Submitted:
31 December 2025
Posted:
01 January 2026
You are already at the latest version
Abstract
Background/Objectives: Antimicrobial peptides (AMPs), evolutionarily conserved components of innate immunity characterized by their broad-spectrum efficacy and minimal resistance development, are increasingly recognized as promising therapeutic candidates. This review aims to integrate current knowledge concerning natural and synthetic antimicrobial peptides and their therapeutic effectiveness in addressing gastrointestinal infections. Methods: A literature review was performed, evaluating recent peer-reviewed studies on AMPs. The research concentrated on their molecular mechanisms of action, antimicrobial spectrum, as well as on their interactions with standard antibiotics. More in detail, the peptide classes examined herein included defensins, cathelicidins, histatins, and various natural peptides such as lactoferricin, protamines, RegIII, and hepcidin, along with synthetic analogs like WR12, D-IK8, MSI-78, IMX942. Results: Natural AMPs demonstrated significant antimicrobial and immunomodulatory effects against Escherichia coli, Klebsiella pneumoniae, Salmonella spp., and Shigella spp. Beyond direct antimicrobial activity, intestinal antimicrobial peptides act as integrated anti-infective agents by modulating host–microbiota interactions, preserving epithelial barrier integrity, and limiting inflammation, thereby offering a multifaceted strategy to control gastrointestinal infections. On the other hand, synthetic peptides showed better stability, less toxicity, and synergistic interactions with antibiotics, which suggests that they could be used either alone or in combination with other treatments. Conclusions: AMPs constitute a promising category endowed with anti-infective activity especially for therapy of intestinal diseases, which is attributed to their distinctive anti-infective mechanisms, immune-modulating characteristics, and a relatively low propensity for resistance development compared to conventional antibiotics. However, more clinical trials and improvements to their formulation are needed to translate promising in vitro results into reliable patient outcomes.
Keywords:
intestinal infections
; antimicrobial peptides
; gut microbiota
; epithelial barrier
; defensins
; cathelicidins
; synthetic antimicrobial peptides
; antibiotic resistance
; anti-infective therapy
1. Introduction
Intestinal infections remain a pressing global health challenge, largely due to their high prevalence and the growing resistance of pathogenic microorganisms to existing antimicrobial agents [1,2,3,4]. In recent years, several Gram-negative and Gram-positive pathogens, including Escherichia coli, Klebsiella pneumoniae, Salmonella spp., and Shigella spp., have developed multi-drug-resistant (MDR) and even extremely drug-resistant (XDR) traits [5,6,7,8] . Consequently, the efficacy of traditional antibiotics has markedly declined. This issue is particularly pronounced in developing countries, where inappropriate antibiotic use and inadequate regulatory enforcement accelerate the dissemination of resistant strains. The resulting increase in morbidity and mortality highlights the urgent need for innovative therapeutic approaches and strengthened public health measures [9,10,11] . With the declining efficacy of conventional antibiotics, antibacterial peptides (AMPs) have emerged as a promising alternative. AMPs are naturally occurring molecules present across nearly all forms of life and constitute a fundamental component of the innate immune system. Beyond their ability to disrupt bacterial membranes, they also modulate host inflammatory responses, with high concentrations of these peptides being found within the intestinal mucosa, a critical protective barrier of the body. Produced primarily by epithelial and Paneth cells, AMPs prevent pathogen adhesion and invasion while simultaneously contributing to the maintenance of intestinal microbiota homeostasis [12,13,14,15,16,17] . Over the past few years, significant progress has been achieved in developing synthetic peptides that can overcome the limitations of natural antibacterial peptides, such as lower stability or reduced activity under normal conditions. In this regard, structural modifications, amino acid substitutions, and modern design methodologies enabled the production of more stable, targeted, and effective peptides, thereby increasing their potential for treating intestinal infections [16,17,18,19,20] . The objective of this review is to provide a comprehensive overview of current knowledge on both natural and synthetic antibacterial peptides, with particular emphasis on their biological functions, mechanisms of action, and potential clinical applications as anti-infective agents. In fact, as we discuss below, antimicrobial peptides are increasingly regarded as safe and effective candidates for next-generation antibiotics, offering promising strategies to combat intestinal infections and address the escalating global crisis of antibiotic resistance
Methods
To ensure a comprehensive analysis of the role of antimicrobial peptides in intestinal infections, a literature search was conducted. The search was performed using the electronic databases PubMed/MEDLINE, Scopus, and Web of Science. The search covered the period from January 2010 to December 2024, with a special focus on studies published in the last 5 years to ensure the relevance of the data. The following keywords and their combinations were used: “antimicrobial peptides”, “AMPs”, “intestinal infections”, “gut microbiota”, “defensins”, “cathelicidins”, “LL-37”, “synthetic peptides”, “antibiotic resistance”, “gastrointestinal tract”, and “peptide engineering”. The inclusion criteria were: original research articles and reviews published in English, studies focusing on the mechanism of action, immunomodulatory effects, and clinical potential of natural and synthetic AMPs in the context of intestinal infections, papers describing interactions between AMPs and common intestinal pathogens (Escherichia coli, Salmonella spp., Shigella spp., Klebsiella pneumoniae). The exclusion criteria were conference abstracts, editorials, and short communications without full text availability, studies focusing solely on non-intestinal infections (e.g., respiratory or skin infections) unless relevant mechanistic parallels were drawn, duplicate publications.
Study quality and relevance were evaluated using predefined criteria, including clarity of experimental design, appropriateness of methods used to assess antimicrobial or immunomodulatory activity, relevance to intestinal pathogens or gut immunity, and robustness of reported outcomes. This process was guided by general principles of systematic literature assessment and informed by elements of the PRISMA framework. As for data selection and analysis, initially a total of 800 records were identified through database searching. After removing duplicates, identified through visual inspection and verification, and screening titles/abstracts for relevance, 250 full-text articles were assessed for eligibility. Finally, 216 studies were selected for inclusion in this review.
2. AMPs as Emerging Therapeutics for Gastrointestinal Pathogens
This review synthesizes current scientific evidence on the structural organization, biological activities, and therapeutic potential of both natural and synthetic AMPs in the prevention and treatment of gastrointestinal infections [14,21,22]. The studies reviewed elucidate not only the molecular mechanisms of action of AMPs, but also features such as their spectrum of antimicrobial activity, synergism with conventional antibiotics, as well as their role in regulating intestinal immunity and preserving the epithelial barrier [23,24,25] . The major classes of natural antimicrobial peptides, such as defensins, cathelicidins, histatins, lactoferricin, protamines, members of the RegIII family, and hepcidin, exhibit potent antimicrobial, anti-inflammatory, and immunomodulatory activities against clinically relevant pathogens, including Escherichia coli, Klebsiella pneumoniae, Salmonella spp., and Shigella spp [26,27,28,29] . On the other hand, synthetic analogues developed through rational peptide-engineering strategies exhibit enhanced physicochemical and functional characteristics relative to natural AMPs and can be specifically designed to achieve increased proteolytic stability together with potentially reduced cytotoxicity and improved biocompatibility [30,31,32,33] . Table 1 presents a list of bioactive antimicrobial peptides currently under investigation at different stages of biological activity research.
Interestingly, multiple studies have observed synergistic effects between synthetic AMPs and antibiotics, thereby enhancing the prospects for combinatorial strategies in the management of antibiotic-resistant infections. The available evidence supports the classification of antimicrobial peptides as a distinct category of biomolecules with considerable clinical potential, owing to their broad spectrum of activity, lower propensity for resistance development when compared to other antibiotics, and ability to maintain the integrity of the intestinal epithelial barrier [85,86,87,88] . These findings suggest the necessity for additional research focused on enhancing their pharmaceutical formulations, together with improving AMP stability, and mitigating the potential toxicity observed in some cases. Understanding these aspects of AMPs is crucial especially in considering their interactions with the gut microbiota, which will be discussed in the next section
2.1. Interaction Between Intestinal Microbiota and Epithelial Antimicrobial Peptides
The intestinal microbiota represents a highly organized consortium of microorganisms, predominantly comprising members of the phyla Bacteroidetes, Firmicutes, Actinobacteria, and Proteobacteria. These bacterial groups play essential roles in host metabolism, regulation of the immune system, defense against pathogenic organisms, and maintenance of intestinal barrier integrity [89,90,91]. Epithelial-derived AMPs, produced primarily by Paneth cells and enterocytes along the crypt–villous axis, constitute a critical component of the gut’s innate defense system. These peptides regulate the dynamic interactions between the intestinal microbiota and epithelial cells. Their functions extend well beyond direct antimicrobial activity and include:
- (i)
- maintaining the spatial structure of microbial communities, forming distinct gastrointestinal niches for various taxa;
- (ii)
- preventing colonization by pathogens by limiting their adhesion and invasion;
- (iii)
- creating a sterile layer over the epithelium (especially due to RegIIIγ), preventing bacterial contact with the mucosa;
- (iv)
- regulating innate immunity, including modulating Toll-like receptors, producing cytokines, and controlling the inflammatory response.
Epithelial antimicrobial peptides work together to control the microbiota on many levels. They achieve this by directly eliminating microorganisms, fine-tuning the immune response, and keeping the barrier intact. However, the contribution of individual classes of AMPs to these processes varies and is determined by their structural and functional properties. Defensins, the most extensively characterized group of epithelial AMPs occupy a particularly prominent role. They have been shown to play a key role in crypt protection, regulation of microbiota composition, and are implicated in the pathogenesis of inflammatory bowel diseases, most notably ileal Crohn’s disease. Accordingly, a detailed examination of the biological properties and mechanisms of action of defensins is warranted, as they represent the principal effector molecules of innate intestinal immunity [92,93,94,95,96,97] . Thus, the anti-infective efficacy of intestinal antimicrobial peptides is tightly linked to their ability to regulate host–microbiota interactions, thereby limiting pathogen overgrowth while maintaining a balanced and protective microbial ecosystem in the gut
2.1.1. Defensins as Key Epithelial Effector Molecules of Intestinal Innate Immunity
As anticipated, defensins are the most studied and important group of epithelial antimicrobial peptides, playing a crucial role in maintaining gut homeostasis. They directly eliminate bacteria, control the arrangement of the microbiota, and prevent pathogens from adhering to and penetrating the intestinal epithelium. The human intestine contains two main types of defensins: α-defensins, which are produced by Paneth cells, and β-defensins, synthesized by enterocytes and mucosal immune cells [98,99]. Figure 1 (below) provides an overview of the cellular origins, antimicrobial mechanisms, and immunological functions of intestinal α- and β-defensins. These peptides, produced by Paneth cells, enterocytes, and mucosal immune cells, are discussed in greater detail below.
Paneth cells represent the principal source of α-defensins in the small intestine, producing human defensin-5 (HD-5) and human defensin-6 (HD-6). These peptides display potent activity against a wide range of gram-negative and gram-positive bacteria, including clinically relevant enteropathogens such as Escherichia coli, Salmonella spp., Klebsiella pneumoniae, and Shigella spp., including multidrug-resistant strains. HD-5 exhibits a potent direct bactericidal effect by disrupting the integrity of bacterial membranes. HD-6, in contrast, acts in a non-standard manner: it forms amyloid-like fibrils that form so-called “nanowebs” that physically trap bacteria and prevent them from penetrating the crypts. This mechanism ensures control of microbiota without excessive inflammation, which is critical for maintaining epithelial integrity [100,101,102,103]. β-Defensins (hBD-1, hBD-2, hBD-3) are produced primarily by enterocytes and immune system cells (macrophages, dendritic cells) in response to bacterial Pathogen-Associated Molecular Patterns (PAMP) molecules: lipopolysaccharide (LPS), lipoteichoic acid, and muramyl peptide derivatives, as well as proinflammatory cytokines (IL-1β, TNF-α). The functional distribution of β-defensins reflects their diverse roles in intestinal immunity. In particular, hBD-1 is a constitutively expressed peptide that provides a basic level of protection; in contrast, hBD-2 is induced during inflammation and has pronounced activity against gram-negative bacteria, while hBD-3 exhibits a broad spectrum of antimicrobial activity, including effectiveness against yeast fungi (Candida spp.). The balance of α- and β-defensins is critical for homeostasis, with a deficiency in their secretion being closely associated with intestinal diseases. For instance, in ileal Crohn’s disease, reduced α-defensin levels arise from Paneth cell dysfunction, while diminished β-defensin expression is linked to dysbiosis, chronic inflammation, and increased epithelial permeability. Collectively, defensins constitute essential components of the innate intestinal immune system, providing direct antimicrobial activity, reinforcing the mucosal barrier, and regulating the composition of the gut microbiota. Their deficiency results in barrier dysfunction, pathogenic alteration of microbiome, and heightened vulnerability to inflammatory diseases [104,105,106,107] .Overall, defensins constitute a first-line anti-infective barrier in the intestine, exerting potent activity against enteric pathogens while shaping microbiota composition and preventing bacterial invasion of the epithelial surface. However, therapeutic manipulation of defense peptides remains challenging due to their cell-type specificity [108] the theoretical risk of microbiota modification, and the limited number of clinical studies validating their safety and efficacy. Although endogenous defensins appear to support commensal colonization and contribute to microbiota recovery after dysbiosis [109], further research is needed to clarify how exogenous defensins or defensin-based therapeutics interact with the intestinal ecosystem when administered in a therapeutic context
2.1.2. Cathelicidin LL-37 in the Gut: Immunomodulatory Mechanisms and Barrier-Protective Functions
Cathelicidins are epithelial antimicrobial peptides with both antimicrobial and immunomodulatory functions. In humans, the sole representative is LL-37, an amphiphilic α-helical peptide. LL-37 exerts broad activity against intestinal pathogens such as Escherichia coli, Salmonella spp., Shigella spp., and Klebsiella pneumoniae by destabilizing bacterial membranes, forming pores, and preventing biofilm formation. It also blocks adherence and invasion of enteropathogenic E. coli, thereby protecting epithelial cells. Importantly, LL-37 displays relatively increased resistance to proteolytic degradation compared to many other natural antimicrobial peptides, allowing it to retain activity in the intestinal environment. Figure 2 illustrates LL-37’s principal activities, including membrane disruption, biofilm inhibition, modulation of inflammatory signaling, and promotion of epithelial repair and tight-junction integrity [110,111] .
The most critical property of LL-37 is its ability to modulate the intensity of the inflammatory response, thereby preventing excessive immune activation. This peptide binds to and neutralizes LPS, inhibiting Toll-like receptor 4 (TLR4)-mediated initiation of the inflammatory cascade. LL-37 further suppresses NF-κB- and MAPK-dependent signaling pathways, resulting in reduced production of proinflammatory cytokines (TNF-α, IL-6, IL-1β) and enhanced secretion of the anti-inflammatory cytokine IL-10. In addition, LL-37 influences the functional activity of macrophages, neutrophils, and dendritic cells. These properties render LL-37 particularly significant in intestinal disorders characterized by excessive inflammation, including infectious colitis and inflammatory bowel disease [112,113,114] . In addition to its bactericidal activity, LL-37 promotes epithelial cell migration, proliferation, and differentiation. It accelerates wound healing and facilitates the restoration of mucosal tissue, while also reinforcing tight junctions between epithelial cells. These actions collectively enhance barrier integrity and prevent the translocation of bacteria and toxins across the intestinal epithelium. In models of infection induced by Citrobacter rodentium or enterohemorrhagic Escherichia coli (EHEC), LL-37 diminishes epithelial damage, reduces neutrophil infiltration, and expedites barrier restoration. Evidence shows that LL-37 acts as both an antimicrobial molecule and a regulator of immune homeostasis. LL-37 prevents excessive inflammation, preserves epithelial barrier integrity, and accelerates the healing of epithelial tissue following infection. Owing to its ability to modulate diverse immune and regenerative processes, LL-37 is regarded as one of the most versatile and multifunctional antimicrobial peptides in the intestine [115,116,117,118] . Thus, LL-37 plays a dual anti-infective role in the intestine by directly eliminating enteric pathogens and modulating inflammatory responses, thereby limiting tissue damage during intestinal infections. From a translational and therapeutic standpoint, future research must address key clinical limitations of LL-37, including its potential cytotoxicity at elevated concentrations, biochemical instability, and dose-dependent constraints. Moreover, overcoming challenges related to oral or luminal delivery remains essential for its effective application in gastrointestinal settings.
2.1.3. Regenerating AMPs (RegIII) and the Spatial Organization of the Gut Microbiota
The RegIII family (Figure 3a) of regenerating-like lectins represents a distinct class of intestinal epithelial AMPs. In contrast to defensins or LL-37, which primarily eliminate microorganisms by disrupting their membranes, RegIII peptides contribute mainly to the spatial organization of the gut microbiota, preserving physical separation between commensal bacteria and host tissues (Figure 3b). This function is increasingly recognized as a key element of mucosal homeostasis [119,120] . Regarding structural features and regulation, RegIIIγ, the best-characterized member of the regenerating gene family in mice, is a C-type lectin–like antimicrobial peptide produced by Paneth cells and enterocytes [121,122] . Its expression is tightly regulated by microbial and immune signals. Microbial-associated molecular patterns trigger TLR/MyD88-dependent signaling in intestinal epithelial cells, resulting in the induction of RegIIIγ expression in response to bacterial colonization. In addition, IL-22, produced predominantly by type 3 innate lymphoid cells (ILC3s) and Th17 cells, markedly upregulates RegIIIγ expression during increased microbial load, inflammation, or infection. This dual regulation enables epithelial cells to rapidly adjust their antimicrobial activity according to environmental conditions. RegIIIγ binds exposed peptidoglycan on the surface of Gram-positive bacteria, which underlies its selective antimicrobial spectrum [123,124] . With respect to spatial barrier formation, one of the most critical functions of RegIIIγ is the maintenance of a bacteria-free exclusion zone immediately above the intestinal epithelium. This antimicrobial lectin thereby enforces physical separation between commensal microbiota and host tissues, preserving epithelial integrity and preventing inappropriate immune activation. Thus, rather than acting solely as a classical membrane-disrupting AMP, RegIIIγ primarily shapes the spatial structure of the microbiota in vivo [119,121]. It helps to prevent direct microbial contact with the apical surface of enterocytes, restricts colonization of the mucosal surface by commensals, and limits the expansion of mucosa-associated bacteria. In various experimental models, deficiency of RegIII lectins leads to increased mucosa-associated bacteria, enhanced bacterial translocation to mesenteric lymph nodes and liver, as well as a higher risk of systemic complications. In other words, RegIIIγ acts as a chemical barrier that maintains a safe distance between the host and its dense microbial community [119,121].
As for the mechanisms of antibacterial activity, RegIIIγ primarily enforces spatial segregation of bacteria from the mucosal surface but also displays direct bactericidal activity against Gram-positive species. It binds peptidoglycan, oligomerizes on the bacterial surface, and forms pore-like structures that disrupt membranes and cause lysis. Gram-negative bacteria are less susceptible due to their protective outer membrane. Beyond antimicrobial effects, RegIIIγ supports intestinal homeostasis by stratifying microbial communities, preserving barrier integrity during inflammation, and limiting bacterial translocation. Reduced expression is linked to dysbiosis, inflammatory bowel disease (IBD), and impaired host–microbiota balance [119,121,125,126] . Overall, RegIII antimicrobial peptides contribute to intestinal anti-infective defense primarily by enforcing spatial segregation between bacteria and the epithelium, reducing pathogen translocation and preventing infection-driven inflammation
2.1.4. The Overall Role of Epithelial Antimicrobial Peptides in Intestinal Homeostasis
Remarkably, epithelial antimicrobial peptides form a multi-level biochemical defense system, providing both direct antimicrobial protection and regulating interactions between the epithelium and microbiota [127,128] . Their coordinated activity encompasses several complementary functions: direct suppression of pathogen growth through membrane disruption, biofilm inhibition, and toxin neutralization. Other functions comprise the stabilization and spatial organization of the commensal microbiota, including control of opportunistic bacterial expansion, the modulation of the inflammatory response to prevent excessive immune activation and tissue damage; as well as the enhancement of mucosal barrier function by strengthening intercellular junctions and preventing bacterial translocation across the epithelium [129,130,131] . Two-way interactions with microbial metabolites, such as short-chain fatty acids and indole derivatives, tightly regulate AMP expression and contribute to the maintenance of the immune tone. Consequently, epithelial AMPs serve as a central link in sustaining a stable bidirectional contact between the host and its microbiota. They ensure a delicate balance between antimicrobial defense, regulation of inflammation, and preservation of intestinal barrier integrity. Impaired AMP secretion disrupts this equilibrium, leading to dysbiosis, heightened susceptibility to infections, and the development of inflammatory bowel diseases, underscoring their fundamental importance for intestinal homeostasis [127,128] . Although intestinal AMPs, such as defensins, cathelicidins, and Reg III, are endowed with unique functions in the mucosa, they represent only part of a much larger family of natural antimicrobial peptides found in various organisms and sharing common structural motifs. Understanding the universal structural classes of natural AMPs, regardless of their tissue origin, not only explains the similarities in their mechanisms of action, but also substantiates their therapeutic potential, ultimately serving as the foundation for the development of synthetic analogues [132,133,134]. Hence, through coordinated antimicrobial, immunomodulatory, and barrier-protective activities, epithelial antimicrobial peptides form an integrated anti-infective system that safeguards intestinal homeostasis during microbial challenge
2.2. Natural Antimicrobial Peptides: Structural Organization and Mechanisms of Action
Most natural AMPs are a heterogeneous group of low-molecular weight cationic peptides that constitute the primary chemical barrier at mucosal surfaces. All natural AMPs share key physicochemical traits, such as a positive charge, amphipathicity, and affinity for negatively charged microbial lipids, despite differing in origin, sequence, and length. This conserved structure provides a rapid, evolutionarily stable defense that limits resistance and makes AMPs essential for intestinal protection against infection [135,136,137] .The conserved structural features of natural antimicrobial peptides underpin their rapid and broad anti-infective activity in the intestine, enabling effective control of enteric pathogens with a reduced risk of resistance development.
2.2.1. Structural Diversity of Natural AMPs
The structural features of AMPs determine the mechanism of their interaction with membranes and the spectrum of their antimicrobial activity. Four architectural classes are most common: α-helical peptides, β-sheet structures, extended (linear) peptides, and α/β-combined molecules. Interestingly, α-helical AMPs such as LL-37 and lactoferricin lack disulfide bonds and display strong amphipathicity. Due to their flexibility and ability to assume a helical shape only upon contact with the membrane, these peptides effectively integrate into the lipid bilayer and form pores, causing rapid osmotic lysis of bacteria. In this context, in addition to its direct antimicrobial function, LL-37 regulates epithelial regeneration, downregulates proinflammatory signaling pathways, and neutralizes endotoxins, making it a unique link between antimicrobial defense and immunomodulation [133,138,139] . On the other hand, β-sheet AMPs, which include α- and β-defensins, are characterized by a stable conformation reinforced by disulfide bridges. The structural configuration of defensins confers high resistance to proteolysis, a property particularly important in the intestinal lumen, which is rich in digestive enzymes and microbial proteases. Defensins exert broad-spectrum antimicrobial activity against gram-positive and gram-negative bacteria, including multidrug-resistant strains, as well as yeasts. Beyond their bactericidal capacity, defensins also contribute to barrier formation; for example, HD-6 assembles into ‘nanonets’ that physically entrap pathogens, preventing their access to intestinal crypts without eliciting excessive inflammation. In contrast, long linear AMPs such as indolicidin, histatin-5, and tritripticin lack defined secondary structure and are enriched in aromatic and cationic amino acids. This structural flexibility enables them to insert into membranes through multiple pathways and disrupt target cell function. Owing to their ability to penetrate bacterial cells, linear AMPs frequently inhibit DNA and protein synthesis, thereby functioning in a manner that parallels traditional chemotherapeutic agents [133,138,139]. Combined α/β structures, including protegrins and thanatin, are compact, disulfide-stabilized molecules that combine mechanical stability and high activity. These peptides exhibit pronounced bactericidal activity and the ability to disrupt biofilms, which is particularly important in the treatment of chronic infections [138,139,140]. Thus, the structural diversity of natural antimicrobial peptides determines their spectrum and mode of anti-infective action in the gut, allowing efficient targeting of intestinal pathogens through membrane disruption and intracellular interference.
2.2.2. Molecular Mechanisms of Action of Natural AMPs
Owing to the physicochemical characteristics of bacterial membranes, their lipid composition is a crucial determinant of membrane behavior and cellular responses. Different bacterial species possess distinct membrane profiles, and even within a single species the membrane makeup is not fixed but adapts to environmental conditions. Bacterial membranes contain a wide array of amphiphilic lipids. Among the most common are phosphatidylglycerol, phosphatidylethanolamine and cardiolipin, while phosphatidylcholine and phosphatidylinositol appear less frequently. In addition to these phospholipids, bacteria may incorporate other lipid classes such as ornithine lipids, glycolipids, sphingolipids, hopanoids and various specialized membrane components [141]. Antimicrobial peptides typically exert their bactericidal effect by direct contact with the cell membranes of pathogens. AMPs interact with negatively charged lipids to trigger a sequence of structural changes that disrupt the integrity of membranes (Figure 4). In this frame, the barrel-stave, toroidal pore, and carpet-like models are the most studied mechanisms describing how AMPs eliminate both Gram-positive and Gram-negative bacteria. Clinically, AMPs perform better when modified with synthetic strategies such as D-amino acids, lipidation, and cyclization, which stabilize pore formation. The process begins with cationic AMPs binding via electrostatic forces to bacterial surfaces, lipopolysaccharides in Gram-negative and peptidoglycan in Gram-positive species, followed by insertion into the lipid bilayer that destabilizes membranes. The final step involves pore formation: cylindrical pores (barrel-stave), toroidal pores (toroidal pore), or carpet-like disintegration, all leading to cell death. These mechanisms apply to both natural and synthetic AMPs, ensuring rapid bacterial clearance. Beyond direct bactericidal activity, natural AMPs interact with host immunological receptors, inhibit Toll-like receptor signaling, reduce proinflammatory cytokines, and protect mucosal tissue, uniquely combining antimicrobial and anti-inflammatory functions [17,133,142,143]
Recent data show that natural AMPs are a promising basis for intestinal therapeutics, being effective against Escherichia coli, Shigella spp., Salmonella spp., and Klebsiella pneumoniae, including multidrug-resistant strains, while protecting and accelerating healing of epithelial cells, blocking pathogen attachment and invasion, and preventing bacterial translocation into the bloodstream and systemic disease. By altering fundamental membrane physicochemical properties, they reduce the likelihood of resistance development, suggesting, thus, their clinical potential; however, instability, rapid proteolysis, and low bioavailability limit their use, leading to the creation of synthetic analogues and peptidomimetics that retain antimicrobial efficacy but exhibit superior pharmacological properties [17, 133. 144,145 ]. Overall, by targeting fundamental membrane properties and key immune signaling pathways, natural antimicrobial peptides exert rapid anti-infective effects against intestinal pathogens while simultaneously limiting inflammatory tissue damage.
2.3. Synthetic Antimicrobial Peptides: A Modern Idea and Their Role in Medicine
Synthetic AMPs represent a robust class of molecules designed to mimic natural peptides of the innate immune system while exhibiting improved drug-like properties. They are regarded as promising alternatives to conventional antibiotics due to their potent bactericidal activity, their ability to disrupt biofilms, as well as for their broad-spectrum efficacy against diverse pathogens, coupled with a reduced propensity for resistance development. Advances in peptide engineering have further enhanced their therapeutic potential by improving stability, bioavailability, and safety through strategies such as incorporation of D-amino acids, cyclization, lipidation, and the design of peptidomimetics. Compared to natural AMPs, which are rapidly degraded by proteases and may exhibit cytotoxicity at high concentrations, synthetic peptides demonstrate greater tolerability and sustained activity in biological environments [142,146,147,148]. The development of synthetic AMPs is one of the most active areas in the development of modern antimicrobial drugs, with numerous AMP-based therapeutic candidates being currently under preclinical and clinical evaluation. Several well-studied molecules, including those known under the names Omiganan, Pexiganan, and Murepavadin, have advanced to Phase II or Phase III clinical trials, and examples of these compounds are presented in the already mentioned Table 1 [149,150,151]. Thus, synthetic antimicrobial peptides extend the anti-infective potential of natural AMPs by combining enhanced stability and potency with targeted activity against multidrug-resistant intestinal pathogens.
Design Strategies and Structural Modifications of Synthetic Antimicrobial Peptides
Modern strategies for the design of synthetic AMPs rely on rational engineering approaches aimed at increasing their stability, biological activity, selectivity, and pharmacokinetic performance, with structural modifications leading to significantly optimized properties of natural peptides by reducing their toxicity and increasing their efficacy against infections caused by drug-resistant pathogens. [12,152,153]. One effective strategy for enhancing peptide stability involves the substitution of L-amino acids with D-amino acids. Because proteases typically recognize and cleave peptides composed of L-amino acids, incorporation of D-amino acids markedly increases peptide resistance to proteolytic degradation and prolongs their half-life in biological environments. In certain cases, this modification may also reduce immunogenicity. Moreover, D-amino acid substitution can stabilize secondary structures while preserving antimicrobial activity. Notable examples include the D-configured antimicrobial peptide D-IK8 and the D-modified immunoregulatory decapeptide RDP58, both of which exhibit improved stability while maintaining potent biological activity [154,155,156]. Another strategy involves cyclization of the polypeptide chain. Cyclization of AMPs, whether head-to-tail, side-chain-to-side-chain, disulfide bond–mediated, or lasso-like, restricts conformational flexibility, stabilizes the bioactive conformation, and protects the peptide backbone from enzymatic degradation. Consequently, cyclic AMPs frequently demonstrate improved bioavailability, enhanced membrane affinity, and prolonged in vivo activity. Representative examples include brilacidin, a non-peptidic defensin mimetic, and lariocidin, a ribosomally synthesized lasso peptide with exceptional structural stability, both of which exhibit broad-spectrum antibacterial activity [157,158,159]. Another strategy involves lipidation and aromatic enhancement. The attachment of fatty acid chains to the N-terminus or specific side chains of a peptide increases hydrophobicity, thereby strengthening interactions with bacterial membranes and promoting rapid membrane disruption. Lipidated AMPs and lipopeptides often display potent activity against otherwise difficult-to-treat strains, including biofilm-forming and multidrug-resistant bacteria, although their hydrophobicity must be carefully balanced to avoid excessive cytotoxicity. A representative example is HB1345, a short synthetic lipohexapeptide that demonstrates broad-spectrum antimicrobial and anti-inflammatory activity against skin pathogens, including Cutibacterium acnes, with relatively low cytotoxicity in preclinical models [160,161,162]. Another promising strategy involves the development of peptidomimetics and β- and γ-peptides. Peptidomimetics are fully or partially non-peptidic structures that replicate the key physicochemical properties of natural antimicrobial peptides, including cationic charge, amphiphilicity, and spatial separation of hydrophobic and hydrophilic domains. These molecules exhibit superior metabolic stability and pharmacokinetic profiles compared to natural AMPs. Incorporation of β- and γ-amino acids, as well as aryl-amide foldamers, enables the formation of stable secondary structures such as β-helices or helical foldamers that resist proteolytic degradation. Brilacidin (Figure 5a), a non-peptidic defensin mimetic based on an aryl-amide foldamer, disrupts bacterial membranes and has progressed to phase II clinical trials.
Similarly, XOMA-629 (Figure 5b), a synthetic peptide derived from bactericidal/permeability-increasing proteins, functions as an AMP mimetic with potent activity against Staphylococcus aureus (including MRSA) and has been clinically evaluated for impetigo [163,164,165,166]. Another methodology involves the use of hybrid and chimeric constructs. Hybrid AMPs are generated by combining functional fragments from different natural peptides, thereby integrating complementary activities such as membrane permeabilization, antibiofilm effects, and immunomodulation into a single molecule. In this context, a notable example is OP-145 (Figure 5c), a rationally designed 24-mer peptide derived from the human cathelicidin LL-37, in which the proteolytically unstable N-terminal region was removed and the remaining sequence optimized. Compared with its parent peptide, OP-145 exhibits improved stability, a more favorable therapeutic index, and potent activity against pathogens such as methicillin-resistant Staphylococcus aureus, while retaining key functional characteristics of LL-37. Collectively, strategies including D-amino acid substitution, cyclization, lipidation, peptidomimetic design (incorporating β- and γ-peptides), and hybrid constructs all concur to modern AMP engineering, driving the development of candidates with enhanced efficacy and drug-like properties suitable for clinical application [167,168,169,170] . The search for novel therapeutic approaches to address diseases of social importance emphasizes nature-inspired methods [171,172,173,174,175,176] , alongside molecular frameworks that integrate both natural and synthetic agents [177,178] . These include peptide- and oligonucleotide-based molecules, as well as hybrid constructs such as nucleopeptides [179,180,181,182,183,184,185,186]. In this context, the targeted synthesis of peptidomimetics and analogues containing non-protein amino acids has become a pivotal strategy to overcome stability issues. The incorporation of specific functional groups, such as thiazole, triazole, or fluorenyl-methoxycarbonyl derivatives, has been shown to significantly enhance biological activity [187,188,189,190,191] . Moreover, advanced synthetic methodologies, including the Michael addition reaction and the use of Ni-complexes, have enabled the creation of novel chiral amino acid derivatives and dipeptides that demonstrate potent antifungal and antibacterial activity against resistant pathogens [192,193]. Investigating the biomolecular interactions of these synthetic derivatives specifically their binding affinity to DNA and serum albumin provides further crucial insights into their therapeutic potential and pharmacokinetics [194,195] . In summary, synthetic AMPs are a critical component of modern strategies to combat infections caused by multidrug-resistant microorganisms, and as global antibiotic resistance grows, the demand for agents with alternative mechanisms and reduced susceptibility to resistance development increases. Synthetic AMPs meet these requirements by combining rapid membrane-disrupting activity, immunomodulatory properties, and a relatively low rate of resistance development, particularly when enhanced through rational design and chemical modifications, and their ability to simultaneously target multiple pathogenic pathways is a major advantage. In addition to pore formation and depolarization, many engineered peptides inhibit biofilm formation and eradicate mature biofilms, which is crucial for treating chronic infections associated with Staphylococcus aureus, Pseudomonas aeruginosa, and Candida species, while synthetically optimized molecules can also achieve pathogen-specific activity, opening therapeutic options against highly resistant P. aeruginosa strains. Several synthetic AMPs have progressed to advanced clinical evaluation, confirming their translational potential, and their multifaceted mechanisms, reduced risk of resistance compared to conventional antibiotics, biofilm-targeted activity, and synergistic effects with traditional antibiotics support their use as both monotherapy and adjunctive therapy for chronic, nosocomial, and recurrent infections. Moreover, modern drug delivery innovations, including nanoformulations, liposomal carriers, inhaled formulations, and transdermal systems, further enhance different AMP properties such as stability, bioavailability, and tissue targeting, thereby expanding their clinical applicability [196,197,198,199]. Thus, rational design and structural optimization of synthetic antimicrobial peptides enable the development of next-generation intestinal anti-infective agents with improved pharmacological properties and sustained efficacy in the gastrointestinal environment.
3. Challenges and Innovations in AMP Development
Antimicrobial peptides are important components of innate immunity, actively participating in the regulation of microbial homeostasis, fighting pathogenic infections, and maintaining the epithelial barrier. More in detail, natural AMPs can disrupt membranes and modify the immune system by neutralizing toxins, reducing inflammation, and accelerating mucosal healing. However, as numerous studies have shown, their clinical use is limited by rapid proteolysis, low stability, and poor bioavailability, particularly in the gastrointestinal tract [15,16,33,200,201]. These limitations have prompted the development of synthetic AMPs that can replicate the key biological effects of natural molecules while being more stable and pharmacologically predictable. Structural and chemical modifications have played a key role in the development of synthetic AMPs, including processes such as the introduction of D-amino acids, the creation of cyclic and lipidated peptides, the design of short cationic molecules, and the formation of hybrid structures and peptidomimetics [16,31,157,202,203]. These approaches have enabled the creation of compounds that are resistant to enzymatic degradation, yet retain activity under conditions of high ionic strength and in the presence of serum, bile, or mucus secretions. The use of these strategies has enabled the development of molecules such as those known under the names Brilacidin, OP-145, Pexiganan, Omiganan, PAC-113, and Murepavadin, which have demonstrated efficacy in phase II–III clinical trials [40,43,58,61]. It is important to emphasize that progress in the creation of synthetic AMPs has been made possible not only by chemical modifications, such us by the introduction of D-amino acids, cyclization, lipidation, and the creation of hybrid structures, but also by the emergence of large specialized databases containing the sequences, structural parameters, and biological activity of thousands of peptides. In particular, APD (Table 2) offers a well-annotated collection of natural AMPs, while CAMPR4 stands out for its very large dataset and integrated prediction tools. DRAMP provides one of the most comprehensive repositories, including detailed activity and toxicity annotations. BACTIBASE focuses specifically on bacteriocins, and MilkAMP compiles bioactive peptides derived from milk proteins. PhytAMP is dedicated to plant AMPs, whereas AVPdb and HIPdb specialize in antiviral and HIV-inhibiting peptides, respectively. Moreover, DBAASP uniquely links antimicrobial activity with structural data, while Hemolytik is particularly useful for evaluating hemolytic potential and safety profiles of peptides (Table 2). These resources provide a foundation for computer modeling, efficacy prediction, optimal motif selection, and in silico screening prior to laboratory synthesis, significantly accelerating the development of new peptides. Modern development of synthetic AMPs would be not straightforward without the use of global bioinformatics resources (Table 2). International antimicrobial peptide databases accumulate information on over 40,000 natural and synthetic molecules, enabling the systematization of structural motifs, analysis of physicochemical parameters, and in silico efficacy prediction. These databases serve as the basis for machine learning algorithms, 3D modeling, structural docking, and computational design of new peptides, significantly reducing the time and cost of searching for active candidates [204,205,206,207,208] .
By integrating sequence–structure–activity relationship analyses, researchers can assess peptide toxicity, hydrophobicity, amphiphilicity, charge distribution, and the structural domains that determine antibacterial spectrum. These resources facilitate the rational design of synthetic molecules with high efficacy, low resistance potential, and improved selectivity for therapeutic applications. However, despite significant progress, several challenges remain, including the high cost of peptide synthesis, limited in vivo efficacy, the need for improved pharmacokinetics, potential cytotoxicity at elevated concentrations, and insufficient knowledge of AMP effects on commensal microbiota during long-term treatment [209,210,211]. In this context, nanotechnology-based delivery systems, such as liposomal formulations, solid lipid nanoparticles (SLNs), polymer capsules, and targeted carriers, represent a promising approach to enhance local AMP concentrations, reduce toxicity, and prolong therapeutic activity. In parallel, artificial intelligence and machine learning will contribute accelerating AMP sequence optimization, thus, reducing attrition rates among candidate molecules. Collectively, the advances summarized in this work demonstrate the growing importance of natural and synthetic AMPs as next-generation antimicrobial drug candidates capable of addressing the global rise in antibiotic resistance. Consequently, continued progress in peptide delivery systems will depend on the synergistic integration of computational methods, bioengineering, clinical trials, and emerging technologies [212,213] .
Structure–Activity Relationships and Selectivity of Antimicrobial Peptides
Understanding the structure–activity relationship (SAR) [214] of antimicrobial peptides is essential for designing molecules with potent activity and acceptable safety profiles. Sequence-based templates have proven particularly useful in this regard, as they guide the rational design of new analogues and enable the identification of promising peptide fragments within protein databases. For example, applying a helical template to database searches allowed the discovery of an N-terminal segment of pilosulin 1 that, once synthesized, exhibited broad-spectrum antimicrobial activity. [215] Subsequent targeted substitutions further enhanced antibacterial potency while reducing hemolysis, illustrating how small structural adjustments can markedly improve selectivity. A central determinant of AMP activity is their interaction with lipid membranes. For α-helical peptides, membrane engagement aligns with carpet-like mechanisms of disruption, and experimental correlations between peptide structuring in membrane-mimetic environments and bacterial permeabilization support this model. Features such as hydrophobic–polar face distribution, insertion depth, and the snorkel effect strongly influence efficacy. These same structural properties also shape cytotoxicity toward host cells, as conformational stability and hydrophobicity can shift the balance between antimicrobial potency and safety. [215]
Selectivity studies further highlight the importance of SAR-guided design. AMPs differ widely in their ability to discriminate between bacterial membranes and mammalian intestinal cells. For example, gallidermin demonstrates high selectivity, combining strong antimicrobial activity with minimal cytotoxicity and negligible hemolysis [216]. On the other hand, nisin A shows moderate selectivity, remaining safe at antimicrobial concentrations but becoming harmful only at much higher doses. Magainins occupy an intermediate position, displaying more cytotoxicity than gallidermin or nisin but without causing major structural damage to intestinal cells. In contrast, melittin exhibits poor selectivity, as its antimicrobial concentrations overlap with those that induce hemolysis, epithelial barrier disruption, and loss of microvilli. These comparisons underscore that only certain AMPs possess the membrane specificity required for therapeutic use, while others, such as melittin, may be too broadly cytolytic for safe antimicrobial application but could still serve alternative purposes, including drug-delivery enhancement [216].
4. Conclusions
Antimicrobial peptides remain one of the most promising classes for the development of next-generation anti-infective agents, particularly in intestinal disease where conventional antibiotics are losing efficacy. Natural and synthetic AMPs combine broad activity against multidrug-resistant pathogens with biofilm disruption and immunomodulatory effects, while modern design strategies, such as incorporation of D-amino acids, cyclization, lipidation, and peptidomimetics, have markedly improved their stability, selectivity, and clinical potential. Despite persisting challenges in delivery and pharmacokinetics, recent advances demonstrate steady progress toward clinical translation, supported by innovations in nanotechnology, combination therapy, and personalized medicine. A central theme emerging from this review is the role of antimicrobial peptides as integrated intestinal anti-infective agents that act beyond simple pathogen killing. Both natural and synthetic AMPs contribute to intestinal defense by combining direct antimicrobial activity against enteric pathogens with immunomodulatory and barrier-protective functions. Epithelial peptides such as defensins, cathelicidin LL-37, and RegIII lectins not only restrict the growth and invasion of gastrointestinal pathogens but also regulate host–microbiota interactions, enforce spatial segregation between microbes and the epithelium, and prevent excessive inflammatory responses that exacerbate intestinal disease. At the molecular level, the conserved structural features of natural AMPs enable rapid targeting of fundamental membrane properties, while synthetic peptides further extend this anti-infective potential through enhanced stability, bioavailability, and activity against multidrug-resistant organisms. Together, these properties position antimicrobial peptides as a promising class of next-generation intestinal anti-infective agents capable of addressing both microbial eradication and preservation of intestinal homeostasis. In our opinion, the anti-infective role of short AMP sequences should be also explored, as compact peptides may retain antimicrobial and immunomodulatory activity while offering superior manufacturability, in some cases a reduced toxicity, and easier integration into delivery systems. This novel direction, together with synergistic use alongside traditional antibiotics, positions AMPs not only as adjuncts but as central components of future infection-control strategies in the face of escalating antibiotic resistance.
Author Contributions
Conceptualization, T.S. and G.N.R.; methodology, A.G., J.S. and K.H.; validation, A.D., L.S., M.I. and R.P.; data curation, A.T. and L.S.; writing—original draft preparation, T.S. and G.N.R.; writing—review and editing, G.N.R., R.P. and T.S.; visualization, A.D., M.I. and K.H.; supervision, G.N.R. All authors have read and agreed to the published version of the manuscript.
Funding
The authors express their gratitude to the Science Committee of the Republic of Armenia for supporting their research within the framework of the project № 24RL-1D014 and № 25SC-CNR-1D008.
Data Availability Statement
Not applicable.
Acknowledgments
All authors would like to express their sincere gratitude to the Higher Education and Science Committee of the RA Ministry of Education, Science, Culture, and Sports of Armenia for their support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Full name / Meaning |
| AMP | Antimicrobial Peptide |
| CAMP | Cationic Antimicrobial Peptide |
| HD-5 / HD-6 | Human Defensin 5 / 6 |
| LL-37 | Human Cathelicidin Peptide |
| HBD | Human Beta-Defensin |
| MDR | Multidrug-Resistant |
| XDR | Extensively Drug-Resistant |
| MRSA | Methicillin-Resistant Staphylococcus aureus |
| SSTI | Skin and Soft Tissue Infections |
| LPS | Lipopolysaccharide |
| LTA | Lipoteichoic Acid |
| QS | Quorum Sensing |
| GI Tract | Gastrointestinal Tract |
| IBD | Inflammatory Bowel Disease |
| NEC | Necrotizing Enterocolitis |
| PRR | Pattern-Recognition Receptor |
| TLR | Toll-Like Receptor |
| NF-κB | Nuclear Factor kappa B |
| MAPK | Mitogen-Activated Protein Kinase |
| CFU | Colony-Forming Unit |
| MIC | Minimum Inhibitory Concentration |
| IDR | Innate Defense Regulator |
| PAMPs | Pathogen-Associated Molecular Patterns |
| APC | Antigen-Presenting Cell |
| IEC | Intestinal Epithelial Cells |
| SCFA | Short-Chain Fatty Acids |
| ROS | Reactive Oxygen Species |
References
- World Health Organization. Global antimicrobial resistance and use surveillance system (GLASS) report: 2022; WHO Press: Geneva, Switzerland, 2022; Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 10 November 2025).
- Wang, B.; Chen, D.; Chen, H.; Wu, W.; Cheng, K.; Tao, Y.; Zhang, L.; Liu, C.; Ou, D.; Zhang, M.; Tang, X.; Wang, S.; Wang, G.; Luo, B. Global, regional, and national incidence and mortality for enteric infections from 1990 to 2019. BMC Public Health. 2025, 25, 100. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; UK Government: London, UK, 2016; Available online: https://amr-review.org (accessed on 9 November 2025).
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; Merkens, W.; Mielke, M.; Oltmanns, P.; Ross, B.; Rotter, M.; Schmithausen, R.M.; Sonntag, H.G.; Trautmann, M. Antibiotic Resistance: What Is So Special about Multidrug-Resistant Gram-Negative Bacteria? GMS Hygiene and Infection Control 2017, 12, Doc05. [Google Scholar] [CrossRef] [PubMed]
- Birlutiu, V.; Birlutiu, R.-M. An Overview of the Epidemiology of Multidrug Resistance and Bacterial Resistance Mechanisms: What Solutions Are Available? A Comprehensive Review. Microorganisms 2025, 13, 2194. [Google Scholar] [CrossRef]
- Morris, S.; Cerceo, E. Trends, Epidemiology, and Management of Multi-Drug Resistant Gram-Negative Bacterial Infections in the Hospitalized Setting. Antibiotics 2020, 9, 196. [Google Scholar] [CrossRef]
- Mouanga-Ndzime, Y.; Bisseye, C.; Longo-Pendy, N.-M.; Bignoumba, M.; Dikoumba, A.-C.; Onanga, R. Trends in Escherichia coli and Klebsiella pneumoniae Urinary Tract Infections and Antibiotic Resistance over a 5-Year Period in Southeastern Gabon. Antibiotics 2025, 14, 14. [Google Scholar] [CrossRef]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial Resistance: Impacts, Challenges, and Future Prospects. Global Medicine 2024, 100081. [Google Scholar] [CrossRef]
- Oliveira, M.; Antunes, W.; Mota, S.; Madureira-Carvalho, Á.; Dinis-Oliveira, R.J.; Dias da Silva, D. An Overview of the Recent Advances in Antimicrobial Resistance. Microorganisms 2024, 12, 1920. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; Salamat, M.K.F.; Baloch, Z. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Ali, M.; Garg, A.; Srivastava, A.; Arora, P.K. The Role of Antimicrobial Peptides in Overcoming Antibiotic Resistance. Microbe 2025, 7, 100337. [Google Scholar] [CrossRef]
- Gong, T.; Fu, J.; Shi, L.; Chen, X.; Zong, X. Antimicrobial Peptides in Gut Health: A Review. Front. Nutr. 2021, 8, 751010. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.I.; Mergani, A.; Aklilu, E.; Kamaruzzaman, N.F. Antimicrobial Peptides: Bringing Solution to the Rising Threats of Antimicrobial Resistance in Livestock. Front. Vet. Sci. 2022, 9, 851052. [Google Scholar] [CrossRef] [PubMed]
- del Olmo, M.; Andreu, C. Current Status of the Application of Antimicrobial Peptides and Their Conjugated Derivatives. Molecules 2025, 30, 3070. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Gao, F.G.; Wang, L.; Zhu, X.Q.; Wang, R.; Qin, Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Sharma, D.; Dhiman, I.; Das, S.; Das, D.K.; Pramanik, D.D.; Dash, S.K.; Pramanik, A. Recent Advances in Therapeutic Peptides: Innovations and Applications in Treating Infections and Diseases. ACS Omega 2025, 10, 17087–17107. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Niekerk, L.-A.; Aina, O.; Abiona, A.; Barker, A.M.; Basson, G.; Nkomo, M.; Otomo, L.; Keyster, M.; et al. Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides. Int. J. Mol. Sci. 2023, 24, 11864. [Google Scholar] [CrossRef]
- Zhang, H.; Lv, J.; Ma, Z.; Ma, J.; Chen, J. Advances in Antimicrobial Peptides: Mechanisms, Design Innovations, and Biomedical Potential. Molecules 2025, 30, 1529. [Google Scholar] [CrossRef]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignani, C.; Rao, P.; Wool, E.; Johnson, S.C.; Browne, A.J.; Chipeta, M.G.; Fell, F.; Hackett, S.; Haines-Woodhouse, G.; Kashef Hamadani, B.H.; Kumaran, E.A.P.; McManigal, B.; Agarwal, R.; Akech, S.; Albertson, S.; Amuasi, J.; Andrews, J.; Aravkin, A.; Ashley, E.; Bailey, F.; Baker, S.; Basnyat, B.; Bekker, A.; Bender, R.; Bethou, A.; Bielicki, J.; Boonkasidecha, S.; Bukosia, J.; Carvalheiro, C.; Castañeda-Orjuela, C.; Chansamouth, V.; Chaurasia, S.; Chiurchiù, S.; Chowdhury, F.; Cook, A.J.; Cooper, B.; Cressey, T.R.; Criollo-Mora, E.; Cunningham, M.; Darboe, S.; Day, N.P.J.; Feasey, N.; Musicha, P. Global burden of bacterial antimicrobial resistance in 2019. a systematic analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Z.; Zhang, Q.; Xu, N.; Lu, T.; Wang, T.; Hong, W.; Fu, Z.; Penuelas, J.; Gillings, M.; Qian, H. Antimicrobial Peptides in the Global Microbiome: Biosynthetic Genes and Resistance Determinants. Environ. Sci. Technol. 2023, 57, 7698–7708. [Google Scholar] [CrossRef]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial Peptides: Features, Action, and Their Resistance Mechanisms in Bacteria. Microb. Drug Resist. 2018, 24. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Hall, K.; Aguilar, M.-I. Antimicrobial Peptide Structure and Mechanism of Action: A Focus on the Role of Membrane Structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; Derengowski, L.S.; Silva-Pereira, I.; Kyaw, C.M. Antibiotic Development Challenges: The Various Mechanisms of Action of Antimicrobial Peptides and of Bacterial Resistance. Front. Microbiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Xiong, L.; Wu, X.; Chu, M.; Bao, P.; Ge, Q.; Guo, X. Bovine Lactoferricin Exerts Antibacterial Activity against Four Gram-Negative Pathogenic Bacteria by Transforming Its Molecular Structure. Front. Cell. Infect. Microbiol. 2025, 15, 1508895. [Google Scholar] [CrossRef]
- De Smet, K.; Contreras, R. Human Antimicrobial Peptides: Defensins, Cathelicidins and Histatins. Biotechnol. Lett. 2005, 27, 1337–1347. [Google Scholar] [CrossRef]
- Morici, P.; Florio, W.; Rizzato, C.; et al. Synergistic Activity of Synthetic N-Terminal Peptide of Human Lactoferrin in Combination with Various Antibiotics against Carbapenem-Resistant Klebsiella pneumoniae Strains. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1739–1748. [Google Scholar] [CrossRef]
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin, S.F.; Raza, S.Q.; Zohaib, S.; Sefat, F.; Zafar, M.S. Histatin Peptides: Pharmacological Functions and Their Applications in Dentistry. Saudi Pharm. J. 2017, 25, 25–31. [Google Scholar] [CrossRef]
- Fassi, E.M.A.; Moretti, R.M.; Montagnani Marelli, M.; Garofalo, M.; Gori, A.; Pesce, C.; Albani, M.; Milano, E.G.; Sgrignani, J.; Cavalli, A.; et al. FYCO1 Peptide Analogs: Design and Characterization of Autophagy Inhibitors as Co-Adjuvants in Taxane Chemotherapy of Prostate Cancer. Int. J. Mol. Sci. 2025, 26, 5365. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Abdelkhalek, A.; Seleem, M.N. Evaluation of short synthetic antimicrobial peptides for treatment of drug-resistant and intracellular Staphylococcus aureus. Sci. Rep. 2016, 6, 29707. [Google Scholar] [CrossRef]
- Parra, A.L.; Bezerra, L.P.; Shawar, D.E.; Neto, N.A.; Mesquita, F.P.; da Silva, G.O.; Souza, P.F. Synthetic antiviral peptides: A new way to develop targeted antiviral drugs. Future Virol. 2022, 17, 577–591. [Google Scholar] [CrossRef]
- Al Tall, Y.; Alkurdi, Y.; Alshraiedeh, N.; Sabi, S.H. Designing Novel Antimicrobial Agents from the Synthetic Antimicrobial Peptide (Pep-38) to Combat Antibiotic Resistance. Pharmaceuticals 2025, 18, 862. [Google Scholar] [CrossRef] [PubMed]
- Mercer, D.K.; O’Neil, D.A. Innate inspiration: Antifungal peptides and other immunotherapeutics from the host immune response. Front. Immunol. 2020, 11, 2177. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.-X.; Zou, Y.-L.; Duan, J.-J.; Jia, Z.-R.; Li, X.-J.; Wang, Z.; et al. The synthetic melanocortin (CKPV)2 exerts anti-fungal and anti-inflammatory effects against Candida albicans vaginitis via inducing macrophage M2 polarization. PLoS ONE 2013, 8, e56004. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjya, S.; Mohid, S.A.; Bhunia, A. Atomic-Resolution Structures and Mode of Action of Clinically Relevant Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 4558. [Google Scholar] [CrossRef]
- Lesiak, A.; Paprocka, P.; Wnorowska, U.; Mańkowska, A.; Król, G.; Głuszek, K.; Piktel, E.; Spałek, J.; Okła, S.; Fiedoruk, K.; Durnaś, B.; Bucki, R. Significance of host antimicrobial peptides in the pathogenesis and treatment of acne vulgaris. Front. Immunol. 2024, 15, 1502242. [Google Scholar] [CrossRef]
- Brouwer, C.; van der Linden, Y.; Carrasco, M.R.; Alwasel, S.; Abalkhail, T.; Al-Otibi, F.O.; Boekhout, T.; Welling, M.M. Synthetic Human Lactoferrin Peptide hLF(1-11) Shows Antifungal Activity and Synergism with Fluconazole and Anidulafungin Towards Candida albicans and Various Non-Albicans Candida Species, Including Candidozyma auris. Antibiotics 2025, 14, 671. [Google Scholar] [CrossRef]
- Seo, M.-D.; Won, H.-S.; Kim, J.-H.; Mishig-Ochir, T.; Lee, B.-J. Antimicrobial Peptides for Therapeutic Applications: A Review. Molecules 2012, 17, 12276–12286. [Google Scholar] [CrossRef]
- Kumar, N.; Bhagwat, P.; Singh, S.; Pillai, S. A Review on the Diversity of Antimicrobial Peptides and Genome Mining Strategies for Their Prediction. Biochimie 2024, 227, 99–115. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Madera, L.; Afacan, N.; Okumura, K.; Ogawa, H.; Hancock, R.E. The Innate Defense Regulator Peptides IDR-HH2, IDR-1002, and IDR-1018 Modulate Human Neutrophil Functions. J. Leukoc. Biol. 2013, 94, 159–170. [Google Scholar] [CrossRef]
- Ming, L.; Huang, J.-A. The Antibacterial Effects of Antimicrobial Peptides OP-145 against Clinically Isolated Multi-Resistant Strains. Jpn. J. Infect. Dis. 2017, 70, 601–603. [Google Scholar] [CrossRef]
- Cheng, K.-T.; Wu, C.-L.; Yip, B.-S.; Chih, Y.-H.; Peng, K.-L.; Hsu, S.-Y.; Yu, H.-Y.; Cheng, J.-W. The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance. Int. J. Mol. Sci. 2020, 21, 2654. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Li, X.S.; Sun, J.N.; Edgerton, M. The P-113 Fragment of Histatin 5 Requires a Specific Peptide Sequence for Intracellular Translocation in Candida albicans, Which Is Independent of Cell Wall Binding. Antimicrob. Agents Chemother. 2008, 52, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Abdelsattar, A.S.; Abutaleb, N.S.; Seleem, M.N. A Novel Peptide Mimetic, Brilacidin, for Combating Multidrug-Resistant Neisseria gonorrhoeae. PLoS ONE 2025, 20, e0325722. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.R.; Varela, C.L.; Pires, A.S.; Tavares-da-Silva, E.J.; Roleira, F.M.F. Synthetic and Natural Guanidine Derivatives as Antitumor and Antimicrobial Agents: A Review. Bioorg. Chem. 2023, 138, 106600. [Google Scholar] [CrossRef]
- Hetta, H.F.; Sirag, N.; Alsharif, S.M.; Alharbi, A.A.; Alkindy, T.T.; Alkhamali, A.; Albalawi, A.S.; Ramadan, Y.N.; Rashed, Z.I.; Alanazi, F.E. Antimicrobial Peptides: The Game-Changer in the Epic Battle Against Multidrug-Resistant Bacteria. Pharmaceuticals 2024, 17, 1555. [Google Scholar] [CrossRef]
- Odunitan, T.T.; Apanisile, B.T.; Afolabi, J.A.; Adeniwura, P.O.; Akinboade, M.W.; Ibrahim, N.O.; Alare, K.P.; Saibu, O.A.; Adeosun, O.A.; Opeyemi, H.S.; Ayiti, K.S. Beyond Conventional Drug Design: Exploring the Broad-Spectrum Efficacy of Antimicrobial Peptides. Chem. Biodivers. 2024, e202401349. [Google Scholar] [CrossRef]
- Edwards, I.A.; Henriques, S.T.; Blaskovich, M.A.T.; Elliott, A.G.; Cooper, M.A. Investigations into the Membrane Activity of Arenicin Antimicrobial Peptide AA139. Biochim. Biophys. Acta Gen. Subj. 2022, 1866, 130156. [Google Scholar] [CrossRef]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Johannes Zuegg, J.; Edwards, I.; K. Cain, A.; J. Boinett, Ch.; Barquist, L.; Lundberg, C.V.; Steen, J.; Butler, M.S.; Mobli, M.; Porter, K.M.; Blaskovich, M.A.T.; Lociuro, S.; Strandh, M.; Cooper, M.A. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef]
- Bourreille, A.; Doubremelle, M.; Raingeard de la Blétière, D.; Segain, J.P.; Toquet, C.; Buelow, R.; Galmiche, J.P. RDP58, a Novel Immunomodulatory Peptide with Anti-Inflammatory Effects: A Pharmacological Study in Trinitrobenzene Sulphonic Acid Colitis and Crohn Disease. Scand. J. Gastroenterol. 2003, 38, 526–532. [Google Scholar] [CrossRef]
- De Vry, C.G.; Valdez, M.; Lazarov, M.; Muhr, E.; Buelow, R.; Fong, T.; Iyer, S. Topical Application of a Novel Immunomodulatory Peptide, RDP58, Reduces Skin Inflammation in the Phorbol Ester-Induced Dermatitis Model. J. Investig. Dermatol. 2005, 125, 473–481. [Google Scholar] [CrossRef]
- Iseganan, I.B. IB 367, Protegrin IB 367. Drugs R D 2002, 3, 52–55. [Google Scholar] [CrossRef]
- Bolosov, I.A.; Panteleev, P.V.; Sychev, S.V.; Khokhlova, V.A.; Safronova, V.N.; Toropygin, I.Y.; Kombarova, T.I.; Korobova, O.V.; Pereskokova, E.S.; Borzilov, A.I.; et al. Design of Protegrin-1 Analogs with Improved Antibacterial Selectivity. Pharmaceutics 2023, 15, 2047. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Gomes, P. Peptides and Peptidomimetics in the Development of Hydrogels toward the Treatment of Diabetic Wounds. Curr. Res. Biotechnol. 2025, 9, 100292. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Janson, H.W.; Wold, H.; Fugelli, A.; Andersson, K.; Håkangård, C.; Olsson, P.; Olsen, W.M. LTX-109 Is a Novel Agent for Nasal Decolonization of Methicillin-Resistant and -Sensitive Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59. [Google Scholar] [CrossRef]
- Midura-Nowaczek, K.; Markowska, A. Antimicrobial Peptides and Their Analogs: Searching for New Potential Therapeutics. Perspect. Med. Chem. 2014, 6, PMC.S13215. [Google Scholar] [CrossRef]
- Wang, S.; Ong, Z.Y.; Qu, S.; Wang, Y.; Xin, J.; Zheng, Z.; Wu, H. Omiganan-Based Synthetic Antimicrobial Peptides for the Healthcare of Infectious Endophthalmitis. ACS Biomater. Sci. Eng. 2024, 10, 7217–7226. [Google Scholar] [CrossRef]
- Rodríguez-Castaño, G.P.; Rosenau, F.; Ständker, L.; Firacative, C. Antimicrobial Peptides: Avant-Garde Antifungal Agents to Fight against Medically Important Candida Species. Pharmaceutics 2023, 15, 789. [Google Scholar] [CrossRef]
- Zhang, X.-L.; Jiang, A.-M.; Ma, Z.-Y.; Li, X.-B.; Xiong, Y.-Y.; Dou, J.-F.; Wang, J.-F. The Synthetic Antimicrobial Peptide Pexiganan and Its Nanoparticles (PNPs) Exhibit the Anti-Helicobacter pylori Activity in Vitro and in Vivo. Molecules 2015, 20, 3972–3985. [Google Scholar] [CrossRef]
- Gottler, L.M.; Ramamoorthy, A. Structure, Membrane Orientation, Mechanism, and Function of Pexiganan—A Highly Potent Antimicrobial Peptide Designed from Magainin. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1680–1686. [Google Scholar] [CrossRef]
- Li, X.; Hao, Y.; Yang, N.; Mao, R.; Teng, D.; Wang, J. Plectasin: From Evolution to Truncation, Expression, and Better Druggability. Front. Microbiol. 2023, 14, 1304825. [Google Scholar] [CrossRef]
- Xiong, Y.Q.; Hady, W.A.; Deslandes, A.; Rey, A.; Fraisse, L.; Kristensen, H.; Yeaman, M.R.; Bayer, A.S. Efficacy of NZ2114, a Novel Plectasin-Derived Cationic Antimicrobial Peptide Antibiotic, in Experimental Endocarditis Due to Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventós, D.S.; Neve, S.; Ravn, B.; Bonvin, A.M.; De Maria, L.; Andersen, A.S.; Gammelgaard, L.K.; Sahl, H.G.; Kristensen, H.H. Plectasin, a Fungal Defensin, Targets the Bacterial Cell Wall Precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef]
- Sader, H.S.; Dale, G.E.; Rhomberg, P.R.; Flamm, R.K. Antimicrobial Activity of Murepavadin Tested against Clinical Isolates of Pseudomonas aeruginosa from the United States, Europe, and China. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- García-Gros, J.; Cajal, Y.; Marqués, A.M.; Rabanal, F. Synthesis of the Antimicrobial Peptide Murepavadin Using Novel Coupling Agents. Biomolecules 2024, 14, 526. [Google Scholar] [CrossRef] [PubMed]
- Hernández-García, M.; Barbero-Herranz, R.; Bastón-Paz, N.; Díez-Aguilar, M.; López-Collazo, E.; Márquez-Garrido, F.J.; Hernández-Pérez, J.M.; Baquero, F.; Ekkelenkamp, M.B.; Fluit, A.C.; Fuentes-Valverde, V.; Moscoso, M.; Bou, G.; del Campo, R.; Cantón, R.; Avendaño-Ortiz, J. Unravelling the Mechanisms Causing Murepavadin Resistance in Pseudomonas aeruginosa: Lipopolysaccharide Alterations and Its Consequences. Front. Cell. Infect. Microbiol. 2024, 14, 1446626. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Song, Y.; Liu, Z.; Tang, X.; Tan, H. LC-AMP-I1, a Novel Venom-Derived Antimicrobial Peptide from the Wolf Spider Lycosa coelestis. Antimicrob. Agents Chemother. 2025, 69, e00424-24. [Google Scholar] [CrossRef]
- Ohtsuka, Y.; Inagaki, H. In Silico Identification and Functional Validation of Linear Cationic α-Helical Antimicrobial Peptides in the Ascidian Ciona intestinalis. Sci. Rep. 2020, 10, 12619. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Liu, X.; Yu, S.; Tang, X.; Tan, H. LC-AMP-F1 Derived from the Venom of the Wolf Spider Lycosa coelestis, Exhibits Antimicrobial and Antibiofilm Activities. Pharmaceutics 2024, 16, 129. [Google Scholar] [CrossRef]
- Jangra, M.; Travin, D.Y.; Aleksandrova, E.V.; Kaur, M.; Darwish, L.; Koteva, K.; Klepacki, D.; Wang, W.; Tiffany, M.; Sokaribo, A.; Chen, X.; Deng, Z.; Tao, M.; Coombes, B.K.; Vázquez-Laslop, N.; Polikanov, Y.S.; Mankin, A.S.; Wright, G.D. A Broad-Spectrum Lasso Peptide Antibiotic Targeting the Bacterial Ribosome. Nature 2025, 640, 1022–1030. [Google Scholar] [CrossRef]
- Thakur, K.M.; Gadewar, C.K. Lariocidin A: A Promising Frontier in the Battle Against Antimicrobial Resistance—A Comprehensive Review. Int. J. Innov. Res. Technol. 2025, 11, 5098. [Google Scholar]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Mietzner, T.A.; Montelaro, R.C. Rational Design of Engineered Cationic Antimicrobial Peptides Consisting Exclusively of Arginine and Tryptophan, and Their Activity against Multidrug-Resistant Pathogens. Antimicrob. Agents Chemother. 2013, 57, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Buccini, D.F.; Cardoso, M.H.; Franco, O.L. Antimicrobial Peptides and Cell-Penetrating Peptides for Treating Intracellular Bacterial Infections. Front. Cell. Infect. Microbiol. 2021, 10, 612931. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Aristizábal, I.; Ocampo-Ibáñez, I.D. Antimicrobial Peptides with Antibacterial Activity against Vancomycin-Resistant Staphylococcus aureus Strains: Classification, Structures, and Mechanisms of Action. Int. J. Mol. Sci. 2021, 22, 7927. [Google Scholar] [CrossRef]
- Ong, Z.Y.; Cheng, J.; Huang, Y.; Xu, K.; Ji, Z.; Fan, W.; Yang, Y.Y. Effect of Stereochemistry, Chain Length and Sequence Pattern on Antimicrobial Properties of Short Synthetic β-Sheet Forming Peptide Amphiphiles. Biomaterials 2014, 35, 1315–1325. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Li, S.; Han, Y.L.; Shi, Y.F.; Wu, Y.Y.; Cheng, J.; Wang, C.Y.; Zhou, X.Y.; Zhang, Y.X. De novo-designed amphiphilic α-helical peptide Z2 exhibits broad-spectrum antimicrobial, anti-biofilm, and anti-inflammatory efficacy in acute Pseudomonas aeruginosa pneumonia. Bioorg. Chem. 2025, 157, 108309. [Google Scholar] [CrossRef]
- Pashinina, O.A.; Kartashova, O.L.; Pashkova, T.M.; Gritsenko, V.A. Influence of ZP2 Peptide, a Synthetic Analogue of the GM-GSF Active Center, on the Anticytokine Activity of Bacteria from Enterococcus Genus and Their Ability to Produce Cytokine-Like Substances. Russ. J. Immunol. 2022, 25, 477–484. [Google Scholar] [CrossRef]
- Nishio, S.; Emori, C.; Wiseman, B.; Fahrenkamp, D.; Dioguardi, E.; Zamora-Caballero, S.; Bokhove, M.; Han, L.; Stsiapanava, A.; Algarra, B.; Lu, Y.; Kodani, M.; Bainbridge, R.E.; Komondor, K.M.; Carlson, A.E.; Landreh, M.; de Sanctis, D.; Yasumasu, S.; Ikawa, M.; Jovine, L. ZP2 Cleavage Blocks Polyspermy by Modulating the Architecture of the Egg Coat. Cell. 2024, 187, 1440–1459.e24. [Google Scholar] [CrossRef]
- Kogoniya, L.M.; Orlova, R.V. Glutoxim: new about known. Supportive Therapy in Oncology (In Russ.). 2024, 1, 43–51. [Google Scholar] [CrossRef]
- Bugaev, P.A.; Basharin, V.A.; Antushevich, A.E. Comparative Effectiveness of Molixan and Glutoxim in the Model of Toxic Liver Lesions with Anti-Tuberculosis Drugs. Rev. Clin. Pharmacol. Drug Ther. 2019, 17, 49–54. [Google Scholar] [CrossRef]
- Du, Q.-L.; Zhang, Z.-X.; Zhang, T.; Dong, C.-M. Research Progress on the Defensin Acipensins1. Microenvironment Microecol. Res. 2022, 4, 10. [Google Scholar] [CrossRef]
- Shamova, O.V.; Orlov, D.S.; Balandin, S.V.; Shramova, E.I.; Tsvetkova, E.V.; Panteleev, P.V.; Leonova, Y.F.; Tagaev, A.A.; Kokryakov, V.N.; Ovchinnikova, T.V. Acipensins—Novel Antimicrobial Peptides from Leukocytes of the Russian SturgeonAcipenser gueldenstaedtii. Acta Naturae 2014, 6, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Meštrović, T.; Juzbašić, M.; Tomas, M.; Erić, S.; Horvat Aleksijević, L.; Bekić, S.; Schwarz, D.; Matić, S.; Neuberg, M.; Škrlec, I. Antimicrobial Peptides—Mechanisms of Action, Antimicrobial Effects and Clinical Applications. Antibiotics 2022, 11, 1417. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Combination Effects of Antimicrobial Peptides. Antimicrob. Agents Chemother. 2016, 60, 1717–1727. [Google Scholar] [CrossRef]
- Sadeeq, M.; Li, Y.; Wang, C.; Hou, F.; Zuo, J.; Xiong, P. Unlocking the Power of Antimicrobial Peptides: Advances in Production, Optimization, and Therapeutics. Front. Cell. Infect. Microbiol. 2025, 15, 1528583. [Google Scholar] [CrossRef]
- Bizzotto, E.; Zampieri, G.; Treu, L.; Filannino, P.; Di Cagno, R.; Campanaro, S. Classification of Bioactive Peptides: A Systematic Benchmark of Models and Encodings. Comput. Struct. Biotechnol. J. 2024, 23, 2442–2452. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World Journal of Gastroenterology 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Kastl, A.J., Jr.; Terry, N.A.; Wu, G.D.; Albenberg, L.G. The Structure and Function of the Human Small Intestinal Microbiota: Current Understanding and Future Directions. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 33–45. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell. 2012, 148, 1258–1270. [Google Scholar] [CrossRef]
- Giuliani, A.; Pirri, G.; Rinaldi, A.C. Antimicrobial Peptides: The LPS Connection. Methods Mol. Biol. 2010, 618, 137–154. [Google Scholar] [CrossRef]
- Gutierrez, A.; Pucket, B.; Engevik, M.A. Bifidobacterium and the Intestinal Mucus Layer. Microbiome Res. Rep. 2023, 2, 36. [Google Scholar] [CrossRef]
- Heimlich, D.R.; Harrison, A.; Mason, K.M. Host Antimicrobial Peptides in Bacterial Homeostasis and Pathogenesis of Disease. Antibiotics 2014, 3, 645–676. [Google Scholar] [CrossRef]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front. Microbiol. 2018, 9, 736. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, K.F.; Herzberg, M.C. Antimicrobial Peptides: Defending the Mucosal Epithelial Barrier. Front. Oral Health 2022, 3, 958480. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Jin, M.; Xu, C.; Gao, Y.; Yang, Y.; Qi, H.; Zhang, Q.; Yang, X.; Ya, W.; Zhang, Y.; Yang, R. FABP4 in Paneth Cells Regulates Antimicrobial Protein Expression to Reprogram Gut Microbiota. Gut Microbes 2022, 14, 2139978. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Bevins, C.L.; Ganz, T. Defensins and Other Antimicrobial Peptides and Proteins. In Mucosal Immunology, 3rd ed.; Mestecky, J., Lamm, M.E., Ogra, P.L., Strober, W., Bienenstock, J., McGhee, J.R., Mayer, L., Eds.; Academic Press: San Diego, CA, USA, 2005; pp. 95–110. [Google Scholar] [CrossRef]
- Fu, J.; Zong, X.; Jin, M.; Min, J.; Wang, F.; Wang, Y. Mechanisms and Regulation of Defensins in Host Defense. Signal Transduct. Target. Ther. 2023, 8, 300. [Google Scholar] [CrossRef]
- Chairatana, P.; Nolan, E.M. Human α-Defensin 6: A Small Peptide That Self-Assembles and Protects the Host by Entangling Microbes. Acc. Chem. Res. 2017, 50, 960–967. [Google Scholar] [CrossRef]
- Sankaran-Walters, S.; Hart, R.; Dills, C. Guardians of the Gut: Enteric Defensins. Front. Microbiol. 2017, 8, 647. [Google Scholar] [CrossRef]
- Cunliffe, R.N. α-Defensins in the Gastrointestinal Tract. Mol. Immunol. 2003, 40, 463–467. [Google Scholar] [CrossRef]
- Ehmann, D.; Wendler, J.; Koeninger, L.; Larsen, I.S.; Klag, T.; Berger, J.; Marette, A.; Schaller, M.; Stange, E.F.; Malek, N.P.; Jensen, B.A.H.; Wehkamp, J. Paneth Cell α-Defensins HD-5 and HD-6 Display Differential Degradation into Active Antimicrobial Fragments. Proc. Natl. Acad. Sci. USA 2019, 116, 3746–3751. [Google Scholar] [CrossRef]
- Cieślik, M.; Bagińska, N.; Andrzej Górski, A.; Jończyk-Matysiak, E. Human β-Defensin 2 and Its Postulated Role in Modulation of the Immune Response. Cells 2021, 10, 2991. [Google Scholar] [CrossRef] [PubMed]
- Esther, R.C.; Chadee, K. Antimicrobial Human β-Defensins in the Colon and Their Role in Infectious and Non-Infectious Diseases. Pathogens 2013, 2, 177–192. [Google Scholar] [CrossRef]
- Danielian, A.; Danielian, M.; Y. Cheng, M.; Burton, J.; S. Han, P.; Kerr, R.P.R. Antiaging Effects of Topical Defensins. Facial Plastic Surgery Clinics of North America 2023, 31, 535–546. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; C Fleming, D.; Critchley, H.; Kelly, W.R. Differential Expression of the Natural Antimicrobials, Beta-Defensins 3 and 4, in Human Endometrium. Journal of Reproductive Immunology 2003, 59, 1–16. [Google Scholar] [CrossRef]
- Riedl, S.; Zweytick, D.; Lohner, K. Membrane-active host defense peptides—challenges and perspectives for the development of novel anticancer drugs. Chemistry and Physics of Lipids 2011, 164, 766–781. [Google Scholar] [CrossRef]
- Ou, J.; Liang, S.; Guo, X.K.; Hu, X. α-Defensins Promote Bacteroides Colonization on Mucosal Reservoir to Prevent Antibiotic-Induced Dysbiosis. Frontiers in Immunology 2020, 11, 2065. [Google Scholar] [CrossRef]
- Duplantier, A.J.; van Hoek, M.L. The Human Cathelicidin Antimicrobial Peptide LL-37 as a Potential Treatment for Polymicrobial Infected Wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef]
- Nijnik, A.; Hancock, R.E.W. The Roles of Cathelicidin LL-37 in Immune Defences and Novel Clinical Applications. Curr. Opin. Hematol. 2009, 16, 41–47. [Google Scholar] [CrossRef]
- Moreno-Angarita, A.; Aragón, C.C.; Tobón, G.J. Cathelicidin LL-37: A New Important Molecule in the Pathophysiology of Systemic Lupus Erythematosus. J. Transl. Autoimmun. 2020, 3, 100029. [Google Scholar] [CrossRef]
- Pahar, B.; Madonna, S.; Das, A.; Albanesi, C.; Girolomoni, G. Immunomodulatory Role of the Antimicrobial LL-37 Peptide in Autoimmune Diseases and Viral Infections. Vaccines 2020, 8, 517. [Google Scholar] [CrossRef]
- Yang, B.; Good, D.; Mosaiab, T.; Liu, W.; Ni, G.; Kaur, J.; Liu, X.; Jessop, C.; Yang, L.; Fadhil, R.; Yi, Z.; Wei, M.Q. Significance of LL-37 on Immunomodulation and Disease Outcome. Biomed. Res. Int. 2020, 2020, 349712. [Google Scholar] [CrossRef] [PubMed]
- Verjans, E.T.; Zels, S.; Luyten, W.; Landuyt, B.; Schoofs, L. Molecular Mechanisms of LL-37-Induced Receptor Activation: An Overview. Peptides 2016, 85, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.-M.; Zdebik, A.-E.; Brand, S.; Chromik, A.M.; Strauss, S.; Schmitz, F.; Steinstraesser, L.; Schmidt, W.E. Effects of the cathelicidin LL-37 on intestinal epithelial barrier integrity. Regul. Pept. 2009, 156, 104–117. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhou, Y.; Liu, N.; Zhang, W.; Chen, X.; Qiu, G.; Shen, Y. Cathelicidin LL-37 in periodontitis: Current research advances and future prospects—A review. Int. Immunopharmacol. 2025, 150, 114277. [Google Scholar] [CrossRef]
- Chinipardaz, Z.; Zhong, J.M.; Yang, S. Regulation of LL-37 in Bone and Periodontium Regeneration. Life 2022, 12, 1533. [Google Scholar] [CrossRef]
- Vaishnava, S.; Yamamoto, M.; Severson, K.M.; Ruhn, K.A.; Yu, X.; Koren, O.; Ley, R.; Wakeland, E.K.; Hooper, L.V. The antibacterial lectin RegIIIγ promotes the spatial segregation of microbiota and host in the intestine. Science 2011, 334, 255–258. [Google Scholar] [CrossRef]
- Shin, J.H.; Seeley, R.J. Reg3 Proteins as Gut Hormones? Endocrinology. 2019, 160, 1506–1514. [Google Scholar] [CrossRef]
- Qi, Z.; Pang, W.; Zha, X.; Liu, Y.; Liu, S.; Xiao, F.; Wang, X.; Zhou, J.; Wang, W.; Liu, X. REG/Reg Family Proteins: Mediating Gut Microbiota Homeostasis and Implications in Digestive Diseases. Gut Microbes 2025, 17, 2568055. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, X.; Zhao, Y.; Chen, F.; Sun, M.; Yang, W.; Chen, L.; Yao, S.; Peniche, A.; Dann, S.M.; Sun, J.; Golovko, G.; Fofanov, Y.; Miao, Y.; Liu, Z.; Chen, D.; Cong, Y. Interleukin-33 Promotes REG3γ Expression in Intestinal Epithelial Cells and Regulates Gut Microbiota. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 21–36. [Google Scholar] [CrossRef]
- Shin, J.H.; Bozadjieva-Kramer, N.; Seeley, R.J. Reg3γ: Current Understanding and Future Therapeutic Opportunities in Metabolic Disease. Exp. Mol. Med. 2023, 55, 1672–1677. [Google Scholar] [CrossRef]
- Docampo, M.D.; Auletta, J.J.; Jenq, R.R. Emerging Influence of the Intestinal Microbiota during Allogeneic Hematopoietic Cell Transplantation: Control the Gut and the Body Will Follow. Biol. Blood Marrow Transplant. 2015, 21, 1360–1366. [Google Scholar] [CrossRef]
- Mukherjee, S.; Zheng, H.; Derebe, M.G.; Callenberg, K.M.; Partch, C.L.; Rollins, D.; Propheter, D.C.; Rizo, J.; Grabe, M.; Jiang, Q.X.; Hooper, L.V. Antibacterial Membrane Attack by a Pore-Forming Intestinal C-Type Lectin. Nature 2014, 505, 103–107. [Google Scholar] [CrossRef]
- Yao, A.; Huang, C.; Wang, X.; Zhou, R.; Hao, W.; Lin, Q. The Reg Protein Family: Potential New Targets for the Treatment of Inflammatory Bowel Disease and Colorectal Cancer. Front. Gastroenterol. 2024, 3, 1386069. [Google Scholar] [CrossRef]
- Muniz, L.R.; Knosp, C.; Yeretssian, G. Intestinal antimicrobial peptides during homeostasis, infection, and disease. Front Immunol. 2012, 3, 310. [Google Scholar] [CrossRef] [PubMed]
- Ra, Y.E.; Bang, Y.J. Balancing Act of the Intestinal Antimicrobial Proteins on Gut Microbiota and Health. J. Microbiol. 2024, 62, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial Peptides and Gut Microbiota in Homeostasis and Pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Fu, J.; Xu, B.; Wang, Y.; Jin, M. Interplay between Gut Microbiota and Antimicrobial Peptides. Anim. Nutr. 2020, 6, 389–396. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A.J.; Yao, S.; Yi Xiao, Y.; Huang, X.; D Eaves-Pyles, T.; Golovko, G.; Fofanov, Y.; D’Souza, W.; Zhao, Q.; Liu, Z.; Cong, Y. GPR43 Mediates Microbiota Metabolite SCFA Regulation of Antimicrobial Peptide Expression in Intestinal Epithelial Cells via Activation of mTOR and STAT3. Mucosal Immunol. 2018, 11, 752–762. [Google Scholar] [CrossRef]
- Khlystova, K.A.; Sarkisyan, N.G.; Kataeva, N.N. Natural and Synthetic Peptides in Antimicrobial Therapy. Russ. J. Immunol. 2023, 26, 105–114. [Google Scholar] [CrossRef]
- Abaturov, A.; Kryuchko, T.; Lezhenko, G. The Drugs Based on Molecular Structures of Antimicrobial Peptides and Their Therapeutic Potential in the Treatment of Infectious Diseases of the Respiratory Tract (Part 1). Child’s Health. 2021, 12, 925–938. [Google Scholar] [CrossRef]
- Sharma, D. Novel Natural and Synthetic Anticandidal Therapeutic Peptides to Combat Drug-Resistant Infections. Med. Sci. Forum 2024, 24, 12. [Google Scholar] [CrossRef]
- Loffredo, M.R.; Nencioni, L.; Mangoni, M.L.; Casciaro, B. Antimicrobial Peptides for Novel Antiviral Strategies in the Current Post-COVID-19 Pandemic. J. Pept. Sci. 2023, 29, e3534. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of Antimicrobial/Cytotoxic Activity and Structure of Peptides as a Resource for Development of New Therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef] [PubMed]
- Dilawari, R.; Chaubey, G.K.; Priyadarshi, N.; Barik, M.; Parmar, N. Antimicrobial Peptides: Structure, Function, Mechanism of Action and Therapeutic Applications in Human Diseases. Explor. Drug Sci. 2025, 3, 1008110. [Google Scholar] [CrossRef]
- Alzain, M.; Daghistani, H.; Shamrani, T.; Almoghrabi, Y.; Daghistani, Y.; Alharbi, O.S.; Sait, A.M.; Mufrrih, M.; Alhazmi, W.; Alqarni, M.A.; Saleh, B.H.; Zubair, M.A.; Juma, N.A.; Niyazi, H.A.; Niyazi, H.A.; Halabi, W.S.; Altalhi, R.; Kazmi, I.; Altayb, H.N.; Ibrahem, K.; Alfadil, A. Antimicrobial Peptides: Mechanisms, Applications, and Therapeutic Potential. Infect. Drug Resist. 2025, 18, 4385–4426. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Q.; Ren, K.; Xu, T.; Zhang, Z.; Xu, M.; Rao, Z.; Zhang, X. A Review of Antimicrobial Peptides: Structure, Mechanism of Action, and Molecular Optimization Strategies. Fermentation 2024, 10, 540. [Google Scholar] [CrossRef]
- Guith, T.; Gourishetti, K.; Banerjee, P.; Ghosh, N.; Rana, M.; Chatterjee, G.; Biswas, S.; Bagchi, D.; Roy, S.; Das, A. An overview of antimicrobial peptides. In Viral, Parasitic, Bacterial, and Fungal Infections, 2nd ed.; Bagchi, D., Das, A., Downs, D.W., Eds.; Academic Press: London, UK, 2023; Volume 3, pp. 101–106. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: diversity in structures and pathways. FEMS Microbiology Reviews 2016, 40, 133–159. [Google Scholar] [CrossRef]
- Yang, R.; Ma, X.; Peng, F.; Wen, J.; Allahou, L.W.; Williams, G.R.; Knowles, J.C.; Poma, A. Advances in antimicrobial peptides: From mechanistic insights to chemical modifications. Biotechnology Advances 2025, 81, 108570. [Google Scholar] [CrossRef]
- Marciano, C.L.; Félix de Lima, J.V.; Couto Rosa, M.S.d.; do Nascimento, R.A.; Ferraz, A.d.O.; Silva, I.C.d.; Chrysostomo-Massaro, T.N.; Rosa-Garzon, N.G.d.; Cabral, H. A Comprehensive Overview of Antimicrobial Peptides: Broad-Spectrum Activity, Computational Approaches, and Applications. Antibiotics 2025, 14, 1115. [Google Scholar] [CrossRef]
- Fontanot, A.; Ellinger, I.; Unger, W.W.J.; Hays, J.P. A Comprehensive Review of Recent Research into the Effects of Antimicrobial Peptides on Biofilms—January 2020 to September 2023. Antibiotics 2024, 13, 343. [Google Scholar] [CrossRef]
- Xuan, J.; Feng, W.; Wang, J.; Wang, R.; Zhang, B.; Bo, L.; Chen, Z.-S.; Yang, H.; Sun, L. Antimicrobial Peptides for Combating Drug-Resistant Bacterial Infections. Drug Resistance Updates 2023, 68, 100954. [Google Scholar] [CrossRef] [PubMed]
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The Multifaceted Nature of Antimicrobial Peptides: Current Synthetic Chemistry Approaches and Future Directions. Chemical Society Reviews 2021, 50, 7820–7880. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Kamau, P.M.; Thuku, R.C.; Lai, R. Design methods for antimicrobial peptides with improved performance. Zoological Research 2023, 44, 1095–1114. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Chen, M.-T.; Stedman, F.; Hernandez, J.; Kumble, G.; Kang, X.; Zhang, C.; Tang, G.; Reed, I.; Daugherty, I.Q.; Liu, W.; Klucznik, K.R.; Ocloo, J.L.; Li, A.A.; Klousnitzer, J.; Heinrich, F.; Deslouches, B.; Tristram-Nagle, S. Cyclization of Two Antimicrobial Peptides Improves Their Activity. ACS Omega 2025, 10, 9728–9740. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B.K.H.L. Review: Lessons learned from clinical trials using antimicrobial peptides (AMPs). Frontiers in Microbiology. 2021, 12, 616979. [Google Scholar] [CrossRef]
- Masihi, K.N.; Schäfer, H. Overview of biologic response modifiers in infectious disease. Infectious Disease Clinics of North America 2011, 25, 723–731. [Google Scholar] [CrossRef]
- da Costa, J.P.; Cova, M.; Ferreira, R.; Vitorino, R. Antimicrobial peptides: An alternative for innovative medicines? Applied Microbiology and Biotechnology 2015, 99, 2023–2040. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M. Antimicrobial Peptides: From Design to Clinical Application. Antibiotics 2022, 11, 349. [Google Scholar] [CrossRef]
- Kang, S.-J.; Nam, S.H.; Lee, B.-J. Engineering Approaches for the Development of Antimicrobial Peptide-Based Antibiotics. Antibiotics 2022, 11, 1338. [Google Scholar] [CrossRef]
- Lu, J.; Xu, H.; Xia, J.; Ma, J.; Xu, J.; Li, Y.; Feng, J. D- and Unnatural Amino Acid Substituted Antimicrobial Peptides withImproved Proteolytic Resistance and Their Proteolytic Degradation Characteristics. Front. Microbiol. 2020, 11, 563030. [Google Scholar] [CrossRef]
- Kapil, S.; Sharma, V. d-Amino Acids in Antimicrobial Peptides: A Potential Approach to Treat and Combat Antimicrobial Resistance. Can. J. Microbiol. 2021, 67, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, X.; Liu, Y.; Geng, Z.; Dai, J.; Guo, Y. Rational Design and Synthesis of Potent Active Antimicrobial Peptides Based on American Oyster Defensin Analogue A3. RSC Adv. 2025, 15, 19954–19965. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, Y.; Song, Z.; Tan, Z.; Cheng, J. Recent Advances in Design of Antimicrobial Peptides and Polypeptides toward Clinical Translation. Adv. Drug Deliv. Rev. 2021, 170, 261–280. [Google Scholar] [CrossRef] [PubMed]
- McCarver, S.J.; Qiao, J.X.; Carpenter, J.; Borzilleri, R.M.; Poss, M.A.; Eastgate, M.D.; Miller, M.M.; MacMillan, D.W.C. Decarboxylative Peptide Macrocyclization through Photoredox Catalysis. Angew. Chem. Int. Ed. 2017, 56, 728–732. [Google Scholar] [CrossRef]
- Fang, P.; Pang, W.-K.; Xuan, S.; Chan, W.-L.; Leung, K.C.-F. Recent Advances in Peptide Macrocyclization Strategies. Chem. Soc. Rev. 2024, 53, 11725–11771. [Google Scholar] [CrossRef]
- Vicente-Garcia, C.; Colomer, I. Lipopeptides as Tools in Catalysis, Supramolecular, Materials and Medicinal Chemistry. Nat. Rev. Chem. 2023, 7, 710–731. [Google Scholar] [CrossRef]
- Rounds, T.; Straus, S.K. Lipidation of Antimicrobial Peptides as a Design Strategy for Future Alternatives to Antibiotics. Int. J. Mol. Sci. 2020, 21, 9692. [Google Scholar] [CrossRef]
- Makowska, M.; Kosikowska-Adamus, P.; Zdrowowicz, M.; Wyrzykowski, D.; Prahl, A.; Sikorska, E. Lipidation of Naturally Occurring α-Helical Antimicrobial Peptides as a Promising Strategy for Drug Design. Int. J. Mol. Sci. 2023, 24, 3951. [Google Scholar] [CrossRef]
- Allison, J.R.; Müller, M.; van Gunsteren, W.F. A Comparison of the Different Helices Adopted by α- and β-Peptides Suggests Different Reasons for Their Stability. Protein Sci. 2010, 19, 2186–2195. [Google Scholar] [CrossRef]
- Shankar, S.; Ur Rahim, J.; Rai, R. Self-Assembly in Peptides Containing β- and γ-Amino Acids. Curr. Protein Pept. Sci. 2020, 21, 584–597. [Google Scholar] [CrossRef]
- Lobo-Ruiz, A.; Tulla-Puche, J. Synthetic Approaches of Naturally and Rationally Designed Peptides and Peptidomimetics. Peptide Applications in Biomedicine, Biotechnology and Bioengineering 2018, 23–49. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Comprehensive Review of γ-Glutamyl Peptides (γ-GPs) and Their Effect on Inflammation Concerning Cardiovascular Health. J. Agric. Food Chem. 2022, 70, 7851–7870. [Google Scholar] [CrossRef] [PubMed]
- Piller, P.; Wolinski, H.; Cordfunke, R.A.; Drijfhout, J.W.; Keller, S.; Lohner, K.; Malanovic, N. Membrane Activity of LL-37 Derived Antimicrobial Peptides against Enterococcus hirae: Superiority of SAAP-148 over OP-145. Biomolecules 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Voronko, O.E.; Khotina, V.A.; Kashirskikh, D.A.; Lee, A.A.; Gasanov, V.A.o. Antimicrobial Peptides of the Cathelicidin Family: Focus on LL-37 and Its Modifications. Int. J. Mol. Sci. 2025, 26, 8103. [Google Scholar] [CrossRef] [PubMed]
- Ridyard, K.E.; Overhage, J. The Potential of Human Peptide LL-37 as an Antimicrobial and Anti-Biofilm Agent. Antibiotics 2021, 10, 650. [Google Scholar] [CrossRef]
- Cheng, Y.; Qin, J.; Huang, Y.; Kang, X.; Wei, J.; Peng, J.; Chen, J. The Antimicrobial Effects of PLGA Microspheres Containing the Antimicrobial Peptide OP-145 on Clinically Isolated Pathogens in Bone Infections. Sci. Rep. 2022, 12, 14541. [Google Scholar] [CrossRef]
- Costanzo, M.; De Giglio, M.A.R.; Gilhen-Baker, M.; et al. The chemical basis of seawater therapies: a review. En-viron. Chem. Lett. 2024, 22, 2133–2149. [Google Scholar] [CrossRef]
- Vicidomini, C.; Roviello, G.N. Therapeutic Convergence in Neurodegeneration: Natural Products, Drug Repur-posing, and Biomolecular Targets. Biomolecules 2025, 15, 1333. [Google Scholar] [CrossRef]
- Costanzo, M.; Roviello, G.N. Precision Therapeutics Through Bioactive Compounds: Metabolic Reprogramming, Omics Integration, and Drug Repurposing Strategies. Int. J. Mol. Sci. 2025, 26, 10047. [Google Scholar] [CrossRef]
- Sargsyan, T.; Simonyan, H.M.; Stepanyan, L.; Tsaturyan, A.; Vicidomini, C.; Pastore, R.; Guerra, G.; Roviello, G.N. Neuroprotective Properties of Clove (Syzygium aromaticum): State of the Art and Future Pharmaceutical Applica-tions for Alzheimer’s Disease. Biomolecules 2025, 15, 452. [Google Scholar] [CrossRef]
- Roviello, G.N. The Multifunctional Role of Salix spp.: Linking Phytoremediation, Forest Therapy, and Phyto-medicine for Environmental and Human Benefits. Forests 2025, 16, 1808. [Google Scholar] [CrossRef]
- Roviello, G.N. Nature-Inspired Pathogen and Cancer Protein Covalent Inhibitors: From Plants and Other Natu-ral Sources to Drug Development. Pathogens 2025, 14, 1153. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, B.T.; Thompson, E.P.; Roviello, G.N.; Gale, T.F. C-Terminal Analogues of Camostat Retain TMPRSS2 Protease Inhibition: New Synthetic Directions for Antiviral Repurposing of Guanidinium-Based Drugs in Res-piratory Infections. Int. J. Mol. Sci. 2025, 26, 6761. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, P.L.; Tesauro, D.; Roviello, G.N. Metallogels as Supramolecular Platforms for Biomedical Appli-cations: A Review. Processes 2025, 13, 3671. [Google Scholar] [CrossRef]
- Fik-Jaskółka, M.A.; Mkrtchyan, A.F.; Saghyan, A.S.; Palumbo, R.; Belter, A.; Hayriyan, L.A.; Simonyan, H.; Rovi-ello, V.; Roviello, G.N. Spectroscopic and SEM evidences for G4-DNA binding by a synthetic alkyne-containing amino acid with anticancer activity. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 229, 117884. [Google Scholar] [CrossRef]
- Roviello, V.; Musumeci, D.; Mokhir, A.; Roviello, G.N. Evidence of protein binding by a nucleopeptide based on a thymine-decorated L-diaminopropanoic acid through CD and in silico studies. Curr. Med. Chem. 2021, 28, 5004–5015. [Google Scholar] [CrossRef]
- Marzano, M.; Falanga, A.P.; Marasco, D.; Borbone, N.; D’Errico, S.; Piccialli, G.; Roviello, G.N.; Oliviero, G. Eval-uation of an Analogue of the Marine ε-PLL Peptide as a Ligand of G-quadruplex DNA Structures. Mar. Drugs 2020, 18, 49. [Google Scholar] [CrossRef]
- Roviello, G.N.; Ricci, A.; Buccia, E.M.; Pedone, C. Synthesis, biological evaluation and supramolecular assembly of novel analogues of peptidyl nucleosides. Mol. BioSyst. 2011, 7, 1331–1339. [Google Scholar] [CrossRef]
- Roviello, G.N.; Musumeci, D.; Buccia, E.M.; Pedone, C. Evidences for supramolecular organization of nucleo-peptides: synthesis, spectroscopic and biological studies of a novel dithymine L-serine tetrapeptide. Mol. BioSyst. 2011, 7, 1073–1081. [Google Scholar] [CrossRef]
- Roviello, G.; Musumeci, D.; Pedone, C.; Bucci, E.M. Synthesis, characterization and hybridization studies of an alternate nucleo-ε/γ-peptide: complexes formation with natural nucleic acids. Amino Acids 2010, 38, 103–111. [Google Scholar] [CrossRef]
- Musumeci, D.; Oliviero, G.; Roviello, G.N.; Bucci, E.M.; Piccialli, G. G-quadruplex-forming oligonucleotide con-jugated to magnetic nanoparticles: synthesis, characterization, and enzymatic stability assays. Bioconjug. Chem. 2012, 23, 382–391. [Google Scholar] [CrossRef]
- Roviello, G.N.; Di Gaetano, S.; Capasso, D.; Cesarani, A.; Bucci, E.M.; Pedone, C. Synthesis, spectroscopic studies and biological activity of a novel nucleopeptide with Moloney murine leukemia virus reverse transcriptase in-hibitory activity. Amino Acids 2010, 38, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Sargsyan, T.; Hakobyan, H.; Mardiyan, Z.; Khachaturyan, N.; Gevorgyan, S.; Jamgaryan, S.; Gyulumyan, E.; Danghyan, Y.; Saghyan, A. Synthesis of (S)-alanyl-(S)-β-(thiazol-2-yl-carbamoyl)-α-alanine, a dipeptide con-taining it, and in vitro investigation of antifungal activity. Farmacia 2022, 70, 1148–1154. [Google Scholar] [CrossRef]
- Dadayan, A.; Sargsyan, A.; Hakobyan, H.; Mardiyan, Z.; Jamharyan, S.; Hovhannisyan, N.; Sargsyan, T. Modeling, synthesis and in vitro screening of unusual amino acids and peptides as protease inhibitors. J Chem Technol Metall. 2023, 615–620. [Google Scholar]
- Sargsyan, A.S.; Sargsyan, T.H.; Hovhannisyan, N.A. Modeling, synthesis and in vitro testing of peptides based on unusual amino acids as potential antibacterial agents. Biomed. Khim 2024, 70, 413–420. [Google Scholar] [CrossRef]
- Sargsyan, T.; Saghyan, A. The synthesis and in vitro study of 9-fluorenylmethoxycarbonyl protected non-protein amino acids antimicrobial activity. Eurasian Chem.-Technol. J. 2024, 25, 235–240. [Google Scholar] [CrossRef]
- Dadayan, A.S.; Mkrtchyan, A.F.; Poghosyan, A.S.; Dadayan, S.A.; Stepanyan, L.A.; Israyelyan, M.H.; Tovmasyan, A.S.; Tsaturyan, A.H.; Hovhannisyan, N.A.; Topuzyan, V.O.; Sargsyan, T.H.; Saghyan, A.S. Unnatural phosphorus-containing α-amino acids and their N-FMOC derivatives: Synthesis and in vitro investigation of anticholinesterase activity. ChemSelect 2024, 9, 3. [Google Scholar] [CrossRef]
- Mkrtchyan, A.F.; Hayriyan, L.A.; Karapetyan, A.J.; Tovmasyan, A.S.; Tsaturyan, A.H.; Khrustalev, V.N.; Maleev, V.I.; Saghyan, A.S. Using the Ni-[(benzylprolyl)amino]benzophenone complex in the Glaser reaction for the synthesis of bis α-amino acids. New J. Chem. 2020, 44. [Google Scholar] [CrossRef]
- Tovmasyan, A.S.; Mkrtchyan, A.F.; Khachatryan, H.N.; Hayrapetyan, M.V.; Hakobyan, R.M.; Poghosyan, A.S.; Tsaturyan, A.H.; Minasyan, E.V.; Maleev, V.I.; Larionov, V.A.; et al. Synthesis, Characterization, and Study of Catalytic Activity of Chiral Cu(II) and Ni(II) Salen Complexes in the α-Amino Acid C-α Alkylation Reaction. Molecules 2023, 28, 1180. [Google Scholar] [CrossRef]
- Stepanyan, L.; Sargsyan, T.; Mittova, V.; Tsetskhladze, Z.R.; Motsonelidze, N.; Gorgoshidze, E.; Nova, N.; Is-rayelyan, M.; Simonyan, H.; Bisceglie, F.; et al. The synthesis, characterization, and biological evaluation of a fluorenyl-methoxycarbonyl-containing thioxo-triazole-bearing dipeptide: Antioxidant, antimicrobial, and BSA/DNA binding studies. Biomolecules 2025, 15, 933. [Google Scholar] [CrossRef]
- Sargsyan, T.; Hakobyan, H.; Simonyan, H.; Soghomonyan, T.; Tsaturyan, A.; Hovhannisyan, A.; Sardaryan, S.; Saghyan, A.; Roviello, G.N. Biomacromolecular interactions and antioxidant properties of novel synthetic ami-no acids targeting DNA and serum albumin. J. Mol. Liq. 2025, 128700. [Google Scholar] [CrossRef]
- Saubenova, M.; Rapoport, A.; Yermekbay, Z.; Oleinikova, Y. Antimicrobial Peptides, Their Production, and Po-tential in the Fight Against Antibiotic-Resistant Pathogens. Fermentation 2025, 11, 36. [Google Scholar] [CrossRef]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; Falabella, P. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632. [Google Scholar] [CrossRef] [PubMed]
- Gagat, P.; Ostrówka, M.; Duda-Madej, A.; Mackiewicz, P. Enhancing Antimicrobial Peptide Activity through Modifications of Charge, Hydrophobicity, and Structure. Int. J. Mol. Sci. 2024, 25, 10821. [Google Scholar] [CrossRef]
- Min, K.H.; Kim, K.H.; Ki, M.-R.; Pack, S.P. Antimicrobial Peptides and Their Biomedical Applications: A Review. Antibiotics 2024, 13, 794. [Google Scholar] [CrossRef]
- Straus, S.K. Tryptophan- and Arginine-Rich Antimicrobial Peptides: Anti-Infectives with Great Potential. Bio-chim. Biophys. Acta Biomembr. 2024, 1866, 184260. [Google Scholar] [CrossRef]
- Zheng, S.; Tu, Y.; Li, B.; et al. Antimicrobial Peptide Biological Activity, Delivery Systems and Clinical Transla-tion: Status and Challenges. J. Transl. Med. 2025, 23, 292. [Google Scholar] [CrossRef]
- Li Petri, G.; Di Martino, S.; De Rosa, M. Peptidomimetics: An Overview of Recent Medicinal Chemistry Efforts toward the Discovery of Novel Small Molecule Inhibitors. J. Med. Chem. 2022, 65, 7438–7475. [Google Scholar] [CrossRef]
- Rezende, B.S.; Oshiro, K.G.N.; Júnior, N.G.O.; Franco, O.L.; Cardoso, M.H. Advances on chemically modified antimicrobial peptides for generating peptide antibiotics. Chemical Communications 2021, 88, 11578–11590. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Zhang, Q.; Nguyen, T.P.T.; Nguyen, T.T.H.; Medhi, H.; Rodrigues, H.L.S.; Canales Carnero, C.S.; Sutherland, D.; Helmy, N.M.; Araveti, P.B.; de la Torre, B.G.; Albericio, F.; Pavan, F.R. Synergistic Combina-tions of Antimicrobial Peptides and Conventional Antibiotics: A Strategy to Delay Resistance Emergence in WHO Priority Bacteria. Pharmacol. Rev. 2025, 78, 100104. [Google Scholar] [CrossRef]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A review on antimicrobial peptides da-tabases and the computational tools. Database 2022, 2022, baac011. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nu-cleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Mondal, R.K.; Sen, D.; Arya, A.; Samanta, S.K. Developing anti-microbial peptide database version 1 to provide comprehensive and exhaustive resource of manually curated AMPs. Sci. Rep. 2023, 13, 17843. [Google Scholar] [CrossRef] [PubMed]
- Feurstein, C.; Meyer, V.; Jung, S. Structure–Activity Predictions from Computational Mining of Protein Databases to Assist Modular Design of Antimicrobial Peptides. Front. Microbiol. 2022, 13, 812903. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing Antimicrobial Peptides: Form Follows Function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Thera-peutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Mishra, B.; Basu, A.; Shehadeh, F.; Felix, L.; Kollala, S.S.; Chhonker, Y.S.; Naik, M.T.; Dellis, C.; Zhang, L.; Gane-san, N.; Murry, D.J.; Gu, J.; Sherman, M.B.; Ausubel, F.M.; Sotiriadis, P.P.; Mylonakis, E. Antimicrobial Peptide Developed with Machine Learning Sequence Optimization Targets Drug-Resistant Staphylococcus aureus in Mice. J. Clin. Invest. 2025, 135, e185430. [Google Scholar] [CrossRef]
- Wiradharma, N.; Khoe, U.; Hauser, C.A.; Seow, S.V.; Zhang, S.; Yang, Y.Y. Synthetic Cationic Amphiphilic α-Helical Peptides as Antimicrobial Agents. Biomaterials 2011, 32, 2204–2212. [Google Scholar] [CrossRef]
- Guha, R. On exploring structure–activity relationships. Methods Mol. Biol. 2013, 993, 81–94. [Google Scholar] [CrossRef]
- Zelezetsky, I.; Tossi, A. Alpha-helical antimicrobial peptides—Using a sequence template to guide structure–activity relationship studies. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1436–1449. [Google Scholar] [CrossRef]
- Maher, S.; McClean, S. Investigation of the cytotoxicity of eukaryotic and prokaryotic antimicrobial peptides in intestinal epithelial cells in vitro. Biochem. Pharmacol. 2006, 71, 1289–1298. [Google Scholar] [CrossRef]
Figure 1.
Intestinal defensins: sources, molecular mechanisms, regulation, and pathological consequences. The benefits of AMP activity are effective bacterial killing through membrane disruption, trapping of microbes via nanonets, modulation of immune signals, and maintenance of epithelial integrity and microbiota balance. The risks associated with AMP deficiency are increased epithelial permeability, dysbiosis, heightened inflammation driven by PAMPs and cytokines, and Paneth cell dysfunction linked to conditions such as Crohn’s disease.
Figure 1.
Intestinal defensins: sources, molecular mechanisms, regulation, and pathological consequences. The benefits of AMP activity are effective bacterial killing through membrane disruption, trapping of microbes via nanonets, modulation of immune signals, and maintenance of epithelial integrity and microbiota balance. The risks associated with AMP deficiency are increased epithelial permeability, dysbiosis, heightened inflammation driven by PAMPs and cytokines, and Paneth cell dysfunction linked to conditions such as Crohn’s disease.
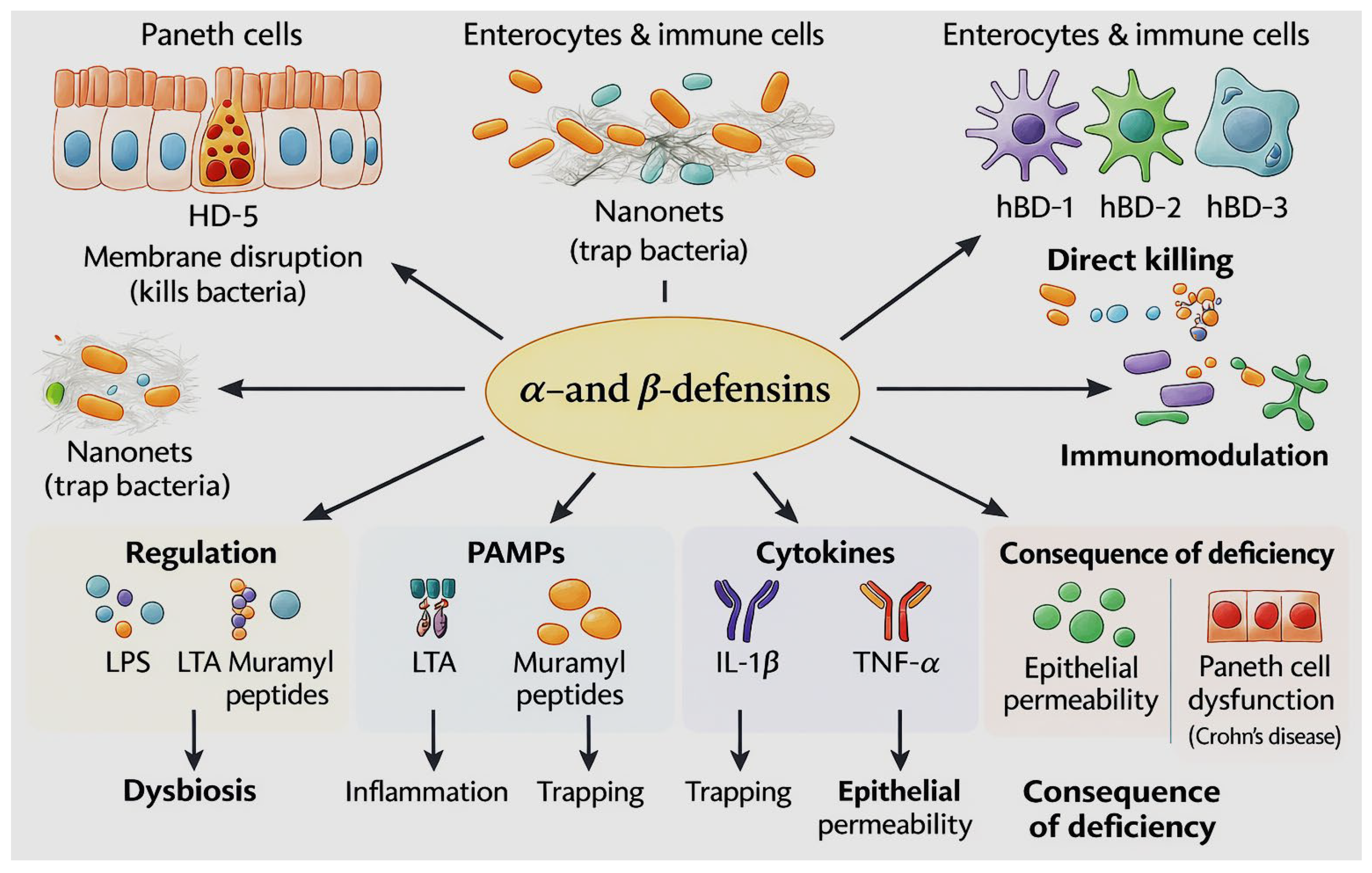
Figure 2.
Illustrative overview of the antimicrobial, immunomodulatory, and barrier-protective functions of LL-37.
Figure 2.
Illustrative overview of the antimicrobial, immunomodulatory, and barrier-protective functions of LL-37.
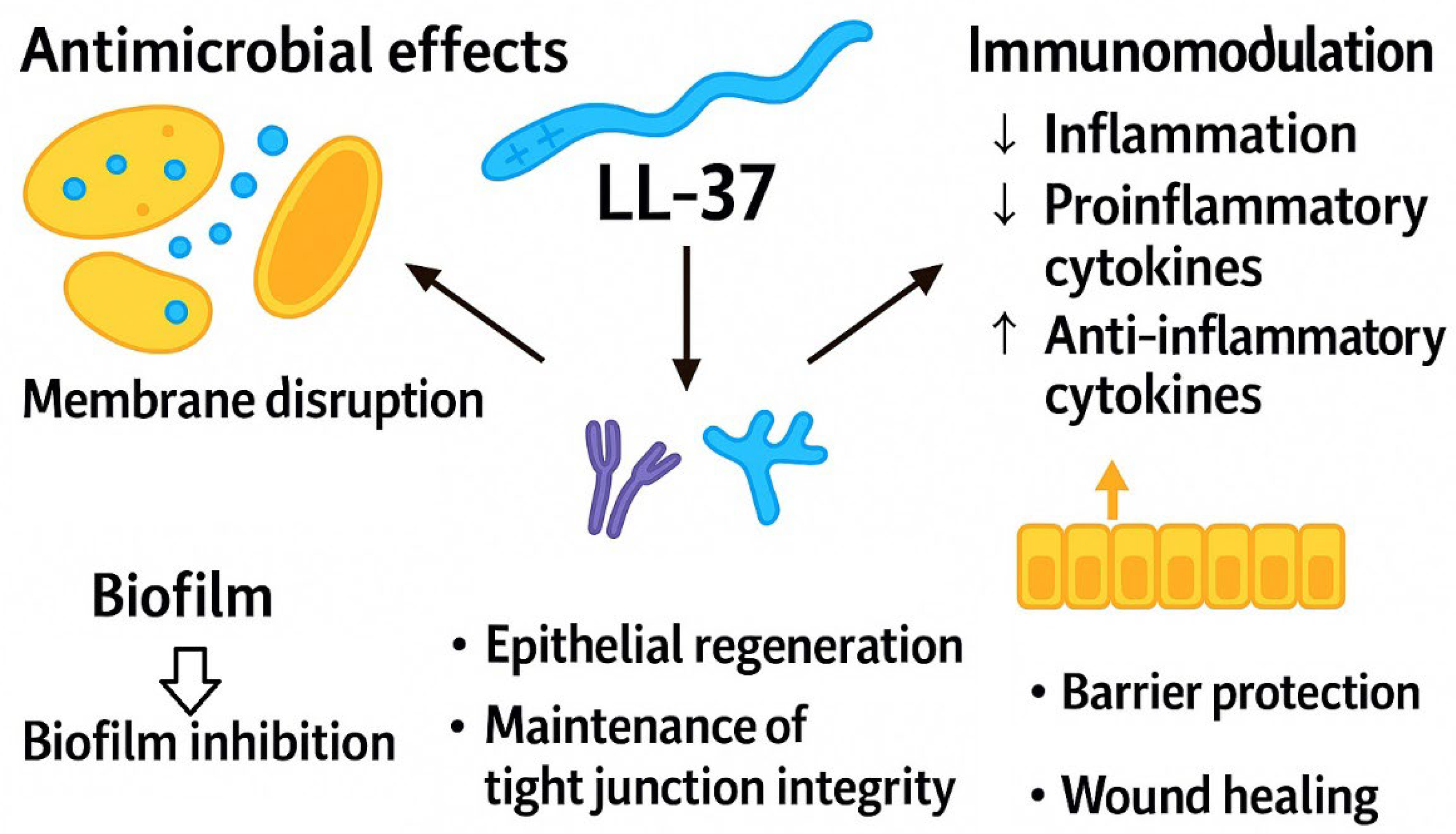
Figure 3.
(a) Three-dimensional views of RegIIIα, one of the members of the RegIII family, shown from different orientations. The structure was visualized and rotated using the 3D viewer available at the Protein Data Bank (https://www.rcsb.org/3d-view/4MTH/1 accessed on 11st December 2025). (b) Scheme illustrating the key functions and regulatory mechanisms of RegIIIγ.
Figure 3.
(a) Three-dimensional views of RegIIIα, one of the members of the RegIII family, shown from different orientations. The structure was visualized and rotated using the 3D viewer available at the Protein Data Bank (https://www.rcsb.org/3d-view/4MTH/1 accessed on 11st December 2025). (b) Scheme illustrating the key functions and regulatory mechanisms of RegIIIγ.
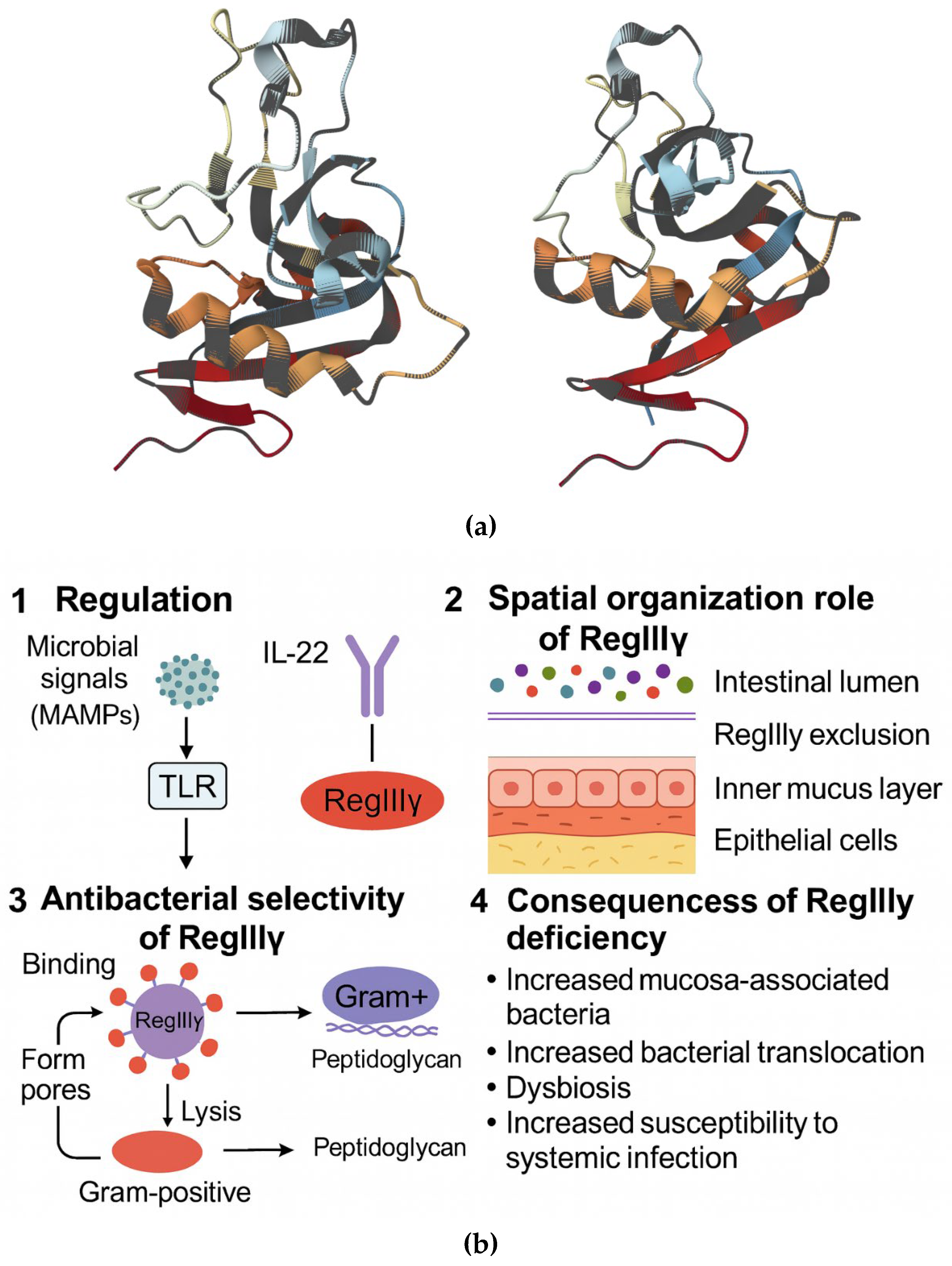
Figure 4.
Schematic overview of the main mechanisms of membrane disruption adopted by antimicrobial peptides.
Figure 4.
Schematic overview of the main mechanisms of membrane disruption adopted by antimicrobial peptides.
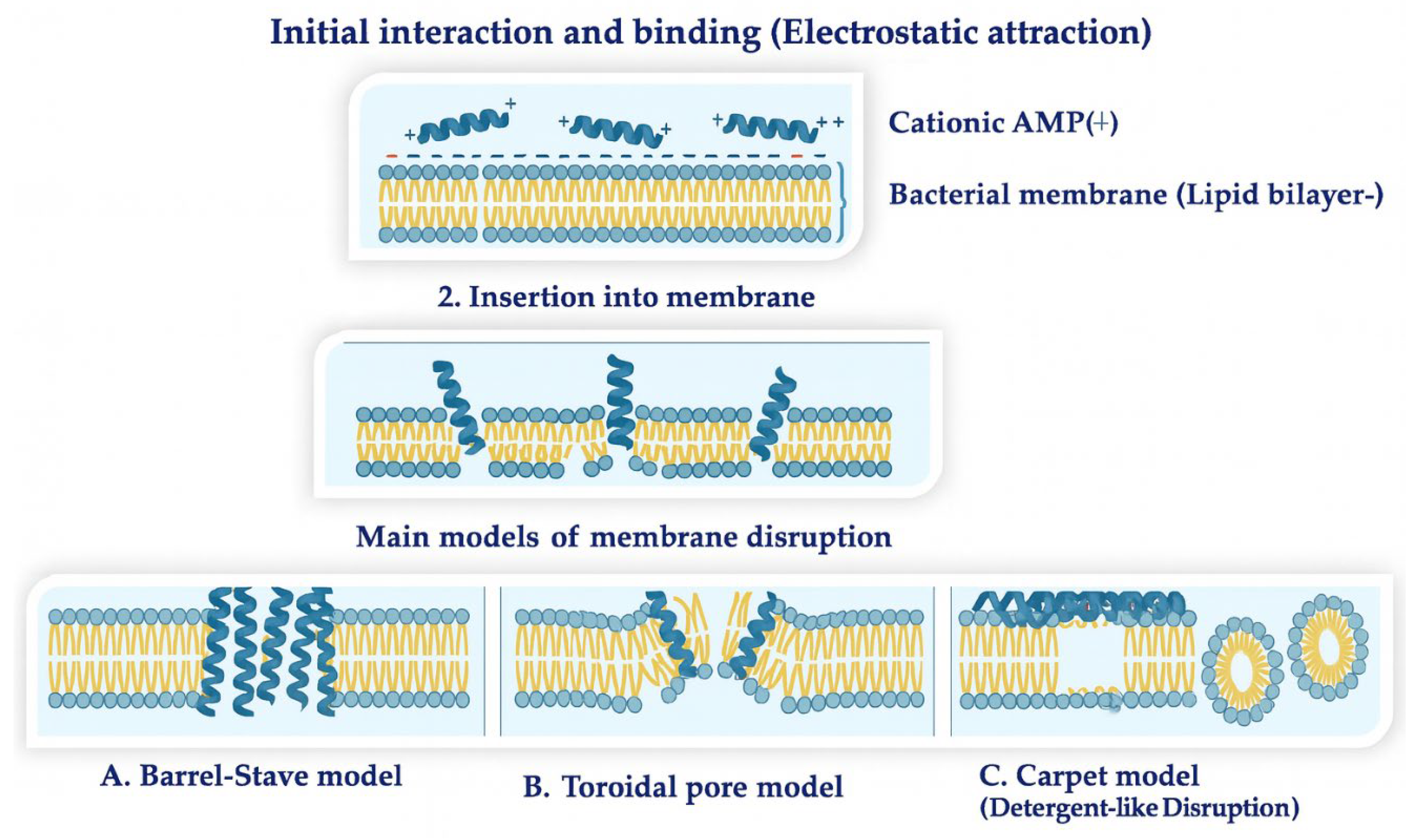
Figure 5.
Structural representation of the defensin.mimetic brilacidin (a), the synthetic peptides XOMA-629 (b) and OP-145 (c).
Figure 5.
Structural representation of the defensin.mimetic brilacidin (a), the synthetic peptides XOMA-629 (b) and OP-145 (c).
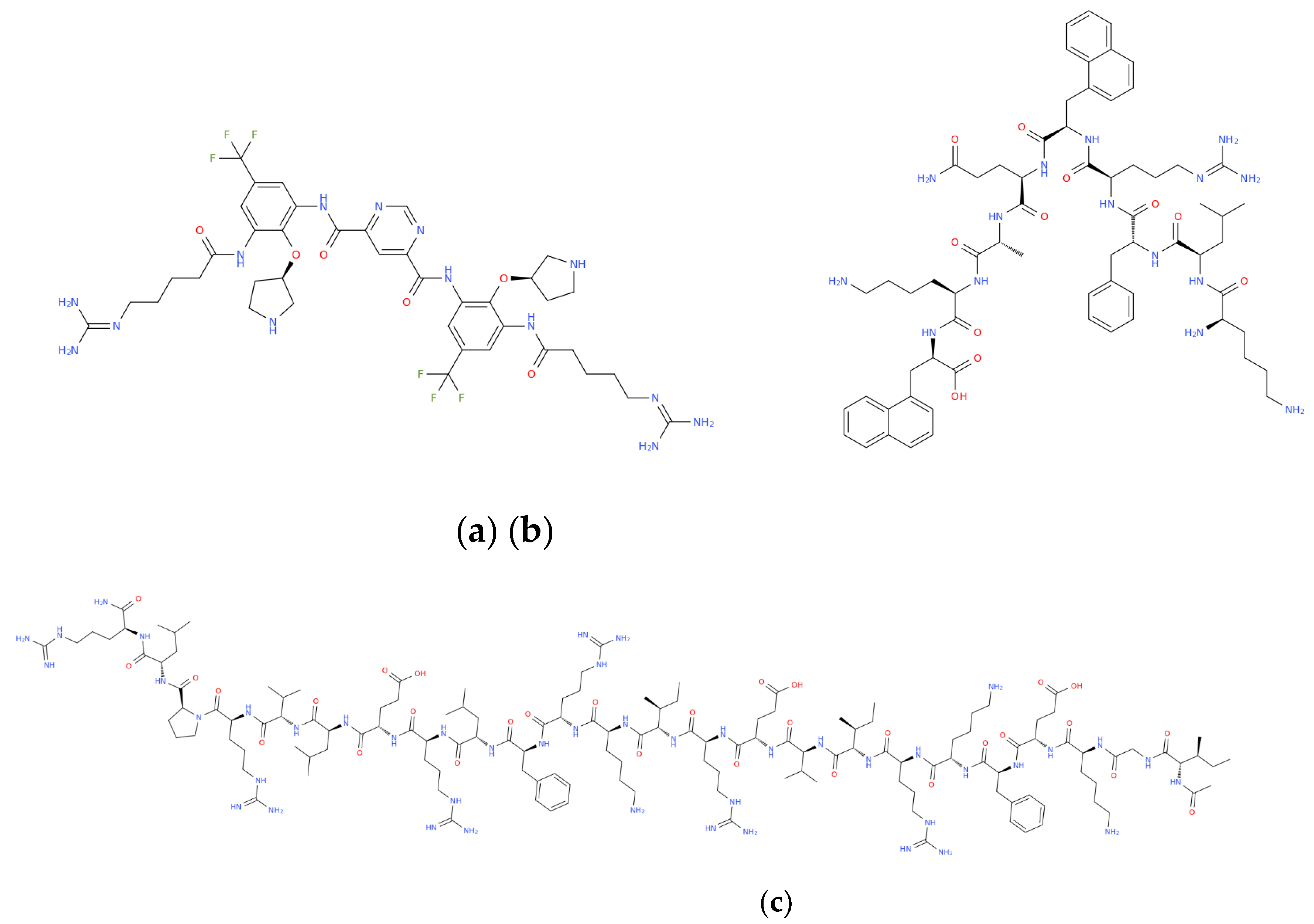

Table 1.
Antimicrobial peptides as drug candidates: clinical and preclinical overview*.
| Name | Sequence/structural details | Activity / Mechanism | Phase | Peptide Type | Developer | Ref. |
| CZEN-002 (CKPV)2 | (CKPV)2 | Antifungal, immunomodulatory (α-MSH analog) | IIb | Synthetic octapeptide | Zengen | [34,35] |
| HB1345 | Decanoyl-KFKWPW | Lipopeptide; membrane disruption | Pre-clinical | Synthetic (lipidated AMP) | Helix BioMedix | [36,37][ |
| hLF1-11 | GRRRRSVQWCA | Lactoferrin fragment; immune-activating | I/II | Synthetic | AM-Pharma | [38,39] |
| IMX-942 (IDR-1) | KSRIVPAIPVSLL | Innate defense regulator (IDR) | I/II | Synthetic cationic peptide | Inimex | [40,41] |
| OP-145 | LL-37 derivative | Anti-biofilm, antibacterial | II | Synthetic | OctoPlus | [40,42] |
| PAC-113 (P-113) | AKRHHGYKRKFH | Anti-Candida (histatin-5 fragment) | IIb | Semi-synthetic | Pacgen | [43,44] |
| Brilacidin | Not disclosed | Defensin-mimetic; membrane disruption | II | Peptidomimetic | Cellceutix / Innovation Pharma | [40,45,46] |
| XOMA-629 | KLFR-(d-naphtho-Ala)-QAK-(d-naphtho-Ala) | Antimicrobial activity via an immunomodulatory mechanism | IIa | Peptidomimetic | XOMA | [40,47,48] |
| Arenicin-3 | β-sheet AMP | Anti-MDR; membrane-active | Pre-clinical | Natural AMP | Adenium | [49,50] |
| RDP58 | D-amino acid decapeptide | TNF-α suppression, anti-inflammatory (immunomodulator) | II | All-D synthetic peptide | Genzyme | [51,52] |
| Iseganan (IB-367) | RGGLCYCRGRFCVCVGR-NH2 | Protegrin analog; antibacterial | III | Semi-synthetic | Ardea Biosciences | [53,54] |
| LTX-109 | Not disclosed | Membrane-active peptidomimetic | II | Peptidomimetic | Lytix Biopharma | [55,56,57] |
| Omiganan | ILRWPWWPWRRK | Indolicidin analog; anti-biofilm | II/III | Semi-synthetic | Cutanea Life Sciences | [58,59,60] |
| Pexiganan (MSI-78) | MSI-78 | Pore-forming antimicrobial | III | Semi-synthetic (magainin analog) | MacroChem | [61,62] |
| Plectasin-212 (NZ2114 class) | Defensin variant | Lipid II binding | Pre-clinical | Engineered defensin | Novozymes | [63,64,65] |
| Murepavadin (POL7080) | β-peptide | LptD inhibitor (P. aeruginosa) | Advanced clinical development (systemic use limited by toxicity) | Peptidomimetic | Polyphor | [66,67,68] |
| LC-AMP-I1 | GRMQEFIKKLK | Ultra-short AMP; MDR activity | Pre-clinical | Natural-derived (spider venom) | Chongqing University | [69,70,71] |
| Lariocidin | Lasso peptide | Pore-forming antimicrobial | Pre-clinical | Natural RiPP | Spanish research groups | [72,73] |
| WR12 | RWKIFKKIEKMGRNIR | Anti-MRSA / VRSA | Pre-clinical | Synthetic de novo peptide | Hong Kong University | [74,75,76] |
| D-IK8 | All-D peptide | Protease-resistant AMP; MDR Gram- | Pre-clinical | All-D synthetic | Korean research groups | [74,77] |
| ZP2 | Synthetic AMP | Quorum-sensing inhibition | Pre-clinical | Synthetic | Chinese research institutes | [78,79,80] |
| Glutoxim | γ-glutamyl-cysteine | Immunomodulator | Marketed | Synthetic (glutathione analog) | Pharmasyntez | [81,82] |
| Acipensin-1 | Fish-derived AMP | Membrane lysis | Pre-clinical | Natural AMP | Far East Biomedical Labs | [83,84] |
* Clinical development stages reported in this table refer to the most advanced phase reached; in some cases, systemic development was limited or discontinued due to safety concerns, while alternative formulations remain under investigation.
Table 2.
Major antimicrobial peptide databases: comprehensive repositories of natural and synthetic AMPs.
Table 2.
Major antimicrobial peptide databases: comprehensive repositories of natural and synthetic AMPs.
| Database | Dataset size* | Content | Website** |
| APD | 2 619 | Natural and some synthetic AMPs | https://aps.unmc.edu/AP/ |
| CAMPR4 | Natural AMPs: 11827 Synthetic AMPs: 12416 |
Collection of Anti-Microbial Peptides | https://camp.bicnirrh.res.in/ |
| DRAMP | 30260 | Data repository of antimicrobial peptides | http://dramp.cpu-bioinfor.org/ |
| BACTIBASE | 177 | Bacteriocins | http://bactibase.pfba-lab.org |
| MilkAMP | 371 | Milk bioactive peptide database | http://mbpdb.nws.oregonstate.edu/ |
| PhytAMP | 273 | Plant AMPs | https://phytamp.hammamilab.org/entrieslist.php |
| AVPdb | 2 683 | Database of antiviral peptides | http://crdd.osdd.net/servers/avpdb/ |
| HIPdb | 981 | Database of experimentally validated HIV inhibiting peptides. | http://crdd.osdd.net/servers/hipdb/index.php |
| DBAASP | ~8 000 | Database of antimicrobial activity and structure of peptides | https://dbaasp.org/home |
| Hemolytik | ~5 000 | Database of hemolytic and non-hemolytic peptides | http://crdd.osdd.net/raghava/hemolytik/ |
* reported dataset sizes are approximate and reflect the number of entries available at the time of access (December 2025); database content is subject to continuous updates and may vary depending on inclusion criteria. ** all links displayed in this table were accessed on 18th December 2025.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



