
Submitted:
31 December 2025
Posted:
02 January 2026
You are already at the latest version
Abstract
Frontotemporal lobar degeneration (FTD) is a proteinopathy that induces neuroinflammation and neurodegeneration; Alzheimer´s disease (AD) is characterized by Abeta-42 deposits, microglia overactivation, astroglial alterations and p-Tau accumulation. Identification of neuroinflammatory mediators as predictors of cognitive cognition have gained attention. We compared several biomarkers in plasma as predictors of cognitive impairment between AD and FTD patients (Nfl, p-Tau217, TDP-43 and CX3CR1 and soluble fractalkine levels) by ELISA (pg/ml) and age-matched controls (without cognitive impairment) or HIV-1 seropositive patients. To our knowledge, this is the first study showing that increased plasma CX3CR1 and soluble fractalkine predict cognitive impairment specifically in FTD. In addition, high plasma p-Tau 271 levels correlate with sFK levels and their mini mental scores in FTD. Thus, fractalkine and TDP-43 are exclusive biomarkers of cognitive impairment in FTD. However, Nfl, GFAP and p-Tau271 levels did not differ between AD or FTD patients. Anatomically, we observed hippocampal involution as well as Tau deposits in human FTD postmortem brains. On the other hand, neuroinflammation contributes to dementia; and chemokines as HIV-1 co-receptors facilitate spread of HIV-1 infection inducing apoptosis in the brain. On the other hand, chemokines promote neuronal survival and regulate neuron-glia interactions. Fractalkine is a delta chemokine (also termed CX3CL1), that binds to its CX3CR1 chemokine receptor, that as a membrane-isoform can be released as a soluble form by damaged neurons. We confirmed that fractalkine prevents LPS (an inflammation inductor)-induced apoptosis by decreasing caspase-3 activation in cortical neurons at 7 DIV LPS exposure. Thus, fractalkine may play a dual role: it is associated with cognitive impairment in both FTD and AD, yet it also exerts neuroprotective effects by reducing LPS-induced neuronal apoptosis at 7 DIV.
Keywords:
CX3CR1/Fractalkine
; chemokines
; Alzheimer and Frontotemporal dementia
; neuroinflammation
; cortical neurons in vitro
1. Introduction
The most common forms of late-onset dementia are AD, and FTD- [1]. Many basic and translational studies have studied the causes, dementia-related mechanisms as well as therapeutic interventions for AD and FTD. Currently, the diagnosis is based on clinical symptoms, neuroimage and biomarkers of cerebrospinal fluid (CSF), which are costly and time demanding. The pathophysiology of dementia of either AD and FTD are broadly characterised by the aggregation of misfolded proteins, such as amyloid-β plaques (Aβ) and hyperphosphorylated Tau forming neurofibrillary tangles (NFT). Furthermore, neuroinflammation, neuronal apoptosis and disrupted synaptic communication are mechanisms that contribute to dementia [2,3].
Clinical subtypes of FTD are: behavioural or frontal variant, progressive nonfluent aphasia, semantic dementia, and logopenic primary progressive aphasia [4,5]. FTD is considered a TDP-43 proteinopathy [6]. TDP-43 protein plays homeostatic roles such as RNA/DNA-binding protein underlying RNA metabolism that can aggregate in neuronal and glial cell in the cytoplasm [7]. Accumulation of Aβ, NFT, or glial fibrillary acidic protein (GFAP) occur in rodent models of AD as well as in AD patients [8,9,10,11,12]. The combination of neuropsychological tests and these biomarkers allow to identify the onset of cognitive decline in dementia (e.g., cortical vs. subcortical dementias) [13]. In this way, Neurofilament M (Nfl) as biomarkers of axonal damage is detected either in blood/erebrospinal fluid (CSF) as well as in altered neuroglial processes [10,11,12]. Nfl increases progressively with age and its affected by subclinical brain injuries and comorbidities [14].
On the other hand, the presence of Tau has been detected in FTD neurofibrillary lesions, cortico-basal degeneration and progressive supranuclear palsy [15,16]. Thus, p-Tau217 reflects peri-plaque synaptic pathology at both pre- and post-synaptic sites surrounding Aβ deposits [17,18]. However, this relationship is not general, since early astroglial alterations, determined by GFAP, may arise first in AD, particularly in the entorhinal cortex [9,10,11,12].
The identification of neuroinflammatory mediators in AD or FTD have gained attention for their potential in early detection and monitoring dementia progression [19]. Since inflammation contributes to dementia, including chemokines. Furthermore, another potential biomarker studied in this study is the CX3CR1/fractalkine axis. Fractalkine is a delta chemokine (also termed CX3CL1) released by neurons and binds to its delta chemokine receptor CX3CR1 in microglia [20]. CX3CL1/fractalkine chemokine is a neuronal transmembrane-anchored protein, which can be cleaved as a soluble isoform by metalloproteases. Fractalkine plays neuroprotective, and chemotactic functions but their roles of both isoforms in neurodegenerative pathophysiology remain elusive [21]. The study of new plasma biomarkers (CX3CR1/sFK and TDP-43) of cognitive disfunction allow to understanding the pathophysiology of dementia as key steps towards finding new and effective teraphies [22,23].
2. Aims
To compare several plasma biomarkers as possible biomarkers of cognitive dysfunction (CX3CR1/soluble fractalkine, p-Tau217, GFAP, TDP-43 and Neurofilament -Nfl- between patients with AD and FTD; comparing them to age-matched controls and HIV patients (with suppressed viral load: < 50 copies/ml) without cognitive impairment.
Once we found that sFK is elevated in dementia patients, we ascertain whether soluble fractalkine recombinant protein might regulate apoptosis caspase-3 dependent level and neuronal survival under inflammatory conditions in cortical neurons at 7 DIV exposure to LPS.
3. Materials and Methods
3.1. Clinical Selection of Participants
A total number of 53 patients with AD, and additional 53 patients with FTD with early diagnosis of disease were enrolled and range of ages between 54 and 77, including a dementia-free aged matched caucassian subjects in a similar rage of age were recruited (n=53, ages between 55 and 75 years old). Thus, we enrolled 227 patients (159 patients for dementia marker determination plus 68 patients for HIV-1 study. Since chemokines are HIV-1 co-receptors, we also included a group of HIV-1 seropositive patients with suppressed viral load (without cognitive impairment (n=34 copies< 50 copies/ml) and their respective controls of similar ages (n=34). Cohort demographics and their mini mental scores to detect possible neuropsychological alterations are showed in Table-1.
3.2. Inclusion and Exclusion Criteria
The inclusion criteria were adults aged 54-77 years old, with at least 6 years of formal education, able to provide informed consent, and with sufficient language, visual and auditory acuity to complete cognitive testing. Patients with psychiatric or other neurological illnesses as alcohol abuse, or disabling stroke were excluded. The patients below < 55 years an anticoagulant treated patients were also excluded.
We included 159 participants plus 68 participants for HIV-1 delta chemokine determination (34 control+34 seropositives, total: 227 participants from the Spanish Hospitals (n=227) for determination by ELISA in plasma samples (TDP-43, CX3CR1, soluble fractalkine, p-Tau271, GFAP, Neurofilament M) and mini mental evaluation. All subjects were evaluated by neuropsychological test scores within the normative range given individual’s age, sex and educational background, that underwent a blood sampling within a maximum interval of 1 year and had at least one clinical follow-up visit available. All volunteers in this study provided informed consent signed by all participants in compliance with the World Medical Association Declaration of Helsinki and approved by a Ethics Committee (code HULP 1032 and PI-837). Blood was collected for measurement of biomarkers and measured by ELISA (see supplementary file of material and methods).
3.3. Neuropsychological Task: Mini mental Task
The Mini-Mental State Examination (MMSE) was used for assessing global cognitive functions and functionality in a validated Spanish population as global index of cognition [24].Higher scores reflect better performmance and low scores means a poor cognitive status in AD (n=53) or FTD patients (n=53); a group of age-matched healthy controls (HCs) underwent MMSE scores of ≥ 27/30 (n=53), which means absence of cognitive impairments.
3.4. ELISAs for Biochemical Markers
Several markers were quantified in plasma by ELISA (CX3CR1 and soluble delta fractalkine, TDP-43, p-Tau271, GFAP Nfl -M, see supplementary methods file).
4. Results
All tested biomarkers are elevated in plasma from AD or FTD (CX3CR1, sFK, TDP-43, p-Tau217, GFAP and Nfl) as compared their respective controls matched patients of similar age (p<0,05 vs cont; see table-2 with Kruskal Wallis values and pos Hoc Mann pos hoc for each marker).

Table 1.
Minimental (MMSE): Demographic and cognitive evaluation of patients.
|
Variables |
Age-matched controls |
AD patients |
FTD patients |
|
Size samples Number of women |
53 21 |
53 18 |
53 19 |
|
BMI index (Kg/m2) |
22.5 |
21.34 |
23 |
|
Education (years) |
7 |
6 |
6,4 |
|
Age of onset (years) |
- |
68 |
70 |
|
MMSE score |
29 |
15+-0,52 |
14.1+-0.46 |
Figure 1 Comparative analysis of plasma biomarkers of cognitive disfunction by ELISA in AD and FTD patients.
CX3CR1/sFK
The Kruskal Walis analysis (KW) confirmed increased CX3CR1/sFK plasma levels not only in AD but also in FTD patients as compare their respective controls (H=104,9; p < 0.001 for CX3CR1 and H=10,01, p = 0.007 for sFK). However, these delta chemokines did not differ between AD and FTD patients (p>0,05, n.s).
TDP-43
The Kruskal Walis analysis (KW) showed increased TDP-43 levels in plasma for FTD only as compare their respective controls (H=13,72, p< 0.001). This was the only marker that differ between AD and FTD participants (p>0,05 in Mann Whitney analysis).
p-Tau217, GFAP and Nfl
The Kruskal Walis analysis (KW) confirmed high p-Tau217 (H=97,3, p < 0.001), GFAP (H=93,9, p < 0.001) as well as Nfl plasma levels (H=84,2, p < 0.001) as compare their respective controls. The pos hoc analysis by Mann Whitney analysis did not show differences between AD and FTD levels (p>0.05 in all tested case, see figure-1 and table-2).
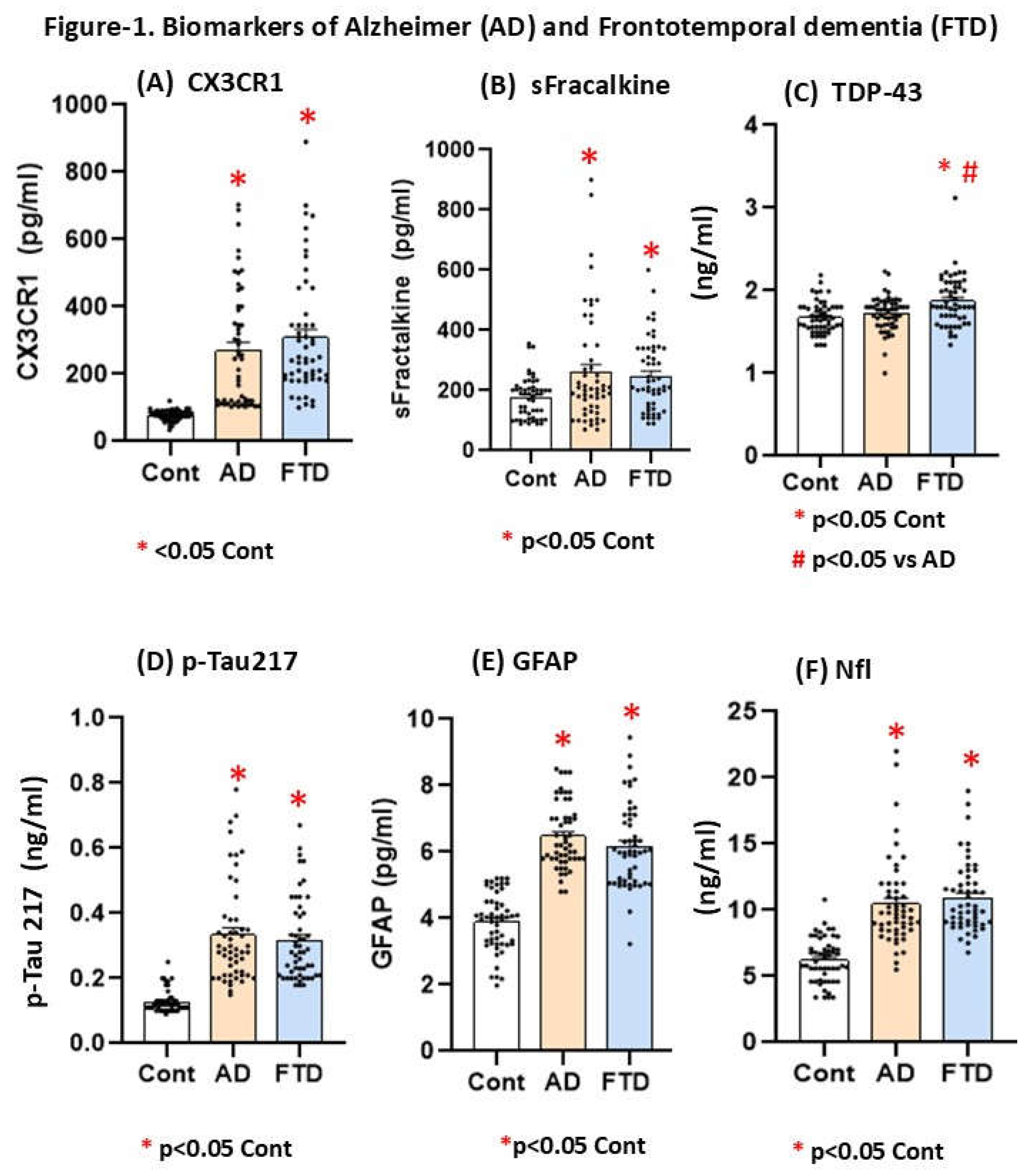
Figure 1.
A-F. Mean plasma values for +- S.E.M (Standard error media) for CX3CR1 (A: pg/ml), (B) soluble fractalkine: pg/ml), (C) TDP-43 (D) p-Tau217 (ng/ml), (E) GFAP(pg/ml), (F) (Nfl: pg/ml) in AD patients AD (70+-6,2 average of this group, n=53), FTD (72+-7 average, n=53) and age-matched controls (average age 68 years old, n=53, without dementia).The table-2 shows Kruskal Wallis values for each marker.
Figure 1.
A-F. Mean plasma values for +- S.E.M (Standard error media) for CX3CR1 (A: pg/ml), (B) soluble fractalkine: pg/ml), (C) TDP-43 (D) p-Tau217 (ng/ml), (E) GFAP(pg/ml), (F) (Nfl: pg/ml) in AD patients AD (70+-6,2 average of this group, n=53), FTD (72+-7 average, n=53) and age-matched controls (average age 68 years old, n=53, without dementia).The table-2 shows Kruskal Wallis values for each marker.
Table 2.
Statistical Kruskal Walis analysis for biomarkers in plasma.
|
Biomarker |
Kruskal Walis (KW) |
Post Hoc (Mann Whitney test) |
|
CX3CR1 |
H=104,9; p < 0.001* |
*p<0.05 vs Cont |
|
Soluble Fractalkine |
H=10,01, p = 0.007* |
*p<0.05 vs Cont |
|
TDP-43 |
H=13,72, p< 0.001* |
#p<0.05 vs AD |
|
p-tau217 |
H=97,3, p < 0.001* |
*p<0.05 vs Cont |
|
GFAP |
H=93,9, p < 0.001* |
*p<0.05 vs Cont |
|
Nfl |
H=84,2, p < 0.001* |
*p<0.05 vs Cont |
|
Seropositive patients - |
||
|
CX3CR1 |
- |
(KW) p = 0.9, n.s |
|
sFractalkine |
(KW) p = 0.99, n.s |
|
Figure 1G. CX3CR1/sFK and p-Tau217 in seropositive patients
The Mann Whitney analysis for CX3CR1/sFK plasma levels did not show differences (p=0.9, n.s for CX3CR1 and p = 0.9, n.s, for sFK) levels between HIV-1 seropositive patients (average ages: 46 years old) with supressed viral load (n=34, <50 copies) and controls of similar range of age (n=34 controls, average age: 38 years old, see figure 1 G
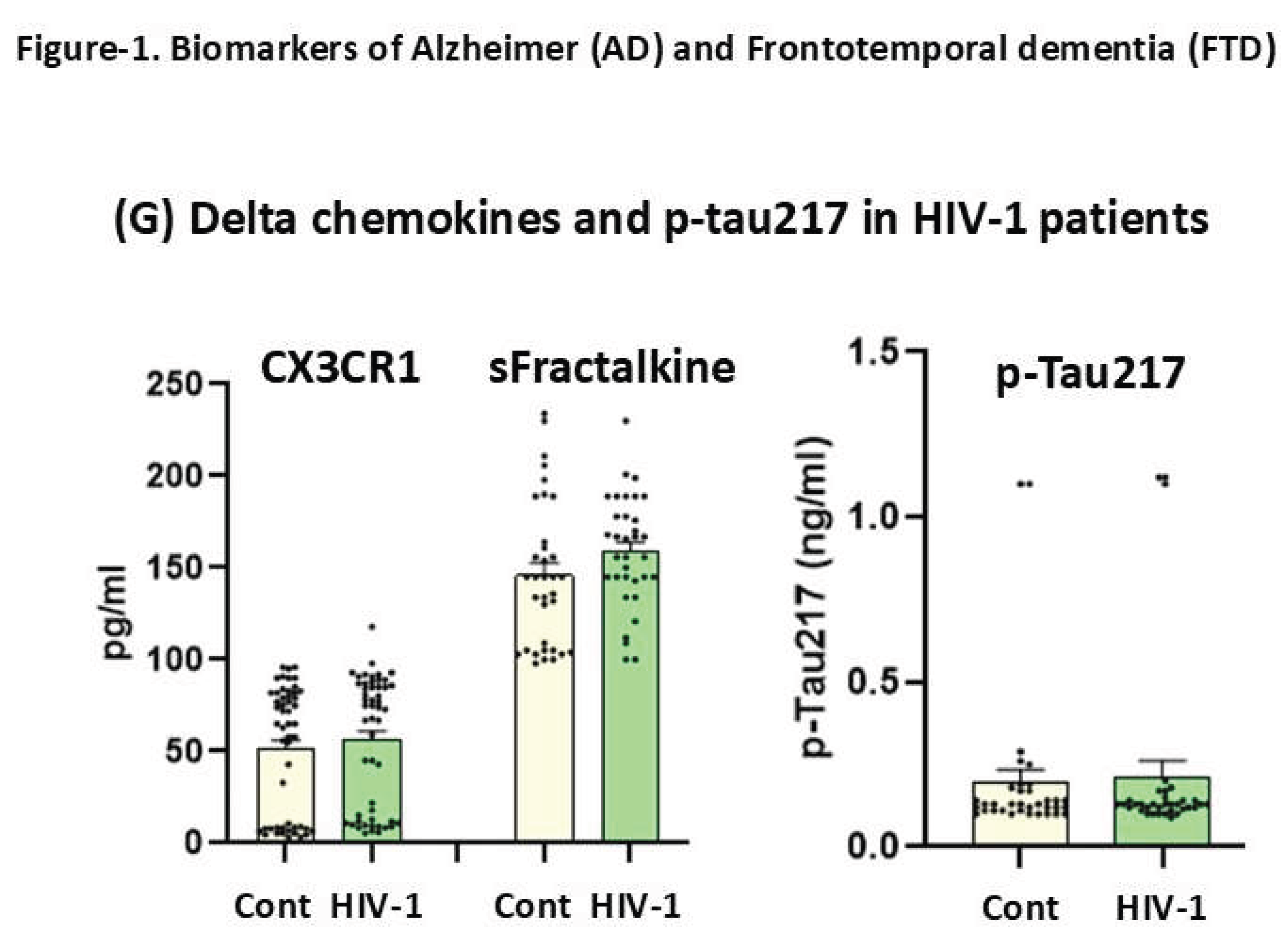
Figure 1.
(1G). Mean plasma values +- S.E.M (Standard error media) for delta chemokines (CX3CR1/sFK,) in seropositive patients with suppressed viral load (n=34: pg/ml) and age-matched controls (N= = 34, 46+/-8.2 average year old, n=34, see table-2).
Figure 1.
(1G). Mean plasma values +- S.E.M (Standard error media) for delta chemokines (CX3CR1/sFK,) in seropositive patients with suppressed viral load (n=34: pg/ml) and age-matched controls (N= = 34, 46+/-8.2 average year old, n=34, see table-2).
Correlations between Mini Mental scores (MMSS) and biomarkers of dementia in plasma
The r Spearman correlations for plasma biomarkers and mini scores were evaluated in AD (n=53) and FTD (n=53). The mini mental scores correlated not only with soluble fractalkine plasma levels (AD, r= 0.36, p=0.022) but also in FTD (TFD, r= 0.36, p=0.0083). Its CX3CR1 delta chemokine receptor correlated with mini mental scores in FTD (FTD, r = 0,39, p = 0.039); additionally, there was a r Spearman correlation between p-Tau217 in AD with CX3CR1 (AD, r = 0,29, p = 0.0038) and p-Tau217 with their mini mental scores (AD, r = 0,59, p = 0.000005). Finaly, p-Tau217 levels correlated not only with their mini mental scores in FTD (FTD, r = 0.38, p = 0.004) but also with CX3CR1 (FTD, r = 0.52, p = 0.000053).
Neuropathological alterations in human postmortem brains of AD and FTD
There are many subtypes of frontotemporal dementia (FTDP-17, alteration of cromosome 17) with different manifestations of neuropathological signs. We also detected by immunohistochemistry certain brain markers in human postmortem brains of AD and FTDP-17 to correlate them with their levels of plasma biomarkers. As in many other neurodegenerative diseases, histopathology varies in different areas of the same region. The subtypes showed positive and negative Tau pathology (Tau and phospho-Tau forms, see supplementary file of methods). There are variable accumulations of various proteins in addition to Tau in this TFD-17 pathology (alpha synuclein, TDP-43, etc) in neurons and some astrocytes, but always in smaller amounts as compared to Alzheimer's disease (AD) and other better defined tauopathies. There do not seem to be amyloid accumulations.
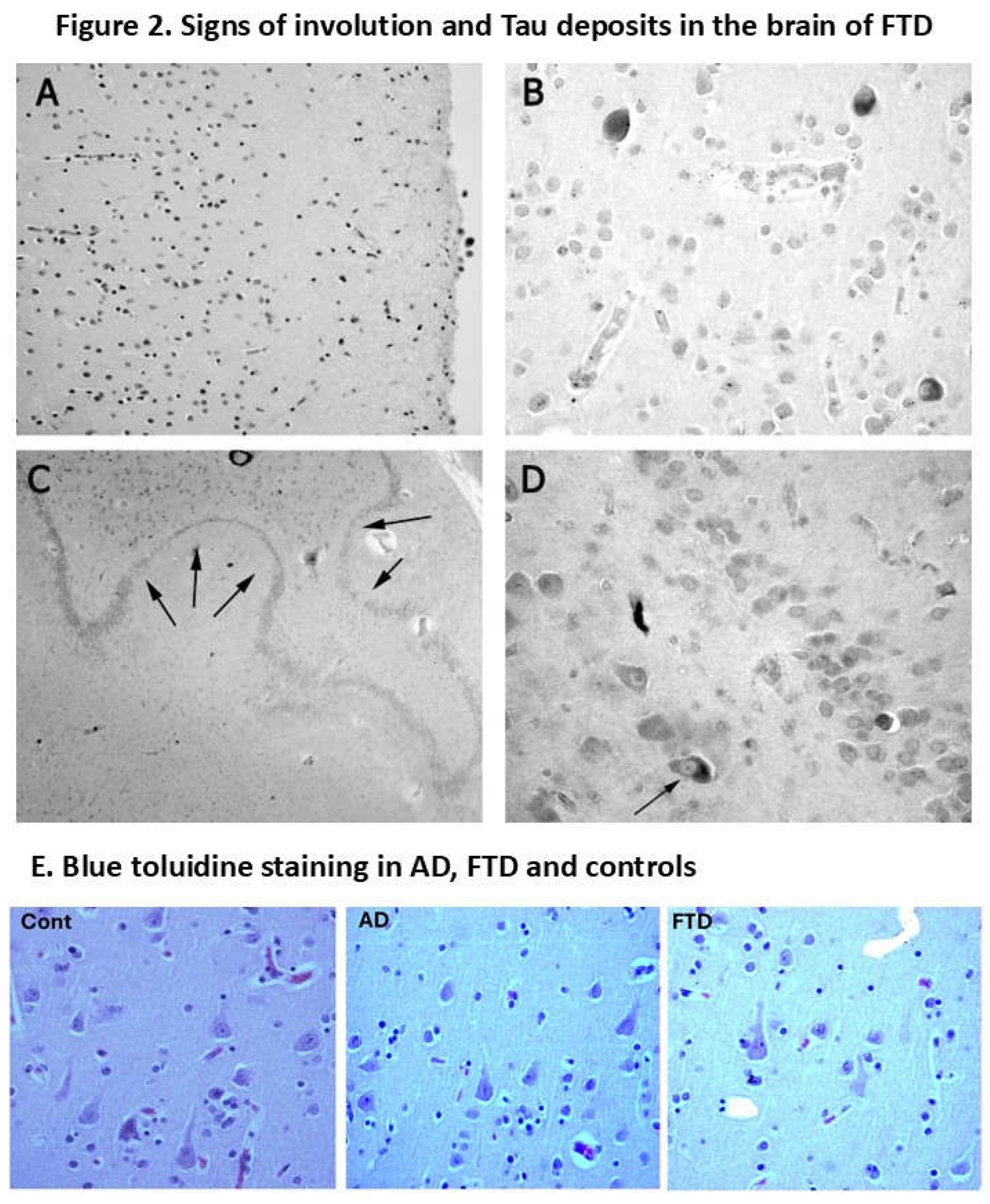
Figure 2.
Immunoreactive neurons in a case of FTD-17. (A) Frontal cortex, layers 1-3, with reduced number of pyramidal neurons. (B) Frontal cortex, layer 5, with reduced number of pyramidal neurons of intense tau immunoreactivity. More of the surviving neurons are atrophic. (C) Panoramic view of dentate gyrus (hippocampus) with areas of neurodegeneration (arrows). (D) Hippocampus. The photo indicates neurons in the CA 4 region (also termed hilus) and glial cells immunoreactive for Tau. The common features of frontotemporal dementia are frontal atrophy (presumed in 3-layer/layer-prefrontal- and 6-layer regions) and temporal atrophy (including hippocampus, subiculum and entorhinal cortex). Atrophy involves neuronal shrinkage and distortion of structures.
Figure 2.
Immunoreactive neurons in a case of FTD-17. (A) Frontal cortex, layers 1-3, with reduced number of pyramidal neurons. (B) Frontal cortex, layer 5, with reduced number of pyramidal neurons of intense tau immunoreactivity. More of the surviving neurons are atrophic. (C) Panoramic view of dentate gyrus (hippocampus) with areas of neurodegeneration (arrows). (D) Hippocampus. The photo indicates neurons in the CA 4 region (also termed hilus) and glial cells immunoreactive for Tau. The common features of frontotemporal dementia are frontal atrophy (presumed in 3-layer/layer-prefrontal- and 6-layer regions) and temporal atrophy (including hippocampus, subiculum and entorhinal cortex). Atrophy involves neuronal shrinkage and distortion of structures.
Figure-2E. Blue toluidine staining for pyramidal neurons from age-matched control of 70 years old (n=53) (C), AD (D) and FTD patients. (Left): Layer 3: normal control 70 years old (high density of pyramidal neurons. (Middle): Layer 3: AD Braak III-IV AD patient of 70 years old (High but less number pyramidal neurons than control). (Right): Layer 3: FTD-17 (very low density of pyramidal neurons).
Figure-3. Astroglial and microglial staining in the enthorrinal cortex of AD and FTD patients
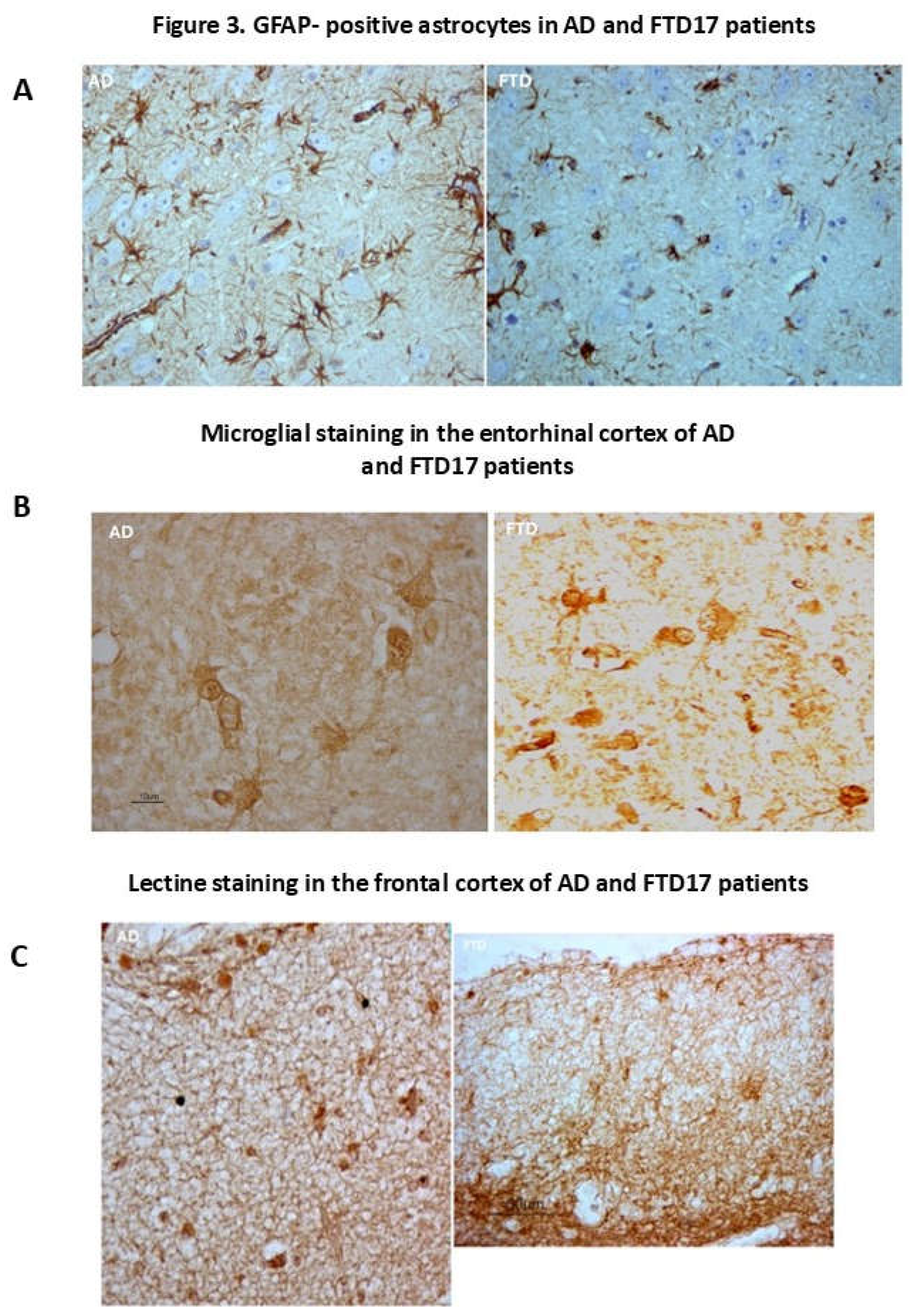
Figure 3.
Neuroglial markers in the cortex and enthorrinal cortex of AD and FTD human brain. Left (GFAP for AD). High immune GFAP-reactive astroglial cells and hypertrophic astroglial cells with long processes in the frontal cortex. (A) Rigth (GAFP for FTD-17). Most GFAP astroglial cells in the frontal cortex are of normal size, with short and thin processes on blood vessels, but a subset present hypertrophic elements (left side in this image). Glial reaction is scarce and smaller than in AD and other neurodegenerative diseases in the brain. We have observed hypertrophic astrocytes but it would be difficult to speak of astrogliosis despite higher GFAP levels found in frontotemporal dementia as compared to age-controls of similar age. It has been demonstrated the existence of extensive areas with astroglial dystrophy or asthenia or lack of GFAP-astroglia. (B) Figure 3 (left, AD). Hypertrophic microglial cells (Lectine staining) of different shape in entorhinal cortex in a case of AD (surroundig an amyloid plaque). (B) Figure 3 (right, FTD-17). Lectine stained microglial cells in the entorhinal cortex. (C) Figure 3 (left, AD). Lectine immunostaining for microglia. Intense network of microglial extension in layers 1, 2 and 3 of the frontal cortex.(C) Figure 3 (right, FTD-17). Lectine immunostaining for microglia. Intense network of microglial extension in layers 1, 2 and, in a special form, in 3 of the frontal cortex.
Figure 3.
Neuroglial markers in the cortex and enthorrinal cortex of AD and FTD human brain. Left (GFAP for AD). High immune GFAP-reactive astroglial cells and hypertrophic astroglial cells with long processes in the frontal cortex. (A) Rigth (GAFP for FTD-17). Most GFAP astroglial cells in the frontal cortex are of normal size, with short and thin processes on blood vessels, but a subset present hypertrophic elements (left side in this image). Glial reaction is scarce and smaller than in AD and other neurodegenerative diseases in the brain. We have observed hypertrophic astrocytes but it would be difficult to speak of astrogliosis despite higher GFAP levels found in frontotemporal dementia as compared to age-controls of similar age. It has been demonstrated the existence of extensive areas with astroglial dystrophy or asthenia or lack of GFAP-astroglia. (B) Figure 3 (left, AD). Hypertrophic microglial cells (Lectine staining) of different shape in entorhinal cortex in a case of AD (surroundig an amyloid plaque). (B) Figure 3 (right, FTD-17). Lectine stained microglial cells in the entorhinal cortex. (C) Figure 3 (left, AD). Lectine immunostaining for microglia. Intense network of microglial extension in layers 1, 2 and 3 of the frontal cortex.(C) Figure 3 (right, FTD-17). Lectine immunostaining for microglia. Intense network of microglial extension in layers 1, 2 and, in a special form, in 3 of the frontal cortex.
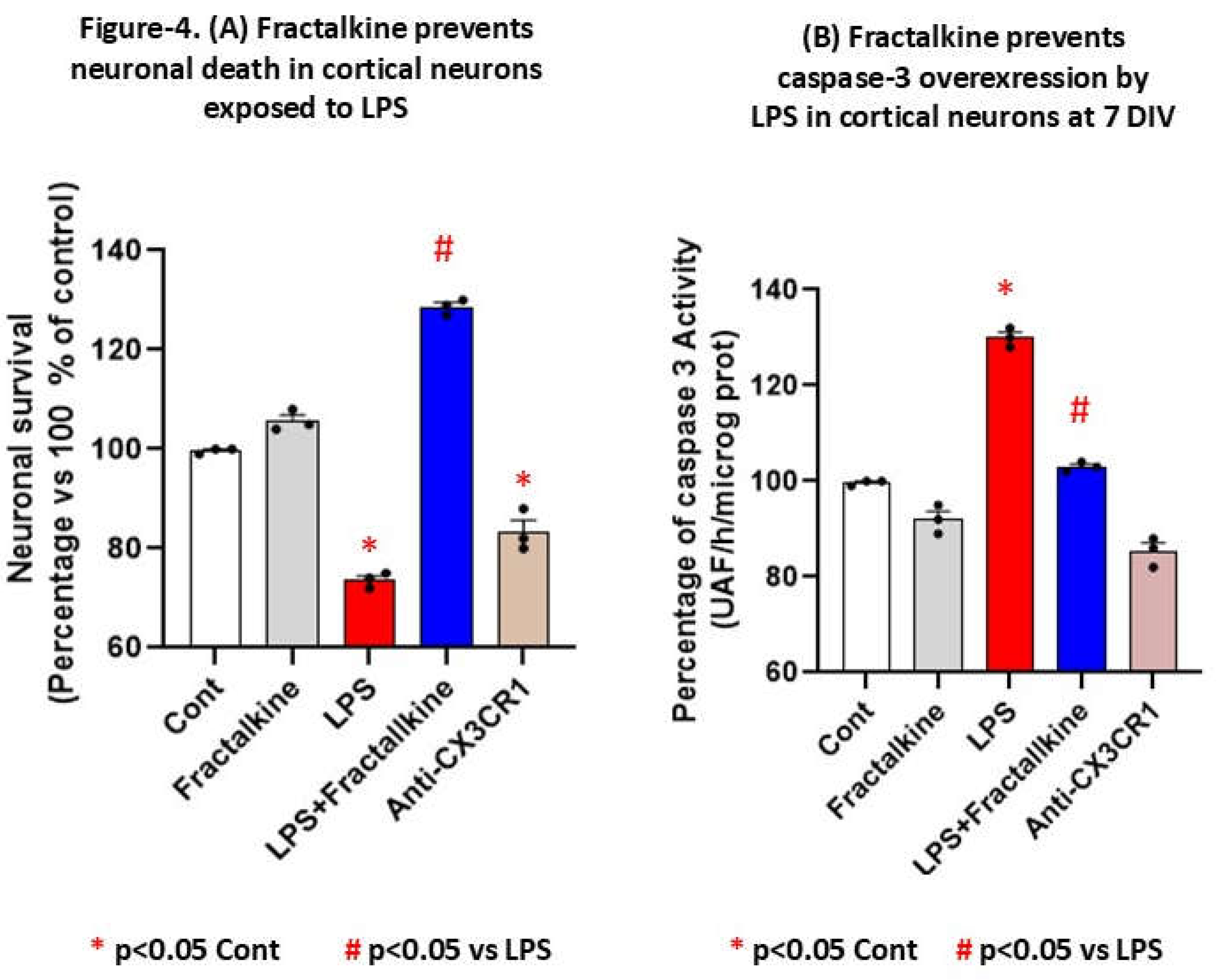
PROTOCOL FOR THE EVALUATION OF ANTIAPOPTOTIC EFFECTS OF FRACTALKINE IN CORTICAL NEURONS AT 7 DIV UNDER INFLAMMATORY CONDITIONS BY LPS TREATMENT.
In vitro protocol for LPS-induced neuroinflammation in cortical neurons at 7 DIV.
Since chemokines were upregulated in patients, we evaluated whether fractalkine could rescue or contribute to induced-apoptosis by LPS (1 µg/mL, an inductor of inflammation) treatment in cortical neurons at 7 DIV (see supplementary file of methods). For this aim, we growth rat cortical neurons at 7 DIV with/without soluble fractalkine recombinant protein (100 ng/ml, R and D company) to prevent apoptosis caspase-3 dependent mediated by LPS. Treatments with LPS take place during 24 hours (1 microg/ml, Sigma) and were started at the 7th day of culture (7 DIV) in the presence of human soluble fractalkine recombinant during 24 hour more (sFK: R & D Systems, 100 ng/ml or without sFK Thus, extract were collected at 48 hours after starting treatments for measurement of caspase-3 activity and neuronal survival were quantified by the XTT assay in all groups 24 h later, including controls (untreated) without any kind of treatment).
Figure 4(B). LPS-induced caspase-3 were prevented by fractalkine at normal levels while blockade of CX3CR1 by a specific CX3CR1 antibody promote apoptosis. This indicates feature indicates that the binding of fractalkine to neuronal CX3CR1 activates an antiapoptotic pathway that is able to counteract LPS-induced neurotoxicity. As shown in fig. 4B, the ability of fractalkine to prevent LPS-induced neuronal death was blocked by a neutralizing antibody to CX3CR1.
Statistical significance compared to controls (untreated neurons) was determined by one-way ANOVA followed by Tukey's post hoc test. * p<0.05 indicate a significant effect as compared to controls (unteated neurons at 7 DIV). # p<0.05 indicate a significant effect as compared to LPS-induced inflammation in cortical neurons at 7 DIV.
4. Discussion
In this study, we investigated whether some plasma biomarkers could distinguish AD from FTD during the prodromal or early symptomatic stages of cognitive decline. NfL is considered a biomarker of neurodegeneration. Plasma and CSF levels rise with advancing age and increase further in neurodegenerative disorders such as AD and FTD, often before cognitive symptoms [5]. However, although NfL is sensitive to neuroaxonal damage, limited disease specificity restricts its utility for differentiating dementia subtypes (https://networkinstitute.org/2020/11/23/neurofilament-light-application-online/). In our study, plasma NfL levels did not differ among AD, FTD, and cognitively unimpaired controls, consistent with its limited etiological specificity [5] and with reports of elevated NfL across neurodegenerative conditions, including increased CSF NfL in AD [27].
Clinically, serum NfL is therefore considered more informative in individuals younger than 60 years, where it can better support or exclude a neurodegenerative diagnosis [28](Willemse EAJ et al., 2021). Moreover, in cognitively healthy individuals, higher plasma NfL is positively associated with age-related comorbidities [29], which may further obscure group differences in older cohorts. [30] Preische et al. (2019) reported that longitudinal increases in serum NfL can distinguish pre-symptomatic mutation carriers from non-carriers up to 10 years before expected symptom onset in AD; these trajectories also correlated with MRI-derived cortical volume loss, supporting NfL as a marker of subclinical neuroaxonal injury prior to symptoms.
We also quantified plasma levels of p-Tau217 in patients with AD, FTD and HIV-1 seropositive patients with supressed viral load (without cognitive impairment) with the aim of evaluating their potential as systemic biomarkers of cognitive impairment, as previously suggested [31, [32]. p-Tau217 levels were elevated in plasma from AD and FTD patients compared to age-matched cognitively healthy controls and HIV-1 seropositive individuals, consistent with previous reports. Plasma p-Tau217 demonstrates diagnostic performance comparable to cerebrospinal fluid (CSF) for detecting amyloid pathology [33,34],including preclinical AD and mild cognitive impairment [35]. These CSF p-tau217 levels could distinguish with 91% of probability between AD and non-AD disorders [32], supporting pTau217 as a blood-based biomarker that predict the progression to dementia. Tauopathy rodent models of AD also demonstrate p-Tau217 accumulation, supporting its relevance across human and animal studies [36,37,38,39,40,41,42].Increased p-Tau217 has also been proposed as a biomarker in individuals with early amyloid-β accumulation [43], and several studies report an association between p-Tau217 and amyloid positron emission tomography [36]).The TRAILBLAZER-ALZ (NCT03367403) clinical trial supports the use of plasma p-Tau217 to monitor neurofibrillary tangle burden and amyloid plaque accumulation during treatment ([44,45,46,47,48]. Accordingly, the elevated plasma p-Tau217 observed in our study in AD and FTD patients is consistent with evidence that p-Tau217 reflects ongoing amyloid pathology, and may indicate increased risk of future cognitive decline [43]. Plasma p-Tau217 can distinguish amyloid-positive/tau-negative individuals from non-AD controls [49] and is associated with amyloid-β positivity in symptomatic individuals [50], while also reflecting tau tangle pathology [51]. Since in our studies p-Tau217 levels did not differ between HIV-1 seropositive individuals and cognitively unimpaired controls, the observed increases appears linked to cognitive impairment in AD and FTD. Brain cellular localization of p-Tau217 has been reported to associate with plasma p-Tau217 levels [18] , supporting for the first time our interpretation that increased plasma p-Tau217 may reflect tau isoform accumulation in FTD brain tissue. Notably, p-Tau217 deposition may help distinguish AD pathology from primary age-related tauopathy and other non-AD tauopathies [52,53].In this regard, p-Tau217 is largely undetectable in normal brain tissue but arises at both pre- and post-synaptic areas surrounding amyloid-β plaques in AD samples [17]. The elevated plasma p-Tau217 levels observed in our cohorts are consistent with its proposed role as a prognostic biomarker of cognitive impairment linked to neurodegeneration and brain atrophy in AD [50,54,55], and tau217 measured in both plasma and CSF can reliably discriminate amyloid-positive from amyloid-negative individuals [56].
A recent study indicates that plasma p-Tau217 predicts brain-wide tau accumulation during the preclinical stages of AD [57]. In line with this, we observed increased plasma p-Tau217 in FTD patients and detected tau immunoreactivity in the hippocampus of postmortem FTD brain. Histopathological analysis further revealed pronounced microglial activation together with p-Tau217 accumulation, supporting hippocampal involvement and broader regional pathology in FTD, including cortical and entorhinal areas (see Figure 3).
p-Tau217 has also been reported to correlate with poorer cognitive performance prior to the onset of explicit symptomatic disease [58]. Among amyloid-β–positive individuals, p-Tau217 predicted cognitive impairment and reduced cortical thickness with greater accuracy than p-Tau181[59,60]; by contrast, no longitudinal increase in p-Tau217 was observed in individuals with mild cognitive impairment who did not subsequently progress to clinically diagnosed AD [61].
Soluble fractalkine (sFK), or CX3CL1, was also analyzed in our studies. FK is a chemokine expressed by neurons that signals via the CX3CR1 receptor on microglia and peripheral leukocytes, playing a role in neuroimmune communication. Under pathological conditions such as inflammation or oxidative stress, FK is cleaved from the neuronal membrane and released as a soluble form (sFK), which can be neuroprotective or neurotoxic depending on context [62,42].
sFK and plasma p-Tau217 have been reported as early clinical indicators of incipient dementia in AD [63] and across other neurological conditions [54,55], [64].FK has been proposed to anticipate initial cognitive decline in AD, potentially by contributing to CNS injury and neuroinflammatory cascades [63,64] while other evidence supports a neuroprotective role of fractalkine signalling [62]. In our study, recombinant sFK attenuated caspase-3 activation in cortical neurons at 7 days in vitro following exposure to LPS, supporting a neuroprotective role under inflammatory stress [42,65].
Here, we provide first evidence that elevated peripheral sFK may serve as a predictor of cognitive dysfunction in FTD. Systemic increases in soluble fractalkine in FTD are consistent with elevated plasma fractalkine reported in AD [63]. The CX3CL1/CX3CR1 axis shows context-dependent effects: CX3CR1 deletion can mitigate neuronal loss in AD transgenic mice [66] and reduce β-amyloid deposition [67](Chen HR., et al., 2022), yet can enhance neuroinflammation [68]. Accordingly, sFK elevations may reflect a compensatory response to inflammatory stress. Peripheral monocytes can amplify neuroinflammatory responses in the brain [67], so metalloprotease-mediated FK cleavage could also exacerbate inflammation and may promote monocyte recruitment in FTD. Consistent with a protective component of this pathway, blocking CX3CR1 increased caspase-3 activation in cortical neurons in our study. Blockade of IL-1β and loss of CX3CR1 signalling enhance tau hyperphosphorylation and aggregation, accompanied by increased sFK release as a potentially compensatory response [68]. In our cohort, CX3CR1 and sFK levels did not differ between HIV-1 seropositive individuals and cognitively unimpaired controls, suggesting that elevations in this chemokine pathway are more closely linked to cognitive dysfunction in FTD. Because soluble fractalkine is produced by endothelial cells and CX3CR1 is expressed on circulating leukocytes even in healthy subjects [69], the CX3CR1/soluble fractalkine axis remains a plausible therapeutic target to modulate neuroinflammation and, indirectly, amyloid-β and tau pathology.
The combined CX3CR1/sFK axis and plasma p-Tau217 in FTD may be more closely linked to astrogliosis, microglial overactivation, and/or neuronal dysfunction in patients with mild cognitive impairment [70]. The correlation between Mini-Mental scores and plasma p-Tau271 (r = 0.4, p = 0.0025, n = 53) may reflect cognitive impairment in FTD. Lower postmortem p-Tau217 was associated with better baseline memory, whereas higher p-Tau217 at preclinical stages predicted worse cognitive trajectories [47]. FK also contributes to trafficking of HIV-infected lymphocytes into the brain in HIV-associated neurocognitive disorders [71], and because sFK is expressed by endothelial cells while CX3CR1 is expressed on leukocytes in healthy subjects, this pathway may have systemic relevance. Finally, we cannot exclude that increased CX3CR1/sFK signaling may influence hippocampal tau accumulation in FTD, given that FK overexpression suppresses tau pathology in a mouse tauopathy model [72]. In our study both AD and FTD patients showed elevated plasma levels of sFK with potential applicability in the stratification of participants for therapeutic interventions in clinical trials targeting AD or FTD pathology. Moreover, the correlation between p-Tau217 and sFK plasma levels suggests the release of sFK in response to inflammatory stimuli, as fractalkine cleavage by ADAM10 and ADAM17 has been demonstrated under conditions of inflammation [64]. This soluble fractalkine release by damaged neurons under inflammation limits microglial neurotoxicity while a sustained fractalkine release promoting neuroprotection and astroglial/microglial surveillance but also contribute to neurodegeneration [62], which agree with the antiapoptotic role of sFK in cortical neurons exposed to LPS-treatment in our in vitro study. In fact, fractalkine protects hippocampal neurons against LPS-induced neurotoxicity, an effect blocked by anti-CX3CR1 antibodies in vitro. Thus, fractalkine is neuroprotective since counteract LPS-induced caspase-3 overactivation and increases neuronal survival in cortical neurons at 7 DIV under inflammatory conditions (see figure 4). Our findings agree with the suppresses tau pathology in a mouse model of tauopathy by fractalkine overexpression [72]. Further studies will analzye a possible role of metalloprotease activation in the dual role (neuroprotective or neurotoxic) by shedding of fractalkine.
The relationship between chemokines dependent inflammation and a possible translocation of TDP-43 to the nucleus, is unknown in patients with FTD dementia. TDP-43 is a DNA/RNA-binding protein involved in RNA metabolism that form cytoplasmic aggregates in both neurons and glial cells and it is accumulated in the hippocampus.TDP-43 appears to be a more specific marker for FTD since its levels are increased in FTD patients [73,74] but are not characteristically altered in AD [75]. In our study, plasma TDP-43 levels did not change between AD patients and cognitively unimpaired controls, supporting TDP-43 as a specific plasma marker of cognitive dysfunction in our FTD cohort. Despite positron emission tomography (PET) tracers of cerebral TDP-43 do not exist, increased TDP-43 brain accumulation is consistently associated with poorer cognition. In this way, levels of sFK correlated with TDP-43 in FTD patients; In addition, TDP-43 correlated with their mini mental scores. Thus, TDP-43 may help to distinguish those cases of FTD with ubiquitin/TDP-43 pathology in their brains from those with tauopathy [76]. Thus, high TDP-43 plasma levels could identify those patients with early cognitive disfunction in FTD. In fact, plasma TDP-43 (total and/or phosphorylated TDP-43) levels correlated with TDP-43 brain pathology in FTD [77,78], suggesting its predictive value as biomarker in plasma of FTD with neurodegeneration in limbic brain regions where its accumulated.
5. Conclusions
Interpretation of short-term changes in patients with minor cognitive impairment is an emerging need for the identification of disease-modifying therapies for FTD, AD or others dementia. Given that AD begins 20 years or more before symptom onset, there is a substantial window to identify early markers of cognitive impairment; in this way, we found for the first time soluble fractalkine rises in plasma as possible predictor of cognitive impairments in FTD together; in addition, TDP-43 elevations in plasma in FTD patients distinguish between FTD and AD patients. Colectivelly, plasma pTau-217 and CX3CR1/sFK chemokines increase may prevent lumbar punctures as valid biomarkers of early cognitive disfunction; these biomarkers could identify those patients in risk of dementia that need PET scans analysis or CSF extraction for concomitant evaluation of biomarkers of dementia.
6. Limitations
Plasma biomarkers have been already included in recent recommendations from the Global CEO Initiative on AD, but they still need to be put into clinical practice to prove their usefulness levels [79]. In our study, all analysis were done in the same plate of ELISA with the same standardization protocol for all collected samples [80]. Plasma p-tau levels and other biomarkers of dementia may variate by age or sex. body mass index -BMI- [81,82] or comorbidities [83].
Author Contributions
Conceptualization, JJM, JJRA, XB, AT. Methodology: JJM (ELISAs), JJRA and AT (Neuroanatomy), formal analysis: JJM, JJRA.; investigation; JJM, XB writing—original draft preparation: JJMM, XB. and M.C.; writing—review and editing, JJM, JJRA, XB, AT supervision, JJRA, AT. All authors have read and agreed to the published version of the manuscript.
Funding
Ramon and Cajal Research program to JJM. IP RYC-2006- 002658. Project: “Role of chemokines, EPO) and stem cells as repair molecules against gp120 neurotoxicity in the hippocampus in vivo and in vitro: behavioural alterations by gp120 ¨ to JJM and SAIOTEK ¨Neurodegeneration associated to Progranuline mutations in Frontotemporal dementia in patients¨ to JJM.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki (1974 and updated in 2000) and approved by the Ethics Committee of the HULP (ID PI-837 and PI-1032).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kelliny, S.; Zhou, X.F.; Bobrovskaya, L. Alzheimer's Disease and Frontotemporal Dementia: A Review of Pathophysiology and Therapeutic Approaches. J. Neurosci. Res. 2025, 103, e70046. [Google Scholar] [CrossRef]
- Chin, K.S. Pathophysiology of dementia. Aust. J. Gen. Pract. 2023, 52, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.; Budson, A. Current understanding of Alzheimer’s disease diagnosis and treatment. F1000Res 2018, 7, F1000 Faculty Rev–1161. [Google Scholar] [CrossRef] [PubMed]
- Kirshner, HS. Frontotemporal dementia and primary progressive aphasia, a review. Neuropsychiatr Dis Treat 2014, 10, 1045–55. [Google Scholar] [CrossRef] [PubMed]
- Zetterberg, H.; Teunissen, C.; van Swieten, J.; Kuhle, J.; Boxer, A.; Rohrer, J.D.; Mitic, L.; Nicholson, A.M.; Pearlman, R.; McCaughey, S.M.; Tatton, N. The role of neurofilament light in genetic frontotemporal lobar degeneration. Brain Communications 2022, 5, fcac310. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, I.R.A.; Rademakers, R.; Neumann, M. TDP-43 and FUS in amyotrophic lateral sclerosis and frontotemporal dementia. Lancet Neurol. 2010, 9, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, X-; Wang, J.; Chen, J.; Zhao, B.; Jin, X.; Jian, g M.; Wang, J.; Liao, W.; Yang, B.; Geng, X. TDP-43: unveiling the hidden key to cellular fate decisions. Cell Commun Signal 2025, 23(1), 453. [Google Scholar] [CrossRef]
- Jiao, B.; Ouyang, Z.; Liu, Y.; Zhang, C.; Xu, T.; Yang, Q.; Zhang, S.; Zhu, Y.; Wan, M.; Xiao, X.; Liu, X.; Zhou, Y.; Liao, X.; Zhang, W.; Luo, S.; Tang, B.; Shen, L. Evaluating the diagnostic performance of six plasma biomarkers for Alzheimer’s disease and other neurodegenerative dementias in a large Chinese cohort. Alzheimers Res. Ther. 2025, 17, 71. [Google Scholar] [CrossRef]
- Rodríguez, J.J.; Zallo, F.; Gardenal, E.; Cabot, J.; Busquets, X. Prominent and Conspicuous Astrocyte Atrophy in Human Sporadic and Familial Alzheimer’s Disease. Brain Struct. Funct. 2023, 228, 2103–2113. [Google Scholar] [CrossRef]
- Rodríguez, J. J.; Gardenal, E.; Zallo, F.; Cabot, J.; Busquets, X. “Early PSA-NCAM reduction in the dentate gyrus and impaired plasticity inthe Alzheimer´s disease 3xTg-mice model”. Acta Histochemica 126(8), 152194. [CrossRef]
- Rodríguez, J. J., S. Terzieva.; E. Gardenal.; F. Zallo.;, C.Y. Yeh.; A. Verkhratsky.; A. Arrue:; Joan Cabot.; and X. Busquets. “Astrocyte S100β expression and selective differentiation to GFAP and GS in the entorhinal cortex during ageing in the 3xTg-Alzheimer´s disease mouse model”. Acta Histochemica. 2024, (126), 152131. [CrossRef] [PubMed]
- Rodríguez, J.J.; Zallo, F.; Gardenal, E.; Cabot, J.; Busquets, X. Entorhinal Cortex Astrocytic Atrophy in Human Frontotemporal Dementia. Brain Struct. Funct. 2024, 229, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Bondi, M.W.; Edmonds, E.C.; Salmon, D.P. Alzheimer's Disease: Past, Present, and Future. J Int Neuropsychol Soc. 2017, 23(9-10), 818–831. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gaur, A.; Gallagher, D.; Herrmann, N.; Chen, J.J.; Marzolini, S.; Oh, P.; Amemiya, Y.; Seth, A.; Kiss, A.; Lanctôt, K.L. Neurofilament Light Chain as a Biomarker of Global Cognition in Individuals With Possible Vascular Mild Cognitive Impairment. J. Geriatr. Psychiatry Neurol. 2025, 38, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.M.; Goedert, M; Trojanowski, JQ. Neurodegenerative tauopathies. Annu Rev Neurosci 2001, 24, 1121–59. [Google Scholar] [CrossRef]
- Creekmore, B.C.; Watanabe, R.; Lee, E.B. Neurodegenerative Disease Tauopathies. Annu. Rev. Pathol. 2023, 19, 345–370. [Google Scholar] [CrossRef]
- Hirota, Y.; Sakakibara, Y.; Ibaraki, K.; Takei, K.; Iijima, K.M.; Sekiya, M. Distinct Brain Pathologies Associated with Alzheimer’s Disease Biomarker-Related Phospho-Tau 181 and Phospho-Tau 217 in App Knock-in Mouse Models of Amyloid-β Amyloidosis. Brain Commun. 2022, 4, fcac286. [Google Scholar] [CrossRef]
- Wennström, M.; Janelidze, S.; Nilsson, K.P.R.; The Netherlands Brain Bank; Serrano, G.E.; Beach, T.G.; Dage, J.L.; Hansson, O. Cellular Localization of P-Tau217 in Brain and Its Association with p-Tau217 Plasma Levels. Acta Neuropathol. Commun. 2022, 10, 3. [CrossRef]
- Roveta, F.; Bonino, L.; Piella, E.M.; Rainero, I.; Rubino, E. Neuroinflammatory Biomarkers in Alzheimer's Disease: From Pathophysiology to Clinical Implications. Int J Mol Sci. 2024, 6;25(22), 11941. [Google Scholar] [CrossRef]
- Noda, M.; Doi, Y.; Liang, J.; Kawanokuchi, J.; Sonobe, Y.; Takeuchi, H.; Mizuno, T.; Suzumura, A. Fractalkine attenuates excito-neurotoxicity via microglial clearance of damaged neurons and antioxidant enzyme heme oxygenase-1 expression. J. Biol. Chem. 2011, 286, 2308–2319. [Google Scholar] [CrossRef]
- Morganti, J.M.; Nash, K.R.; Grimmig, B.A.; Ranjit, S.; Small, B.; Bickford, P.C.; Gemma, C. The soluble isoform of CX3CL1 is necessary for neuroprotection in a mouse model of Parkinson's disease. J. Neurosci. 2012, 32, 14592–14601. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Meier, S.; Schaedelin, S.; Manouchehrinia, A.; Yaldizli, Ö.; Maceski, A.; Oechtering, J.; Achtnichts, L.; Conen, D.; Derfuss, T.; Lalive, P.H.; Mueller, C.; Müller, S.; Naegelin, Y.; Oksenberg, J.R.; Pot, C.; Salmen, A.; Willemse, E.; Kockum, I.; Blennow, K.; Zetterberg, H.; Gobbi, C.; Kappos, L.; Wiendl, H.; Berger, K.; Sormani, M.P.; Granziera, C.; Piehl, F.; Leppert, D.; Kuhle, J. NfL Reference Database in the Swiss Multiple Sclerosis Cohort Study Group. Serum neurofilament light chain for individual prognostication of disease activity in people with multiple sclerosis: A retrospective modelling and validation study. Lancet Neurol. 2022, 21, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.J.; Lee, V.M.; Trojanowski, J.Q. TDP-43 functions and pathogenic mechanisms implicated in TDP-43 proteinopathies. Trends Mol Med. 2011, 17(11), 659–67. [Google Scholar] [CrossRef] [PubMed]
- Llamas-Velasco, S.; Llorente-Ayuso, L.; Contador, I.; Bermejo-Pareja, F. Versiones en español del Minimental State Examination (MMSE). Cuestiones para su uso en la práctica clínica. Rev. Neurol. 2015, 61, 363–371. [Google Scholar] [CrossRef]
- Merino, J. J.; Cabaña-Muñoz, M. E.; Toledano Gasca, A.; Garcimartín, A.; Benedí, J.; Camacho-Alonso, F.; Parmigiani-Izquierdo, J. M. Elevated Systemic L-Kynurenine/L-Tryptophan Ratio and Increased IL-1 Beta and Chemokine (CX3CL1, MCP-1) Proinflammatory Mediators in Patients with Long-Term Titanium Dental Implants. Journal of Clinical Medicine 2019, 8(9), 1368. [Google Scholar] [CrossRef]
- Merino, J. J.; Garcimartín, A.; López-Oliva, M. E.; Benedí, J.; González, M. P. The Impact of CXCR4 Blockade on the Survival of Rat Brain Cortical Neurons. International Journal of Molecular Sciences 2016, 17(12), 2005. [Google Scholar] [CrossRef]
- van der Ende, E.L.; Meeter, L.H.; Poos, J.M.; Panman, J.L.; Jiskoot, L.C.; Dopper, E.G.P.; Papma, J.M.; de Jong, F.J.; Verberk, I.M.W.; Teunissen, C.; Rizopoulos, D.; Heller, C.; Convery, R.S.; Moore, K.M.; Bocchetta, M.; Neason, M.; Cash, D.M.; Borroni, B.; Galimberti, D.; Sanchez-Valle, R.; Laforce, R., Jr.; Moreno, F.; Synofzik, M.; Graff, C.; Masellis, M.; Tartaglia, M.C.; Rowe, J.B.; Vandenberghe, R.; Finger, E.; Tagliavini, F.; de Mendonça, A.; Santana, I.; Butler, C.; Ducharme, S.; Gerhard, A.; Danek, A.; Levin, J.; Otto, M.; Frisoni, G.B.; Cappa, S.; Pijnenburg, Y.A.L.; Rohrer, J.D.; van Swieten, J.C. Genetic Frontotemporal Dementia Initiative (GENFI). Serum neurofilament light chain in genetic frontotemporal dementia: A longitudinal, multicentre cohort study. Lancet Neurol. 2019, 18, 1103–1111. [Google Scholar] [CrossRef]
- Willemse, E.A.J.; Scheltens, P.; Teunissen, C.E.; Vijverberg, E.G.B. A neurologist’s perspective on serum neurofilament light in the memory clinic: A prospective implementation study. Alzheimers Res. Ther. 2021, 13, 101. [Google Scholar] [CrossRef]
- Nyberg, L.; Lundquist, A.; Nordin Adolfsson, A.; Andersson, M.; Zetterberg, H.; Blennow, K.; Adolfsson, R. Elevated plasma neurofilament light in aging reflects brain white-matter alterations but does not predict cognitive decline or Alzheimer’s disease. Alzheimers Dement. (Amst). 2020, 12, e12050. [Google Scholar] [CrossRef]
- Preische, O.; Schultz, S.A.; Apel, A.; Kuhle, J.; Kaeser, S.A.; Barro, C.; Gräber, S.; Kuder-Buletta, E.; LaFougere, C.; Laske, C.; Vöglein, J.; Levin, J.; Masters, C.L.; Martins, R.; Schofield, P.R.; Rossor, M.N.; Graff-Radford, N.R.; Salloway, S.; Ghetti, B.; Ringman, J.M.; Noble, J.M.; Chhatwal, J.; Goate, A.M.; Benzinger, T.L.S.; Morris, J.C.; Bateman, R.J.; Wang, G.; Fagan, A.M.; McDade, E.M.; Gordon, B.A.; Jucker, M. Dominantly Inherited Alzheimer Network. Serum Neurofilament Dynamics Predicts Neurodegeneration and Clinical Progression in Presymptomatic Alzheimer’s Disease. Nat. Med. 2019, 25, 277–283. [Google Scholar] [CrossRef]
- Honey, M.I.J.; van Maurik, I.S.; van Harten, A.C.; Gouda, M.; van Leeuwenstijn, M.; Mank, A.; Trieu, C.; Bouteloup, V.; Chêne, G.; Pellegrin, I.; Dufouil, C.; Doecke, J.D.; Fowler, C.J.; Masters, C.L.; Pijnenburg, Y.; Wilson, D.; van der Flier, W.M.; Teunissen, C.E.; Verberk, I.M.W. Individualized Prediction of Clinical Progression to Dementia Using Plasma Biomarkers in Non-Demented Elderly. Alzheimers Res. Ther. 2025, 17, (published online 3 Dec 2025). [CrossRef] [PubMed]
- Janelidze, S.; Stomrud, E.; Smith, R.; Palmqvist, S.; Mattsson, N.; Airey, D.C.; Proctor, N.K.; Chai, X.; Shcherbinin, S.; Sims, J.R.; Triana-Baltzer, G.; Theunis, C.; Slemmon, R.; Mercken, M.; Kolb, H.; Dage, J.L.; Hansson, O. Cerebrospinal fluid p-tau217 performs better than p-tau181 as a biomarker of Alzheimer's disease. Nat. Commun. 2020, 11, 1683. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ortiz, F.; Kac, P.R.; Brum, W.S.; Zetterberg, H.; Blennow, K.; Karikari, T.K. Plasma Phospho-Tau in Alzheimer’s Disease: Towards Diagnostic and Therapeutic Trial Applications. Molecular Neurodegeneration 2023, 18, 18. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ortiz, F.; Turton, M.; Kac, P.R.; Smirnov, D.; Premi, E.; Ghidoni, R.; Benussi, L.; Cantoni, V.; Saraceno, C.; Rivolta, J.; Ashton, N.J.; Borroni, B.; Galasko, D.; Harrison, P.; Zetterberg, H.; Blennow, K.; Karikari, T.K. Brain-Derived Tau: A Novel Blood-Based Biomarker for Alzheimer’s Disease-Type Neurodegeneration. Brain 2023, 146, 1152–1165. [Google Scholar] [CrossRef]
- Ashton, N.J.; Janelidze, S.; Mattsson-Carlgren, N.; et al. Differential roles of Aβ42/40, p-tau231 and p-tau217 for Alzheimer’s trial selection and disease monitoring. Nat. Med. 2022, 28, 2555–2562. [Google Scholar] [CrossRef]
- Therriault, J.; Pascoal, T.A.; Lussier, F.Z.; Tissot, C.; Chamoun, M.; Bezgin, G.; Servaes, S.; Benedet, A.L.; Ashton, N.J.; Karikari, T.K.; Lantero-Rodriguez, J.; Kunach, P.; Wang, Y.T.; Fernandez-Arias, J.; Massarweh, G.; Vitali, P.; Soucy, J.P.; Saha-Chaudhuri, P.; Blennow, K.; Zetterberg, H.; Gauthier, S.; Rosa-Neto, P. Biomarker modeling of Alzheimer’s disease using PET-based Braak staging. Nat. Aging 2022, 2, 526–535. [Google Scholar] [CrossRef]
- Palmqvist, S.; Janelidze, S.; Quiroz, Y.T.; Zetterberg, H.; Lopera, F.; Stomrud, E.; Su, Y.; Chen, Y.; Serrano, G.E.; Leuzy, A.; Mattsson-Carlgren, N.; Strandberg, O.; Smith, R.; Villegas, A.; Sepulveda-Falla, D.; Chai, X.; Proctor, N.K.; Beach, T.G.; Blennow, K.; Dage, J.L.; Reiman, E.M.; Hansson, O. Discriminative accuracy of plasma phospho-tau217for Alzheimer disease vs other neurodegenerative disorders. JAMA 2020, 324, 772–781. [Google Scholar] [CrossRef]
- Barthélemy, N.R.; Horie, K.; Sato, C.; Bateman, R.J. Blood plasma phosphorylated-tau isoforms track CNS change in Alzheimer’s disease. J. Exp. Med. 2020, 217, e20200861. [Google Scholar] [CrossRef]
- Barthélemy, N.R.; Li, Y.; Joseph-Mathurin, N.; Gordon, B.A.; Hassenstab, J.; Benzinger, T.L.S.; et al. Dominantly Inherited Alzheimer Network. A soluble phosphorylated tau signature links tau, amyloid and the evolution of stages of dominantly inherited Alzheimer’s disease. Nat. Med. 2020, 26, 398–407. [Google Scholar] [CrossRef]
- Suárez-Calvet, M.; Karikari, T.K.; Ashton, N.J.; Lantero Rodríguez, J.; Milà-Alomà, M.; Gispert, J.D.; Salvadó, G.; Minguillon, C.; Fauria, K.; Shekari, M.; Grau-Rivera, O.; Arenaza-Urquijo, E.M.; Sala-Vila, A.; Sánchez-Benavides, G.; González-de-Echávarri, J.M.; Kollmorgen, G.; Stoops, E.; Vanmechelen, E.; Zetterberg, H.; Blennow, K.; Molinuevo, J.L. ALFA Study. Novel Tau Biomarkers Phosphorylated at T181, T217 or T231 Rise in the Initial Stages of the Preclinical Alzheimer’s Continuum When Only Subtle Changes in Aβ Pathology Are Detected. EMBO Mol. Med. 2020, 12, e12921. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, W.; Ming, C.; Gao, X.; Yuan, H.; Lin, X.; Mao, X.; Wang, C.; Guo, X.; Du, Y.; Shao, L.; Yang, R.; Lin, Z.; Wu, X.; Huang, T.Y.; Wang, Z.; Zhang, Y.W.; Xu, H.; Zhao, Y. P-tau217 correlates with neurodegeneration in Alzheimer’s disease, and targeting p-tau217 with immunotherapy ameliorates murine tauopathy. Neuron 2024, 112, 1676–1693.e12. [Google Scholar] [CrossRef]
- Merino, J.J.; Muñetón-Gómez, V.C.; Álvarez, M.I.; Toledano-Díaz, A. Effects of CX3CR1 and Fractalkine Chemokines in Amyloid Beta Clearance and p-Tau Accumulation in Alzheimer’s Disease. Curr. Alzheimer Res. 2016, 13, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.S.; Therriault, J.; Jonaitis, E.M.; Wilson, R.; Langhough, R.E.; Rahmouni, N.; Benedet, A.L.; Ashton, N.J.; Tissot, C.; Lantero-Rodriguez, J.; Macedo, A.C.; Servaes, S.; Wang, Y.T.; Arias, J.F.; Hosseini, S.A.; Betthauser, T.J.; Lussier, F.Z.; Hopewell, R.; Triana-Baltzer, G.; Kolb, H.C.; Jeromin, A.; Kobayashi, E.; Massarweh, G.; Friese, M.A.; Klostranec, J.; Vilali, P.; Pascoal, T.A.; Gauthier, S.; Zetterberg, H.; Blennow, K.; Johnson, S.C.; Rosa-Neto, P. Identification of late-stage tau accumulation using plasma phospho-tau217. EBioMedicine 2024, 109, 105413. [Google Scholar] [CrossRef] [PubMed]
- Pontecorvo, M.J.; Lu, M.; Burnham, S.C.; Schade, A.E.; Dage, J.L.; Shcherbinin, S.; Collins, E.C.; Sims, J.R.; Mintun, M.A. Association of Donanemab Treatment with Exploratory Plasma Biomarkers in Early Symptomatic Alzheimer Disease: A Secondary Analysis of the TRAILBLAZER-ALZ Randomized Clinical Trial. JAMA Neurol. 2022, 79, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Shcherbinin, S.; Andersen, S.W.; Evans, C.D.; Lo, A.C.; Lu, M.; Navitsky, M.; Collins, E.C.; Sims, J.R.; Brooks, D.A.; Mintun, M.A. TRAILBLAZER-ALZ Study: Dynamics of Amyloid Reduction after Donanemab Treatment. Alzheimer’s Dement. 2021, 17, e057492. [Google Scholar] [CrossRef]
- Sims, J.R.; Lu, M.; Schade, A.E.; Brooks, D.A.; Mintun, M.A. TRAILBLAZER-ALZ Studies: Plasma P-Tau Assays and the Initial Performance in Clinical Trials. J. Prev. Alzheimer’s Dis. 2021, 8 Suppl. 1, S2. [Google Scholar]
- Gicas, K.M.; Honer, W.G.; Petyuk, V.A.; Wilson, R.S.; Boyle, P.A.; Leurgans, S.E.; Schneider, J.A.; De Jager, P.L.; Bennett, D.A. Primacy and Recency Effects in Verbal Memory Are Differentially Associated with Post-Mortem Frontal Cortex p-Tau 217 and 202 Levels in a Mixed Sample of Community-Dwelling Older Adults. J. Clin. Exp. Neuropsychol. 2023, 45, 770–785. [Google Scholar] [CrossRef]
- Mattsson-Carlgren, N.; Salvadó, G.; Ashton, N.J.; Tideman, P.; Stomrud, E.; Zetterberg, H.; Ossenkoppele, R.; Betthauser, T.J.; Cody, K.A.; Jonaitis, E.M.; Langhough, R.; Palmqvist, S.; Blennow, K.; Janelidze, S.; Johnson, S.C.; Hansson, O. Prediction of Longitudinal Cognitive Decline in Preclinical Alzheimer Disease Using Plasma Biomarkers. JAMA Neurol. 2023, 80, 360–369. [Google Scholar] [CrossRef]
- Devanarayan, V.; Doherty, T.; Charil, A.; Sachdev, P.; Ye, Y.; Murali, L.K.; Llano, D.A.; Zhou, J.; Reyderman, L.; Hampel, H.; Kramer, L.D.; Dhadda, S.; Irizarry, M.C. Plasma pTau217 Predicts Continuous Brain Amyloid Levels in Preclinical and Early Alzheimer’s Disease. Alzheimers Dement. 2024, 20, 5617–5628. [Google Scholar] [CrossRef]
- Aguillon, D.; Langella, S.; Chen, Y.; Sanchez, J.S.; Su, Y.; Vila-Castelar, C.; Vasquez, D.; Zetterberg, H.; Hansson, O.; Dage, J.L.; Janelidze, S.; Chen, K.; Fox-Fuller, J.T.; Aduen, P.; Martinez, J.E.; Garcia, G.; Baena, A.; Guzman, C.; Johnson, K.A.; Sperling, R.A.; Blennow, K.; Reiman, E.M.; Lopera, F.; Quiroz, Y.T. Plasma p-tau217 predicts in vivo brain pathology and cognition in autosomal dominant Alzheimer's disease. Alzheimers Dement. 2023, 19, 2585–2594. [Google Scholar] [CrossRef]
- Mattsson-Carlgren, N.; Janelidze, S.; Bateman, R.J.; Smith, R.; Stomrud, E.; Serrano, G.E.; Reiman, E.M.; Palmqvist, S.; Dage, J.L.; Beach, T.G.; Hansson, O. Soluble P-tau217 reflects amyloid and tau pathology and mediates the association of amyloid with tau. EMBO Mol. Med. 2021, 13, e14022. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.; Li, B.; Bishnoi, R. P-tau217 as a Reliable Blood-Based Marker of Alzheimer’s Disease. Biomedicines 2024, 12, 1836. [Google Scholar] [CrossRef] [PubMed]
- Burnham, S.C.; Hu, H.; Tang, Y.; Sireci, A.; Pontecorvo, M.J.; Schulz, P.E.; Laird, R.D.; Schreiber, C.P.; Beck, R.C. P-tau217 Testing Impact on Intended Management of Patients Presenting with Cognitive Impairment: A Randomized Clinical Utility Study. Alzheimers Dement. (Amst). 2025, 17(4), e70201. [Google Scholar] [CrossRef] [PubMed]
- Leuzy, A.; Smith, R.; Cullen, N.C.; Strandberg, O.; Vogel, J.W.; Binette, A.P.; Borroni, E.; Janelidze, S.; Ohlsson, T.; Jögi, J.; Ossenkoppele, R.; Palmqvist, S.; Mattsson-Carlgren, N.; Klein, G.; Stomrud, E.; Hansson, O. Biomarker-Based Prediction of Longitudinal Tau Positron Emission Tomography in Alzheimer Disease. JAMA Neurol. 2022, 79, 149–158. [Google Scholar] [CrossRef]
- Barthélemy, N.R.; Salvadó, G.; Schindler, S.E.; He, Y.; Janelidze, S.; Collij, L.E.; et al. Highly accurate blood test for Alzheimer’s disease is similar or superior to clinical cerebrospinal fluid tests. Nat. Med. 2024, 30, 1085–1095. [Google Scholar] [CrossRef]
- Martínez-Dubarbie, F.; Guerra-Ruiz, A.; López-García, S.; Lage, C.; Fernández-Matarrubia, M.; Nevado-Cáceres, Á.; Rivera-Sánchez, M.; Valera-Barrero, A.; Pozueta-Cantudo, A.; García-Martínez, M.; Corrales-Pardo, A.; Bravo, M.; López-Hoyos, M.; Irure-Ventura, J.; de Lucas, E.M.; Drake-Pérez, M.; Cahuana-Santamaría, N.H.; García-Unzueta, M.T.; Sánchez-Juan, P.; Rodríguez-Rodríguez, E. Diagnostic performance of plasma p-tau217 in a memory clinic cohort using the Lumipulse automated platform. Alzheimers Res. Ther. 2025, 17, 68. [Google Scholar] [CrossRef]
- Moon, H.; Chen, X. Alzheimer’s Disease Neuroimaging Initiative. J Prev Alzheimers Dis. 2025, 12, 100252. [Google Scholar] [CrossRef]
- Weston, P.S.J.; Poole, T.; O’Connor, A.; Heslegrave, A.; Ryan, N.S.; Liang, Y.; Druyeh, R.; Mead, S.; Blennow, K.; Schott, J.M.; Frost, C.; Zetterberg, H.; Fox, N.C. Longitudinal measurement of serum neurofilament light in presymptomatic familial Alzheimer’s disease. Alzheimers Res. Ther. 2019, 11, 19. [Google Scholar] [CrossRef]
- Mielke, M.M.; Aakre, J.A.; Algeciras-Schimnich, A.; Proctor, N.K.; Machulda, M.M.; Eichenlaub, U.; Knopman, D.S.; Vemuri, P.; Graff-Radford, J.; Jack, C.R., Jr.; Petersen, R.C.; Dage, J.L. Comparison of CSF Phosphorylated Tau 181 and 217 for Cognitive Decline. Alzheimers Dement. 2022, 18, 602–611. [Google Scholar] [CrossRef]
- Mielke, M.M.; Dage, J.L.; Frank, R.D.; Algeciras-Schimnich, A.; Knopman, D.S.; Lowe, V.J.; Bu, G.; Vemuri, P.; Graff-Radford, J.; Jack, C.R., Jr.; Petersen, R.C. Performance of Plasma Phosphorylated Tau 181 and 217 in the Community. Nat. Med. 2022, 28, 1398–1405. [Google Scholar] [CrossRef]
- Mattsson-Carlgren, N.; Janelidze, S.; Palmqvist, S.; Cullen, N.; Svenningsson, A.L.; Strandberg, O.; Mengel, D.; Walsh, D.M.; Stomrud, E.; Dage, J.L.; Hansson, O. Longitudinal plasma p-tau217 is increased in early stages of Alzheimer’s disease. Brain 2020, 143, 3234–3241. [Google Scholar] [CrossRef] [PubMed]
- Lauro, C.; Catalano, M.; Trettel, F.; Limatola, C. Fractalkine in the nervous system: neuroprotective or neurotoxic molecule? Ann. N. Y. Acad. Sci. 2015, 1351, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-S.; Lim, H.-K.; Lee, J.Y.; Kim, D.-J.; Park, S.; Lee, C.; Lee, C.-U. Changes in the Levels of Plasma Soluble Fractalkine in Patients with Mild Cognitive Impairment and Alzheimer’s Disease. Neurosci. Lett. 2008, 436, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Iemmolo, M.; Ghersi, G.; Bivona, G. The Cytokine CX3CL1 and ADAMs/MMPs in Concerted Cross-Talk Influencing Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 8026. [Google Scholar] [CrossRef]
- Finneran, D.; Li, Q.; Subbarayan, M.S.; Joly-Amado, A.; Kamath, S.; Dengler, D.G.; Gordon, M.N.; Jackson, M.R.; Morgan, D.; Bickford, P.C.; Smith, L.H.; Nash, K.R. Concentration and Proteolysis of CX3CL1 May Regulate the Microglial Response to CX3CL1. Glia 2023, 71, 245–258. [Google Scholar] [CrossRef]
- Fuhrmann, M.; Bittner, T.; Jung, C.K.; Burgold, S.; Page, R.M.; Mitteregger, G.; Haass, C.; LaFerla, F.M.; Kretzschmar, H.; Herms, J. Microglial Cx3cr1 knockout prevents neuron loss in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2010, 13, 411–413. [Google Scholar] [CrossRef]
- Chen, H.R.; Chen, C.W.; Kuo, Y.M.; Chen, B.; Kuan, I.S.; Huang, H.; Lee, J.; Anthony, N.; Kuan, C.Y.; Sun, Y.Y. Monocytes promote acute neuroinflammation and become pathological microglia in neonatal hypoxic-ischemic brain injury. Theranostics 2022, 12, 512–529. [Google Scholar] [CrossRef]
- Bhaskar, K.; Konerth, M.; Kokiko-Cochran, O.N.; Cardona, A.; Ransohoff, R.M.; Lamb, B.T. Regulation of tau pathology by the microglial fractalkine receptor. Neuron 2010, 68, 19–31. [Google Scholar] [CrossRef]
- Imai, T.; Hieshima, K.; Haskell, C.; Baba, M.; Nagira, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Nomiyama, H.; Schall, T.J.; Yoshie, O. Identification and molecular characterization of fractalkine receptor CX3CR1, which mediates both leukocyte migration and adhesion. Cell 1997, 91, 521–530. [Google Scholar] [CrossRef]
- Karikari, T.K.; Emeršič, A.; Vrillon, A.; Lantero-Rodriguez, J.; Ashton, N.J.; Kramberger, M.G.; Dumurgier, J.; Hourregue, C.; Čučnik, S.; Brinkmalm, G.; Rot, U.; Zetterberg, H.; Paquet, C.; Blennow, K. Head-to-head comparison of clinical performance of CSF phospho-tau T181 and T217 biomarkers for Alzheimer's disease diagnosis. Alzheimers Dement. 2021, 17, 755–767. [Google Scholar] [CrossRef]
- Letendre, S.L.; Zheng, J.C.; Kaul, M.; Yiannoutsos, C.T.; Ellis, R.J.; Taylor, M.J.; Marquie-Beck, J.; Navia, B. HIV Neuroimaging Consortium. Chemokines in cerebrospinal fluid correlate with cerebral metabolite patterns in HIV-infected individuals. J. Neurovirol. 2011, 17, 63–69. [Google Scholar] [CrossRef]
- Nash, K.R.; Lee, D.C.; Hunt, J.B., Jr.; Morganti, J.M.; Selenica, M.L.; Moran, P.; Reid, P.; Brownlow, M.; Yang, G.-Y.; Savalia, M.; Gemma, C.; Bickford, P.C.; Gordon, M.N.; Morgan, D. Fractalkine overexpression suppresses tau pathology in a mouse model of tauopathy. Neurobiol. Aging 2013, 34, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Özdemir, S.; Fritz, C.; Möbius, W.; Kleineidam, L.; Mandelkow, E.; Biernat, J.; Doğdu, C.; Peters, O.; Cosma, N.C.; Wang, X.; Schneider, L.S.; Priller, J.; Spruth, E.; Kühn, A.A.; Krause, P.; Klockgether, T.; Vogt, I.R.; Kimmich, O.; Spottke, A.; Hoffmann, D.C.; Fliessbach, K.; Miklitz, C.; McCormick, C.; Weydt, P.; Falkenburger, B.; Brandt, M.; Guenther, R.; Dinter, E.; Wiltfang, J.; Hansen, N.; Bähr, M.; Zerr, I.; Flöel, A.; Nestor, P.J.; Düzel, E.; Glanz, W.; Incesoy, E.; Bürger, K.; Janowitz, D.; Perneczky, R.; Rauchmann, B.S.; Hopfner, F.; Wagemann, O.; Levin, J.; Teipel, S.; Kilimann, I.; Goerss, D.; Prudlo, J.; Gasser, T.; Brockmann, K.; Mengel, D.; Zimmermann, M.; Synofzik, M.; Wilke, C.; Selma-González, J.; Turon-Sans, J.; Santos-Santos, M.A.; Alcolea, D.; Rubio-Guerra, S.; Fortea, J.; Carbayo, Á.; Lleó, A.; Rojas-García, R.; Illán-Gala, I.; Wagner, M.; Frommann, I.; Roeske, S.; Bertram, L.; Heneka, M.T.; Brosseron, F.; Ramirez, A.; Schmid, M.; Beschorner, R.; Halle, A.; Herms, J.; Neumann, M.; Barthélemy, N.R.; Bateman, R.J.; Rizzu, P.; Heutink, P.; Dols-Icardo, O.; Höglinger, G.; Hermann, A.; Schneider, A. Plasma Extracellular Vesicle Tau and TDP-43 as Diagnostic Biomarkers in FTD and ALS. Nat. Med. 2024, 30(6), 1771–1783.
- Chatterjee, M.; Özdemir, S.; Fritz, C.; et al. Plasma extracellular vesicle tau and TDP-43 as diagnostic biomarkers in FTD and ALS. Nat Med 2024, 30(6), 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Solje, E.; Benussi, A.; Buratti, E.; Remes, A.M.; Haapasalo, A.; Borroni, B.; Brugnolo, A. State-of-the-Art Methods and Emerging Fluid Biomarkers in the Diagnostics of Dementia—A Short Review and Diagnostic Algorithm. Diagnostics 2021, 11, 788. [Google Scholar] [CrossRef]
- Bauer, C.E.; Zachariou, V.; Sudduth, T.L.; Van Eldik, L.J.; Jicha, G.A.; Nelson, P.T.; Wilcock, D.M.; Gold, B.T. Plasma TDP-43 levels are associated with neuroimaging measures of brain structure in limbic regions. Alzheimers Dement (Amst) 2023, 31;15(2), e12437. [Google Scholar] [CrossRef]
- Foulds, P.; McAuley, E.; Gibbons, L.; Davidson, Y.; Pickering-Brown, S.M.; Neary, D.; Snowden, J.S.; Allsop, D.; Mann, D.M. TDP-43 protein in plasma may index TDP-43 brain pathology in Alzheimer's disease and frontotemporal lobar degeneration. Acta Neuropathol. 2008, 116(2), 141–6. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Foulds, P; Davidson, Y; Mishra, M; et al. Plasma phosphorylated-TDP-43 protein levels correlate with brain pathology in frontotemporal lobar degeneration. Acta Neuropathol. 2009, 118, 647–658. [Google Scholar] [CrossRef]
- Schindler, S.E.; Galasko, D.; Pereira, A.C.; Rabinovici, G.D.; Salloway, S.; Suárez-Calvet, M.; Khachaturian, A.S.; Mielke, M.M.; Udeh-Momoh, C.; Weiss, J.; Batrla, R.; Bozeat, S.; Dwyer, J.R.; Holzapfel, D.; Jones, D.R.; Murray, J.F.; Partrick, K.A.; Scholler, E.; Vradenburg, G.; Young, D.; Algeciras-Schimnich, A.; Aubrecht, J.; Braunstein, J.B.; Hendrix, J.; Hu, Y.H.; Mattke, S.; Monane, M.; Reilly, D.; Somers, E.; Teunissen, C.E.; Shobin, E.; Vanderstichele, H.; Weiner, M.W.; Wilson, D.; Hansson, O. Acceptable Performance of Blood Biomarker Tests of Amyloid Pathology-Recommendations from the Global CEO Initiative on Alzheimer’s Disease. Nat. Rev. Neurol. 2024, 20, 426–439. [Google Scholar] [CrossRef]
- Minic, R.; Zivkovic, I. Optimization, validation and standardization of ELISA. In Norovirus (pp. 9-28). London, UK: IntechOpen. 2020, 9-28.
- Vila-Castelar, C.; Chen, Y.; Langella, S.; Lopera, F.; Zetterberg, H.; Hansson, O.; Dage, J.L.; Janelidze, S.; Su, Y.; Chen, K.; Pluim McDowell, C.; Martinez, J.E.; Ramirez-Gomez, L.; Garcia, G.; Aguillon, D.; Baena, A.; Giraldo-Chica, M.; Protas, H.D.; Ghisays, V.; Rios-Romenets, S.; Tariot, P.N.; Blennow, K.; Reiman, E.M.; Quiroz, Y.T. Sex differences in blood biomarkers and cognitive performance in individuals with autosomal dominant Alzheimer’s disease. Alzheimers Dement. 2023, 19, 4127–4138. [Google Scholar] [CrossRef]
- Pichet Binette, A.; Janelidze, S.; Cullen, N.; Dage, J.L.; Bateman, R.J.; Zetterberg, H.; Blennow, K.; Stomrud, E.; Mattsson-Carlgren, N.; Hansson, O. Confounding Factors of Alzheimer’s Disease Plasma Biomarkers and Their Impact on Clinical Performance. Alzheimer’s Dement. 2023, 19, 1403–1414. [Google Scholar] [CrossRef]
- Daniilidou, M.; Öhlund-Wistbacka, U.; Hagman, G.; Rosenberg, A.; Ashton, N.; Zetterberg, H.; Blennow, K.; Matton, A.; Kivipelto, M. Enhancing diagnostic precision in Alzheimer's disease: Impact of comorbidities on blood biomarkers for clinical integration. Alzheimers Dement. 2025, 21(12), e70931. [Google Scholar] [CrossRef]
Figure 4.
(A). Percentage of cell viability in LPS (1 µg/mL)-treated neurons for 24 hours, as determined by the MTT assay, and sFK recombinant protein were added during 24 hour more in the presence/absence of LPS treatment. An antibody against CX3CR1 was also included in order to block CX3CR1 chemokine receptor. (A) Data are presented as mean ± S.E.M of 3 different experiments in vitro (n = 3) of XTT assay for neuronal survival stimation. Additionally, caspase-3 activity as index of apoptosis were stimated in extracts from neurons under these inflammatory conditions in order to ascertain whether fractalkine could prevent caspase-3-induced apoptosis in cortical neurons at 7 DIV. The controls are expressed as 100 % of percentage.
Figure 4.
(A). Percentage of cell viability in LPS (1 µg/mL)-treated neurons for 24 hours, as determined by the MTT assay, and sFK recombinant protein were added during 24 hour more in the presence/absence of LPS treatment. An antibody against CX3CR1 was also included in order to block CX3CR1 chemokine receptor. (A) Data are presented as mean ± S.E.M of 3 different experiments in vitro (n = 3) of XTT assay for neuronal survival stimation. Additionally, caspase-3 activity as index of apoptosis were stimated in extracts from neurons under these inflammatory conditions in order to ascertain whether fractalkine could prevent caspase-3-induced apoptosis in cortical neurons at 7 DIV. The controls are expressed as 100 % of percentage.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



