1. Introduction
The immune system can protect one from pathogens or from cancer but can also drive anti-host immune responses to cause disease. Therefore, modulating the activity of immune cells is key to generating robust vaccine responses and to blunt pathogenic immune responses stimulated by an overactive immune system. One approach to modulate the immune system is to transiently or permanently genetically modify immune cells to activate or inhibit their responses with RNA or DNA therapeutics.
Many lessons were learned with RNA and DNA therapeutics and the immune system during the COVID-19 pandemic [
1]. Many different vaccines were deployed and were administered by diverse vaccination routes in efforts to control and contain SARS-CoV-2. mRNA-lipid nanoparticle (LNP) vaccines expressing the SARS-CoV-2 spike protein were advanced by Moderna and Pfizer and these became the numerically favored vaccine during the pandemic [
1]. While these were rolled out quickly and show great merits, other vaccines like DNA-based adenovirus (Ad) vaccines were also deployed globally. Ad vaccines were initially deployed more broadly in developed and less developed countries owing to their lower cost, but they became disfavored due to early observations of vaccine-induced thrombotic thrombocytopenia (VITT) [
2,
3].
While VITT is certainly a concerning side effect, mRNA-LNP vaccines also had their own side effect: cardiomyopathy. Pfizer or Moderna mRNA vaccines were reported early to cause cardiomyopathy at a rate of 146 cases per 100 000 by the CDC, but later passive reporting in the United States by the Vaccine Adverse Event Reporting System (VAERS) suggested that cardiomyopathy in young males occurred at 105.9 cases per million doses. Authors of the review [
4], calculated the incidence at 77 cases per million vaccinations. For comparison, the incidence of VITT is now estimated to be 3 to 15 cases per million vaccinations [
5]. Therefore, Ad vaccines do not in fact have a worse safety record than more popular mRNA vaccines.
Potency is also a significant consideration in selecting vaccines. Of the most utilized COVID-19 vaccines, only the Johnson and Johnson Ad26 vaccine was given as a single immunization. All others required at least two immunizations as part of their clinical regimen. This need for one versus two vaccinations in the simplest comparison suggests that this DNA-based adenoviral vaccine might have more intrinsic potency than other vaccines that need two or more immunizations.
There seems to be some merit to this conclusion. For example, in the BOOST study, volunteers received the BNT162b2 or mRNA-1273 mRNA vaccines two times or they received the Ad26.COV2.S vaccine only once [
6]. Neutralizing antibody (NAb) titers at 1- and 6-months showed that nABs declined in recipients of BNT162b2 and mRNA-1273, while nABs in recipients of Ad26.COV2.S showed a significant increase. At 6-months, NAbs were significantly higher in the single vaccination Ad26.COV2.S cohort than in the BNT162b2 or mRNA-1273 groups [
6].
While the Ad26 vaccine was given once, the similar ChAdOx1 was given two times like the two mRNA vaccines. When ChAdOx1 was compared to the mRNA vaccines in the ZOE COVID Study, SARS-CoV-2 NAbs waned over 5 months in the mRNA groups, but climbed in the Ad group over the same time period [
7].
These data demonstrate that mRNA and DNA-based Ad vaccines have both their own strengths and their own particular side effects, so both should be considered in future vaccine strategies.
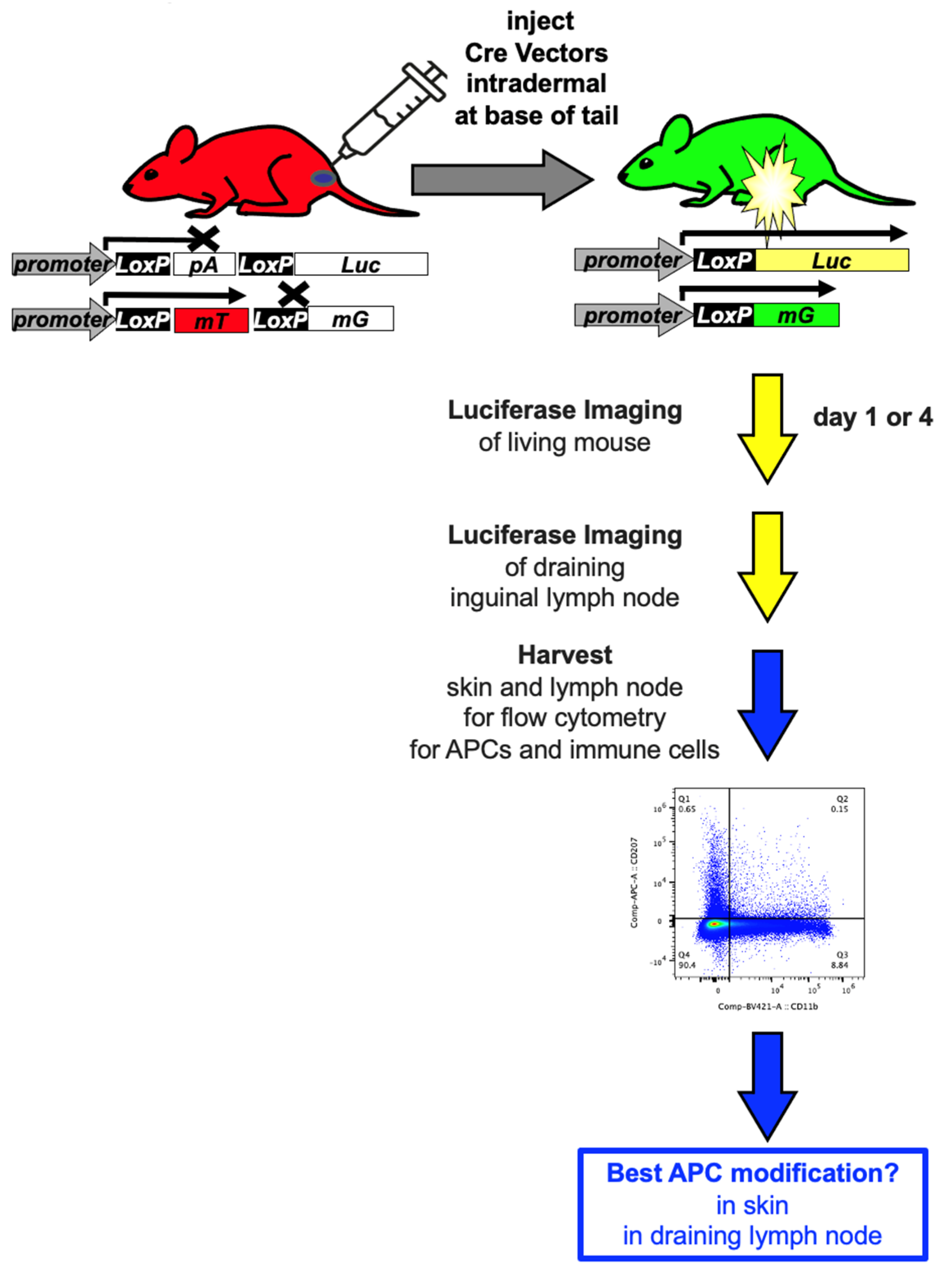
Given this, the current study aimed to compare and benchmark different vaccine platforms for their ability to deliver transgenes into antigen presenting cells in vivo. To quantify the cell specificity of vector delivery to antigen presenting cells (APCs), we utilized a reporter system we developed that “fingerprints” gene delivery in vivo by monitoring luciferase (Luc), green fluorescent protein (GFP), and red fluorescent protein (RFP) expression in the same mouse (
Figure 1 and (2)). Here, we focused on APC modification, since these are key to the initiation of immune responses. Recognition and presentation of foreign antigens is performed by a variety of antigen-presenting cells (APCs) including macrophages, B cells and a variety of phenotypically diverse dendritic cells (DCs) [
8,
9,
10,
11,
12]. Immature DCs can mediate antigen uptake in peripheral organs or in lymph nodes. In the skin are epidermal resident DCs called Langerhans cells (LCs) [
10]. LCs surveil environmental antigens and pathogens that pierce the epidermis of the skin. Once LCs capture and process antigens, these DCs migrate to draining lymph nodes where they mature into DCs that can present antigens to T or B cells [
12]. Antigens can also themselves be transported to draining lymph nodes via lymphatics where they can be sampled by APCs and processed for presentation to T and B cells.
While intradermal vaccines are intentionally injected into the skin to interact with LCs and other APCs, it should be noted that any of the many intramuscular (IM) vaccines also coincidentally deliver antigens into the skin since the needles for IM injection must pierce the skin to reach the muscle. Therefore, LCs and the skin may actually play a role in not only ID, but also in IM vaccine strategies.
To track APC modification efficiently in this study, we compared these vectors by the intradermal (ID) route. This route allowed the surface site of injection to be readily observed. It also allowed the known inguinal draining lymph node for the injection site to also be easily imaged and its cells to be examined.
2. Materials and Methods
2.1. Vectors
Cre recombinase cDNA was expressed from pTRS-CBh-Cre plasmid DNA [
13], or an E1-deleted adenovirus serotype 5 vector [
14], or as an mRNA produced by in vitro transcription (IVT) from a T7 Cre recombinase plasmid. Plasmids were purified on Endotoxin-free Maxiprep kits (Machery-Nagel). Adenovirus was produced from 10 Plate CellStacks (Corning) by banding on two CsCl gradients as in [
15].
2.2. In Vitro Transcription (IVT) of mRNA
The T7-Cre recombinase plasmid was linearized and purified by phenol/chloroform/isoamyl alcohol extraction followed by ethanol precipitation as in [
16,
17]. This was used as a template for IVT using a T7 RNA synthesis kit (New England Biolabs HiScribe E2040S). The mRNA was synthesized with N1-Methylpseudouridine-5’-Triphosphate (m1ΨTP, Trilink N-1081) and co-transcriptionally capped with Trilink CleanCap reagent AG N-7113. DNA template was removed with DNase I and mRNA was purified with NEB Monarch RNA Cleanup Kit T2050. mRNA was assessed by agarose gel electrophoresis and nanodrop.
2.3. Lipid Nanoparticle Assembly
Lipid nanoparticles were formulated as in [
16,
17] using cholesterol (Sigma C3045), 1,2-dimyristoyl-rac-glycero-3-methoxypolyethylene glycol-2000 (DMG-PEG2000, Avanti polar lipids 880151P), 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC, Avanti polar lipids 850365P), and dilinoleylmethyl-4-dimethylaminobutyrate (D-Lin-MC3-DMA or “MC3”, MedChemExpress HY-112251) on a NanoAssemblr Ignite (Cytiva) microfluidics system. Concentrated LNPs were filtered with a 0.2 µM syringe filter (Pall Acrodisc 4602) and size, polydispersity index, and particle concentration were measured using a Zetasizer Advance Ultra Red (Malvern Panalytical ZSU3305). Encapsulation efficiency and total encapsulated mRNA was quantified using a Quant-iT RiboGreen RNA assay kit (Invitrogen R11490).
2.4. In Vivo Experiments
All animal experiments were performed at Mayo Clinic and were approved by the Mayo Clinic Animal Care and Use Committee. All experiments followed guidance of the Public Health Service Animal Welfare Policy, Animal Welfare Act and NIH guidelines in the Guide for the Care and Use of Laboratory Animals.
2.5. mT/mG: LSL-Lux Cre Reporter Mice
mT/mG mice (aka. B6.129(Cg)-Gt(ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo/J strain # 007676) and LSL-Luc mice (aka FVB.129S6(B6)-Gt(ROSA)26Sortm1(Luc)Kael/J mice strain # 005125) were purchased from The Jackson Laboratory (Bar Harbor, ME). mT/mG and LSL-Luc mice were crossed as F1 progeny for experiments involving luciferase imaging and flow cytometry.
2.6. Vector Injections and Tissue Harvest
Cre reporter mice were injected intradermally on the right side at the base of the tail with the indicated vectors 1) RD-Ad5-Cre (5 × 1010 vp); 2) Cre mRNA-LNPs (159 µg/mL); 3) Cre plasmid DNA-LNPs (93.5 µg/mL); and 4) naked Cre plasmid DNA (93.5 µg/mL) with total volume 50 µL. The solvent PBS was injected as a control group in the same volume 50 µL. The Cre plasmid DNA-LNPs and naked Cre plasmid DNA used the same pTRS-CBh-Cre plasmid.
4 days after injection, animals were imaged using a Lumina S5 IVIS imager (Revvity) to measure luciferase expression. After imaging, the animals were euthanized and injection site skin and the draining inguinal lymph node were collected. Skin tissues were washed by PBS and then digested by the Epidermis Dissociation Kit (Epidermis Dissociation Kit, mouse; Miltenyi Biotec) for enzymatic epidermal-dermal separation. In brief, scrape off the subcutaneous fat, then place tissue pieces on top of the enzyme G solution mix with the dermal side facing downwards, incubate at 4 °C for 16 hours. Peel off the epidermis from the dermis, transfer to Buffer S and cut into smaller pieces followed by transfer to enzyme P/A mix. After incubating for 20 minutes at 37 °C in a water bath. Stop enzymatic reaction by adding PB buffer (PBS containing 0.5% bovine serum albumin (BSA)). Then dissociate the epidermis pieces with gentleMACS Dissociator (Miltenyi). Apply samples to 70 µm MACS SmartStrainer and washed by PB buffer. Discard the strainer and centrifuge samples at 300× g for 10 minutes at room temperature. Resuspended supernatant with PB buffer for further flow cytometry analysis. The right side inguinal lymph node was harvested and smashed onto a 100 µm cell strainer with the back of a syringe, rinse the strainer with PBS. Collect passed through cells with 40 µm cell strainer and rinse cell strainer with PBS. Centrifuge cells at 800× g, 4 °C for 5 min. Remove supernatant, wash with PBS twice and resuspend cells with 2 ml PBS for further study.
2.7. Flow Cytometry
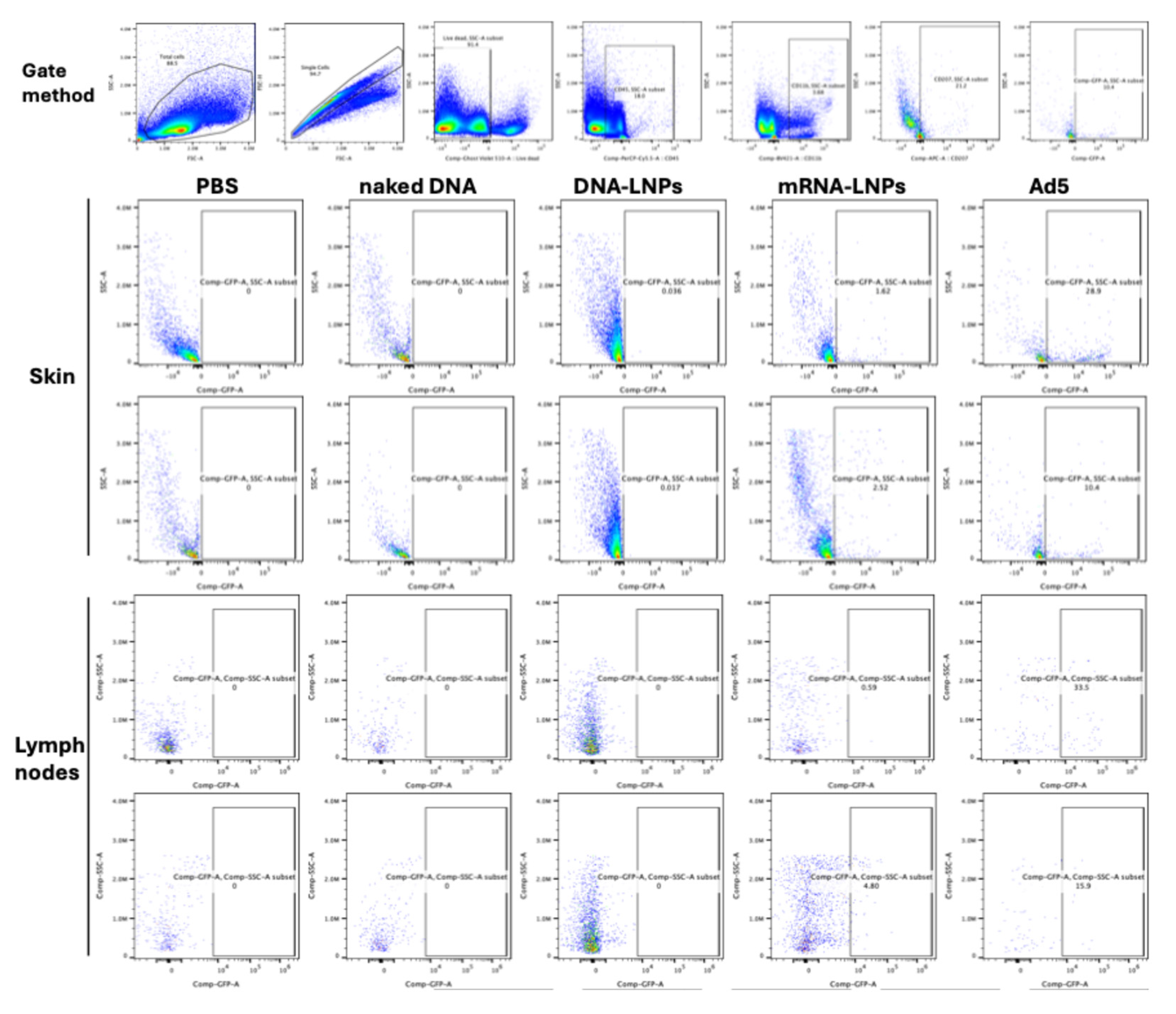
Single-cell suspensions were prepared from mouse skin and inguinal lymph nodes. Cells were washed and resuspended in staining buffer (PBS supplemented with 2% FBS). Cell viability was assessed using Ghost Violet 510 Viability Dye (Tonbo Biosciences, San Diego, CA, USA) according to the manufacturer’s instructions. After viability staining, cells were incubated with fluorochrome-conjugated monoclonal antibodies for surface marker staining for 30 min at 4 °C in the dark. The following antibodies: anti-mouse CD11b (BV421) and CD207/Langerin (APC) were from BioLegend (San Diego, CA, USA). Anti-mouse CD45 (PerCP-Cy5.5), CD3 (PE-Cy7) were from Tonbo Biosciences. After staining, cells were fixed with 4% paraformaldehyde (PFA) incubate on ice for overnight, then washed twice with staining buffer and resuspended for acquisition. Data were acquired on a Cytek northern lights spectral flow cytometer (Cytek Biosciences, Fremont, CA, USA). Spectral unmixing was performed using single-stained reference controls, prepared with either antibody cells. Fluorescence minus one (FMO) controls were included for CD207 analysis. Flow cytometry data were analyzed using FlowJo software. Doublets were excluded by FSC-A versus FSC-H gating, and dead cells were excluded based on Ghost Violet 510 staining. Immune cells were identified as CD45+, followed by analysis of CD3+ T cells, CD11b+ cells, and CD207+ dendritic cell and GFP+ subsets. The experiments were performed 3 times independently (lymph nodes n = 6, for skin tissues n = 4).
2.8. Statistical Analysis
Flow cytometry data are presented as percentages of immune cell subsets. Statistical analysis for this study was performed using GraphPad Prism version 10.4.1.3. For comparisons among multiple groups, the nonparametric Kruskal–Wallis test followed by Dunn’s multiple comparisons test was applied, with all pairwise group comparisons performed. Data are presented as individual points representing each mouse, with median and interquartile range (IQR). A p-value < 0.05 was considered statistically significant.
3. Results
3.1. Cre Reporter Mice
Cre reporter mice were used to track and “fingerprint” gene and mRNA delivery in vivo (
Figure 1 and (2)). These mice are transgenic in the Rosa26 locus for Cre-activated reporter genes. In LSL-Luc mice, firefly luciferase’s expression is blocked by a LoxP-flanked (floxed) poly-adenylation (PolyA) cassette between the CAG promoter and the luciferase gene (
Figure 1).
In the absence Cre expression, no luciferase is expressed. When Cre is delivered, it deletes the floxed PolyA to activate luciferase (
Figure 2 and (2)). mT/mG mice instead have a floxed membrane-targeted red fluorescent protein mTomato (mT) cDNA followed by membrane-targeted GFP (mG) cDNA also in the Rosa26 locus (
Figure 1).
In the absence of Cre, mT is expressed on the membranes of all cells in the mouse. When Cre is delivered, mT is deleted and membrane-targeted GFP is expressed (2). These membrane-targeted reporter proteins provide exquisite discrimination of which cells are modified by vectors when examined by confocal microscopy. When LSL mice are crossed with mT/mG mice, these hybrid mice have exactly one copy of Cre-activatable luciferase and exactly one copy of Cre-activatable mT/mG. This allows 1) in vivo imaging; 2) cell-specific transduction monitoring via mG, and 3) on/off confirmation of transduction by coordinated loss of mT. Beverly Davidson’s lab later reported a similar “foot printing” system without luciferase imaging that was used to track gene delivery of NLS-Cre-GFP and Cas9 in Ai14 mT mice (16). In this work, they demonstrated that the Cre reporter system was approximatley 8 times more sensitive than GFP delivery for detecting genetically modified cells. Thus, the Cre reporter system allows sensitive tracking of gene or mRNA delivery to cells in vivo.
3.2. In Vivo Luciferase Imaging
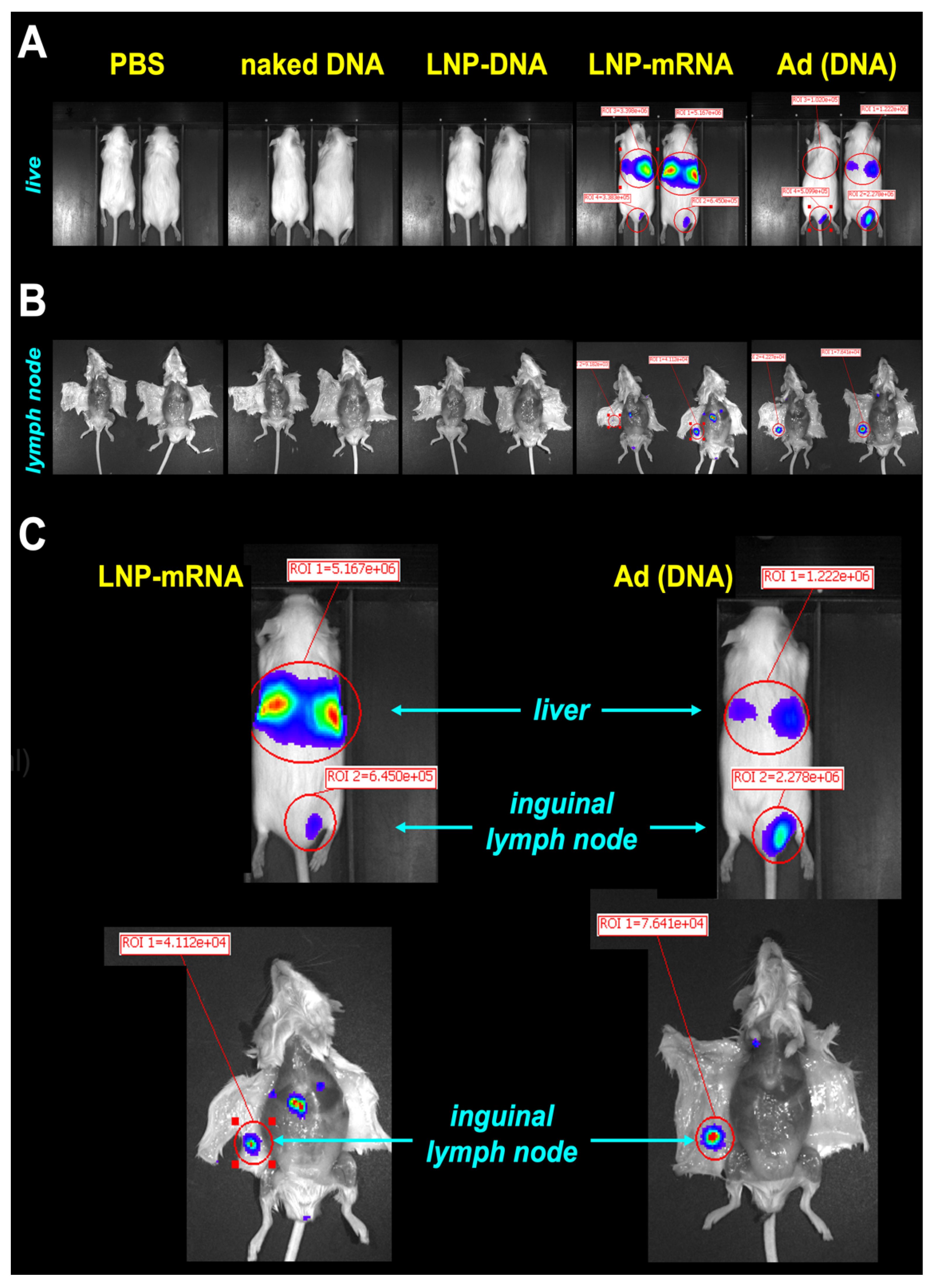
Cre reporter mice were injected intradermally (ID) with RNA and DNA-based Cre expression vectors at the base of their tail at a site that is drained by the inguinal lymph node. Four days later, the living animals were imaged for luciferase activity on an IVIS (
Figure 2A). Naked plasmid DNA and DNA in LNPs did not produce detectable luciferase activity under these conditions, In contrast, mRNA in LNPs and DNA-based Ad Cre vectors both produced IVIS signals (
Figure 2A,C). Both vectors produced luciferase at the ID site of injection. Notably, both vectors also produced stronger luciferase activity in the livers of the mice consistent with prior observations that both vectors leak into the blood [
16,
18]. The animals were euthanized and their inguinal lymph nodes were visualized on the underside of their abdominal skin (
Figure 2B,C). Imaging demonstrated visible luciferase activity in the nodes by the mRNA-LNP and Ad vectors, but not by the others.
3.3. Cre Delivery to Immune Cells in the Skin and in the Draining Lymph Node
Skin injection sites and the inguinal lymph nodes were harvested at day 4 and immune cells were isolated from each. These cells were stained with cell phenotyping antibodies against CD45 to identify immune cells, CD3 to identify T cells, CD11b to observe antigen-presenting cells, and with CD207 to identify skin Langerhans DCs in the skin and that have migrated to the draining lymph node. When skin cells were examined, flow cytometry, revealed marked activation of mG GFP in Langerhans cells by mRNA-LNP and DNA-based Ad vectors (
Figure 3). No Langerhans cells were marked by ID injection of naked DNA. When this same DNA was delivered in LNPs, there was detectable mG activation in Langerhans cells, but this was approximately 100-fold lower than Cre mRNA in LNPs and 300-fold lower than by DNA-based Ad vector.
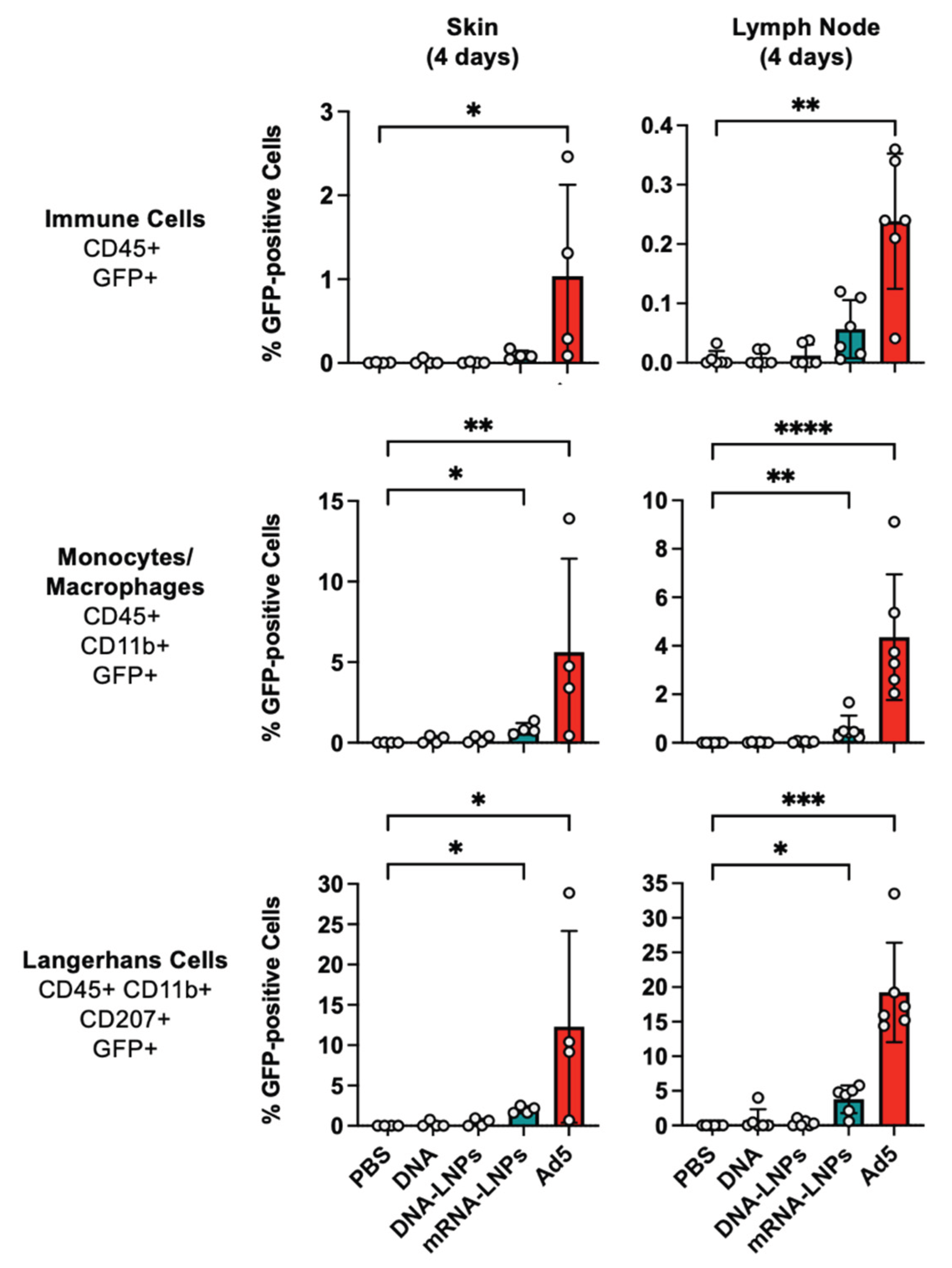
When Langerhans cells and other immune cells were examined in the skin and draining lymph node, it was apparent that the mRNA-LNP and Ad vectors were superior to the other injections (
Figure 4). In the skin, Ad activated GFP in approximately 1% of all CD45+ immune cells (p < 0.05 by Kruskal–Wallis test). CD45
+GFP
+ cells were observed in mRNA-LNP treated skin, but this did not reach significance. In the draining lymph node, Ad activated GFP in approximately 0.2% of all CD45
+ immune cells (p < 0.01 by Kruskal–Wallis test). Up to 0.1% of CD45
+ in the lymph node became GFP
+ after mRNA-LNP treatment, but this did not reach statistical significance. Occasional CD45
+GFP
+ cells were generated by DNA groups, but these were lower than in the mRNA and Ad groups.
Ad activated GFP in 4 to 5% of CD45+CD11b+GFP+ monocyte/macrophages at the injection site and in the draining lymph node (p < 0.01 and p < 0.0001). mRNA-LNPs activated GFP in approximately 1% of these monocyte/macrophage cells (p < 0.05 and p < 0.01). GFP+ monocyte/macrophages were infrequently observed in the skin.
When Langerhans cells were examined 4 days after injection, mRNA-LNPs activated GFP in 2-5% of these cells in the skin and lymph node (p < 0.05). In contrast, Ad activated GFP in up to 30% of these cells in both sites (p < 0.05 and 0.001). Rare GFP+ Langerhans cells were observed in either site after DNA delivery.
4. Discussion
The goal of this work was to map the initial transgene delivery paths of vectors relevant to gene or mRNA-based vaccines. This was tested after intradermal injection to enable simple tracking to the known inguinal draining lymph node. Under these conditions, naked DNA and DNA in LNPs mediate weak to no gene delivery to cells as determined by luciferase imaging and by flow cytometry. In contrast, mRNA delivered LNPs and DNA-based Ad vectors mediated significant delivery in antigen presenting cells and Langerhans cells in the skin. In general, the Ad vector mediated higher genetic modification of immune cells than the mRNA-LNP vector in the skin and in the draining lymph node. Notably, both of these robust vectors also mediated off-target delivery to the liver as evidenced by luciferase imaging in the living animals. This off target delivery was not unexpected for either vector and might explain to some degree certain side effects during genetic immunization.
Author Contributions
Conceptualization, M.A.B.; methodology, J.L., D.T.H., M.H., V.P.V.K., B.J.P. and L.P.; formal analysis, J.L. and M.A.B.; investigation, J.L.; writing—original draft preparation, J.L. and M.A.B.; writing—review and editing, J.L., D.T.H., M.H., V.P.V.K., B.J.P., L.P. and M.A.B.; funding acquisition, L.P. and M.A.B. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by Mayo IACUC (protocol code A00003440-18-R24 approved 5/22/24).
Data Availability Statement
Data will be provided upon request.
Acknowledgments
We would like to acknowledge the technical assistance of Mary E. Barry, Jacqulyn Jurgensen, and Zoe Dubin.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Ad |
adenovirus |
| APC |
antigen presenting cell |
| ID |
intradermal |
| GFP |
green fluorescent protein |
| Luc |
luciferase |
| mG |
membrane targeted GFP |
| RFP |
red fluorescent protein |
| mT |
membrane targeted mTomato RFP |
| VAERS |
vaccine adverse event reporting system |
| VITT |
vaccine-induced thrombotic thrombocytopenia |
References
- Prubeta, B.M. Current State of the First COVID-19 Vaccines. Vaccines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A.; Langer, F.; Makris, M.; Pai, M.; Pavord, S.; Tran, H.; Warkentin, T.E. Vaccine-induced immune thrombotic thrombocytopenia (VITT): Update on diagnosis and management considering different resources. J Thromb Haemost 2021. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.V.C.; Bouazza, F.Z.; Dauby, N.; Mullier, F.; d’Otreppe, S.; Jissendi Tchofo, P.; Bartiaux, M.; Sirjacques, C.; Roman, A.; Hermans, C.; et al. Fatal vaccine-induced immune thrombotic thrombocytopenia (VITT) post Ad26.COV2.S: first documented case outside US. Infection 2021. [Google Scholar] [CrossRef] [PubMed]
- Heidecker, B.; Dagan, N.; Balicer, R.; Eriksson, U.; Rosano, G.; Coats, A.; Tschope, C.; Kelle, S.; Poland, G.A.; Frustaci, A.; et al. Myocarditis following COVID-19 vaccine: incidence, presentation, diagnosis, pathophysiology, therapy, and outcomes put into perspective. A clinical consensus document supported by the Heart Failure Association of the European Society of Cardiology (ESC) and the ESC Working Group on Myocardial and Pericardial Diseases. Eur J Heart Fail 2022, 24, 2000–2018. [Google Scholar] [CrossRef] [PubMed]
- Petito, E.; Gresele, P. VITT Pathophysiology: An Update. Vaccines 2025, 13. [Google Scholar] [CrossRef] [PubMed]
- Prather, A.A.; Dutcher, E.G.; Robinson, J.; Lin, J.; Blackburn, E.; Hecht, F.M.; Mason, A.E.; Fromer, E.; Merino, B.; Frazier, R.; et al. Predictors of long-term neutralizing antibody titers following COVID-19 vaccination by three vaccine types: the BOOST study. Sci Rep 2023, 13, 6505. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; May, A.; Polidori, L.; Louca, P.; Wolf, J.; Capdevila, J.; Hu, C.; Ourselin, S.; Steves, C.J.; Valdes, A.M.; et al. COVID-19 vaccine waning and effectiveness and side-effects of boosters: a prospective community study from the ZOE COVID Study. Lancet Infect Dis 2022, 22, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Akyol, R.; Dalod, M. Identity, Functions, and the Spatiotemporal Maturation of Type 1 Conventional Dendritic Cells. Immunol Rev 2025, 336, e70079. [Google Scholar] [CrossRef] [PubMed]
- Vine, E.E.; Austin, P.J.; O’Neil, T.R.; Nasr, N.; Bertram, K.M.; Cunningham, A.L.; Harman, A.N. Epithelial dendritic cells vs. Langerhans cells: Implications for mucosal vaccines. Cell reports 2024, 43, 113977. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu Rev Immunol 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Melero, I.; Vile, R.G.; Colombo, M.P. Feeding dendritic cells with tumor antigens: self-service buffet or a la carte? Gene Ther 2000, 7, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Timares, L.; Takashima, A.; Johnston, S.A. Quantitative analysis of the immunopotency of genetically transfected dendritic cells. Proc Natl Acad Sci U S A 1998, 95, 13147–13152. [Google Scholar] [CrossRef] [PubMed]
- Hillestad, M.L.; Guenzel, A.J.; Nath, K.A.; Barry, M.A. A vector-host system to fingerprint virus tropism. Hum Gene Ther 2012, 23, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.D.; Nguyen, T.V.; Allen, K.L.; Ayasoufi, K.; Barry, M.A. Comparison of Gene Delivery to the Kidney by Adenovirus, Adeno-Associated Virus, and Lentiviral Vectors After Intravenous and Direct Kidney Injections. Hum Gene Ther 2019, 30, 1559–1571. [Google Scholar] [CrossRef] [PubMed]
- Crosby, C.M.; Barry, M.A. IIIa deleted adenovirus as a single-cycle genome replicating vector. Virology 2014, 462-463, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Parrett, B.J.; Yamaoka, S.; Barry, M.A. Reducing off-target expression of mRNA therapeutics and vaccines in the liver with microRNA binding sites. Mol Ther Methods Clin Dev 2025, 33, 101402. [Google Scholar] [CrossRef] [PubMed]
- Betageri, K.R.; Meridew, J.A.; Parrett, B.J.; Gilbert, R.M.; Link, P.A.; Schussler, N.A.; Mercado-Perez, A.; Caporarello, N.; Barry, M.A.; Tschumperlin, D.J. Lung-targeted Lipid Nanoparticle Delivery of a Matricellular mRNA Promotes Fibrotic Lung Repair. Am J Respir Cell Mol Biol 2025. [Google Scholar] [CrossRef] [PubMed]
- Weaver, E.A.; Nehete, P.N.; Buchl, S.S.; Senac, J.S.; Palmer, D.; Ng, P.; Sastry, K.J.; Barry, M.A. Comparison of replication-competent, first generation, and helper-dependent adenoviral vaccines. PloS one 2009, 4, e5059. [Google Scholar] [CrossRef] [PubMed]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).