiota and SCFAs improve the treatment efficacy of chemotherapy and immunotherapy in NSCLC. NPJ Biofilms Microbiomes 2025, 11, 1–14.
- Liu, X.F.; Shao, J.H.; Liao, Y.T.; Wang, L.N.; Jia, Y.; Dong, P.J.; et al. Regulation of short-chain fatty acids in the immune system. Front. Immunol. 2023, 14, 1186892. [CrossRef]
- Zhang, C.; Zhang, C.; Wang, S.; Xu, Z.; Zhang, D.; Zhou, L.; et al. Characterizations of multi-kingdom gut microbiota in immune checkpoint inhibitor-treated hepatocellular carcinoma. J. Immunother. Cancer 2024, 12, eXXXX. [CrossRef]
- Noorbakhsh Varnosfaderani, S.M.; Ebrahimzadeh, F.; Akbari Oryani, M.; Khalili, S.; Almasi, F.; Mosaddeghi Heris, R.; et al. Potential promising anticancer applications of beta-glucans: A review. Biosci. Rep. 2024, 44, BSR2023XXXX. [CrossRef]
- Geller, A.; Shrestha, R.; Yan, J. Yeast-derived beta-glucan in cancer: Novel uses of a traditional therapeutic. Int. J. Mol. Sci. 2019, 20, 3618. [CrossRef]
- Wang, M.; Pan, J.; Xiang, W.; You, Z.; Zhang, Y.; Wang, J.; et al. Beta-glucan: A potent adjuvant in immunotherapy for digestive tract tumors. Front. Immunol. 2024, 15, 1424261. [CrossRef]
- Song, Q.; Xu, Y.; Zhang, M.; Wu, L.; Liu, S.; Lv, Y.; et al. A beta-1,3/1,6-glucan enhances anti-tumor effects of PD1 antibody by reprogramming tumor microenvironment. Int. J. Biol. Macromol. 2024, 279, 134660. [CrossRef]
- Wang, M.; Bai, Y.; Pei, J.; Li, D.; Pu, X.; Zhu, W.; et al. Beta-glucan combined with PD-1/PD-L1 checkpoint blockade for immunotherapy in patients with advanced cancer. Front. Pharmacol. 2022, 13, 887457. [CrossRef]
- Chu, Y.; He, X.; Xue, Y.; Jiang, H.; Zhu, C.; Qi, C.; et al. An exploratory clinical study of beta-glucan combined with camrelizumab and SOX chemotherapy as first-line treatment for advanced gastric adenocarcinoma. Front. Immunol. 2024, 15, 1448485. [CrossRef]
- Kim, E.; Ahn, H.; Park, H. A review on the role of gut microbiota in immune checkpoint blockade therapy for cancer. Mamm. Genome 2021, 32, 223–231. [CrossRef]
- Iglesias, L.; Prathikanti, R.; Ma, B.; Mattson, P.; Kedrowski, D.; Lowe, J.; et al. A multicenter, open-label, phase II study of PGG beta-glucan and pembrolizumab in patients with advanced melanoma (MEL) following progression on treatment with checkpoint inhibitors (CPI) or triple negative breast cancer (TNBC) failing front-line chemotherapy for metastatic disease. J. Clin. Oncol. 2017, 35, TPS3105. [CrossRef]
- Kang, X.; Lau, H.C.; Yu, J. Modulating gut microbiome in cancer immunotherapy: Harnessing microbes to enhance treatment efficacy. Cell Rep. Med. 2024, 5, 101478. [CrossRef]
- Szostak, N.; Handschuh, L.; Samelak-Czajka, A.; Tomela, K.; Pietrzak, B.; Schmidt, M.; et al. Gut mycobiota dysbiosis is associated with melanoma and response to anti-PD-1 therapy. Cancer Immunol. Res. 2024, 12, 427–439. [CrossRef]
- Bernardes, M.; Hohl, T. Fungal infections associated with the use of novel immunotherapeutic agents. Curr. Clin. Microbiol. Rep. 2020, 7, 142–149. [CrossRef]
- Cao, Z.; Sugimura, N.; Burgermeister, E.; Ebert, M.P.; Zuo, T.; Lan, P. The gut virome: A new microbiome component in health and disease. eBioMedicine 2022, 81, 104113. [CrossRef]
- Lathakumari, R.H.; Vajravelu, L.K.; Gopinathan, A.; Vimala, P.B.; Panneerselvam, V.; Ravi, S.S.S.; et al. The gut virome and human health: From diversity to personalized medicine. Eng. Microbiol. 2025, 5, 100191. [CrossRef]
- Taylor, V.L.; Fitzpatrick, A.D.; Islam, Z.; Maxwell, K.L. The diverse impacts of phage morons on bacterial fitness and virulence. Adv. Virus Res. 2019, 103, 1–31.
- Mahmud, M.R.; Tamanna, S.K.; Akter, S.; Mazumder, L.; Akter, S.; Hasan, M.R.; et al. Role of bacteriophages in shaping gut microbial community. Gut Microbes 2024, 16, 2390720.
- Duerkop, B.A.; Hooper, L.V. Resident viruses and their interactions with the immune system. Nat. Immunol. 2013, 14, 654–659. [CrossRef]
- Kennedy, E.A.; Holtz, L.R. Gut virome in early life: Origins and implications. Curr. Opin. Virol. 2022, 55, 101233. [CrossRef]
- Kernbauer, E.; Ding, Y.; Cadwell, K. An enteric virus can replace the beneficial function of commensal bacteria. Nature 2014, 516, 94–98. [CrossRef]
- Huang, H.; Yang, Y.; Wang, X.; Wen, B.; Yang, X.; Zhong, W.; et al. Gut virome dysbiosis impairs antitumor immunity and reduces 5-fluorouracil treatment efficacy for colorectal cancer. Front. Oncol. 2024, 14, 1501981. [CrossRef]
- Pescarmona, R.; Mouton, W.; Walzer, T.; Dalle, S.; Eberhardt, A.; Brengel-Pesce, K.; et al. Evaluation of TTV replication as a biomarker of immune checkpoint inhibitors efficacy in melanoma patients. PLoS ONE 2021, 16, e0255972. [CrossRef]
- Sanyour, J.; Awada, B.; Mattar, A.; Matar, R.; Yaqoub, N.; Al Haddabi, I.; et al. Cytomegalovirus and Epstein–Barr virus reactivation in steroid-refractory immune checkpoint inhibitor colitis. J. Infect. Dev. Ctries. 2025, 19, 1276–1282. [CrossRef]
- Harper, H.; Newman, J.; Sun, W.; MacKenzie, E. Cytomegalovirus hepatitis after treatment with dostarlimab for colorectal cancer. BMJ Case Rep. 2025, 18, eXXXX. [CrossRef]
- Iwamuro, M.; Tanaka, T.; Makimoto, G.; Ichihara, E.; Hiraoka, S. Two cases of cytomegalovirus colitis during the treatment of immune checkpoint inhibitor-associated colitis. Cureus 2024, 16, e63308. [CrossRef]
- Xia, Z.; Zhang, J.; Chen, W.; Zhou, H.; Du, D.; Zhu, K.; et al. Hepatitis B reactivation in cancer patients receiving immune checkpoint inhibitors: A systematic review and meta-analysis. Infect. Dis. Poverty 2023, 12, 87. [CrossRef]
- Anastasopoulou, A.; Samarkos, M.; Diamantopoulos, P.; Vourlakou, C.; Ziogas, D.C.; Avramopoulos, P.; et al. Cytomegalovirus infections in patients treated with immune checkpoint inhibitors for solid malignancies. Open Forum Infect. Dis. 2023, 10, ofad164. [CrossRef]
- Burns, E.A.; Muhsen, I.N.; Anand, K.; Xu, J.; Umoru, G.; Arain, A.N.; et al. Hepatitis B virus reactivation in cancer patients treated with immune checkpoint inhibitors. J. Immunother. 2021, 44, 132–139. [CrossRef]
- Hutchinson, J.A.; Kronenberg, K.; Riquelme, P.; Wenzel, J.J.; Glehr, G.; Schilling, H.L.; et al. Virus-specific memory T cell responses unmasked by immune checkpoint blockade cause hepatitis. Nat. Commun. 2021, 12, 1439.
- Seki, M.; Kitano, S.; Suzuki, S. Neurological disorders associated with immune checkpoint inhibitors: An association with autoantibodies. Cancer Immunol. Immunother. 2022, 71, 769–775. [CrossRef]
- He, L.; Yu, L.; Zhang, C.; Jia, A.; Zhang, L.; Ma, W.; et al. Microbiome meets immunotherapy: Unlocking the hidden predictors of immune checkpoint inhibitors. NPJ Biofilms Microbiomes 2025, 11, 1–14.
- Zhou, J.; Chen, G.; Wang, J.; Zhou, B.; Sun, X.; Wang, J.; et al. Anti-PD-1 therapy achieves favorable outcomes in HBV-positive non-liver cancer. Oncogenesis 2023, 12, 22. [CrossRef]
- Chesney, J.A.; Puzanov, I.; Collichio, F.A.; Singh, P.; Milhem, M.M.; Glaspy, J.; et al. Talimogene laherparepvec in combination with ipilimumab versus ipilimumab alone for advanced melanoma: 5-year final analysis of a multicenter, randomized, open-label, phase II trial. J. Immunother. Cancer 2023, 11, eXXXX. [CrossRef]
- Robert, C.; Gastman, B.; Gogas, H.; Rutkowski, P.; Long, G.V.; Chaney, M.F.; et al. Open-label, phase II study of talimogene laherparepvec plus pembrolizumab for the treatment of advanced melanoma that progressed on prior anti-PD-1 therapy: MASTERKEY-115. Eur. J. Cancer 2024, 207, 114120. [CrossRef]
- Ribas, A.; Dummer, R.; Puzanov, I.; VanderWalde, A.; Andtbacka, R.H.I.; Michielin, O.; et al. Oncolytic virotherapy promotes intratumoral T cell infiltration and improves anti-PD-1 immunotherapy. Cell 2017, 170, 1109–1119.e10. [CrossRef]
- Harrington, K.; Freeman, D.J.; Kelly, B.; Harper, J.; Soria, J.-C. Optimizing oncolytic virotherapy in cancer treatment. Nat. Rev. Drug Discov. 2019, 18, 689–706. [CrossRef]
- Reynolds, K.; Thomas, M.; Dougan, M. Diagnosis and management of hepatitis in patients on checkpoint blockade. Oncologist 2018, 23, 991–997. [CrossRef]
- Tagliamento, M.; Remon, J.; Giaj Levra, M.; De Maria, A.; Bironzo, P.; Besse, B.; et al. Immune checkpoint inhibitors in patients with cancer and infection by hepatitis B or C virus: A perspective through the results of a European survey. JTO Clin. Res. Rep. 2023, 4, 100446. [CrossRef]
- Pandey, A.; Ezemenari, S.; Liaukovich, M.; Richard, I.; Boris, A. A rare case of pembrolizumab-induced reactivation of hepatitis B. Case Rep. Oncol. Med. 2018, 2018, 5985131. [CrossRef]
- Cianci, R.; Caldarelli, M.; Brani, P.; Bosi, A.; Ponti, A.; Giaroni, C.; et al. Cytokines meet phages: A revolutionary pathway to modulating immunity and microbial balance. Biomedicines 2025, 13, 5. [CrossRef]
- Champagne-Jorgensen, K.; Luong, T.; Darby, T.; Roach, D.R. Immunogenicity of bacteriophages. Trends Microbiol. 2023, 31, 1058–1071.
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vétizou, M.; Cogdill, A.P.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [CrossRef]
- Pala, B.; Trzeciak, K.; Olejnik-Schmidt, A.; Gapska, Ł.; Mackiewicz, J.; Kaczmarek, M.; et al. A clinical outcome of the anti-PD-1 therapy of melanoma in Polish patients is mediated by population-specific gut microbiome composition. Cancers 2022, 14, 1–20.
- Grafanaki, K.; Mavridis, A.; Anastogianni, A.; Boumpas, A.; Pappa, E.; Sideris, C. Nutrition and diet patterns as key modulators of metabolic reprogramming in melanoma immunotherapy. J. Clin. Med. 2025, 14, 1–20.
- Severino, A.; Tohumcu, E.; Tamai, L.; Dargenio, P.; Porcari, S.; Rondinella, D.; et al. The microbiome-driven impact of western diet in the development of noncommunicable chronic disorders. Best Pract. Res. Clin. Gastroenterol. 2024, 72, 101923. [CrossRef]
- Garcia-Montero, C.; Fraile-Martinez, O.; Gomez-Lahoz, A.M.; Pekarek, L.; Castellanos, A.J.; Noguerales-Fraguas, F.; et al. Nutritional components in western diet versus Mediterranean diet at the gut microbiota–immune system interplay: Implications for health and disease. Nutrients 2021, 13, 2. [CrossRef]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The impact of Western diet and nutrients on the microbiota and immune response at mucosal interfaces. Front. Immunol. 2017, 8, 838. [CrossRef]
- Zeb, F.; Osaili, T.; Obaid, R.S.; Naja, F.; Radwan, H.; Cheikh Ismail, L.; et al. Gut microbiota and time-restricted feeding/eating: A targeted biomarker and approach in precision nutrition. Nutrients 2023, 15, 2. [CrossRef]
- Popa, A.D.; Nita, O.; Gherasim, A.; Enache, A.I.; Caba, L.; Mihalache, L.; et al. A scoping review of the relationship between intermittent fasting and the human gut microbiota: Current knowledge and future directions. Nutrients 2023, 15, 9. [CrossRef]
- Liu, J.; Shao, N.; Qiu, H.; Zhao, J.; Chen, C.; Wan, J.; et al. Intestinal microbiota: A bridge between intermittent fasting and tumors. Biomed. Pharmacother. 2023, 167, 115484. [CrossRef]
- Zhang, X.; Li, H.; Lv, X.; Hu, L.; Li, W.; Zi, M.; et al. Impact of diets on response to immune checkpoint inhibitors (ICIs) therapy against tumors. Life 2022, 12, 3. [CrossRef]
- Kazmierczak-Siedlecka, K.; Stachowska, E.; Kucharski, R.; Wisniewski, P.; Ulasinski, P.; Stanczak, M.; et al. The impact of enteral immunonutrition on gut microbiota in colorectal cancer and gastric cancer patients in the preoperative period—Preliminary results of randomized clinical trial. Front. Immunol. 2025, 16, 1606187. [CrossRef]
- Bolte, L.A.; Lee, K.A.; Bjork, J.R.; Leeming, E.R.; Campmans-Kuijpers, M.J.E.; de Haan, J.J.; et al. Association of a Mediterranean diet with outcomes for patients treated with immune checkpoint blockade for advanced melanoma. JAMA Oncol. 2023, 9, 705–709. [CrossRef]
- Aziz, T.; Hussain, N.; Hameed, Z.; Lin, L. Elucidating the role of diet in maintaining gut health to reduce the risk of obesity, cardiovascular and other age-related inflammatory diseases: Recent challenges and future recommendations. Gut Microbes 2024, 16, 2297864. [CrossRef]
- Wang, Y.; Wymond, B.; Tandon, H.; Belobrajdic, D.P. Swapping white for high-fibre bread increases faecal abundance of short-chain fatty acid-producing bacteria and microbiome diversity: A randomized, controlled, decentralized trial. Nutrients 2024, 16, 7. [CrossRef]
- McCrory, C.; Lenardon, M.; Traven, A. Bacteria-derived short-chain fatty acids as potential regulators of fungal commensalism and pathogenesis. Trends Microbiol. 2024, 32, 1106–1118. [CrossRef]
- Zhang, F.; Aschenbrenner, D.; Yoo, J.Y.; Zuo, T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe 2022, 3, e969–e983. [CrossRef]
- Gașpar, B.S.; Roșu, O.; Enache, R.; Manciulea, M.; Pavelescu, L.; Crețoiu, S. Gut mycobiome: Latest findings and current knowledge regarding its significance in human health and disease. J. Fungi 2025, 11, 1–20. [CrossRef]
- Xin, Y.; Liu, C.-G.; Zang, D.; Chen, J. Gut microbiota and dietary intervention: Affecting immunotherapy efficacy in non–small cell lung cancer. Front. Immunol. 2024, 15, 1–14. [CrossRef]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [CrossRef]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [CrossRef]
- Perrone, P.; D’Angelo, S. Gut microbiota modulation through Mediterranean diet foods: Implications for human health. Nutrients 2025, 17, 6. [CrossRef]
- Abrignani, V.; Salvo, A.; Pacinella, G.; Tuttolomondo, A. The Mediterranean diet, its microbiome connections, and cardiovascular health: A narrative review. Int. J. Mol. Sci. 2024, 25, 9. [CrossRef]
- Mitsou, E.K.; Kakali, A.; Antonopoulou, S.; Mountzouris, K.C.; Yannakoulia, M.; Panagiotakos, D.B.; et al. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br. J. Nutr. 2017, 117, 1645–1655. [CrossRef]
- Agus, A.; Denizot, J.; Thevenot, J.; Martinez-Medina, M.; Massier, S.; Sauvanet, P.; et al. Western diet induces a shift in microbiota composition enhancing susceptibility to adherent-invasive E. coli infection and intestinal inflammation. Sci. Rep. 2016, 6, 19032. [CrossRef]
- Buttar, J.; Kon, E.; Lee, A.; Kaur, G.; Lunken, G. Effect of diet on the gut mycobiome and potential implications in inflammatory bowel disease. Gut Microbes 2024, 16, 1–18. [CrossRef]
- Musumeci, S.; Coen, M.; Leidi, A.; Schrenzel, J. The human gut mycobiome and the specific role of Candida albicans: Where do we stand, as clinicians? Clin. Microbiol. Infect. 2021, 27, 1–8. [CrossRef]
- Gupta, Y.; Ernst, A.L.; Vorobyev, A.; Beltsiou, F.; Zillikens, D.; Bieber, K.; et al. Impact of diet and host genetics on the murine intestinal mycobiome. Nat. Commun. 2023, 14, 834. [CrossRef]
- Castaldo, G.; D’Elia, M.; De Prisco, M.; Folliero, V.; Marino, C.; D’Ursi, A.; et al. Modulation of gut bacterial and fungal microbiota in fibromyalgia patients following a carb-free oloproteic diet: Evidence for Candida suppression and symptom improvement. Microorganisms 2025, 13, 9. [CrossRef]
- Yu, D.; Xie, L.; Chen, W.; Qin, J.; Zhang, J.; Lei, M.; et al. Dynamics of the gut bacteria and fungi accompanying low-carbohydrate diet-induced weight loss in overweight and obese adults. Front. Nutr. 2022, 9, 846378. [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [CrossRef]
- Ducarmon, Q.R.; Grundler, F.; Le Maho, Y.; Wilhelmi de Toledo, F.; Zeller, G.; Habold, C.; et al. Remodelling of the intestinal ecosystem during caloric restriction and fasting. Trends Microbiol. 2023, 31, 832–844. [CrossRef]
- van der Merwe, M.; Sharma, S.; Caldwell, J.L.; Smith, N.J.; Gomes, C.K.; Bloomer, R.J.; et al. Time of feeding alters obesity-associated parameters and gut bacterial communities, but not fungal populations, in C57BL/6 male mice. Curr. Dev. Nutr. 2020, 4, nzz145. [CrossRef]
- Kadyan, S.; Park, G.; Singh, P.; Arjmandi, B.; Nagpal, R. Prebiotic mechanisms of resistant starches from dietary beans and pulses on gut microbiome and metabolic health in a humanized murine model of aging. Front. Nutr. 2023, 10, 1106463. [CrossRef]
- Boucher, E.; Plazy, C.; Richard, M.-L.; Suau, A.; Mangin, I.; Cornet, M.; et al. Inulin prebiotic reinforces host cancer immunosurveillance via γδ T cell activation. Front. Immunol. 2023, 14, 1104224. [CrossRef]
- Hughes, R.L.; Alvarado, D.A.; Swanson, K.S.; Holscher, H.D. The prebiotic potential of inulin-type fructans: A systematic review. Adv. Nutr. 2022, 13, 492–529. [CrossRef]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [CrossRef]
- Zhu, X.; Li, K.; Liu, G.; Wu, R.; Zhang, Y.; Wang, S.; et al. Microbial metabolite butyrate promotes anti-PD-1 antitumor efficacy by modulating T cell receptor signaling of cytotoxic CD8 T cell. Gut Microbes 2023, 15, 2249143. [CrossRef]
- Chen, G.; Ran, X.; Li, B.; Li, Y.; He, D.; Huang, B.; et al. Sodium butyrate inhibits inflammation and maintains epithelium barrier integrity in a TNBS-induced inflammatory bowel disease mice model. EBioMedicine 2018, 30, 317–325. [CrossRef]
- Li, J.; Chen, D.; Yu, B.; He, J.; Zheng, P.; Mao, X.; et al. Fungi in gastrointestinal tracts of human and mice: From community to functions. Microb. Ecol. 2018, 75, 821–829. [CrossRef]
- Galus, L.; Michalak, M.; Lorenz, M.; Stoinska-Swiniarek, R.; Tusien Malecka, D.; Galus, A.; et al. Vitamin D supplementation increases objective response rate and prolongs progression-free time in patients with advanced melanoma undergoing anti-PD-1 therapy. Cancer 2023, 129, 2047–2055. [CrossRef]
- Yu, Y.; Guo, Y.; Liu, Y.; Jiang, Q.; Yu, C.; Li, Z.; et al. Vitamin D, gut microbiota, and cancer immunotherapy—A potentially effective crosstalk. Int. J. Mol. Sci. 2025, 26, 1–20.
- Wu, X.Z.; Wen, Z.G.; Hua, J.L. Effects of dietary inclusion of Lactobacillus and inulin on growth performance, gut microbiota, nutrient utilization, and immune parameters in broilers. Poult. Sci. 2019, 98, 4656–4663. [CrossRef]
- Sheng, W.; Ji, G.; Zhang, L. Immunomodulatory effects of inulin and its intestinal metabolites. Front. Immunol. 2023, 14, 1224092. [CrossRef]
- Li, X.; Zhang, S.; Gao, G.; Hu, J.; Yu, J. Gut microbiome in modulating immune checkpoint inhibitors. eBioMedicine 2022, 82, 104162. [CrossRef]
- Han, Z.Y.; Fu, Z.J.; Wang, Y.Z.; Zhang, C.; Chen, Q.W.; An, J.X.; et al. Probiotics functionalized with a gallium–polyphenol network modulate the intratumor microbiota and promote anti-tumor immune responses in pancreatic cancer. Nat. Commun. 2024, 15, 7096. [CrossRef]
- Danne, C.; Sokol, H. Butyrate, a new microbiota-dependent player in CD8+ T cells immunity and cancer therapy? Cell Rep. Med. 2021, 2, 100328. [CrossRef]
- David, A.; Lev-Ari, S. Targeting the gut microbiome to improve immunotherapy outcomes: A review. Integr. Cancer Ther. 2024, 23, 1–18. [CrossRef]
- Dahiya, D.; Nigam, P. Biotherapy using probiotics as therapeutic agents to restore the gut microbiota to relieve gastrointestinal tract inflammation, IBD, IBS and prevent induction of cancer. Int. J. Mol. Sci. 2023, 24, 1–20. [CrossRef]
- Tripathy, A.; Dash, J.; Kancharla, S.; Kolli, P.; Mahajan, D.; Senapati, S.; et al. Probiotics: A promising candidate for management of colorectal cancer. Cancers 2021, 13, 1–25. [CrossRef]
- Rukavina, L.; Golčić, M.; Mikolasevic, I. The relationship between the modulation of intestinal microbiota and the response to immunotherapy in patients with cancer. Biomedicines 2025, 13, 1–20. [CrossRef]
- Chen, K.; Zhu, Y.; Zhang, Y.; Hamza, T.; Yu, H.; Saint Fleur, A.; et al. A probiotic yeast-based immunotherapy against Clostridioides difficile infection. Sci. Transl. Med. 2020, 12, eaba0375. [CrossRef]
- Pakbin, B.; Dibazar, S.P.; Allahyari, S.; Javadi, M.; Amani, Z.; Farasat, A.; et al. Anticancer properties of probiotic Saccharomyces boulardii supernatant on human breast cancer cells. Probiotics Antimicrob. Proteins 2022, 14, 1130–1138. [CrossRef]
- Rebeck, O.N.; Wallace, M.J.; Prusa, J.; Ning, J.; Evbuomwan, E.M.; Rengarajan, S.; et al. A yeast-based oral therapeutic delivers immune checkpoint inhibitors to reduce intestinal tumor burden. Cell Chem. Biol. 2025, 32, 98–110.e7. [CrossRef]
- Cheng, D.; Xie, M.Z. A review of a potential and promising probiotic candidate—Akkermansia muciniphila. J. Appl. Microbiol. 2021, 130, 1813–1822.
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [CrossRef]
- Paaske, S.E.; Baumwall, S.M.D.; Rubak, T.; Birn, F.H.; Ragard, N.; Kelsen, J.; et al. Real-world effectiveness of fecal microbiota transplantation for first or second Clostridioides difficile infection. Clin. Gastroenterol. Hepatol. 2025, 23, 602–611.e8. [CrossRef]
- Yang, Y.; An, Y.; Dong, Y.; Chu, Q.; Wei, J.; Wang, B.; et al. Fecal microbiota transplantation: No longer cinderella in tumour immunotherapy. EBioMedicine 2024, 100, 104967. [CrossRef]
- Wang, W.; Fan, J.; Zhang, C.; Huang, Y.; Chen, Y.; Fu, S.; et al. Targeted modulation of gut and intra-tumor microbiota to improve the quality of immune checkpoint inhibitor responses. Microbiol. Res. 2024, 282, 127668. [CrossRef]
- Chacon, J.; Faizuddin, F.; McKee, J.C.; Sheikh, A.; Vasquez, V.M., Jr.; Gadad, S.S.; et al. Unlocking the microbial symphony: The interplay of human microbiota in cancer immunotherapy response. Cancers 2025, 17, 5. [CrossRef]
- Li, A.; He, L.; Jia, A.; Zhang, L.; Ma, W.; Yu, L.; et al. Microbiota boost immunotherapy? A meta-analysis dives into fecal microbiota transplantation and immune checkpoint inhibitors. BMC Med. 2025, 23, 1–15.
- Oh, B.; Boyle, F.; Pavlakis, N.; Clarke, S.; Eade, T.; Hruby, G.; et al. The gut microbiome and cancer immunotherapy: Can we use the gut microbiome as a predictive biomarker for clinical response in cancer immunotherapy? Cancers 2021, 13, 1–21. [CrossRef]
- Kang, Y.; Cai, Y. Faecal microbiota transplantation enhances efficacy of immune checkpoint inhibitors therapy against cancer. World J. Gastroenterol. 2021, 27, 5362–5375. [CrossRef]
- Derosa, L.; Routy, B.; Favier, M.; Iebba, V.; Alou, M.T.; Pessione, E.; et al. Gut bacteria composition drives primary resistance to cancer immunotherapy in renal cell carcinoma patients. Eur. Urol. 2020, 78, 195–206. [CrossRef]
- Elkrief, A.; Waters, N.R.; Smith, N.; Dai, A.; Slingerland, J.; Aleynick, N.; et al. Immune-related colitis is associated with fecal microbial dysbiosis and can be mitigated by fecal microbiota transplantation. Cancer Immunol. Res. 2024, 12, 308–321. [CrossRef]
- Chen, M.; Liu, M.; Li, C.; Peng, S.; Li, Y.; Xu, X.; et al. Fecal microbiota transplantation effectively cures a patient with severe bleeding immune checkpoint inhibitor-associated colitis and a short review. Front. Oncol. 2022, 12, 913217. [CrossRef]
Figure 1.
Mechanism of action of anti CTLA-4 antibody (ICIs) in cancer treatment.
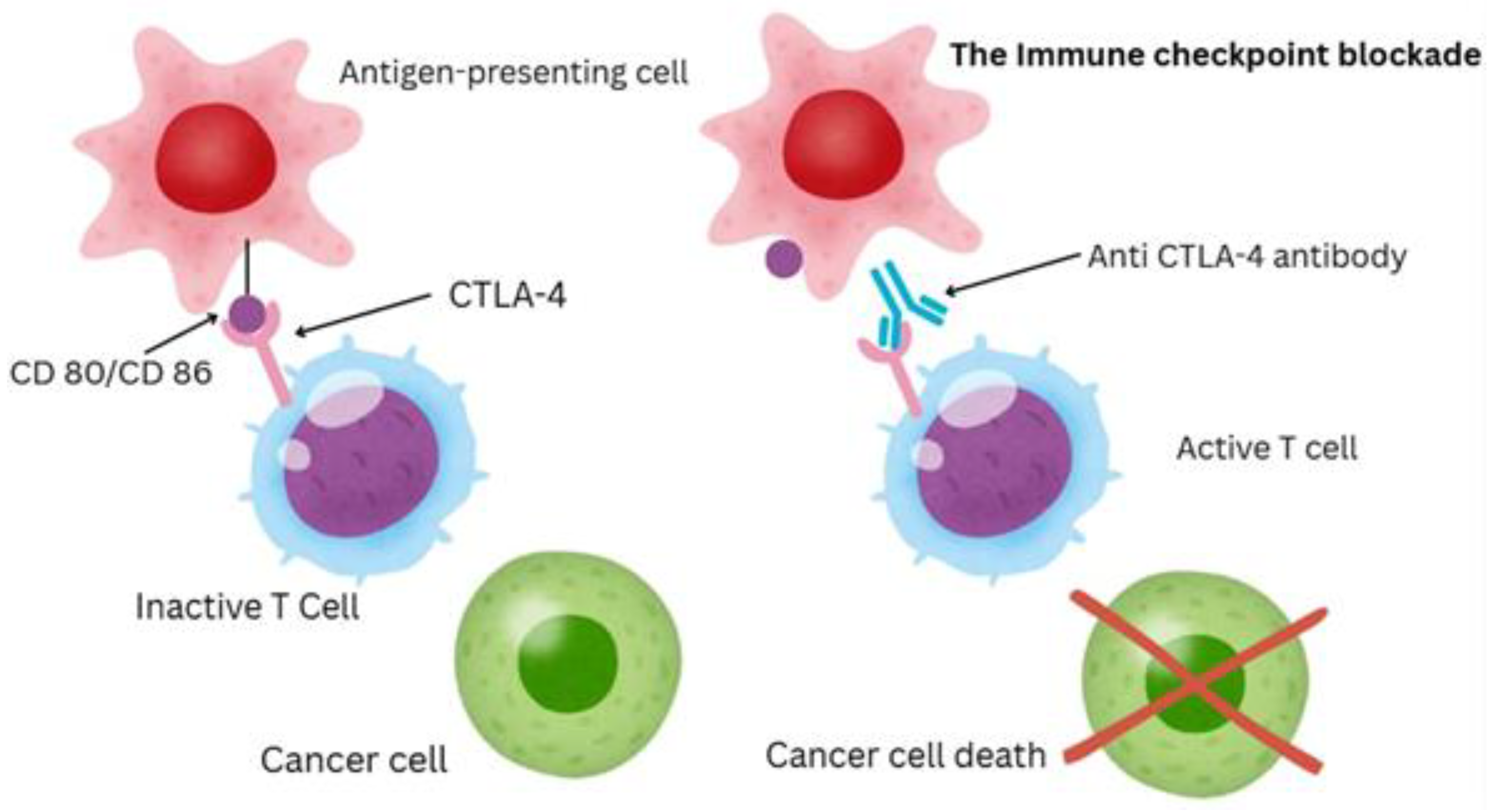
Figure 2.
Fecal microbiota transplant in patients with C. difficile infection.
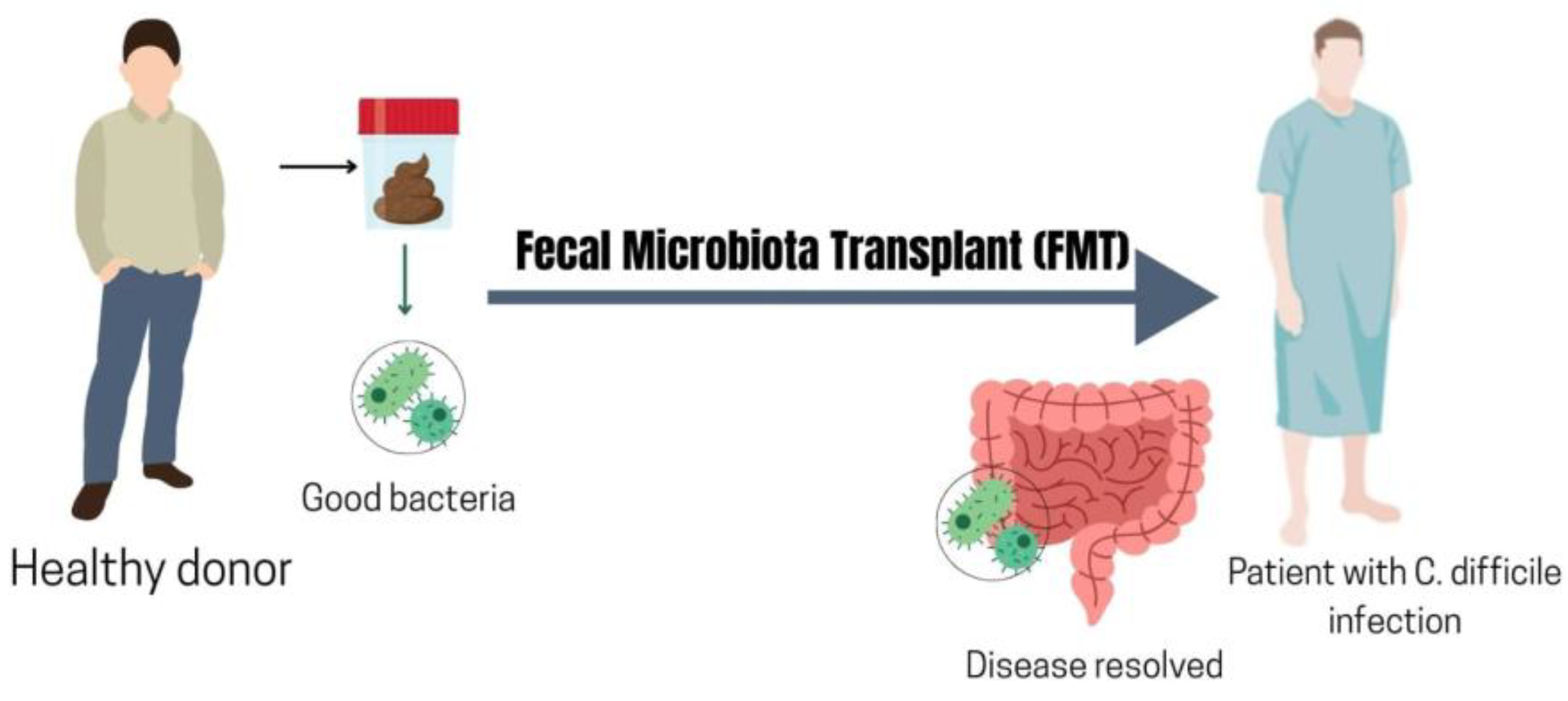

Table 1.
Gut Bacterial taxa or features associated with ICI efficacy and toxicity.
| Bacterial Taxon/Feature | Association with ICI Outcomes | Key References |
| Akkermansia muciniphila | Enriched in responders to PD-1/PD-L1 therapy; absence linked to inferior ICI efficacy. Fecal transfer of Akkermansia-rich microbiota improved anti-PD-1 responses in mice. | [24,29,32,94,95,96,97,98,99,102,108,117,118,119] |
| Faecalibacterium prausnitzii | Consistently enriched in ICI responders across multiple cancers. Produces SCFAs that support T effector and T reg cell metabolism. High baseline abundance is linked to better tumor control (pro-immune environment) but also associated with higher risk of ipilimumab-colitis in melanoma. Illustrates immunity–toxicity trade-off. | [30,31,32,39,98,99,106,114] |
| Bacteroides spp. | Certain Bacteroides are linked to lower GI toxicity: Patients with Bacteroidetes-rich microbiomes had less CTLA-4–induced colitis. B. fragilis produces polysaccharide A which induces regulatory T-cells and was shown to protect against colitis in mice. B. fragilis (and other Bacteroides) also enhanced anti-CTLA-4 tumor responses in preclinical models. However, in some anti–PD-1 studies, non-responders showed higher relative abundance of Bacteroidales showing a context-dependent effect. | [20,39,82,89,90,91,92,105,106,110,111,120,121,122,123,124] |
| Bifidobacterium spp. | Several Bifidobacterium species correlate with improved anti-tumor efficacy. In preclinical models, Bifidobacterium augmented anti–PD-L1 therapy by enhancing dendritic cell function. In patients, higher Bifidobacterium abundance has been noted in melanoma responders. Produces metabolites that stimulate T-cell activity. Not prominently linked to irAEs; generally considered immunostimulatory yet gut friendly. | [21,23,82,93,99,100,101,111,118,122,125] |
| High gut microbiota diversity | Diverse, well-balanced microbiomes are associated with better clinical response to ICIs (observed in melanoma, lung, kidney cancers). Such microbiomes typically include multiple SCFA producers and immunomodulatory taxa. High diversity may also be protective against single-species overgrowth that triggers colitis. Low diversity, dysbiosis – often caused by antibiotics or illness – correlates with poor response and higher risk of complications. | [25,29,39,97,106] |
| Pathobiont enrichment | Overrepresentation of opportunistic bacteria is a red flag for toxicity. Patients with baseline gut expansion of pathobionts had higher odds of severe irAEs. These organisms can provoke inflammation and thrive when beneficial flora are depleted. A post-ICI bloom of pathobionts (and concurrent drop in Clostridial commensals) has been observed at colitis onset. Such dysbiosis likely both predisposes to and results from mucosal inflammation. | [29,116] |
Table 2.
Gut Fungal findings associated with ICI outcomes.
| Fungal Taxon | Association with ICI Efficacy/Toxicity | Key References |
| Schizosaccharomyces octosporus | Enriched in ICI responders in a pan-cancer analysis. A fungal signature including S. octosporus predicted treatment response with high accuracy as this yeast was largely absent in non-responders. It may ferment dietary fibers into short-chain fatty acids, indirectly boosting anti-tumor immunity. Not known to cause disease; its presence is considered beneficial. | [32,143,155] |
| Candida albicans | Mixed effects: Identified as part of a multi-kingdom predictor mode. Moderate colonization might help “prime” immunity, but overgrowth is often a sign of dysbiosis. In responder/non-responder comparisons, C. albicans was generally higher in non-responders, suggesting that an overabundance could be a negative factor for efficacy. | [32,143,156] |
| Core fungal set (n=26) predicted response (AUC 0.87), and multi-kingdom (20 fungi + 17 bacteria) improved AUC to 0.89 | Clear non-responder fungi: Pseudocercospora musae, Daedalea quercina, Lachancea mirantina, Lomentospora prolificans, Enterocytozoon bieneusi, Verticillium nonalfalfae; Clear responder fungi: Trichophyton benhamiae, Cryptococcus amylolentus, Suillus clintonianus, Pseudogymnoascus sp. 05NY08, Schizosaccharomyces octosporus, Podospora anserina, (Verticillium longisporum modestly) |
[32] |
| Saccharomyces paradoxus, Malassezia restricta | Saccharomyces paradoxus - ↑Efficacy (better anti-PD-1 response), Malassezia restricta - ↓Efficacy (worse response) | (143) |
| Overall fungal diversity | Higher mycobiome diversity before ICI therapy has been associated with better treatment outcomes, although the effect size is modest. A rich fungal community might indicate a resilient gut ecosystem. There is no firm evidence that higher fungal diversity increases irAE risk; rather, loss of fungal diversity often accompanies antibiotic use and may be a marker of microbiome perturbation. | (142, 146) |
| Opportunistic fungal blooms | Some reports note that patients who develop severe irAEs have expansion of fungi like Candida albicans, Aspergillus spp., Pneumocystis jirovecii or Fusarium spp., leading to ↑Toxicity. Typically occurred with high-dose immunosuppression for irAEs and/or lymphopenia | (157) |
Table 3.
Gut Virome and viral factors associated with ICI outcomes.
| Viral Factor | Association with ICI Efficacy/Toxicity | References |
| High gut virome diversity | Unlike bacteria/fungi, no specific enteric virus has been identified as a positive or negative predictor of ICI response in humans. Some hypothesize that a diverse gut virome could continually stimulate the immune system, potentially aiding tumor surveillance. However, studies integrating virome analysis have not found strong, consistent differences in viral populations between responders and non-responders. | [32,33,146,159] |
| Blood Anellovirus load/diversity | Anellovirus (TTV) load tracks immune competence—it typically rises with immunosuppression and can reflect host immune tone. In a melanoma anti–PD-1 cohort, TTV load did not differ between responders and non-responders at baseline and didn’t change with therapy, arguing against a simple anellovirus–ICI efficacy link. | [166] |
| Oncolytic virus therapy in combination | Oncolytic viruses can inflame the tumor and attract immune cells. Trials combining oncolytic viruses with ICIs have shown synergistic efficacy. The success of talimogene laherparepvec (T-VEC, a modified HSV) in melanoma supports the concept that introducing certain viruses to the tumor microenvironment can enhance checkpoint inhibitor activity. This underscores how viruses can modulate ICI outcomes by provoking anti-tumor immune responses. | [177,178,179,180] |
| Latent virus reactivation | Risk factor for toxicity: ICIs can remove immune checks on virus-specific T cells, leading to reactivation of latent infections. For example, PD-1 blockade in hepatitis B carriers can trigger fulminant hepatitis from viral reactivation. Similarly, cases of immune-related hepatitis and myocarditis have been linked to T-cell responses against latent viruses like EBV or CMV in affected tissues. These events suggest that part of irAE pathology in some patients may be an immune attack on cells harboring latent viruses. Patients with high viral antigen burden in certain organs could be predisposed to organ-specific irAEs when treated with ICIs. | [4,167,168,169,170,171,172,181,182,183] |
| Bacteriophages | Emerging interest in bacteriophages which shape the bacterial microbiome. Shifts in the phage community could indirectly affect bacterial composition and thus ICI outcomes. For instance, a rise in phages that target beneficial bacteria might precipitate dysbiosis associated with poor ICI response or colitis. Research in this area is in early stages, and specific phage dynamics in ICI patients remain largely uncharacterized. |
[25,155,184,185] |
Table 4.
Key microbial species associated with dietary interventions that may impact ICI responses.
| Dietary Intervention | Microbial Effects | Key Refrences |
| High-fiber, plant-rich diet | Increases SCFA-producers: Faecalibacterium prausnitzii, Bifidobacterium spp., Akkermansia muciniphila, Ruminococcaceae. Reduces overgrowth of Candida albicans; supports a balanced mycobiome. | [78,186,198,199,200,201,202,203] |
| Mediterranean diet | Promotes microbial diversity and beneficial taxa similar to high-fiber diets. Associated with lower pro-inflammatory biomarkers; although some studies found an increase in Candida albicans growth. | [78,98,190,198,202,204,205,206,207,208] |
| Western diet | Increases bile-tolerant and potentially harmful bacteri; decreases beneficial fiber-utilizers. Fuels Candida spp. overgrowth – higher gut Candida load can drive Th17 inflammation and gut toxicity. | [69,189,190,191,202,209,210,211,212] |
| Ketogenic / very low-carb diet | May reduce abundance of fiber-dependent commensal; some increase in protein/fat metabolizers, increases total anaerobic microflora and counts of Bacteroides. Starves Candida of carbs. | [202,213,214,215] |
| Intermittent Fasting / Caloric restriction | Enriches beneficial taxa – fasting has been shown to increase Akkermansia muciniphila and other mucin-degraders. Also boosts Ruminococcaceae and SCFA output when refeeding occurs. Possibly favors fungal commensal balance, not a clear signification has been shown. | [98,192,193,194,212,216,217] |
Table 5.
Key microbes that are promoted by prebiotics and related supplements.
| Prebiotic/Supplement | Microbial Implications | Key Refrences |
| Inulin, FOS | ↑Bifidobacterium, ↑Lactobacillus, and ↑Faecalibacterium prausnitzii – associated with better ICI responses. | [186,219,220,227,228] |
| Resistant starch | Ruminococcus bromii, ↑Eubacterium rectale(116); ↑overall SCFA levels which enhance cytotoxic T-cell function. | [78,155,218,229] |
| Polyphenols | ↑Lactobacillus and ↑Akkermansia ↓Enterobacteriacea. These shifts correlate with reduced tumor growth in preclinical studies. | [69,202,203,221,230] |
| Vitamin D | ↑Lachnospiraceae and ↑Ruminococcaceae after supplementation. Also associated with ↑Prevotella and ↑Alistipes in some studies – these genera have been linked to responders. | [78,100,226] |
| Butyrate | Directly provides SCFA to host – can compensate for lack of Faecalibacterium/Roseburia. Butyrate in the colon expands Tregs but also can boost CD8⁺ T-cell memory. | [69,155,231,232] |
| β-glucan | Not a prebiotic but a biotic that activates immunity. Still, it may subtly shift microbiota by enhancing pathogen clearance. In mice it can increase Bacteroidetes and decrease Proteobacteria. | [148,149,150,151,152,155,224] |
Table 6.
Examples of probiotic organisms and their potential relevance alongside immunotherapy treatment.
Table 6.
Examples of probiotic organisms and their potential relevance alongside immunotherapy treatment.
| Probiotic Organism | Relevance to ICI Outcomes | Key Refrences |
| Akkermansia muciniphila | Enriched in responders. Stimulates mucosal immunity and dendritic cell activation. Under development as a therapeutic. May not help if patient already has high baseline Akkermansia (excess could erode mucus). | [24,95,98,117,239,240] |
| Faecalibacterium prausnitzii | Associated with favorable outcomes. Produces butyrate, fostering regulatory immune balance and potentially enhancing CD8 T-cell function via metabolic support. Strict anaerobe, challenging to deliver alive. Being formulated in some consortia. Might protect against colitis. | [186] |
| Bifidobacterium longum and B. breve | Shown to enhance anti–PD-L1 therapy in mice [21] by increasing dendritic cell activity and CD8 infiltration. Some human data suggest higher bifido is beneficial. Available in some supplements, but typical doses may not colonize effectively in adults. Next-gen products might include specific immunostimulatory bifido strains. | [98,100,101] |
| Bacteroides fragilis | Required for CTLA-4 response in preclinical models (induces Th1 and maturation of dendritic cells in lymph nodes). Some responder microbiomes are high in Bacteroides. Could be part of a cocktail; mono-strain probiotic use would need caution as Bacteroides can cause infection in frail patients – hence likely delivered as part of a controlled mix. | [105,155] |
| Saccharomyces boulardii | Not yet studied in ICI patients but widely used to prevent antibiotic-related dysbiosis. Produces factors that inhibit pathogens and possibly modulate macrophage cytokines. Engineered Saccharomyces cerevisiae var. boulardii probiotics that secrete miniature anti-PD-L1 checkpoint proteins can be orally delivered to gastrointestinal tumors, where they reduced tumor burden and reshaped immune and microbiome profiles in an ICI-refractory colorectal cancer mouse model, demonstrating a modular, customizable platform for targeted GI cancer therapy. Generally safe except in critically ill. Could be given during antibiotic courses to preserve microbiome and prevent Candida overgrowth. | [236,237,238] |
Table 7.
Key species involved in FMT success in the context of immunotherapy.
| FMT Donor Microbes (examples) | Role in Recipient after FMT | Key Refrences |
| Akkermansia muciniphila | Engrafts and increases mucosal immunogenicity, linked to improved T-cell responses and sensitivity to PD-1 blockade. In responder-to-patient FMT, Akkermansia often blooms in recipients whose baseline microbiome lacked it. Its presence post-FMT associated with re-sensitization to therapy. | [24,27,94,95,242,245,247] |
| Faecalibacterium prausnitzii | Drives anti-inflammatory milieu in gut, yet systemically its metabolite butyrate can boost CD8 T cells. Post-FMT increase of Faecalibacterium correlated with reduced colitis and possibly better tumor control. Key butyrate producer; often depleted in antibiotic-treated or ICI-failing patients and restored by FMT. It also indicates overall gut health. | [25,26,242] |
| Ruminococcaceae spp. | Contribute to sustained engraftment of a diverse anaerobic community; produce SCFAs and modulate Tregs/Th17 balance. Higher Ruminococcus post-FMT was observed in responders. Broadly reflects restoration of diversity. Some specific members might enhance antigen presentation indirectly via metabolite cross-talk. | [27,242,247] |
| Bacteroides spp. | Some donors have high Bacteroides; these can take hold in recipients and have been linked to favorable immune changes. Bacteroides can also help exclude enteropathogens. Need to monitor, as overabundance might cause diarrhea in some cases. | [242,247] |
| Overall diversity | The cumulative effect of FMT is to greatly expand microbial diversity in the recipient. High diversity is consistently associated with better ICI outcome. Diversity is likely to ensure multiple pathways of immune stimulation are intact. FMT donors tend to have richer microbiomes than cancer patients who have been heavily treated with antibiotics. | [27,242,248] |
| Candida spp. | Interesting case: FMT also transfers the donor’s mycobiome. C. albicans was detected in donors and subsequently in the patients – but the patients responded to therapy. The role of fungal transfer is unclear; ideally, donors lack overt fungal overgrowth. Donor screening currently focuses on bacteria, but perhaps selecting donors with balanced fungal communities could be beneficial. The ideal FMT would correct bacterial and fungal dysbiosis together. | [26,27,156] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



