
Submitted:
24 December 2025
Posted:
31 December 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Respiratory syncytial virus (RSV) resurged in many regions after the relaxation of stringent non-pharmaceutical interventions (NPIs) implemented during the COVID-19 pandemic. Here, we characterized the epidemiological patterns and molecular evolution of RSV among pediatric inpatients with acute respiratory tract infections (ARTIs) on tropical Hainan Island, China. We retrospectively analyzed 32,329 children (≤18 years) hospitalized at Hainan Women and Children’s Medical Center from January 2021 to December 2024. RSV positivity was determined using targeted next-generation sequencing. In total, 4483/32,329 (13.86%) patients were RSV-positive, with a high positivity in 2021 (20.27%, 957/4721), marked suppression in 2022 (2.03%, 106/5227) during intensive NPIs, and a rebound in 2023–2024 (15.31%, 1490/9732; 15.26%, 1930/12,649). RSV positivity was higher in boys than girls (14.42% vs. 13.00%). Seasonality shifted from a summer–autumn peak in 2021 to a spring–summer predominance in 2023–2024. Among 56 sequenced RSV-positive specimens (29 RSV-A; 27 RSV-B), all RSV-A strains belonged to genotype ON1 (lineages A.D.3 and A.D.5.2), and all RSV-B strains belonged to genotype BA9 (lineages B.D.4.1.1, B.D.E.1, and B.D.E.2). Subtype dominance transitioned from RSV-A (2021–2023; mainly A.D.3) to RSV-B in 2024 (all B.D.E.1). Lineage-specific amino-acid and predicted N-glycosylation changes were observed, including loss of the N179 site in A.D.5.2 and acquisition of N258 in B.D.E.1. These findings indicate that RSV circulation on tropical Hainan was strongly suppressed during intensive NPIs and re-established after policy relaxation, accompanied by earlier seasonal activity and clear lineage replacement, underscoring the need for sustained genomic surveillance to inform locally tailored clinical preparedness and immunization strategies.
Keywords:
respiratory syncytial virus (RSV)
; pediatric acute respiratory tract infections
; seasonality shift
; non-pharmaceutical interventions (NPIs)
; post–Zero-COVID period
; molecular epidemiology
; lineage replacement
; G glycoprotein glycosylation
1. Introduction
Human respiratory syncytial virus (RSV) is the most commonly identified viral pathogen in pediatric acute lower respiratory infections (ALRIs), with an estimated annual incidence exceeding 33 million cases worldwide, resulting in over 3 million hospitalizations and nearly 100,000 deaths in children under five [1]. Despite the recent availability of long-acting monoclonal antibodies and maternal vaccines, RSV continues to impose a heavy burden on health-care systems, particularly in low- and middle-income countries where access to preventive interventions remains limited [2].
RSV is classified into two major antigenic and genetic groups, RSV-A and RSV-B, based on variability in the attachment (G) glycoprotein gene [3]. Within each group, multiple genotypes and lineages have been recognized, many of which are characterised by a partial duplication in the second hypervariable region of the G gene—designated ON1 for RSV-A and BA for RSV-B [4]. These duplications, together with extensive O- and N-linked glycosylation, shape the antigenic surface of the G protein and may facilitate immune evasion. Structural studies have shown that antibodies targeting the central conserved region of G can mediate broad neutralisation, underscoring the need for continued molecular surveillance of circulating strains.
Before the COVID-19 pandemic, RSV epidemics in temperate regions typically peaked in winter. In contrast, in tropical and subtropical areas RSV usually circulated year-round with one or two broad peaks, often coinciding with the rainy or humid season rather than a distinct winter period. In many tropical settings, including southern China and Southeast Asia, RSV activity increases during warm, rainy months and exhibits less pronounced off-season troughs than in temperate climates. These geographical differences in seasonality are critical for the design and timing of immunoprophylaxis and other RSV control strategies.
The COVID-19 pandemic and associated non-pharmaceutical interventions (NPIs)—such as masking, school closures, travel restrictions and lockdowns—profoundly altered the circulation of seasonal respiratory viruses. Many countries reported an almost complete disappearance of RSV and influenza in 2020–2021, followed by atypical, often off-season resurgences after relaxation of NPIs [5,6,7,8,9]. In China, stringent containment policies and the Dynamic Zero-COVID strategy were maintained until 7 December 2022, when nationwide restrictions were abruptly lifted [10]. Subsequent reports from Beijing and other regions described intense RSV epidemics with altered seasonality and age distribution, as well as the emergence of novel lineages such as A.D.5.2 and B.D.E.1 that contributed to large post–zero-COVID RSV waves [11]. However, most available data originate from temperate, highly urbanised settings, and the impact of NPIs and policy changes on RSV dynamics in tropical environments remains poorly defined.
Hainan Island, located at the southernmost tip of China, is the country’s only tropical island province. Its warm, humid climate, distinct monsoon pattern and role as a free-trade-port tourism hub create unique conditions for respiratory virus transmission. Hainan experienced relatively limited but clearly demarcated waves of COVID-19, accompanied by stringent, time-bound NPIs and a synchronised withdrawal of the Dynamic Zero-COVID policy with the rest of mainland China. This combination of tropical climate, geographic semi-isolation and well-documented policy shifts provides a natural setting in which to examine how NPIs and their relaxation influenced RSV transmission dynamics and viral evolution in a paediatric population. Our previous work on influenza virus and rhinovirus among children on Hainan has shown that both viruses exhibited altered seasonal patterns and genetic diversity before and after the Dynamic Zero-COVID period, yet comparable data for RSV are lacking despite its central role in paediatric respiratory disease [12,13,14].
In this study, we conducted a retrospective analysis of paediatric inpatients with acute respiratory tract infections (ARTIs) at Hainan Women and Children’s Medical Center from January 2021 to December 2024. Using targeted next-generation sequencing (tNGS) for RSV detection and G-gene sequencing for lineage, phylogenetic and amino-acid analyses, we aimed to: (i) describe temporal trends and age distribution of RSV infection before and after the end of the Dynamic Zero-COVID policy; (ii) characterise shifts in RSV seasonality and subtype predominance; and (iii) investigate molecular lineage turnover and lineage-specific glycosylation patterns of circulating RSV-A and RSV-B strains on tropical Hainan Island. By integrating epidemiological and genomic data across a period of intensive NPIs and their withdrawal, this study seeks to inform local RSV prevention and clinical management and to enhance understanding of how major public-health interventions reshape RSV transmission and evolution in tropical settings.
2. Materials and Methods
2.1. Sample Collection and RSV Detection
This hospital-based retrospective surveillance included 32,329 pediatric inpatients (≤18 years old) with ARTIs admitted to Hainan Women and Children’s Medical Center between January 2021 and December 2024. Eligible children presented at admission with clinical manifestations consistent with ARTIs, such as fever, headache, rhinorrhoea, tachypnoea, cough, sore throat, sputum production, chest pain, or radiological evidence of pneumonia or bronchitis. Nasopharyngeal swabs were obtained from each patient on admission according to standard clinical procedures and were immediately placed in viral transport medium. Specimens were stored at −80 °C until further analysis. RSV infection was detected using a targeted next-generation sequencing (tNGS) respiratory panel (KingMed Diagnostics, Guangzhou, China). The panel targets conserved genomic regions of common respiratory pathogens; sequencing data were processed through a dedicated bioinformatics pipeline for quality control, alignment to reference databases, read counting and normalization. RSV positivity was determined according to the manufacturer’s recommended reporting thresholds. The study protocol was reviewed and approved by the Institutional Review Board of Hainan Women and Children’s Medical Center.
2.2. RSV Classification Using G Gene Sequences
To characterize RSV lineages, 56 RSV-positive samples representing different time points across the study period were selected for G-gene sequencing. Viral RNA was extracted from 200 μL of each specimen using the QIAamp Viral RNA Mini Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s protocol. Partial G-gene fragments encompassing the second hypervariable region were amplified by nested reverse-transcription PCR (RT-PCR) following the method described by Agoti et al [15]. The first-round RT-PCR was performed using the SuperScript™ III One-Step RT-PCR System (Invitrogen, USA). Cycling conditions were 94 °C for 5 min; 40 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 5 min; and a final extension at 72 °C for 10 min. The second-round PCR was carried out with GoTaq™ Colorless Master Mix (Promega, USA) using the same conditions except for an annealing temperature of 56 °C. A negative control was included in each run. PCR products from positive reactions were purified and subjected to Sanger sequencing on an ABI Prism 3730XL DNA Analyzer (Sangon Biotech, Guangzhou, China). Raw sequences were assembled and edited using SeqMan (DNASTAR, Madison, WI, USA), and viral identity was confirmed by BLAST searches against the National Center for Biotechnology Information (NCBI) database.
2.3. Phylogenetic Analysis
Nextclade (version 3.3.1; https://clades.nextstrain.org/) was used to assign RSV lineages and to identify amino-acid substitutions. All amino-acid changes were reported relative to the default reference sequences (RSV-A: hRSV/A/England/397/2017, EPI_ISL_412866; RSV-B: hRSV/B/Australia/VIC-RCH056/2019, EPI_ISL_1653999). Phylogenetic trees were constructed in MEGA X using the neighbor-joining method, with branch support evaluated by 1,000 bootstrap replicates. Genetic diversity, pairwise sequence identity, and amino-acid variation were analyzed using BioEdit software. All sequences generated in this study were deposited in GenBank under accession numbers OQ248592–OQ248607, OR140542–OR140554, and PV800154–PV800198.
2.4. Statistical Analysis
All statistical analyses were performed using IBM SPSS Statistics version 26.0 (IBM Corp., Armonk, NY, USA). Categorical variables were compared using the χ² test. Two-tailed P values <0.05 were considered statistically significant. Graphs were prepared using GraphPad Prism version 6.0 (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. Sample Information and Demographics
From January 2021 to December 2024, 32,329 children with ARTIs were hospitalized at Hainan Women and Children’s Medical Center and included in the analysis. Patient ages ranged from 1 month to 18 years (mean ± SD, 2.97 ± 2.86 years). Boys accounted for 19,633 (60.73%) admissions and girls for 12,696 (39.27%), indicating a male predominance in the study population.
3.2. Analysis of RSV Infection Trends Before and After the Ending of Dynamic Zero-COVID Policy
Over the four-year period, 4,483 of 32,329 pediatric inpatients with ARTIs tested positive for RSV, yielding an overall positivity rate of 13.86% (Table 1). In 2021, 957 of 4,721 ARTIs inpatients were RSV-positive (20.27%). In 2022, although the number of ARTIs hospitalizations increased slightly to 5,227, only 106 patients were RSV-positive, and the annual positivity rate dropped sharply to 2.03%. This sharp decline coincided with two local COVID-19 outbreaks in March and August 2022 and the implementation of stringent non-pharmaceutical interventions. Following relaxation of the Dynamic Zero-COVID policy in December 2022, RSV circulation rebounded in 2023, with 1,490 of 9,732 ARTIs inpatients were positive (15.31%). The infection rate remained high in 2024, with 1,930 of 12,649 ARTIs inpatients were positive (15.26%), reaching levels comparable to those observed in 2021 and indicating re-establishment of endemic transmission. The distribution of RSV positivity differed significantly by year (χ² = 812.97, p < 0.001), indicating a profound temporal impact of COVID-19 control measures on RSV activity (Table 1, Figure 1).
RSV prevalence also differed by sex. Overall, the positivity rate among ARTIs inpatients was higher in boys than in girls (14.42% [2,832/19,633] vs 13.00% [1,651/12,696]; χ² = 13.03, p < 0.001). This male predominance persisted from 2022 to 2024, with significantly higher RSV positivity in boys than in girls in each year (Table 1).
Marked seasonal variation in RSV positivity was observed (Table 2). In 2021, detection rates rose from 5.85% in spring to 37.54% in summer, then declined to 29.20% in autumn and 1.61% in winter (χ² = 636.76, p < 0.001). In 2022, RSV circulation was strongly suppressed throughout the year, with positivity rates of 0.93% in spring, 0.07% in summer, 1.57% in autumn and 5.61% in winter (χ² = 126.29, p < 0.001). By 2023, the seasonal pattern had shifted: positivity peaked in spring (24.14%) and summer (29.09%), then decreased in autumn (7.76%) and winter (1.07%) (χ² = 917.55, p < 0.001). A similar spring–summer predominance persisted in 2024, with rates of 23.56%, 21.64%, 11.77% and 3.51% in spring, summer, autumn and winter, respectively (χ² = 721.83, p < 0.001). Together with the monthly curves in Figure 1, these findings indicate a shift in RSV seasonality on Hainan Island from a summer–autumn peak before 2022 to a spring–summer predominance after the end of the Dynamic Zero-COVID policy. (Table 2, Figure 1).
3.3. Age-Based Analysis of RSV Infections
RSV-positive patients were categorized into four age groups commonly used in pediatric epidemiology in China: 0–1 years (Group 1), 1–3 years (Group 2), 3–7 years (Group 3), and 7–18 years (Group 4). Between 2021 and 2024, the highest RSV prevalence was observed in infants 0–1 years old, with a positivity rate of 22.55% (2,228/9,882). This was followed by children 1–3 years (17.86%; 1,438/8,051). RSV prevalence was lower in older age groups, at 7.06% (728/10,314) in children 3–7 years and 2.18% (89/4,082) in those 7–18 years (Table 3). Differences among age groups were statistically significant (χ² = 1,597.85, p < 0.001).
As illustrated in Figure 2, RSV infections were concentrated in younger children across all four years. Both the absolute number of RSV-positive cases and the infection rate showed a clear decreasing gradient with increasing age, with the majority of infections occurring in children under 3 years of age. These data confirm that infants and toddlers bear the greatest burden of RSV infection and are likely key drivers of RSV transmission in this tropical setting.
3.4. Phylogenetic Analysis of RSV
Partial G-gene sequences were successfully obtained from 56 RSV-positive specimens, including 29 RSV-A and 27 RSV-B strains. All RSV-A sequences belonged to the ON1 genotype and clustered within A.D lineages; specifically, they grouped into A.D.3 and A.D.5.2 (both GA2.3.5-related). All RSV-B sequences belonged to the BA9 genotype and were assigned to B.D.4.1.1, B.D.E.1 or B.D.E.2 lineages (GB5.0.5a-related) (Figure 3 A and B).
From 2021 to 2023, RSV-A predominated in Hainan, with an overall RSV-A : RSV-B ratio of approximately 3:1 and A.D.3 representing the most frequently detected lineage. In 2024, a marked shift in subtype predominance was observed: RSV-B became dominant, with an RSV-A : RSV-B ratio of approximately 1:3.8. During the 2021–2023 epidemic seasons, A.D.3 was the main RSV-A lineage, whereas in 2024 A.D.5.2 gradually replaced A.D.3. For RSV-B, only a single B.D.4.1.1 strain was detected in 2021–2022. In 2023, RSV-B sequences were distributed among three lineages, with B.D.4.1.1 being predominant and B.D.E.1 and B.D.E.2 also present. By 2024, all sequenced RSV-B strains belonged to the B.D.E.1 lineage, indicating complete turnover of RSV-B from B.D.4.1.1 to B.D.E.1 (Figure 3 A and B).

3.5. Amino Acid Analysis
To characterize lineage-specific amino-acid changes, we compared the deduced G-protein sequences of RSV strains from Hainan with prototype ON1 (JN257693) and BA9 (AY333364) strains. The analysed regions encompassed the second hypervariable region (HVR2), including the 23-amino-acid duplication in RSV-A and the 20-amino-acid duplication in RSV-B (Figure 4A,B).
Among RSV-A strains, all A.D.3 sequences shared H258Q and H266L substitutions. Other frequent substitutions included L274P (10/18), L289I (7/18), L298P (6/18), E262K (6/18), Y305H (5/18), T239A (4/18) and T319K (3/18). In contrast, all A.D.5.2 strains exhibited a highly conserved pattern of concurrent mutations at L248I, L274P, S283P, L298P, V303A, Y304H, L310I and T320A (Figure 4A).
Predicted N-linked glycosylation also differed by RSV-A lineage. Relative to the ON1 prototype, nearly all A.D.3 strains were predicted to harbour five N-glycosylation sites (N85, N103, N135, N179 and N237). The N179 site likely arose through an N178G substitution that created a novel N-X-S/T motif. In addition, an S100N substitution detected in five strains collected between July and August 2023 introduced an extra predicted N-glycosylation site at N100. In contrast, almost all A.D.5.2 strains retained four N-glycosylation sites (N85, N103, N135 and N237) but lacked the N179 site observed in A.D.3 strains (Table 4).
For RSV-B, all B.D.4.1.1 strains carried a shared set of substitutions—K218T, L223P, S247P, T254I, T270I, V271A, H287Y, T290I, N296Y and T312I—with additional frequent changes at I286T (4/5, 82%) and T302I (4/5, 82%) (Figure 4B). All B.D.E.1 strains showed a distinct, relatively uniform mutation profile involving P216S, K218T, S247P, K258N, S277P, I281T, T290I and T312I, with common additional substitutions at V271A (20/21), S267P (16/21) and I229T (4/21).
Predicted N-glycosylation patterns of RSV-B also differed by lineage. B.D.4.1.1 strains had a single N-glycosylation site at N230; the combination of N296Y and T312I abolished predicted glycosylation sites in the C-terminal region of the G protein. In contrast, B.D.E.1 strains were predicted to contain three N-linked glycosylation sites at N230, N258 and N296. The K258N substitution, present in all B.D.E.1 sequences, created a novel N258 site. None of the B.D.E.1 strains retained the N310 site, likely due to T312I disrupting the N-X-S/T consensus motif required for N-linked glycosylation (Table 5).
4. Discussion
This four-year study from tropical Hainan Island shows that RSV epidemiology and viral population structure in hospitalized children were profoundly reshaped across the implementation and withdrawal of China’s Dynamic Zero-COVID policy. RSV activity was markedly suppressed in 2022, then rapidly re-established in 2023–2024 to near-pre-pandemic levels, accompanied by an advance of epidemic timing from a summer–autumn peak to spring–summer predominance and by clear lineage turnover within both RSV-A and RSV-B. Together, these findings highlight how non-pharmaceutical interventions (NPIs), population immunity, and viral evolution jointly determine RSV transmission dynamics, even in a tropical climate.
The abrupt reduction of RSV positivity in 2022 is temporally consistent with intensive NPIs implemented during two local COVID-19 waves, supporting the interpretation that decreased contact rates and mobility can interrupt RSV circulation. Following nationwide relaxation of NPIs in late 2022, RSV rebounded quickly and peaked earlier in the year. Similar post-NPI “off-season” or advanced RSV epidemics have been widely documented, and are most commonly explained by (i) immunity debt, whereby reduced exposure during NPI periods increases the pool of susceptible infants and toddlers, and (ii) ecological interactions among respiratory viruses, including both interference (e.g., interferon-mediated) and facilitation related to co-circulation and altered host susceptibility [16,17].
This pattern is also consistent with broader respiratory virus re-organization in Hainan observed in our team’s prior studies: influenza virus, rhinovirus and Human parainfluenza virus showed pronounced NPI-related perturbations and altered seasonal timing after relaxation, whereas rhinovirus circulation was comparatively resilient [12,13,14]. Such differences underscore virus-specific sensitivity to behavioural interventions and the likelihood that RSV resurgence reflects both restored contact networks and a widened susceptible population.
Despite major temporal shifts, the demographic signature of RSV in Hainan remained typical. Infants <1 year carried the highest burden, followed by children 1–3 years, with a steep decline in older age groups. This age gradient aligns with global evidence that RSV hospitalization burden is concentrated in the first two years of life, plausibly driven by immature immunity, smaller airway caliber, and limited prior exposure [2,18].
RSV positivity was consistently higher in boys than girls in most years. Male predominance has been repeatedly reported in RSV cohorts and likely reflects a combination of biological susceptibility and behavioural or care-seeking factors [19,20]. These stable demographic patterns, observed across periods of suppression and resurgence, reinforce the need to prioritize very young children—particularly infants—for prevention strategies as RSV immunization tools become more widely implemented.
All sequenced RSV-A strains belonged to genotype ON1 and all RSV-B strains to BA9, matching their long-standing global predominance [20,21]. Within these genotypes, however, we observed clear lineage replacement. RSV-A predominated during 2021–2023, mainly as A.D.3, while RSV-B became dominant in 2024, driven almost entirely by B.D.E.1. Concurrently, A.D.5.2 increased and replaced A.D.3 as the major RSV-A lineage by 2024, and B.D.E.1 replaced B.D.4.1.1 within RSV-B. This sequence of events is consistent with recent reports from other parts of China indicating that A.D.5.2 and B.D.E.1 expanded rapidly after relaxation of travel and control measures [11,22].
Mechanistically, several non-exclusive processes may explain the observed turnover: founder effects linked to re-introduction after prolonged suppression; altered immune landscapes created by “gaps” in exposure during NPIs; and lineage-specific fitness advantages related to transmissibility, immune escape, or attachment [13,23]. RSV evolution is characterized by strong purifying selection across most of the genome but repeated adaptive change in surface glycoproteins—particularly G—making lineage replacement a plausible outcome when susceptible pools expand and introductions resume.
Lineage-specific amino-acid substitutions and predicted N-glycosylation patterns in the G protein provide a molecular correlate to these epidemiological shifts. In RSV-A, A.D.3 strains shared conserved H258Q and H266L substitutions plus additional variable changes (e.g., E262K, L274P, L298P, Y305H), consistent with ongoing intra-lineage diversification [4,24]. A.D.5.2 strains exhibited a more stereotyped substitution set (L248I, L274P, S283P, L298P, V303A, Y304H, L310I, T320A), suggesting a comparatively conserved profile. Predicted glycosylation differed meaningfully between these RSV-A lineages. Most A.D.3 strains carried N-linked sites at N85, N103, N135, N179, and N237, with a subset gaining N100 (consistent with S100N). In contrast, A.D.5.2 strains lacked N179 while retaining N85, N103, N135, and N237. Because N179 lies within the cysteine-noose region embedded in broadly neutralizing epitope landscapes, its absence in A.D.5.2 could alter epitope accessibility and antibody recognition [25,26]. Although our analysis is predictive, these patterns support the hypothesis that A.D.5.2 may differ antigenically from A.D.3 in ways that could influence population-level susceptibility.
In RSV-B, B.D.4.1.1 strains carried a characteristic panel of substitutions (including N296Y and T312I) and were predicted to retain N230 as the dominant C-terminal glycosylation site, consistent with loss of other C-terminal sites [21,27,28]. The emergent B.D.E.1 lineage—predominant in 2024—was characterized by changes including P216S and K258N, with predicted glycosylation typically at N230, N258, and N296. K258N introduces a novel N258 site, while T312I disrupts the N310 site. We also observed minor heterogeneity within B.D.E.1 (a small subset lacking N296 and/or N258), indicating ongoing fine-scale diversification even within a rapidly expanding lineage.
Collectively, these substitutions and glycosylation changes imply re-configuration of the G-protein glycan shield, a plausible mechanism for immune evasion and improved fitness in partially immune populations [18]. Functional studies—confirming glycosylation, mapping antigenicity, and assessing binding/neutralization—will be needed to establish causal links between these molecular signatures and transmission success.
The spring–summer predominance observed in 2023–2024 suggests that the optimal timing of future RSV prophylaxis on Hainan may differ from pre-pandemic patterns and from schedules derived in temperate climates. As long-acting monoclonal antibodies and maternal vaccination strategies become available, locally calibrated seasonality data will be important to maximize effectiveness. Although current leading interventions primarily target the prefusion F protein (rather than G), continued genomic surveillance remains essential. G-protein evolution can shape transmissibility and antigenic context, and expanding immunization may introduce new selection pressures that are best captured by integrated whole-genome surveillance and linkage to clinical outcomes.
This study has several limitations. It was conducted at a single tertiary center and included only hospitalized children, potentially under-representing mild/community infections and biasing toward severe disease. The number of sequenced specimens was limited, and analysis focused on partial G-gene regions rather than whole genomes, restricting resolution of genome-wide evolutionary dynamics and limiting assessment of genotype–severity associations. Future work should expand multicenter sampling across Hainan, include outpatient/community cohorts, increase sequencing density with whole-genome approaches, and integrate clinical severity metrics, co-infection data, and (ideally) longitudinal serology to disentangle the contributions of immunity debt, viral interactions, and viral evolution.
In summary, RSV circulation on tropical Hainan rebounded rapidly after NPI relaxation with earlier seasonal peaks, a switch from RSV-A to RSV-B predominance, and lineage replacement within ON1 and BA9 genotypes. Distinct lineage-specific amino-acid and glycosylation signatures in the G protein suggest ongoing antigenic remodeling, underscoring the value of sustained genomic and epidemiological surveillance to inform timing and targeting of RSV immunization strategies in tropical regions.
Author Contributions
Conceptualisation, F.Y. and Y.G.; methodology, M.C., M.X. and Y.C.; experimental operations, Y.J., S.C. and S.W.; data collection and analysis, R.P., Y.H. and G.W.; visualization, S.C.; resources, M.C., M.X. and Y.C.; data curation, R.P., Y.H. and G.W.; writing—original draft preparation, Y.J. and S.C.; writing—review and editing, F.Y. and Y.G.; supervision, Y.G. and F.Y.; project administration, F.Y.; funding acquisition, F.Y. and M.X. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (82360658) and the Hainan Province Clinical Medical Center (QWYH202175). The funding sources had no role in the study design, data collection, analysis, interpretation, or writing of this report.
Institutional Review Board Statement
This study received approval from the Ethics Committee of the Hainan Women and Children’s Medical Center (protocol code HNWCMC 111, approved on 9 November 2023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data: Availability Statement:All sequences analyzed during this study are available from the NCBI database (GenBank accession No. OQ248592-OQ248607, OR140542-OR140554 and PV800154-PV800198).
Acknowledgments
The authors wish to thank the patient for participating in this study, and the support from the Hainan Tropical Infectious Diseases Biobank and the Open Foundation of Key Laboratory of Tropical Translational Medicine of Ministry of Education, Hainan Medical University.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Wang, X.; Li, Y.; Shi, T.; Bont, L.J.; Chu, H.Y.; Zar, H.J.; Wahi-Singh, B.; Ma, Y.; Cong, B.; Sharland, E.; et al. Global disease burden of and risk factors for acute lower respiratory infections caused by respiratory syncytial virus in preterm infants and young children in 2019: a systematic review and meta-analysis of aggregated and individual participant data. Lancet 2024, 403, 1241–1253. [Google Scholar] [CrossRef]
- I Mazur, N.; Caballero, M.T.; Nunes, M.C. Severe respiratory syncytial virus infection in children: burden, management, and emerging therapies. Lancet 2024, 404, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Escalante, J.C.; Comas-García, A.; Bernal-Silva, S.; Robles-Espinoza, C.D.; Gómez-Leal, G.; Noyola, D.E. Respiratory syncytial virus A genotype classification based on systematic intergenotypic and intragenotypic sequence analysis. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, Y.; Wang, W.; Li, C.; An, S.; Lu, G.; Jin, R.; Xu, B.; Zhou, Y.; Chen, A.; et al. A multi-center study on Molecular Epidemiology of Human Respiratory Syncytial Virus from Children with Acute Lower Respiratory Tract Infections in the Mainland of China between 2015 and 2019. Virol. Sin. 2021, 36, 1475–1483. [Google Scholar] [CrossRef]
- Chow, E.J.; Uyeki, T.M.; Chu, H.Y. The effects of the COVID-19 pandemic on community respiratory virus activity. Nat. Rev. Microbiol. 2022, 21, 1–16. [Google Scholar] [CrossRef]
- Poole, S.; Brendish, N.J.; Clark, T.W. SARS-CoV-2 has displaced other seasonal respiratory viruses: Results from a prospective cohort study. J. Infect. 2020, 81, 966–972. [Google Scholar] [CrossRef]
- Baker, R.E.; Park, S.W.; Yang, W.; Vecchi, G.A.; Metcalf, C.J.E.; Grenfell, B.T. The impact of COVID-19 nonpharmaceutical interventions on the future dynamics of endemic infections. Proc. Natl. Acad. Sci. 2020, 117, 30547–30553. [Google Scholar] [CrossRef] [PubMed]
- Huh, K.; Jung, J.; Hong, J.; Kim, M.; Ahn, J.G.; Kim, J.-H.; Kang, J.-M. Impact of Nonpharmaceutical Interventions on the Incidence of Respiratory Infections During the Coronavirus Disease 2019 (COVID-19) Outbreak in Korea: A Nationwide Surveillance Study. Clin. Infect. Dis. 2020, 72, e184–e191. [Google Scholar] [CrossRef]
- Tang, J.W.; Bialasiewicz, S.; Dwyer, D.E.; Dilcher, M.; Tellier, R.; Taylor, J.; Hua, H.; Jennings, L.; Kok, J.; Levy, A.; et al. Where have all the viruses gone? Disappearance of seasonal respiratory viruses during the COVID-19 pandemic. J. Med Virol. 2021, 93, 4099–4101. [Google Scholar] [CrossRef]
- Liang, W.N.; Yao, J.H.; Wu, J.; Liu, X.; Liu, J.; Zhou, L.; Chen, C.; Wang, G.F.; Wu, Z.Y.; Yang, W.Z.; et al. Experience and thinking on the normalization stage of prevention and control of COVID-19 in China. 2021, 101, E001. [Google Scholar]
- Wei, X.; Wang, L.; Li, M.; Qi, J.; Kang, L.; Hu, G.; Gong, C.; Wang, C.; Wang, Y.; Huang, F.; et al. Novel imported clades accelerated the RSV surge in Beijing, China, 2023-2024. J. Infect. 2024, 89, 106321. [Google Scholar] [CrossRef]
- Chang, M.; Shi, S.; Jin, Y.; Wang, G.; Peng, R.; An, J.; Huang, Y.; Hu, X.; Tang, C.; Niu, Y.; et al. Epidemiology and Genetic Evolutionary Analysis of Influenza Virus Among Children in Hainan Island, China, 2021–2023. Pathogens 2025, 14, 142. [Google Scholar] [CrossRef]
- Xiao, M.; Banu, A.; Jia, Y.; Chang, M.; Wang, G.; An, J.; Huang, Y.; Hu, X.; Tang, C.; Li, Z.; et al. Circulation pattern and genetic variation of rhinovirus infection among hospitalized children on Hainan Island, before and after the dynamic zero-COVID policy, from 2021 to 2023. J. Med Virol. 2024, 96, e29755. [Google Scholar] [CrossRef]
- Xiao, M.; Banu, A.; Zeng, X.; Shi, S.; Peng, R.; Chen, S.; Ge, N.; Tang, C.; Huang, Y.; Wang, G.; et al. Epidemiology of Human Parainfluenza Virus Infections among Pediatric Patients in Hainan Island, China, 2021–2023. Pathogens 2024, 13, 740. [Google Scholar] [CrossRef] [PubMed]
- Agoti, C.N.; Mwihuri, A.G.; Sande, C.J.; Onyango, C.O.; Medley, G.F.; Cane, P.A.; Nokes, D.J. Genetic Relatedness of Infecting and Reinfecting Respiratory Syncytial Virus Strains Identified in a Birth Cohort From Rural Kenya. J. Infect. Dis. 2012, 206, 1532–1541. [Google Scholar] [CrossRef]
- Cohen, R.; Ashman, M.; Taha, M.-K.; Varon, E.; Angoulvant, F.; Levy, C.; Rybak, A.; Ouldali, N.; Guiso, N.; Grimprel, E. Pediatric Infectious Disease Group (GPIP) position paper on the immune debt of the COVID-19 pandemic in childhood, how can we fill the immunity gap? Infect. Dis. Now 2021, 51, 418–423. [Google Scholar] [CrossRef]
- Piret, J.; Boivin, G. Viral Interference between Respiratory Viruses. Emerg. Infect. Dis. 2022, 28, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Umar, S.; Yang, R.; Wang, X.; Liu, Y.; Ke, P.; Qin, S. Molecular epidemiology and characteristics of respiratory syncytial virus among hospitalized children in Guangzhou, China. Virol. J. 2023, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Phyu, W.W.; Htwe, K.T.Z.; Saito, R.; Kyaw, Y.; Lin, N.; Dapat, C.; Osada, H.; Chon, I.; Win, S.M.K.; Hibino, A.; et al. Evolutionary analysis of human respiratory syncytial virus collected in Myanmar between 2015 and 2018. Infect. Genet. Evol. 2021, 93, 104927. [Google Scholar] [CrossRef]
- Li, J.; Chon, I.; Phyu, W.W.; Kyaw, Y.; Aye, M.M.; Setk, S.; Win, S.M.K.; Yoshioka, S.; Wagatsuma, K.; Sun, Y.; et al. Molecular epidemiological surveillance of respiratory syncytial virus infection in Myanmar from 2019 to 2023. Sci. Rep. 2025, 15, 1–12. [Google Scholar] [CrossRef]
- Tramuto, F.; Maida, C.M.; Mazzucco, W.; Costantino, C.; Amodio, E.; Sferlazza, G.; Previti, A.; Immordino, P.; Vitale, F. Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons. Pathogens 2023, 12, 1099. [Google Scholar] [CrossRef]
- Zhang, Y.; Pu, D.; Liu, Q.; Li, B.; Lu, B.; Cao, B. Epidemic Outbreak of Respiratory Syncytial Virus Infection After the end of the Zero-COVID-19 Policy in China: Molecular Characterization and Disease Severity Associated With a Novel RSV-B Clade. J. Med Virol. 2025, 97, e70343. [Google Scholar] [CrossRef]
- Langedijk, A.C.; Vrancken, B.; Lebbink, R.J.; Wilkins, D.; Kelly, E.J.; Baraldi, E.; Santos, A.H.M.d.L.; Danilenko, D.M.; Choi, E.H.; Palomino, M.A.; et al. The genomic evolutionary dynamics and global circulation patterns of respiratory syncytial virus. Nat. Commun. 2024, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.S.; Cowling, B.J.; Peiris, J.M.; Chan, E.L.; Wong, W.H.; Lee, K.P. Effects of Nonpharmaceutical COVID-19 Interventions on Pediatric Hospitalizations for Other Respiratory Virus Infections, Hong Kong. Emerg. Infect. Dis. 2022, 28, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Ritschel, T.; Pascual, G.; Brakenhoff, J.P.J.; Keogh, E.; Furmanova-Hollenstein, P.; Lanckacker, E.; Wadia, J.S.; A Gilman, M.S.; Williamson, R.A.; et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLOS Pathog. 2018, 14, e1006935. [Google Scholar] [CrossRef]
- Fedechkin, S.O.; George, N.L.; Wolff, J.T.; Kauvar, L.M.; DuBois, R.M. Structures of respiratory syncytial virus G antigen bound to broadly neutralizing antibodies. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Escalante, J.C.; Comas-García, A.; Bernal-Silva, S.; Noyola, D.E. Respiratory syncytial virus B sequence analysis reveals a novel early genotype. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Kim, H.N.; Hwang, J.; Yoon, S.-Y.; Lim, C.S.; Cho, Y.; Lee, C.-K.; Nam, M.-H. Molecular characterization of human respiratory syncytial virus in Seoul, South Korea, during 10 consecutive years, 2010–2019. PLOS ONE 2023, 18, e0283873. [Google Scholar] [CrossRef]
Figure 1.
Monthly RSV activity among hospitalized pediatric patients with ARTIs in Hainan, January 2021–December 2024. Monthly numbers of hospitalized children with ARTIs (yellow shaded area), RSV-positive cases (orange shaded area), and RSV positivity (%) (black line, right y-axis). Blue shaded intervals indicate the periods of local COVID-19 outbreaks in Hainan. Abbreviations: RSV, respiratory syncytial virus; ARTI, acute respiratory tract infection.
Figure 1.
Monthly RSV activity among hospitalized pediatric patients with ARTIs in Hainan, January 2021–December 2024. Monthly numbers of hospitalized children with ARTIs (yellow shaded area), RSV-positive cases (orange shaded area), and RSV positivity (%) (black line, right y-axis). Blue shaded intervals indicate the periods of local COVID-19 outbreaks in Hainan. Abbreviations: RSV, respiratory syncytial virus; ARTI, acute respiratory tract infection.
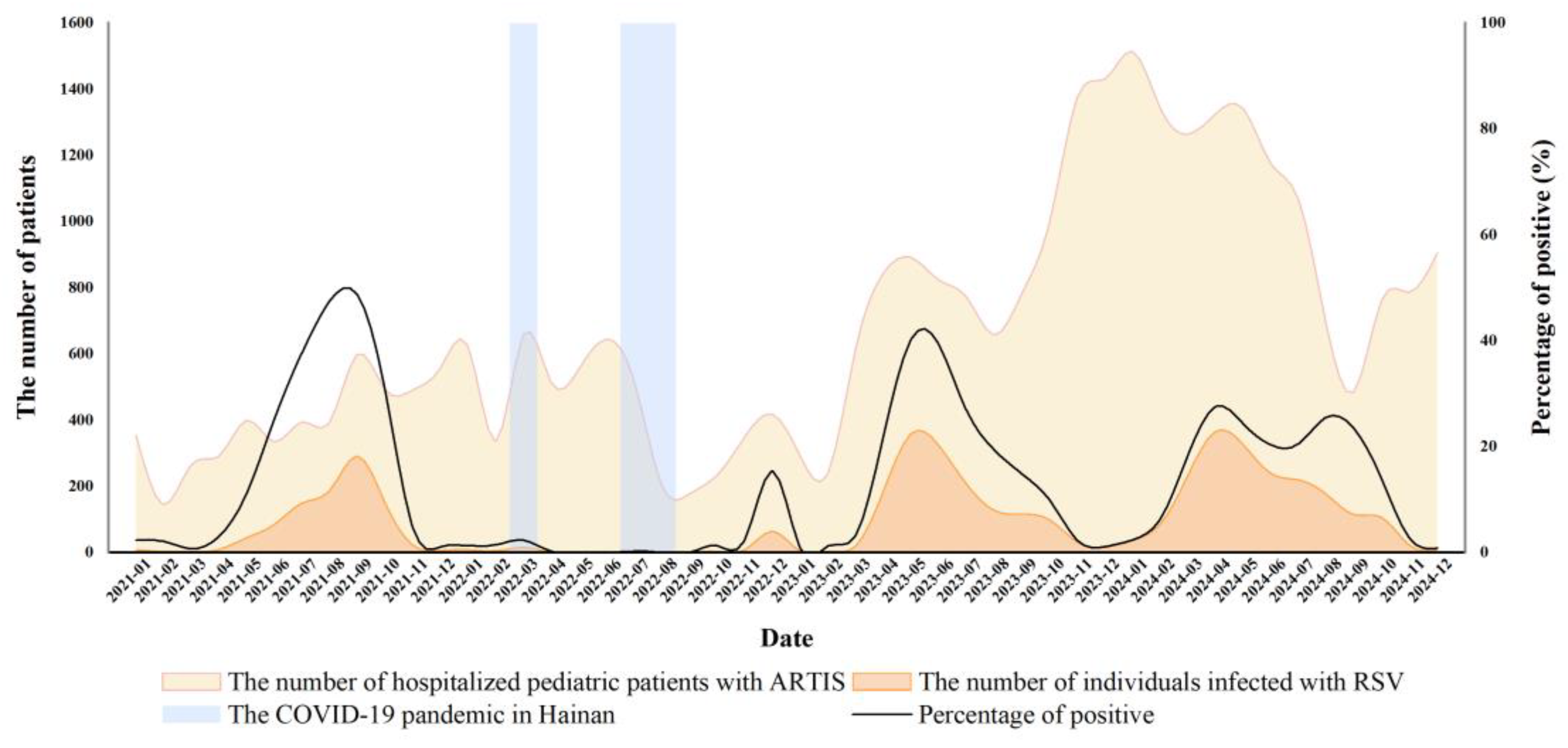
Figure 2.
Age distribution and RSV positivity among hospitalized pediatric patients with ARTIs, January 2021–December 2024. RSV-positive cases (bars, left y-axis) and RSV positivity (%) (lines, right y-axis) by age group (0–1, 1–3, 3–7, and 7–18 years) across study years.
Figure 2.
Age distribution and RSV positivity among hospitalized pediatric patients with ARTIs, January 2021–December 2024. RSV-positive cases (bars, left y-axis) and RSV positivity (%) (lines, right y-axis) by age group (0–1, 1–3, 3–7, and 7–18 years) across study years.
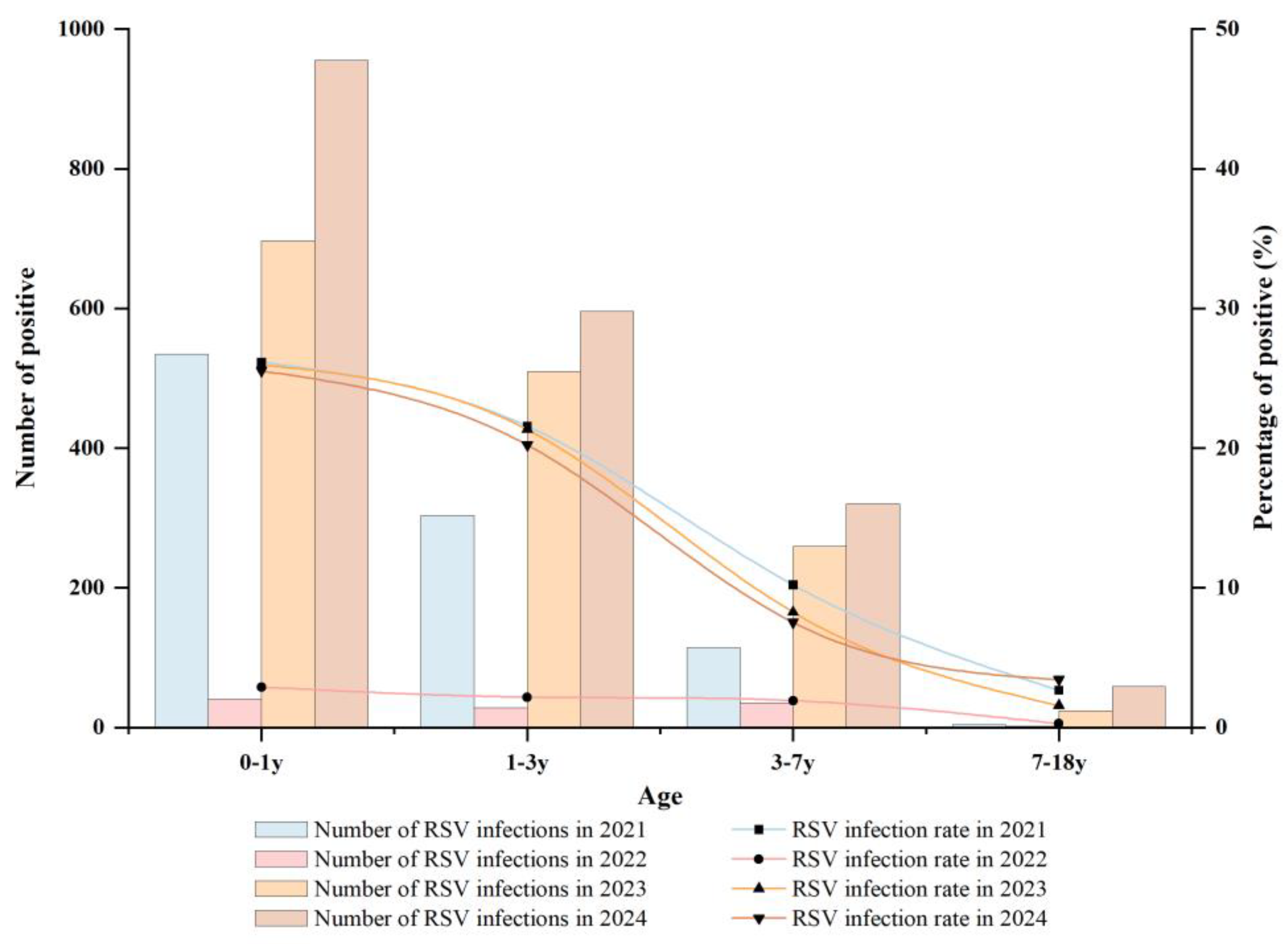
Figure 3.
Phylogenetic analysis of RSV strains detected on Hainan Island based on partial G-gene sequences. (A) RSV-A; (B) RSV-B. Trees were constructed using partial G-gene sequences from this study together with representative reference sequences. Genotypes/lineages are labeled as shown. Hainan sequences are highlighted in the figure. Scale bars indicate the number of substitutions per site.
Figure 3.
Phylogenetic analysis of RSV strains detected on Hainan Island based on partial G-gene sequences. (A) RSV-A; (B) RSV-B. Trees were constructed using partial G-gene sequences from this study together with representative reference sequences. Genotypes/lineages are labeled as shown. Hainan sequences are highlighted in the figure. Scale bars indicate the number of substitutions per site.
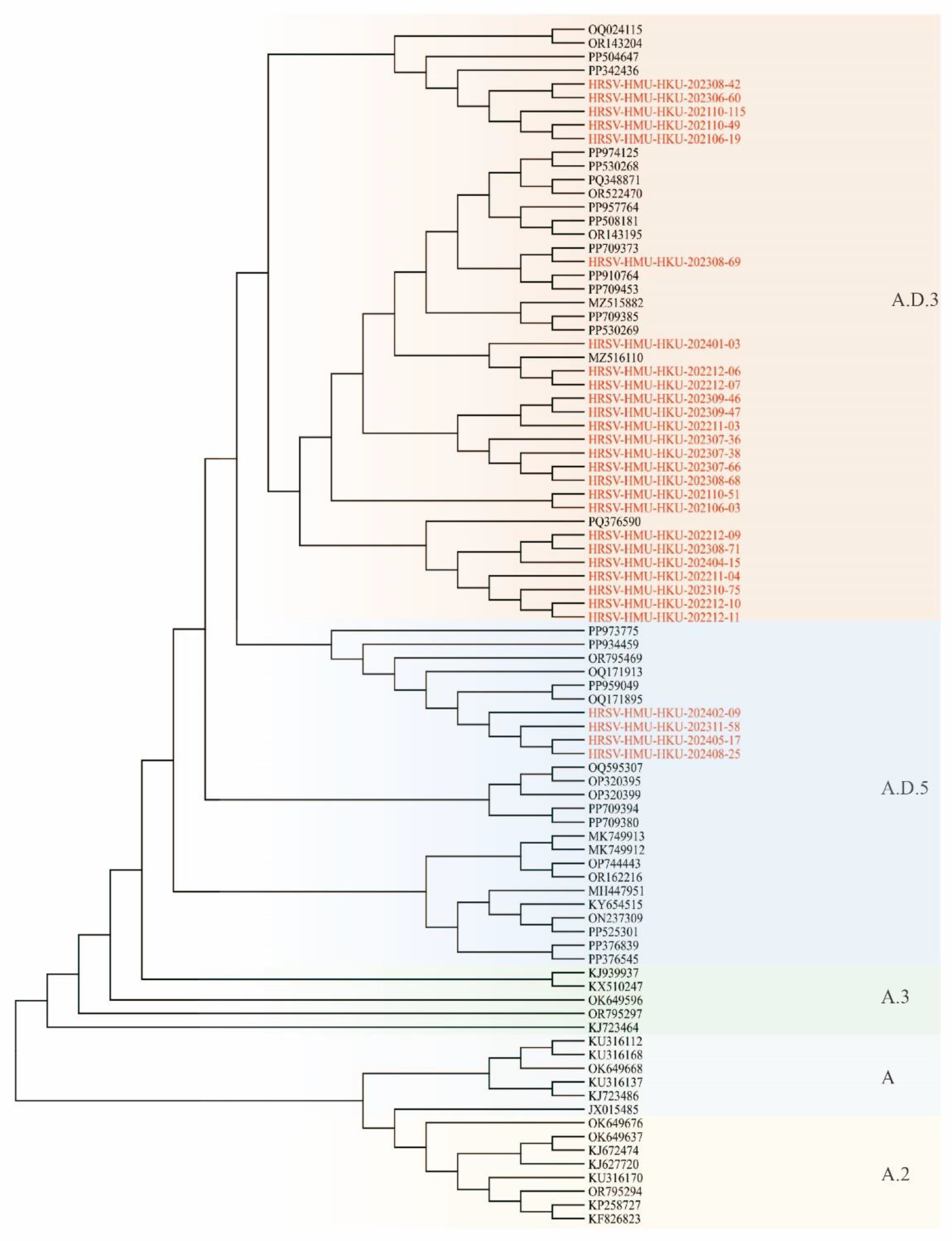
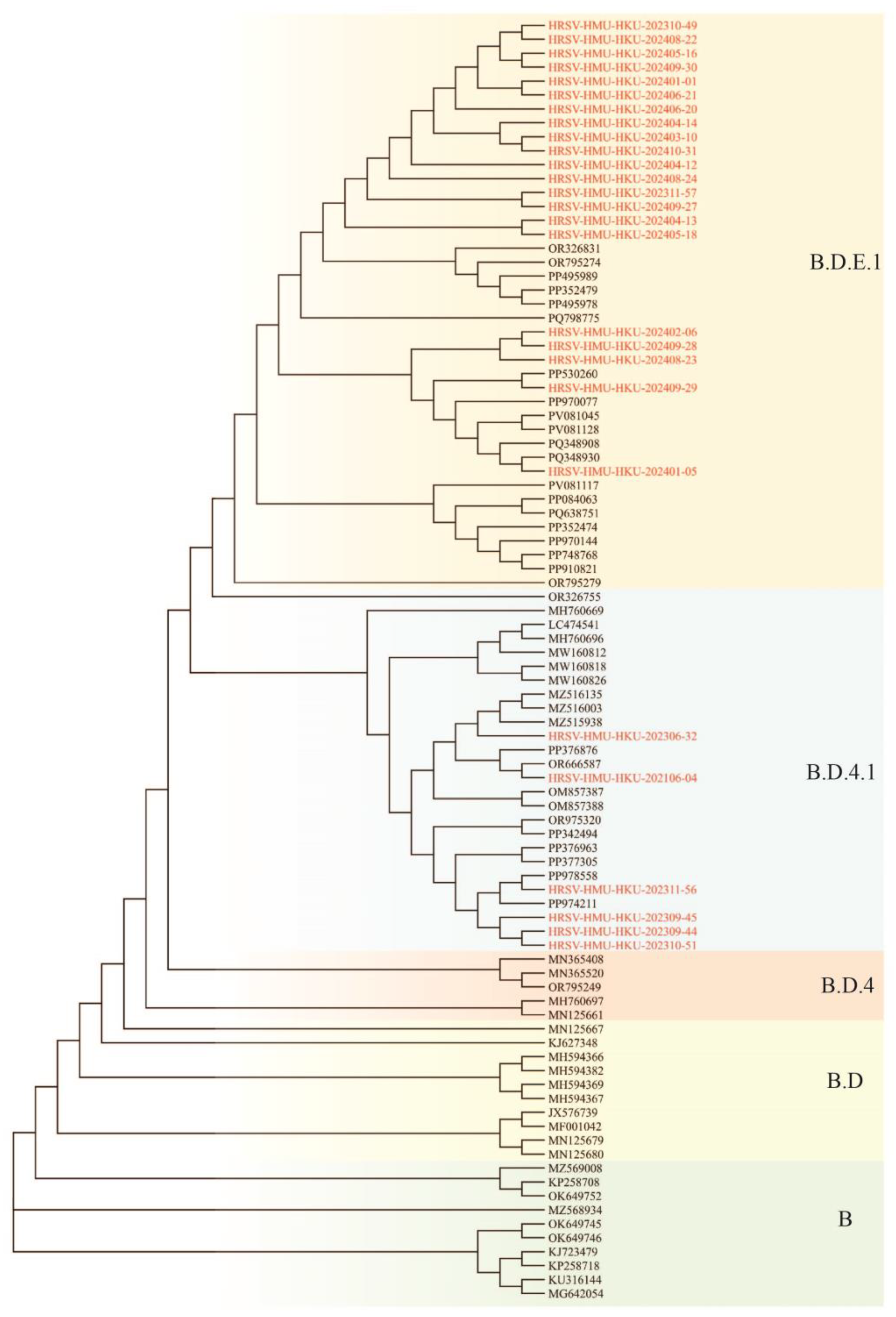
Figure 4.
Amino-acid alignments of the second hypervariable region (HVR2) of the RSV G protein. (A) RSV-A strains aligned to the ON1 prototype strain (GenBank: JN257693); alignment corresponds to amino-acid positions 227–321 of the G protein. (B) RSV-B strains aligned to the BA9 prototype strain (GenBank: AY333364); alignment corresponds to amino-acid positions 214–315. Identical residues are indicated by dots. The duplicated segments are marked by dotted boxes (23 amino acids for RSV-A and 20 amino acids for RSV-B).
Figure 4.
Amino-acid alignments of the second hypervariable region (HVR2) of the RSV G protein. (A) RSV-A strains aligned to the ON1 prototype strain (GenBank: JN257693); alignment corresponds to amino-acid positions 227–321 of the G protein. (B) RSV-B strains aligned to the BA9 prototype strain (GenBank: AY333364); alignment corresponds to amino-acid positions 214–315. Identical residues are indicated by dots. The duplicated segments are marked by dotted boxes (23 amino acids for RSV-A and 20 amino acids for RSV-B).
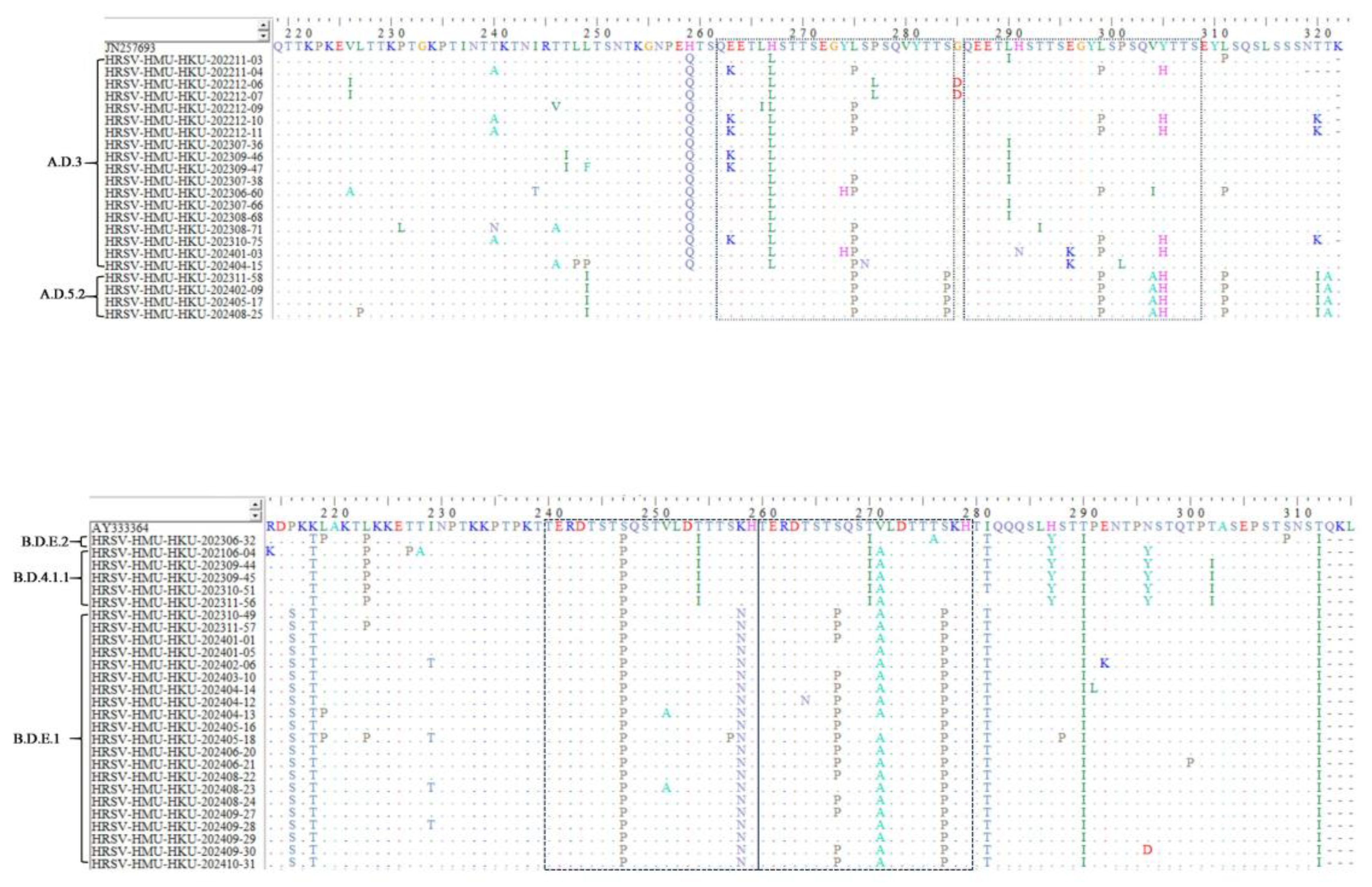

Table 1.
RSV prevalence among hospitalized pediatric patients with ARTIs by sex, 2021–2024.
| Gender | 2021 | 2022 | 2023 | 2024 | Total |
|---|---|---|---|---|---|
| Male | 618/3002(20.57) | 54/3173(1.70) | 968/5951(16.27) | 1192/7507(15.88) | 2832/19633(14.42) |
| Female | 339/1719(19.72) | 52/2054(2.53) | 522/3781(13.81) | 738/5142(14.35) | 1651/12696(13.00) |
| χ2 value | 0.51 | 4.32 | 10.79 | 5.50 | 13.03 |
| p value | 0.477 | 0.038 | 0.001 | 0.019 | <0.001 |
Table 2.
RSV prevalence among hospitalized pediatric patients with ARTIs by season, 2021–2024.
| season | 2021 | 2022 | 2023 | 2024 |
|---|---|---|---|---|
| Spring | 56(5.85) | 16(0.93) | 569(24.14) | 926(23.56) |
| Summer | 422(37.54) | 1(0.07) | 655(29.09) | 632(21.64) |
| Autumn | 462(29.20) | 12(1.57) | 245(7.76) | 240(11.77) |
| Winter | 17(1.61) | 77(5.61) | 21(1.07) | 132(3.51) |
| χ2 value | 636.76 | 126.29 | 917.55 | 721.83 |
| p value | <0.001 | <0.001 | <0.001 | <0.001 |
Table 3.
RSV prevalence among hospitalized pediatric patients with ARTIs by age, 2021–2024.
| Age | 2021 | 2022 | 2023 | 2024 | Total | χ2 value | p value |
|---|---|---|---|---|---|---|---|
| 0-1y | 535/2045(26.16) | 41/1413(2.90) | 697/2684(25.97) | 955/3740(25.53) | 2228/9882(22.55) | 364.70 | <0.001 |
| 1-3y | 304/1409(21.57) | 28/1305(2.15) | 510/2391(21.33) | 596/2946(20.23) | 1438/8051(17.86) | 263.83 | <0.001 |
| 3-7y | 114/1117(10.20) | 35/1809(1.93) | 259/3140(8.25) | 320/4248(7.53) | 728/10314(7.06) | 97.50 | <0.001 |
| 7-18y | 4/150(2.67) | 2/700(0.29) | 24/1517(1.58) | 59/1715(3.44) | 89/4082(2.18) | 27.26 | <0.001 |
| Total | 957/4721(20.27) | 106/5227(2.03) | 1490/9732(15.31) | 1930/12649(15.25) | 4483/32329(13.86) | 812.97 | <0.001 |
| χ2 value | 144.17 | 16.293 | 643.24 | 743.12 | 1597.85 | / | |
| p value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
Table 4.
Prediction of N-glycosylation sites in the G gene of RSV-A strains found in Hainan Province.
Table 4.
Prediction of N-glycosylation sites in the G gene of RSV-A strains found in Hainan Province.
| lineage | Sample number | N85 | N100 | N103 | N135 | N179 | N237 |
|---|---|---|---|---|---|---|---|
| ON1 | |||||||
| JN257693 | + | + | + | + | |||
| A.D.3 | |||||||
| HRSV-HMU-HKU-202211-03 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202211-04 | + | + | + | + | |||
| HRSV-HMU-HKU-202212-06 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202212-07 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202212-09 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202212-10 | + | + | + | + | |||
| HRSV-HMU-HKU-202212-11 | + | + | + | ||||
| HRSV-HMU-HKU-202401-03 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202404-15 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202307-36 | + | + | + | + | + | + | |
| HRSV-HMU-HKU-202309-46 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202309-47 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202307-38 | + | + | + | + | + | + | |
| HRSV-HMU-HKU-202306-60 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202307-66 | + | + | + | + | + | + | |
| HRSV-HMU-HKU-202308-68 | + | + | + | + | + | + | |
| HRSV-HMU-HKU-202308-71 | + | + | + | + | + | ||
| HRSV-HMU-HKU-202310-75 | + | + | + | + | |||
| A.D.5.2 | |||||||
| HRSV-HMU-HKU-202408-25 | + | + | + | + | |||
| HRSV-HMU-HKU-202405-17 | + | + | + | + | |||
| HRSV-HMU-HKU-202311-58 | + | + | + | + | |||
| HRSV-HMU-HKU-202402-09 | + | + | + | + |
Table 5.
Prediction of N-glycosylation sites in the G gene of RSV-B strains found in Hainan Province.
Table 5.
Prediction of N-glycosylation sites in the G gene of RSV-B strains found in Hainan Province.
| lineage | Sample number | N230 | N258 | N296 | N310 |
|---|---|---|---|---|---|
| BA9 | |||||
| AY333364 | + | + | + | ||
| B.D.4.1.1 | |||||
| HRSV-HMU-HKU-202306-32 | + | + | |||
| HRSV-HMU-HKU-202106-04 | + | ||||
| HRSV-HMU-HKU-202309-44 | + | ||||
| HRSV-HMU-HKU-202309-45 | + | ||||
| HRSV-HMU-HKU-202310-51 | + | ||||
| B.D.E.1 | |||||
| HRSV-HMU-HKU-202311-56 | + | ||||
| HRSV-HMU-HKU-202310-49 | + | + | + | ||
| HRSV-HMU-HKU-202311-57 | + | + | + | ||
| HRSV-HMU-HKU-202401-01 | + | + | + | ||
| HRSV-HMU-HKU-202401-05 | + | + | + | ||
| HRSV-HMU-HKU-202402-06 | + | + | + | ||
| HRSV-HMU-HKU-202403-10 | + | + | + | ||
| HRSV-HMU-HKU-202404-14 | + | + | + | ||
| HRSV-HMU-HKU-202404-12 | + | + | + | ||
| HRSV-HMU-HKU-202404-13 | + | + | + | ||
| HRSV-HMU-HKU-202405-16 | + | + | + | ||
| HRSV-HMU-HKU-202405-18 | + | + | + | ||
| HRSV-HMU-HKU-202406-20 | + | + | + | ||
| HRSV-HMU-HKU-202406-21 | + | + | + | ||
| HRSV-HMU-HKU-202408-22 | + | + | + | ||
| HRSV-HMU-HKU-202408-23 | + | + | + | ||
| HRSV-HMU-HKU-202408-24 | + | + | + | ||
| HRSV-HMU-HKU-202409-27 | + | + | + | ||
| HRSV-HMU-HKU-202409-28 | + | + | + | ||
| HRSV-HMU-HKU-202409-29 | + | + | + | ||
| HRSV-HMU-HKU-202409-30 | + | + | |||
| HRSV-HMU-HKU-202410-31 | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



