
Submitted:
30 December 2025
Posted:
31 December 2025
You are already at the latest version
Abstract
This study investigates the effects of physical activity on serum cortisol levels and phagocytic capacity of the innate immune system in 8 captive bottlenose dolphins. Analysis of paired samples (n=16) revealed a significant increase in cortisol during periods of physical activity (mean increase of 1.27 µg/dL, 122% elevation), accompanied by decreased phagocytosis in granulocytes (92% reduction) and monocytes (52% reduction). Statistical analyses demonstrated consistent negative correlations between cortisol levels and phagocytic function, suggesting that physical activity influences hypothalamic-pituitary-adrenal axis activation and, consequently, innate immune system function. Sex-differentiated responses were observed, with the male showing attenuated cortisol response but maintained monocyte sensitivity. These findings highlight the complex interplay between the neuroendocrine cortisol response and immune function in cetaceans, with important implications for controlled environments management and animal welfare assessment. A multi-method statistical framework incorporating Bayesian analysis, bootstrapping, and traditional approaches ensured robust inference despite limited sample size.
Keywords:
animal welfare
; Tursiops truncatus
; controlled environments
; cortisol
; immune system
; monocytes
; granulocytes
; phagocytosis
1. Introduction
Cortisol is a glucocorticoid synthesized from cholesterol, secreted by the adrenal cortex and released into the blood [1]. In blood plasma, most cortisol (65%) binds with high intensity to corticosteroid-binding globulin (transcortin), 30% binds to albumins, while 3–5% remains in a metabolically active form (free) [1,2,3].
Cortisol secretion follows a circadian rhythm, with highest levels occurring in the early morning hours (30–50 minutes after waking) [1,2]. Additionally, certain factors such as physical activity episodes, exercise, food intake, or individual stress can generate an increase in blood cortisol levels, peaking 10–30 minutes after stimulus interaction [1,2].
In general, this neuroendocrine response can affect immune system (IS) functioning. When an individual perceives any of the described factors, a physiological response is triggered in the hypothalamus (central nervous system), which converts nerve impulses into hormonal signals, releasing cortisol into the blood, that subsequently acts on primary and secondary lymphoid organs, T and B lymphocytes, neutrophils, monocytes, and macrophages, triggering alterations in the IS [4,5,6].
The immune system communicates and modulates through cell contact or low molecular weight proteins called interleukins (IL) or cytokines. Studies in mammals have observed that corticoids can inhibit immune cell migration and proliferation, as well as numerous immune responses, such as increased chemotaxis and IL production, especially proinflammatory ones like IL-8 [7,8]. In addition, studies with free-ranging cetaceans observed that when individuals are subjected to stressful stimuli such as capture and subsequent release, proinflammatory IL-8 increases [9], potentially causing IS suppression. This provides evidence of the relationship between prolonged or constant stressor exposure, blood cortisol levels, and IS functionality.
Although analysis of cortisol levels and their relationship to stress in captive cetaceans has been addressed in several previous studies, immune system functioning and its correlation with blood cortisol levels remains a poorly explored research line. Understanding general IS functioning and how certain factors influence participating cells is crucial, as previous studies with free-ranging dolphins observed that individuals typically do not present observable clinical disease manifestations until very advanced stages, thus showing no weakness signs and avoiding predator attraction [10]. In captivity, although predators are absent, an apparently asymptomatic individual exposed to certain exogenous or endogenous factors could experience increased blood cortisol levels, potentially altering IS cell functioning [10].
The present study employs robust statistical analysis to provide evidence of the relationship between the factor “physical activity”, cortisol dynamics, and the functionality of certain cells of the innate IS in captive bottlenose dolphins, aiming to better understand these mechanisms and their implications for animal welfare.
2. Materials and Methods
2.1. Study Subjects
The study involved 8 bottlenose dolphins (Tursiops truncatus), comprising 1 male and 7 females, maintained under controlled conditions at Zoo Aquarium Madrid, Spain (Table 1).
The dolphinarium facilities include three interconnected pools: two indoor pools without public access, connected to an outdoor pool with public access. All eight dolphins could move freely among the three pools throughout the day, except during physical activities performed during training and educational interactions, when they remained in the main pool.
The amount of physical activity performed by all study subjects during public educational sessions was 15 minutes twice daily. Additionally, in the mornings before the zoo opened to visitors, caretakers conducted entertainment games and physical activities.
This demographic composition provides ecological validity, reflecting typical under controlled conditions population structures, while allowing examination of potential sex-specific responses. The broad age range allows observation of stress response patterns across different life stages, although limited male representation requires cautious interpretation of sex-based differences.
2.2. Sample Collection and Stress Minimization Protocol
Since cortisol levels are known to be higher in the morning, to assess how cortisol increases influence IS function, blood samples were collected at two different times of day:
- Time 1 (WPA—Without Physical Activity): Early morning (approx. 10 AM), before first meal and without physical activity.
- Time 2 (DPA—During Physical Activity): Late morning (approx. 12 PM), after physical training and before educational encounter activity.
Importantly, before initiating research, all individuals underwent physical and medical evaluations to assess health status and rule out any pathology that could influence results. Additionally, samples were always collected without prior feeding, as previous studies demonstrated that fish contain high cortisol concentrations that could alter results [11].
Sample collection was always carried out within the pool, without the use of any lifting platform, and was performed voluntarily by all study subjects. All individuals had been previously trained for blood sample collection. Keepers used hand signals to indicate to the individuals that they should turn slightly within the pool so that blood could be drawn from the caudal fin. During sampling, if an animal did not approach or cooperate after a couple of attempts, it was left alone without further prompting. Because it was not a forced process, the stress component was minimized, and consequently, its potential effect on blood cortisol levels was reduced.
As a result, sample collection took longer, as some days obtaining samples from all individuals was not possible, since they were never forced, restricted, restrained, or isolated for collection. A total of 96 blood samples were obtained for cortisol level analysis and 96 samples for innate immune response assessment, distributed as shown in Table 2:
2.3. Blood Cortisol Analysis
At the Biomedical Research Unit (UIB) of Alfonso X el Sabio University in Madrid, Spain, serum cortisol concentrations were evaluated using the Demeditec Cortisol ELISA Kit, a competitive immunoenzymatic assay for quantitative cortisol measurement.
Multi-well plates were coated with anti-cortisol antibody. The unknown cortisol amount in our sample competes with a cortisol-horseradish peroxidase (HRP) conjugate for binding. The bound peroxidase conjugate amount is inversely proportional to cortisol concentration in the sample. Therefore, after substrate addition, the developed color intensity is inversely proportional to sample cortisol concentration.
2.4. Innate Immune Response Analysis
To evaluate innate immune response, we analyzed granulocyte and monocyte phagocytic capacity at the UIB of Alfonso X El Sabio University and Fundación Jiménez Díaz (Madrid, Spain) using the IngoFlowEx® kit. This kit evaluates Escherichia coli (E. coli) bacterial ingestion by these immune cells, as this bacterium can infect cetaceans, including dolphins [10,12,13]. The IngoFlowEx® kit was selected because it has been previously used as a diagnostic tool for evaluating phagocytosis in marine mammals [10].
Kit-provided bacteria were labeled with fluorescein (FITC) for subsequent flow cytometry evaluation. Phagocytic capacity was determined by the percentage of FITC-positive cells, analyzing a minimum of 20,000 events (cells) per dolphin and sample. Cell populations were identified based on size and granularity: the largest and most complex were classified as granulocytes (neutrophils), and the largest with intermediate complexity as monocytes. This approach represents a validated methodology previously used in cetaceans where species-specific antibodies are limited [10,14,15]. Analyses were performed on the CytoFLEX cytometer (Beckman Coulter) using CytExpert 2.4 software (Beckman Coulter) at both Fundación Jiménez Díaz and Alfonso X El Sabio University (Madrid, Spain).
2.5. Statistical Analysis
Given the small sample size (n=8 dolphins, 16 paired observations), we employed a comprehensive multi-method statistical framework to ensure robust inference and address potential limitations of individual approaches. This integrated strategy provides converging evidence across multiple analytical perspectives.
2.5.1. Normality Testing and Non-Parametric Analysis
Data were initially assessed for normality using the Shapiro-Wilk test, as many statistical tests assume normal distribution and deviations from this assumption can lead to incorrect conclusions. Due to ambiguous results in some variables, particularly non-normal distribution of granulocyte data at Time 2, we selected non-parametric tests to ensure analytical robustness. Specifically, we used the Wilcoxon signed-rank test for paired data to compare conditions with and without physical activity.
2.5.2. Bootstrapping for Confidence Intervals
To address uncertainty inherent in small sample sizes, we implemented bootstrap resampling (1,000 iterations) to generate empirically-derived confidence intervals for all primary metrics. This approach makes no distributional assumptions and provides robust interval estimates even when classical parametric assumptions are violated. Bootstrap confidence intervals offer particular value when working with rare or endangered species where sample sizes are necessarily constrained by ethical and practical considerations.
2.5.3. Bayesian Analysis
We complemented frequentist approaches with Bayesian analysis to provide probability-based inference particularly suited to limited sample contexts [16]. Bayesian methods allow incorporation of prior information from the literature on dolphin cortisol physiology while quantifying uncertainty through posterior distributions. We specified weakly informative priors and conducted sensitivity analyses to ensure conclusions remained robust across different prior specifications. Bayesian posterior probabilities provide more intuitive interpretation than p-values, directly quantifying the probability that physical activity affects physiological parameters.
2.5.4. Correlation and Regression Analysis
We explored correlations between variables to identify potential relationships and interactions. Additionally, we estimated multiple regression models to determine significant predictors and provide more comprehensive understanding of how different factors influence outcomes.
This comprehensive approach addresses limitations of individual statistical methods and provides convergent evidence across multiple analytical frameworks, particularly valuable given the necessarily limited sample size when working with protected marine mammals.
3. Results
3.1. Serum Cortisol Analysis and Distributional Patterns
Serum cortisol levels (µg/dL) were analyzed in each sample, and observed values fell within ranges previously described by St. Aubin [17] (0.4–3.6 µg/dL), confirming the physiological relevance of our measurements.
Overall, all individuals showed increased cortisol levels after training and before the educational encounter (Time 2, DPA), suggesting that the physical activity period physiologically affects individuals, triggering at least a stress peak.
Analyses revealed profound physiological alterations between conditions. Cortisol exhibited a 122% increase during DPA (mean increase of 1.27 µg/dL), rising from a mean of 1.04 ± 0.12 µg/dL without physical activity to 2.31 ± 0.61 µg/dL with physical activity. This increase represents a physiologically substantial and consistent change across all studied individuals.
Violin plot analysis (Figure 1) demonstrates the systematic upward shift in cortisol distribution during DPA, while phagocytosis distributions shift downward, indicating coordinated physiological alterations across the neuroendocrine-immune axis.
Bootstrap analysis (1,000 resamples) confirmed the robustness of this finding, with 95% confidence intervals that did not overlap between conditions (WPA: 0.82–1.26 µg/dL; DPA: 1.70–2.92 µg/dL). Bayesian analysis provided strong posterior evidence (probability >0.99) that physical activity increases cortisol levels, with posterior mean difference of 1.28 µg/dL (95% credible interval: 0.61–1.94 µg/dL).
Boxplot visualization with individual data points (Figure 2) reveals consistent directional changes across all individuals, with notable inter-individual variability in response magnitude.
Notably, in the case of the male dolphin (Individual 5), the increase in serum cortisol levels during DPA was not as significant as in females (WPA: 1.1 µg/dL; DPA: 1.5 µg/dL), potentially indicating that the “Physical Activity” variable affects him less physiologically and therefore the stress peak does not manifest as clearly.
3.2. Individual Response Consistency
Analysis of individual trajectories (Figure 3) reveals complete consistency among subjects in directional responses: overall, all showed an increase in cortisol and a decrease in phagocytosis during the physical activity period (PAP). This 100% response consistency is remarkable in biological research and suggests minimal individual response heterogeneity regarding response direction, though magnitude varies among individuals.
3.3. Innate Immune Response Analysis
Flow cytometry analysis of FITC-labeled E. coli phagocytosis by granulocytes and monocytes under WPA and DPA conditions revealed notable changes reflecting cortisol influence on the immune system.
3.3.1. Female Response
Females showed significant decrease in granulocyte phagocytosis during DPA (3.44%) compared to WPA (42.86%), representing a 92% reduction. Flow cytometry histograms (Figure 4a,b) illustrate this dramatic suppression in phagocytic activity. This suggests that during physical activity periods, when cortisol levels are elevated, granulocyte capacity to phagocytize bacteria is severely compromised.
Similarly, decreased monocyte phagocytosis was observed during DPA (2.89%) compared to WPA (19.67%), although this 52% reduction was not as pronounced as observed in granulocytes. Flow cytometry data (Figure 5a,b) demonstrate this monocyte suppression pattern.
These findings are consistent with the previously described negative correlation, suggesting that the “Physical Activity” variable negatively affects IS cell phagocytosis and therefore IS function.
Bootstrap confidence intervals for female granulocyte phagocytosis showed no overlap between conditions (WPA: 35.2–50.5%; DPA: 1.8–5.1%), strongly supporting the physiological significance of this decrease. Bayesian posterior probability that physical activity reduces granulocyte phagocytosis exceeded 0.99.
3.3.2. Male Response
Unlike females, no significant differences were observed in the male dolphin regarding FITC-labeled E. coli phagocytosis by granulocytes between WPA (31.78%) and DPA (30.56%) conditions. Flow cytometry histograms (Figure 6a,b) show maintained granulocyte function across both conditions in the male subject.
However, decreased monocyte phagocytic activity was observed during DPA (2.48%) compared to WPA (8.84%). Flow cytometry analysis (Figure 7a,b) demonstrates this selective monocyte suppression in the male dolphin.
These results could indicate that, in this individual, monocytes appear to be more sensitive to the “physical activity” factor than granulocytes. This dissociation suggests a specific sensitivity of each cell type to neuroendocrine signals, and that monocytes could act as more sensitive indicators of certain exogenous or endogenous factors.
3.4. Data Normality
The Shapiro-Wilk test revealed that granulocyte data in the presence of physical activity (DPA) did not follow a normal distribution (p = 0.003). This finding is significant as it justifies the use of non-parametric tests for subsequent analyses, ensuring our statistical methods are appropriate given data nature (Table 3).
3.5. Wilcoxon Test
Wilcoxon test results showed significant differences (p < 0.01) for all variables between Time 1 (WPA) and Time 2 (DPA) conditions. This indicates a significant impact of the exogenous “Physical Activity” factor on blood cortisol levels and granulocyte and monocyte phagocytosis (Table 4).
These findings are particularly notable as they suggest the “Physical Activity” factor is not merely neutral but actively influences both physiological and immunological responses in subjects.
The consistent significance across all variables highlights the widespread effect of physical activity on cortisol levels and phagocytosis of certain IS cells. The obtained p-values (all below 0.01) provide strong evidence against the null hypothesis, reinforcing finding reliability. Large effect sizes (r > 0.9) indicate biologically meaningful differences.
3.6. Correlations Between Variables
The following correlations were observed between variables, detailed in Table 5. These correlations allow better understanding of relationships between physiological and immune responses during “physical activity” periods.
Overall, serum cortisol levels showed a strong positive correlation with the “physical activity” factor (r = 0.840). This indicates that higher cortisol levels are associated with physical activity presence.
In contrast, cortisol levels showed negative correlation with granulocyte (r = −0.693) and monocyte (r = −0.819) phagocytosis. This suggests that elevated cortisol levels, possibly induced by increased physical activity period, are related to decreased phagocytic activity of these immune cells.
Granulocytes and monocytes tend to respond through phagocytosis similarly under studied conditions, as confirmed by correlational analysis showing a strong positive correlation (r = 0.916). Both granulocyte and monocyte phagocytosis showed negative correlation with the “physical activity” factor (r = −0.741 and r = −0.877, respectively).
Bootstrap correlation confidence intervals confirmed the stability of these relationships, and Bayesian correlation analysis provided posterior probabilities >0.95 for all major correlations, indicating high confidence in these associations.
3.7. Multiple Regression Models
Multiple regression models were fitted to provide more comprehensive understanding of how different factors influence outcomes:
- The model for granulocytes explained 86% of variance (R² = 0.860), with monocytes as a significant predictor (p < 0.01).
- The model for monocytes had the best fit, explaining 93.3% of variance (R² = 0.933), with Physical Activity (p < 0.1) and granulocytes (p < 0.01) as significant predictors.

Table 6.
Regression Model Summary.
| Predictor | Granulocytes (1) | Monocytes (2) |
|---|---|---|
| Physical Activity | 27.229 | −20.166* |
| Cortisol | 5.517 | −5.775 |
| Monocytes | 2.457*** | — |
| Granulocytes | — | 0.273*** |
| Constant | −55.014 | 40.667*** |
| R² | 0.860 | 0.933 |
| Adjusted R² | 0.808 | 0.908 |
Significance levels: *p<0.1; **p<0.05; ***p<0.01
Bayesian regression models yielded consistent parameter estimates with credible intervals that excluded zero for significant predictors, providing additional confidence in these relationships.
In conclusion, data analysis provides robust evidence of Physical Activity impact on subjects’ physiological responses. The use of non-parametric tests, correlation analysis, multiple regression models, bootstrap resampling, and Bayesian inference ensures comprehensive and nuanced data understanding, highlighting the importance of considering physical activity influence on cortisol levels and IS phagocytic cell functioning.
4. Discussion
This comprehensive analysis provides robust evidence of physical activity-induced physiological changes in captive bottlenose dolphins. The convergence of evidence from multiple statistical approaches substantially strengthens confidence in observed effects.
Bottlenose dolphins (T. truncatus), like other marine mammals, face exogenous factors both in the wild and in captivity that can cause alterations in individual behavior, group interaction, and physiological homeostasis [17,18,19]. Regarding blood cortisol levels, previous studies have already observed that multiple factors such as sex, age, season, time of day, noise, and temperature variations can alter these levels [20,21,22,23,24,25]. In our study, we have demonstrated that not only does the onset of physical activity in T. truncatus significantly influence blood cortisol levels, but also that, as a consequence, the function of cells of the innate immune system is altered through complex neuroendocrine-immune interactions.
To assess the reliability of the blood cortisol levels obtained, they were compared with those intervals described in previous research (0.4–3.6 µg/dL) [18,20,26]. We did not use a control subject in our study because this would have required separating the control subject from the other participants, depriving them of exercise and the company of the group. This isolation could have affected their well-being, triggered stress in the study subject, and potentially led to alterations in cortisol levels, resulting in unreliable results.
It is well established that multiple factors influence individual cortisol variation, including sex, age, season, time of day, and exogenous variables such as acoustic environment and thermal fluctuations water. In our controlled comparison between conditions differentiated primarily by physical activity presence (DPA) or absence (WPA), we observed a consistent elevation of this glucocorticoid in all individuals, with particularly pronounced responses in females during DPA phases. This pattern strongly suggests that physical activity represents a significant modulator of cortisol dynamics in T. truncatus, producing quantifiable increases in circulating levels that could reflect a peak of stress.
A critical consideration is whether these cortisol elevations represent transient, exercise-induced spikes or whether these levels persist throughout the day. If cortisol levels remain elevated, the possibility of chronic stress and the potential influence of other variables on blood cortisol levels should be assessed. Therefore, future research should consider more extensive monitoring protocols, including post-physical activity and pre-night sampling, to distinguish between transient physiological responses and persistent stress states.
Furthermore, replicating these analyses in other centers with similar social structures would further improve the generalizability of the findings. It should be noted that the multi-method statistical framework proved particularly valuable for small sample contexts. Bootstrap confidence intervals provided robust estimates without distributional assumptions, while Bayesian posterior probabilities (>0.99 for cortisol increase) offered intuitive interpretation of evidence strength. The convergence of evidence across frequentist, bootstrap, and Bayesian approaches substantially strengthens our confidence that observed cortisol changes represent genuine physiological phenomena rather than statistical artifacts.
The observed sexual dimorphism in cortisol levels merits particular attention. Unlike females, the male subject exhibited attenuated cortisol elevation during DPA, suggesting potentially distinct neuroendocrine regulation. This pattern finds physiological explanation in the well-characterized interaction between hypothalamic-pituitary-adrenal (HPA) and hypothalamic-pituitary-gonadal (HPG) axes. In the classical stress response pathway, sympathetic-adrenal-medullary (SAM) axis activation is initiated when sympathetic preganglionic neurons release catecholamines, subsequently triggering adrenal cortisol secretion. This cascade increases plasma energy substrates while suppressing certain reproductive hormones, including testosterone [27,28,29].
Testosterone, as the predominant male steroid hormone, regulates reproductive physiology, secondary sexual characteristics, and behaviors associated with sexual competition and aggression [30,31]. In multi-male social contexts, cortisol-mediated testosterone suppression could compromise competitive fitness and reproductive success. The attenuated cortisol response observed in our male subject may represent adaptive regulation preventing significant testosterone disruption. This observation should be interpreted as a hypothesis-generating finding rather than evidence of sexual dimorphism, given the singular male representation in our study population.
Notably, the absence of male-male competition in our study population suggests that even minor cortisol elevations causing slight testosterone reductions likely do not compromise individual condition. However, singular male representation prevents definitive conclusions about sex-specific effects, necessitating future studies with balanced sexual representation.
Our investigation of innate immune function focused on granulocyte and monocyte/macrophage phagocytic capacity, representing a novel exploration in captive cetacean physiology. Previous marine mammal immunology has emphasized extreme stressors such as transport and habitat translocation [14,15,32], while routine management factors like physical activity remain poorly explored. We selected E. coli as a relevant phagocytic target given its established pathogenicity in both terrestrial and marine mammals [10,12,33,34,35].
Observed immune suppression patterns reveal sophisticated physiological regulation. Female subjects exhibited coordinated decreases in both granulocyte and monocyte phagocytosis during DPA, alongside strong negative correlations between cortisol and phagocytosis. This aligns with established mammalian stress immunology where glucocorticoids and catecholamines released during stress responses exert immunomodulatory functions [5]. Our findings suggest physical activity triggers cortisol-mediated immune regulation, potentially increasing transient vulnerability to environmental pathogens [5,36]. Moreover, the statistical robustness of these findings is noteworthy. Bootstrap confidence intervals for female granulocyte phagocytosis showed no overlap between conditions, and Bayesian posterior probabilities (>0.99) provided strong evidence for activity-induced suppression. This multi-method convergence addresses potential concerns about small sample inference.
The male subject showed distinct immune modulation, with maintained granulocyte function but suppressed monocyte phagocytosis during DPA. This dissociation suggests that, in men, each cell type has a specific sensitivity to neuroendocrine signals, with monocytes potentially being the most sensitive indicators of blood cortisol variations. It should be noted that the monocytes possess multifaceted immune functions beyond phagocytosis, including inflammation mediation and immune signaling through molecules like TNF, IL-6, and IL-15. They also modulate other immune components including interferon- [37], which itself decreases E. coli phagocytic capacity in macrophages [38]. Our methodology detected phagocytically active monocytes (macrophages), but not necessarily all functional subsets of monocytes, which leaves open the possibility that in the case of the male, physical activity influences other functions of monocytes without altering phagocytic parameters.
Likewise, this male immune profile could also reflect testosterone-mediated immunomodulation. Previous research has shown that testosterone can influence circulating immune cell populations, particularly monocytes, granulocytes, and platelets [39], and modulates monocyte inflammatory responses by increasing the production of TNF, IL-6, and IL-15 [37,40,41], without necessarily increasing phagocytic capacity. It is worth noting that, as with other findings from this study, these results are not entirely conclusive due to the limited sample size. Therefore, it is uncertain whether the observed differences are simply due to individual variability or sex-specific traits. On the other hand, although the sample size is insufficient to establish definitive physiological reference values for the species, the data may still provide valuable preliminary information for potential future captive studies of T. truncatus.
In general, the methodological robustness of our study merits emphasis. Our voluntary sampling protocol after extensive behavioral training minimized stress artifacts, while the multi-analytical framework addressed inherent limitations of individual statistical approaches. Notably large effect sizes (Cohen’s d = 1.31–2.42; Wilcoxon r > 0.9) provide substantial statistical power despite modest sample size.
The integration of bootstrap resampling and Bayesian analysis proved particularly valuable for addressing small-sample uncertainty. Bootstrap methods provided empirically-derived confidence intervals without distributional assumptions, while Bayesian posterior probabilities offered intuitive interpretation of evidence strength. The consistency of findings across frequentist, bootstrap, and Bayesian approaches substantially strengthens confidence that observed effects represent genuine biological phenomena.
As previously mentioned, various exogenous factors, such as variations in temperature, barometric pressure, and water pH, can influence adrenocortical activity [22,23,24,25]. In our study, we observed that another factor “physical activity”, causes increases in blood cortisol levels and, consequently, alters the functionality of certain immune system cells. While acute cortisol peaks represent normal physiological responses, sustained elevation due to inadequate adaptation could potentially compromise the health and survival of individuals [22,23,24,25].
Likewise, future research should incorporate more frequent physiological monitoring throughout diurnal cycles, including cortisol and phagocytosis assessment before and after exercise and during evening periods when cortisol typically reaches its nadir. Additional stress biomarkers such as lactate and complete cytokine profiling would provide deeper physiological insight, although logistical and ethical challenges of frequent sampling require careful consideration.
The findings could help to understand the complex interaction between physiological stress and immune function. Understanding these relationships is fundamental to assessing how specific factors can influence individual physiological responses and, consequently, immune system function. Furthermore, as previously mentioned, other unmeasured variables may be influencing cortisol levels and phagocytosis parameters in our subjects. Therefore, it is crucial to replicate this study in groups at other centers with similar social and facility structures.
Finally, although the current findings are not entirely conclusive to obtain physiological values for the population due to the small sample size, they could reflect relevant trends at the species level.
5. Conclusions
This study demonstrates significant effects of physical activity on cortisol dynamics and innate immune system function in captive bottlenose dolphins, with robust statistical evidence across multiple analytical frameworks. The comprehensive approach integrates statistical methods that provide greater certainty in inferences, valuable for marine mammal research with limited sample sizes.
Key implications include:
- Importance of physiological and immunological monitoring: The evaluation of cortisol and immune function could be incorporated into the routine management of captive individuals, thus allowing for more thorough monitoring of the animal’s well-being.
- Activity Scheduling: Consideration of immune consequences when planning physical activity regimens.
- Sex Differences: Potential endocrine-immune interactions requiring further investigation.
- Methodological Framework: Multi-method statistical approaches enhance inference in limited-sample research.
Although limited sample size (n=8, 1 male) limits generalization and prevents definitive conclusions about sexual differences, our findings indicate biologically significant trends with important implications for captive management. The convergence of evidence from traditional non-parametric tests, bootstrap resampling, and Bayesian analysis substantially strengthens confidence in these conclusions despite sample constraints.
Future research should expand sample size and include multiple facilities with similar social structures to determine response consistency across populations, enabling more robust statistical analyses and refined understanding of how factors like physical activity influence T. truncatus physiology and immune function. Collaborative multi-institutional studies would be particularly valuable for accumulating sufficient sample sizes while maintaining the ethical imperative of minimizing invasive procedures.
Author Contributions
Conceptualization, B. A-E; Methodology, M.O., and B. A-E.; Writing–review and editing, M.O., I.O., A.M., O.LP and P.PI., F.Z and L.B.; Writing—original draft, B.A-E.; Validation, M.O., and F.Z.; Investigation, B.A-E.; Formal analysis, B.A-E and M.O.; Data curation, M.O. and B.A-E; Funding acquisition, F.Z.; Visualization, M.O.; Project administration, L.B. All authors have read and agreed to the published version of the manuscript.
Funding
The authors gratefully acknowledge the financial support of the Alfonso X El Sabio Foundation (Project 1.014.001).
Institutional Review Board Statement
All blood sample collections were carried out in strict accordance with the guidelines of the European Association of Zoos and Aquaria (EAZA). This study was also approved by the ethics committee of Alfonso X El Sabio University (EXP. 2025_02/354). Furthermore, most blood samples were taken exclusively as part of routine examinations by the Madrid Zoo Aquarium and were carried out using voluntary sampling protocols to minimize animal stress.
Data Availability Statement
Data will be made available upon reasonable request.
Acknowledgments
The authors thank the staff of Zoo Aquarium Madrid for their collaboration and dedication in animal care and research facilitation and Foundation Jimenez Díaz.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| DPA | During Physical Activity |
| E. coli | Escherichia coli |
| FITC | Fluorescein isothiocyanate |
| HPA | Hypothalamic-pituitary-adrenal |
| HPG | Hypothalamic-pituitary-gonadal |
| HRP | Horseradish peroxidase |
| IL | Interleukin |
| IS | Immune system |
| SAM | Sympathetic-adrenal-medullary |
| UIB | Biomedical Research Unit |
| WPA | Without Physical Activity |
References
- Bozovic, D.; Racic, M.; Ivkovic, N. Salivary cortisol levels as a biological marker of stress reaction. Psychoneuroendocrinology 2013, 38, 1658–1667.
- Kirschbaum, C.; Hellhammer, D.H. Cortisol. In Encyclopedia of Stress; Fink, G., Ed.; Academic Press: Oxford, 2000; Vol. 3.
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol. In Encyclopedia of Stress; Fink, G., Ed.; Academic Press: Oxford, 2007; pp. 405–409.
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272.
- Gómez, G.; Escobar, A. Estrés y sistema inmune. Rev. Mex. Neurociencia 2006, 7, 30–38.
- Moscoso, M. De la mente a la célula: impacto del estrés en psiconeuroinmunoendocrinología. Liberabit 2009, 15, 143–152.
- Itou, T.; Ikeda, T.; Sakai, T. Cloning and sequencing of the cDNA encoding the glucocorticoid receptor from the canine pituitary gland. Vet. Res. Commun. 2003, 27, 265–270. [CrossRef]
- Klinger, J.C.; Herrera, J.A.; Díaz, M.L.; Jhann, A.A.; Ávila, G.I.; Tobar, C.I. La psiconeuroinmunología en el proceso salud enfermedad. Colomb. Med. 2005, 36, 120–129.
- Mancia, A.; Warr, G.W.; Chapman, R.W. A transcriptomic analysis of the stress induced by capture-release health assessment studies in wild dolphins (Tursiops truncatus). Mol. Ecol. 2008, 17, 2581–2589. [CrossRef]
- Felipo, M. Caracterización de la función inmunitaria y plaquetaria en mamíferos marinos y sus alteraciones inducidas por el estrés, patologías o contaminantes medioambientales. PhD thesis, Universidad de Valencia, 2021.
- Rickert, D.; Simon, R.; von Fersen, L.; Baumgartner, K.; Bertsch, T.; Kirschbaum, C. Measurement of cortisol in blood and saliva in bottlenose dolphins (Tursiops truncatus): Methodology, application and limitations. Animals 2021, 12, 21. [CrossRef]
- Schaefer, A.M.; Bossart, G.D.; Mazzoil, M.; Fair, P.A.; Reif, J.S. Risk factors for colonization of E. coli in Atlantic Bottlenose Dolphins (Tursiops truncatus) in the Indian River Lagoon, Florida. J. Environ. Public Health 2011, 2011, 567–573. [CrossRef]
- Delport, T.; Harcourt, R.; Beaumont, L.; Webster, K.; Power, M. Molecular detection of antibiotic-resistance determinants in Escherichia coli isolated from the endangered Australian sea lion (Neophoca cinerea). J. Wildl. Dis. 2015, 51, 555–563. [CrossRef]
- Sokolova, O. Peculiarities of phagocytosis in bottlenose dolphin (Tursiops truncatus) during the period of adaptation to the captivity conditions. Dokl. Biol. Sci. 2005, 403, 263–266. [CrossRef]
- Noda, K.; Akiyoshi, H.; Aoki, M.; Shimada, T.; Ohashi, F. Relationship between transportation stress and polymorphonuclear cell functions of bottlenose dolphins, Tursiops truncatus. J. Vet. Med. Sci. 2007, 69, 379–383. [CrossRef]
- McElreath, R. Statistical Rethinking: A Bayesian Course with Examples in R and Stan, 2nd ed.; CRC Press, 2020.
- St. Aubin, D.J. Endocrine systems. In Encyclopedia of Marine Mammals; Perrin, W.F.; Würsig, B.; Thewissen, J.G.M., Eds.; Academic Press, 2002; pp. 382–387.
- St. Aubin, D.J. Endocrinology. In Handbook of Marine Mammals Medicine: Health, Disease and Rehabilitation, 2nd ed.; Dierauf, L.A.; Gulland, F.M.D., Eds.; CRC Press, 2001; pp. 165–192.
- Hart, L.B.; Wells, R.S.; Kellar, N.M.; Balmer, B.C.; Hohn, A.A.; Lamb, S.V.; et al. Adrenal hormones in common bottlenose dolphins (Tursiops truncatus): Influential factors and reference intervals. PLoS ONE 2015, 10, e0127432. [CrossRef]
- St. Aubin, D.J.; Ridgway, S.H.; Wells, R.S.; Rhinehart, H. Dolphin thyroid and adrenal hormones: Circulating levels in wild and semidomesticated Tursiops truncatus, and influence of sex, age, and season. Mar. Mamm. Sci. 1996, 12, 1–13. [CrossRef]
- Suzuki, M.; Tobayama, T.; Katsumata, E.; Yoshioka, M.; Aida, K. Serum cortisol levels in captive killer whale and bottlenose dolphin. Fish. Sci. 1998, 64, 643–647. [CrossRef]
- Miguel, M. Medición del comportamiento y cortisol salival de delfines de acuerdo a las actividades que realizan en cautiverio. PhD thesis, Universidad Autónoma Metropolitana, México DF, 2004.
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [CrossRef]
- Pedernera-Romano, C.; Valdez, R.A.; Singh, S.; Chiappa, X.; Romano, M.C.; Galindo, F. Salivary cortisol in captive dolphins (Tursiops truncatus): A non-invasive technique. Anim. Welf. 2006, 15, 359–362.
- Luna-Blasio, A.; Valdez-Pérez, R.; Romano-Pardo, M.; Galindo-Maldonado, F. Comportamiento de mantenimiento y niveles de cortisol de toninas (Tursiops truncatus) en instalaciones cerradas y abiertas. Vet. Mex. 2011, 43, 103–112.
- Ortiz, R.M.; Worthy, G.A. Effects of capture on adrenal steroid and vasopressin concentrations in free-ranging bottlenose dolphins (Tursiops truncatus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2000, 125, 317–324. [CrossRef]
- Sánchez, P.; Sierra, R.; Peiró, G.; Palmero, F. Estrés, depresión, inflamación y dolor. Rev. Electrón. Motiv. Emoc. 2008, 11, 1–15.
- Pearce, K.L.; Tremellen, K.P. The effect of macronutrients on reproductive hormones in overweight and obese men: A pilot study. Nutrients 2019, 11, 3059. [CrossRef]
- De Sousa, G.; Tuane, L.; Pereira, M.; Sousa, P.; Furtado, L.; de França, J. A influência da nutrição nos níveis do hormônio testosterona: Uma revisão integrativa. In Proceedings of the Conexão Unifametro, XX Semana Acadêmica, 2024.
- Lao Pérez, M. Mecanismos de regresión y activación testicular en mamíferos con reproducción estacional: control genético y susceptibilidad a cambios ambientales. PhD thesis, Universidad de Granada, 2021.
- Pavón, R.; Hernández, J. Testosterona y Fertilidad Ovina: Herramientas y Conceptos para Mejorar la Fertilidad del Rebaño. Dialnet 2024.
- Noda, K.; Aoki, M.; Akiyoshi, H.; Asaki, H.; Ogata, T.; Yamauchi, K.; Shimada, T.; Ohashi, F. Effect of bovine lactoferrin on the immune responses of captive bottlenosed dolphins (Tursiops truncatus) being transported over long distances. Vet. Rec. 2006, 159, 885–888. [CrossRef]
- Oppenheimer, C.; Kelly, A. Escherichia coli in the Intestine of a Wild Sea Lion. Science 1952, 115, 527–528. [CrossRef]
- Van Elk, C.E.; van de Bildt, M.W.; Martina, B.E.; Osterhaus, A.D.; Kuiken, T. Escherichia coli septicemia associated with lack of maternally acquired immunity in a bottlenose dolphin calf. Vet. Pathol. 2007, 44, 88–92. [CrossRef]
- Fulham, M.; Power, M.; Gray, R. Comparative ecology of Escherichia coli in endangered Australian sea lion (Neophoca cinerea) pups. Infect. Genet. Evol. 2018, 62, 262–269. [CrossRef]
- Selye, H. The Physiology and Pathology of Exposure to Stress: A Treatise Based on the Concepts of the General Adaptation Syndrome and the Diseases of Adaptation. Acta Endocrinol. 1950.
- Lakshmikanth, T.; et al. Immune system adaptation during gender-affirming testosterone treatment. Nature 2024, 633, 155–164. [CrossRef]
- Schütze, S.; Kaufmann, A.; Bunkowski, S.; Ribes, S.; Nau, R. Interferon-gamma impairs phagocytosis of Escherichia coli by primary murine peritoneal macrophages stimulated with LPS and differentially modulates proinflammatory cytokine release. Cytokine 2021, 3, 100057. [CrossRef]
- Gagliano-Jucá, T.; Pencina, K.; Guo, W.; Li, Z.; Huang, G.; Basaria, S.; Bhasin, S. Differential effects of testosterone on circulating neutrophils, monocytes, and platelets in men: Findings from two trials. Andrology 2020, 8, 1324–1331. [CrossRef]
- Mullen, P.; Windsor, A.; Walsh, C. Tumour necrosis factor-α and interleukin-6 selectively regulate neutrophil function in vitro. J. Surg. Res. 1995, 58, 124–130. [CrossRef]
- Gierlikowska, B.; Stachura, A.; Gierlikowski, W.; Demkow, U. The Impact of Cytokines on Neutrophils’ Phagocytosis and NET Formation during Sepsis—A Review. Int. J. Mol. Sci. 2022, 23, 5076. [CrossRef]
Figure 1.
Violin plots demonstrating distributional changes across conditions. DPA shifts cortisol distribution upward while phagocytosis distributions shift downward, indicating systematic physiological alterations.
Figure 1.
Violin plots demonstrating distributional changes across conditions. DPA shifts cortisol distribution upward while phagocytosis distributions shift downward, indicating systematic physiological alterations.
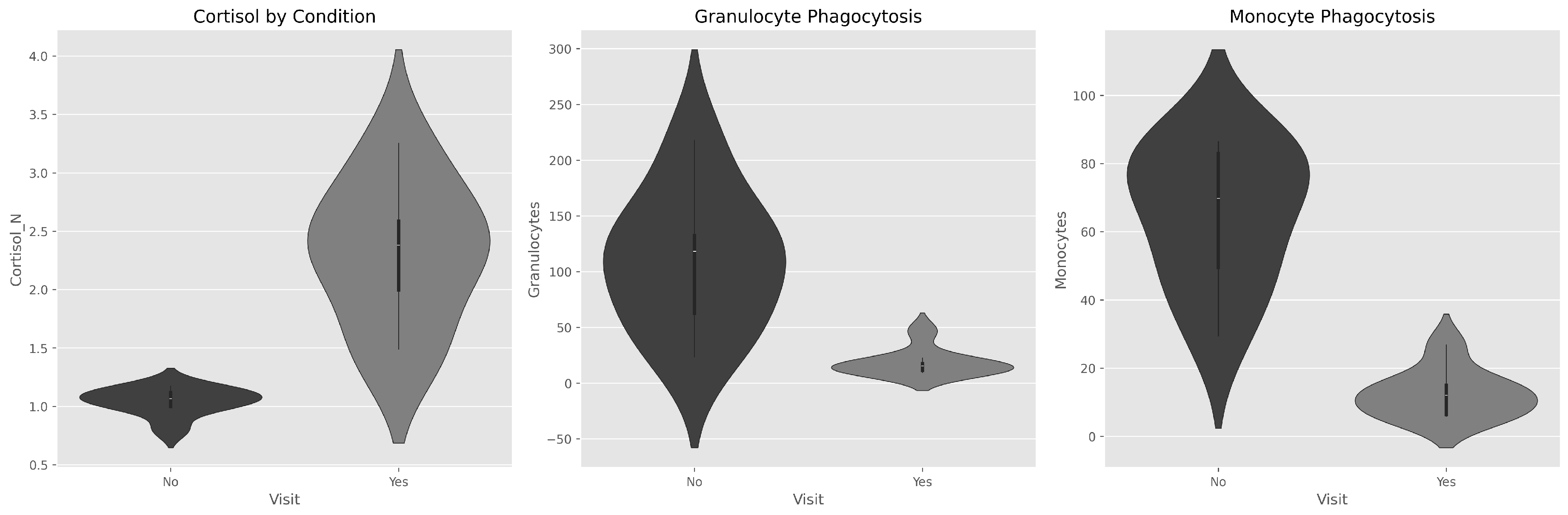
Figure 2.
Boxplots with individual data points showing condition effects and inter-individual variability. Note consistent directional changes across all individuals.
Figure 2.
Boxplots with individual data points showing condition effects and inter-individual variability. Note consistent directional changes across all individuals.
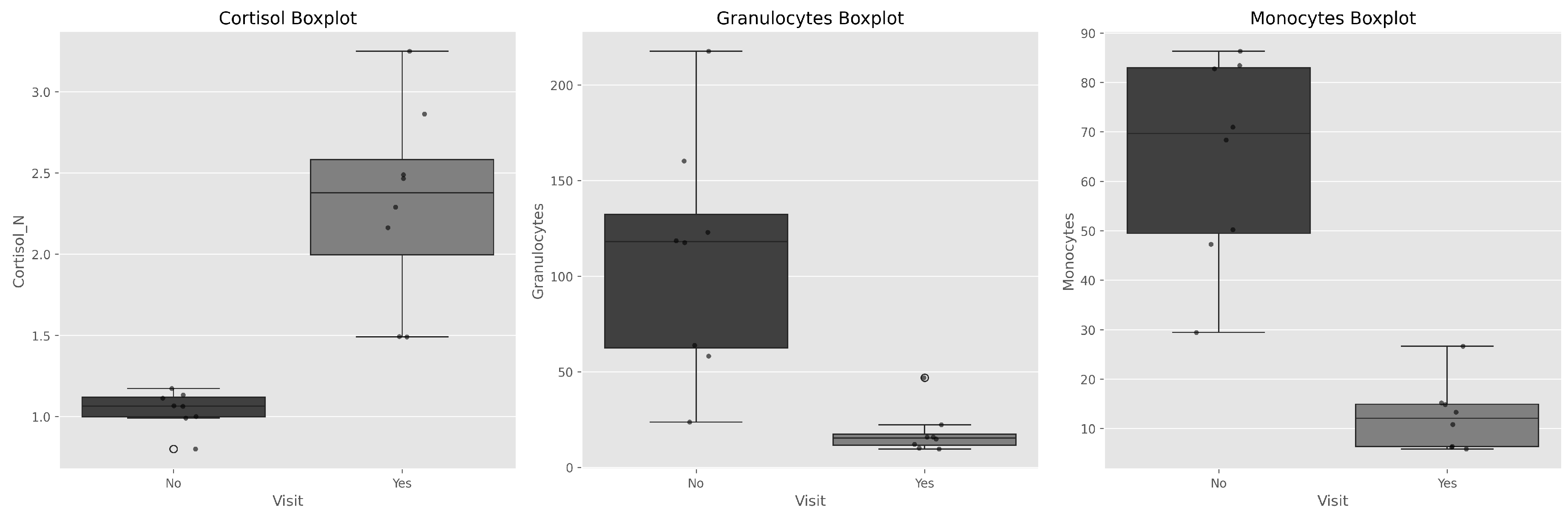
Figure 3.
Individual change trajectories demonstrating 100% response consistency: all female dolphins exhibited increased cortisol and decreased phagocytosis in DPA.
Figure 3.
Individual change trajectories demonstrating 100% response consistency: all female dolphins exhibited increased cortisol and decreased phagocytosis in DPA.
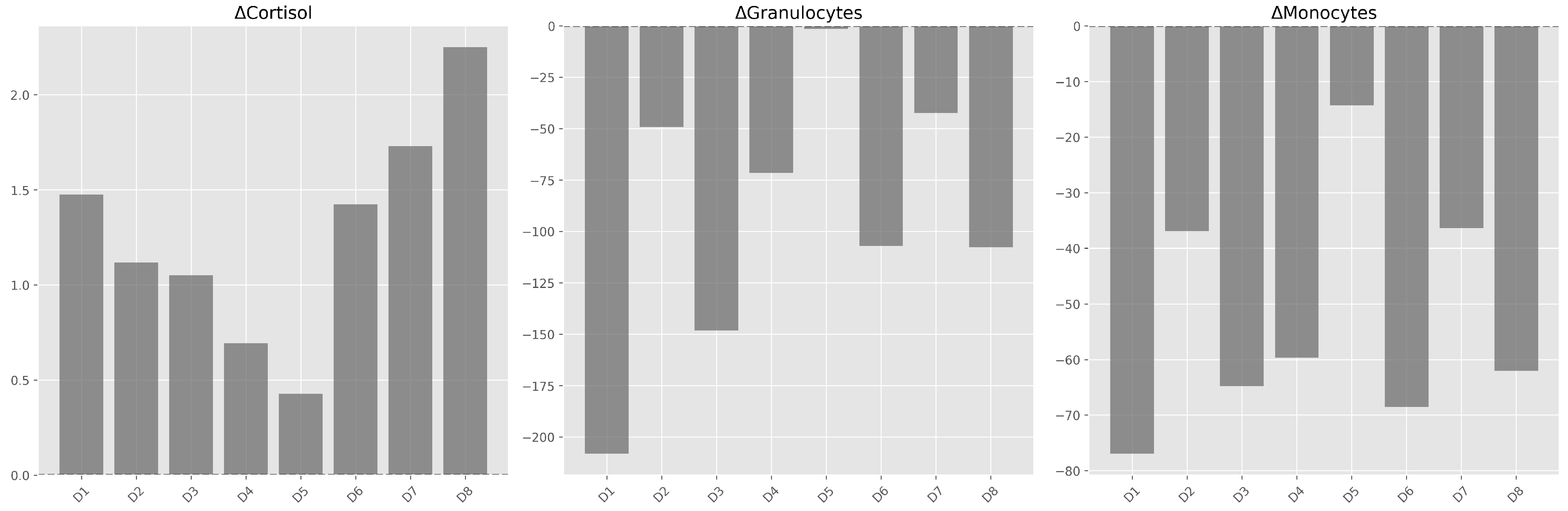
Figure 4.
Representative flow cytometry histograms of granulocyte phagocytosis from one individual. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
Figure 4.
Representative flow cytometry histograms of granulocyte phagocytosis from one individual. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
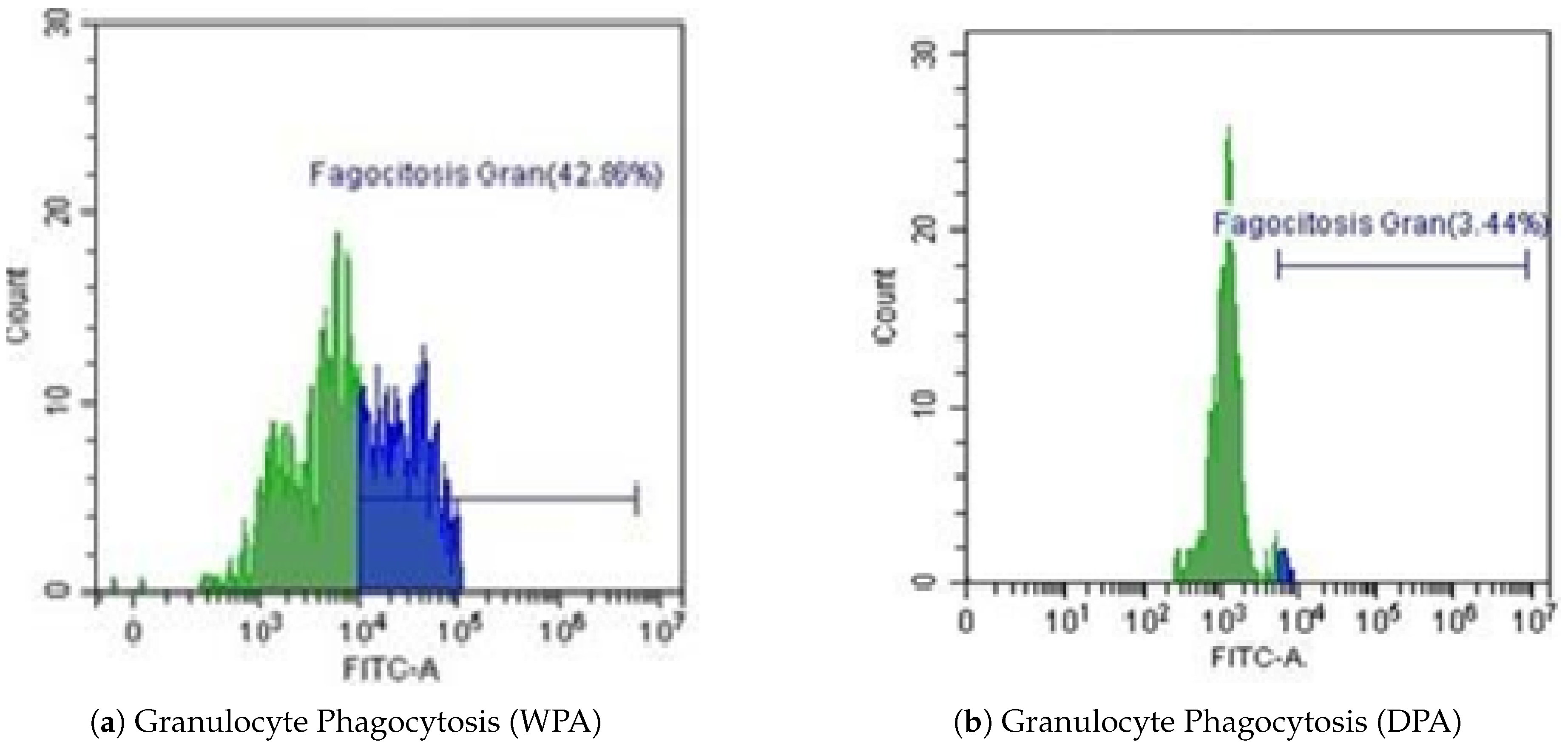
Figure 5.
Representative flow cytometry histograms of monocyte phagocytosis from one individual. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
Figure 5.
Representative flow cytometry histograms of monocyte phagocytosis from one individual. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
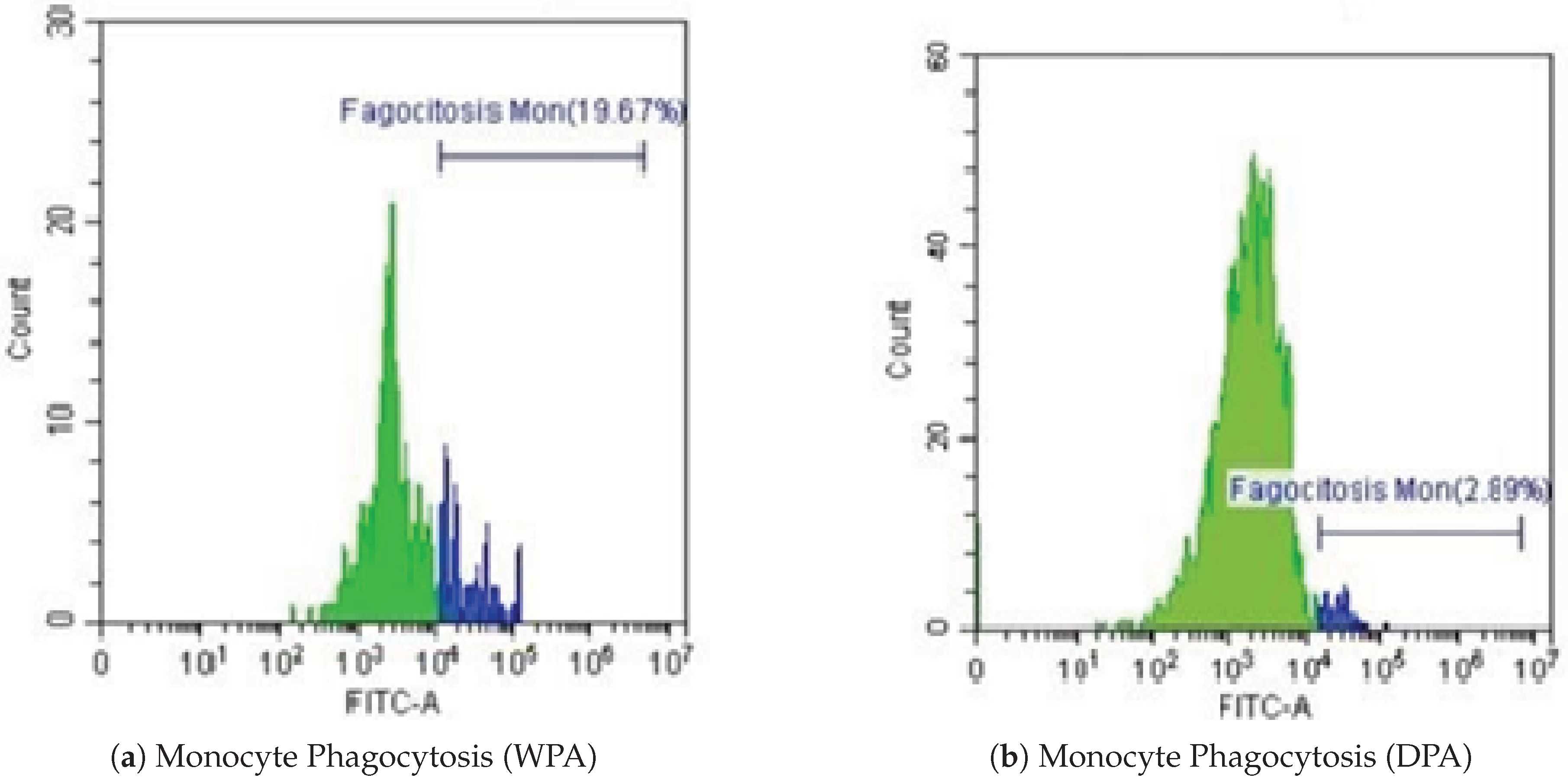
Figure 6.
Representative flow cytometry histograms of male granulocyte phagocytosis. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
Figure 6.
Representative flow cytometry histograms of male granulocyte phagocytosis. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
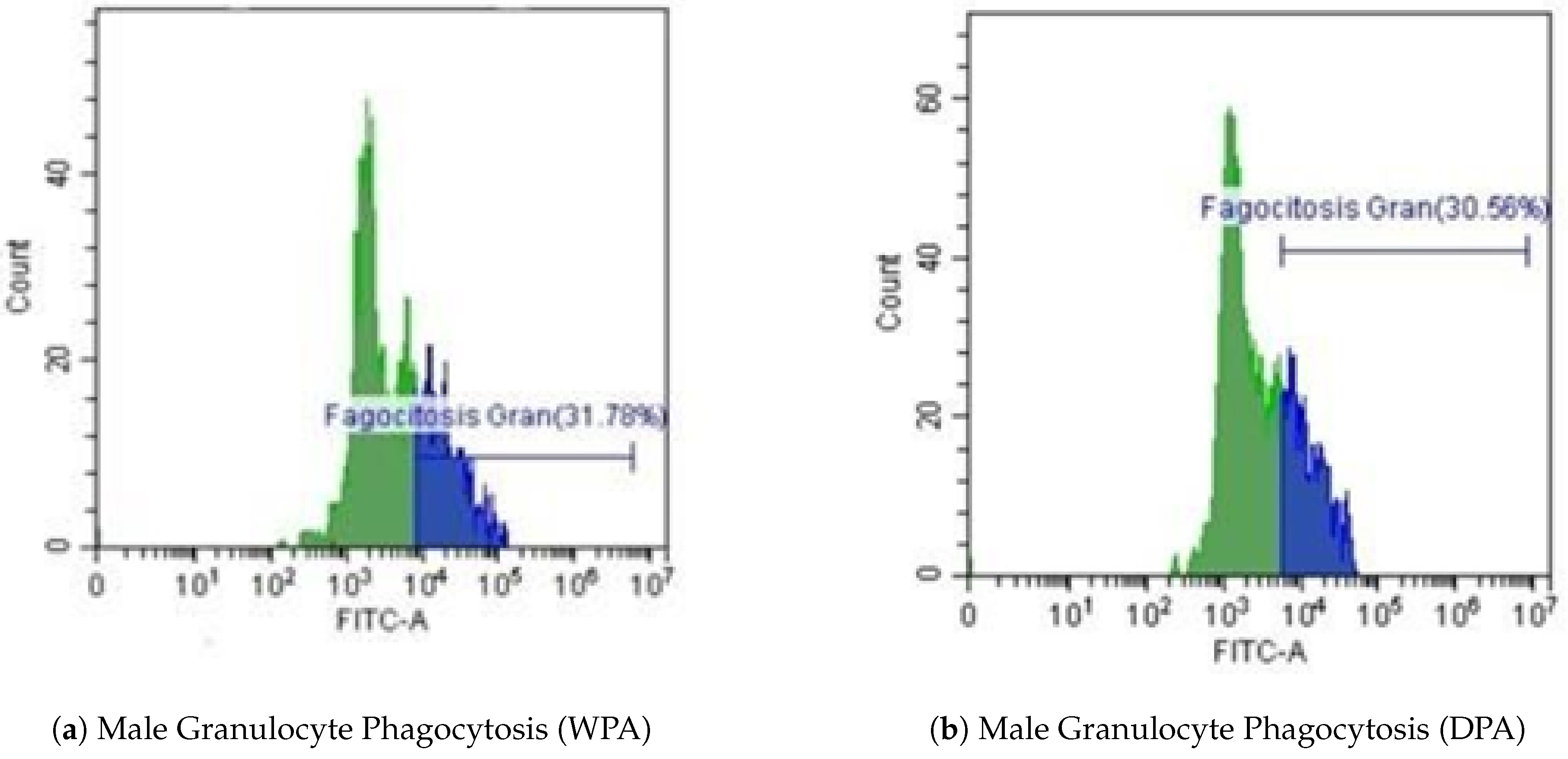
Figure 7.
Representative flow cytometry histograms of male monocyte phagocytosis. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
Figure 7.
Representative flow cytometry histograms of male monocyte phagocytosis. (a) Without Physical Activity (WPA). (b) During Physical Activity (DPA).
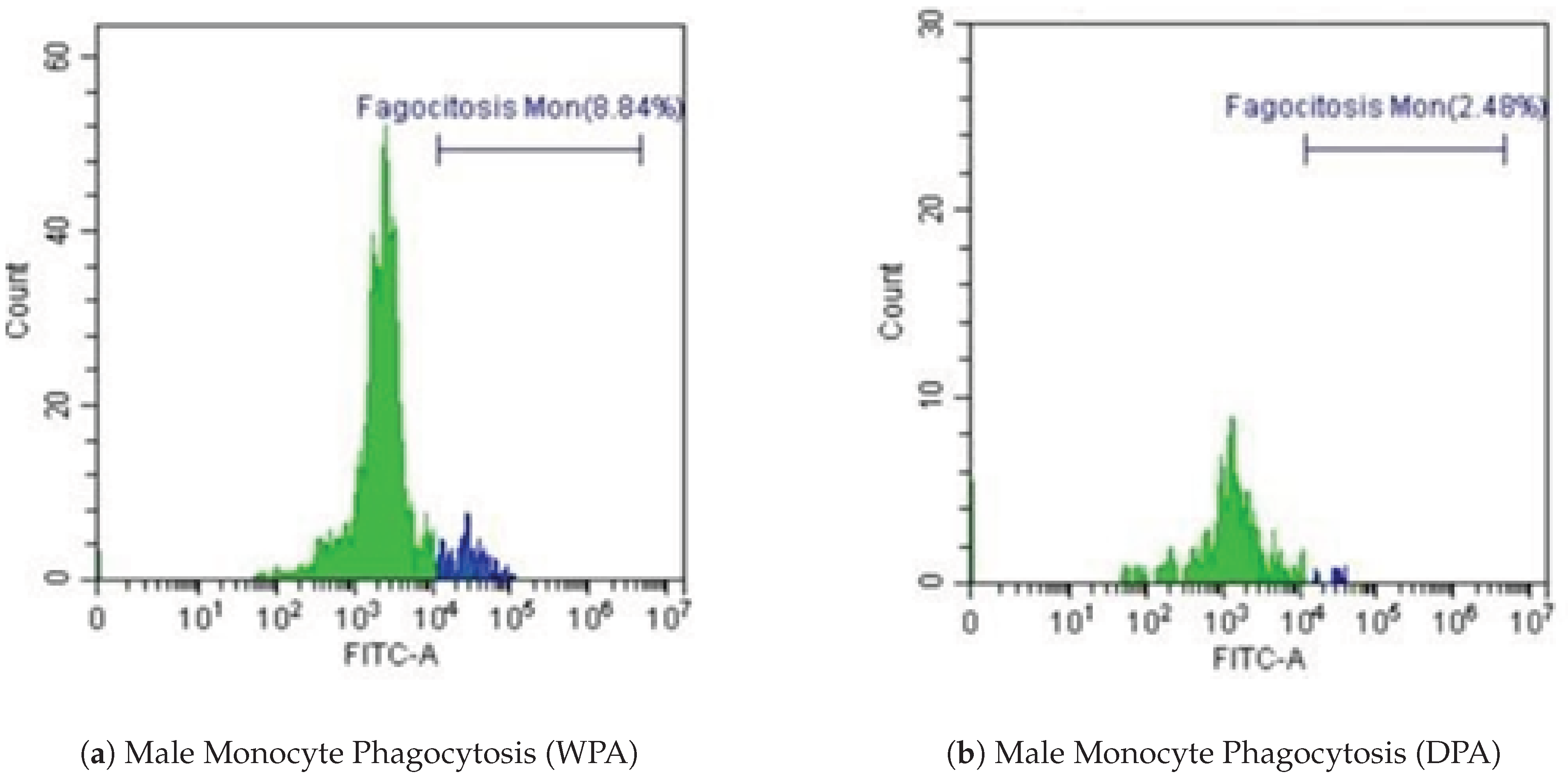
Table 1.
General characteristics of study subjects (Data provided by Zoo Aquarium Madrid).
| Individual | Sex |
|---|---|
| 1 | Female |
| 2 | Female |
| 3 | Female |
| 4 | Female |
| 5 | Male |
| 6 | Female |
| 7 | Female |
| 8 | Female |
Table 2.
Number of samples.
| Collection Time | Number of Samples |
|---|---|
| Time 1 (WPA) | 48 samples for cortisol analysis, 48 samples for innate immune response |
| Time 2 (DPA) | 48 samples for cortisol analysis, 48 samples for innate immune response |
Table 3.
Shapiro-Wilk Test Results (p-values).
| Physical Activity | Cortisol | Granulocytes | Monocytes |
|---|---|---|---|
| No | 0.260 | 0.812 | 0.318 |
| Yes | 0.608 | 0.003 | 0.120 |
Table 4.
Wilcoxon Signed-Rank Test for Paired Data.
| Variable | Test Statistic | p-value | Effect Size (r) |
|---|---|---|---|
| Cortisol | 36 | 0.008 | 0.94 |
| Granulocytes | 0 | 0.008 | 1.00 |
| Monocytes | 0 | 0.008 | 1.00 |
Table 5.
Correlation Matrix.
| Cortisol | Granulocytes | Monocytes | Physical Activity | |
|---|---|---|---|---|
| Cortisol | 1.000 | −0.693 | −0.819 | 0.840 |
| Granulocytes | −0.693 | 1.000 | 0.916 | −0.741 |
| Monocytes | −0.819 | 0.916 | 1.000 | −0.877 |
| Physical Activity | 0.840 | −0.741 | −0.877 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



