Submitted:
02 January 2026
Posted:
06 January 2026
You are already at the latest version
Abstract
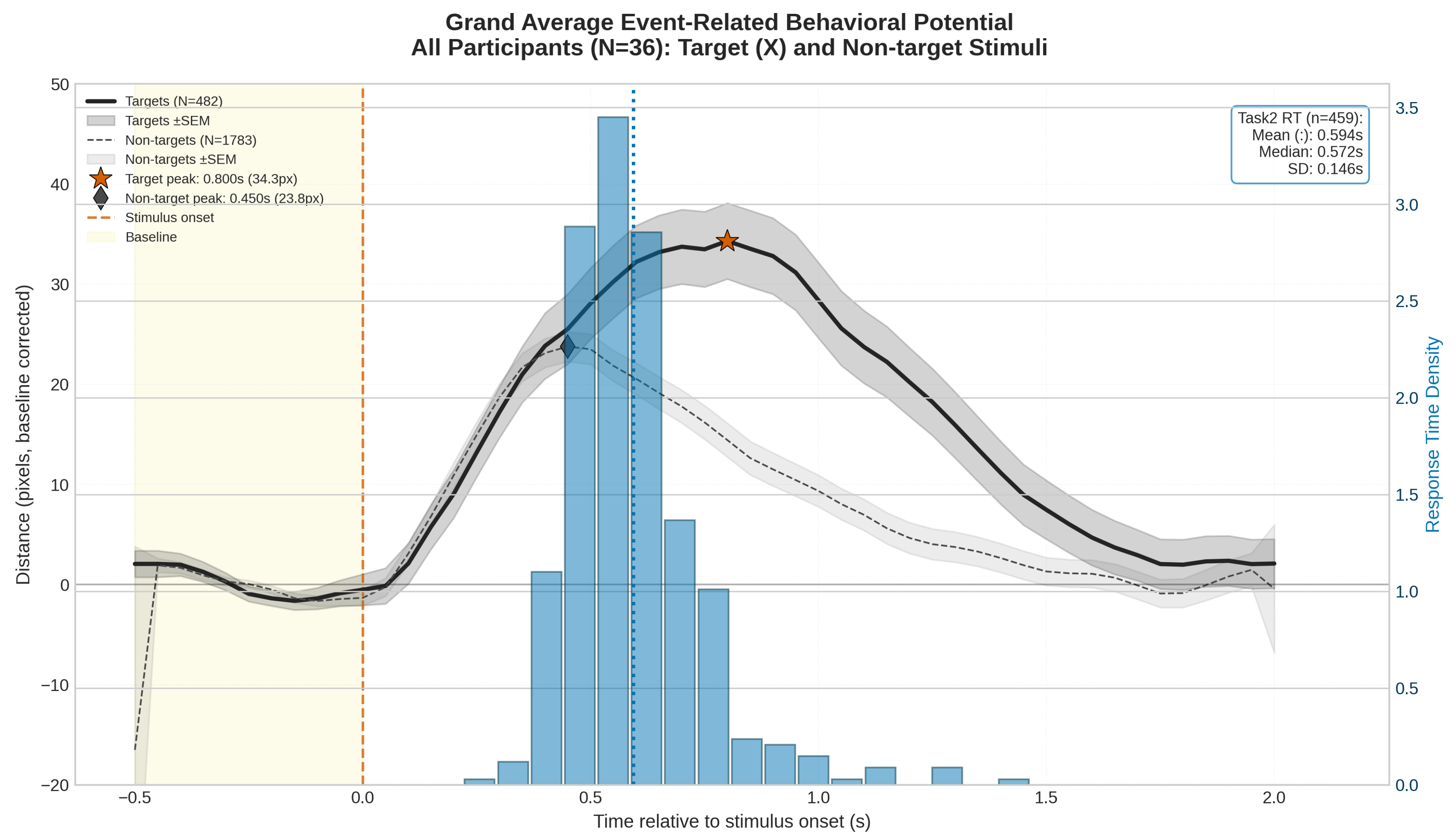
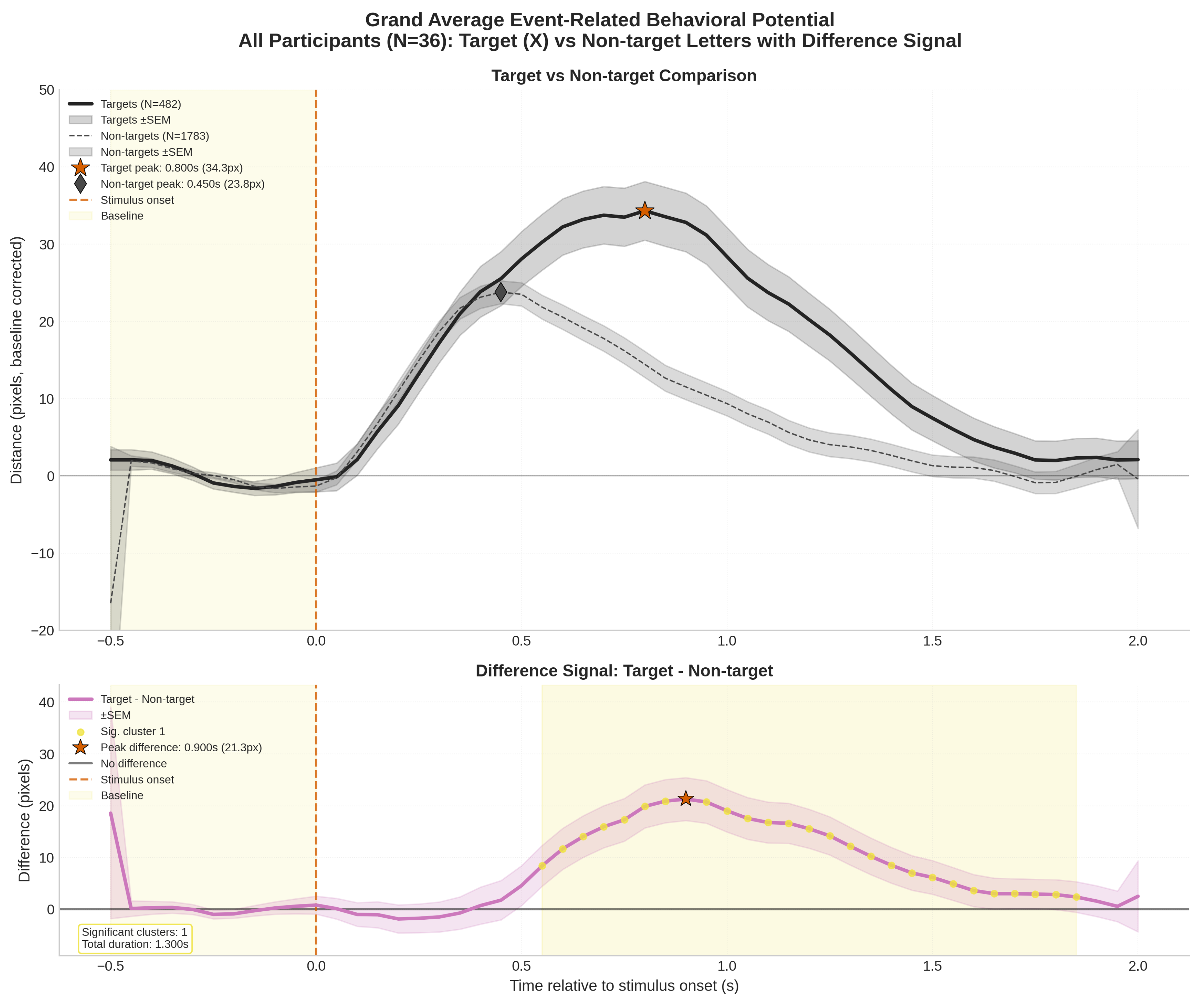
High-performance athletes operate in demanding environments requiring simultaneous coordination of multiple cognitive and motor tasks. This study developed a novel dual-task protocol combining continuous visuomotor tracking with discrete attentional vigilance to investigate temporal dynamics of dual-task interference in young athletes. Thirty-six participants from interceptive and static sports performed the dual-task paradigm while behavioral performance metrics were continuously recorded. Adapting event-related potential methodology to behavioral data, we computed Event-Related Behavioral Potentials (ERBPs) to characterize time-locked performance changes. Results revealed a significant Dual-Task Effect (DTE) with distinct temporal components: an early perceptual interference phase around 450 ms post-stimulus and a later decision-execution phase extending to 1400 ms. Friedman tests confirmed significant performance differences across temporal windows (\( \chi^2 \)(4) = 85.32, p < 0.001), with performance returning to baseline by 1500 ms. The ERBP analysis enabled quantification of DTE amplitude, latency, and duration—providing novel metrics for continuous assessment of cognitive-motor interference. Target events elicited pronounced performance degradation compared to non-target events (peak difference: 10.5 px, latency difference: 350 ms), indicating sensitivity to decision-making processes beyond motor execution. Exploratory comparisons between sport groups revealed trends suggesting differential interference patterns, though no significant between-group differences emerged. These findings demonstrate that ERBP analysis offers a powerful framework for dissecting temporal dynamics of dual-task performance, with implications for understanding attentional resource allocation in high-demand environments and potential applications in sports training and cognitive assessment.
Keywords:
1. Introduction
2. Materials and Methods
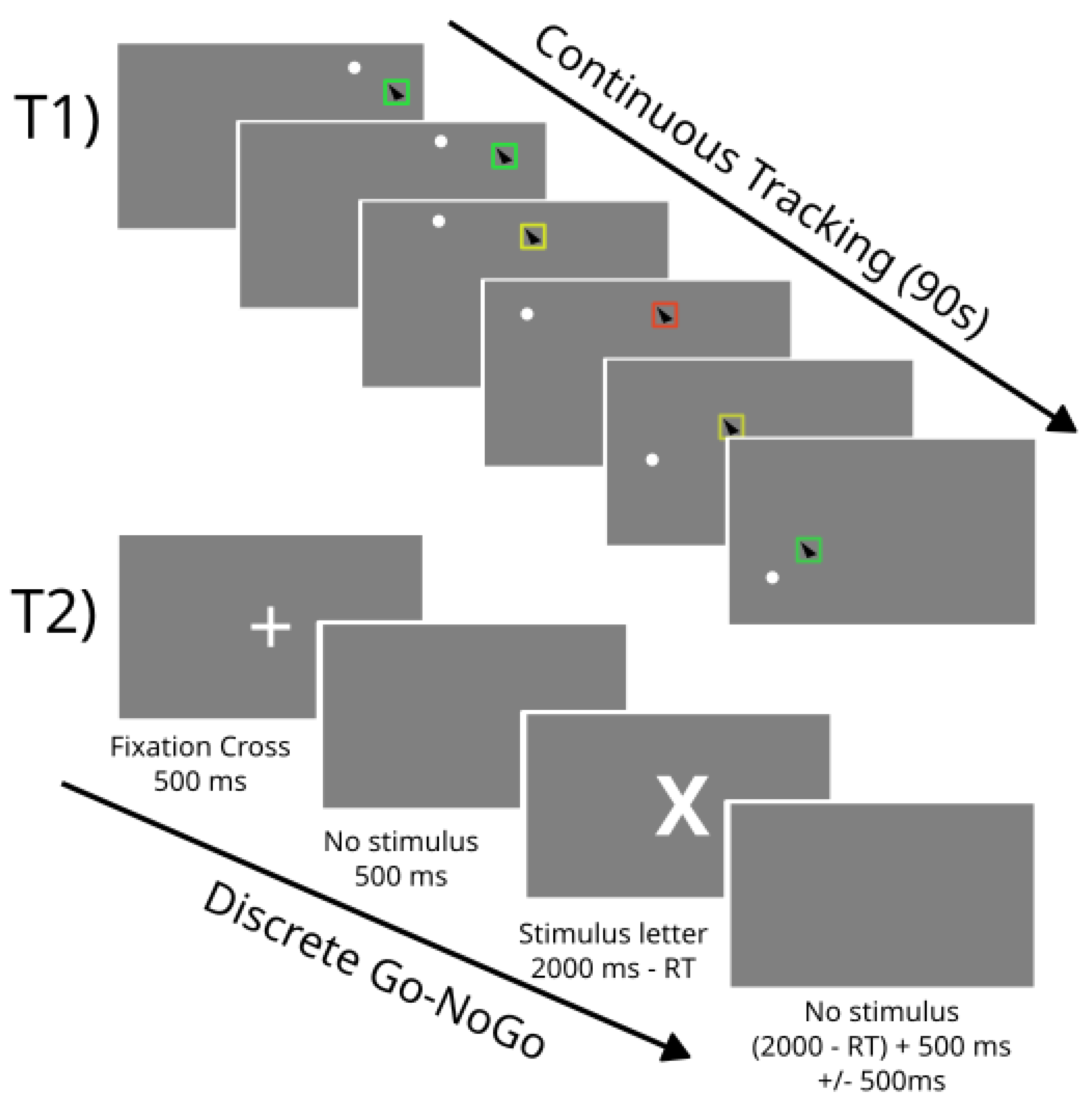
3. Results
3.1. Event Related Behavioral Potentials
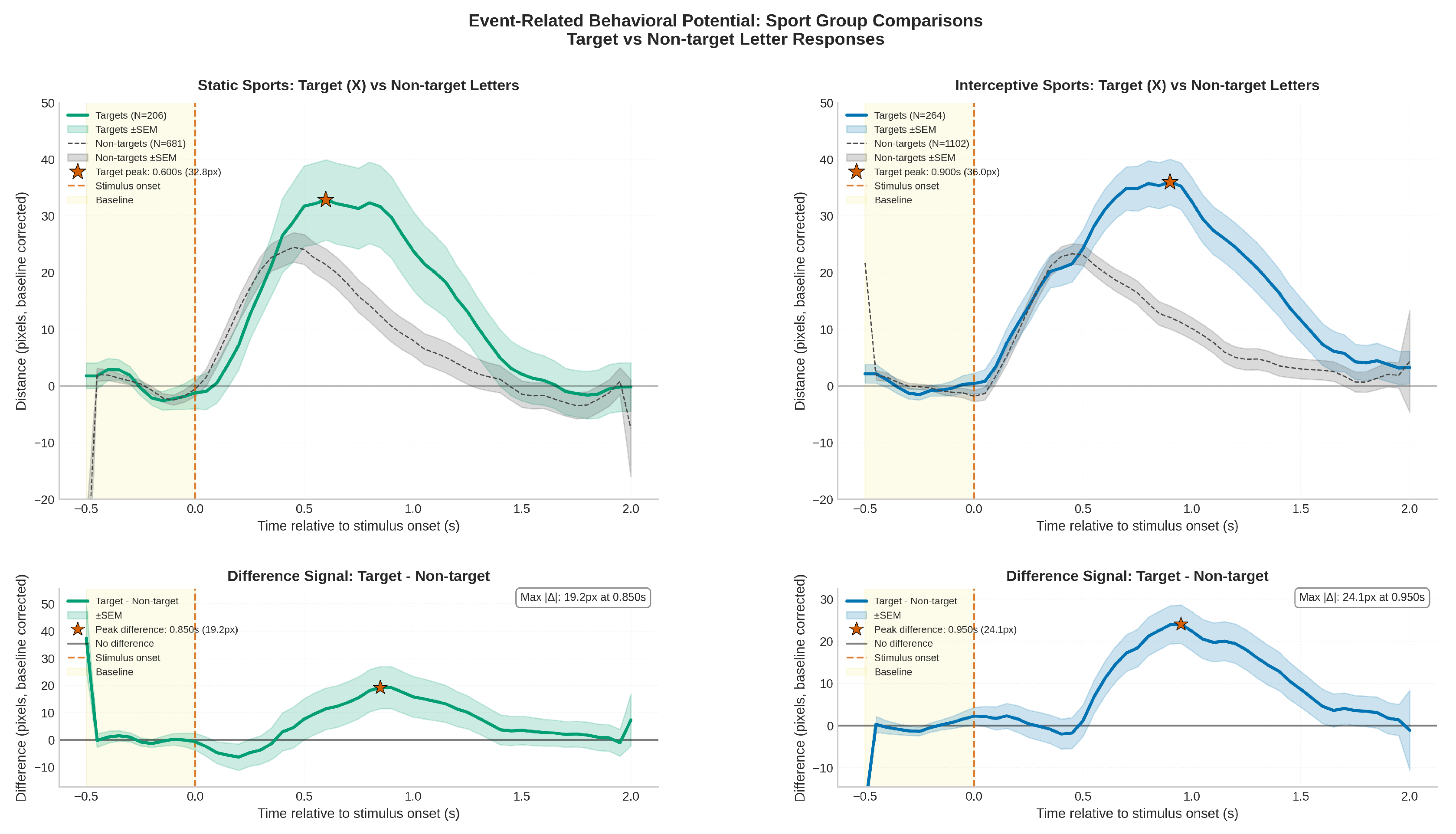
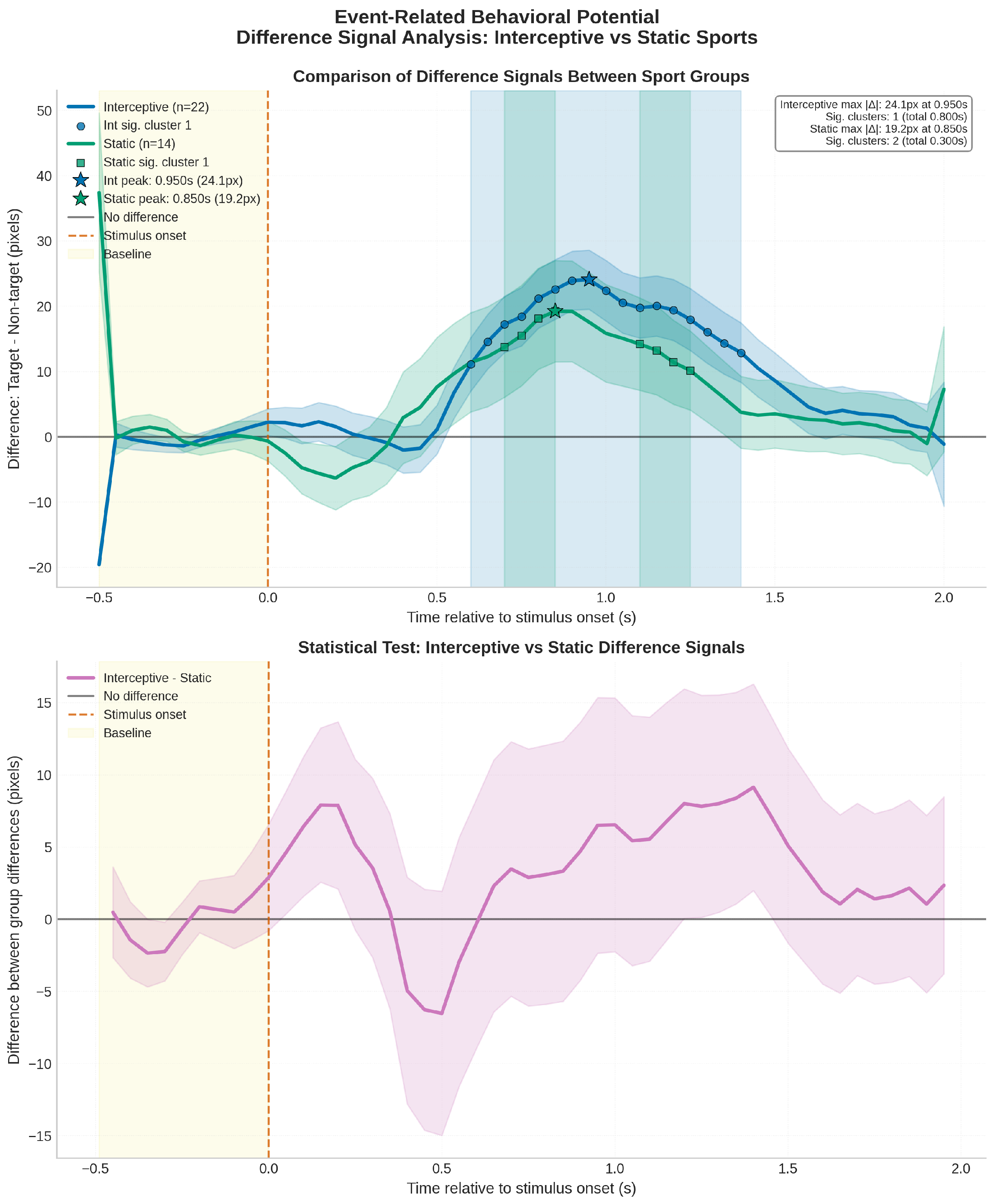
3.2. Exploratory analysis: Sport Group Comparison
4. Discussion
4.1. Temporal Dynamics of Dual-Task Interference
4.2. Adaptation Through Resource Redistribution
4.3. Continuous Assessment of DTE
4.4. Aligment with Multitasking Models
4.5. Limitations
4.6. Outlook
5. Conclusions
Supplementary Materials
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez Rosales, A.d.J.; Morquecho Méndez, A.; Ródenas Cuenca, L.T. Memoria de trabajo y control inhibitorio en beisbolistas universitarios. Retos: nuevas tendencias en educación física, deporte y recreación 2021, 939–946. [Google Scholar]
- Jacobson, J.; Matthaeus, L. Athletics and Executive Functioning: How Athletic Participation and Sport Type Correlate with Cognitive Performance. Psychology of Sport and Exercise 2014, 15, 521–527. [Google Scholar] [CrossRef]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cognitive Psychology 2000, 41, 49–100. [Google Scholar] [CrossRef]
- Vestberg, T.; Reinebo, G.; Maurex, L.; Ingvar, M.; Petrovic, P. Core Executive Functions Are Associated with Success in Young Elite Soccer Players. PLOS ONE 2017, 12, e0170845. [Google Scholar] [CrossRef]
- Yu, M.; Liu, Y. Differences in Executive Function of the Attention Network between Athletes from Interceptive and Strategic Sports. Journal of Motor Behavior 2021, 53, 419–430, [32654658. [Google Scholar] [CrossRef] [PubMed]
- Ayoubi, S.; Behgam, N.; Sadeghi-Bahmani, D.; Heidari, H.; Maghbooli, Z.; Eskandarieh, S. Response Inhibition, Attention and Processing Speed in Male Athlete and Non-athlete Adolescents. Caspian Journal of Neurological Sciences 2024, 10, 306–311. [Google Scholar] [CrossRef]
- Logan, N.E.; Henry, D.A.; Hillman, C.H.; Kramer, A.F. Trained Athletes and Cognitive Function: A Systematic Review and Meta-Analysis. International Journal of Sport and Exercise Psychology 2023, 21, 725–749. [Google Scholar] [CrossRef]
- Ishak, M.A.; Kassim, A.F.M.; Miswan, M.S.; Zainuddin, N.F. The Comparison of Decision-Making Skills between Athletes and Non-athletes among University Students. Journal of Human Centered Technology 2023, 2, 73–77. [Google Scholar] [CrossRef]
- Yongtawee, A.; Park, J.; Kim, Y.; Woo, M. Athletes Have Different Dominant Cognitive Functions Depending on Type of Sport. International Journal of Sport and Exercise Psychology 2022, 20, 1–15. [Google Scholar] [CrossRef]
- Qizi, Y.D.I. TECHNICAL AND TACTICAL SKILLS IN SPORTS. American Journal Of Social Sciences And Humanity Research 2023, 3, 105–116. [Google Scholar] [CrossRef]
- MacPherson, S.E. Definition: Dual-tasking and Multitasking. Cortex 2018, 106, 313–314. [Google Scholar] [CrossRef] [PubMed]
- Pashler, H. Dual-Task Interference in Simple Tasks: Data and Theory. Psychological Bulletin 1994, 116, 220–244. [Google Scholar] [CrossRef]
- Plummer, P.; Eskes, G. Measuring Treatment Effects on Dual-Task Performance: A Framework for Research and Clinical Practice. Frontiers in Human Neuroscience 2015, 9. [Google Scholar] [CrossRef]
- Künstler, E.C.S.; Penning, M.D.; Napiórkowski, N.; Klingner, C.M.; Witte, O.W.; Müller, H.J.; Bublak, P.; Finke, K. Dual Task Effects on Visual Attention Capacity in Normal Aging. Frontiers in Psychology 2018, 9. [Google Scholar] [CrossRef]
- Strobach, T. Cognitive Control and Meta-Control in Dual-Task Coordination. Psychonomic Bulletin & Review 2024, 31, 1445–1460. [Google Scholar] [CrossRef]
- Sigman, M.; Dehaene, S. Brain Mechanisms of Serial and Parallel Processing during Dual-Task Performance. Journal of Neuroscience 2008, 28, 7585–7598, [18650336. [Google Scholar] [CrossRef]
- Sánchez-Cubillo, I.; Periáñez, J.A.; Adrover-Roig, D.; Rodríguez-Sánchez, J.M.; Ríos-Lago, M.; Tirapu, J.; Barceló, F. Construct Validity of the Trail Making Test: Role of Task-Switching, Working Memory, Inhibition/Interference Control, and Visuomotor Abilities. Journal of the International Neuropsychological Society 2009, 15, 438–450. [Google Scholar] [CrossRef]
- Tokgöz, O.; Korkmaz, N.; Pancar, S. The Comparison of Trail Making Test Scores of Open and Closed Skill Sports Athletes. The Journal of Eurasia Sport Sciences and Medicine 2021, 3, 33–40. [Google Scholar]
- Holfelder, B.; Klotzbier, T.J.; Eisele, M.; Schott, N. Hot and Cool Executive Function in Elite- and Amateur- Adolescent Athletes From Open and Closed Skills Sports. Frontiers in Psychology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Klotzbier, T.J.; Schott, N. Skillful and Strategic Navigation in Soccer – a Motor-Cognitive Dual-Task Approach for the Evaluation of a Dribbling Task under Different Cognitive Load Conditions. Frontiers in Psychology 2024, 15. [Google Scholar] [CrossRef]
- Sarter, M.; Givens, B.; Bruno, J.P. The Cognitive Neuroscience of Sustained Attention: Where Top-down Meets Bottom-Up. Brain Research Reviews 2001, 35, 146–160. [Google Scholar] [CrossRef]
- Scolari, M.; Byers, A.; Serences, J.T. Optimal Deployment of Attentional Gain during Fine Discriminations. Journal of Neuroscience 2012, 32, 7723–7733, [22649250. [Google Scholar] [CrossRef]
- Kerlin, J.R.; Shahin, A.J.; Miller, L.M. Attentional Gain Control of Ongoing Cortical Speech Representations in a “Cocktail Party”. Journal of Neuroscience 2010, 30, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, T.W.; Dixon, M.L.; Anderson, A.K.; De Rosa, E. Distinguishing Attentional Gain and Tuning in Young and Older Adults. Neurobiology of Aging 2014, 35, 2514–2525. [Google Scholar] [CrossRef]
- Wulf, G. Attentional Focus and Motor Learning: A Review of 15 Years. International Review of Sport and Exercise Psychology 2013, 6, 77–104. [Google Scholar] [CrossRef]
- Koch, I.; Poljac, E.; Müller, H.; Kiesel, A. Cognitive Structure, Flexibility, and Plasticity in Human Multitasking—An Integrative Review of Dual-Task and Task-Switching Research. Psychological Bulletin 2018, 144, 557–583. [Google Scholar] [CrossRef] [PubMed]
- Boag, R.J.; Strickland, L.; Heathcote, A.; Loft, S. The Dynamics of Stability and Flexibility: How Attentional and Cognitive Control Support Multitasking under Time Pressure. Journal of Experimental Psychology: General 2025, 154, 1699–1728. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.C.; Strayer, D.L.; Matzke, D.; Heathcote, A. Cognitive Workload Measurement and Modeling under Divided Attention. Journal of Experimental Psychology: Human Perception and Performance 2019, 45, 826–839. [Google Scholar] [CrossRef]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in Behavior Made Easy. Behavior Research Methods 2019, 51, 195–203. [Google Scholar] [CrossRef]
- Reips, U.D.; Krantz, J.H. Conducting True Experiments on the Web. In Advanced Methods for Conducting Online Behavioral Research; American Psychological Association, 2010; pp. 193–216. [Google Scholar] [CrossRef]
- Luck, S.J. An Introduction to the Event-Related Potential Technique, 2 ed.; MIT Press, 2014. [Google Scholar]
- Sanchez-Lopez, J.; Silva-Pereyra, J.; Fernandez, T. Sustained Attention in Skilled and Novice Martial Arts Athletes: A Study of Event-Related Potentials and Current Sources. PeerJ 2016, 4, e1614. [Google Scholar] [CrossRef]
- Nakamoto, H.; Mori, S. Sport-Specific Decision-Making in a Go/Nogo Reaction Task: Difference among Nonathletes and Baseball and Basketball Players. Perceptual and Motor Skills 2008, 106, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.W.; Collier, S.A.; Night, J.C. Timing and Executive Resources: Dual-task Interference Patterns between Temporal Production and Shifting, Updating, and Inhibition Tasks. Journal of Experimental Psychology: Human Perception and Performance 2013, 39, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Boag, R.J.; Strickland, L.; Loft, S.; Heathcote, A. Strategic Attention and Decision Control Support Prospective Memory in a Complex Dual-Task Environment. Cognition 2019, 191, 103974. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.M.; Shaw, T.H.; Helton, W.S. Evaluating the Dual-Task Decrement within a Simulated Environment: Word Recall and Visual Search. Applied Ergonomics 2023, 106, 103861. [Google Scholar] [CrossRef]
- Asuako, P.A.G.; Stojan, R.; Bock, O.; Mack, M.; Voelcker-Rehage, C. Multitasking: Does Task-Switching Add to the Effect of Dual-Tasking on Everyday-like Driving Behavior? Cognitive Research: Principles and Implications 2025, 10, 5. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Q. Dual-Task Interference: Bottleneck Constraint or Capacity Sharing? Evidence from Automatic and Controlled Processes. Attention, Perception, & Psychophysics 2024, 86, 815–827. [Google Scholar] [CrossRef]
- Kahneman, D.; Tversky, A. On the Psychology of Prediction. Psychological Review 1973, 80, 237–251. [Google Scholar] [CrossRef]
- Strayer, D.L.; Johnston, W.A. Driven to Distraction: Dual-Task Studies of Simulated Driving and Conversing on a Cellular Telephone. Psychological Science 2001, 12, 462–466. [Google Scholar] [CrossRef]
- Strayer, D.L.; Castro, S.C.; Turrill, J.; Cooper, J.M. The Persistence of Distraction: The Hidden Costs of Intermittent Multitasking. Journal of Experimental Psychology: Applied 2022, 28, 262–282. [Google Scholar] [CrossRef]

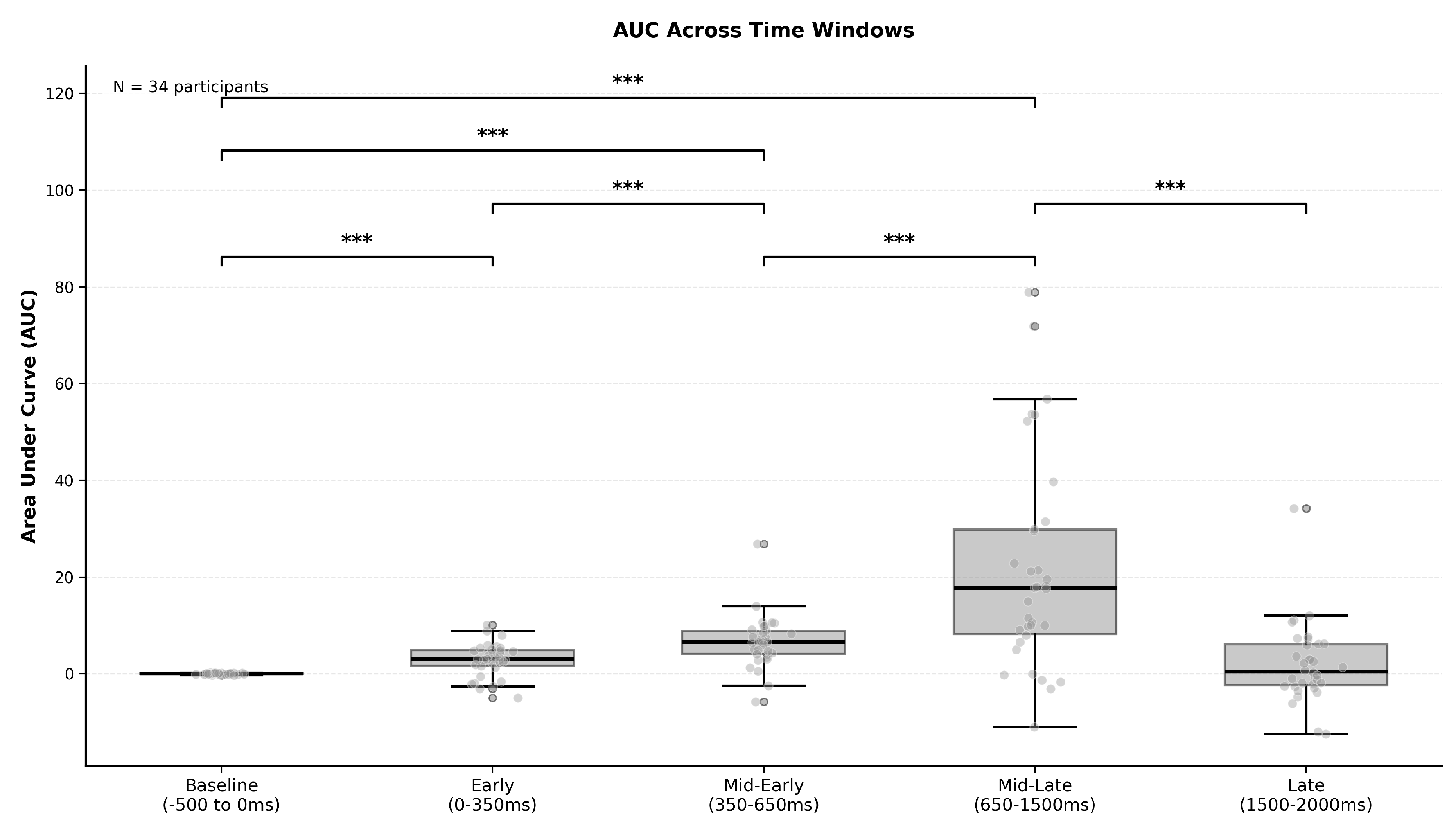
| Time Window | Mean ± SD | Median | Pairwise Comparisons | |
| (pixels) | (pixels) | Comparison | p-value (Bonf.) | |
| Baseline (-500 to 0 ms) | 0.01 ± 0.14 | 0.03 | – | – |
| Early (0–350 ms) | 2.82 ± 3.38 | 2.97 | Baseline vs Early | |
| Mid-Early (350–650 ms) | 6.62 ± 5.28 | 6.55 | Baseline vs Mid-Early | |
| Early vs Mid-Early | ||||
| Mid-Late (650–1500 ms) | 21.53 ± 21.93 | 17.73 | Baseline vs Mid-Late | |
| Mid-Early vs Mid-Late | ||||
| Late (1500–2000 ms) | 1.92 ± 8.11 | 0.46 | Baseline vs Late | |
| Mid-Late vs Late | ||||
| Overall test: Friedman (33) = 67.08, p < 0.001***, Kendall’s W = 0.493 | ||||
| Sample:N = 34 participants with complete data | ||||
| Post-hoc: 7 Wilcoxon signed-rank tests, Bonferroni-corrected = 0.0071 | ||||
| Measure | Mean | SD |
|---|---|---|
| Response Accuracy (%) | ||
| Correct Responses | 77.01 | 30.92 |
| Omissions | 22.99 | 30.92 |
| Commissions | 0.00 | 0.00 |
| Task 1: Tracking Performance | ||
| Average Distance (px) | 76.00 | 16.86 |
| Task 2: GA ERBP Measures | ||
| Reaction Time (s) | 0.594 | 0.146 |
| Peak Amplitude (px) | 97.94 | 89.14 |
| Peak Latency (s) | 0.828 | 0.489 |
| AUC Measures (px·s) | ||
| 0–350 ms | 2.89 | 14.03 |
| 350–650 ms | 7.23 | 18.80 |
| 650–1500 ms | 18.69 | 52.33 |
| 1500–2000 ms | 1.31 | 23.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



