
Submitted:
26 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
In this study, three distinct hydrolysates which designated Dur-I, Dur-II, and Dur-III, were generated from extrusion-pretreated Durvillaea antarctica biomass by applying viscozyme, cellulase, and α-amylase, respectively. Dur-III had a higher proportion of low-molecular-weight polysaccharides as compared to Dur-I and Dur-II. Chemical composition determination and FTIR analyses revealed that Dur-I, Dur-II, and Dur-III contained fucose-containing sulfated polysaccharides. To investigate neuroprotective properties of Dur-I, Dur-II, and Dur-III, rotenone (Rot) was added to SH-SY5Y cells that had been pretreated with Dur-I/II/III. Here, flow cytometry was employed to assess changes in mitochondrial membrane potential (MMP), Bcl-2 expression, cytochrome c release, caspase-9, -8, and -3 activation, as well as DNA fragmentation. The protective effect of Dur-I/II/III pretreatment of SH-SY5Y cells on the Rot-induced death process was further investigated using cell cycle and annexin V-fluorescein isothiocyanate (FITC) / PI (propidium iodide) double staining analyses. The results reveal that the Rot-induced apoptotic factors were all recovered by the pretreatment of Dur-I/II/III. Moreover, cell cycle and annexin V-FITC/PI double staining analyses also indicated that Dur-I/II/III were capable of protecting SH-SY5Y cells from Rot-induced cytotoxicity. Therefore, these Dur extracts are considered as good candidates for the prevention and treatment of neurodegeneration induced by oxidative stress.
Keywords:
1. Introduction
2. Results and Discussion
2.1. Preparation of Enzyme Extracts (Dur-I, Dur-II, and Dur-III) from Extrusion-Pretreated D. antarctica and Physicochemical Characteristics of Dur-I, Dur-II, and Dur-III
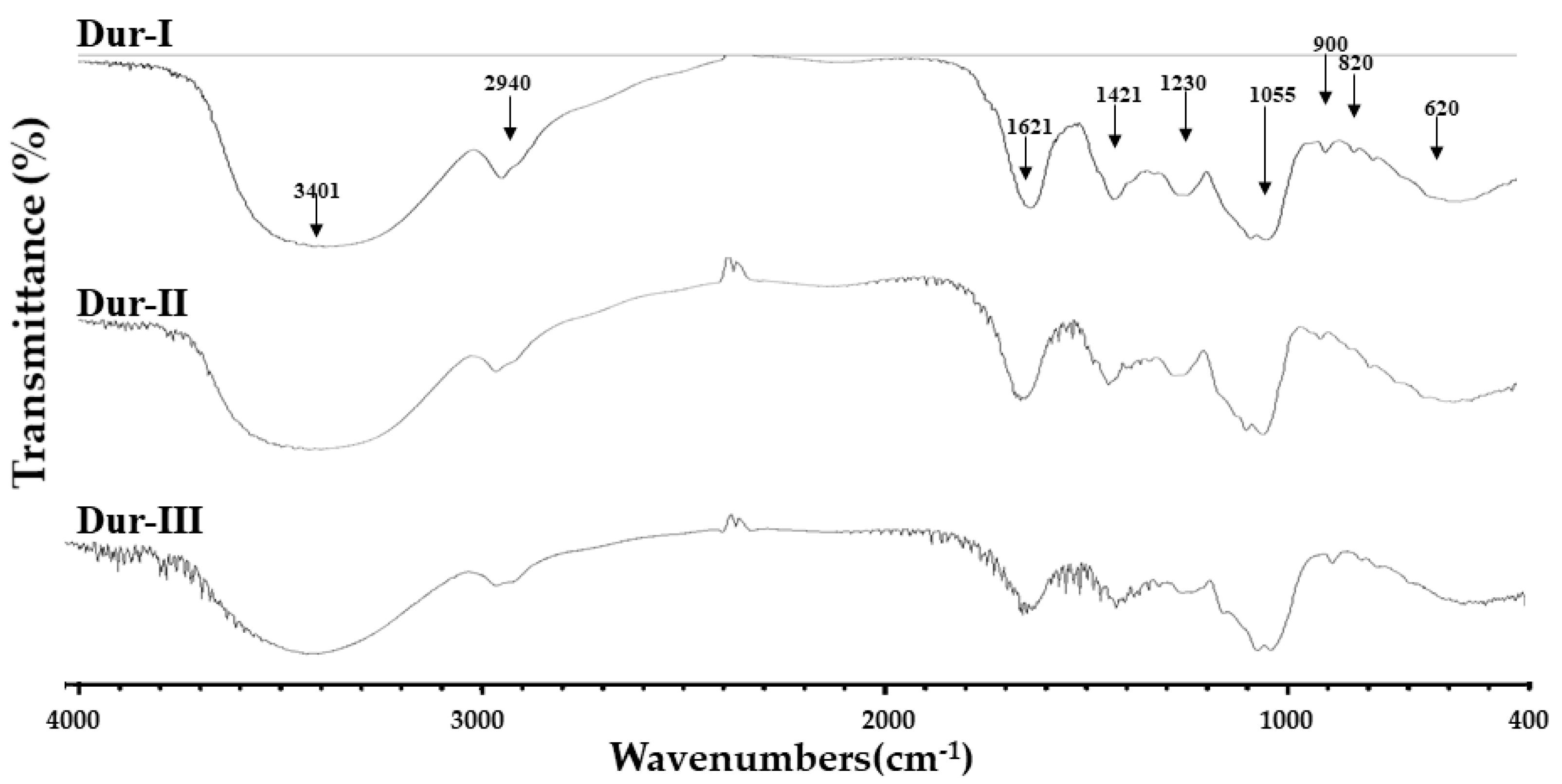
2.2. Elucidation of Structural Characterization of Dur-I, Dur-II, and Dur-III by FTIR and NMR Techniques
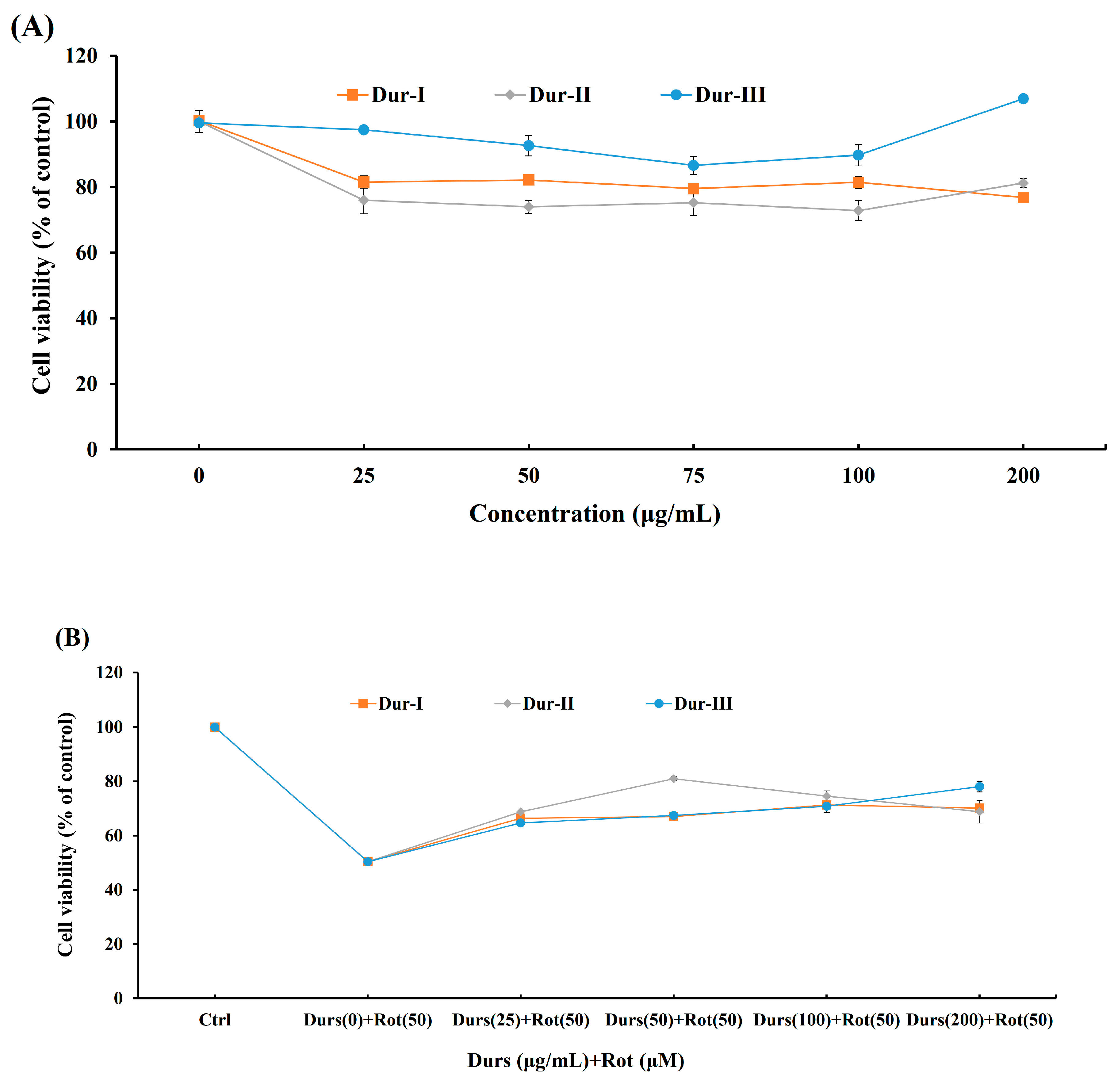
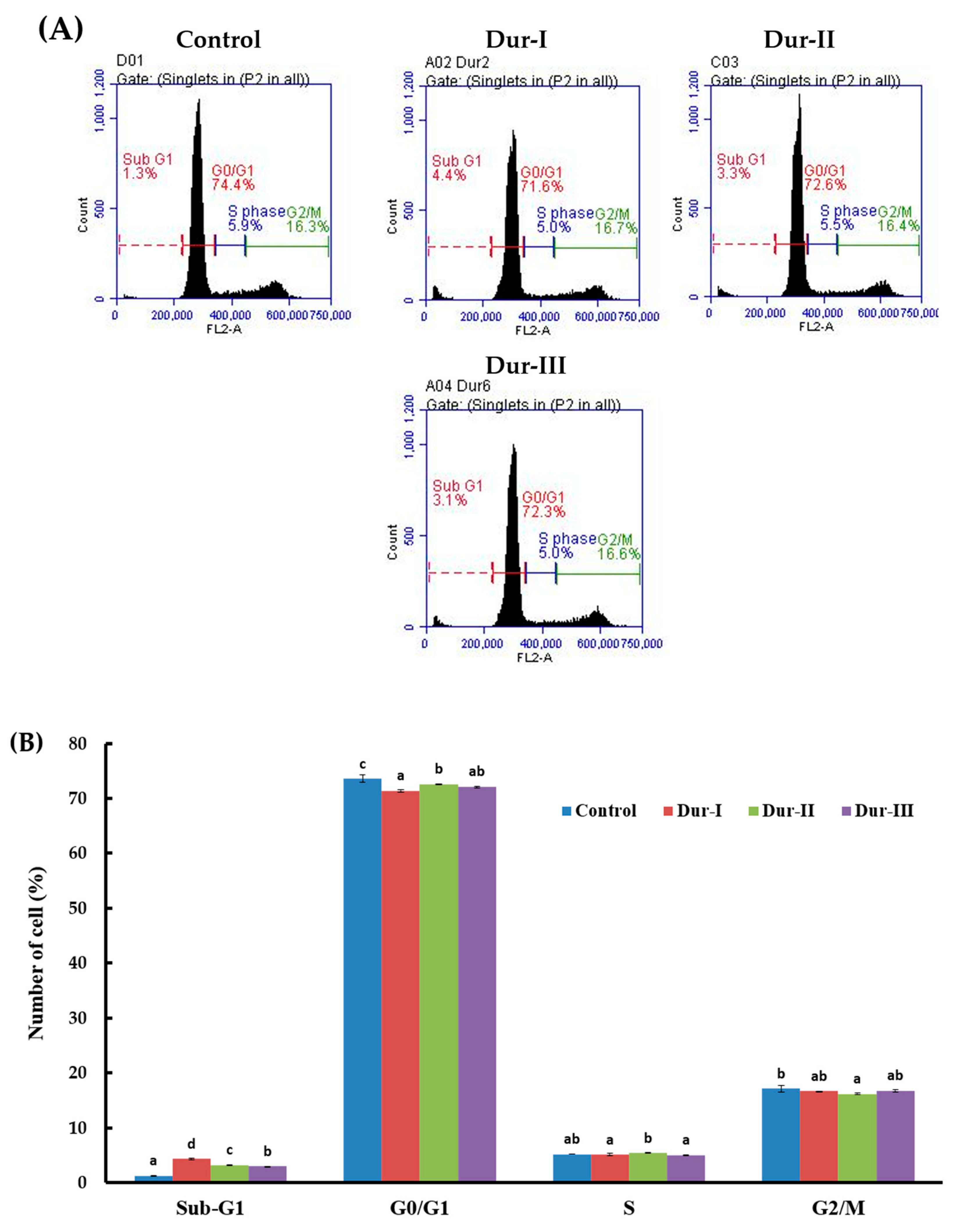
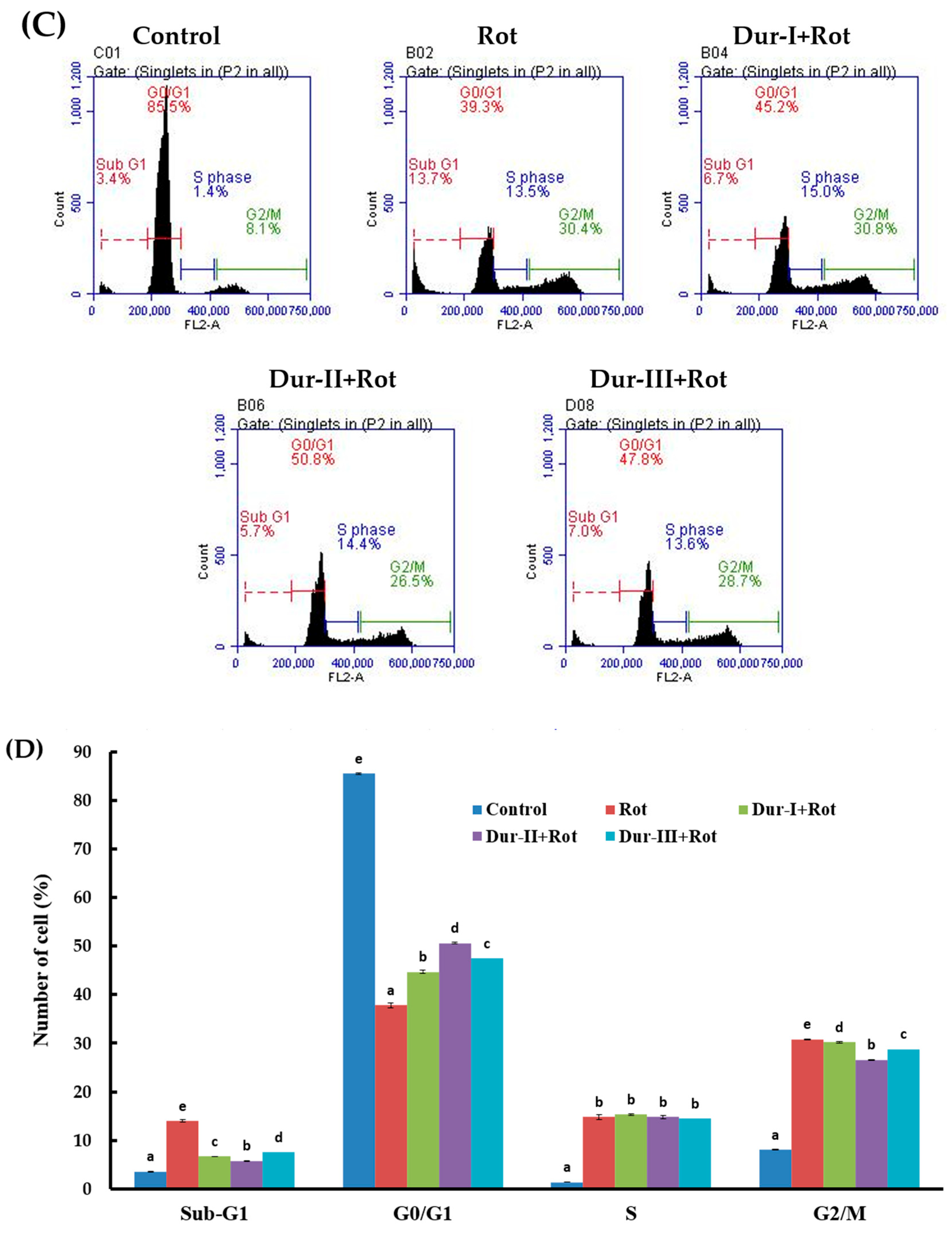
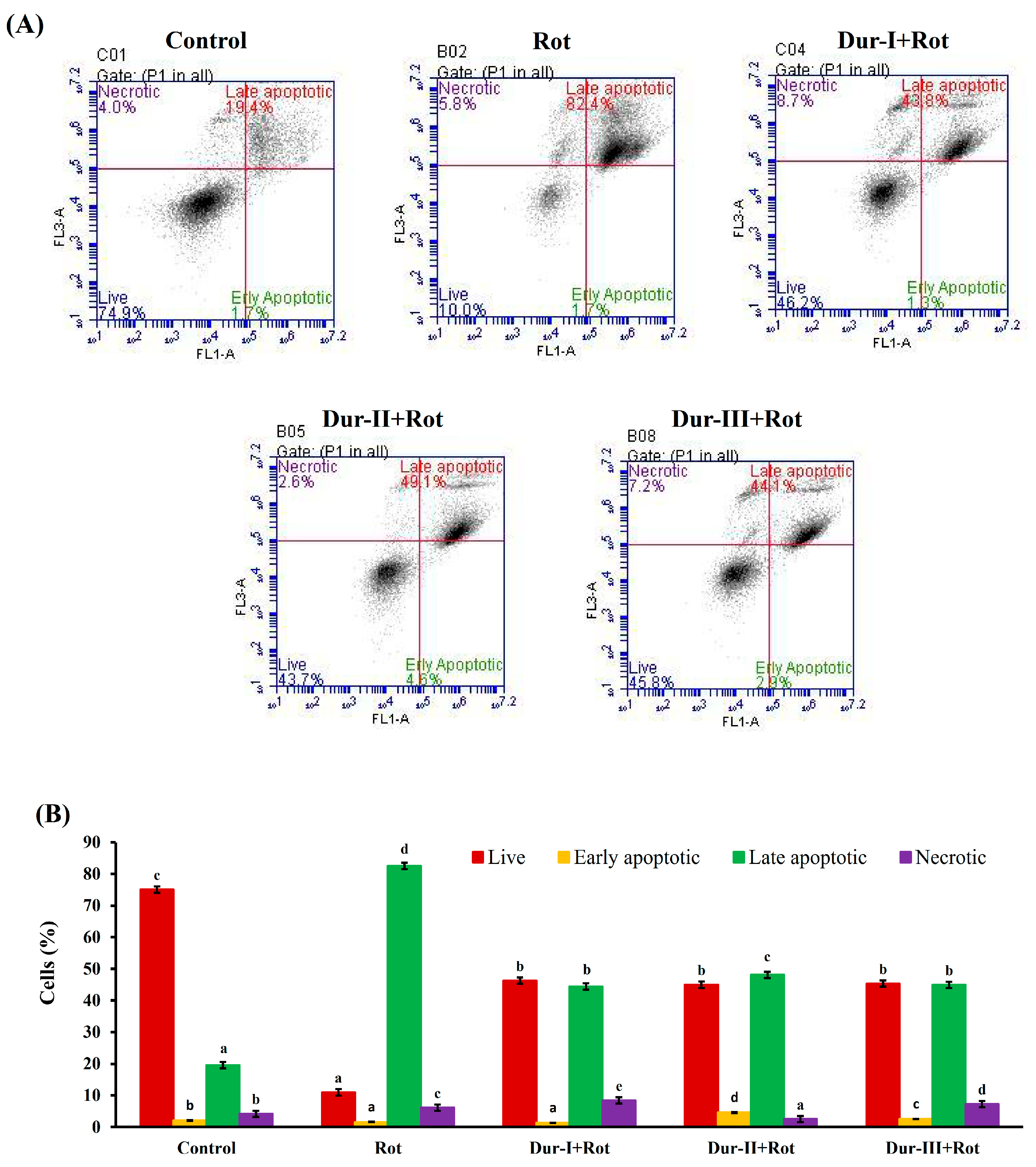
2.3. Dur-I, Dur-II, and Dur-III attenuated Rotenone-Induced Apoptosis in SH-SY5Y Neuronal Cells
3. Materials and Methods
3.1. Materials
3.2. Extrusion Method
3.3. Seaweed Extraction by Enzymes
3.4. Molecular Weight Analysis
3.5. Chemical Methods
3.6. Analysis of Monosaccharide Composition
3.7. FTIR Spectroscopy
3.8. Nuclear Magnetic Resonance (NMR) Spectroscopy
3.9. Cell Culture
3.10. Cell Viability Analysis
3.11. Flow Cytometry-Based Analyses
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| MMP | Mitochondrial membrane potential |
| FITC | Fluorescein isothiocyanate |
| PI | Propidium iodide |
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| ROS | Reactive oxygen species |
| SEC | Size-exclusion chromatography |
| HPSEC | High-performance size exclusion chromatography |
| HSQC | Heteronuclear single quantum coherence |
| Rot | Rotenone |
| mPTP | Mitochondrial permeability transition pore |
| mitoK ATP | Mitochondrial ATP-sensitive potassium |
| TMRE | Tetramethylrhodamine ethyl ester |
| Bcl-2 | B-cell lymphoma-2 |
| AIF | Apoptosis-inducing factor |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| PS | Phosphatidylserine |
| BSA | Bovine serum albumin |
| KBr | Potassium bromide |
| TFA | Trifluoroacetic acid |
| NMR | Nuclear magnetic resonance |
| FTIR | Fourier transform infrared spectroscopy |
References
- Li, H.; Wang, N.; Mao, X. Incidental vs. engineered nanoparticles in Alzheimer’s and Parkinson’s disease: Pathological pathways and therapeutic interventions. Nano Res. 2025, 18. [CrossRef]
- Olufunmilayo, E.O.; Gerke-Duncan, M.B.; Holsinger, R.M.D. Oxidative Stress and Antioxidants in Neurodegenerative Disorders. Antioxidants 2023, 12, 517. [CrossRef]
- Goel, P.; Chakrabarti, S.; Goel, K.; Bhutani, K.; Chopra, T.; Bali, S. Neuronal cell death mechanisms in Alzheimer’s disease: An insight. Front. Mol. Neurosci. 2022, 15, 937133. [CrossRef]
- Kumari, S.; Dhapola, R.; Sharma, P.; Paidlewar, M.; Vellingiri, B.; Medhi, B.; HariKrishnaReddy, D. Unravelling neuronal death mechanisms: The role of cytokines and chemokines in immune imbalance in Alzheimer’s disease progression. Ageing Res. Rev. 2025, 112, 102883. [CrossRef]
- Li, Y.-Y.; Qin, Z.-H.; Sheng, R. The Multiple Roles of Autophagy in Neural Function and Diseases. Neurosci. Bull. 2023, 40, 363–382. [CrossRef]
- Zhou, Q.-L.; Wang, Z.; Chen, W.-T.; Liu, X.-F.; Cheong, K.-L.; Zou, Y.-X.; Zhong, S.-Y.; Li, R. The structural characteristics, biological activities and mechanisms of bioactive brown seaweed polysaccharides: A review. J. Funct. Foods 2024, 119. [CrossRef]
- Wu, J.; Wang, H.; Liu, Y.; Xu, B.; Du, B.; Yang, Y. Effect of Ultrasonic Irradiation on the Physicochemical and Structural Properties of Laminaria japonica Polysaccharides and Their Performance in Biological Activities. Molecules 2022, 28, 8. [CrossRef]
- Matsuzaki, K.; Tatsumi, D.; Sugiyama, T.; Hiraoka, M.; Igura, N.; Tsubaki, S. Accelerating Sulfated Polysaccharides Extraction from Fast-Growing Ulva Green Seaweed by Frequency-Controlled Microwaves. ACS Omega 2024, 9, 29896–29903. [CrossRef]
- Otero, P.; Carpena, M.; Garcia-Oliveira, P.; Echave, J.; Soria-Lopez, A.; Garcia-Perez, P.; Fraga-Corral, M.; Cao, H.; Nie, S.; Xiao, J.; et al. Seaweed polysaccharides: Emerging extraction technologies, chemical modifications and bioactive properties. Crit. Rev. Food Sci. Nutr. 2021, 63, 1901–1929. [CrossRef]
- Alfinaikh, R.S.; Alamry, K.A.; Hussein, M.A. Sustainable and biocompatible hybrid materials-based sulfated polysaccharides for biomedical applications: a review. RSC Adv. 2025, 15, 4708–4767. [CrossRef]
- Shih, M.-K.; Hou, C.-Y.; Dong, C.-D.; Patel, A.K.; Tsai, Y.-H.; Lin, M.-C.; Xu, Z.-Y.; Perumal, P.K.; Kuo, C.-H.; Huang, C.-Y. Production and Characterization of Durvillaea antarctica Enzyme Extract for Antioxidant and Anti-Metabolic Syndrome Effects. Catalysts 2022, 12, 1284. [CrossRef]
- Sanjeewa, K.A.; Herath, K.; Kim, Y.-S.; Jeon, Y.-J.; Kim, S.-K. Enzyme-assisted extraction of bioactive compounds from seaweeds and microalgae. TrAC Trends Anal. Chem. 2023, 167. [CrossRef]
- Jiang, G.; Ameer, K.; Ramachandraiah, K.; Feng, X.; Tan, C.; Cai, N. Effects of synergistic application of Viscozyme L–wet ball milling on structural, physicochemical and functional properties of insoluble dietary fiber from ginseng residue. LWT 2024, 209. [CrossRef]
- Thanh, H.D.T.; Nguyen, T.T. High gravity enzymatic hydrolysis of non–gelatinized starch from black - purple rice. Vietnam. J. Sci. Technol. 2023, 61, 787–797. [CrossRef]
- Silva, G.H.; Flores-Alarcón, M.A.; Durán, L.M.; Roberto, I.C. Enzyme supplementation as a strategy for enhancing the cellulose and hemicellulose conversion yields at high solids content of pretreated rice straw. Process. Biochem. 2025, 157, 207–215. [CrossRef]
- Zhang, Z.; Xing, J.; Li, X.; Lu, X.; Liu, G.; Qu, Y.; Zhao, J. Review of research progress on the production of cellulase from filamentous fungi. Int. J. Biol. Macromol. 2024, 277, 134539. [CrossRef]
- Sahu, P.K.; Singh, R.; Shrivastava, M.; Darjee, S.; Mageshwaran, V.; Phurailtpam, L.; Rohatgi, B. Microbial production of α-amylase from agro-waste: An approach towards biorefinery and bio-economy. Energy Nexus 2024, 14. [CrossRef]
- Worth, A.J.; Basu, S.S.; Snyder, N.W.; Mesaros, C.; Blair, I.A. Inhibition of Neuronal Cell Mitochondrial Complex I with Rotenone Increases Lipid β-Oxidation, Supporting Acetyl-Coenzyme A Levels. J. Biol. Chem. 2014, 289, 26895–26903. [CrossRef]
- Ganguly, U.; Bir, A.; Chakrabarti, S. Cytotoxicity of mitochondrial Complex I inhibitor rotenone: A complex interplay of cell death pathways. Bioenerg. Commun. 2022, 2022, 14-14.
- Fu, X.; Zhan, Y.; Li, N.; Yu, D.; Gao, W.; Gu, Z.; Zhu, L.; Li, R.; Zhu, C. Enzymatic Preparation of Low-Molecular-Weight Laminaria japonica Polysaccharides and Evaluation of Its Effect on Modulating Intestinal Microbiota in High-Fat-Diet-Fed Mice. Front. Bioeng. Biotechnol. 2022, 9, 820892. [CrossRef]
- Jayawardena, T.U.; Nagahawatta, D.P.; Fernando, I.P.S.; Kim, Y.-T.; Kim, J.-S.; Kim, W.-S.; Lee, J.S.; Jeon, Y.-J. A Review on Fucoidan Structure, Extraction Techniques, and Its Role as an Immunomodulatory Agent. Mar. Drugs 2022, 20, 755. [CrossRef]
- Akter, A.; Alam Sobuj, M.K.; Islam, S.; Chakroborty, K.; Tasnim, N.; Ayon, M.H.; Hossain, F.; Rafiquzzaman, S. Seaweed polysaccharides: Sources, structure and biomedical applications with special emphasis on antiviral potentials. Futur. Foods 2024, 10. [CrossRef]
- Baltrusch, K.; Torres, M.D.; Domínguez, H. Characterization, ultrafiltration, depolymerization and gel formulation of ulvans extracted via a novel ultrasound-enzyme assisted method. Ultrason. Sonochemistry 2024, 111, 107072. [CrossRef]
- Barzkar, N.; Rungsardthong, V.; Jahromi, S.T.; Laraib, Q.; Das, R.; Babich, O.; Sukhikh, S. A recent update on fucoidonase: source, Isolation methods and its enzymatic activity. Front. Mar. Sci. 2023, 10. [CrossRef]
- Zhang, S.; Chen, L.; Shang, N.; Wu, K.; Liao, W. Recent Advances in the Structure, Extraction, and Biological Activity of Sargassum fusiforme Polysaccharides. Mar. Drugs 2025, 23, 98. [CrossRef]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Fourier Transform Infrared (FTIR) Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [CrossRef]
- Shao, P.; Pei, Y.P.; Fang, Z.X.; Sun, P.L. Effects of partial desulfation on antioxidant and inhibition of DLD cancer cell of Ulva fasciata polysaccharide. Int. J. Biol. Macromol. 2014, 65, 307–313. [CrossRef]
- Soto-Vásquez, M.R.; Alvarado-García, P.A.A.; Youssef, F.S.; Ashour, M.L.; Bogari, H.A.; Elhady, S.S. FTIR Characterization of Sulfated Polysaccharides Obtained from Macrocystis integrifolia Algae and Verification of Their Antiangiogenic and Immunomodulatory Potency In Vitro and In Vivo. Mar. Drugs 2022, 21, 36. [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [CrossRef]
- Chen, S.; Sathuvan, M.; Zhang, X.; Zhang, W.; Tang, S.; Liu, Y.; Cheong, K.-L. Characterization of polysaccharides from different species of brown seaweed using saccharide mapping and chromatographic analysis. BMC Chem. 2021, 15, 1–11. [CrossRef]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: Structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [CrossRef]
- Ammar, H.H.; Lajili, S.; Ben Said, R.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Physico-chemical characterization and pharmacological evaluation of sulfated polysaccharides from three species of Mediterranean brown algae of the genus Cystoseira. DARU J. Pharm. Sci. 2015, 23, 1–8. [CrossRef]
- Amin, M.N.G.; Mischnick, P.; Rosenau, T.; Böhmdorfer, S. Refined linkage analysis of the sulphated marine polysaccharide fucoidan of Cladosiphon okamuranus with a focus on fucose. Carbohydr. Polym. 2024, 342. [CrossRef]
- Vijayabaskar, P.; Vaseela, N.; Thirumaran, G. Potential antibacterial and antioxidant properties of a sulfated polysaccharide from the brown marine algae Sargassum swartzii. Chin. J. Nat. Med. 2012, 10, 421–428. [CrossRef]
- Immanuel, G.; Sivagnanavelmurugan, M.; Marudhupandi, T.; Radhakrishnan, S.; Palavesam, A. The effect of fucoidan from brown seaweed Sargassum wightii on WSSV resistance and immune activity in shrimp Penaeus monodon (Fab). Fish Shellfish. Immunol. 2012, 32, 551–564. [CrossRef]
- Mensah, E.O.; Kanwugu, O.N.; Panda, P.K.; Adadi, P. Marine fucoidans: Structural, extraction, biological activities and their applications in the food industry. Food Hydrocoll. 2023, 142. [CrossRef]
- Usoltseva, R.V.; Zueva, A.O.; Malyarenko, O.S.; Anastyuk, S.D.; Moiseenko, O.P.; Isakov, V.V.; Kusaykin, M.I.; Jia, A.; Ermakova, S.P. Structure and Metabolically Oriented Efficacy of Fucoidan from Brown Alga Sargassum muticum in the Model of Colony Formation of Melanoma and Breast Cancer Cells. Mar. Drugs 2023, 21, 486. [CrossRef]
- Castro-Riquelme, C.L.; Maldonado, E.A.L. Multifunctional Characterization of Fucoidan: Structural Insights and Efficient Removal of Toxic Metal Ions. J. Res. Updat. Polym. Sci. 2025, 14, 54–61. [CrossRef]
- Ohmes, J.; Mikkelsen, M.D.; Nguyen, T.T.; Tran, V.H.N.; Meier, S.; Nielsen, M.S.; Ding, M.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Depolymerization of fucoidan with endo-fucoidanase changes bioactivity in processes relevant for bone regeneration. Carbohydr. Polym. 2022, 286, 119286. [CrossRef]
- Dhahri, M. Cystoseira myrica: from beach-cast seaweed to fucoidan with antioxidant and anticoagulant capacity. Front. Mar. Sci. 2023, 10, 1327408. [CrossRef]
- Sichert, A.; Le Gall, S.; Klau, L.J.; Laillet, B.; Rogniaux, H.; Aachmann, F.L.; Hehemann, J.-H. Ion-exchange purification and structural characterization of five sulfated fucoidans from brown algae. Glycobiology 2020, 31, 352–357. [CrossRef]
- Saravana, P.S.; Karuppusamy, S.; Rai, D.K.; Wanigasekara, J.; Curtin, J.; Tiwari, B.K. Elimination of Ethanol for the Production of Fucoidans from Brown Seaweeds: Characterization and Bioactivities. Mar. Drugs 2024, 22, 493. [CrossRef]
- Ferrante, L.; Rajpoot, K.; Jeeves, M.; Ludwig, C. Automated analysis for multiplet identification from ultra-high resolution 2D-1H, 13C-HSQC NMR spectra. Wellcome Open Res. 2023, 7, 262.
- Kirsten, N.; Ohmes, J.; Mikkelsen, M.D.; Nguyen, T.T.; Blümel, M.; Wang, F.; Tasdemir, D.; Seekamp, A.; Meyer, A.S.; Fuchs, S. Impact of Enzymatically Extracted High Molecular Weight Fucoidan on Lipopolysaccharide-Induced Endothelial Activation and Leukocyte Adhesion. Mar. Drugs 2023, 21, 339. [CrossRef]
- Yong, S.J.; Veerakumarasivam, A.; Teoh, S.L.; Lim, W.L.; Chew, J. Lactoferrin Protects Against Rotenone-Induced Toxicity in Dopaminergic SH-SY5Y Cells through the Modulation of Apoptotic-Associated Pathways. J. Mol. Neurosci. 2024, 74, 1–18. [CrossRef]
- Kaya, Z.B.; Santiago-Padilla, V.; Lim, M.; Boschen, S.L.; Atilla, P.; McLean, P.J. Optimizing SH-SY5Y cell culture: exploring the beneficial effects of an alternative media supplement on cell proliferation and viability. Sci. Rep. 2024, 14, 1–11. [CrossRef]
- Guo, D.; Liu, Z.; Zhou, J.; Ke, C.; Li, D. Significance of Programmed Cell Death Pathways in Neurodegenerative Diseases. Int. J. Mol. Sci. 2024, 25, 9947. [CrossRef]
- Han, X.; Han, B.; Zhao, Y.; Li, G.; Wang, T.; He, J.; Du, W.; Cao, X.; Gan, J.; Wang, Z.; et al. Rosmarinic Acid Attenuates Rotenone-Induced Neurotoxicity in SH-SY5Y Parkinson’s Disease Cell Model through Abl Inhibition. Nutrients 2022, 14, 3508. [CrossRef]
- Kent, A.C.; El Baradie, K.B.Y.; Hamrick, M.W. Targeting the Mitochondrial Permeability Transition Pore to Prevent Age-Associated Cell Damage and Neurodegeneration. Oxidative Med. Cell. Longev. 2021, 2021. [CrossRef]
- Bernardi, P.; Gerle, C.; Halestrap, A.P.; Jonas, E.A.; Karch, J.; Mnatsakanyan, N.; Pavlov, E.; Sheu, S.-S.; Soukas, A.A. Identity, structure, and function of the mitochondrial permeability transition pore: controversies, consensus, recent advances, and future directions. Cell Death Differ. 2023, 30, 1869–1885. [CrossRef]
- Gottschalk, B.; Koshenov, Z.; Malli, R.; Graier, W.F. Implications of mitochondrial membrane potential gradients on signaling and ATP production analyzed by correlative multi-parameter microscopy. Sci. Rep. 2024, 14, 1–12. [CrossRef]
- Dadsena, S.; Jenner, A.; García-Sáez, A.J. Mitochondrial outer membrane permeabilization at the single molecule level. Cell. Mol. Life Sci. 2021, 78, 3777–3790. [CrossRef]
- Vianello, C.; Bello, F.D.; Shin, S.H.; Schiavon, S.; Bean, C.; Rebelo, A.P.M.; Knedlík, T.; Esfahani, E.N.; Costiniti, V.; Lacruz, R.S.; et al. High-Throughput Microscopy Analysis of Mitochondrial Membrane Potential in 2D and 3D Models. Cells 2023, 12, 1089. [CrossRef]
- Huang, C.-Y.; Kuo, C.-H.; Chen, P.-W. Compressional-Puffing Pretreatment Enhances Neuroprotective Effects of Fucoidans from the Brown Seaweed Sargassum hemiphyllum on 6-Hydroxydopamine-Induced Apoptosis in SH-SY5Y Cells. Molecules 2017, 23, 78. [CrossRef]
- Mitra, S.; Rauf, A.; Sutradhar, H.; Sadaf, S.; Hossain, J.; Soma, M.A.; Bin Emran, T.; Ahmad, B.; Aljohani, A.S.; Al Abdulmonem, W.; et al. Potential candidates from marine and terrestrial resources targeting mitochondrial inhibition: Insights from the molecular approach. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2022, 264, 109509. [CrossRef]
- Nguyen, T.T.; Wei, S.; Nguyen, T.H.; Jo, Y.; Zhang, Y.; Park, W.; Gariani, K.; Oh, C.-M.; Kim, H.H.; Ha, K.-T.; et al. Mitochondria-associated programmed cell death as a therapeutic target for age-related disease. Exp. Mol. Med. 2023, 55, 1595–1619. [CrossRef]
- Nguyen, T.T.M.; Gillet, G.; Popgeorgiev, N. Caspases in the Developing Central Nervous System: Apoptosis and Beyond. Front. Cell Dev. Biol. 2021, 9, 702404. [CrossRef]
- Araya, L.E.; Soni, I.V.; Hardy, J.A.; Julien, O. Deorphanizing Caspase-3 and Caspase-9 Substrates In and Out of Apoptosis with Deep Substrate Profiling. ACS Chem. Biol. 2021, 16, 2280–2296. [CrossRef]
- Yang, W.-N.; Chen, P.-W.; Huang, C.-Y. Compositional Characteristics and In Vitro Evaluations of Antioxidant and Neuroprotective Properties of Crude Extracts of Fucoidan Prepared from Compressional Puffing-Pretreated Sargassum crassifolium. Mar. Drugs 2017, 15, 183. [CrossRef]
- Costigan, A.; Hollville, E.; Martin, S.J. Discriminating Between Apoptosis, Necrosis, Necroptosis, and Ferroptosis by Microscopy and Flow Cytometry. Curr. Protoc. 2023, 3, e951. [CrossRef]
- Olivares-Molina, A.; Fernández, K. Comparison of different extraction techniques for obtaining extracts from brown seaweeds and their potential effects as angiotensin I-converting enzyme (ACE) inhibitors. J. Appl. Phycol. 2016, 28, 1295–1302. [CrossRef]

| Variables of Hydrolysis | Dur-I | Dur-II | Dur-III |
| Variables for extrusion-pretreatment | Feed moisture content: 35%, feed rate: 10.4 kg/h, barrel temperature: 115 °C, screw speed: 360 rpm, rounded die with 5 mm opening | ||
| Enzymes | Viscozyme | Cellulase | α-Amylase |
| Hydrolysis conditions | pH 6.0, 40 °C, 17 h | pH 6.0, 40 °C, 17 h | pH 6.0, 40 °C, 17 h |
| Extraction yield | Dur-I | Dur-II | Dur-III |
| Extraction yield (%) | 41.3±2.3 1, a | 45.5±3.0 a | 44.2±0.8 a |
| Molecular weight | Dur-I | Dur-II | Dur-III |
| Peak 1 (MW (kDa) / Peak area (%)) | 201.4/ 96.8 | 213.5/ 99.9 | 170.5 /46.7 |
| Peak 2 (MW (kDa) / Peak area (%)) | 10.9 /3.2 | 15.6 / 0.1 | 3.68 / 54.3 |
| Chemical composition | Dur-I | Dur-II | Dur-III |
| Total sugar (%) 1 | 43.8 ± 0.2 b | 36.9 ± 0.4 a | 73.7 ± 0.3 c |
| Fucose (%) 1 | 14.6 ± 0.5 a | 15.8 ± 0.5 b | 22.5 ± 0.5 c |
| Sulfate (%) 1 | 38.1 ± 0.0 b | 41.9 ± 0.0 c | 25.6 ± 0.0 a |
| Uronic acid (%) 1 | 15.6 ± 0.2 a | 15.6 ± 0.3 a | 18.2 ± 0.4 b |
| Alginic acid (%) 1 | 4.54 ± 0.15 a | 4.59 ± 0.70 a | 6.78 ± 0.56 b |
| Polyphenols (%) 1 | 0.39 ± 0.02 a | 0.33 ± 0.00 a | 0.36 ± 0.06 a |
| Proteins (%) 1 | 0.93 ± 0.03 a | 1.13 ± 0.05 c | 1.03 ± 0.00 b |
| Monosaccharide composition (molar ratio) | Dur-I | Dur-II | Dur-III |
| Fucose | 1 | 1 | 1 |
| Rhamnose | 2.08 | 1.85 | 1.56 |
| Glucuronic acid | 0.06 | 0.06 | 0.04 |
| Galacturonic acid | 0.32 | 0.03 | 0.30 |
| Glucose | 0.01 | 1.10 | 0.95 |
| Galactose | 0.02 | 0.01 | 0.00 |
| Xylose | 0.17 | 0.10 | 0.18 |
| Factors | Control | Rot | Dur-I+Rot | Dur-II+Rot | Dur-III+Rot |
| Low mitochondrial membrane potential (%) 1 | 9.43 ± 0.12 a | 27.2 ± 0.2 c | 20.0 ± 0.4 b | 19.6 ± 0.5 b | 19.8 ± 0.1 b |
| Level of Bcl-2 (%) 1 | 87.7 ± 0.6 e | 65.2 ± 0.7 a | 69.3 ± 0.3 b | 73.5 ± 0.4 d | 71.4 ± 0.4 c |
| Release of cytochrome c (%) 1 | 55.6 ± 1.2 e | 19.2 ± 0.5 a | 39.3 ± 2.2 c | 50.2 ± 0.9 d | 35.9 ± 2.0 b |
| Caspase-9 activity (%) 1 | 31.3 ± 0.2 a | 63.8 ± 0.7 d | 49.6 ± 0.5 b | 51.4 ± 1.2 c | 50.7 ± 0.1 bc |
| Caspase-8 activity (%) 1 | 35.8 ± 0.9 a | 67.5 ± 2.3 d | 51.6 ± 0.4 b | 37.7 ± 1.1 a | 61.9 ± 0.7 c |
| Caspase-3 activity (%) 1 | 32.0 ± 0.1 a | 67.3 ± 0.6 d | 50.8 ± 0.5 b | 54.1 ± 1.2 c | 52.0 ± 0.1 b |
| DNA fragmentation (%) 1 | 43.0 ± 0.4 a | 69.2 ± 0.6 e | 59.2 ± 3.1 c | 63.0 ± 0.6 d | 48.6 ± 0.9 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



