Submitted:
03 May 2026
Posted:
04 May 2026
Read the latest preprint version here
Abstract
Background: Autism spectrum disorder (ASD) affects approximately 1-2% of children worldwide, yet its etiology remains incompletely understood. Emerging evidence suggests that offspring of parents with autoimmune diseases show elevated autism prevalence. Notably, children of parents with psoriasis (OR 1.59), type 1 diabetes (OR 1.49-2.36), and rheumatoid arthritis (OR 1.51) demonstrate particularly strong associations.Hypothesis: I propose that autism may be conceptualized as an immune-metabolic disorder in which multiple pro-inflammatory cytokines—including TNF-α, IL-6, IL-1β, and IFN-γ—act through distinct molecular pathways yet converge on a common endpoint of mitochondrial dysfunction and cerebral energy deficiency. This convergence implies that it is the cumulative prenatal inflammatory burden, rather than any single cytokine, that drives the energy deficit. The resulting energy shortage may impair three critical processes: (1) synaptic pruning during neurodevelopment, (2) real-time social cognition including gaze processing and emotion recognition, and (3) protein synthesis of critical synaptic scaffolding molecules.The proposed mechanism is a chronic low-grade pro-inflammatory cytokine state—clinically silent, yet biologically consequential—arising from inherited inflammatory susceptibility and/or direct fetal exposure to elevated maternal inflammatory signaling during pregnancy. Unlike high-grade inflammatory states in which maternal and fetal survival are acutely threatened, low-grade cytokine elevations may proceed without conspicuous symptoms or detectable clinical signs, particularly when chronic. Although seemingly quiet, such a state may be insufficient to endanger maternal or fetal survival, yet sufficient to disrupt fetal brain bioenergetics during sensitive gestational windows—producing neonates who appear outwardly healthy at term while their neurodevelopmental trajectories have already been altered.I further propose that the well-documented "firstborn effect" in autism reflects maternal immune maladaptation during primigravid pregnancies. Additionally, for cases without parental autoimmune history, a speculative secondary mechanism is proposed: mitonuclear immune conflict, where paternal immune genes may partially recognize maternal mitochondria as non-self, generating endogenous pro-inflammatory signaling.Implications: This framework may provide an integrative account of disparate observations about autism pathophysiology and suggests that pro-inflammatory immune pathways and mitochondrial protection strategies merit further investigation for potential risk modification, particularly in pregnancies identified as high-risk through parental autoimmune or inflammatory disease. If supported by sufficient subsequent evidence, prenatal cytokine monitoring and corresponding clinical management—currently not part of routine obstetric care—may merit consideration by the medical community as a candidate strategy for autism risk reduction.
Keywords:
pro-inflammatory cytokines
; mitochondrial dysfunction
; synaptic pruning
; neuroinflammation
; maternal immune activation
; SHANK3
; birth order effect
; maternal obesity
; decidua basalis
; microglial reprogramming
; prenatal origin
; placental amplification
; cerebral energy deficit
Lay Summary
Children of parents with certain immune-related conditions—such as psoriasis, type 1 diabetes, and rheumatoid arthritis—have a higher chance of being autistic. This paper proposes that a chronic, clinically silent, pro-inflammatory immune state during pregnancy may interfere with the energy-producing structures inside developing brain cells (mitochondria), creating an energy shortage in the fetal brain. Several different inflammatory signals, each traveling through different molecular routes, may converge on the same endpoint—compromising these energy-producing structures. Because the brain requires enormous amounts of energy during development, this shortage may disrupt processes critical for typical neurodevelopment, including the refinement of brain connections and real-time social processing such as eye contact. If confirmed, this framework could help identify pregnancies at higher risk and guide future research into potential preventive strategies.
1. Introduction
Autism spectrum disorder (ASD) is a neurodevelopmental condition characterized by deficits in social communication and interaction, restricted interests, and repetitive behaviors. Despite decades of research, the fundamental biological mechanisms underlying autism remain elusive. While genetic factors contribute substantially to autism risk, environmental and immunological factors increasingly appear to play critical roles [1].
A growing body of evidence indicates that offspring of parents with autoimmune diseases show elevated autism prevalence. Meta-analyses have demonstrated that family history of autoimmune disease is associated with a 28-50% higher autism prevalence. Importantly, these conditions involve broader pro-inflammatory immune dysregulation and may expose the developing fetus to a chronic inflammatory milieu during pregnancy.
In this paper, I propose that autism may be conceptualized as an immune-metabolic disorder in which prenatal pro-inflammatory cytokine signaling contributes to mitochondrial dysfunction and cerebral energy deficiency, impairing three critical processes: synaptic pruning during neurodevelopment, real-time social cognitive processing, and protein synthesis of synaptic scaffolding molecules. This framework may help explain core autism symptoms from a unified energetic perspective and generates testable predictions.
A central feature of this framework is convergence: multiple pro-inflammatory cytokines—including TNF-α, IL-6, IL-1β, and IFN-γ—act through distinct molecular pathways yet converge on a shared endpoint of mitochondrial dysfunction and reduced cerebral ATP production. This convergence implies that the cumulative prenatal inflammatory burden, rather than any single cytokine, drives the energy deficit central to this hypothesis. The model therefore moves beyond previous proposals centered on a single mediator (such as TNF-α or IL-6 alone) and instead positions multiple cytokine pathways as additive contributors to a common bioenergetic failure.
A central tenet of this hypothesis is that autism-related neural circuit abnormalities are established prenatally, during in utero brain development, rather than arising de novo after birth. In this view, the autistic brain is not a typically developed brain that subsequently malfunctions; it is a brain whose foundational architecture—synaptic density, connectivity patterns, and microglial programming—has been shaped by an adverse intrauterine inflammatory and metabolic environment. The clinical manifestations observed postnatally represent the downstream expression of developmental trajectories that were altered before birth.
A central implication of the present hypothesis is that ASD may arise not only from overt maternal inflammatory disease, but also from a “clinically silent, chronic low-grade pro-inflammatory cytokine state” that is insufficient to endanger maternal or fetal survival, yet sufficient to disrupt fetal brain development during sensitive gestational windows. This distinguishes the present model from acute, clinically obvious conditions such as preeclampsia or cytokine storm: in the cases addressed here, pregnancy may proceed to term with normal birth weight and outwardly healthy neonates, while the cumulative impact on fetal brain bioenergetics has already shaped neurodevelopmental trajectories.
2. Epidemiological Evidence: Parental Autoimmune Diseases and Autism Risk
2.1. Large-Scale Studies
Multiple large-scale epidemiological studies have established associations between parental autoimmune diseases and offspring autism risk. Table 1 summarizes key findings from major studies.
2.2. A Pro-Inflammatory Cytokine Common Denominator
A critical observation is that the parental diseases most strongly associated with offspring autism risk involve a broader pro-inflammatory cytokine milieu rather than a single isolated immune factor. Across these conditions, dysregulated inflammatory signaling has been implicated in:
- Psoriasis: TNF-α drives keratinocyte proliferation and inflammatory cascade; anti-TNF biologics are first-line therapy [2]
- Type 1 Diabetes: TNF-α directly induces β-cell apoptosis and promotes autoimmune destruction of pancreatic islets [7]
- Rheumatoid Arthritis: TNF-α orchestrates synovial inflammation and joint destruction; anti-TNF therapy revolutionized treatment [5]
- Normal-Tension Glaucoma: pro-inflammatory and neurodegenerative signaling has been implicated in NTG pathogenesis, and experimental glaucoma models support cytokine-related mechanisms of retinal ganglion cell loss [8]
3. Sources of Prenatal Pro-Inflammatory Cytokine Exposure
Multiple pathways can lead to elevated prenatal pro-inflammatory cytokine exposure during critical periods of neurodevelopment. This section examines four distinct sources: parental autoimmune diseases, maternal obesity, maternal immune maladaptation during first pregnancies, and endogenous mitonuclear immune conflict.
3.1. Parental Autoimmune Diseases
As detailed in Section 2, offspring of parents with autoimmune diseases including psoriasis, type 1 diabetes, and rheumatoid arthritis show elevated autism prevalence. These conditions share broader pro-inflammatory immune dysregulation as a common pathogenic feature. Children may be exposed to elevated inflammatory signaling through genetic inheritance of immune susceptibility variants and/or direct fetal exposure to maternal cytokine imbalance during pregnancy.
3.2. Maternal Obesity: An Additional Source of Prenatal Pro-Inflammatory Cytokine Exposure
Beyond parental autoimmune diseases, maternal obesity represents another condition associated with elevated offspring autism prevalence. Multiple large-scale epidemiological studies have consistently demonstrated that children of obese mothers show higher autism rates.
3.2.1. Epidemiological Evidence
3.2.2. Obesity as a Chronic Inflammatory State
Obesity is fundamentally a state of chronic low-grade inflammation [11]. Adipose tissue is not merely an energy storage organ but an active endocrine tissue that produces pro-inflammatory cytokines:
- Adipose tissue is a major source of pro-inflammatory cytokine production, including TNF-α; obese individuals express approximately 2.5-fold more TNF-α mRNA in adipose tissue than lean controls [12]
- Other inflammatory markers (IL-6, CRP, leptin) are also elevated
- Inflammation correlates with degree of adiposity [11]
3.2.3. Mechanism of Fetal Exposure
During pregnancy, maternal obesity creates multiple pathways for fetal pro-inflammatory cytokine exposure [13]:
- Transplacental passage: Maternal inflammatory cytokines can cross the placenta and directly affect fetal brain development
- Placental inflammation: The placenta itself becomes inflamed in obese pregnancies, producing additional local cytokines
- Metabolic stress: Maternal hyperglycemia and insulin resistance further compromise fetal mitochondrial function
- Oxidative stress: Obesity-associated oxidative stress damages both maternal and fetal mitochondria
3.2.4. Convergence with the Energy-Deficit Model
Maternal obesity thus represents another route to the same pathogenic endpoint: pro-inflammatory cytokine-mediated mitochondrial dysfunction in the developing fetal brain. Whether inflammatory signaling originates from parental autoimmune disease, maternal obesity, or both, the downstream consequences—impaired synaptic pruning, compromised social cognition, and protein synthesis deficits—may remain the same. I propose that this convergence may explain why maternal obesity and parental autoimmune conditions show similar effect sizes for autism risk and could have additive effects when co-occurring.
3.3. The Birth Order Effect: Maternal Immune Maladaptation
Epidemiological studies consistently demonstrate that firstborn children have significantly elevated autism risk compared to later-born siblings. This “firstborn effect” has been dismissed as reproductive stoppage (parents not having more children after an autistic child), but this explanation conflates correlation with causation.
3.3.1. Epidemiological Evidence for the Firstborn Effect

Table 2.
Birth Order and Autism Risk: Evidence Summary.
| Finding | Effect Size | Reference |
| Firstborn autism risk (Utah) | OR 1.8 | Bilder et al. (2009) [14] |
| Short interpregnancy interval (<12 mo) | OR 3.39 | Cheslack-Postava et al. (2011) [15] |
| Preeclampsia in nulliparous women | Significantly higher | Robillard et al. (1994) [16] |
| Partner change resets preeclampsia risk | Returns to primigravid | Dekker & Robillard (2007) [17] |
Note: OR = Odds Ratio. Preeclampsia data included as parallel evidence for primigravid immune maladaptation.
3.3.2. Primigravid Immune Maladaptation Mechanism
The maternal immune system must achieve tolerance to semi-allogeneic fetal antigens. This tolerance develops progressively across the first pregnancy as paternal antigen-specific regulatory T cells (Tregs) expand [16,17]. During the first pregnancy, the maternal immune system encounters paternal antigens for the first time, resulting in a Th1-dominant response with elevated pro-inflammatory cytokines. Subsequent pregnancies (with the same partner) benefit from immunological memory, with rapid Treg expansion providing enhanced tolerance [17].
3.3.3. Preeclampsia as Parallel Paradigm
Preeclampsia—characterized by placental inflammation and elevated pro-inflammatory signaling—has been called “the disease of primigravidae” since 1902. Nulliparous women have significantly higher preeclampsia risk than multiparous women, and critically, this protective effect is lost when women change partners [16]. Prior abortion with the same partner reduces preeclampsia risk by half (OR 0.54), but abortion with a different partner confers no protection [16]. These findings demonstrate that paternal antigen-specific tolerance develops during first pregnancy and provides lasting protection [17]—precisely the mechanism I propose underlies the autism birth order effect.
3.4. Mitonuclear Immune Conflict: An Endogenous Source of Pro-Inflammatory Signaling
While the preceding sections describe how pro-inflammatory cytokine-mediated mitochondrial dysfunction may contribute to autism, an important question remains: what about cases where parents have no autoimmune disease? I propose that mitonuclear immune conflict may represent an endogenous source of pro-inflammatory signaling that activates the same pathogenic pathway. This mechanism remains highly speculative and is presented as a hypothesis-generating extension of the broader cytokine-mitochondrial framework; it requires independent empirical validation.
3.4.1. The Gap in the Cytokine-Mitochondrial Hypothesis
The present energy-deficit hypothesis explains autism risk in offspring of parents with autoimmune diseases such as psoriasis, type 1 diabetes, and rheumatoid arthritis. However, autism also occurs in families with no history of autoimmune disease. Additionally, studies show that a substantial proportion of autistic individuals exhibit mitochondrial dysfunction biomarkers without carrying classical mitochondrial disease mutations.
This raises a critical question: if pro-inflammatory cytokine-mediated mitochondrial dysfunction is central to autism pathophysiology, what is the source of inflammatory signaling in cases without parental autoimmune disease?
3.4.2. The Unique Inheritance Pattern of Mitochondria
Mitochondria possess a unique inheritance pattern. Mitochondrial DNA (mtDNA) is inherited exclusively from the mother—paternal mitochondria are actively eliminated from the fertilized egg. Every mitochondrion in an individual’s body carries only maternal genetic information.
In contrast, the nuclear genome—including genes governing immune function and self/non-self recognition—is inherited from both parents. This creates an asymmetry: the immune system is shaped by both parental genomes, but the mitochondria it must tolerate are exclusively maternal.
3.4.3. The Conflict Hypothesis: Paternal Immune Genes vs. Maternal Mitochondria
I hypothesize that in some individuals, paternally inherited immune genes may fail to fully recognize maternal mitochondria as “self.” This could result in:
- Immune misrecognition: The paternal contribution to immune recognition machinery (HLA genes, innate immune pathways) may be calibrated to recognize mitochondrial signatures that differ from those inherited from the mother.
- Chronic immune attack: The immune system may mount persistent inflammatory responses against the individual’s own mitochondria, treating them as partially foreign.
- Endogenous cytokine production: This chronic immune activation would result in sustained pro-inflammatory cytokine release—activating the same pathogenic cascade described in previous sections, even without external inflammatory exposure from parental autoimmune disease.
The biological plausibility of this mechanism is supported by two established principles. First, mitochondria retain their bacterial ancestry and, when damaged or released extracellularly, their components—including mtDNA, cardiolipin, and formyl peptides—function as damage-associated molecular patterns (DAMPs) that potently activate innate immune receptors, particularly Toll-like receptor 9 (TLR9) and formyl peptide receptors [18]. Second, mitochondrial stress is known to activate NF-κB signaling, which in turn promotes production of pro-inflammatory cytokines, while released mtDNA can also activate the NLRP3 inflammasome [19]. Thus, even partial immune misrecognition of mitochondrial components could initiate a self-amplifying cycle: immune-mediated mitochondrial damage releases DAMPs, which activate NF-κB, which drives further inflammatory signaling, which causes additional mitochondrial dysfunction.
3.4.4. Two Pathways to the Same Outcome
The mitonuclear immune conflict hypothesis does not replace the parental autoimmune disease hypothesis—it complements it by providing a second pathway to pro-inflammatory cytokine-mediated mitochondrial dysfunction:
This framework is consistent with the following observations:
Table 3.
Two Pathways to Pro-Inflammatory Cytokine-Mediated Mitochondrial Dysfunction.
| Pathway 1: External | Pathway 2: Internal | |
| Source of Inflammatory Signaling | Parental autoimmune disease | Mitonuclear immune conflict |
| Mechanism | Genetic inheritance + fetal exposure during pregnancy | Paternal immune attack on maternal mitochondria |
| Parental disease required? | Yes | No |
| Final common pathway | Pro-inflammatory cytokine signaling → Mitochondrial dysfunction → Energy deficit → Autism | |
Note: Both pathways converge on the same final mechanism of pro-inflammatory cytokine-mediated mitochondrial dysfunction.
- Parental autoimmune disease is associated with elevated autism prevalence (Pathway 1)
- Autism also occurs without parental autoimmune disease (Pathway 2)
- Only a subset of children with autoimmune parents develop autism (variable mitonuclear compatibility may be protective or additive)
3.4.5. Testable Predictions and Preliminary Evidence
The mitonuclear immune conflict hypothesis generates testable predictions:
- Anti-mitochondrial antibodies or mitochondria-targeted immune markers may be elevated in autistic individuals without parental autoimmune history
- Inflammatory cytokines may be elevated even in autism cases without parental autoimmune disease
- Specific HLA haplotype combinations from parents may show associations with autism risk
Notably, the first prediction has already received preliminary support. Zhang et al. [20] reported that serum from young autistic children contains significantly elevated levels of anti-mitochondrial antibody Type 2 (AMA-M2; p = 0.001) and extracellular mitochondrial DNA (cytochrome B, p = 0.0002) compared to neurotypical controls. The authors noted that AMA-M2 had previously been detected clinically only in primary biliary cholangitis (PBC)—an autoimmune disease characterized by immune-mediated destruction of mitochondria-rich biliary epithelial cells. The presence of this antibody in autistic children is consistent with the possibility that the immune system can mount responses against mitochondrial components in autism, aligning with the mitonuclear immune conflict mechanism proposed here. Whether parental PBC is associated with elevated offspring autism risk remains untested and represents an additional testable prediction of this hypothesis.
3.5. The Decidual-Placental Transmission Pathway: How Pro-Inflammatory Signals Reach the Fetal Brain
The preceding sections have identified multiple sources of prenatal pro-inflammatory signaling relevant to autism risk. However, a critical spatial question remains insufficiently addressed: through what anatomical pathway do maternal or endogenous inflammatory signals reach the developing fetal brain? I propose that the decidua basalis—the specialized maternal tissue forming the immunological core of the placenta—represents a key intermediate locus in this transmission chain.
3.5.1. The Decidual Immune Tolerance Circuit
The decidua basalis harbors a uniquely high density of immune cells, including uterine natural killer cells (uNK, ~50–70% of decidual leukocytes), decidual macrophages (~20–25%), and regulatory T cells (Tregs) [21]. In normal pregnancy, these populations maintain a coordinated tolerogenic circuit: Tregs suppress maternal immune responses against paternally derived fetal antigens, macrophages adopt an anti-inflammatory M2 phenotype producing IL-10 and TGF-β, and uNK cells promote spiral artery remodeling through controlled IFN-γ secretion rather than cytotoxic activity [22]. This circuit ensures that the local immune environment at the maternal-fetal interface remains anti-inflammatory and supportive of placental development.
3.5.2. Disruption of the Tolerogenic Circuit
I hypothesize that in mothers with autoimmune disease, chronic inflammation, or other predisposing conditions described in Section 3.1, Section 3.2, Section 3.3 and Section 3.4, this tolerogenic circuit may become dysregulated through three converging mechanisms: (1) Treg insufficiency, leading to inadequate suppression of pro-inflammatory responses at the maternal-fetal interface; (2) macrophage polarization shift from M2 (anti-inflammatory) toward M1 (pro-inflammatory), increasing local TNF-α, IL-6, IL-1β, and IFN-γ production; and (3) uNK cell dysfunction, shifting from controlled IFN-γ-mediated vascular remodeling toward excessive pro-inflammatory cytokine release, impairing spiral artery remodeling and contributing to chronic placental hypoperfusion [21,22]. These mechanisms are not independent but mutually reinforcing: Treg insufficiency promotes macrophage M1 polarization, which further destabilizes the local immune environment.
3.5.3. Placental Amplification and Fetal Exposure
A critical feature of this pathway is that the placenta does not function as a passive filter. Placental trophoblast cells express receptors for TNF-α and IL-6, and respond actively to inflammatory stimulation by increasing their own cytokine production [23]. This means that a relatively modest inflammatory signal originating in the decidual compartment can be amplified through placental relay, resulting in disproportionate cytokine exposure at the fetal level. This amplification mechanism may explain a clinically important observation: why ASD occurs in offspring of mothers who appear outwardly healthy during pregnancy.
3.5.4. Subclinical Decidual Inflammation: Why Healthy-Appearing Mothers Can Have Affected Offspring
A central implication of this model is that clinically significant immune dysregulation at the decidual level need not manifest as overt maternal illness. The decidual immune environment is relatively compartmentalized from the systemic maternal circulation [21,22]; its immune cell composition and cytokine profile are shaped by local factors including progesterone signaling, trophoblast-derived HLA-G molecules, and paracrine interactions among resident immune cells. Consequently, a mother may exhibit normal or subclinical systemic inflammatory markers while harboring a substantially dysregulated immune environment at the decidual level. This dissociation between systemic and local immune states is fundamentally different from conditions such as preeclampsia or cytokine storm, in which maternal systemic inflammation is clinically apparent. The present model proposes that even this “quiet” decidual inflammation—operating below the threshold of clinical detection—may generate sufficient pro-inflammatory signaling, amplified through placental relay, to impair fetal brain mitochondrial function during critical developmental windows. Although immune dysregulation at the decidual-placental interface may exert effects throughout gestation, the second trimester may represent a particularly important convergence window, as this period coincides with the peak of microglial colonization of the fetal brain, the maturation of placental transport function, active synaptogenesis and early circuit formation, and the height of decidual immune activity [21,22,23]. However, the precise gestational timing of maximal vulnerability likely varies across individuals and may depend on the nature, severity, and duration of the maternal immune perturbation.
This framework helps resolve a persistent puzzle in autism epidemiology: why the majority of mothers who deliver autistic children have unremarkable pregnancy histories. If the operative locus of immune dysregulation is the decidua rather than the systemic maternal compartment, then conventional prenatal screening—which relies on systemic markers—would be expected to miss these cases.
3.5.5. Microglial Reprogramming: The Bridge from Placental Inflammation to Aberrant Synaptic Pruning
The pathway from decidual-placental inflammation to disrupted fetal brain development requires a cellular intermediary within the fetal brain itself. Microglia—the resident immune cells of the central nervous system—serve this role. During normal fetal development, microglia originating from the yolk sac colonize the brain primarily during the second trimester, where they perform essential functions including synaptic pruning, neuronal circuit refinement, and clearance of apoptotic cells [24,25]. These functions are activity-dependent and energetically costly, requiring precise calibration of microglial activation state.
When the placental relay transmits elevated pro-inflammatory cytokines to the fetal compartment, fetal microglia may undergo functional reprogramming toward a chronically activated, pro-inflammatory phenotype [24,25]. This reprogramming has two consequences directly relevant to autism: first, chronically activated microglia may execute synaptic pruning aberrantly—either excessively or insufficiently—disrupting the normal refinement of neural circuits during critical developmental windows. This aligns with the excess synaptic density observed in postmortem autism brain tissue (Section 5.1). Second, activated microglia may increase local pro-inflammatory signaling within the fetal brain itself, potentially creating a self-sustaining neuroinflammatory loop that persists beyond the initial prenatal insult. This mechanism provides a cellular explanation for how a transient gestational exposure can produce permanent alterations in brain architecture—the microglia are reprogrammed during a critical window, and their altered pruning behavior shapes the synaptic landscape of the developing brain irreversibly.
Such low-grade immune dysregulation—below the clinical threshold of conditions such as preeclampsia or cytokine storm, yet above the level compatible with normal fetal neurodevelopment—may define an “autism-risk immune activation zone” in which clinically silent decidual inflammation remains biologically consequential for fetal neurodevelopment. The precise immunological parameters defining this zone, including cytokine thresholds, Treg counts, and decidual macrophage polarization ratios, remain to be established through future investigation.
4. Pro-Inflammatory Cytokines and Mitochondrial Dysfunction: The Mechanistic Link
Before detailing the mechanisms by which inflammatory signaling may impair mitochondrial function, it is important to note that elevated pro-inflammatory cytokine activity in autism is not merely hypothetical—it is supported by converging evidence. Vargas et al. [26] demonstrated active neuroinflammation in postmortem brain tissue from autistic individuals, with marked activation of microglia and astroglia and elevated pro-inflammatory cytokine profiles including TNF-α in both brain parenchyma and cerebrospinal fluid. Li et al. [27] independently confirmed elevated immune responses in the brains of autistic patients, reporting increased expression of pro-inflammatory cytokines in frontal cortex tissue. At the peripheral level, Ashwood et al. [28] and Xie et al. [29] identified dysregulated cytokine profiles in autistic children compared to typically developing controls, with inflammatory markers correlating with greater behavioral symptom severity. These converging lines of evidence—from postmortem brain tissue, cortical immune profiling, and peripheral blood—support the relevance of inflammatory signaling to autism [30], providing an empirical basis for the mechanistic model that follows.
An important caveat is warranted: the cross-sectional nature of existing cytokine studies cannot establish whether elevated pro-inflammatory signaling is a cause of autistic neurodevelopment, a consequence of ongoing neuroinflammatory processes, or both. The present hypothesis proposes a causal role for prenatal inflammatory signaling based on three lines of reasoning: (1) parental autoimmune diseases characterized by chronic inflammatory dysregulation precede and predict offspring autism risk, establishing temporal precedence; (2) pro-inflammatory signaling has plausible inhibitory effects on mitochondrial function, providing mechanistic plausibility; and (3) immune-modifying interventions in inflammatory disorders show that these pathways are biologically actionable. Nevertheless, definitive causal evidence would require prospective longitudinal studies tracking prenatal inflammatory states and subsequent neurodevelopmental outcomes.
4.1. Illustrative Effects of TNF-α on Mitochondrial Function
The following sections use TNF-α as one illustrative example of cytokine-induced mitochondrial dysfunction; however, the broader hypothesis developed here does not depend on TNF-α alone and may apply to multiple pro-inflammatory mediators, including IL-6, IL-1β, and IFN-γ.
TNF-α exerts profound inhibitory effects on mitochondrial function through multiple mechanisms. Table 4 summarizes the key pathways by which TNF-α impairs cellular energy production.
4.2. Rapid Neurotoxicity as an Illustrative Cytokine Example
Critically, TNF-α-induced mitochondrial dysfunction illustrates how inflammatory cytokines can rapidly impair neuronal bioenergetics. Studies using pathophysiologically relevant concentrations demonstrate [31]:
- Reduction in mitochondrial basal respiration within 1.5 hours of TNF-α exposure
- Decreased ATP production preceding neuronal cell death
- Effects mediated specifically through TNF-R1 receptor signaling
- Cascade involving caspase-8 activation, membrane potential collapse, and cytochrome c release
4.3. Beyond TNF-α: Convergent Mitochondrial Effects of Other Pro-Inflammatory Cytokines
While TNF-α provides the most detailed mechanistic illustration of cytokine-induced mitochondrial dysfunction, other pro-inflammatory cytokines implicated in maternal immune activation converge on similar bioenergetic endpoints through distinct molecular pathways. This convergence strengthens the broader hypothesis that it is the cumulative inflammatory burden—rather than any single cytokine—that drives prenatal mitochondrial dysfunction.
4.3.1. IL-6: JAK-STAT3 Pathway and Mitochondrial Respiration
IL-6 is the most extensively studied cytokine in maternal immune activation (MIA) models of autism. Smith et al. (2007) demonstrated that IL-6 alone is sufficient to induce autism-like behavioral phenotypes in mouse offspring exposed during gestation [44]. Mechanistically, IL-6 signals through the gp130-JAK1-STAT3 axis, and STAT3 has been shown to localize to mitochondria where it modulates Complex I activity and mitochondrial respiration [45]. A 2025 study in Cell Reports demonstrated that IL-6 directly suppresses mitochondrial respiration through the gp130-JAK1/STAT1/3-HIF1α/ERRα axis, and that this suppression is additive with other gp130 family cytokines including IL-11 and oncostatin M [46]. Furthermore, IL-6 has been shown to disrupt protein translation and radial glia development in human brain organoid models of maternal immune activation [43], directly linking this cytokine to the type of neurodevelopmental disruption central to the present hypothesis.
4.3.2. IL-1β: NLRP3 Inflammasome and Mitochondrial-Immune Crosstalk
IL-1β is produced through activation of the NLRP3 inflammasome, a multiprotein complex that senses cellular stress signals including mitochondrial dysfunction itself [47]. This creates a bidirectional relationship: mitochondrial damage (through release of mtROS and mtDNA) activates the NLRP3 inflammasome, which produces IL-1β, which in turn exacerbates mitochondrial dysfunction—establishing a self-amplifying inflammatory loop [47]. In the context of the developing brain, this feedback cycle is particularly concerning: chronic low-grade NLRP3 activation in decidual or placental tissue could sustain IL-1β production that impairs fetal mitochondrial function without triggering overt maternal symptoms. Additionally, IL-1β increases blood-brain barrier permeability, potentially facilitating the entry of other inflammatory mediators into the fetal brain compartment.
4.3.3. IFN-γ: Metabolic Reprogramming and Complex II Inhibition
IFN-γ affects mitochondrial function through a mechanism distinct from TNF-α and IL-6. Rather than primarily targeting Complex I, IFN-γ reprograms host mitochondrial metabolism through inhibition of Complex II (succinate dehydrogenase) and promotes itaconate production, which further suppresses oxidative phosphorylation [48]. In macrophages, IFN-γ combined with pathogen-associated signals can collapse mitochondrial respiration entirely [49]. Chronic low-level IFN-γ expression has been shown to disrupt Complex I activity specifically in macrophages, driving a metabolic shift from oxidative phosphorylation to glycolysis [50]. In the decidual context, where uNK cells are a major source of IFN-γ, dysregulated IFN-γ production could simultaneously impair spiral artery remodeling (Section 3.5.1) and contribute to local mitochondrial stress in placental and fetal tissues.
Taken together, these findings indicate that multiple pro-inflammatory cytokines—each through distinct molecular mechanisms—converge on the common endpoint of mitochondrial dysfunction and reduced ATP production. Table 4a summarizes these convergent pathways.
Table 4.a.
Pro-Inflammatory Cytokines and Their Mitochondrial Targets.
| Cytokine | Primary Signaling Pathway | Mitochondrial Target | Functional Consequence |
| TNF-α | NF-κB / caspase cascade | Complex I, III; membrane potential | ETC inhibition, ROS ↑, ATP ↓ |
| IL-6 | gp130-JAK1-STAT3-HIF1α | Complex I (via mitoSTAT3); respiration | Respiration suppression, metabolic shift |
| IL-1β | NLRP3 inflammasome / NF-κB | mtROS/mtDNA feedback loop | Self-amplifying inflammation, BBB permeability ↑ |
| IFN-γ | JAK-STAT1 / itaconate | Complex II (SDH); OXPHOS → glycolysis | Metabolic reprogramming, OXPHOS collapse |
Note: Each cytokine reaches the same functional endpoint—reduced mitochondrial ATP production—through distinct molecular mechanisms, supporting the hypothesis that cumulative pro-inflammatory burden rather than any single cytokine drives the energy deficit.
4.4. Tissue-Specific Vulnerability: Why the Brain Bears the Burden
A potential objection to the energy-deficit hypothesis is the clinical observation that many autistic individuals exhibit normal or even exceptional physical strength. If mitochondrial dysfunction is central to autism pathophysiology, why does skeletal muscle appear largely unaffected? The answer lies in the fundamentally different energy metabolic profiles of these tissues, and closer examination reveals that muscle is not entirely spared.
4.4.1. Divergent Energy Dependencies of Brain and Skeletal Muscle
The brain consumes roughly 20% of total body energy despite constituting only 2% of body weight [32], and depends almost exclusively on mitochondrial oxidative phosphorylation (OXPHOS). Skeletal muscle, by contrast, possesses robust glycolytic capacity: fast-twitch fibers can generate ATP rapidly through anaerobic glycolysis independent of mitochondrial function. This asymmetry means that partial OXPHOS impairment preferentially affects the brain while leaving gross motor function relatively intact (Table 5). Notably, the perception of exceptional strength in autistic individuals may in some cases reflect catecholamine-driven fight-or-flight responses during emotional dysregulation—transient sympathetic activation rather than evidence of sustained muscle power.
4.4.2. Hypotonia and Hyperlactatemia: Evidence That Muscle Is Not Entirely Spared
Despite preserved gross motor strength, skeletal muscle does show signs of mitochondrial compromise. Hypotonia is one of the most frequently reported motor findings in autistic children, with prevalence estimates of 30–51% [33]—comparable to rates seen in primary mitochondrial diseases such as MELAS and Leigh syndrome. Additionally, Rossignol and Frye [34] found that approximately 30% of autistic children exhibit elevated blood lactate, indicating compensatory upregulation of anaerobic glycolysis in response to impaired OXPHOS. This metabolic signature is precisely what the energy-deficit hypothesis predicts. The pattern mirrors established mitochondrial disorders, where neurological symptoms typically precede and dominate over myopathic features due to the brain’s extreme OXPHOS dependence.
5. Consequences of Cerebral Energy Deficit
The present model posits that chronic prenatal pro-inflammatory signaling, regardless of its precise upstream composition, leads to persistent mitochondrial dysfunction and cerebral energy deficiency. As established in Section 4.4, the brain’s near-exclusive dependence on oxidative phosphorylation renders it uniquely vulnerable to this energy deficit, which manifests in four critical domains that explain core autism symptoms and associated features.
5.1. Impaired Synaptic Pruning
The Energy Cost of Synaptic Pruning: The developing brain undergoes massive synaptic pruning, eliminating approximately 50% of synapses from infancy to adolescence. This process is extraordinarily energy-intensive because:
- Microglia actively phagocytose synapses, requiring substantial ATP
- The infant brain consumes 40% of total body energy [32]—far exceeding adult proportions
- Complement cascade activation and autophagy pathways require ATP
Evidence of Pruning Deficits in Autism: Postmortem studies reveal striking differences in synaptic density between autistic and neurotypical brains:
Table 6.
Synaptic Pruning in Neurotypical vs Autistic Brains.
| Parameter | Neurotypical | Autism |
| Synaptic density reduction (childhood→adolescence) | ~50% | ~16% |
| Dendritic spine density | Normal | Elevated |
| mTOR pathway activity | Normal | Hyperactive |
| Autophagy function | Normal | Impaired |
Source: Tang et al. [35], Neuron (Columbia University study).
Consequences of Excess Synapses: The failure to prune synapses results in:
- Local over-connectivity: Excess short-range connections creating “neural noise”
- Long-distance under-connectivity: Insufficient resources for developing major “highway” connections between brain regions
- Reduced signal-to-noise ratio: Difficulty filtering relevant from irrelevant information
- Sensory overload: Heightened sensitivity due to failure to attenuate sensory inputs
5.2. Impaired Social Cognition and Gaze Avoidance
The Energy Demands of Social Processing: Social cognition—including face recognition, gaze processing, and emotion interpretation—is among the most computationally and energetically demanding brain functions. It requires simultaneous activation of:
- Fusiform Face Area (FFA): Face identity processing
- Superior Temporal Sulcus (STS): Gaze direction and biological motion
- Amygdala: Emotional salience and threat detection
- Prefrontal Cortex: Social context integration and decision-making
Eye Contact as Energy Conservation: This framework suggests that gaze avoidance in autism may represent an adaptive energy conservation strategy. Several lines of evidence are consistent with this interpretation:
Table 7.
Self-Reported Experiences of Eye Contact in Autism.
| Experience Category | Representative Quote |
| Energy Exertion | “Eye contact feels like I’m using up a lot of energy. Maximum 2-6 seconds.” |
| Audiovisual Integration Failure | “I cannot listen to someone while making eye contact at the same time.” |
| Cognitive Trade-off | “When I focus on eye contact, I can’t process what’s being said.” |
| Recovery Requirement | “The longer I maintain eye contact, the more recovery time I need afterward.” |
Source: Trevisan et al. [36], PLOS ONE - Qualitative analysis of first-hand accounts.
Neural Evidence: Functional neuroimaging studies demonstrate that in autism, eye contact triggers amygdala hyperactivation, suggesting heightened metabolic demand [37]. Gaze avoidance may thus function to reduce this hyperarousal and conserve limited neural energy for other cognitive tasks.
5.3. Epilepsy Comorbidity as Supporting Evidence
The high comorbidity between autism and epilepsy provides additional support for the pro-inflammatory cytokine-mediated energy deficit hypothesis. Approximately 20-30% of autistic individuals experience epileptic seizures, compared to 1-2% in the general population. In autism with intellectual disability, prevalence rises to approximately 40% [38].
Pro-Inflammatory Cytokines and Seizure Susceptibility: Pro-inflammatory cytokines, particularly TNF-α, can directly increase neuronal excitability through multiple mechanisms:
- TNF-α, for example, has been reported to increase AMPA receptor surface expression, enhancing excitatory transmission
- Pro-inflammatory signaling may promote GABA receptor internalization, reducing inhibitory tone
- The resulting excitation/inhibition imbalance may lower seizure threshold
Energy Deficit and Seizure Vulnerability: Mitochondrial dysfunction further predisposes to seizures through:
- Impaired Na+/K+-ATPase function due to ATP deficit, destabilizing membrane potential
- Compromised GABAergic inhibition, which is highly energy-dependent
- Notably, primary mitochondrial diseases (e.g., MELAS, Leigh syndrome) frequently present with epilepsy
The autism-epilepsy comorbidity thus reflects converging consequences of pro-inflammatory cytokine-mediated neuroinflammation and mitochondrial energy deficit: neuroinflammation increases excitability while energy deficit impairs the inhibitory circuits required to prevent seizures.
5.4. Impaired Protein Synthesis: The Critical Energy Bottleneck
Protein synthesis is the most energy-intensive cellular process, consuming approximately 25-30% of total cellular ATP [39]. Each amino acid incorporation requires ~4 ATP equivalents, making the synthesis of large synaptic proteins extraordinarily energy-demanding. During neurodevelopment, when neurons must produce vast quantities of synaptic proteins, any ATP deficit creates a critical bottleneck.
5.4.1. Mitochondrial Protein Synthesis as Rate-Limiting Step
The electron transport chain requires both nuclear- and mitochondrial-genome-encoded subunits. Critically, mitochondrial protein synthesis by the 55S ribosome is the rate-limiting step in ETC synthesis [40]. Of the 230 genes in the Mitochondrial Central Dogma, 59 are associated with neurodevelopmental delay, representing a 2-fold enrichment (p < 8.95E-9) [40]. This age of onset coincides with the brain’s peak glutamatergic synapse density, emphasizing the developmental linkage between energy consumption and brain maturation.
Table 8.
Critical Synaptic Proteins Vulnerable to Energy Deficit.
| Protein | Function | ASD Association | Energy Cost |
| SHANK3 | Synaptic scaffold | 0.5-2% of ASD cases | Large (1,731 aa) |
| NRXN/NLGN | Synaptic adhesion | Multiple variants | Transmembrane |
| PSD-95 | Postsynaptic density | Altered in ASD | Scaffold assembly |
| BDNF | Neuron survival | Reduced in ASD | Activity-dependent |
| FMRP | mRNA regulation | Fragile X syndrome | Translation control |
Note: aa = amino acids. All listed proteins are critical for synaptic formation and function.
5.4.2. The SHANK3-Mitochondria Connection
Phelan-McDermid syndrome, caused by SHANK3 deletion, may illustrate the intimate connection between synaptic proteins and mitochondrial function. The 22q13.3 region containing SHANK3 also harbors several mitochondrial genes, including SCO2 (cytochrome c oxidase assembly), NDUFA6 (Complex I), TYMP, TRMU (mtDNA maintenance), CPT1B (fatty acid metabolism), and ACO2 (TCA cycle). Deletions affecting SHANK3 may therefore simultaneously disrupt mitochondrial function, potentially creating a dual vulnerability.
6. Integrated Pathophysiological Model
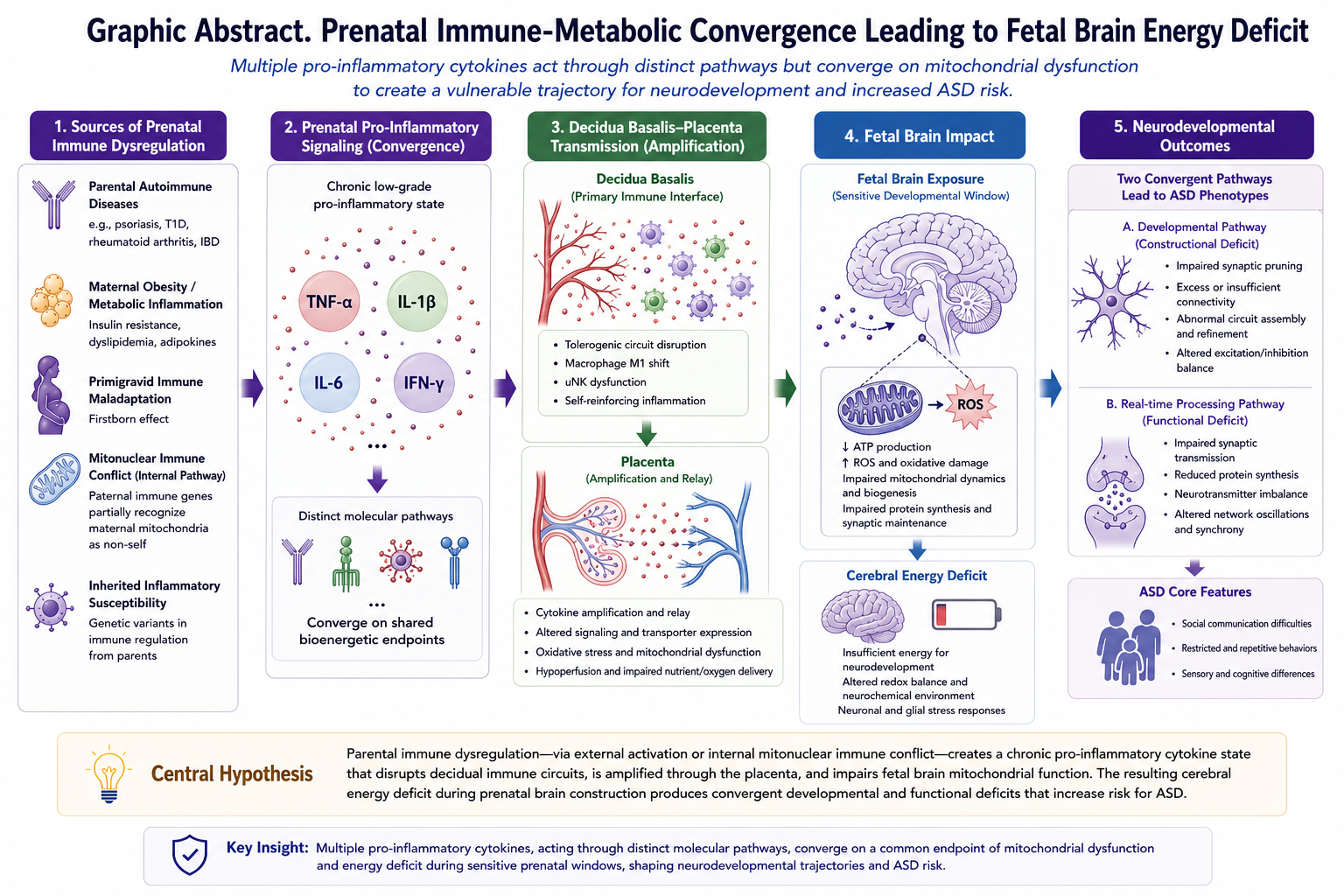
Figure 1 presents the unified model linking parental immune dysregulation—via both external (maternal systemic inflammation) and internal (mitonuclear immune conflict) pathways—to offspring autism through decidual immune circuit disruption, placental amplification, fetal brain mitochondrial dysfunction, cerebral energy crisis, and two convergent neurodevelopmental pathways leading to ASD phenotypes.
Figure 2 provides a detailed view of the decidual-placental transmission pathway proposed in Section 3.5, illustrating how subclinical decidual immune dysregulation may relay inflammatory signals to the developing fetal brain during a vulnerable gestational window.
7. Exploratory Prediction: Normal-Tension Glaucoma and Autism
Normal-tension glaucoma (NTG) offers a potentially useful test case for the present hypothesis. Emerging evidence implicates TNF-α and other pro-inflammatory mediators in NTG pathogenesis, yet NTG has never been examined in the context of offspring neurodevelopment—the present framework predicts that parental NTG should be associated with elevated autism risk if the pro-inflammatory cytokine–mitochondrial pathway is operative. NTG affects retinal ganglion cells (RGCs) and shares overlapping inflammatory and neurodegenerative features with the autoimmune conditions linked to autism risk.
7.1. NTG as a Condition with Reported Inflammatory Involvement
Evidence consistent with inflammatory involvement in NTG:
- Elevated TNF-α levels have been reported in aqueous humor and serum of NTG patients [8]
- TNF-α directly induces RGC apoptosis via TNF-R1 signaling [8]
- Immune-modulating interventions have shown protective effects in some animal models [8]
- Disease progression occurs despite normal intraocular pressure, suggesting the involvement of IOP-independent neurodegenerative mechanisms
7.2. The Untested Association
If the pro-inflammatory cytokine energy deficit hypothesis is correct, children of parents with NTG may show elevated autism prevalence compared to the general population.
To my knowledge, no published study has examined the association between parental NTG and offspring autism. This remains an untested question that may be worth investigating.
Table 9.
Conditions with Reported Inflammatory Involvement and Autism Association Studies.
| Parental Condition | Reported Inflammatory Involvement | Autism Association Studied? |
| Psoriasis | Substantial | Yes (OR 1.59) |
| Type 1 Diabetes | Documented | Yes (OR 1.49-2.36) |
| Rheumatoid Arthritis | Documented | Yes (OR 1.51) |
| Normal-Tension Glaucoma | Reported | No studies identified |
Note: The absence of studies on parental NTG and offspring autism represents a critical research gap and a testable prediction of the present hypothesis.
8. Therapeutic Implications
The energy-deficit hypothesis suggests several therapeutic approaches:
8.1. Anti-Inflammatory Interventions
Among available anti-inflammatory biologics, anti-TNF agents provide the most relevant precedent for evaluating the therapeutic implications of the present hypothesis. Existing anti-TNF biologics (etanercept, infliximab, golimumab, adalimumab) have proven efficacy in TNF-α-mediated diseases. In type 1 diabetes, golimumab preserved β-cell function in a phase 2 trial [7]. These observations raise the possibility that anti-inflammatory strategies—including but not limited to TNF-α-targeted agents—could eventually be evaluated for risk modification in carefully defined high-risk pregnancies, although substantial safety, ethical, and regulatory issues would first need to be resolved.
Preclinical evidence is consistent with this therapeutic rationale. Liu et al. (2023) demonstrated in a preeclampsia mouse model that maternal TNF-α elevation drives ASD-like phenotypes in offspring through fetal NFκB signaling, and that TNF-α neutralization during pregnancy ameliorated these ASD-like behaviors and restored NFκB activation to normal levels [41]. This study provides preclinical support for a causal contribution of maternal TNF-α signaling to offspring ASD-like phenotypes and suggests that this pathway may be pharmacologically modifiable. Although Liu et al. examined a preeclampsia model specifically, their findings are consistent with the possibility that diverse prenatal inflammatory exposures converge on a shared TNF-α-dependent pathway that perturbs fetal neurodevelopment.
In humans, existing pregnancy-exposure studies of anti-TNF biologics are reassuring with respect to developmental safety. The PIANO registry and other cohort studies have not shown increased long-term neurodevelopmental risk among children exposed to anti-TNF therapy in utero [42]. However, these studies were designed to assess safety rather than ASD-specific outcomes, and none has demonstrated a reduction in autism risk. An especially informative future test of the present hypothesis would be a well-powered, disease-severity-adjusted registry analysis comparing ASD rates in offspring of biologic-treated versus untreated pregnancies in women with inflammatory disorders.
8.2. Early Identification
If validated, parental inflammatory disorders (psoriasis, T1D, RA, NTG) could serve as clinically accessible markers of pregnancies at elevated autism risk, potentially enabling earlier surveillance and stratified intervention.
8.3. Toward Future Risk Stratification: Prenatal Cytokine Monitoring
A core implication of the present hypothesis is that the majority of pregnancies producing autistic offspring may involve subclinical, low-grade inflammatory states that escape detection by conventional prenatal screening, which relies on systemic markers and clinical symptomatology. If supported by sufficient subsequent evidence, prenatal cytokine monitoring—currently not part of routine obstetric care—may merit consideration by the medical community as a candidate strategy for autism risk reduction.
Such monitoring would face substantial methodological challenges. These include: (1) establishing population-specific reference ranges for pregnancy-relevant cytokines, given that some degree of pro-inflammatory signaling is physiologically necessary across gestation; (2) distinguishing physiologically necessary cytokine fluctuations from pathogenic ones, particularly across the implantation, mid-gestation, and parturition windows in which immune profiles normally shift; and (3) identifying which cytokines, or cytokine combinations, carry the strongest predictive value for offspring neurodevelopmental outcomes. Furthermore, the appropriate clinical response to detected elevations remains undefined, as targeted anti-inflammatory interventions during pregnancy require careful consideration of fetal safety.
Nevertheless, the cumulative evidence reviewed here suggests that systematic investigation of this approach—beginning with longitudinal cohort studies linking maternal cytokine profiles across gestation to offspring neurodevelopmental outcomes—is warranted as a foundation for any future clinical translation. Such studies would also help establish whether the hypothesized “autism-risk immune activation zone” (Section 3.5.5) can be empirically defined.
9. Limitations and Future Directions
Limitations: This paper synthesizes existing evidence into a theoretical framework and does not present new experimental data. Several specific limitations should be acknowledged. First, the epidemiological associations between parental autoimmune diseases and offspring autism risk are based on observational studies and may be subject to confounding by shared genetic factors; sibling comparison designs that could better control for familial confounding have not been applied to most of the specific associations cited here. Second, while pro-inflammatory cytokine elevation is consistently observed in autistic individuals, the causal direction of this association remains unestablished. Third, the effect sizes for parental autoimmune disease–autism associations (OR 1.28–2.36) are moderate and are compatible with a cytokine-mediated mechanism but do not exclude the contribution of parallel or partially overlapping pathways. Fourth, the mitonuclear immune conflict hypothesis (Section 3.4) remains speculative, and while preliminary evidence from Zhang et al. [20] is consistent with this mechanism, larger studies with parental autoimmune stratification are needed. Fifth, although anti-TNF exposure during pregnancy would represent a natural human quasi-intervention relevant to the hypothesis, current clinical data primarily address safety outcomes and composite neurodevelopmental endpoints rather than ASD-specific risk reduction; thus, the present hypothesis is not yet supported by direct human interventional evidence. Sixth, the present hypothesis does not assign primacy to any single cytokine, but instead proposes that cumulative prenatal pro-inflammatory signaling may converge on mitochondrial dysfunction and cerebral energy deficit as a common pathogenic pathway. Cytokine networks are bidirectional and highly interactive; for example, IL-6/JAK-STAT signaling may amplify TNF-α production in some contexts, whereas TNF-α can reciprocally induce IL-6. The relative contribution of each cytokine to prenatal mitochondrial dysfunction remains to be established. Seventh, the decidual-placental transmission pathway proposed in Section 3.5 remains a hypothesis built on indirect evidence; direct human studies linking decidual immune parameters specifically to offspring ASD outcomes are extremely limited, and the distinction between decidual-level and systemic-level immune contributions to fetal neurodevelopmental risk has not yet been empirically resolved.
Future Directions: Key studies needed include: (1) Epidemiological investigation of parental NTG and offspring autism risk; (2) Longitudinal studies of mitochondrial function in infants at high autism risk; (3) Clinical trials of mitochondrial support interventions; (4) Mechanistic studies of how prenatal pro-inflammatory cytokine states alter synaptic pruning and bioenergetics in animal models; (5) disease-severity-adjusted registry analyses comparing offspring outcomes across pregnancies with differing inflammatory burdens and immune-modifying treatments; and (6) targeted investigation of decidual immune parameters—including macrophage polarization state, Treg density, and local cytokine concentrations—in pregnancies followed prospectively for offspring neurodevelopmental outcomes. Accordingly, the present model should be interpreted as a convergent, hypothesis-generating framework rather than a definitive causal account of ASD.
10. Conclusions
I propose that autism spectrum disorder may be understood as an immune-metabolic disorder in which multiple pro-inflammatory cytokines—each acting through distinct molecular mechanisms—converge on a common pathogenic pathway of mitochondrial dysfunction and cerebral energy deficiency. The hypothesis does not assign primacy to any single cytokine; rather, it proposes that cumulative prenatal pro-inflammatory signaling, regardless of its precise composition, may impair fetal brain bioenergetics during sensitive developmental windows. This energy deficit may impair synaptic pruning during development and compromise real-time social cognitive processing, potentially explaining core autism symptoms from a unified mechanistic perspective.
The hypothesis also generates an exploratory, testable prediction: that parents with normal-tension glaucoma—a neurodegenerative condition in which inflammatory signaling has been implicated, but not previously linked to offspring autism—may show elevated prevalence of autistic children. Confirmation of this prediction would be consistent with the broader hypothesis.
This hypothesis therefore locates the origin of pathology not primarily in postnatal dysfunction, but in disrupted prenatal brain construction. In the subgroup of cases addressed here, aberrant immune signaling during pregnancy may impair the formation of neural circuits before birth, such that vulnerability is built into early brain development itself.
If validated, this framework carries a fundamental implication: autism, in the subgroup of cases addressed here, is a condition of prenatal origin. The neural circuit abnormalities that underlie autistic traits—excess synaptic density, altered connectivity, and compromised real-time social processing—are not acquired postnatally but are established during fetal brain development through in utero inflammatory and metabolic insult. This reframing suggests that the most effective window for risk modification is not after diagnosis in early childhood, but during pregnancy itself—through identification of high-risk pregnancies based on parental immune profiles and, if future research supports it, through targeted anti-inflammatory or mitochondrial support interventions during the prenatal period.
References
- Meltzer A, Van de Water J. The role of the immune system in autism spectrum disorder. Neuropsychopharmacology. 2017;42(1):284-298.
- Wu S, Ding Y, Wu F, et al. Family history of autoimmune diseases is associated with an increased risk of autism in children: A systematic review and meta-analysis. Neurosci Biobehav Rev. 2015;55:322-332.
- Xiang AH, Wang X, Martinez MP, et al. Maternal Type 1 Diabetes and Risk of Autism in Offspring. JAMA. 2018;320(1):89-91.
- Persson M, et al. Maternal type 1 diabetes, pre-term birth and risk of autism spectrum disorder. Int J Epidemiol. 2023;52(2):377-386.
- Keil A, Daniels JL, Forssen U, et al. Parental autoimmune diseases associated with autism spectrum disorders in offspring. Epidemiology. 2010;21(6):805-808.
- Atladóttir HO, Pedersen MG, Thorsen P, et al. Association of family history of autoimmune diseases and autism spectrum disorders. Pediatrics. 2009;124(2):687-694.
- Quattrin T, Haller MJ, Steck AK, et al. Golimumab and Beta-Cell Function in Youth with New-Onset Type 1 Diabetes. N Engl J Med. 2020;383(21):2007-2017.
- Nakazawa T, et al. Tumor necrosis factor-alpha mediates oligodendrocyte death and delayed retinal ganglion cell loss in a mouse model of glaucoma. J Neurosci. 2006;26(49):12633-12641.
- Li YM, Ou JJ, Liu L, Zhang D, Zhao JP, Tang SY. Association between maternal obesity and autism spectrum disorder in offspring: a meta-analysis. J Autism Dev Disord. 2016;46(1):95-102.
- Wang Y, Tang S, Xu S, Weng S, Liu Z. Maternal body mass index and risk of autism spectrum disorders in offspring: a meta-analysis. Sci Rep. 2016;6:34248.
- Cawthorn WP, Sethi JK. TNF-α and adipocyte biology. FEBS Lett. 2008;582(1):117-131.
- Kern PA, Saghizadeh M, Ong JM, Bosch RJ, Deem R, Simsolo RB. The expression of tumor necrosis factor in human adipose tissue: regulation by obesity, weight loss, and relationship to lipoprotein lipase. J Clin Invest. 1995;95(5):2111-2119.
- Pantham P, Aye IL, Powell TL. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta. 2015;36(7):709-715.
- Bilder D, Pinborough-Zimmerman J, Miller J, McMahon W. Prenatal, perinatal, and neonatal factors associated with autism spectrum disorders. Pediatrics. 2009;123(5):1293-1300.
- Cheslack-Postava K, Liu K, Bearman PS. Closely spaced pregnancies are associated with increased odds of autism in California sibling births. Pediatrics. 2011;127(2):246-253.
- Robillard PY, Hulsey TC, Perianin J, Janky E, Miri EH, Papiernik E. Association of pregnancy-induced hypertension with duration of sexual cohabitation before conception. Lancet. 1994;344(8928):973-975.
- Dekker G, Robillard PY. Pre-eclampsia: Is the immune maladaptation hypothesis still standing? J Reprod Immunol. 2007;76(1-2):8-16.
- Zhang Q, Raoof M, Chen Y, et al. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature. 2010;464(7285):104-107.
- Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469(7329):221-225.
- Zhang B, Angelidou A, Alysandratos KD, et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J Neuroinflammation. 2010;7:80.
- Moffett A, Colucci F. Co-evolution of NK receptors and HLA ligands in humans is driven by reproduction. Immunol Rev. 2015;267(1):283-297.
- Erlebacher A. Immunology of the maternal-fetal interface. Annu Rev Immunol. 2013;31:387-411.
- Mor G, Aldo P, Alvero AB. The unique immunological and microbial aspects of pregnancy. Nat Rev Immunol. 2017;17(8):469-482.
- Paolicelli RC, Bolasco G, Pagani F, et al. Synaptic pruning by microglia is necessary for normal brain development. Science. 2011;333(6048):1456-1458.
- Cunningham CL, Martínez-Cerdeño V, Noctor SC. Microglia regulate the number of neural precursor cells in the developing cerebral cortex. J Neurosci. 2013;33(10):4216-4233.
- Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann Neurol. 2005;57(1):67-81.
- Li X, Chauhan A, Sheikh AM, et al. Elevated immune response in the brain of autistic patients. J Neuroimmunol. 2009;207(1-2):111-116.
- Ashwood P, Krakowiak P, Hertz-Picciotto I, et al. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav Immun. 2011;25(1):40-45.
- Xie J, Huang L, Li X, et al. Immunological cytokine profiling identifies TNF-α as a key molecule dysregulated in autistic children. Oncotarget. 2017;8(47):82390-82398.
- Morris G, Berk M. The many roads to mitochondrial dysfunction in neuroimmune and neuropsychiatric disorders. BMC Med. 2015;13:68.
- Deng W, et al. Rapid mitochondrial dysfunction mediates TNF-alpha-induced neurotoxicity. J Neurochem. 2015;132(4):443-451.
- Kuzawa CW, Chugani HT, Grossman LI, et al. Metabolic costs and evolutionary implications of human brain development. Proc Natl Acad Sci U S A. 2014;111(36):13010-13015.
- Lopez-Espejo MA, Nuñez AC, Moscoso OC, Escobar RG. Clinical characteristics of children affected by autism spectrum disorder with and without generalized hypotonia. Eur J Pediatr. 2021;180(10):3243-3246.
- Rossignol DA, Frye RE. Mitochondrial dysfunction in autism spectrum disorders: a systematic review and meta-analysis. Mol Psychiatry. 2012;17(3):290-314.
- Tang G, Gudsnuk K, Kuo SH, et al. Loss of mTOR-dependent macroautophagy causes autistic-like synaptic pruning deficits. Neuron. 2014;83(5):1131-1143.
- Trevisan DA, Roberts N, Lin C, Birmingham E. How do adults and teens with self-declared Autism Spectrum Disorder experience eye contact? PLOS ONE. 2017;12(11):e0188446.
- Dalton KM, Nacewicz BM, Johnstone T, et al. Gaze fixation and the neural circuitry of face processing in autism. Nat Neurosci. 2005;8(4):519-526.
- Amiet C, Gourfinkel-An I, Bouzamondo A, et al. Epilepsy in autism is associated with intellectual disability and gender: evidence from a meta-analysis. Biol Psychiatry. 2008;64(7):577-582.
- Buttgereit F, Brand MD. A hierarchy of ATP-consuming processes in mammalian cells. Biochem J. 1995;312(Pt 1):163-167.
- Bülow P, Patgiri A, Faundez V. Mitochondrial protein synthesis and the bioenergetic cost of neurodevelopment. iScience. 2022;25(9):104920.
- Liu X, Liu H, Gu N, Pei J, Lin X, Zhao W. Preeclampsia promotes autism in offspring via maternal inflammation and fetal NFκB signaling. Life Sci Alliance. 2023;6(8):e202301957.
- Mahadevan U, Long MD, Kane SV, et al. Pregnancy and neonatal outcomes after fetal exposure to biologics and thiopurines among women with inflammatory bowel disease. Gastroenterology. 2021;160(4):1131-1139.
- Sarieva K, Kagermeier T, Khakipoor S, et al. Human brain organoid model of maternal immune activation identifies radial glia cells as selectively vulnerable. Mol Psychiatry. 2023;28(12):5077-5089.
- Smith SE, Li J, Garbett K, Mirnics K, Patterson PH. Maternal immune activation alters fetal brain development through interleukin-6. J Neurosci. 2007;27(40):10695-10702.
- Wegrzyn J, Potla R, Ber YJ, et al. Function of mitochondrial Stat3 in cellular respiration. Science. 2009;323(5915):793-797.
- Ahn J, Kim M, et al. The pro-inflammatory cytokine IL6 suppresses mitochondrial function via the gp130-JAK1/STAT1/3-HIF1α/ERRα axis. Cell Rep. 2025;44(3):115358.
- Shi J, Bhargava P, Zhang Q. NLRP3 inflammasome in neuroinflammation and central nervous system diseases. Cell Mol Immunol. 2025;22(4):341-355.
- Jessop F, Buntyn R, Schwarz B, Priest S, Bosio CM. Interferon gamma reprograms host mitochondrial metabolism through inhibition of complex II to control intracellular bacterial replication. Infect Immun. 2020;88(2):e00744-19.
- Garabatos N, Alvarez-Sierra D, et al. Pro-inflammatory macrophage activation does not require inhibition of oxidative phosphorylation. EMBO Rep. 2025;26(4):e59465.
- Kim S, et al. Chronic low-level IFN-γ expression disrupts mitochondrial complex I activity in renal macrophages: An early mechanistic driver of lupus nephritis pathogenesis. Int J Mol Sci. 2025;26(1):63.
Figure 1.
Pro-Inflammatory Cytokine–Driven Mitochondrial Dysfunction and Cerebral Energy Crisis Pathway Leading to ASD. Integration of external and internal pathways from parental immune dysregulation to core neurodevelopmental and cognitive phenotypes.
Figure 1.
Pro-Inflammatory Cytokine–Driven Mitochondrial Dysfunction and Cerebral Energy Crisis Pathway Leading to ASD. Integration of external and internal pathways from parental immune dysregulation to core neurodevelopmental and cognitive phenotypes.

Figure 2.
Decidua Basalis–Placenta–Fetal Brain Transmission Pathway. Proposed pathway by which prenatal immune-metabolic dysregulation may converge on fetal brain energy deficit and altered neurodevelopment.
Figure 2.
Decidua Basalis–Placenta–Fetal Brain Transmission Pathway. Proposed pathway by which prenatal immune-metabolic dysregulation may converge on fetal brain energy deficit and altered neurodevelopment.

Table 1.
Parental Autoimmune Disease and Offspring Autism Risk.
| Parental Disease | Odds Ratio | 95% CI | Key Reference |
| Psoriasis | 1.59 | 1.21-2.10 | Wu et al. (2015) [2] |
| Type 1 Diabetes (T1D) | 1.49-2.36 | 1.21-4.12 | Xiang et al. (2018); Persson et al. (2023) [3,4] |
| Rheumatoid Arthritis | 1.51 | 1.14-2.00 | Keil et al. (2010) [5] |
| Hypothyroidism | 1.64 | 1.16-2.32 | Atladóttir et al. (2009) [6] |
| Any Autoimmune Disease | 1.28-1.50 | 1.11-1.75 | Wu et al. (2015) [2] |
Note: All listed conditions involve inflammatory pathways in which TNF-α and other pro-inflammatory mediators have been implicated. CI = Confidence Interval.
Table 4.
Illustrative TNF-α Effects on Mitochondrial Function.
| Mechanism | Effect on Energy Metabolism |
| ETC Complex I Inhibition | Blocks electron transfer at the first step of oxidative phosphorylation |
| ETC Complex III Inhibition | Disrupts cytochrome bc1 complex function |
| Cytochrome c Oxidase (COX) | Reduces terminal electron transfer and oxygen consumption |
| Membrane Depolarization | Collapses mitochondrial membrane potential (ΔΨm), halting ATP synthesis |
| PDH Suppression | Inhibits pyruvate dehydrogenase, blocking glucose entry into TCA cycle |
| ROS Overproduction | Increases reactive oxygen species, causing oxidative damage to mitochondrial components |
| Warburg Effect Induction | Shifts metabolism to inefficient aerobic glycolysis (2 vs 36 ATP per glucose) |
Note: ETC = Electron Transport Chain; PDH = Pyruvate Dehydrogenase; TCA = Tricarboxylic Acid Cycle; ROS = Reactive Oxygen Species.
Table 5.
Energy Metabolic Profiles: Brain vs. Skeletal Muscle.
| Parameter | Brain (Neurons) | Skeletal Muscle |
| Primary ATP source | OXPHOS (~95%) | OXPHOS + glycolysis (variable) |
| Glycolytic capacity | Very limited | High (type II fibers) |
| Glycogen reserves | Minimal | Substantial |
| Tolerance to OXPHOS impairment | Very low | Moderate to high |
Note: OXPHOS = oxidative phosphorylation. Brain energy consumption during infancy is even higher (~40% of total body energy) [32], coinciding with peak vulnerability to mitochondrial dysfunction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



